biosynthesis indoleacetic from trvntsnhan-'4c in cell-free ... · plant physiol. (1967) 42,...
TRANSCRIPT

Plant Physiol. (1967) 42, 1787-1796
Biosynthesis of Indoleacetic Acid from Trvntsnhan-'4Cin Cell-free Extracts of Pea Shoot Tips'
Thomas C. Moore and Coralie A. ShanerDepartment of Botany and Plant Pathology, Oregon State University, Corvallis, Oregon 97331
Received August 14. 1967.
Summary. A 2-step, 1-d:,mens-ional thn-layer chromatograph-c proceduire fnrisolating indoleacetic ac:d (IAA) was developed and util:zed in inve,t at:ons of thebiosynthesis of JAA from tryl)to,)han-_4C in cell-fre'- extracts of ipea (Pi.sun sativitmnL.) shoot tips. Identfcat;on of a 14C-prodnict as IAA was bv (a) co-chromatograT)hvof atitheiiitc IAA and '4C-pro(duict on th.:n-layer chromatography, and (b) gas-liqu-;dand thin-laver chromategraphy of authentic and presumptive IAA methyl esters.Dalysis of enzyme extracts and addt;on of a-keto-li1tqr`c ac d anl pvr:doxal phos-phate to reactioni m;xtl'res re,ulited in ap)prox:mately 2- to 3-f-ld increases in nety;elds of IAA over yields in non-d:alyzed react-on m:xtres wW:ch d:d not conta;nad,d:tives esqent:al to a translaminace reaction of trvptophan. Add`t:on of thiam:nepyro0hoonbh,,te to reacton m;xtl,req fiurther enhanced npt b;h,vnth-s-s of I'A. It isconclhlded that the format;on of indolepyrtiv:c ac;d and its silhbenlent decarboxvlat:onprohablv are seeventtal react:onis in the maior pathway of IAA bocvnfhsis fromtrvntonhan in cell-free extracts of Pisutnt short t:pS. ComDar;-on of max:mtlm n't-IAA biosynthes;s in extracts of shoot t:ps of etiolated anid leht-grown dwvarf andtall pea !dluui;nzs reveal-id. aln order, on a uin:t T)rofe:n N ba:, of: 1`lo-4-rowntall > Pght-zrown dwarf > etiolated tall _ etolated dwarf. It is conclrxded thatthe d:fferent rates of stpm elonzation among et;el"ted and l:ght-pr-wrn dwarf andtal!l pea seedl:ngs are correlated, in general, with differences in net TAA biosynthesisaild setisiitivity of the tissules to IAA.
Ex'olan.utions for the (Ifferer(nces in stemi elomioa-tioIl betweeil dwarf and(l tall varne;es of a sneciese.g. Pisumin sativ'mi, and betweon et;olatod atndtlc-ht-grown l)lants of the sam- var;ety have he-nsought repeatedly throu"gh inve-t;zat:ons of pols):bVe(orrelat;ve difference,s in endogen-onls aulx;n an1dib,berell;n relationships. The maiori'ty of these
investigations have been either: (a) studies ofcomparat:ve resp)onses of intact see(linzs or ex-cised stem sectioins to exogenotus hormones: or(b) extraction and bWoassay of enidozeniouis hor-mones. Another kin(d of exoer:mmntal avnroachwhich is tusefult in attempts to elu1cidate comparativeg,rowth hormone relationships is invest:gation ofhormone bWosvntheses tsi;ng cell-free enzymne prep-atrations. Tn 1958, Gor(lon (12) f:rqt 'describe-d anenzyme system from shoot ti'ps of Phascoluts autreusseedlings w,hich catalvzed the coniversion of trvnto-phan to indole-3-acetic ac&d (IAA). Wghtman;ind Cohen (39) recently described in detail theintermediary steps in the enzymic conlversion oftrvptophan to TA.A in cell-free preparation;s from
1 This investigation was supported in part by GrauitGBI-4613 from the National Science Founidation.
mntg be-an (Ph1 rscolls (1a1 remis) seedlPl;ngs. So)mepropert e- of an TAA-syntheq;z;ng syvtem isol-te-dfrom ani:ces of Avona. coleoptiles and otllel- W'antmrn-er:akc were de-cr'be,l by Lant.can and Mui:r(17). WhIle cell-free bWos-wnthesis of ghbberell nshas not vet he-n reportled, sutbstantial progresstowaqrrl this objective has been made recenitly(1, 14, 28).
The present report describes inno'vationi illthmn-layer chro-matographc procedulres for isolatinigIAA from plant preparations and comparative net3 ields of IAA-14C formed from try'ptophan-_4C incell-free extracts of shoot t ps of etiolated andlight-grown pea seedlings.
Materials and Methods
Develop/ment of a 2-step Thin-layer Chronwa-tographic Procedutre for Isolating IAA. None ofthe reported methods for isolating IAA from plantpreparations was readily adaptable to the investiga-tions here described. Hence considerable attentioinwas devoted to the development of a suitable thin-layer chromatographic (TLC) procedure.
One problem was to find a stuitable meanis ofseparating indolepyruivic acid (IPyA) from IAA
1787 www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
and also of prevent:ing noin-enizymic conversion ofIPyA to IAA during the separation. Samples of5 or 10 kg each of these 2 in(lole compouinds wereapplied separately on 5 X 20 cm glass plates coatedwith 250 M of silica gel G. Development was con-duicted in closed glass cylinders (ca. 6 X 23 cm),with the solvent front being allowed ito adlvance10 cm above the origin. Detectioin of non-radio-acitive indoles wvas achieved by meains of theirfluiorescence in ultraviolet light (ca. 3660 A) orby s,praying the chromatograms with Elhrlich's re-agent, which was prepared and(I uise(l as (lescril)edby Stahl (33).
Sam,ples of 0.5 Kc each of IAA-14C were appliedto TLC plates and (leveloped in vari/ouis solvent sys-tems singly ancd also successively in combinationsof 2 solvent systems. The 2-ste,p TLC techniquewas conduicted as follows: a) after initial dlevelop-men;t in 1 solvent system, a band of silica gel wasremove(l from the origin-end of the plate to within1 cm of the center of the positioin of the compoundof interest; an(d then, b) taking the ceintelr of thecompound as a seconcd origin, tihe chromatogramwas dlevelope(d in a second solvent, system, movingin the same dlimensioni as the first, \N ith the solventfron,t again advancing 10 cm fromii the orig in.Five ,ug of auithentic iunilabeleld IAA Nwere routinelyap,plie(l oin top of, or to 1 side of, the radloacti'VeIAA oin each plate, and the inlabieled IAA -wasco-chronmatographed wxith the ra(dioactive sal)ple.\VNrhen chro,matogranms were (levelope(l in 2 solventsvstemlls, anI a(dlitional 5 /sg of uinlabele( IAA wereapplied to 1 si(le of the secon(l origin.
Followin-g development, the chromnatogramn werescannlle(l oIn a ITackard Radiochromatogram Scanner,Mo(lel 7201, to locate the posit:Ons of radioactivesuibstalnces, and(I the mnlabeled IAA was (letectedl byits fluorescence in ultraviolet light. habi( ofsilica gel 2.0 cIm widle and centered on the positioof IAA-14C was remov-ed from each of severalchromatograms after (development in 1 or 2 solventSy'stelims, an(d the radlioact:vit\v N'xas miieasuiired b\-liqulld( scintillation coulntinlg. Saples of .iAalso w\ere apl)pliedl to silica gel plates, remove(l in2-cm banids of gel, andl couintel oii a I 'ackard 'I'ri-carl) -icquiid Scinitillation Spectrometer, Model314EX, to pirovide standards for comparison ofyield after chromatography. T'he fluid usedl forliqi1i,d scintillation coulntillng 'was Bra\ys solultionlwhich consists of 60 g of naphthalene, 200 nig of1,4-bis-2'- (5'- )henyloxcazolyl)1)bein7ene, 4 - of 2,5-diphenylloxazo1e, 100 ml of imiethainol, ainld 20 ml ofethylene glycol, made uip to 1 liter volume withpara-(lioxane. Each radioactive saml)le x\ as (lis-perse(h in 10 ml of scintillation fluiid.
Cailtiure of Seedlings. See(ds of Dwarf Tele-phone an(d Tall Telephone ctltivars of Pisi inisatizvon ('W. Atlee Buirpee Company, Riverside,California) were surface-disinfected vith a 0.5 %soluition of NaOCl, rinsed 3 times with distilledwater, and planted in plastic containers filled w\vith
vermicuilite. The root medium was irrigated awneeded with a complete mineral nutrient solution.Environlmental colndlitioins in the growth chamberswere: 16-hotir photoperiod at 20 ± 10 and inci-dent intenisity- of approximately 900 to 1000 ft-c,and an 8 houir (lark p)eriod at 16 +- 1°. Etiolatedseedlings experienced( the same temperature regimeas light-grow-%NIn see(lings. OIn the twelfth (lay afterplanting, shoot tips were excised immediately abovethe fotirth nodle of etiolated seedlings and abovethe fifth nlode of light-groxxn see(lliigs. All nleces-sary handling of etiolatedl seedlings xxas conductedin dim green light. M\easureimients of to,tal shootlength were nia(lde oI 20 to 25 seedlings of eachgroup at the time shoot tips xvcre excisedl.
Prepa ration of Cell-frece Enz3:ymie E.I-tricts andReaction Conzditions. Elach sanmple of shoot tips(routinely 80-100 tips Wveighling apl)proximatelN5-10 g) wxas hoimioge-nized immle(liately after ex-cision in 4 voluimes (4 ml:1 g fr- x\t) of IKH.-,P1,-Na.,HPO, buffer (0.1 :v, pH 7.4) containin1lg 0.3 Msulcrose and 200 eg/ml eachi of penicillin G andstreptomycin sfll fate. Holmogenization was coIn-(Ieicted iisin,g a Thomas teflon-to-glass homogenizerin a cold room (approxiniiately 50), aild the homog-cnate x\-as kepl in ani ice bath throughout thepreparative proce(dllre. 'T'he cl-iide homogeniate \Nascenitr filged at 10,000 X g for 20 millutes at 40,and the resultilln slihperlatailt wxas ilse( as theenzyme souirce. Dialv sis of enzyme extracts wasagainst 0.1 I pl)hosphate buffer for 22 to 24 houirsat approx:mately 2 to 5 x\ ith a total of 3 chalngesof extel-rnal buiffer. Duplicate determillationis ofproteininitrogenl coilteiit xvere ma(le on 1-ml ali-(1iiots of eachi dialyzed enizymiie extract bx a mIticro-Kjeldahl procedlure (16).
Reaction mixtures each contained 3.0 ml ofeizvme extract, boiled enzynie extract, or btifferp)hils additives only; an(l 3.0 nil of 0.1 lph)osphatebuiffer (pH 7.4) containinig ,6 lc of DLtryptophanl-2-14C ( TTP-I 4C; ring-labeled; specific activity21 mc/mmole), 30 juiioles of unlabeled FTTP, anld,w\here in(d:cate(l, 30 /iimolcs of a-ketoghutaric acidand 0.6 tksmiole of l)yridoxal phosphate. Experinmentswere also con(ltlcted in x\ hich the effects of addinga-ketoglutaric aci(d anId( pyridoxal phosp)hiate xvereinvestigated independeintl\. In somiie a(h(ditional ex-1)Criment-, Inot rel)orte(l b re in (Icetal, 0.6 LImole of
th.ianline piyrol)hospl)hate and 0.6 ,umole of nlico-tinani:de aIdellillc (Iililelcotide xvcre adlded to eaclhof certain reactioln miiixtuires. Reactioni mixtulresx\'ere incuibaited for 120 minuiites at 300 in larknlessin 12 nil conical, glass-stoppered tuhies. Reactionswere stopped bv addlillfg a predheterminedh amount ofconcentrated I1TPOL a(ldequlate to low\er the pHto 3.0.
Isolaitionz ad(1 AIcMasureinent of Presumptivef.44 lt4C. UInlabeled IAA (1 ,Ug/ml of reactionmixture) \vas added to each ttubc immediately afterincubation. Then each reactionl mixttire \vas ex-tracted twvice with 6 ml of redistilled methyvene
1788
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

MOORE AND SHANER-CELL-FREE BIOSYNTHESIS OF IAA
chloride (MVJeCl,). The combined extracts fromeach reaction mixture were evaporated to drynessunder red-uced 'pressure in a ro,tary-film evaporatorat 300. Each residue was dissolved in 2.0 ml of95 % ethanol, and 500-,A a,liquots of each extractwere applied to the origins of 5 X 20 cm TLCplates coated with 250 ju of silica gel G. Ten uxgof unlabeled IAA were applied at the same positionas extract on each plate, and the chromatogramswere developed, according to the procedure de-scribed previotusly, suiccessively in chloroform-96 %acetic acid (95:5, v/v) (CHAc) and methyl ace-tate-isopropanol-28 % ammonium hydroxide (45:35:20, v/v/v) (MIA). The position of carrier IAAwas determined after each stage of development bvexamination of the plates in tilitraviolet light.Chromatograms were scanned to detect radioactivityco,inciding in position to carrier IAA. Then a bandof silica gel 2 cm wide and centered on the positionof carrier IAA was removed from each plate, addledto 10 ml of Bray's solution and counted1 by theliquid scintillation technique.
Methyl Esterification of 14C-Product (anid Aut-the utic IAA. Two samples of 14C-product, eachcontaining several .thouisand cpm of radioactivIty,were isolated from chromatograms of ac di1c ex-tract of complete reaction mixtures. Silica gelcontaining the 14C-product was elutted twice withMeCI2 for 1 preparation an-d with 95 % ethanolfor the second preparation, since 95 % ethanol wasfotund to eltute the product more efficielntly thanMeCl2. The ellting solvent contained suffic&entunlabeled IAA to allow for a concentratioln ofapproximately 1 ug/ul in 'the final preparat on.T'he eluates were evaporated to dryness unlder astream of N, while holding the ttube in an icebath. The residue was re-dissolved in a nn:rmnalvoluime of acetone, with the final preparation con-tamining approximately 1 ug/,ul of unlal)eledl 1J1kAand approximately 200 cpm/ul of radioactivitypresent in 14C-producit. Sample's of authentic IAA-14C were putrified by TLC and finally dissolvedalso in acetone.
To acetone solutions of 14C-product, authentitcIAA-14C and autihentic unlabeled IAA were a(ldedaliquots of diazomelthane solution (kindly suppliedby Dr. Te May Ching). Amounts ,of diazomethanesufficient to cause the solutions 'to remaini yellowafter 1 to 2 hours were added to insure anl excessof diazomethane. When diazomethylation was pre-sumed to be complete, the remaining diazomethanewas elim,inated by blowing a stream of N2 over thesurface of each solution. When further conden-sation was desired, the soluitions were condensed byevaporation un'der N,.
Gas Chromatography of Methyl Esters. Ali-quots of 10 jLl of each methyl ester so-luition werechromatographed on an F and M Gas Chroma-tograph equipped with dual hydrogen flame detec-tors, using a procedure modified from those de-scribed by Dedio and Zalik (6) and Powell (26).
Samples were injected onto a coiled metal column(6 feet long X one-eighth inch in diameter) packedwith 10 % SE-52 on Gas Chromosorb Q (80-100mesh) (Applied Science Laboratories) and devel-*o,ped at a column temperature of 2000 with anargon flow rate of 60 ml/minute. Injection portand detector temperatures were 2500 and 2000,respectively.
Using a micrometer valve to regulate the pro-portions of carrier gas going to the detectors andeffluient, a flow rate of 40 ml/minute of argon wasshunted to an efffluent, where samples were .collectedin liquiid scintillation vials at 1-minute intervals.Each vial contained 10 ml of toluene in which weredissolved 30 mg of para-terphenyl and 0.3 mg of1 ,4-bis-2'- ( 5'-phenyloxazolyl) -benzene. A problemwas encotuntered in trapping the methyl ester effi-ciently. Using amouints of 25 to 50 ug of uinlabeledIAA methyl ester, it was apparent that the esterwas not being efficiently absorbed in the scintilla-tion solution. This problem was partially overcomeby connecting 3 serum-capped vials in series, bultstill only 25 % or less of a measured alinuot ofauthentic radioactive ester couild be trapped.
Thin-layer Chronm(atogralphy of Authe ttic (in(lPresumiptive IAAA-14C Methyl Esters. In fu r'therefforts to verify the identification of 14C-produictas IAA-14C, samples of authentic and presuimptiveIAA-1lC and unlabeled IAA methyl esters werechromatographed on thin-layer plates in severalsolvent systems, including CHAc (95 :5, v/v),hexane-diethyl ether (1:4, v/v) an'd 100 % chloro-form.
Reagents. The critical reagents uised and thesouirces from which they were obtained are: DL-tryptophan-2-14C (labeled in indole ring; sp act -21mc/mmole, Schwarz BioResearch, Orangebuirg, NewYork); IAA-2-14C (side-chain labeled; sp act 2mc/mmole, New England Ntuclear, Boston); IAA(Mann Research Laboratories, New York); DL-trvpto,phan an(d indole-3-pyruvic ac'id (Sigma Chem-ical Company, St. Louis) ; pyridoxal phosphatemonohydrate, a-ketoglutaric acid, thiamine pyro-phosphate chloride and nicotinamide adenine dinii-cleotide (CalBiochem, Los Angeles). All organ:csolvents uised in extraction an-d chromatographicprocedures were redistilled.
Results
Thin-layer Chromnatography of Au1ltenttic IAAand IPyA. Of several solvent systems tested, the3 which proved most useful were (a) methyl ace-tate-isopropanol-28 % ammoniuim hydroxide (45:35:20) (MITA), (b) chloroform-96 % acetic acid(95:5) (CHAc), and (c) isopro'panol-28 % ammo-nium hydroxide-water (8:1:1) (IAW). The aver-age RF vallues of IAA in MIA, CHAc and IAWwere 0.48, 0.42 and 0.55, respectively. The RFvaluies of IAA in the 2 basic solvent systems variedwith significant variations in the amount of ammo-
1789
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
1a actually p)resent in the chromatography Qx- inder.I Py.`\ (dd nioit in grate from the or- gnIl in CII Ac,l)tit wvas very IIIustable tlle 2 l)basc solvent s >teIis.I1y means of spraying Nwith Ehrlich's reagent andeXamnat 0on tilndeCl nltrav'o,et ght, at least S spo,tswere fotind on the IPyA chromatograms developedin e thler basic solvent sysenl. I AA wvas solmewhatInIIusIable tindler alIl cli romatograph"Ib C()oiil toiIIstestedl, andl data based Oll recovery of 1 A A-'t, to
be presented later, ind ate (jiantitatively thd ilegr-eeof instablity for each set of coml t'oils. A fter
levelkpment in CHAc, the position of FAA iicilb
wvas d-cernible without spray recigeIIt or Illtilavioletlight by a pinuk color representing an (i 'xdatlonproducit of IAA. If the gel were removed fr-om
the p'ate lI)elow\ the pink spot and the plate re-
chro,matographed a-a<n in CHlAc oir in I of thebas:c solvent systems, thl remaining 1AA separate(lfroni the position of the pink ox dation proliict.[n b,)th basic solvent systems l)rodiicts formed b)vthe breakdown of IAA m grated to tlh solventfront. wvhereas if re-chromatc-ralphed ill Cl lAc,
the maior breakdown pro(ltict or proliiuts remained
att or near the origin.Indlolepyruvic ac(l generally is consi(lered to be
aln intermed'ate proldict in the formation of I\Afromii tryptophan (5, 12, 13). If lresent in thetissule or cell-free enzyimie preparation, IPvA is
exlrcactedl -<tmultaneonisly with IAA (2, 5). Becatiseof the pronotinced instab lity of IPyA, on chromna-tography in many solvent systems it is cos e rtedto a niuimber of products, oine of which rel)orte(llv isIAA ( 13). Henice, in or(ler to dleterminiie vicll ofenzym:cally formecd IAA, it is iiecessary to lprevenltnon-enzym:c conversion of IPy\A to iAA (dtinrmigChromatography. TlI the auithors' cXl)e ricnce. it
possible to accomplish this objective by: (a) firstchromatograph'lig an extract or mixttire of ati-thentlc IJPyA and IlAA in CHAc; and(I (b) removingthe silica gel 1.0 cimi below the ceinter of the IAAspot ani(l chronimaographiing the residtual I.\.\ in a
seconid solvent system.Chroaotogrlo ll oof . otlci tic 1T1.1 (. 'lable
I shows the qluant:tative recover-ies of _I\C14(after single chromatography in CIlAc and a ftcr2-stag,e chromatcgraphy f;rst in CIJAc and secouldl)in each of 2 basic antid( 1 atcidlic (CHIAC ) solventsystems. Approxmniately 30 % of each slample of
IAA-_ 'C was lost throliolgh oxidation as a resiult ofbeinig chromatograplhed only oiice in ClIAc. ol
lowing a secotl( chromatography, only 37 to °
of the org:inal sampleis remained as [.\AA- C.
\Vhile tlhese amounts of loss are h:-gl, thle,, autlhors'experience is that the 2-stage chromaltography ofIAA- 1C vieltls qtiamititativelv reproducible i-esuilts.'lIe data for 2 selparate experiments iii table 1 are
ill goo(l agreement, except for a sigilificanit liis-p)arity' in the recovery after seconId chromato-raplhxin IAW. Resiults obtained mIIore recentlv reveal
that reprodtic We recoveries of 45 to 50% also areattainable tising IAN\V as the second(I solvenit system.
Str p chart tracings of radiochromatograms, offAA_'4C aft er the second(I develol)ment in the 2-stage'I'LC p)rocedtire revealed 2 major lpcaks of raldio-activity oni each chroniatogram. The large lp-akon each chromatograin coincided in posit:on withaiuthentic tiinlabele(d IAA, and the smaller l)eakeither remamned at the orignin (CHAc) or moved'to the solveint froiit (A\IA\ and IAW).
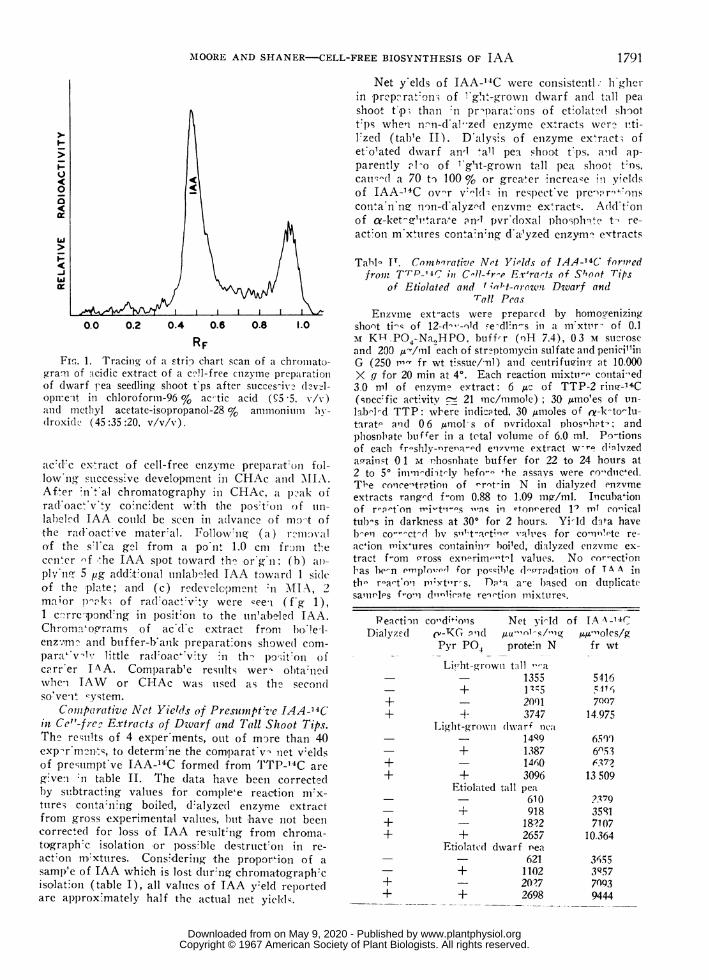
Isol(itiofl of Presumnptize IAA-' 'C fronti Cell-fre)Reaction M.iXtures. Presente(l in f.giire 1 is a
tracing(of a strip) chart scaii of a clhromatogrami (f
Table I. Recov'ery of .-1 Ithen1tic I.1J.,-'4C Aftcr C/iroollato.yraphy in I amluiois .Solvent SystenilsAni aliquot of 0.5 u.c of IAA-2-14C was applied at the first ori,gin of each l)late; all samples were (lilute(l to
20 % of in'tial concentration before they were counited. Ahbreviations are as follows: chloroform-96 % acetic acid(95 :5, v/v) (CHAc) methyl acetate-isopropanol-28 % ammllioniumlil hydroxide (45 :35 :20, v/v/v) (MIIA), isopro-patiol-28 % amnionium1 hydroxide-water (8:1 :1. v-/v/v) ( IA'V ). Data are base(d onl duplicate samples.
Treatment ofIAA- I'IC cpili
.Application to silica gel only2. Chromiatography in first
solvent svsteiii (CHA-Ac)3. Chroniatography iii secodI
solvenit systenill
(a) MNIIA(b) IA\W(c) CHA\c
1. Application to silica gel only2. Chromatographx in first
solvent systeni (CHAc')3. Chromiatograplih il secoll (I
solvent system:a) MIIA
(b) IA\\(c) CH-Ac
Ex )t 1
96,700
44,44653,18137,699
xp)t 2
116,568
78.452
53.20948,47243,144
1.-\.\-14C recovered
YGJ/c of original samiple
170.0
70).6
46.055.039.0
100.0
68.7
46.441.937.3
1790
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.
MOORE AND SHANER-CELL-FREE BIOSYNTHESIS OF IAA
I-.
uoc0C
w
-JtiJ
CZ~ ~
gra' ofof (dwaiopire itand nilE(d roxi d-
a c'('d. c (
lowr ngAfterrad.loaclabeledthe ra]of thecenerplynll-.of the
Net y'elds of IAA-"C were consistentL h'gbiin prep?rat:on; of -'gt-grown dwarf and( tall peashoot t,p. than :n prl)parat:ons of et:olciat 'd slott p-s when nwn-d al-zed enzyme extracts ver? vti-l.zed (tab'e II). D'alysis of enzyme ex ract; ofet'o'ated dxvarf an, 'all pet shoot tCps, a'id ap-parently al-o of 1 g'it-grown tall pea shvoot t:os,cailq'(] a 70 to 100 % or grea,ler nicrea,e in yieldsof IAA-'4C ov'-r v:eHz in respect've pre aro+.nscontVanono non-d'alyznd enzvme ex&ract,. A(1d't:onof a-ket'&rvara,e an-1 pvrd(loxal phoisphh..te t- re-acton m xtires conta:nng d'a,'yzed enzym- extracts
Tabl- T Cornhorative Net Yields of IAA-'4C formedfromz' TTP_14r itl C11--r- E.rrarfs of S'ioot Tips
of Etiolated and ro;7t-orown Dwarf andToall Peas
,I,v I-
I I It | lX, Enizvme ext-acts were preparc(l b)y homov'enizing
0.0 0.2 0.4 0.6 0.8 1.0 shoot tic of 12-d'- ol d ce dlins iii a mixttur of 0.1RF Nf KT .PO4-Na,HPO, buff-r (nH 7.4), 03 M sucrose
and 200 Au/mnl each of str?ptomycin sulfate and penicillin1. Tracing of a stri) chart scan of a chroniato- G (250 tn" fr wt tissue/mIl) and centrifuvin'e at 10.000
f Icidic extract of a c-l-free cnzyme preparation X g for 20 min at 40. Each reaction mixturlo contai-edrf pea seedlinig slhoot t ps after succes,i (l2v2l- 3.0 mnl of enzym- extract: 6 ,uO of TTP-2 rinc-1"Cin chloroform-96% ac-tic acid (C5-5, v/) (snec:fic activityv 21 mc/mmole); 30 gnmo'es of tin-
etlhyl acetate-isopropanol-28 % amniiionitulni 'hY\ lab]Wd TTP: wlIere indicated, 30 /tmoles of rX-kto'-iu-(45 :35 :20, v/v/v ). taratp and 0 6 miol- s of nvridoxal Thos-hat-; and
phosonlate buffer in a total volume of 6.0 ml. Po-tionsof each fr-shly-nrena--e( en7zvne extract w-r- (d;ilvzed
extract of cell-free enzyme preparatton fol- ai ai'vt 0 1 M rhosnhate buffer for 22 to 24 hours atsticcessive development in CHAc and Ml A: 2 to 50 invmdi'trly hefo he assays were coIdAced.. . Tie conce-tration of *rot-in N in dialyzed enzvmental chlromatographly in CHAc, a p 'ak; of extracts raing-d fom 0.88 to 1.09 mi/ml. Incubasionvty co:nc:dent w:th the pos ton of 1in- of r-aYton mi-i-s -.1s ii 0ton'rered 1' mnl co-icalIAA cotiul be seen in advance of ma) t of tub-s in darkness at 300 for 2 hours. Yi-ld dita have
Iloactfve mater al. Follow lg (a ) rnim al b-en co-et-d bv s"1' taet;- valt'es for com' )te re-sl cal gel from al pOntlF 1.0 cm frmi t -ieac*ion rnix'ures containin- hoiled, di-lyzed enzvtne ex-
-f -he IAA spot toward th' or'g-n; (b) ai,- tract f-om cross exn'irim,-tl values. No cor-ect;on5 pg add.t onal unlab-led IAA toward 1 si(le las be-n enpllo',-l for po-zible d-qrndition of TA A inpla-wte; anid (c) r"develeic;ment n M\11 2, thb r-aeton Prixt11r s. nTai a-e based on duplicate.. . . . samirles f"ooi dItnlirgte reirtion mixturesmanor p '!i ot ra(n oac: v ty were ceeei (t g 1),
1 c"rre-pondong in poSIt0on to the un'ab"led IAA.ChromnVoe'rams of ac d c extract from bo te l-enzz,m, and buiffer-bank preparat.ons showe(d com-para' v-i little radl oac; v.ty in th- po-,itoon ofcarr er IAA. ComparabWe resiults wer- olbta:nedvhien JAW or CHAc was uised as the second(lso'vet1c ystem.
Comiparative Net Yields of Presumpt ve IAA-'4Cin Ce"'-free Extracts of Dwarf and Tall Shoot Tips.Thb results of 4 exper ments, ouit of more than 40exp-r mnntc, to determne the comparat'v- net v:eldsof prestimpt've IAA-14"C formed from 'T'TP-' 4C aregive:I n table II. The data have b-een correctedby subtracting valuies for complee reaction m-x-ttires conta1n:ng boiled, d:alyzed enizyime extract-from gross experimental values, but have niot beencorrected for loss of IAA result:nig from chroma-tograph c isolation or p)oss'l)le destructon in re-act!on ml xttlres. Considering the proportion of asamlp'e of IAA which is lost dur ng chrornatogra<ph:cisolation (table I), all valties of IAA y:eld rel)orteclare approx:mately half the actuial net yieldl.
PReactihni codi+;olisDialyzedl (V-KG Pand
Pyr P04
+
+
Net yibld of IA .014Cprotei N/Sng frwtoles/gProtein N fr wt
Li"-ht-grown t'll --a1355
+ 1C- 209 1± 3747
Light-growno (ldwarf oea14R9
+ 1387- 14604 3096
Etiol,ated tall pea610
+ 918-18?2
+ 2657Etiolatd(1 dWarf Pea
621+ 1102- 2v037+ 2698
54165.116)7007
14.975
65'Y)
6<37313509
?.37935R17107
10.364
36553Q5770939444
1791
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

ILANT PHYSIOLOGY
further increased net y ields of IAA-''C. \NVheieither the a-keto acid or pyridoxal phosphate was
added independently o,f the other, the net yield ofIAA was substantially less than in the preseice ofthe 2 compotinds combined. Py-ridoxal phosphateandl a-ketogltitarate had nio significanit effect on
yield of IAA-14,C when added to non-dialyzedenzyme extracts. The apparent order of maximtim
net yield of IAA-'4C in the 4 t%ypes of preparationswas: light-grown tall > light-grown dwarf > etio
lated tall - etiolated dxx arf. For comparison, itcan be noted that the mean shoot heights for the4 kinds of seedlinigs at the time shoot tips were
excised wvere : etiolated tall, 23.1 etiolate(d nlwNarf,15.4; light-grown tall, 11.7; and light-grown dwarf,4.8 cm.
AMIorphologically equivalent samples of shoots
were taken for dwarf and tall seedl,ings in eachexperiment; that is, shoot tips ere excised iimime-(liatelv above the fifth node from all light-grownplanits and above the fouirth no,de from all etiolatedlplants. However, the relative amounts of leaf andstem tissue in the different kinds of shoot tips vary
qtiite significantly. A (letailedl analysis revealedthat: (a) light-grown tall shoot tips contaiin ap-
proximately twice the fresh weight of stem tisstuesa!s dwarf shoot tips, ancd that the dwarf and talltips have the same amotunt of leaf tissule; aiid((b) etiolated dwarf and tall shoot segments are
closely comparable in amotints of leaf and stemtissue, but the ratio o,f leaf :stem is mtuch lowver foretiolated types than for respective light-growvn types.The mean fresh weights of shoot tips were not
greatly variable among the 4 kinds of seedlings,ranging from 67 !mg for etio-lated -tall seedlings to
88 mg for etiolated dwarf planits. HeInce, if it isasstumedl that aulxin prodtiction is most intense inthe terminal btid and that leaf tissties are higherin nitrogen content than elongated stem tissties,the acttial net yields of IAA per tinit of N in themost intensely active auixin-prodticiing shoot apicaltissules may possibly be more nearly: light-growntall > etiolated tall > etiolated dlwarf light-grown (lxx'arf. Tn the opinion of the atithors, com-
parisotns on a fresh-weight basis have less meaningin a physiological sense becauise of the differencesin relative amnoulnits of stemii ai1d leaf tisstues amiongthe 4 kinds of samples.
Perceintages of stibstrate radioactixvitvr which were
extractable in acidic (pH 3.0), neutral (pH 7.4)andcl basic (pH 9.4) fractioins w,ere determniined in
3 experimenlts in which non-dialyzed enizyme ex-
tract was uised and there were no add;tions ofa-ketogluttarate or pyridoxal phosphate. The per-
centages wNere 0.06 to 0.10, 0.04 to 0.12, annl 0.02for the acid, neuitral and basic fractions, respec-
tively. The order of extractioin was varied fromacidic to neuitral to basic and in reverse order, andit was found that the order of extraction (lid notappreciably alter the percentage of initial radio-activity extracted at each pH. Nor w,-as there any
evidence on thin-laycr chromatograams for anymeasturable 'ITTP-14C being extracted froml reactionmixtures. Hence it is beliexed that the actual.percentage of substrate converted t)iacid extract-able compounds tinder these conditions wxas ap-proximately 0.1 %. 't'he maximumii percentag-es ofsubstrate radioactiv\ity which w\,ere extractable inacidic, netutral and basic fractions of reaction mix-tuires contain,ing(dialy'-ed enzytme extract, a-Xketo-glIutarate, px ridoxal phosphate and thiaiminie pyro-phosp)hate wvere 0.23, 0.50, and 0.13, respectively.TI'he rate of convxersioni of 'TI"TP-1 C to 1AA-\ cwxas nearly constant with time ilp to 120 to 150miinuttes in a nlumber of experimiienits in which thiswas investigated.
A problem encountered throuighouit the plresentinvestigations wvas the presence of signiificant radio-activity in acidic extracts of boiled-enzyme anidchuffer-blank reaction mixtulres. It ,\,as Iolt tn-usual for the amoulnts olf radioactivity extractedfrom btiffer-blank and boiled-enzyme tuibes to ap-proximate 50 % of experimenital valuies. In earlyexperiments it was fotind that part of the blankradioactivity couild be attributed to radioactive im-pulrities in the stockl solu1tion o,f T11P-14C. HeInce,in all subseqtient experimenits, the TTP-14C stocksoluition was checked for radliopuirity by TILC priorto use, can, if necessary, extracted( twice to 3 t meswith AMeCL. . Even wvith this precauition, however,blank valuies for those experiments were neverreduiced to less than 10 % of experimeital valuies.
Thin-lavcr and Gais Chroinatographyl. of Aiatthen-tic IAA Jlcthvl Estcr and Prcsumiptize IAtA-"1CMethyl Ester. Additionial evidelnce for the iden.ti-fication of the 14C-,product as IAA was obtainedby co-chromatograp,hy of atuthentic tinlabele(l IAAmethyl ester and presuimptive IAA-_4C methyl ester.Of the several solvent systems tested usiling authenticmethyl ester, co-c,hromato,graphy of the 2 sampleswas conduicted in only 3, namely CHAc, hexaine-diet,hyl ether (1 :4) and 100 % chloroform. Inthese solvent systems, the mean RF valuies for theatilthentic and presuimptive IAA methyl esters were,respectively: CHAc, 0.68 ancd 0.69; hexane-diethylether, 0.76 and 0.76: and 100 % chloroforni, 0.29and 0.28. In other solvent systems that were testedl,the aulthentic methyl ester either moved to thesolvent fronit, exhibited taililng, or appeared to beinstable.
In figulre 2 are presented dlata from gas chroma-tography of IAA methyl ester. When 10 to 50 ,tgsamiples of auithelntic uinlabeled TAA methyl esterwere chroimatographed, a single peak representingthe inilole compolinld was seeni (fig 2, A). Theretentioni time for the authentic TAA methyl esterwas 6 to 7 mintutes. \When samples of authenticIAA-14C methyl ester (fig 2, B) anld diazomethane-treated 14C-pro,dtuct (fig 2, C) xvere injected intothe gas chromatograph, anid samples collect'-(l offthe coltumn at 1-mintlte intervals, the nmtiir w) rt on
1792
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

MOORE AND SHANER-CELL-FREE BIOSYNTHESIS OF IAA
v)z0
cn
gr
0I--
La
250
a. 200u
I, ISOI-.
< 1000
so
100O
5:
I-.
0
so
w
th
)N-
I-
IC
I-
z
1 2 3 4 5 6 7
K
601-
40_
20 -
1-
ag
O I I
,-I yLXO1 2 3 4 5 6 7
RETENTION TIME.FIG. 2. Data from gas chromatography
ester of IAA. A, gas chromatogram of non
IAA methyl ester. B and C, histograms s
tention times for authentic IAA-14C methvdiazomethane-treated 14C-product (presump14C methyl ester), respectively, on the gas
graph. The data for B and C were obtainescintillation counting.
of the radioactivity in each sample was collectedA also between 6 and 7 mintutes post-injection.
Discussion
Rapid and precise procedures for the isolationand quantitative analysis of IAA from plant prep-arations by purely cheimical means, i.e., withoutreliance on bioassays, are long re,cogn,ized needs inauxinology. The occurrence of IAA in infini-tesimal concentrations in most hi'gher plant tissuies,the relaitively small yields obtained from in vitrobiosynthesis, and the lability of the compotund inliight, 02 atmosphere and strong acid soluitionis pose
=, problems that have never been solved entirely8 9 10 satisfactorily. Problems in identification and anal-
ysis of IAA continue to exist despite the develop-ment of many valuable proce(lures involving paper
B (2, 5, 7, 12, 13, 15, 23, 33, 34), thin-layer (17, 21, 22)and partition column (15, 24, 25, 26) chromatog-raphy; electrophoresis (15, 23); gas chromatog-raphy (26); s9pectrophotofltiorometry (26); color-imetry (2, 5, 7, 12, 13, 15, 23, 24, 26, 33, 34) andspectrophotometry (7).
Obviously the kinds of analytical methods whichcan be em-ployed depend uipon the type of investiga-tion conducted. The 2-step TLC technique heredescribed for the isolation of IAA appears to bea relatively rapid procedure which yields qtuantita-tively reproduciible results. The method is par-ticcularly useful in investigations wherein smallyields of IAA must be analyzed in a nuimber of
' ' ' separate reaction mixtures. Advantaiges of this8 9 10 method for the kind of investigation here described
incltude: (a) relative ease anid rapid.ity with whichC the procedure can be performed; (b) good chroma-
tographic resolution of IAA; (c) quantitativelyreproducible recovery of IAA dutring chromatog-raphy; and (d) excltusioln of non-enzymic conver-sion of IPyA to IAA during chromatography. Thechief disadvantage is the consistent loss of approxi-mately 50 % of a sample of IAA dulring the chro-matography. Tafuri (35) claimed to have recov-ered 95 % of samples of IAA from TLC platescoated with silica gel G and developed in MIA,but ithe methods of analysis which he emiployed aren4ot specific for IAA. Destruction of IAA dutringTLC apparently is partially attributable to thesilica gel G and reportedly can be redtuced by uisingceillullose as the adsorbent (22). While the auithors
8 9 10 have not yet investigated the possibility extensively,it is probable that the percentage loss of IAA
mUTES could be substantially re(dutced also if during repeti-of methyl tive application of extract conta-ining IAA to thin-
-radioactive layer plates a stream of N., in!stead of forced air
Iester and as here uised, were tised to hasten evaporation oftive JAA- tthe solvent. It might be advantageouis also tochromato- employ carboxymethylcelluilose as a binder for
d by liquid silicic acid and to employ some other innovationsrecommended by Obreiter and Stowe (21).
1793
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY'
The chromatographic method was developedspec fecally in connectioni with the isolat on of'4C-labe'el IAA, which accordls the advantage ofbe ng able to measure the amount of IAA by liquidscintillatF on couint'ng or radlochromatcgram scan-ning. However, by stiff c'ently c-ncentrat ng ex-tracts it should be possible to iso&'ate non-radoact veIAA by the proceduire and to mea^;tre the amountof IAA by 1 of several reported proceduires. Itlikely woufld be advantageouis, when applying theprocedulre to chromatography of n-on-rad oact veIAA, to uise small amouints of IAA-'4C as marl-eran(d to employ chromatogram scanning to detectthe position of IAA on the chromatograms. Th:salternative wotuld make possible determ nation ofthe p2rcentage loss of IAA org nally present inthe extract during each chromatographic develop-ment.
'I'he identf.cat on of IAA as a prodtict formedenzvm:cal'y from TTP in cell-free extracts of peasecdl:ng shoot tips is ba-ed on 3 lFnes of ev dence:(a) co:ncident RF value-; of authent c IAA and"4C-product in 3 solvenit systems on TLC, (b) coIn-cid(lenit retent on t:mes of atuthentic IAA methylester an(d d'azomethane-treated 14C-prodlict on gaschromatography, anld (c) co:nciden,t R,, values ofauthentic anid presumptive IAA methyl esters onTLC.
Net yields of IAA-14C in the presenit ;i vest:ga-tion are of sim:lar magnituide, per 1lin:It N, to theyields reported by Gordon (12) for cell-free ex-tracts of !hoot t:ps of Ph-'seo'i.v aureus seedl:nzs.Thuis while recogn;zing the theoretical l)o05 1) l ttvthat other compotunds may serve as precuirsors inIAA bo,ynthesis by a pathwvav :ndlpende i, ofTTP (19, 40), it is concd'ded fr-m the ev:dlnechere presented that cell-frne extracts of Pisu",seedll:n( -hoot tfps canl convert s-gnfcant cuant'-ties of TTP to IAA. The auithors dl- no be! cv-that bacter:al contaminat:on was responsble forthe obs.erved synthesis of IAA, a- a'naret11vwlywsthe case n other inve. t'gat:ons (18, 19, 40). Thecentr ft gal force itsed in the preparat:%)n of the-enzym? extracts was adeutiate to sed ment bacter'alcells wh ch may have been precen- in or onl 'hetissules. Hence interferenc byv ,act-r.al met,bo-lism would have hacd to resuilt from either (a) solln-b:l:zat:on of bacterial enzrmes dulring homovee:za-tionl of the t:ssues or (b) inirodiict-on of wh<'ebacter:a in the preparat:on of r-act-on m:xtulres.The f:rst p5ss:b:l:tv seems uinlkely in that thehomeg-n:zer tused would not be expected to beeff-ct:ve in breaking bacter:a. The second pogg:-bility also seems tinl:kely. The lag t:me for de-velopment of a potental bacler:al popullat:on woukldbe exuected to exceed the 2-ho,' r incl-hation ft:mefor the renction m:xttures, and 100 ,ug/ml each ofl)en;c:llin G and streplomyc:n stulfate shool'd sulp-press potential bacter:al act'v:ty stlbqtant:al;lv.
D:alVsis of enzyme extrac,s and add:t;on ofa-ketoglutarate and pyridoxal phosiphate to reaction
mixtures st'mnulated the conversion olf TTP to IAAquite sebstanti.ally. Hence a transamniase reactIonlof TTP to form JPyA probabily is the int'al react on in the pa'thway of conversion of TTP toIAA uinder these cond t ons. This find'ng agreeswell with the report by Wightman and Coheni (39).wh ch was presen,'ed wh:le the present invest gationwas tunderway. W ghtman and Cohen (39) pr)ro-v'ded convunc'ng ev dence for the en7ymic coniver-s:on of 'T"rP' to JPyA in the presence of addeda-ketog'vtarat,e and pyr d!oxal phosphate in par-tally purifed enzyme preparations from mung, beanseedllings;. Notab!y, in the present investgaat oln.acddtions of a-ketoglhtarate and pyrdoxal p)hos-phate st mnilated net IAA b osynthesis only if theenzyme extract had I emn d'alyzed.
In later exper mentQ, not reported here in d1taialaddlt on of th anm:ne p .rophosphate has been foutndto flrrher enhanice the net yelds of IAA overyields obtained in the presence of a-ketoglultarateand pyr'doxal phos;phate. This find,lng agrees wellwth the re-iults of W:ghtmlu and Cohen (39)wh ch showe(d th'at the yie'cd of indoleacetaldehydewas ncreazed by acldlt on of thiamine pyrophos-phate, and whech the auithors interpret"2d as ev denue,for act vitsy of IPyA decarboxylase in their en7ymeextract,. InexlAicably, in the experments thus farconduicted by the present aulthors, addition of n c()-t nam'de adlen'ne d'nlicleot'de has n )t st:mnllated ne:b ocynthe-,s of IAA over that obcerved in thepre-ence of a-ketoglultarate, pyr:,aoxal phosphatcand -h'am:ne pyrophoFphate.
Th- r'<sults hlr- rep-orted are compat:ble w.ththe pathway for IAA bosvnthesis jrnos< foriXn.n h,- *lt-edl' t"g-- by \Vo'btrnan anwl Cohen (39).in wh:ch IPyA is an intermediate. However, theposs 'blbtv cannot at pre;ent be excludNl the't 1 ormore a.l"rnate pa'llwvays may bie s m'lta1eo)usl\oPera' ve :n n)ea-. Tlh're is -v\:(l-,ce for a pithwiay:,iv, v no d-cP rb^xvlat'on of TTP to fIli tiy-tam ne in a var:e-v of plants includ'ng, P:v7-isct'v'm"t (17, 23, 36, 37, 40). However, tbh p -dlictre tXi'nws from the (leTarboxvlaton (f TFTP inP:-ijtl eiz',-ne exV-ract-. :nve-t`gated by Reed an(dCre"lYug (27) roi"'d not be :del,tfed as trv,1tsun;'ncn.Prel m:nerv attempts 'to isiola±e tryptam:ne from re-act'on m x-i'rs :n tle pre ent ;nvest'vt:on-, haveb^eninconcllqsv-. Obvxously, additional work willbe reoti:red to eliicd(la'e the total comnplemelnt ofract onc. nvilv'd ;n IAA b:osynthesis inl Pisuiitiand ol¢her seedI plants.
Attent:on in the present investigations as di-r'cted aolely t-ward comparicon of net yielids ofTAA :i reacr:o, m x.tureq, a",d tlO eff-rt was madeto determine factors, e.g. IAA oxidase act:vitv,r,'snon b'e for ob-erved d fferences. However, o01the bazi-s of the extensive investigations of cofactorsand ;nhIlb:tors of TAA ox.dase in Pisiunyt by Galctollet al. (3,4,8,9,29), itt is probable that the simtm-lat:nz effect of d alysis of enzyme extracts otn netIAA b osynthesis in preparations of etiolated shoot
1794
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

MOORE AND SHANER-CELL-FREE BIOSYNTHESIS OF IAA
tips resulted from the removal of a cofactor ofIAA oxidase, perhaps a kaempferol derivative.The apparent stimulating effect of dialysis of en-
zyme extract of light-grown tall pea shoot tips may
also be due to removal of a substance or substanceswhich modify IAA oxidase activity.
Maximtum net IAA-14C biosynthesis in cell-free
exitracts of shoot tips of etiolated and light-growndwarf and tall pea seedlings corresponded to theorder: ilight-grown tall > light-grown dwarf >etiolated tall _ etiolated dwarf. This findingagrees with the reported observations that: (a)the concentration of inhibitor of IAA oxidase ishigher in light-grown than in etiolated pea seed-lings, and higher in some tall cultivars of peas thanin some dwarf cultivars (9, 11, 20); (b) light-grown Alaska pe,a seedlings contain more auxinthan etiolated seedlings (30,31, 32); and (c) theoptimuim concentration of IAA for the growth ofstem sections from green pea seedlings is 100- to1000-fold higher than for stem sections from etio-lated seedlings (9, 10, 11). Hence it may be hypo-thesized that the differenit rates of stem elongationamong light-grown and etiolated dwarf and tallpea seedlings are correlated with differences in netIAA synthesis and sensitivity to IAA. Accordingto this hypothesis, pea seedllings exhibiit higher net
auxin production in the 'light than in the dark, andboth dwarf and tall seedlings exhibit reduced sensi-tivity to auxin when grown in the light, 'as com-
pare,d to their 'respective etiola,ted counterparts.Interestingly, the data here reported for etiolatedpea seedlings are only slightly at variance with thereport by von Abrams (38) who, using the lesssensitive methods available to him in 1953, fouindno notable differences in the capacities of etiolatedDwarf and Tall Telephone pea seedlings to syn-
thesize atuxin from TTP or to enzymically destroyauxin. Neither did von Abrams detect a -differencein the amounts of extrac.table or diffusible auxinin epicotyls of etiolated dwarf and tall seedlings.
In summary, the authors believe that the dif-ferent rates of stem elongation among etiolated andIlight-grown dwarf 'and tall peas are correlated withdifferences in auxin relations. However, the ob-served differences in stem growth clearly are not
explainable excltusively oIn an aulxin basis, sincecorrela,tive differences in endogen-ons gilherellinrelations are known to exist also.
Acknowledgments
The authors express their gratitude for the valuableassistance and criticisms of Dr. D. J. Reed and thetechnical assistance of Mrs. Lynda Ty,l.
Literature Cited
1. ANDERSON, J. D. AND T. C. MOORE. 1967. Bio-synthesis of (-)-kaurene in cell-free extractsof immature pea seeds. Plant Physiol. 42: 1527-34.
2. BENTLEY, J. A., K. R. FARRAR, S. HOUSLEY, G. F.SMITH, AND W. C. TAYLOR. 1956. Some chemi-cal and physiological properties of 3-indolylpyruvicacid. Biochem. J. 64: 44 49.
3. BOTTOMLEY, W., H. SMITH, AND A. W. GALSTON.1965. A phytochrome mediated effect of lighton the hydroxylation pattern of flavonoids inPisuni sativum var. 'Alaska'. Nature 207: 1311-12.
4. BOTTOMLEY, W., H. SMITH, AND A. W. GALSTON.1966. Flavonoid complexes in Pisum sativum. III.The effect of light on the synthesis of kaempferoland quercetin complexes. Phytochemistry 5:117-23.
5. DANNENBURG, W. N. AND J. L. LlVERMAN. 1957.Conversion of tryptophan-2-14C to indoleaceticacid by watermelon tissue slices. Plant Physiol.32: 263-69.
6. DEDIO, W. AND S. ZALIK. 1966. Gas chromatog-raphy of indole compounds. Anal. Biochem. 16:36-52.
7. FLETCHER, R. A. AND S. ZALIK. 1963. Quantita-tive spectrophotometric determination of indolyl-acetic acid. Nature 199: 903-04.
8. FURUYA, M., A. W. GALSTON, AND B. B. STOWE.1962. Isolation from peas of co-factors and in-hibitors of indolyl-3-acetic acid oxidase. Nature193: 456-57.
9. GALSTON, A. W. 1959. Studies on indoleaceticacid oxidase inhibitor and its relation to photo-morphogenesis. In: Photoperiodism, R. B. With-row, ed. Am. Assoc. Adv. Sci., Washington,D. C. p 137-57.
10. GALSTON, A. W. AND R. S. BAKER. 1951. Studieson the physiology of light action. IV. Lightenhancement of auxin-induced growth in greenpeas. Plant Physiol. 26: 311-17.
11. GALSTON, A. W. AND R. KAUR. 1961. Compara-tive studies on the growth and light sensitivity ofgreen and etiolated pea stem sections. In: Lightand Life. W. D. McElroy and B. Glass, eds.Johns Hopkins Press, Baltimore. p 687-705.
12. GORDON, S. A. 1958. Intracellular localization ofthe tryptophan-indoleacetate enzyme system. PlantPhysiol. 33: 23-27.
13. GORDON, S. A. AND L. G. PALEG. 1961. Formationof auxin from tryptophan through action of poly-phenols. Plant Physiol. 36: 838-45.
14. GRAEBE, J. E., D. T. DENNIS, C. D. UPPER, ANDC. A. WEST. 1965. Biosynthesis of gibberellins.I. The biosynthesis of (-) -kaurene, (-) -kauren-19-ol, and trans-geranylgeraniol in endospermnucleus of Echinocystis macrocarpa Greene. J.Biol. Chem. 240: 1847-54.
15. HAMILTON, R. H., R. S. BANDURSKI, AND B. H.GRIGSBY. 1961. Isolation of indole-3-acetic aci(dfrom corn kernels and etiolated corn seedlings.Plant Physiol. 36: 354-59.
16. HORWITZ, W., ed. 1965. Micro-Kjeldahl-official,final action. In: Official Methods of the Associa-tion of Agricultural Chemists, 10th ed. p 744 45.
17. LANTICAN, B. P. AND R. M. MUIR. 1967. Isola-tion and propertieis of the enzyme system formingindoleacetic acid. Plant Physiol. 42: 1158-60.
18. LIBBERT, E. AND S. WICHNER. 1963. Tryptophan-abbau zu Indolessigsaure durch pflanzenbesie-delnde Mikroorganismen. Naturwiss. 50: 451.
1795
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
19. LIBBERT, E., S. NVICHNER, U. SCHIEFWER, H. RiscH,AND \V. KAISER. 1966. The inifluenice of epi--phytic bacteriae on autxini metabolism. Planta68: 327-34.
20. AiICCUNE, D. C. AND A. W. GALSTON. 1959. In-verse effects of gibberellin on peroxidase activityand growth in dwarf strains of peas anld corn.Plant Physiol. 34: 416-18.
21. OBREITER, J. B. AND B. B. STOWA-E. 1964. Car1oxy-methvlcellulose: a binder for thin-laver chromiia-tography of lipids and inidoles. J. Chromatog.16: 226-28.
22. OHWAKI, Y. 1966. Thii-laaer chromatography ofdiffusible auxin of corn coleoptiles. Botan. 'Mag.Tokyo 79: 200-01.
23. PERLEY, J. E. AND B. B. STOWN E. 1966. On theability of Taphrina deforuzianis to produce indole-acetic acid from tryptophan by way of tryptamine.Plant Physiol. 41: 234-37.
24. POWELL, L. E. 1960. Separationi of plaint growthiregulating substanices on silica gel columns. PlantPhysiol. 35-: 256-61.
25. POWELL, L. E. 1963. Solvent systems for silica-gel columin chromatography of indole (lerivatives.Nature 200: 79.
26. POWELL, L. E. 1964. Preparation of in(lole ex-tracts from plants for gas chromatography aidspectrophotofluorometry. Plant Physiol. 39: 836-42.
27. REED, D. J. AND J. CRECELIUS. 1967. Light-inducedinhibition of tryptophan decarboxylation in cell-free extracts of pea epicotyls. Plant PhIysiol. 42:130306.
28. ROBINsON, D. R. AND C. A. WEST. 1967. Biosyn-thesis of (-)-kaurene and other diterpenes inextracts of germinating castor beatn seeds. Fed-eration Proc. 26: 1099.
29. RUSSELL, D. W. AND A. W. GALSTON. 1967. Fla-vonoid complexes in Pisu7n sativZumni. IV. Theeffect of red light on synthesis of kaempferol
colmiplexes and( oni growthl in sub-apical internodetissues. Phytochemistrv 6: 791-97.
30. SCOTT, T. K. AND W. R. BRIGGS. 1960. Auxinrelationships in the Alaska pea (Pisumi sativumll).Am. J. Botany 47: 492-99.
31. SCOTT, T. K. AND W. R. BRIGGS. 1962. Recoveryof native and applied atixin fromii the light-growni'A\laska' pea seedling. Am. J. Botany 49: 1056-63.
32. SCOTT. T. K. AND W. R. BRIGGS. 1963. Recoveryof native and applied aulxini froml the dark-grown'Alaska' pea seedling. Am. J. Botany 50: 652-57.
33. STAHL, E., ed. 1965. Thin-Layer Chromatography.Springer-Verlag, Berlin. Heidelberg, New York.553 p.
34. STOWNE. B. B. AND K. . THIMNIANN. 1954. Thepaper chromatography of indole compounds andsome indole-containing auxins of pl)ant tissues.Arch. Biochem. Biophys. 51: 499-516.
35. T'AFURI. F. 1966. IAA determination in the ker-nels of 4 lines of corn can(I of their hybrids.Plhvtochemistrv 5: 999-1003.
.36. V-ALDOVINOS, J. G. AND L. C. ERNEST. 1966. Gi)-berellin-enhanced CO, rielease from tryptophan-1-'4C in plant apical tissuie. Planit Physiol. 41:1551-52.
37. VALDOVINOS, J. (G. ANI) J. H. PERLEY. 1966. Me-tabolism of tryptoplhan in petioles of Colcus. PlantPhvsiol 41: 1632-36.
38. VON ABRAMS, G. J. 1953. Auxin relations of a
dlwarf pea. Plant PhVsiol. 28: 443-56.39. WIGHTMAN, F. ANT) D. COHEN. 1968. Intermedi
ary steps in the enzymatic conversion of trypto-ph-in to IAA ini cell-free systems from higlherplants. In: The Physiology and Biochemistrv ofPlant Grow!tlh Substances. G. Setterfield and F.Wi-ightmal1 C(es. Runiige Press, Ottawa. In press.
40. WVINTER, A. 1966. A hypothetical route for thebiogenesis of IA A. Plainta 71: 229-39.
1796
www.plantphysiol.orgon May 9, 2020 - Published by Downloaded from Copyright © 1967 American Society of Plant Biologists. All rights reserved.