biological damage induced by ionizing cosmic...
TRANSCRIPT

9
Nucl. Tracks Radiat. Meas., Vol. 17, No. 2, pp. 155-165, 1990 In!. J. Radiat. Appl. Instrum., Part D Printed in Great Britain
d
0735-245x190 $3.00 + .00 Pergamon Press plc
BIOLOGICAL DAMAGE INDUCED BY IONIZING COSMIC RAYS IN DRY AMBIDOPSIS SEEDS*
A. R. KRANZ,~ U. BORK,~ H. BUCKER$ and G. REITZ$ fBotanica1 Institute, Plantgenetics, J. W. Goethe-University, POB 111932, D-6000 FrankfurtIMain,
F.R.G. and $Institute for Aerospace Medicine, Biophysics Division, DLR-German Aerospace Research, KolnlPorz, F.R.G.
(Received 30 October 1989)
Abstract-In September 1987 dry seeds containing embryos of the crucifer plant Arabidopsis tlraliarza (L.) Heynh, were flown in orbit for 13 days on the Kosmos 1887 satellite. The seeds were fixed on CNd detectors and stored in units of Biorack type 110. One unit was exposed inside, another one outside the satellite. The temperature profile of the flown seeds inside the satellite was simulated on earth in an identical backup control sample (BC). An additional control (SC) was studied with the original seeds sample. By use of the CNd-detector, HZE-tracks were measured with a PC-assisted microscope. The biological damages were investigated by growing the seeds under controlled climatic conditions. The following biological endpoints of the cosmic radiation damage were studied: germination, radicle length, sublethality, morphological aberrations, flower development, tumorization, embryo lethality inside the siliques. The summarized damage (D) and the mutation frequencies of embyronic lethal genes were calculated. The following results were obtained: the damages increase significantly in orbit at all biological endpoints; germination and flowerings especially, as well as embryo lethality of fruits and lethal mutation frequency, were maximum mostly for HZE-hit seeds. Additionally, an increase of damage was observed for the seeds of the outside-exposed Biorack in comparison to the inside ones, which was probably caused by less radiation shielding and free space vacuum. The significance of the results obtained is discussed with respect to stress and risk and, thus, the quality of the RBE-factors and heavy ionizing radiation all needed for the very definition of radiation protection standards in space.
1. INTRODUCTION
FROM THE very beginning of space flights, cosmic ray studies related to energy transfer were focused on biological target systems, i.e. spores, eggs and seeds (Bucker et al., 1973; Ivanov, 1974). This was essential since densely ionizing heavy ions, the so-called HZE- particles, were a new and particular component of radiation in orbit, to which cells and organisms had never been exposed before during their evolution on
) earth. Thus, less than two decades ago, experience on the biological effect of this component of cosmic rays was nearly nil.
In the meantime, several space flight experiments have been executed with the experimental equipment of Biostack (Biicker et al., 1984) and Biobloc (Gaubin et al., 1983) which consisted of variant stacks of sandwiched detector sheets enclosing single layers of different biological objects between the sheets. In this way microscopical measurement of the nuclear tracks and the biological target region was performed by computer-aided x/y/z-graphs in order to correlate the nuclear particle parameters (2, LET, energy) with
well as of fertilized animal eggs and dormant plant seeds, was observed. With the availability of a 1 g- centrifuge in the U.S. Spacelab Shuttle, effects of HZE-particles and of microgravity, and the com- bined effects, were shown by use of locust Carattsius cells flown during the German Dl-Mission (Bucker et al., 1986). The proper interpretation of the space data is only possible if these data are compared to control experiments on Earth, including the evaluation of samples irradiated with heavy ions at accelerators. Obviously, recent availability of both experimental conditions was very rare and, therefore, not intended with the experiment described below.
In this study results of an advanced Biostack experiment performed with outside and inside exposed units of Biokosmos 8 aboard the Kosmos 1887 biosatellite are presented. Since no 1 g- centrifuge was available, only combined effects of microgravity, cosmic ionizing radiation, and/or space vacuum on dry seeds of Arabidopsis were investigated.
The aims of this study are focused on the following:
the biological effect. (1) Improvement of the correlation between physi- Severe damage, mainly expressed by the reduced cal data of densely ionizing HZE-particles and the
survival of HZE-hit bacterial and fungus spores, as biological endpoints of damage.
*This study is dedicated to Prof. Dr E. Schopper on the occasion of his 80th birthday.
NT 1712-F 155

156 A. R. KRANZ et al.
(2) Presentation of detailed information on the 2.3. Storaae conditions type, spectrum and specificity of the damage.
(3) Gathering of data which can contribute to an estimation of the quality factor(s) of sparsely as well as densely ionizing radiation in space.
(4) Investigation of early and late damage effects induced in orbit on a living model system which will allow the evaluation of the biological risk and stress
., All samples were stored during pre- and post-flight
preparation at the laboratory in Frankfurt, F.R.G., in an exsiccator equipped with silica gel. Silica gel absorber was used to achieve a dry and pure atmo- sphere. The exsiccator was stored in the dark at room temperature ( + 22°C & 3°C).
of the space environment-both essential for the 2,4. Flight harhore and biological objects future evaluation of radiation protection standards in mace. The flight hardware consisted of standard Biorack
containers of type I/O and type 1110. Cellulose nitrate foils (CNd Daicel) with a thickness of 250 pm were
2. MATERIALS AND METHODS
2.1. Flight conditions
In performing the Biokosmos 8 mission, the Kosmos 1887 biosatellite was launched on 29 September 1987 from the space flight center Baikonur, U.S.S.R. All experiments were integrated in the satellite 2 days before launch under tempera- ture-controlled conditions. Initial orbital parameters were: duration 90.5 min, apogee 406 km, perigee 224 km and inclination 62.8". The mission was stopped on the 13th day. Due to a deviation from the intended landing site, the satellite was exposed to a temperature of -25°C for about 10 min upon return to Earth. After this time the satellite was stored at a temperature range of +25-+30°C in an artificial house. Recovery of all of the material on board took one day. The temperature during the transport from landing place to IBMP (Moscow) was never below + 10°C.
2.2. Dosimetric conditions
The dosimetric conditions are shown in Table 1. In total only about 10 of approximately 1000 seeds could be hit by HZE-particles (LETco > 1.7 GeV cm-I), which coresponds to the registration threshold of cellulose nitrate Daicel (CNd).
used as carriers for the biological objects and as plastic detectors for highly energetic and highly charged particles with Z 2 6(HZE-particles) of cos- mic radiation.
Biological objects were dry seeds (embryos) of the crucifer plant (Arabidopsis thaliana (L.) Heynh). Two samples of ecotypes and one mutant line obtained from Arabidopsis Information Service seed bank, Frankfurt, F.R.G. (AIS seed bank, Frankfurt, access No. in parentheses) were used: En-2 (1982), Col-0 (1986), ch2 (1989). The dry seeds were glued on the CNd-foils using the adhesive Luviskol VA64 (10 g in 100 ml of ethanol 50%). Samples size for each type was 5000 seeds at a minimum.
One Biorack-unit (type 110) containing seeds of En-2 only was exposed to free space (in the following, called "outside"). The other one (type 1110) was placed inside the satellite.
2.5. Hardware for nzeasurements
The following hardware was used for the determi- nation of the distance between a nuclear track and the biological object (i.e. impact parameter). A Leitz Orthoplan microscope equipped with a stepmotor driven table was used for visual recognition and measurement of tracks and biological objects. The position of the table (x-, y- and z-coordinates) was measured by a table controller-unit with a precision
Table 1. Dosimetric conditions of the Biokosmos 8 experiment with Arabidopsis seeds - 1 Exposure Events per seed
Radiation inside outside and day approx.
HZE with LET, > 1.7 GeV cm-I 0.38 p ~ m - ~ d-I - 0.5 x lo-' I
(threshold CNd)
HZE with LET, > 1.7 GeVcm-I 0.24 pcm-'d-' 0.87 p ~ m - ~ d-l 0.3 rsp.l.0 x lo-' and 2 2 8
HZE with LET, > 1.3 GEV cm-I 11.8 pcm-'d-I - 15 x lo-' (threshold) CNK
Protons, electrons 290 pGy d- ' 0.41.8 mGy d-' - y -radiation
Fast neutrons 2 x 105pcm-2d-i - 250 = 12 pGy d-l
Disintegration - 2000 -5000 0.1 stars stars cm-3 d-'

IONIZING COSMIC RAYS 157
of 0.15 pm for the x- and y-axis and 0.10 pm for the (4) BC backup control: not flown seeds fastened z-axis. The corresponding data were transferred via on CNd-sheets and stacked, stored by IBMP at an interface to a Victor V120 personal computer. Moscow, used for temperature simulation on Earth; Appropriate self-developed software did the calcu- no = 300, n, = 100 lation of all data as well as the control of the (5) SC seed control: original sample of AIS-seed microscope table. Although the personal computer bank; rz,, = 100, n, = 100. - is not IBM/PC-compatible its operation system is Hit and non-hit seeds were determined on the basis
Version 2.11' The is of computer data for tracks and seeds as described written in Microsoft-BASIC, using Compiler Version
above, These seeds were investigated separately, i.e. 5.32 and Linker Version 1.10. This compiler/linker- the data for the biological endpoints were also regis- combination was used for further programming of tered per plant. Individual evaluation was performed
software as as a 'pecial only for seeds of group H and NH. In the case of interpreter called MegaBasic Version 5.273. Output groups F, BC and SC, a representative amount of of the programs was printed on a Siemens PT88 seed was evaluated by sample. inkjet printer.
2.6. Graphic representation of data and results
All measured and estimated data obtained by use of the personal computer were printed either for graphics using screen hardcopy software or by normal sequential printing of the inkjet printer. Additional graphic representation was performed using PC/AT-compatible personal computers with appropriate printers and software.
2.7. PostTfliglzt actiuities
After etching, CNd-sheets showed the tracks of densely ionizing particles (Z 2 6). For the evaluation all the HZE-tracks with a distance smaller than 1 mm to the biological objects and the corresponding seeds were measured individually. Based on the as- sumption that the oval seed shape approximately equals an ellipsoid, the position of the four points defining the long and the short axes were measured up to four times. The mean values and standard deviation were used for the calculation of the pos- itions of the seed center and the embryonic meristems of the shoot and radicle. Up to ten points were measured for each track and from these the three- dimensional regression was calculated. These data enabled us to decide by graphic interpretation whether a seed was really hit by the respective HZE- particle or not. Measured seeds were removed from CNd-sheets with a moistened brush and stored sep- arately in Eppendorf tubes under the same conditions as mentioned above.
With respect to different treatments of the exper- iment the seed samples were arranged in classes abbreviated by letters as follows. Small letters i and o following the abbreviation letters stand for inside and outside exposure (in the following called "insiders" and "outsiders" respectively):
(1) H hit seeds: measured seeds hit by the HZE- particles; no = 23, 17, = 7
(2) NH non-hit seeds: seeds measured non-hit by the HZE-particles as defined above; 11, = 156, n, = 53
(3) F flight sample: flown seeds from CNd-sheets without nearby HZE-tracks and therefore not measured; n, = 300, n, = 100
2.8. Culture conditions
All seeds were sown under sterile conditions in petri dishes (diameter 90mm) on GELRITE- medium, adapted to mineral and nutrient conditions (composition per liter distilled water: NH,NO, 200 mg; MgSO, x 7H, 0 500 mg; CaHPO, x 2H,O 100 mg; KH2P0, 100 mg; K2HP04 50 mg; FeC,H,O, x 3H,O (ferricitrate) 2.5 mg; HCI- hydrolysate 1 g; dextrose 20 g; GELRITE Gellan Gum (SCOTT Laboratories, Inc., Carson, CA 90746, U.S.A.) 5 g; pH 6.2-6.5). Seeds were vernalized for 120 h at + 4 & 2°C. Afterwards the petri dishes were transferred to a growth cabinet (light: fluorescent lamps 113 Osram Fluora, 213 Osram Daylight 5000, C z 54 x 1018 Q m-2 s-I, 16L/8D, temperature + 25 k l0C, moisture 75 &- 5% r.h.). Seedlings were kept there until transfer to soil. Six days after germi- nation the seedlings were transferred to plastic pots with standard soil (loam or compost/peat/sand 8: 2: 1 v/v/v). The seedlings of the H- and NH-group were single set into separate pots (4 5 cm), while seedlings of the F-, BC- and SC-groups were planted in groups of 25 plants/pot (9 cm x 9 cm). Further growing of the plants took place in a growth chamber (light: fluorescent + Hg-high pressure lamps, C z 136.7 x Q m-2 s-I, 16L/8D; temperature 22 i 2°C; moisture 70 IfI 10% r.h.).
2.9. Determination and measurement of damage end- points
One day after vernalization, control of seed germi- nation was started at intervals of one day under a microscope. The criterion of germination was the emergence of the radicle. Seeds showing no germi- nation after 33 days were called "lethal". Radicle length was measured on the third day after germi- nation with the microscope equipped for micro- positioning. For a period of 11 days we noted the number of plants flowering on a given date. Thus, flowering is estimated on the basis of days from sowing to appearance of the first open flower per plant.
For the test of embryonic sterility of the fruits (Miiller, 1963) five consecutive siliques from the basal
158 A. R. KRANZ et al.
part of the main inflorescence, three from the inside and five from the outside sample, were evaluated. We carefully opened the siliques under a stereo- microscope and counted the number of normal seeds, shrunken seeds, albinos and rudiments ( = lethal embryos). From these data the frequencies of lethal mutations m,, m, and m, (Redei, 1969) were estimated.
2.10. Statistical tests
For statistical tests of different frequencies the &-test (Sokal and Rohlf, 1987) was used, which is a t-test adapted to data rates of percentage values. In case of absolute values (size of siliques, radicle length) the normal I-test for difference of means was used. Furthermore, we used Wilcoxon's signed-rank test (Sokal and Rohlf, 1987) for comparing the germi- nation development between the definitely hit and non-hit seeds. The significance is indicated by differ- ent symbols. The triple symbol stands for the signifi- cance of having different samples at more than 99.9% probability, the double symbol stands for more than 9976, and the single symbol for more than 95% probability.
3. RESULTS
3.1. Germination and flowering delay
In order to obtain quick information about the general condition of the bio-objects, soon after flight we tested the dynamics of germination of the seeds placed inside and outside the satellite. The samples flown in space were compared with stacked and non-stacked control seeds stored on Earth. Evidently, flight-samples germinated later and less than the control. Furthermore, frequency of flowering plants
was significantly reduced among the flown seeds (Zimmermann et al., 1988).
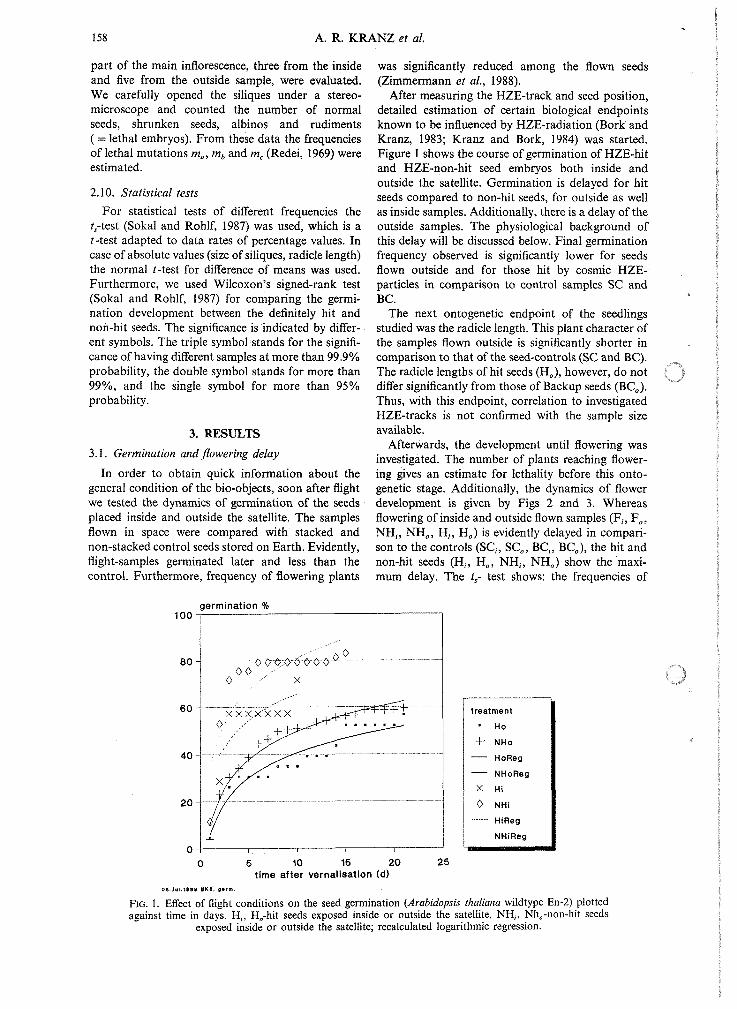
After measuring the HZE-track and seed position, detailed estimation of certain biological endpoints known to be influenced by HZE-radiation (Bork and Kranz, 1983; Kranz and Bork, 1984) was started. Figure 1 shows the course of germination of HZE-hit and HZE-non-hit seed embryos both inside and outside the satellite. Germination is delayed for hit seeds compared to non-hit seeds, for outside as well as inside samples. Additionally, there is a delay of the outside samples. The physiological background of this delay will be discussed below. Final germination frequency observed is significantly lower for seeds flown outside and for those hit by cosmic HZE- particles in comparison to control samples SC and BC.
The next ontogenetic endpoint of the seedlings studied was the radicle length. This plant character of the samples flown outside is significantly shorter in comparison to that of the seed-controls (SC and BC). The radicle lengths of hit seeds (H,), however, do not '1
8
differ significantly from those of Backup seeds (BC,). Thus, with this endpoint, correlation to investigated HZE-tracks is not confirmed with the sample size available.
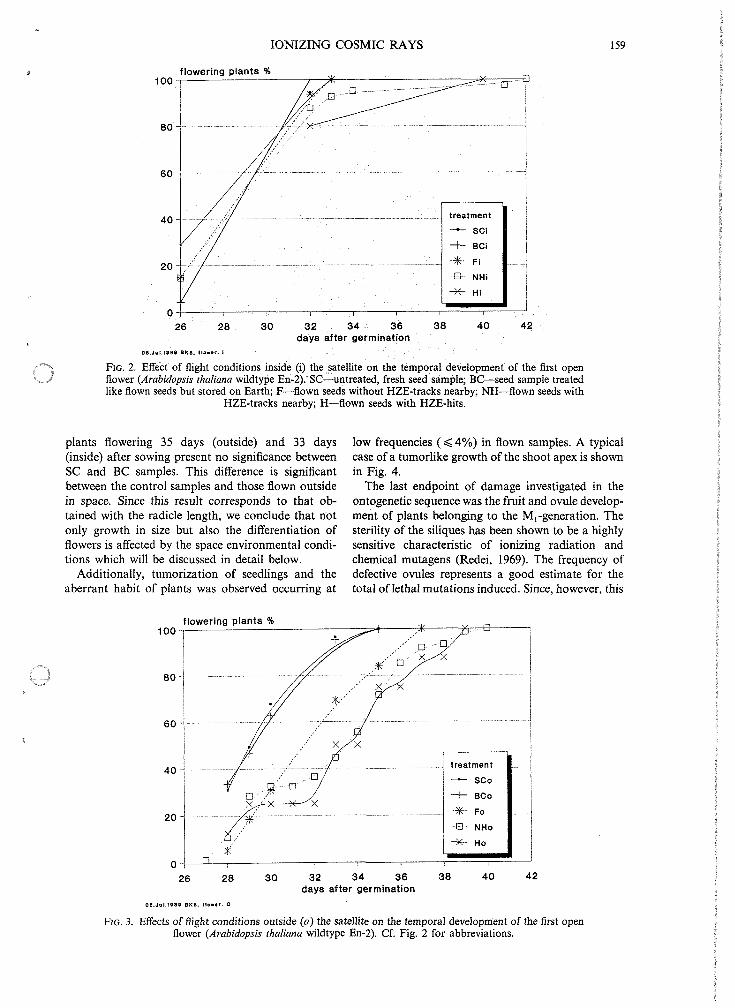
Afterwards, the development until flowering was investigated. The number of plants reaching flower- ing gives an estimate for lethality before this onto- genetic stage. Additionally, the dynamics of flower development is given by Figs 2 and 3. Whereas flowering of inside and outside flown samples (F,, Fa, NH,, NH,, H,, H,) is evidently delayed in compari- son to the controls (SC,, SC,, BC,, BC,), the hit and non-hit seeds (H,, H,, NH,, NH,) show the maxi- mum delay. The t,- test shows: the frequencies of
0 5 10 15 20 25 time after vernalisation (d)
FIG. 1. Effect of flight conditions on the seed germination (Arabidopsis thaliana wildtype En-2) plotted against time in days. H i , H,-hit seeds exposed inside or outside the satellite, NH,, Nh,-non-hit seeds
exposed inside or outside the satellite; recalculated logarithmic regression.
treatment
" Ho
+ NHo - HoReg
- NHoReg
X Hi
0 NHi
HiReg
NHiReg
I
IONIZING COSMIC RAYS
- --
- - - -
26 28 30 32 34 36 38 40 42 days after germination
06 JUI 1089 B I ~ . 1lox.r I
FIG. 2. Effect of flight conditions inside (i) the satellite on the temporal development of the first open flower (Arabidopsis thaliana wildtype En-2). SC-untreated, fresh seed sample; BC--seed sample treated like flown seeds but stored on Earth; F-flown seeds without HZE-tracks nearby; NH-flown seeds with
HZE-tracks nearby; H-flown seeds with HZE-hits.
plants flowering 35 days (outside) and 33 days (inside) after sowing present no significance between SC and BC samples. This difference is significant between the control samples and those flown outside in space. Since this result corresponds to that ob- tained with the radicle length, we conclude that not only growth in size but also the differentiation of flowers is affected by the space environmental condi- tions which will be discussed in detail below.
Additionally, tumorization of seedlings and the aberrant habit of plants was observed occurring at
low frequencies (< 4%) in flown samples. A typical case of a tumorlike growth of the shoot apex is shown in Fig. 4.
The last endpoint of damage investigated in the ontogenetic sequence was the fruit and ovule develop- ment of plants belonging to the M,-generation. The sterility of the siliques has been shown to be a highly sensitive characteristic of ionizing radiation and chemical mutagens (Redei, 1969). The frequency of defective ovules represents a good estimate for the total of lethal mutations induced. Since, however, this
26 28 30 32 34 36 38 40 42 days after germination
FIG. 3. Effects of flight conditions outside ( 0 ) the satellite on the temporal development of the first open flower (Arabidopsis thaliana wildtype En-2). Cf. Fig. 2 for abbreviations.

A. R. KRANZ et al.
FIG. .4 Normal seedling (left) and an aberrant seedling with tumor-like growth of the shoot (right) found in the cosmic HZE-hit seed sample (H,) of Arabidopsis thaliana wildtype En-2. Bar 1.5 mm.
is a quantitative value influenced by non-controlled environmental factors during plant culture, e.g. nutri- ent and water supply, the results must be interpreted with care.
Under control conditions, i.e. when irradiation is missing, the different amounts of defective ovules, which are obtained in a silique, will follow a typical Poisson-distribution of probabilities for fruit sterility. However, the different samples for "insiders" and "outsiders" (Figs 5 and 6 ) show that fruits with
higher sterility, i.e. embryonic lethality, are signifi- cantly more frequent in the flown samples (F, NH, H) than in the controls (SC, BC), except for the sample of BC, which will be discussed below.
The reason we present the frequencies in bar charts (Figs 5 and 6) is based on a genetic evaluation of this plant character. According to Mendelian ratios ob- tained after recombination, a mutation to a recessive lethal gene will produce 25 + 12.5% defective ovules in a fruit, and mutation to a dominant lethal gene
pel
all
rcentage
siliques
SCi BCi F i NHi Hi treatment
lU .JU I . IS8S BKS. ~ ler l l . . l l iq . I
lethal embr
per silique
FIG. 5. Effect of flight conditions inside (i) the satellite on the frequency distribution (% of all siliques) of lethal embryos per silique (Arabidopsis thaliarza wildtype En-2). SC-untreated, fresh seed sample; BC-seed sample treated like flown seeds but stored on earth; F-flown seeds without HZE-tracks nearby;
NH-flown seeds with HZE-tracks nearby; H-flown seeds with HZE-hits.

IONIZING COSMIC RAYS
pel all
lethal embryos
per silique
SCo BCo Fo NHo Ho treatment
L IO.Ju1.198P B X I . .I.rll.slllQ 0
\, FIG. 6. Effect of flight conditions outside ( 0 ) the satellite on the frequency distribution (% of all siliques)
_ 1 of lethal embryos per silique (Arabidopsis thaliana wildtype En-2). Cf. Fig. 5 for abbreviations.
75 f 12.5%. If, however, only one gamete contributes Cf',, ma=- to the production of a lethal ovule through a domi- n',,,' nant gene mutation, about 50 Ifr 12.5% sterility is obtained. If energy deposition and thus ionization of the DNA molecule increases, double strand breakage of this molecule can be expected preferen- tially, which produces the highest class of sterility (87.5-100%). Figures 5 and 6 show very clearly the increase of defective ovule production in the flown samples (F ,,,, NH,,,, H,,,) with bars represent- ing the highest sterility classes. Among the control samples (SC,,,, BC,,,), evidently, the bars of the outside backup-control (BC,) are runaways because frequencies of 12.5-62.5% are over-represented to the debit of the lowest frequencies (0.0-12.5%). This deviation comes from certain sheets of this sample which were stored during the mission period in a vacuum chamber at room temperature in order to simulate the space vacuum on Earth. Since the out- side conditions of the satellite in orbit were not recorded, simulation with the BC, sample was incom- plete.
Nevertheless, comparison between flown "insid- ers" and "outsiders" shows in total higher sterility of the outside exposed sample where the HZE-particle flux was stronger; all bars representing frequencies probably influenced by an increase of lethal mutations ( a 12.5%) are larger. The background of this late biophysical effect will be discussed below.
A popular method of estimating the consequences of mutagenic treatments in Arabidopsis is that of the so-called embryo test mentioned above. The test has been used for the estimation of three different though related parameters of lethal mutants (Redei, 1969). It is not really an inheritance test since embry- onic malformations are affected also by physiological factors.
.".
P,,,: segregating M , plants, i.e. with 2 12.5% lethal embryos PI,,: all the M I plants examined based on 3 or 5 consecutive siliques per M I plant (1)
S,,,: segregating silique progenies, i.e. with 2 12.5% lethal embryos S,,,: all the silique progenies examined based on 3 or 5 siliques per M , plant (2)
El,,: lethal embryos, E,,,: all embryos examined. (3)
The m, formula expresses mutation frequency as the " M , method", whereas m, procedure is very similar to the " M , method". The m, approach gives some information on the chimerical nature of the M I plants with respect to the mutation though loaded with various degrees of errors depending on the method of sampling as well as on the general con- dition of the M I mother plant which is influenced by the culture conditions etc. The bar graphs of Figs 7 and 8 as well as the &-tests presented in Table 2 show the significant increase of the three mutation par- ameters mainly in the inside flown samples NH, and Hi except BC,. Evidently, the m, parameter is highest for the hit seed samples inside and outside exposed (H,,). The radiogenetic conclusion of this result will be discussed below.
The broad, nevertheless particular spectrum of damages obtained by the investigated endpoints

A. R. KRANZ et al.
A
SCi BCi F i NHi Hi t r ea tment
l l .Ju l . IS89 BK8, Uut .Fr .~u. I
FIG. 7. Effect of flight conditions inside (i) the satellite on the mutation frequency of Arabidopsis thaliana wildtype En-2 expressed by n7,, m,, m, (formulae 1-3). SC-untreated, fresh seed sample; BC-seed sample treated like flown seeds but stored on Earth; F-flown seeds without HZE-tracks nearby;
NH-flown seeds with HZE-tracks nearby; H-flown seeds with HZE-hits.
argues for summarizing the effects of the space environment as well as those of the cosmic heavy ion hits. In order to characterize the whole damage induced, the total damage parameter D given by the formula
was defined (Bork et al., 1986); D was obtained by classifying and summarizing all the biological effects observed in the M,-generation. However, for biological reasons, a better estimate of D is
obtained if the single damages are weighted by certain factors as follows: dl = 1 for curved radicle or hypocotyl, defective cotyledon, d, = 2 for ageotropism, d, = 3 for tumor, d, = 7 for sublethal- ity, and d, = 10 for lethality in which n, = number of damaged plants, no = number of examined plants. The factors have been estimated with regard to the assumed damage degree induced in the number of DNA strand breakages which control the defective plant characters. Data obtained are shown by bar graphs of Fig. 9. Evidently, D is estimated maximum for hit seeds
SCo BCo Fo NHo H o t r ea tment
FIG. 8. Effect of flight conditions outside (0) the satellite on the mutation frequency Arabidopsis tkaliana wildtype En-2 expressed by ma, m,, m, (formulae 1-3). Cf. Fig. 7 for abbreviations.

IONIZING COSMIC RAYS 163
Table 2. Mutation frequencies of lethal embryos in M, generation inside and outside exposed Arabidopsis seeds
"Insiders" SC, BC, Fi NH, H;
"Outsiders" s c "
+ + + , + + , + : significance level of t, compared with Fi, F, x x x significance level of t, compared with BC,, BC,
*** significance level of t, compared with NH,, NH,
exposed outside the satellite (H,) , and also for hit embryo with developed radicle, two cotyledons seeds among the "insider" samples (H,). and a shoot apex. During germination its growth
will proceed from two meristems, one for the root
4. DISCUSSION AND CONCLUSION and bne for the shoot, which consists of only 2-4, and 8-10 initial cells respectively, forming the
The test subject used for this study, the dry seed stem cells and, thus, the main sensitive target sites of Arabidopsis thaliana, contains a complete plant for the ionizing radiation. Additionally, significant
summarized damage D 700
treatment
m sc BC
E2 F
NH @g
Outside Inside
FIG. 9. Calculated summarized damage (D) (formula 4) for the different treatments of Arabidopsis thaliana wildtype En-2 seeds inside or outside the Biokosmos 8 satellite. Cf. Fig. 7 for abbreviations.

advantages for using Arabidopsis seeds in space stud- ies (Ivanov, 1974; Kranz, 1986) are:
(i) smallness at every level of biological organiz- ation (plant, seed, embryo, meristem, initial cell, cell nuclei and organelles, chromosomes, DNA per genome and gene);
(ii) large genetic variability available in well- defined classical and physical gene maps;
(iii) reconlmended test subject of radiation and chemical mutagenicity tests, and model plant of mutation and molecular genetics;
(iv) comparatively high resistance to ionizing radiation investigated for several decades.
In our Biokosmos 8 experiment, the increase of damage found for early and late biological endpoints is evident. The damage increases in about all the investigated biological endpoints from control to flown specimens; within the latter, damage induced is maximum mostly for HZE-hit seeds. We find a similar spectrum of harmful effects induced in inside and outside exposed seeds, but an increase of damage with the outside exposure, i.e. with reduced shielding and vacuum. Comparison with the results obtained from an earlier experiment performed during the SL-I mission, even though different in flight con- ditions and material exposure, shows similar tenden- cies of damage (Kranz, 1986).
The determination of the impact parameter of cosmic HZE-tracks to the biological subject (center of the seed embryo) is significantly improved in comparison with the data obtained in the earlier experiments of the SL-1 mission (Kranz, 1986); how- ever, more precise estimates of this kind to the radicle and shoot meristem (stem cells) failed, because the sample size obtained was too small. The microdosi- metric data of Table 1 reveal that only about 1 HZE hit (Z 2 8) among 100 seeds can be expected for the whole mission of Biokosmos 8. Therefore, space flight experiments of longer duration (e.g. ERA 1, LDEF) are needed in future to overcome the sample size problem.
Information on the damage endpoints is enhanced in quality and quantity. Besides the less frequent malformations of tumorlike growth, the aberrant habit of plants in M, generation, the defects in germination and growth (flowering) are affected sig- nificantly by the detected HZE-particles (Figs 1, 2 and 3). Since, however, the plant growth characters are also affected for flown but non-hit seeds, addi- tional effects by factors of the space environment have to be considered. Protons, fast neutrons, and disintegration stars (cf. Table 1) form a source for added radiation damage. Single events of this type of radiation can only be measured with the detectors of nuclear emulsion and of silver halide type presently improved for future missions. Furthermore, micro- gravity of the space environment is known to be harmful for seedling growth (Volkman et al., 1986); its effect, however, can only be separated if a I g-
.ANZ et al.
centrifuge is available which was not the case for this *
Biokosmos 8 experiment. Decrease in growth is probably caused by stress
and aberrant control of phytohormones, since for instance gibberellic acid is able to reduce germination decrease induced by X-rays (Reinholz, 1967). This is an interesting aspect which has to be studied particularly in future spaceflight and accelerator experiments.
A genetic approach to the deleterious effect of the cosmic HZE-radiation is presented by the result of the fruit sterility and the estimation of lethal mutation frequencies. This defect is really produced by lethal genes after meiotic recombination in the second generation (M,) and thus it is a very late effect of ionizing radiation. Damage effects of this kind induced by cosmic HZE-particles have not yet been studied with other biological objects, showing once more the exceptional utility of Arabidopsis in space. The observed increase of higher sterility of the flown specimens (Figs 5 and 6) proves the stay in space to 3 be genetically risky for the plant species. Outside * exposure is more critical than inside exposing of seeds because of less shielding and the vacuum condition of the first (Figs 7 and 8).
Characterization of the total damage induced in space by introducing the parameter D which classifies the sensitive biological effects in the first generation M, is a biophysical approach to the preliminary estimation of the quality factor(s) of densely ionizing radiation in space. The weight of the damage criteria used in this study can only be preliminary as long as better differentiation between developmental i.e. physiological stress, and genetic, i.e. DNA-damage risk, will be achieved. Nevertheless, the results of our calculations presented by Fig. 9 are a good step towards this.
Furthermore, the nuclear physical bases of the primary heavy ion effect in the biological target matter has to be considered in more detail. Our experiments with specific heavy ions accelerated by UNILAC to 10-20 MeV u-', which is significantly 1 lower than those in space, have shown (Bork et al., 9
1986, 1989) that the damage effect in quality and quantity depends on the charge, particle fluence and energy. If dose is considered, the dependence of the radiation effect on the particular properties of the HZE-particles is ambiguous. This indicates that for the dose effect particular RBE-factors exist for every ion and not a single factor for all of them. The very dense ionization caused by cosmic heavy ions extends, however, to such large values including the production of secondary electrons where both the spatial and temporal pattern of energy deposition become extremely inhomogeneous, that the very definition of absorbed dose as a measure of radiation exposure and also the concept of the quality factor for ionizing radiation in general becomes meaning- less. A unified theory of the HZE-radiation effect especially with reference to the produced secondary

IONIZING COSMIC RAYS 165
D electrons has yet to be achieved (Chadwick and Leenhouts, 1981; Kraft, 1988; Kiefer and Straaten, 1989).
Acknowledgements-Thanks are due to the following people: E. Schopper, who has contributed sound ideas through his professional competence and enthusiasm; to J. U. Schott for taking care of the experimental hardware at Moscow before launch and after retrieval; to Brigitte Kirchheim and the student team, Monika Pickert, Sabine Sommer, and M. Zimmermann, for pre-flight preparation of the biological subjects and their post-flight investigations. The production of the manuscript is attributable to the cooperative activity of the whole group of investigators mentioned above.
We also wish to thank our colleagues at IBMP, Moscow, who made the experiment successful by their helpful tech- nical assistance, and the colleagues at DLR, Koln and ESA,
i Paris, who were responsible for the project management.
This study was supported by the German Ministry of Research and Technology, Bonn (Grant No. 01 QV 85650).
REFERENCES
Bork U., Gartenbach K., Koch C. and Kranz A. R. (1986) Biological effects of heavy ions in Arabidopsis seeds. Ado. Space Res. 6, 149-152.
Bork U., Gartenbach K. and Kranz A. R. (1989) Early and late damages induced by heavy charged particle irradi- ation in embryonic tissue of Arabidopsis seeds. Proc. XXVZZth COSPAR Mtg, Espoo, Finland, 1988, Adv. Space Res. 9, 1 17-12 1.
Bork U. and Kranz A. R. (1983) A computerized system for microdosimetric measurements of HZE-particles with seeds and spores. Arabidopsis It$ Seru. 20, 19-26.
Bucker H., Facius R., Horneck G., Reitz G., Graul E. H., Berger H., HofFken H., Ruther W., Heinrich W., Beaujean R. and Enge W. (1986) Embryogenesis and organogenesis of Carausius ntorosus under spaceflight conditions. Adu. Space Res. 6, 115-124.
Bucker H., Horneck G., Facius R., Reitz G., Schafer M., Schott J. U., Beaujean R., Enge W., Schopper E., Heinrich H., Beer J., Wiegel B., Pfohl R., Francois H., Portal G., Bonting S. L., Graul H., Ruther W., Kranz A. R., Bork U., Koller-Lampert K., Kirchheim B., Starke M. E., Planel H. and Delpoux M. (1984) Radiobiological advanced Biostack equipment. Science 225, 222-224.
Bucker H., Horneck G. and Hildebrand D. (1973) Effects of individual HZE-particles in the BIOSTACK exper- iment. Proc. 4th Symp. Microdosim., pp. 10-71. Euratom EUR. 51222 d-e-f, September 1973.
Chadwick K. H. and Leenhouts H. P. (1981) The Molecular Tlteory of Radiation Biology, pp. 205-215. Springer, Berlin.
Gaubin Y., Planel H., Gasset G., Pianezzi B., Clegg J., Kovalev E. E., Nevzgodina L. V., Maximova E. N., Miiller A. T. and Delpoux M. (1983) Results of Arternia cysts, lettuce and tobacco seeds in the Biobloc 4 experiments flown aboard the Soviet biosatellite Cosmos-1 129. Adv. Space Res. 3, 8-135.
Ivanov V. I. (1974) Radiobiologiya e Getzetika Arabidopsisa, Vol. 27, pp. 1-191. Nauka, Moskva.
Kiefer J. and Straaten H. (1989) Quantitative interpretation of heavy ion effects: models for the biological effects of heavy ions. Proc. XXVZZth COSPAR Mtg, Espoo, Finland, 1988. Adv. Space Res. 9, 8-16.
Kraft G. (1988) On the interpretation of radiobiological experiments performed with heavy charged particles. GSZ 88-33 preprint, pp. 1-21.
Kranz A. R. (1986) Genetic and physiological damage induced by cosmic radiation on dry plant seeds during space flight. Adv. Space Res. 6, 135-138.
Kranz A. R. and Bork U. (1984) Biotests for heavy ion effects and preliminary total evaluation of cosmic radiation damage in Arabidopsis seeds flown during the first mission of Spacelab on STS 9. Arabidopsis Znf: Sew. 21, 3140.
Muller A. (1963) Embryonentest zum Nachweis rezessiver Letalfaktoren bei Arabidopsis thaliana. Biol. Zbl. 83, 133-163.
Redei G. P. (1969) Arabidopsis thaliana (L.) Heynh. A review of the genetics and biology. Bibliogr. Genet. 21, 1-151.
Reinholz G. (1967) The influence of gibberellic acid on the germination of irradiated and non-irradiated Arabidopsis seeds. Arabidopsis I?$ Serv. 4, 16-17.
Sokal R. R. and Rohlf F. J. (1987) Introduction to Biostatistics, pp. 127-129; 225-227. Freeman, New York.
Volkmann D., Behrens H. M. and Sievers A. (1986) Devel- opment and gravity sensing of cress roots under micro- gravity. Naturwisset~schaften 73, 438-441
Zimmermann M., Sommer S., Pickert M., Kranz A. R., Gartenbach K. and Bork U. (1988) Interim results of an experiment on Kosmos 1887 satellite with Arabidopsis seeds exposed in space. Arabidopsis Zt7f: Serv. 26, 47-54.
