bruno ernande ifremer, port-en-bessin, france fishace, mallorca 2006 quantitative genetics: analysis...
Post on 19-Dec-2015
219 views
TRANSCRIPT

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Quantitative genetics:Analysis of phenotypes and evolutionary dynamics
Bruno Ernande
Laboratoire Ressources Halieutiques
IFREMER
Port-en-Bessin, France

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Quantitative genetics
∎ Quantitative genetics are a macroscopic and statistical description of the effect of genes on phenotypes in the same manner that thermo-dynamics describe the macroscopic properties of gases emerging from the behavior of molecules behavior
∎ In QG, the genes are actually never observed, only their effect at the phenotypic level can be experimentally assessed
∎ Focus on quantitative traits determined by a large number of genes with small effects: multi-locus genetics
∎ Can be used analyze the sources of phenotypic variation and particularly genetic inheritance as well as to describe evolutionary dynamics

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Content
1. QG analysis of phenotypes: Basic model
2. QG analysis of phenotypes: Generalized models
3. QG analysis of phenotypes: Practice
4. Case study: using multivariate QG to infer the energetics underlying life history traits
5. QG analysis of probabilistic maturation reaction norms
6. Quantitative genetic models of evolutionary dynamics

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
1. QG analysis of phenotypes: Basic model

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning phenotypes: Basic model
∎ The phenotype of an individual can be viewed as resulting from a genetic and environmental contribution
z = g + ε
with g the genotypic value and ε the micro-environmental contribution
∎ The average micro-environmental contribution is assumed to be zero, E(ε)=0, so thatthe mean phenotype in the population is equal to the mean genotypic value
E(z) = E(g)
∎ Phenotypic variation within the population can be decomposed as
V(z) = V(g) + V(ε)
∎Micro-environmental contributions are assumed independent from each others and from genotypic values, so that
cov(z1,z2) = cov(g1,g2)
The covariance between phenotypes (i.e. phenotypic resemblance) is the covariance between individuals’ genotypic values.

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning genotypic value: One autosomal locus
∎ An individual with paternal and maternal alleles Bp and Bm at a given loci has a genotypic values g described as:
g = μ + ap + am + apam = μ + a + d
witha = ap + am the additive effect of alleles
d = apam the dominance interaction between alleles
B1B1 B1B2 B2B2
Genotypic value
+a
-a
+d
Genotype

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning genotypic value: One autosomal locus
∎ The expectations of additive and dominance effect are assumed to be zero, E(a) = 0 and E(d) = 0, and both effects are assumed independent, COV(a,d) = 0, so that
E(g) = μ
∎ Genotypic variation is then decomposed as
V(g) =V(a) + V(d)
∎ Assumptions:Mendelian inheritanceRandom matingSelection does not affect genotype frequencies

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Covariance between genotypic values: One autosomal locus
∎ Individual 1 with alleles B1p and B1m and individual 2 with alleles B2p and B2m
g1 = μ + a1p + a1m + a1pa1m and g2 = μ + a2p + a2m + a2pa2m
∎ There are three possible patterns of allelic identities in two individuals, each pattern occurring with respective probability p0, p1 and p2:
1. No identical alleles:
cov(g1, g2|no identical alleles) = 0
2. One pair of identical genes.For instance, B1p = B1p and maternal alleles independent yields a1p = a2p. Therefore,
cov(g1,g2|a1p = a2p)=1/2 V(a)
3. Two pairs of identical alleles, B1p = B2p and B1m = B2m:
cov(g1,g2|a1p = a2p and a1m = a2m) = V(g),
cov(a1,a2|a1p = a2p and a1m = a2m) = V(a)
cov(d1,d2|a1p = a2p and a1m = a2m) = V(d)
∎Averaging over these three possibilities gives
cov(g1,g2) = (0.5 p1 + p2) V(a) + p2 V(d)
∎ 0.5 p1 + p2 is twice the coefficient of co-ancestry of the two individuals

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning genotypic values: multi-locus
∎ Quantitative traits, such has morphological or life history traits, are generally determined by a large number of genes physically located at different loci dispersed within the genome.

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning genotypic values: multi-locus
∎ Quantitative traits, such has morphological or life history traits, are generally determined by a large number of genes physically located at different loci dispersed within the genome.
∎ In such case, the linear model used for one autosomal locus can be generalized:
Note the introduction of epistatic effects owing to the fact that alleles located at different physical loci can interact
i i i ip m p mi i
g a a a a( )
i j i j j i i jp p p m p m m mj i j
a a a a a a a a( )
i j j i j jp p m m p mj i j
a a a a a a( )
...
a d
aa
ad
Witha: additive effectd: dominance effectaa: epistatic additive effectad: epistatic dominance effect…

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning genotypic values: multi-locus
∎ In summary:
g = μ + a + d + aa + ad + dd + aaa +…
with all effects having expectation zero E(a) = E(d) = E(aa) = … = 0 and being independent
∎ Genotypic variation is then decomposed as
V(g) = V(a) + V(d) + V(aa) + V(ad) + V(dd) + V(aaa) +…
∎ Due the central limit theorem, all constituent components of the genotypic value are normally distributed, and thus
g follows N(μ,V(g)) and z follows N(μ,V(z)=V(g) + V(ε))
∎ Assumptions:Diploid autosomal loci with Mendelian inheritanceRandom matingSelection does not affect genotype frequenciesNo linkage between loci, i.e. free recombinationGametic phase equilibrium Independence of effects within-individuals

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Covariance between genotypic values: multi-locus
∎ Phenotypic covariance between two individuals is equal to their genetic covariance or the covariance of their genotypic values:
cov(z1,z2) = cov(g1,g2)
Due to the independence of effects,
cov(g1,g2) = cov (a1,a2) + cov (d1,d2) + cov (aa1,aa2) + cov (ad1,ad2) +…
∎ As for one autosomal locus, it can be shown that
cov(g1,g2) = (2Θ12)iΔ12jV(aidj)
with Θ12 the coefficient of co-ancestry between individual 1 and 2, and
Δ12 the coefficient of fraternity
Phenotypic resemblance is thus related to the genetic components of variance via individuals relatedness

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Relatedness between individuals
∎ The essential measures of relatedness between individuals are based on the concept of identity by descent
∎ Identity by descent: alleles are identical by descent if they are direct descendants of the same gene carried by a common ancestral individual (different from identity by state)
∎ Coefficient of co-ancestry: the probability that single genes drawn randomly from two individuals but at the same locus are identical by descent (single alleles: additivity)
∎ Coefficient of fraternity: the probability that the genotypes of two individuals at a given locus are identical by descent (pairs of alleles: dominance)
∎ In general, the following approximation is used:
cov(g1,g2) ≈ 2Θ12V(a) + Δ12V(d)
∎ Examples Θ Δ
Parent-offspring 1/4 0
Grandparent-grandchild 1/8 0
Half-sibs 1/8 0
Full-sibs 1/4 1/4

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Heritability
∎ The covariance between phenotypic and genotypic value
cov(z,g) = cov((g + ε),g) = V(g) + cov(g, ε) = V(g)
∎ The squared correlation coefficient between phenotypic and genotypic value is then:
ρ2(z,g) = V2(g)/(V0.5(g)V0.5(z))2=V(g)/V(z)=hb2
the broad sense heritability, the portion of phenotypic variance that is genetic.
∎ The narrow sense heritability is defined as the portion of phenotypic variance that is due to additive genetic effects only:
h2 = V(a)/V(z)
in other terms, those allelic effects that will be transmitted to the next generation with certainty

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Summary
∎ An individual’s phenotype z can be described as:
z = g + ε
with g = μ + a + d + aa + ad + dd +…
∎ All effects are independents and normally distributed
∎ z follows N(E(z) = E(g), V(z))
g follows N(E(g) = μ, V(g))
ε follows N(0, V(ε))
∎ Phenotypic variation can then be decomposed as :
V(z) = V(g) + V(ε) = V(a) + V(d) + V(aa) +…+ V(ε)
∎ Phenotypic resemblance is given by
cov(z1,z2)= cov(g1,g2) ≈ 2Θ12V(a) + Δ12V(d)
∎ Narrow sense heritability is defined as:
h2 = V(a)/V(z)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
2. QG analysis of phenotypes: Generalized models

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning phenotypes: introducing macro-environment
∎ Phenotypic plasticity: A given genotype can produce different phenotypes according to the environment it experiencesThis applies to both spatial and temporal variation in the environment
∎ Reaction norm: It is the systematic profile of phenotypes zij a genotype gi produces in response to a given range of environments ej. g1
g2
Environment e
Phen
otyp
e z

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Partitioning phenotypes: introducing macro-environment (Via 1984)
∎ An individual’s phenotype can then be described according to the following model:
z = g + e + g×e+ ε
∎ z follows N(E(z) = E(g) + E(e), V(z))
g follows N(E(g) = μg, V(g))
e follows N(E(e) = μe, V(e))
g×e follows N(0, V(g×e))
ε follows N(0, V(ε))
∎ Phenotypic variation can then be decomposed as
V(z) = V(g) + V(e) + V(g×e) + V(ε)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Macro-environment: Interpreting the model (Scheiner & Lyman 1989)
V(g)
Environnement e
z
g1
g2
V(z) = V(g) + V(e) + V(ε)
The two genotypes have the same plastic response: there is no genetic variance for plasticity
V(e)V(z)
Environnement e
z
g1
g2
V(z) = V(g) + V(e) + V(g×e)+ V(ε)
The two genotypes exhibit different plastic responses: there is some genetic variability for plasticity
V(e)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Heritability of trait mean and plasticity (Scheiner & Lyman 1989)
∎ The genetic variance of trait mean and plasticity are then defined as V(g) and V(g×e) respectively with
V(g) = V(a) + V(d) + V(aa) +…
V(g×e) = V(a×e) + V(d×e) + V(aa×e) +…
∎ Then, trait mean heritability is
hbm2 = V(g)/V(z) broad sense
hm2 = V(a)/V(z) narrow sense
∎ And trait plasticity heritability is
hbpl2 = V(g×e)/V(z) broad sense
hpl2 = V(a×e)/V(z) narrow sense

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Generalizing to multivariate phenotypes
∎ The basic model for univariate phenotypes can be generalized to multivariate phenotypes. Scalars are then replaced by vectors (lowercase) and matrices (uppercase):
z = g + e + g×e + ε
with
g = μ + a + d + aa + ad + dd +…
∎ So that a single trait zi is described as
zi = gi + ei + g×ei+ εi
∎ z follows a multivariate normal distribution MVN(E(z) = E(g) + E(e), Z) with Z being the phenotypic co-variance matrix
g follows MVN(E(g) = μg, G), G genetic co-variance matrix
e follows MVN(E(e) = μe, E), E environmental co-variance matrix
g×e follows N(0, G×E), G×E interaction co-variance matrixε follows N(0, ε), ε micro-environmental co-variance matrix

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Generalizing to multivariate phenotypes (Lande, 1979)
∎ As for univariate phenotypes, multivariate phenotypic co-variation can be decomposed in co-variance components
Z = G + E + G×E + ε
withG = A + D + AA + AD + DD +…
∎ This leads to phenotypic, genetic and environmental covariance and correlations between traits 1 and 2:
Co-variances:
covz(1,2) = covg(1,2) + cove(1,2) + covg×e(1,2) + covε(1,2)
Correlations
ρz(1,2) = covz(1,2) /(σz(1)σz(2))
ρA(1,2) = covg(1,2)/(σg(1)σg(2))
Relationship between correlations
ρz(1,2) = h1h2 ρA(1,2) + ρe(1,2) ((1-h12)(1-h2
2))0.5
ρe(1,2) = ρz(1,2) – h1h2 ρA(1,2)/((1-h12)(1-h2
2))0.5

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Generalizing to function-valued traits (Kirkpatrick & Heckman, 1989)
∎ Function-valued traits are traits that can be represented as functions of a given argument:Reaction norm: phenotype as a function of the environmentAllometric relationships: one morphological trait as a function of another oneDispersal kernel: rate of dispersion as a function of distance …
∎ A functional phenotype can be described as:z(e) = g(e)+ε(e)
∎ z(·) follows a Gaussian process with mean function mz(·) and covariance function Z(·) g(·) follows a Gaussian process with mean function mg(·) and covariance function G(·) ε(·) follows a Gaussian process with mean function mε(·) and covariance function E(·)
∎ Phenotypic covariance function can be decomposed in several covariance functions:
Z(e,e’) = G(e,e’) + E(e,e’)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Different approaches to plasticity: Univariate reaction norm
∎The historical view:Characters state approach
∎Correlation of the character states across environments, Falconer 60’s, Via and Lande 1985, Kawecki and Stearns 1993
{z1, z2, z3, z4}
g
1 2 3 4
z1
z2
z3
z4
Environment
Phenotype
∎Alternative view:Polynomial approach
∎Intercept and slope (z0,s) are considered as correlated traits, Gavrilets and Scheiner 1993a,b
slope, s
intercept
e0
z0
Environment
Phenotype
g
{z0,s}

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Different approaches to plasticity: Univariate reaction norm
∎The historical view:Characters state approach
∎Correlation of the character states across environments, Falconer 60’s, Via and Lande 1985, Kawecki and Stearns 1993
{z1, z2, z3, z4}
g
1 2 3 4
z1
z2
z3
z4
Environment
Phenotype
∎Alternative view:Functional or infinite-dimensional approach
∎Reaction norm is viewed as function of the environment characterizing a given genotype, Gomulkiewicz and Kirkpatrick 1992
Environment
Phenotype
z(e)
g
z(e)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Different approaches to plasticity: Bivariate reaction norm
grow
th 1
grow
th 2
growth 3
size
age
e.g. age and size at maturation
∎Correlated traits approach
∎Environmental dimension omitted: each point is the expression of the two traits in one environment (Stearns & Crandall, 1984)
∎Reaction norms with individual status as a determinant
∎Reaction norm is viewed as one trait being a function of the second one: functional approach of bivariate reaction norms (Ernande et al. 2004)
g
y(x)
y(x)
Phenotype x
Phenotype y
{(x1,y1),(x2,y2),(x3,y3)}
e1e2
e3
Phenotype x
Phenotype y
g

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
3. QG analysis of phenotypes: Practice

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Estimating components of phenotypic value
∎ The estimation of quantitative genetic parameters is based on similarities between relatives. In theory, as soon as the coefficients of co-ancestry Θ12 and fraternity Δ12
between two individuals 1 and 2 are known, most parameters can be estimated since
COV(z1,z2)= COV(g1,g2) ≈ 2Θ12V(a) + Δ12V(d)
∎ There is a number of classical breeding experiments that can be used for this purpose:Parent-offspring regressionNested mating designFactorial nested mating design

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Parent-offspring regression
∎ The offspring of several controlled crosses between pairs of mates are reared and followed during their life time.
∎ The phenotypic value of the offspring zo can then be regressed against the phenotypic value of one parent zp (mixed linear model):
zoi = α + βopzpi + εi

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Parent-offspring regression
∎ The regression slope gives an estimate of heritability
βop = COV(zo, zp)/V(zp) ≈ 0.5 V(a)/ V(zp) = 0.5 h2
∎ The phenotypic value of the offspring can also be regressed against the mid-parent value when both parents are measured, which increases precision:
zoi = α + βop’0.5 (zmi + zfi) + εi
Then,
βop’ = COV(zo, 0.5 (zm + zf))/V(zp) = 2 COV(zo, zp)/V(zp) = 2 βop

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Nested mating design: half-sibs and full-sibs
∎ Several full-sib and half-sib families are produced according to a nested mating design.
∎ A nested analysis of variance (mixed linear model) can then be used to estimate the parameters
zoijk = μ + mi + f(m)ij + εijk
with m male effect and f(m) female effect nested within males
∎ Trick: an important relationship here is that the covariance between individuals within the same groups equals the variance among groups, e.g. covariance between half-sibs sharing the same father:
COV(zij,zik) = COV(μ + mi + εij, μ + mi + εik )
= COV(mi,mi) + COV(mi, εik) + COV(εij, mi) + COV(εij, εik)
= V(m)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 1512345
Females
Mal
es
Half-sib familyFull-sib families

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Nested mating design: half-sibs and full-sibs
∎ Using the nested model and the previous relationship, one can then estimate the various variance components following:
V(z) = V(m) + V(f) + V(e)
V(m) = COV(Half-Sibs) ≈ 0.25 V(a)
V(f) = COV(Full-Sibs) – COV(Half-Sibs) ≈ 0.25 V(a) + 0.25 V(d)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Factorial nested mating design
∎ In addition to the nested mating design, the macro-environment can be included in the experiment, in order to estimate the components of phenotypic variation related to plasticity or the macro-environmental effect.
∎ In this case, families are divided in several groups reared in different environmental conditions, so as to produce a factorial design between families and environmental conditions. A mixed ANOVA allows to partition the variance in the following way:
zoijkl = μ + mi + f(m)ij + el + m×eil + f(m)×eijl + εijkl
V(z) = V(m) + V(f(m)) + V(e) + V(m×e) + V(f(m)×e) + V(ε)
∎With
V(m) = 0.25 V(a)
V(f(m)) = 0.25 V(a) + 0.25 V(d)
V(m×e) = 0.25 V(a×e)
V(f(m)×e) = 0.25 V(a×e) + 0.25 V(d×e)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Binary traits or threshold characters
Liability scale, lμp
Freq
uenc
y
μr
Freq
uenc
y
xp
xr
i
∎ The mean of any group is expressed as deviation in standard deviation unit from the threshold, e.g. xp = 1.6σp so that μp=-1.6σp and μr = -0.8σr
∎ Assumption: σp = σr = σ, then μr–μp = 0.8σ
∎ The correlation of liability between relatives of any specified sort is given by
βrp = COV(lp,lr)/V(lp)
= (μr-μp)/(μpa-μp)
= (xr-xp)ppa/(2π-0.5exp(-xp2/2))
= 2 Θ12 h2
∎ Based on any classical design, one can then estimate variance components on the liability scale using generalized linear mixed models for binomial data.
ppa = 5%
pra = 20%
Base population
Relatives of affected members of the base population
Liability scale, l
μpa
Threshold

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
4. Case study: using multivariate QG to infer the energetics underlying life history traits
in the Pacific oyster Crassostrea gigas

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Problematic
Constraints
Bivariate reaction norm:
plastic correlation
e
Trait x
Trait y
R(1-u)
RuLife history traits (survival, growth and reproductive effort) result from the allocation of energy between 3 physiological compartments.
R
Ru
Rv
R(1-u-v)maintenance
somatic growth
reproduction
Ernande et al. (2003, 2004) Journal of Evolutionary Biology

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Constraints
g
e
RuGiven these constraints
1. What are the plastic responses in terms of energy allocation to variation in the abundance of trophic resources?
2. Are there different strategies of energy allocation that are genetically determined?
R
Ru
Rv
R(1-u-v)maintenance
somatic growth
reproductionGenetic
correlation
Problematic
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
Trait x
Trait y
R(1-u)
Bivariate reaction norm:
plastic correlation

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
.
.
F1
F2
F3
F4
F5
F1
F2
F3
F4
F5
.
.
Nested breeding design:15 full-sib families distributed across 5 half-sib families.Laboratoire de Génétique et Pathologie, IFREMER
Richenvironment
Poorenvironment
Phytoplankton–
Phytoplankton+++
Site 1 : Controlled conditions
Laboratoire Conchylicole des Pays de Loire, IFREMER
Tidal zoneSalt marshSite 2 :
Fieldconditions
CREMACNRS-IFREMER
Experiment
Ernande et al. (2003, 2004) Journal of Evolutionary Biology

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
.
.
F1
F2
F3
F4
F5
F1
F2
F3
F4
F5
.
.
Richenvironment
Poorenvironment
Phytoplankton–
Phytoplankton+++
Site 1 : Controlled conditions
Laboratoire Conchylicole des Pays de Loire, IFREMER
Tidal zoneSalt marshSite 2 :
Fieldconditions
CREMACNRS-IFREMER
Experiment
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
Genetic
variability
Environmental heterogeneity

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
.
.
F1
F2
F3
F4
F5
F1
F2
F3
F4
F5
.
.
Richenvironment
Poorenvironment
Experiment
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
Genetic
variability
Environmental heterogeneity
During 6 months, every two weeks:- Survival : 120 individuals/ family/treatment;- Growth : 30 individuals/ family/treatment.
Once during summer:- Reproductive effort: 30 individuals/family/treatment.

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
0.00
reproductive effortsu
rviv
al
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
growth
-1.5
-1.5
rep
rod
uct
ive
effo
rt
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
growth
surv
ival
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Poor environmentRich environment
Plasticity of energy allocation
Ernande et al. (2003, 2004) Journal of Evolutionary Biology

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
surv
ival
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
-1.5
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Environnement pauvreEnvironnement riche
survivalP < 0.01
growthP < 0.01R. E.
P = 0.03
Plasticity of energy allocation
∎Plastic shift of energy allocation from survival towards growth and reproduction when the abundance of trophic resources increases.
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
reproductive effort growth
rep
rod
uct
ive
effo
rt
growth
surv
ival

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
surv
ival
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
-1.5
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Environnement pauvreEnvironnement riche
survivalP < 0.01
growthP < 0.01R. E.
P = 0.03
∎Plastic shift of energy allocation from survival towards growth and reproduction when the abundance of trophic resources increases.
One can show theoretically that this strategy maximizes the life reproductive success of individuals in each environment.
Maximizing growth by farming stocks in a rich environment is likely to lead to an increase in mortality.
Plasticity of energy allocation
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
reproductive effort growth
rep
rod
uct
ive
effo
rt
growth
surv
ival

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
-1.5
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Genetic determinism of energy allocation
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
Poor environmentRich environment
reproductive effortsu
rviv
algrowth
rep
rod
uct
ive
effo
rt
growth
surv
ival

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
-1.5
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Environnement pauvreEnvironnement riche
r = 1.12P = 0.01
r = -1.56P = 0.03
r = 0.34P = 0.39
r = 0.33P = 0.41
r = -0.60P = 0.17
∎ The negative genetic correlations observed in the poor environment reveal a trade-off between the 3 traits, but they turn positive in the rich environment some genotypes are "superior" when trophic resources are abundant.
Genetic determinism of energy allocation
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
r = 1.52P = 0.14
reproductive effortsu
rviv
algrowth
rep
rod
uct
ive
effo
rt
growth
surv
ival

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
-1.5
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
-1.0 -0.5 0.0 0.5 1.01.0
0.60
0.00
0.10
0.20
0.30
0.40
0.50
-1.5 -1.0 -0.5 0.0 0.5
Environnement pauvreEnvironnement riche
r = 1.12P = 0.01
r = -1.56P = 0.03
r = 0.34P = 0.39
r = 0.33P = 0.41
r = -0.60P = 0.17
r = 1.52P = 0.14
∎ The negative genetic correlations observed in poor environment reveal a trade-off between the 3 traits, but they turn positive in the rich environment some genotypes are "superior" when trophic resources are abundant.
Despite the fact that some genotypes are superior in a rich environment, genetic variability is maintained because of the negative genetic correlations in poor environments
A selective breeding program aiming at improving survival could result in a decrease in reproductive effort and thus recruitment (and reciprocally)
Genetic determinism of energy allocation
Ernande et al. (2003, 2004) Journal of Evolutionary Biology
reproductive effortsu
rviv
algrowth
rep
rod
uct
ive
effo
rt
growth
surv
ival

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Conceptual interpretation
Trait x
Trait y
(1-u)
u
R
R
u(g1)
u(g2)
u(g3)
u(g4)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
R(g3)
Conceptual interpretation
(1-u)
u
R(g3)
u
R(g1) R(g2) R(g4)
R(g1)
R(g2)
R(g4)
Trait x
Trait y

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Conceptual interpretation
Trait x
Trait y
Trait x
Trait y
∎ The ratio between genetic variability in energy allocation and energy acquisition determines the sign of the genetic correlation observed
In the poor environment, trophic resource abundance was so low that genetic variability in energy acquisition could not be expressed: negative genetic correlations
In the rich environment, genetic variability in energy acquisition was expressed and stronger that genetic variability in energy allocation: positive genetic correlation
acqu
isiti
on
allocation
allocation
acqu
isiti
on

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
5. QG analysis of probabilistic maturation reaction norms

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Objectives
∎ Evaluate/disentangle genetic and environmental variation in maturation tendency of exploited fish stocks
∎ Backward estimation of genetic variance erosion and/or shift of genetic mean in maturation tendency
∎ Forward prediction of future evolution of maturation tendency
forward
backward
t0
t0genotypic values for maturation tendency
freq
uenc
y

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The concept of probabilistic reaction norm
4
5
6
7 30
50
80
40
60
70
0.000.250.500.751.00
Age, a
Size
, s
Probability of maturing

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The concept of probabilistic reaction norm
5 6 7
40
50
60
70
80
430
75%50%25%
Age, a
Size
, s

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Turning PMRNs into classical quantitative traits
40 50 60 70
0.2
0.4
0.6
0.8
1.0
0.030
Size, s
Mat
urat
ion
prob
abili
ty
Age 6
Size, s40 50 60 70
0.2
0.4
0.6
0.8
1.0
0.030
Mat
urat
ion
prob
abili
ty
Individual level
40 50 60 7030
Size at maturation, sm
freq
uenc
y
Population level

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Turning PMRNs into classical quantitative traits
40 50 60 70
0.2
0.4
0.6
0.8
1.0
0.030
Size, s
Mat
urat
ion
prob
abili
ty
Age 6
Size, s40 50 60 70
0.2
0.4
0.6
0.8
1.0
0.030
Mat
urat
ion
prob
abili
ty
Individual level
40 50 60 7030
Size at maturation, sm
freq
uenc
y
Population level
The distribution of age and size at maturation can be inferred from the probabilistic reaction norm
FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
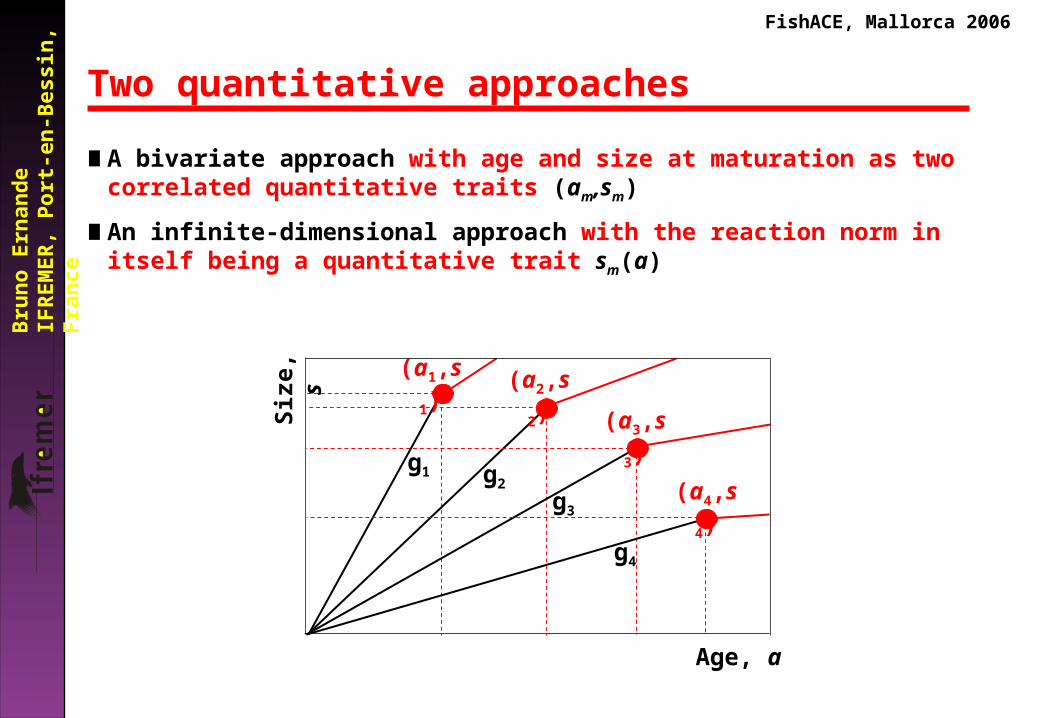
Two quantitative approaches
∎ A bivariate approach with age and size at maturation as two correlated quantitative traits (am,sm)
∎ An infinite-dimensional approach with the reaction norm in itself being a quantitative trait sm(a)
Age, a
Size
, s
g1
(a1,s1)
g2
(a2,s2)
g3
(a3,s3)
g4
(a4,s4)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Two quantitative approaches
∎ A bivariate approach with age and size at maturation as two correlated quantitative traits (am,sm)
∎ An infinite-dimensional approach with the reaction norm in itself being a quantitative trait sm(a)
Age, a
Size
, s

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Two quantitative approaches
∎ A bivariate approach with age and size at maturation as two correlated quantitative traits (am,sm)
∎ An infinite-dimensional approach with the reaction norm in itself being a quantitative trait sm(a)
Age, a
Size
, s sm(a)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Bivariate phenotype : the basic model
m
m ijkl
a
s
g gr ei j k
interactions
g gr g e gr e g gr ei j i k j k i j k
igkl
micro-environment
genotypegrowth
environment beyond growth
Individualphenotype

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Bivariate phenotype : (co)variance components
Zm
genotypegrowth
environment beyond growth
Phenotypicvariance
G GR G E GR E G GR Ek k k
G GR Ek
micro-environment
interactions

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What can we extract using the bivariate approach?
)( 1gam )( 2gam )( 3gam )( 3gsm)( 2gsm)( 1gsm
Distribution of age at maturation conditional on growth
Distribution of size at
maturation conditional on
growth

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What can we extract using the bivariate approach?
∎ The growth-related environmental (co)variance can be estimated as the (co)variance of the mean age and size at maturation conditional to growth
―
∎ The distribution of age and size at maturation can be averaged over growth rates, which gives access to an upward biased estimate of genetic (co)variance,
―
∎ An upward biased estimate of the variance related to the genotype-growth interaction can be obtained by substracting the twoi previous estimates from total phenotypic variance,
―
G Ε G Ek k
GR
G GR GR E G GR Ek k

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Infinite-dimensional phenotype: the basic model
m i k iklikla a a a g es
genotypeenvironment beyond growth
micro-environment
Individualphenotype

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Infinite-dimensional phenotype: variance components
m ka a a a ES G
genotypeenvironment beyond growth
micro-environment
Phenotypicvariance

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What can we extract using the infinite-dimensional approach?
4
5
6
730
50
80
40
60
70
0.000.250.500.751.00
Age, a
Size
, s
Probability of maturing

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What can we extract using the infinite-dimensional approach?
4
5
6
7 30
50
80
40
60
70
Age, a
Size
, s
freq
uenc
y
The whole distribution of age and size at maturation can be inferred from the probabilistic maturation reaction norm

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What can we extract using the infinite-dimensional approach?
∎ Since the effect of growth is already removed in the infinite-dimensional approach, the phenotypic variance of the infinite-dimensional approach is already an upward biased estimate of genetic variance
― m ka a a a S G E

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What are future needs of research?
∎ The coefficient of relatedness between individuals is needed to obtain unbiased estimates of
―
―
∎ Classical quantitative genetics experiments with controlled mating designAdvantage: high statistical powerDisadvantages:
― long experiments (maturity of most commercially exploited fish occurs late in life), ― experimental environmental variation might be not representative of natural
environmental variation
aGG G GR ,

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
What are future needs of research?
∎ Using micro-satellites to determine the coefficient of relatedness between individuals in the wild
Advantage:― representative of natural environmental variation― information available immediately
Disadvantages: ― is it possible?,― low statistical power

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
6. Quantitative genetic modelsof evolutionary dynamics

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Response to selection

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Response to selection seen through the parent-offspring regression
μo*/μp* = βop’
μo* = βop’ μp* = V(a) μp*/ V(zp)
mid-parent
offspring
μo*
μp*

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Principle:
z
Freq
uenc
y
μz(n) μz*(n) μz(n+1)μz(n)
Generation n Generation n+1
z

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Principle:
∎ Equation:
z
Freq
uenc
y
μz(n) μz*(n) μz(n+1)μz(n)
Generation n Generation n+1
Selectiondifferential, s
Heritability, h2
az z z z
z
n n n n2
*2
( 1) ( ) ( ( ) ( ))
z

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Equation:
Selection gradient, β
z zz z
n nn n V a
V z
*( ( ) ( ))( 1) ( ) ( )
( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Equation:
∎ Selection differential s, intensity of selection i and selection gradient β
s = (μz(n)-μz*(n)) = i σz
β = s/ V(z) = i/σz
Selection gradient, β
z zz z
n nn n V a
V z
*( ( ) ( ))( 1) ( ) ( )
( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Response to selection (Falconer, 1960)
∎ Equation:
∎ Selection differential s, intensity of selection i and selection gradient β
s = (μz(n)-μz*(n)) = i σz
β = s/ V(z) = i/σz
∎ Recasting the breeder’s equation
Δμz = h2s = V(a)β = V(a)i/σz
Selection gradient, β
z zz z
n nn n V a
V z
*( ( ) ( ))( 1) ( ) ( )
( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎ The breeder’s equation was derived for the purpose of artificial selection. In case of natural selection, the selection gradient has to be derived from fitness W(z) and the distribution of phenotypes p(z,n)

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎ The breeder’s equation was derived for the purpose of artificial selection. In case of natural selection, the selection gradient has to be derived from fitness W(z) and the distribution of phenotypes p(z,n)
∎Mean phenotype
z n zp z n z( ) ( , )d

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎ The breeder’s equation was derived for the purpose of artificial selection. In case of natural selection, the selection gradient has to be derived from fitness W(z) and the distribution of phenotypes p(z,n)
∎Mean phenotype
∎Mean fitness
z n zp z n z( ) ( , )d
W p z n W z z( , ) ( )d

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎ The breeder’s equation was derived for the purpose of artificial selection. In case of natural selection, the selection gradient has to be derived from fitness W(z) and the distribution of phenotypes p(z,n)
∎Mean phenotype
∎Mean fitness
∎Mean phenotype after natural selection
z n zp z n z( ) ( , )d
W p z n W z z( , ) ( )d
z n zp z n W z zW
* 1( ) ( , ) ( )d

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎ The breeder’s equation was derived for the purpose of artificial selection. In case of natural selection, the selection gradient has to be derived from fitness W(z) and the distribution of phenotypes p(z,n)
∎Mean phenotype
∎Mean fitness
∎Mean phenotype after natural selection
∎ Phenotypes’ distribution
z n zp z n z( ) ( , )d
W p z n W z z( , ) ( )d
z
zz
z np z n
2
22
2( ( ))1( , ) exp( )
22
z n zp z n W z zW
* 1( ) ( , ) ( )d

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎Mean fitness derivative
z z zz z
W p z nW z z W n n
n n2 *( , )
( )d ( / )( ( ) ( ))( ) ( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎Mean fitness derivative
∎ Breeder’s equation under natural selection
Note that fitness is here frequency-independent.
zz z
V a W WV a
W n n
( ) ln( )
( ) ( )
z z zz z
W p z nW z z W n n
n n2 *( , )
( )d ( / )( ( ) ( ))( ) ( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
The breeder’s equation: Natural selection (Lande, 1976)
∎Mean fitness derivative
∎ Breeder’s equation under natural selection
Note that fitness is here frequency-independent.
∎ Extension to the frequency-dependent case
zz z
V a W WV a
W n n
( ) ln( )
( ) ( )
z z zz z
W p z nW z z W n n
n n2 *( , )
( )d ( / )( ( ) ( ))( ) ( )
zz z
V a W W zp z n z
W n n
( ) ( )( ( , ) d )
( ) ( )

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Multivariate evolution: Dynamics (Lande, 1979)
∎ The breeder’s equation can be generalized to multivariate phenotypes
with
W1zμ AZ s Aβ A ln
m
W
z
W
W
z
1
ln
ln ...
ln

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Multivariate evolution: Dynamics (Lande, 1979)
∎ The breeder’s equation can be generalized to multivariate phenotypes
with
∎ In other words, the evolution of a particular character i is given by
with Aij the additive genetic covariance between trait i and j
m
W
z
W
W
z
1
ln
ln ...
ln
i j
m
z ij zj i
A Wln /
W1zμ AZ s Aβ A ln

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Multivariate evolution: Genetic constraints (Lande, 1979)
∎ Evolutionary dynamics may come to a halt becauseThe selection gradient vanishes
for all zi
The additive genetic co-variance matrix is singular, so that
whatever the selection gradient
WA ln 0
izWln / 0
Wln

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Multivariate evolution: Genetic constraints (Lande, 1979)
∎ Evolutionary dynamics may come to a halt becauseThe selection gradient vanishes
for all zi
The additive genetic co-variance matrix is singular, so that
whatever the selection gradient
∎ The rate and direction of evolution determined by the selection gradient are modified by the genetic co-variance matrix. The rate of evolution of a given trait is a sum of
the response to direct selection on that trait and the correlated responses to selection on genetically correlated traits
WA ln 0
izWln / 0
Wln

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Multivariate evolution: Genetic constraints (Lande, 1979)
∎ Evolutionary dynamics may come to a halt becauseThe selection gradient vanishes
for all zi
The additive genetic co-variance matrix is singular, so that
whatever the selection gradient
∎ The rate and direction of evolution determined by the selection gradient are modified by the genetic co-variance matrix. The rate of evolution of a given trait is a sum of
the response to direct selection on that trait and the correlated responses to selection on genetically correlated traits
∎ Genetic constraintsLack of genetic variance: The rate of response to direct selection is scaled by the additive genetic
varianceTrade-offs: the rate and direction of responses to correlated selection depends on additive
genetic covariance with other traits, so that the direction may be opposite to the actual force of selection thus highlighting trade-offs between traits
WA ln 0
izWln / 0
Wln

FishACE, Mallorca 2006 B
runo
Ern
ande
IFR
EMER
, Por
t-en-
Bes
sin,
Fra
nce
Advantage and drawbacks
∎ Quantitative genetic evolutionary dynamicsallow studying both genetic and phenotypic evolution,
predict both evolutionary transient states and equilibrium
accounts for genetic constraints such as the lack of variance or trade-offs
∎ Drawbacksno detailed ecological considerations or population dynamics: fitness and selection gradient are
not extracted from the ecological setting
― not geared toward identifying explicit selective pressures,
― no complex population dynamics and frequency-dependent selection
constant additive genetic variance-covariance matrices: limited to short term evolution or “weak” selection, so that the mutation-selection balance can maintain additive genetic (co-)variance (Lande 976, 1979).