biological responses of tropical cladoceran, ceriodaphnia ... · biological responses of tropical...
TRANSCRIPT

Biological Responses of Tropical Cladoceran,
Ceriodaphnia Cornuta to Different Algae Diets
Hasnun Nita Ismail Faculty of Applied Sciences/Universiti Teknologi MARA (Perak) Kampus Tapah, 34500 Tapah Road, Perak, Malaysia
Email: [email protected]
Umi Wahidah Ahmad Dini and Chia Chay Tay Faculty of Applied Sciences/Universiti Teknologi MARA (Perlis), 02600 Arau, Perlis, Malaysia
Email: [email protected]; [email protected]
Abstract—The biological responses of tropical cladoceran,
Ceriodaphnia cornuta fed on different algae diets were
investigated under laboratory conditions. Two types of
microalgae, Chlorella (C) and Scenedesmus (S) were given
as mono-algal diet and multi-algal diet (CxS). The different
types of algae diets have significantly altered the biological
responses of C. cornuta. Diet contained Chlorella alone
promoted high longevity, fast egg development, early
maturation, increased egg clutch number and increased egg
and offspring production. As a result, animals showed
higher survival, faster development and greater
reproductive output. On the other hand, diet with
Scenedesmus alone supports for higher growth rate more
than Chlorella. Despite of the opposite algal effect, diet
combination of both algal (Chlorella x Scenedesmus)
enhanced all biological responses. Therefore, this study
suggests that the diet combination of both algae is the
optimum diet to boost up the survival, development and
reproduction of C. cornuta. This outcome has provides the
guideline to choose the suitable algal species which can
maximize the biological performance of C. cornuta.
Index Terms—tropical cladoceran, algae diets, growth rate,
development, reproduction, survival
I. INTRODUCTION
Cladoceran forms the most dominating group of
micro-crustacean in tropical freshwaters especially in
lakes, ponds and rivers. Although microscopic in size,
cladoceran plays a dual role in food web as a primary
consumer and a prey for higher consumers [1]. They
influence energy and organic matter transfer from
primary producers to the highest consumers [2]. As a
primary consumer, most cladoceran are herbivorous.
They have the ability to control the production of
producer species in aquatic systems especially the
microalgae [3]. In fact, microalgae have been the primary
diet chosen by the aquatic systems especially the
microalgae [3]. In fact, microalgae have been the primary
diet chosen by the herbivorous cladocerans beside of
alternative diets like bacteria, debris, unicellular fungi
and humus.
Manuscript received August 19, 2014; revised December 1, 2014.
Tropical region such as Malaysia is well known to
support growth of many species due to it warm
environments [4]. Indeed, tropical cladoceran should be
more diverse with many selections of food diet here.
However, reported studies on tropical cladoceran in
South East Asia especially from Malaysia are limited [5].
Many studies are restricted to taxonomy, distribution and
zoogeographic analysis [6], [7]. Thus far, none of the
studies reveal the important parts related to biological
and ecological requirements in the life history of
Malaysian cladoceran.
The genus Ceriodaphnia is classified under the family
of Daphniidae [8]. In their life cycle, Ceriodaphnia
displays cyclic parthenogenesis which is typical in nearly
all groups of cladoceran. Cyclic parthenogenesis consists
of alternation between asexual and sexual reproduction
[9]. Asexual reproduction is the major mode of
reproduction conducted under favourable conditions by
means of parthenogenesis. Meanwhile, sexual
reproduction begins as the environments start to
deteriorate and challenge the population’s survival. The
outcome from sexual reproduction is the resting eggs
which are resistant under harsh environment and hatch
once the environments back to favourable conditions.
In many diet studies, the biological responses of
cladoceran vary when fed to algae of different taxa. The
cladoceran, C. silvestrii lived longer when fed on
Chlorella lacustris but showed the shortest longevity
when fed on Scenedesmus bijugus [10]. The growth of
euryhaline cladoceran, Daphniopsis australis grew faster
when fed on Nannochloropsis oculata than fed to
Tetraselmis suecica [11]. Other euryhaline cladoceran,
Moina mongolica matured earlier with N. oculata but
maturation was delayed by Dunaliella salina [12]. The
reproductive performance of freshwater cladoceran,
Simocephalus vetulus was significantly higher when fed
to Scenedesmus acutus then fed to other algae such as
Microcyctis and Planktothrix [13]. In general, the life
responses of cladoceran are species-specific and likely
dependent on the types of algae.
This study covers the Malaysian cladoceran species, C.
cornuta isolated from wild freshwater environments. C.
cornuta is a small tropical cladoceran with a body size
less than 0.50 mm [8]. The first objective is to examine
482014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014
doi: 10.12720/jolst.2.2.48-54

the biological responses of C. cornuta fed to different
types of microalgae; Chlorella and Scenedesmus. The
second objective is to determine the optimal food for C.
cornuta cultured under laboratory conditions. The finding
will show the impact of algal diet to the survival,
development and reproduction and provide a guideline in
feeding selection of this cladoceran under man-made
conditions. Overall, it might shed light in introducing this
species as an alternative live food in fish aquaculture.
II. MATERIALS AND METHODS
A. Culture of Algae
The freshwater green algae used in this study include
Chlorella and Scenedesmus. Both algal species were
cultured in BBM medium in 2 L glass bottle provided
with aeration. The air was filtered using Nylon air filter,
0.45 µm pore size. Light was continuously provided at
1900 lux with 12 h light: 12 h dark photoperiod.
Temperature was controlled at 25°C. Algae were
harvested for every two days at exponential growth phase
to feed the stock culture of C. cornuta.
B. Stock Culture of C. Cornuta
C. cornuta was obtained from Biological Research
Laboratory, Universiti Teknologi MARA (Perlis),
Malaysia. Originally, the species was collected from
freshwater Tasik Elham around the campus in July, 2013.
Since then, the animals have been cultured for more than
100 generations under laboratory condition. C. cornuta
have been maintained in aged tap water in 5L beaker.
The stock culture condition was maintained at 25 oC
under 12 h light:12 h dark regime at 1900 lux. The
population was fed on Chlorella and Scenedesmus
separately at the density of 106
algal cells/mL. The
population density of C. cornuta was maintained below
1000 individual/L to reduce overcrowding.
C. Experimental Design
The response of C. cornuta to different diet was tested
with two mono-algal diets which were Chlorella (C),
Scenedesmus (S) and one multi-algal diet which was the
mixture of both Chlorella and Scenedesmus (CS). Algal
cells were harvested at exponential growth phase for
feeding regimes. Enumeration of algal density was
carried by using Improved Neubaeur haemocytometer
under the light microscope (Olympus) at 400
magnifications. Since the size of two algae was highly
variable, the biovolume measurement was used to
standardize the concentration of food [14].
Experiment on the life cycle of C. cornuta was started
by isolating a single parthenogenetic female from the
stock culture and acclimatized in a culture bottle
containing 5 mL medium from each corresponding food
diet Chlorella, Scenedesmus and a mixture of them
respectively. The parthenogenetic females were left until
the appearance of the third generation. During
acclimatization, the medium was renewed daily to avoid
confounding effects from dead microalgae. The
acclimatization procedure is important and is a standard
procedure to offset the maternal effect [15]. Only the
animals after the third generation and above were used
for observation on the life cycle responses.
In the life cycle experiment, the animals younger than
24h were individually inoculated into a bottle containing
5mL of medium for each food treatment. Temperature
and photoperiod was maintained similar to the culture
stock condition. 25 replicates were conducted for each
food treatment and the culture media were renewed on
daily basis. The life cycle parameters recorded for each
individual include longevity (day), growth rate (mm/day),
age at first reproduction (day), egg development time
(day), total clutch (female/life time), total egg per clutch
(female/life time) and total offspring (female/life time).
Daily observation was conducted until the death of the
last individual. The Olympus SZ51 zoom
stereomicroscope (up to 80x total magnification) was
used for observation on the individual response to each
different diet.
D. Statistical Analysis
All data were analyzed using statistical software, SPSS
version 20.0. In prior, data were explored for
homogeneity variance and assessing normality
assumptions. Only data for longevity were inversely
transformed to achieve the normality assumption. One-
way analysis of variance (ANOVA) will be used to test
the treatment effects on each life cycle variable to find
the significant difference among treatments. The
difference between groups will be compared using
Tukey’s test. All significant level will be set at P<0.05.
III. RESULTS
A. Longevity
The individuals longevity significantly differed
between the diet treatments (ANOVA, P<0.05; Table I).
Further analysis revealed that individuals fed on the
combination of Chlorella and Scenedesmus showed the
longest longevity rather than those fed on other diet
treatments (Tukey's HSD, P<0.05; Fig. 1a). The
longevity was significantly shortened as individuals fed
on the Scenedesmus alone (P<0.05). However, no
significant difference in longevity was found when
individuals fed on Chlorella alone and fed on
combination of Chlorella and Scenedesmus (P<0.05).
B. Age at First Reproduction (AFR)
The age at first reproduction (AFR) was significantly
affected by the diet treatments (ANOVA; P<0.05; Table
I). Specifically, Scenedesmus alone significantly
prolonged the AFR of individuals (Tukey's HSD, P<0.05;
Fig. 1b). In comparison, individuals significantly
shortened the AFR when fed on combination of
Chlorella and Scenedesmus (P<0.05). However, AFR
was not significantly different between individuals fed on
Chlorella alone and fed on the combination of Chlorella
and Scenedesmus (P<0.05). Regarding the diet treatments,
the time to reach maturity is significantly faster for those
individuals fed on the combination of Chlorella and
Scenedesmus than those fed on the Scenedesmus.
492014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014
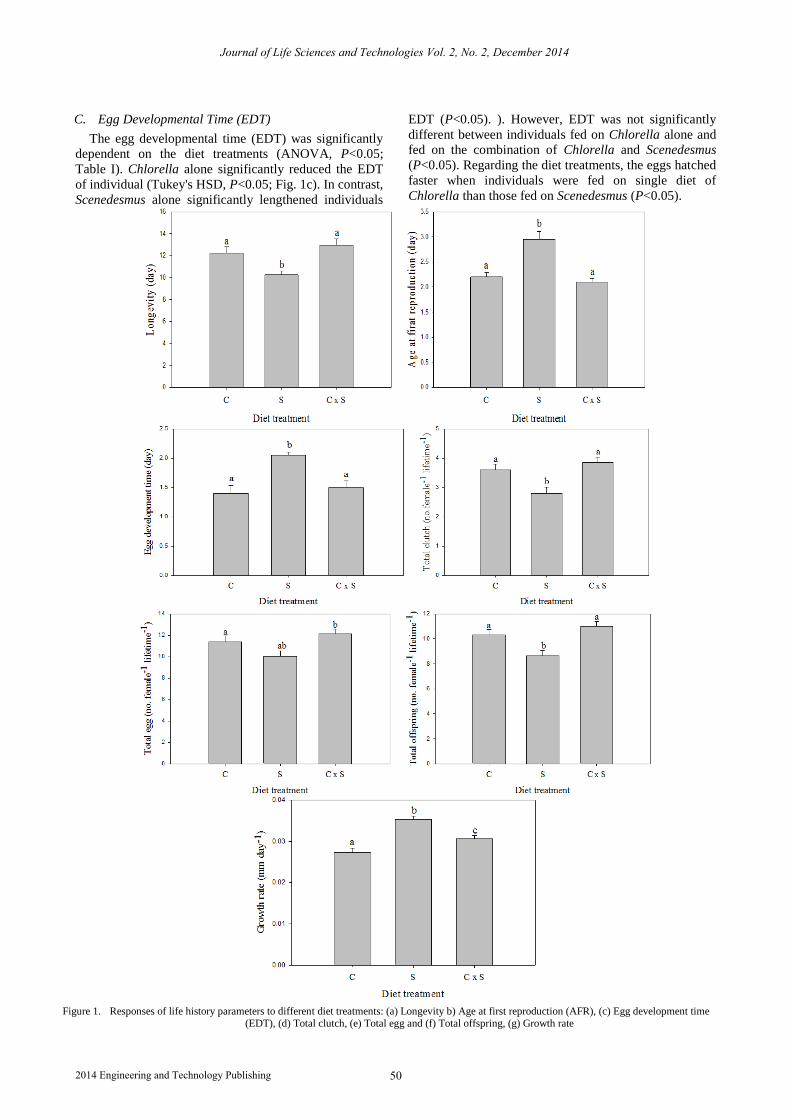
C. Egg Developmental Time (EDT)
The egg developmental time (EDT) was significantly
dependent on the diet treatments (ANOVA, P<0.05;
Table I). Chlorella alone significantly reduced the EDT
of individual (Tukey's HSD, P<0.05; Fig. 1c). In contrast,
Scenedesmus alone significantly lengthened individuals
EDT (P<0.05). ). However, EDT was not significantly
different between individuals fed on Chlorella alone and
fed on the combination of Chlorella and Scenedesmus
(P<0.05). Regarding the diet treatments, the eggs hatched
faster when individuals were fed on single diet of
Chlorella than those fed on Scenedesmus (P<0.05).
Figure 1. Responses of life history parameters to different diet treatments: (a) Longevity b) Age at first reproduction (AFR), (c) Egg development time (EDT), (d) Total clutch, (e) Total egg and (f) Total offspring, (g) Growth rate
502014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014

D. Total Clutch
Diet treatments significantly affected the total number
of egg clutch of individuals (ANOVA, P<0.05; Table I).
The highest number of egg clutch occurred in females fed
on combination of Chlorella and Scenedesmus (Tukey's
HSD, P<0.05; Fig. 1d). Meanwhile, Scenedesmus alone
significantly lowered the total egg clutch of individuals
(P<0.05). However, no significant differences were
observed on the number of egg clutch between females
fed on Chlorella alone and fed on mix diet of Chlorella
and Scenedesmus (P<0.05). Analysis revealed that food
affected significantly on the reproductive frequency of C.
cornuta.
TABLE I. RESULTS OF ANOVA FOR LIFE CYCLE PARAMETERS OF C. CORNUTA FED ON DIFFERENT DIET TREATMENTS
One-way ANOVA df SS MS F P
AFR (day)
Diet treatment
Error
2
57
8.63
13.95
4.32
0.24
17.64
0.000*
EDT (day)
Diet treatment
Error
2 57
4.90 12.75
2.45 0.22
10.95
0.000*
Total egg clutch
(no.female-1 lifetime-1)
Diet treatment
Error
2
57
12.03
42.55
6.02
0.75
8.06
0.001*
Total egg
(no. female-1 lifetime-1 )
Diet treatment
Error
2
57
45.30
268.30
22.65
4.71
4.81
0.012*
Total offspring
(no. female-1 lifetime-1)
Diet treatment
Error
2
57
58.23
188.75
29.12
3.31
8.79
0.000*
Longevity (day)
Diet treatment
Error
2
57
0.00
0.16
0.00
0.00
7.90
0.001*
Growth rate
(mm day-1)
Diet treatment
Error
2 57
0.01 0.01
0.00 0.00
19.75
0.000*
df — degree of freedom, SS — sum of squares, MS — mean square
*significant different at P < 0.05
E. Total Egg
Diet treatments had a significant effect on the number
of egg production (ANOVA; P< 0.05; Table I). The
highest numbers of total number of egg were obtained by
the individuals fed on mix diet, Chlorella and
Scenedesmus whereas the lowest values were recorded in
those fed on Scenedesmus. alone (Tukey's HSD, P<0.05;
Fig. 1e). Total number of egg produced was influenced
significantly among each diet treatment (P<0.05).
Individuals fed Scenedesmus produced a total number of
egg that was significantly lower than that obtained with
single diet, Chlorella and combination of Chlorella and
Scenedesmus(P<0.05).
F. Total Offspring
There was a significantly effect of diet treatments on
the number of offspring production (ANOVA; P<0.05;
Table I). Total number of offspring production
significantly reduced when individuals fed on
Scenedesmus alone while the highest production of
offspring obtained by individuals fed on combination of
Chlorella and Scenedesmus (Tukey's HSD, P<0.05; Fig.
1f). However, total number of offspring production was
not significantly different between individuals fed on
Chlorella alone and combination of Chlorella and
Scenedesmus (P<0.05).
G. Growth Rate
Dietary types significantly affect the growth rate of C.
cornuta (ANOVA, P<0.05; Table 2). Growth rate
differed at each dietary treatment. Individuals fed on
Scenedesmus alone showed the highest growth rate while
the lowest growth rate was found in individuals fed on
Chlorella alone (Tukey's HSD, P<0.05; Fig. 1g).The
growth rate showed by single diet treatment is
significantly differ from those fed on multi-algal species
Chlorella and Scenedesmus (P<0.05).
IV. DISCUSSION
A. Longevity
In this study, the longevity of C. cornuta was
lengthened when fed on the multi-diet (Chlorella x
Scenedesmus) and the mono-algae, Chlorella. The
shortest longevity displayed when C. cornuta was fed on
Scenedesmus alone. The same finding was reported on
the other species of Ceriodaphnia, C. dubia where the
longevity was enhanced in similar combination algal diet
(Chlorella x Scenedesmus) and the longevity was
hampered with Scenedesmus alone [16]. Other study on
C. silvestrii reported that the highest longevity was
obtained with Chlorella alone rather than Scenedesmus
alone [10]. However, the longevity of C. rigaudi was the
shortest when fed on Chlorella alone [17]. Likely, the
longevity of Ceriodaphnia species is affected by the
types of diet and the addition of Chlorella in diet mostly
provides positive effect to their survival.
B. Development Performance
The development performance by C. cornuta was
evaluated by referring to their age at first reproduction
(AFR) and egg developmental time (EDT). The present
result showed that the fastest maturity observed in C.
cornuta fed on multi-diet (Chlorella x Scenedesmus) as
well as in Chlorella alone. The trend of eggs
development is also similar with AFR using the same
diets. However, C. cornuta matured later and their eggs
developed longer with Scenedesmus alone.
Correspondingly, study on other cladoceran, Daphnia
magna showed faster maturation when fed on Chlorella
than Scenedesmus alone [18]. Likewise, C. cornuta and
M. macrocopa matured at early age and showed fastest
EDT when fed on Chlorella alone [19]. Other study also
showed slowest AFR and longest EDT of C. dubia when
fed on Scenedesmus alone [20]. Therefore, the
development of most cladoceran species are supported by
Chlorella compared to Scenedesmus.
C. Reproductive Performance
In this study, C. cornuta obtained the maximum
reproductive output when fed on multi-diet (Chlorella x
512014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014

Scenedesmus) and Chlorella alone. This showed by the
highest total number of egg clutch, egg number and total
offspring. However, less number of egg clutch, egg
number and total offspring produced in C. cornuta fed on
Scenedesmus alone. This finding is parallel when the
same diet was applied to C. dubia [16] and Daphnia
magna [18].Similarly, the other study showed that the
highest reproduction output displayed by M. macrocopa
[21] and M. micrura [22] fed on Chlorella alone rather
than Scenedesmus alone. Again, diet contained with
Chlorella supports reproduction of many tropical
cladoceran more than Scenedesmus.
D. Growth Rate Performance
In contrast to other life cycle parameters, the growth
rate of our species, C. cornuta was highly promoted
when fed on Scenedesmus alone. However, the mono-diet,
Chlorella resulted with the shortest growth rate andthe
intermediate growth rate was observed when C. cornuta
fed on multi-diet (Chlorella x Scenedesmus). The growth
rate of C. dubia also the fastest when fed on diet with
Scenedesmus alone while diet with Chlorella alone
produced the lowest growth rate [16], [21]. Likewise, the
growth rate of D. gessneri was the fastest using diet with
Scenedesmus alone [22]. Most of the Daphnia spp.
displayed the shortest growth rate with diet contained
Chlorella alone [23]. Hence, the growth rate of
cladoceran can be maximized with Scenedemus rather
than Chlorella.
In overall, our finding on C. cornuta found that
Chlorella seems to be the best diet for survival,
development and reproduction. However, Scenedesmus is
the best diet to promote for species growth. In many diet
studies, the life performance of cladoceran can be
improved when the diet contained combination of poor
algae and the good algae. For instance, the survival of
Daphnia pulicaria was shortest when supplied with
Cyclotella meneghiniana but the combination with
Scenedesmus resulted in better survival and development
of this cladoceran [24]. Moreover, Cryptomonas are
considered as the high-quality food for Daphnia pulex
due to high production of offspring but the production of
offspring was impeded when fed on Scenedesmus [25].
Interestingly, the combination of Cryptomonas and
Scenedesmus boosted the reproduction of the D. pulex.
Study on different cladoceran, Daphniopsis australis also
found that Tetraselmis suecica was the best diet to
improve growth while Nannochloropsis oculata was the
worst diet for growth. However, combination of these
algae increased the growth rate significantly [11].
Therefore, while Scenedesmus is suitable for growth and
Chlorella support more on development, survival and
reproduction, the combination of both algae managed to
improve all life parameters involved in the life history of
C. cornuta.
The positive results showed by Chlorella on the
development and reproductive performance of C. cornuta
may due to the highest lipid contents in comparison to
Scenedesmus [26]-[28]. The essential lipid content to
determine the cladocerans reproduction refers to the
group of PUFAs [29]. Many studies have shown that
Chlorella contained 43.37 to 47.12 % PUFAs while
Scenedesmus contained 29.56 to 30.17 % PUFAs [30]-
[32]. The higher amount of PUFAs in Chlorella than
Scenedesmus might be the reason that led to better
development and reproduction of C. cornuta in this study.
Specifically, the growth rate of cladoceran is related to
the amount of α-linolenic acid (α -LA) in algal diet where,
algae with high α-LA content contribute most to the body
growth [33], [34]. Previous studies have shown that
Scenedesmus contained 25.61 % of α-LA content while
Chlorella contained 2.28 to 4.21 % of α-LA content [35],
[36]. Therefore, the difference of α-LA composition in
both algal species could be considered as one of the
factor that affects the growth rate of this C. cornuta.
Overall, the variation in life cycle responses of C.
cornuta to different algal diets in this study may have
been caused by the differences in nutritional quality of
algal diet. The evidence for the effects of nutritional
quality comes from the experiment with multi-diet which
showed enhancement in all life cycle parameters. This is
because different algae can complement to each other
with respect to nutritional quality [37]. Hence, the
combination of different food quality of algal diets
contributed to the better life cycle performance of C.
cornuta.
V. CONCLUSION
Our results showed that different types of alga diet
significantly affect the life cycle of C. cornuta. Diet
contained Chlorella alone enhanced higher survival,
faster development and greater reproductive output in
comparison to Scenedesmus. On the other hand,
Scenedesmus supports for higher growth rate more than
Chlorella. Despite of opposite effects, combinations of
both algal enhanced all life cycle parameters. Therefore,
this study suggests that the combination of both algal
diets is the optimal food for C. cornuta. This study has
provides the guideline to choose the suitable algal species
which can maximize the performance of C. cornuta.
Further study should be conducted in detail to examine
the nutritional composition of algal species which needed
by the cladoceran to meet their life requirement. In
addition, further study has to be conducted to assess the
ingestion rate and food selectivity in this cladoceran. Conclusively, this study has reveals the performance
of thiscladoceran species as well as indicate the most
optimal condition for the better production. By knowing
all the requirements to achieve the maximal production,
this species can possibly be reared in man-made
condition as the alternative live food in aquaculture.
Therefore, the continuous supplying of live food for the
fish larvae can give beneficial impact to the aquaculture
industry which then can supports the demand for
aquaculture products from time to time.
ACKNOWLEDGEMENT
Authors would like to thank UniversitiTeknologi Mara
for supporting this project technically and financially.
This project was granted under the Academic Excellent
522014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014

Fund with project number was 600-
UiTMPs/PJIM&A/ST/DCKP 5/3 (07/2012).
REFERENCES
[1] A. F. Alva-Martínez, S. S. S. Sarma, and S. Nandini, “Effect of
mixed diets (cyanobacteria and green algae) on the population
growth of the cladocerans ceriodaphnia dubia and moina macrocopa,” Aquatic Ecology, vol. 41, no. 4, pp. 579-585, 2007.
[2] A. Infante and A. H. Litt, “Differences between two species of
daphnia in the use of 10 species of algae in Lake Washington,” Limnology and Oceanography, vol. 30, no. 5, pp. 1053-1059,
1985.
[3] A. S. Ferrao-Filho and M. S. Arcifa, “Growth responses of tropical cladocerans to seston from Lake Monte Alegre (Brazil)
supplemented with phosphorus, fatty acids, a green algae and a cyanobacterium,” Hydrobiologia, vol. 563, no. 1, pp. 155-165,
2006.
[4] A. S. Ferrão-Filho, M. S. Arcifa, and C. Fileto, “Resource limitation and food quality for cladocerans in a tropical Brazilian
lake,” Hydrobiologia, vol. 491, no. 1-3, pp. 201-210, 2003.
[5] B. A. G. Idris, Freshwater Zooplankton of Malaysia, Crustacea, Cladocera, Universiti Pertanian Malaysia, Serdang, Selangor,
Malaysia, 1983, pp. 153.
[6] B. Czeczuga, M. Kozlowska, and B. Kiziewicz, “Effect of various types of phytoplankton on fertility, egg size and duration of
postembryonic growth of a few plankton representatives of
cladocera (Crustacea),” Polish Journal of Environmental Studies, vol. 12, no. 5, pp. 545-556, 2003.
[7] D. J Innes and D. R. Singleton, “Variation in allocation to sexual
and asexual reproduction among clones of cyclically parthenogenetic daphnia pulex (Crustacea: Cladocera),”
Biological Journal of the Linnean Society, vol. 71, no. 4, pp. 771-
787, 2000. [8] D. Taghavi, O. Farhadian, N. M. Soofiani, and Y. Keivany,
“Effects of different light/dark regimes and algal food on growth,
fecundity, ephippial induction and molting of freshwater cladoceran, ceriodaphnia quadrangular,” Aquaculture, vol. 410, pp.
190-196, 2013.
[9] D. Tang, W. Han, P. Li, X. Miao, and J. Zhong, “CO2 biofixation and fatty acid composition of scenedesmus obliquus and chlorella
pyrenoidosa in response to different CO2 levels,” Bioresource
technology, vol. 102, no. 3, pp. 3071-3076, 2011. [10] E. von Elert, “Determination of limiting polyunsaturated fatty
acids in daphnia galeata using a new method to enrich food algae
with single fatty acids,” Limnology and Oceanography, vol. 47, no. 6, pp. 1764-1773, 2002.
[11] E. W. Becker, “Micro-algae as a source of protein,”
Biotechnology advances, vol. 25, no. 2, pp. 207-210, 2007. [12] F. Peña-Aguado, S. Nandini, and S. S. S. Sarma, “Differences in
population growth of rotifers and cladocerans raised on algal diets
supplemented with yeast,” Limnologica-Ecology and Management of Inland Waters, vol. 35, no. 4, pp. 298-303, 2005.
[13] F. Martinez-Jeronimo and C. Ventura-Lopez, “Population
dynamics of the tropical cladoceran ceriodaphnia rigaudi Richard, 1894 (Crustacea: Anomopoda). Effect of food type and
temperature,” Journal of Environmental Biology, vol. 32, no. 4, pp.
513-521, 2011. [14] G. Ahlgren, L. Lundstedt, M. Brett, and C. Forsberg, “Lipid
composition and food quality of some freshwater phytoplankton
for cladoceran zooplankters,” Journal of Plankton Research, vol. 12, no. 4, pp. 809-818, 1990.
[15] G. Muñoz-Mejía and F. Martínez-Jerónimo, “Impact of algae and
their concentrations on the reproduction and longevity of cladocerans,” In Annales de Limnologie. vol. 43, no. 30, pp. 167-
177, 2007. [16] H. N. Ismail, J. G. Qin, and L. Seuront, “Dietary responses of the
brackish cladoceran daphniopsis australis fed on different marine
microalgae,” Journal of Experimental Marine Biology and Ecology, vol. 409, no. 1-2, pp. 275-282, 2011.
[17] J. Karlsson and P. Byström, “Littoral energy mobilization
dominates energy supply for top consumers in subarctic lakes,” Limnology and Oceanography, vol. 50, no. 2, pp. 538-543, 2005.
[18] J. Pratoomyot, P. Srivilas, and T. Noiraksar, “Fatty acids
composition of 10 microalgal species,” Songklanakarin Journal of
Science and Technology, vol. 27, no. 6, pp. 1179-1187, 2005.
[19] J. Y. Lee, C. Yoo, S. Y. Jun, C. Y. Ahn, and H. M. Oh, “Comparison of several methods for effective lipid extraction
from microalgae,” Bioresource Technology, vol. 101, no. 1, pp.
75-77, 2010. [20] L. N. De Senerpont Domis, J. J. Elser, A. S. Gsell, V. L. Huszar,
et al., “Plankton dynamics under different climatic conditions in
space and time,” Freshwater Biology, vol. 58, no. 3, pp. 463-482, 2013.
[21] M. Lynch and R. Ennis, “Resource availability, maternal effects,
and longevity,” Experimental Gerontology, vol. 18, no. 2, pp. 147-165, 1983.
[22] M. Santos, M. G. G. Melão, and A. T Lombardi, “Life history
characteristics and production of ceriodaphnia silvestrii Daday (Crustacea, Cladocera) under different experimental conditions,”
Acta Limnologica Brasiliensia, vol. 18, no. 3, pp. 199-212, 2006.
[23] N. M. Korovchinsky, “Cladocera (Crustacea: Branchiopoda) of South East Asia: History of exploration, taxon richness and notes
on zoogeography,” Journal of Limnology, vol. 72, no. 2, pp. 109-
124, 2013. [24] O. Farhadian, M. H. Khanjani, Y. Keivany, and E. Ebrahimi,
“Culture experiments with a freshwater cladoceran, ceriodaphnia
quadrangula (Of Müller, 1785), as suitable live food for Mayan cichlid (Cichlasoma urophthalmus) larvae,” Brazilian Journal of
Aquatic Science and Technology, vol. 16, no. 2, pp. 1-11, 2012.
[25] P. I. Leonardi, C. A. Popovich, and M. C. Damiani, “Feedstocks for second-generation biodiesel: Microalgae’s biology and oil
composition,” Economic Effects of Biofuel Production, no 16, pp.
317-346, 2011. [26] R. De Bernardi, G. Giussani, and M. Manca, “Cladocera:
Predators and prey,” Hydrobiologia, vol. 145, no. 1, pp. 225-243,
1987. [27] R. Fernandez, S. Nandini, and S. S. S. Sarma, “A comparative
study on the ability of tropical micro-crustaceans to feed and grow
on cyanobacterial diets,” Journal of plankton research, vol. 34, no. 8, pp. 719-731, 2012.
[28] R. Kumar and J. S. Hwang, “Ontogenetic shifts in the ability of the cladoceran, moina macrocopa straus and ceriodaphnia cornuta
sars to utilize ciliated protists as food source,” International
Review of Hydrobiology, vol. 93, no. 3, pp. 284-296, 2008.
[29] R. P. Lim, “Population changes of some aquatic invertebrates in
ricefields” Tropical Ecology and Development, pp. 971-980, 1980.
[30] S. Jun and D. Liu, “Geometric models for calculating cell biovolume and surface area for phytoplankton,” Journal of
Plankton Research, vol. 25, no. 11, pp. 1331-1346, 2003.
[31] T. Zaret, “Predators, invisible prey, and the nature of polymorphism in the cladocera (Class Crustacea),” Limnology and
Oceanography, vol. 17, no. 17, pp. 171-184, 1972.
[32] U. Koch, E. Von Elert, and D. Straile, “Food quality triggers the reproductive mode in the cyclical parthenogen daphnia
(Cladocera),” Oecologia, vol. 159, no. 2, pp. 317-324, 2009.
[33] U. Sommer and H. Stibor, “Copepoda–Cladocera–Tunicata: The role of three major mesozooplankton groups in pelagic food
webs,” Ecological Research, vol. 17, no. 2, pp. 161-174, 2002.
[34] V. Ördög, W. A. Stirk, P. Bálint, C. Lovász, O. Pulz, and J. van Staden, “Lipid productivity and fatty acid composition in chlorella
and scenepdesmus strains grown in nitrogen-stressed conditions,”
Journal of Applied Phycology, vol. 25, no. 1, pp. 233-243, 2013. [35] W. R. DeMott, “Discrimination between algae and detritus by
freshwater and marine zooplankton,” Bulletin of Marine Science,
vol. 43, no. 3, pp. 486-499, 1988. [36] Y. Carotenuto, T. Wichard, G. Pohnert, and W. Lampert, “Life-
history responses of daphnia pulicaria to diets containing
freshwater diatoms: Effects of nutritional quality versus polyunsaturated aldehydes,” Limnology and Oceanography, vol.
50, no. 2, pp. 449-454, 2005.
[37] Z. H. He, J. G. Qin, Y. Wang, H. Jiang, and Z. Wen, “Biology of moina mongolica (Moinidae, Cladocera) and perspective as live
food for marine fish larvae: review,” Hydrobiologia, vol. 457, no.
1-3, pp. 25-37, 2001.
532014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014

Hasnun Nita Ismail was born in the quiet
area in Perak, Malaysia on 31 August 1970.
She graduated from Universiti Malaya Kuala
Lumpur for her BSc (Entomology Toxicity) and MSc (Limnology & Aquatic Toxicity) in
1994 and 2004 respectively. She pursued for
her PhD (Zooplankton Ecology) at Flinders University, South Australia and had graduated
in 2010.
Previously, she has been under training as the Research Officer at Institute of Medical Research. Presently, she
attached with Universiti Teknologi MARA (Perak) as a SENIOR
LECTURER and involved in teaching various levels of student from diploma, first degree until post graduates.
Dr Hasnun Nita Ismail has been acting as the Chief-Editor for the
university journal publication and had been appointed as invited reviewer for many national and international journal articles. She has
produced several national and international publications as a book
chapter, academic magazines including journal articles in impact factor journals such as Hydrobiologia, Journal of Plankton Research and
Journal of Experimental Marine Biology and Ecology. Her field of
interests cover the water quality studies and plankton ecology.
Umi Wahidah Ahmad Dini was born in
the
small town of Malaysia, Selangor on 19
January 1992. She just finiished her study at
Universiti Teknologi MARA (Perlis) for her BSc (Zooplankton Biology). She is now
looking forward to pursue for her MSc in the
same field of interest.
Chia Chay Tay was born in the smoothest
state of Malaysia, Johor on 11 November 1978. She graduated from Universiti
Kebangsaan Malaysia for her BSc (Hons) in
marine sciences and MSc. in environmental sciences in 2002 and 2005. respectively.
Subsequently, she obtained PhD in
environmental sciences, technology and engineering from Universiti Malaya in 2012.
Currently, she joined Universiti Teknologi
MARA as a SENIOR LECTURER. She has been working as a lecturer in INTEC for AUSMAT and ADFP program. Besides, she has working
experience in multi-national company in global product development
and customer service. She has published her research in ISI and SCOPUS indexed journals including Water Science Technology and
Korean Journal of Chemical Engineering. Her research interests include
biosorption, environmental management and pollution control. Dr. Tay is a professional member of International Water Association
(IWA). She has been awarded several research grants and medals in
research output exhibitions.
542014 Engineering and Technology Publishing
Journal of Life Sciences and Technologies Vol. 2, No. 2, December 2014