biogeochemical assessment of paleoproductivity …g131/dbsubmit.pdf2 biogeochemical assessment of...
TRANSCRIPT

Biogeochemical assessment of paleoproductivity during the Early AptianOceanic Anoxic Event at Shatsky Rise, ODP Leg 198
Mirela Dumitrescu, Simon C. Brassell*Biogeochemical Laboratories, Department of Geological Sciences, Indiana University, Bloomington,
IN 47405-1403, U.S.A.
Abstract
This study focuses on the biogeochemical investigation of organic-rich sedimentary sequences deposited during the early AptianOceanic Anoxic Event (OAE1a) at Sites 1207 and 1213 on Shatsky Rise (ODP Leg 198) in the west-central Pacific. Biomarkeranalyses provide evidence of the algal and bacterial origin of organic matter (OM) from the sediments corresponding to OAE1a. Theabundance of steroidal components, particularly sterenes and sterones, suggests that the OM includes major contributions fromeukaryotic sources in an environment characterized by high phytoplankton productivity. The presence of alkenones at Site 1213B isdiagnostic of OM derived from representatives of haptophyte algae among the calcareous nannoplankton and their δ13C values(average -31.6‰) are consistent with those expected during elevated pCO2. The occurrence and prominence of 2β-methylhopanes and2β-methylhopanones indicates significant contributions to the OM from cyanobacteria, which are also likely contributors of hopanoidsbased on their δ13C compositions. Theses biomarker data suggest oceanic conditions, such as nitrate- or iron-limited conditions,conducive to the prevalence of cyanobacteria during deposition of OAE1a. Comparisons with biomarker distributions associated withother Cretaceous OAE suggest that the importance of cyanobacteria among phytoplankton may represent a critical distinguishingcharacteristic of OAE1a.
1. Introduction
Significant changes took place in the ocean-climate systemduring the mid-Cretaceous, including global warming, massivesubmarine volcanism, formation of oceanic plateaux, andincreased rates of seafloor spreading (e.g., Leckie et al., 2002).In addition, there are discrete intervals of time (typically lessthan one million years) when widespread organic-rich sedimentswere deposited and preserved under oxygen-deficient conditions(Schlanger and Jenkyns, 1976; Leckie et al., 2002). The termOceanic Anoxic Events (OAE) has been used to describe theseepisodes of increased sequestration of organic matter. Two ofthese events are observed on a global scale and the earliest ofthese, designated OAE1a, occurs during the early Aptian.Sediments corresponding to OAE1a were recovered at threelocations, Sites 1207B (37°47.433’N 162°45.053’E, 3101 mwater depth), 1213B (31°34.649’N 157°17.861’E; 3883 m waterdepth) and 1214A (31°52.025’N 157°28.717’E; 3402 m waterdepth), drilled on Shatsky Rise in the west-central Pacific duringOcean Drilling Program (ODP) Leg 198 (ODP Leg 198:Bralower et al., 2002a). The lithology, thickness and continuityof organic-rich units corresponding to OAE1a varies betweenthe sites, but the dominant microfossils in all of them areradiolaria (Bralower et al., 2002a). These sediments offer apelagic record of OAE1a that has rarely been recovered from thePacific (Sliter, 1989), although it is well recognized in theTethyean realm (Menegatti et al., 1998; Hochuli et al., 1999;Luciani et al., 2001; de Gea et al., 2003; Erba and Tremolada,2004).
This contribution provides an assessment of the biomarkercompositions of these cores focused on determination of thesources of the organic matter (OM) accumulated and preserved
in these sediments, and on interpretation of the evidence theyprovide regarding temporal changes in both local and globalenvironmental conditions.
2. Methods
The organic carbon (Corg) content was determined with a CarloErba 1500 CNS analyzer and calculated as difference betweentotal carbon from CNS analysis and the carbonate contentquantified by coulometry (Bralower et al., 2002a).δ13Corg values were determined using a ThermoFinnigan Delta
Plus XP mass-spectrometer connected on-line to a CostechElemental Analyzer. Powdered and weighed sediment sampleswithin foil capsules were placed into the autosampler of theelemental analyzer and combusted. The carbon isotopecomposition of the resulting CO2 gas was calibrated against aninternal laboratory standard (acetanilide, δ13Corg= -24.34‰),expressed relative to the Vienna Peedee Belemnite (VPDB)international standard.
Powdered samples (~2 g) were lyophilized and extractedultrasonically using CH2Cl2 (10 mL) for 30 min. The extract wasreduced to dryness under N2 and eluted through silica in aPasteur pipette using hexane (4 mL) and CH2Cl2 (5 mL) aseluates. The hydrocarbon and ketone fractions resulted weretaken to near dryness under N2 and dissolved in hexane (50µL)for analysis by GC/MSD (Bralower et al., 2002a).
A Hewlett-Packard 6973 GC/MSD system and a FinniganTSQ700 coupled to a HP 5890 GC were employed forbiomarker analyses (shipboard and shorebased, respectively).GC/MSD consists of a HP 6890 GC with a Mass SelectiveDetector (MSD) and a HP 7683 Automatic Liquid Sampler(ALS). The GC is equipped with an EPC split-splitless injectorand an HP capillary column (5% phenyl methyl siloxane; 30m x0.25mm) programmed from 40°C to 130°C at 20°C/min, then at4°C/min to 320°C, and held isothermally at 320°C for 20 min.Helium was used as the carrier gas. The transfer line was set at
* Corresponding author. Tel.: +1-812-855-3786; fax: +1-812-855-7961.
E-mail address: [email protected] (S.C. Brassell).

2 Biogeochemical assessment of paleoproductivity
2
280°C and the source of the MS at 230°C. The MSD wasscanned from m/z 27-550. An HP MS Chemstation was used fordata acquisition and processing (Brassell et al., 2004). Theidentity of individual components was determined from theirmass spectral characteristics and retention times by comparisonwith the literature. The HP 5890 GC was equipped with an on-column injector and a Restek Rtx-1 capillary column (60m x0.32mm) programmed from 60°C to 320°C at 2.5°C/min, andheld isothermally at 320°C for 80 min. Helium is used as thecarrier gas. The transfer line was set at 320°C and the massspectrometer was scanned from m/z 50-600 in 0.8s.
Carbon isotope compositions of individual biomarkers weredetermined by irmGC-MS (Hayes et al., 1990) using a FinniganMAT 252 instrument coupled to a HP 5890 GC via acontinuous-flow combustion interface that combusted andoxidized compounds to CO2. The GC was equipped with an on-column injector and a Chrompack CP-Sil 8 CB capillary column(50m x 0.32mm) programmed from 60°C to 320°C at 3°C/min,and held isothermally at 320°C for 140 min. The massspectrometer measured m/z 44, 45 and 46 and was calibratedwith both external and internal standards using deuterated n-alkanes and CO2 peaks of known isotopic composition,respectively (Brassell et al., 2004).
3. Results
3.1. Lithology/Stratigraphy
The lower Aptian sediments comprise 45 cm of finelylaminated, dark brown radiolarian claystone (Site 1207B at 566mbsf), intermittent organic-rich units that include clayey andradiolarian porcellanites (Site 1213B at 257 mbsf), andradiolarian porcellanite (Site 1214A; at 207 mbsf Bralower etal., 2002a). An early Aptian age for these intervals wasconfirmed by biostratigraphy, which placed the sediments withinZone NC6 (Table 1).
3.2. Bulk geochemistry
Intervals within the lower Aptian sequence corresponding toOAE1a contained high organic carbon contents at both Sites1207B and 1213B (Table 1). The Corg values range from 1.7% to34.7% at Site 1207B, and from 2.87% to 25.2% at Site 1213B(Bralower et al., 2002a; Brassell et al., 2004), accompanied by
sulfur contents that vary from 0.23% to 10.56%. The C/N valuesfor the samples are high, ranging from 20.1 to 35.4 (Table 1).The hydrogen indices of the samples determined by shipboardRock-Eval pyrolysis (Bralower et al., 2002a, b; Table 2) rangefrom 365 to 523 mg HC/g Corg, and the Tmax values fall in therange of 401°C to 422°C. Analysis of the δ13C composition forOM in the three samples from Site 1207B (Table 1) yields alower value (–27.7‰) at the base of the recovered organic-richinterval (103-104 cm) compared to those (–25.7‰, –25.9‰)higher in the section.
3.3. Hydrocarbon Distributions
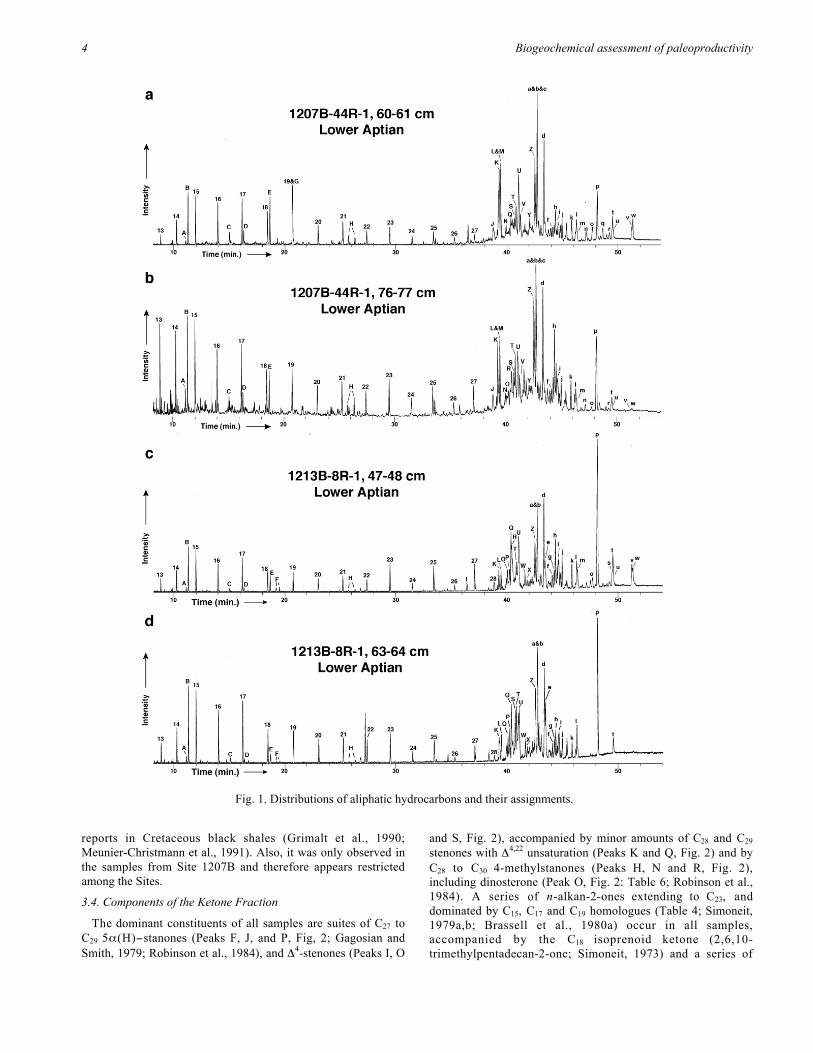
The dominant biomarkers in the organic-rich sediments aresuites of steroidal and hopanoidal alkanes and alkenesaugmented by a prominent series of short-chain n-alkanes (C13 -C19; Fig. 1; Table 3). The distribution patterns of thesecomponents within the individual organic-rich intervals aregenerally similar, as shown by the chromatograms (Fig. 1) andthe consistent dominant compounds within each compound class(Table 4). although their relative abundance varies among thesamples. For example, 17β(H),21β(H)-homohopane (Peak p,Fig. 1; Table 3) is more pronounced within the samples fromSite 1213B and 1214A.
3.3.1. Acyclic alkanesn-Pentadecane (C15) is typically (Table 4) the major n-alkane
(C13 -C19) within a series that ranges to C27+ and exhibits apronounced odd/even preference. Suites of branched alkanes arepresent in sample 1207B-44R-1 76-77cm, but are generallyminor constituents of the samples. The dominant acyclicisoprenoid alkane of all the samples (Table 4) is the C16
component (2,6,10-trimethyltridecane; Peak B, Fig. 1; Table 3).Phytane (Peak E, Fig. 1) is more abundant than pristane (PeakD, Fig. 1), which low Pr/Ph ratios (Table 5). The samples allcontain minor amounts of phytenes (Peaks F, Fig. 1) andphytenyl thiophenes (Peaks H, Fig. 1; Brassell et al., 1986).Lycopane (Peak u, Fig. 1; Brassell et al., 1981; Wakeham et al.,1993) was identified in several samples during shorebasedanalysis aided by an improved separation from its co-elutinghopane, and the regular C30 component (Peak I, Fig. 1c) occursin one sample from Site 1213B.
Table 1
Carbon and carbonate contents of organic-rich sediments from OAE1a at Shatsky Rise.
Interval Nannofossil Ccarb Corg S C/N ratioSample (cm) Lithology Age Zone (%) (%) (%) (atomic) δ13Ca
Site 1207B44R-1 60-61 radiolarian claystone early Aptian NC6 0.01 34.74 10.56 34.1 -25.744R-1 76-77 radiolarian claystone early Aptian NC6 0.00 10.40 1.92 30.3 -25.944R-1 103-104 claystone early Aptian NC6 0.00 1.70 1.30 22.0 -27.7
Site 1213B8R-1 47-48 claystone early Aptian NC6 0.50 10.23 2.00 32.3 n.d.8R-1 63-64 claystone early Aptian NC6 0.75 25.20 4.36 35.4 n.d.8R-1 96-97 porcellanite early Aptian NC6 0.42 2.87 0.76 23.9 n.d.
Site 1214A23R-1 5-7 radiolarian porcellanite early Aptian NC6 0.75 1.38 0.23 20.1 n.d.
n.d. = not determined.a The δ13C values were determined on samples collected from the core repository at the same intervals as those sampled for
the shipboard analyses.

Biogeochemical assessment of paleoproductivity 3
3
Table 2
Rock-Eval results for organic-rich sediments from OAE1a atShatsky Risea.
S1 S2 S3 HI (mg OI (mgInterval peak peak peak Tmax HC/ CO2/
Sample (cm) (mg/g) (mg/g) (mg/g) (°C) g TOC) g TOC)
Site 1207B44R-1 60-61 14.0 147 7.64 401 423 2144R-1 76-77 3.24 54.5 2.69 403 523 2544R-1 103-104 0.49 7.45 0.94 417 438 55
Site 1213B8R-1 47-48 2.54 51.3 2.14 418 501 208R-1 63-64 6.97 128 5.87 404 506 238R-1 96-97 0.39 10.5 6.08 422 365 211
n.d. = not determined.a No sample from Site 1214A was analyzed by Rock-Eval pyrolysis.
3.3.2. Polycyclic alkanes and alkenesThe dominant steroidal hydrocarbons are series of C27-C29 Δ
4-
and Δ5-sterenes (Peaks K, L, Z and a, Fig. 1; Table 4; Rhead etal., 1971; Dastillung and Albrecht, 1977), plus C28 and C29
components with Δ4,22 and Δ5,22 unsaturation (Peaks O, P, W andX, Fig. 1; Table 3; McEvoy and Maxwell, 1983). 5α(H)- and5β(H)-steranes (Peaks J, M, V, Y, and c, Fig. 1) are minorconstituents of samples from Site 1207B (Fig. 1a, b), but aretrace components barely above detection limits in samples fromSite 1213B and 1214A. Samples from Site 1213B containedΔ3,5-steradienes (Peak e, Fig. 1; Gagosian and Farrington, 1978).
The dominant hopane constituents possess 17β(H),21β(H)configurations (Table 4), occurring as a series that extends to C33
(Peaks i, l, p, t and w, Fig. 1). The most abundant hopane in allsamples is 17β(H),21β(H)-homohopane (Peak p, Fig. 1), and itis the dominant hydrocarbon in all the samples from Site 1213B.Minor amounts of 17β(H),21α(H)- and 17α(H),21β(H)-hopanesalso occur in all samples (Fig. 1, Table 3), together with hop-17(21)-enes (C27, C29 to C32; Peaks Q, b, d, j and l, Fig. 1; Barneset al., 1979; Brassell et al., 1980a; ten Haven et al., 1985;McEvoy and Giger, 1986), neohop-13(18)-enes (C27, C30; PeaksN and h, Fig. 1; Ensminger et al., 1977; Brassell et al., 1980a,b)and fern-8-ene (Peak g, Fig. 1; Brassell and Eglinton, 1983).
The most variable characteristic of the hydrocarbondistributions is the relative amounts of C32 - C34 2β-methyl-17β(H),21β(H)-hopanes (Peaks m, s, and v in Fig. 1: McEvoyand Giger, 1986), which are prominent constituents of specificsamples from Site 1207B (i.e. 44R-1 60-61 cm: Fig. 1a) and Site1213B (8R-1 47-48 cm; Fig. 1c; also 8R-1 96-97 cm). 2β-methyl-17β(H)-22,29,30-trisnorhop-17(21)-ene (Peak R, Fig. 2;cf. McEvoy and Giger, 1986) also occurs in these samples. Incontrast, none of these components is present 1207B abovedetection limits in samples 44R-1 103-104 cm, 1213B 8R-1 63-64 cm (Fig. 1d), and 1214A 23R-1 5-7 cm (Table 5). Thus,despite the broad similarity in the distributions of aliphatichydrocarbon among the sites there are some noteworthydifferences. In particular, hopanes appear more prevalent at Site1213B, as shown by the proportion of 17β(H),21β(H)-homohopane relative to C29 Δ
4-sterene (Table 3). The occurrenceof only one dammarene (Peak N, Fig. 1) is consistent with
Table 3
Identities of biomarkers from the hydrocarbon fractions, labeled in Figure 1.Peak Identification Peak Identification
A 3-Me C15 [3-methylpentadecane] a C29 Δ5 sterene [24-ethylcholest-5-ene]
B 2,6,10-trimethyltridecane b C29 Δ17(21) hopene [30-norhop-17(21)-ene]
C 2,6,10-trimethylpentadecane c C29 5α sterane [24-ethyl-5α(H)-cholestane]D pristane [2,6,10,14-tetrapentadecane] d C30 Δ
17(21) hopene [hop-17(21)-ene]E phytane [2,6,10,14-tetrahexadecane] e C29 Δ
3,5 steradiene [24-ethylcholesta-3,5-diene]F phytenes [2,6,10,14-tetrahexadecenes] f C30 17α,21β hopane [17α(H),21β(H)-hopane]G C20 tricyclic terpane g fern-8-eneH phytenyl thiophenes h C30 Δ
13(18) neohopene [neohop-13(18)-ene]I hexamethyltetracosane [2,6,10,14,18,22-hexamethyltetracosane] i C29 17β,21β hopane [30-nor-17β(H),21β(H)-hopane]J C27 5β sterane [5β(H)-cholestane] j C31 Δ
17(21) hopenes [22S, 22R homohop-17(21)-enes]K C27 Δ
4 sterene [cholest-4-ene] k C31 17α,21β hopane [17α(H),21β(H)-homohopane]L C27 Δ
5 sterene [cholest-5-ene] l C30 17β,21β hopane [17β(H),21β(H)-hopane]M C27 5α sterane [5α(H)-cholestane] + C32 Δ
17(21) hopenes [22S, 22R bishomohop-17(21)-enes]N Δ13(17) dammarene [dammar-13(17)-ene] m C31 17β,21β Me hopane [2β-methyl-17β(H),21α(H)-homohopane]
+ C27 Δ13(18) neohopene [22,29,30-trisnorneohop-13(18)-ene] n C32 17α,21β hopane [17α(H),21β(H)-bishomohopane]
O C28 Δ4,22 steradiene [24-methylcholesta-4,22-diene] o C32 17β,21α hopane [17β(H),21α(H)-bishomohopane]
P C28 Δ5,22 steradiene [24-methylcholesta-5,22-diene] p C31 17β,21β hopane [17β(H),21β(H)-homohopane]
Q C27 Δ17(21) hopene [22,29,30-trisnorhop-17(21)-ene] q C33 17α,21β hopane [17α(H),21β(H)-trishomohopane]
R C28 Δ17(21) Me hopene [2β-methyl-22,29,30-trisnorhop-17(21)-ene] r C33 17β,21α hopane [17β(H),21α(H)-trishomohopane]
S C27 17α hopane [22,29,30-trisnor-17α(H)-hopane] s C33 17β,21β Me hopane [2β-methyl-17β(H),21α(H)-homohopane]T C29 triterpene t C32 17β,21β hopane [17β(H),21β(H)-bishomohopane]U C27 17β hopane [22,29,30-trisnor-17β(H)-hopane] u lycopane [2,6,10,14,19,23,27,31-octamethyldotriacontane]V C28 5α sterane [24-methyl-5α(H)-cholestane] v C34 17β,21β Me hopane [2β-methyl-17β(H),21α(H)-homohopane]W C29 Δ
4,22 steradiene [24-ethylcholesta-4,22-diene] w C33 17β,21β hopane [17β(H),21β(H)-trishomohopane]X C29 Δ
5,22 steradiene [24-ethylcholesta-5,22-diene]Y C29 5β sterane [24-ethyl-5β(H)-cholestane]Z C29 Δ
4 sterene [24-ethylcholest-4-ene]
* numbers = n-alkanes
4 Biogeochemical assessment of paleoproductivity
4
reports in Cretaceous black shales (Grimalt et al., 1990;Meunier-Christmann et al., 1991). Also, it was only observed inthe samples from Site 1207B and therefore appears restrictedamong the Sites.
3.4. Components of the Ketone Fraction
The dominant constituents of all samples are suites of C27 toC29 5α(H)−stanones (Peaks F, J, and P, Fig, 2; Gagosian andSmith, 1979; Robinson et al., 1984), and Δ4-stenones (Peaks I, O
and S, Fig. 2), accompanied by minor amounts of C28 and C29
stenones with Δ4,22 unsaturation (Peaks K and Q, Fig. 2) and byC28 to C30 4-methylstanones (Peaks H, N and R, Fig. 2),including dinosterone (Peak O, Fig. 2: Table 6; Robinson et al.,1984). A series of n-alkan-2-ones extending to C23, anddominated by C15, C17 and C19 homologues (Table 4; Simoneit,1979a,b; Brassell et al., 1980a) occur in all samples,accompanied by the C18 isoprenoid ketone (2,6,10-trimethylpentadecan-2-one; Simoneit, 1973) and a series of
Fig. 1. Distributions of aliphatic hydrocarbons and their assignments.
Biogeochemical assessment of paleoproductivity 5
5
abundant tocopherols (α-, β-, γ- and δ-; Peaks A and E, Fig. 2,Table 6; Brassell et al., 1983b; Goossens et al., 1984). C36 to C39
alkenones (Peaks a, d, e, and j, Fig. 2c: Volkman et al., 1980; deLeeuw et al., 1980: Xu et al., 2001) were recognized in a singlesample (Site 1213B-8R-1 96-97 cm; Brassell et al., 2004).
All samples contain 17β(H),21β(H)-hopanones (some or all ofthe C30, C33, C34, and C35 components, Peaks U, c, f, and i,respectively, Fig. 2; Dastillung et al., 1980b), although the majordifference among the samples in terms of the presence orabsence of biomarkers is the sporadic appearance and variableabundance of 2β-methyl-17β(H),21β(H)-hopanones (C34, C35,C36; Peaks b, e and h, Fig. 2). All three components areprominent constituents in two of the samples from Site 1203B(8R-1 47-48 cm and 8R-1 96-97 cm; Fig. 2a, c), but only minoramounts of the C34 homologue (b in Fig. 2) are observed in thethird sample from Site 1213B (8R-1 63-64 cm; Fig. 2b) and inthe sample from Site 1207B (44R-1 76-77 cm, Fig. 2d). In eachof the samples that contain 2β-methylhopanones a componenttenta t ive ly ident i f ied as 2β-methyl-17β(H),21β(H)-homohopanyl methyl ether is also present (Peak V, Fig. 2). 2β-methylhopanones were not detected in the sample from Site1214A (23R-1 5-7 cm; Fig. 2e).
3.5. Carbon Isotopic Composition of Biomarkers
The δ13C compositions of various hydrocarbons (hopanes, 2β-methylhopanes and lycopane) and ketones (sterones andalkenones) were determined in two samples from Site 1213B(8R-1 47-48 cm and 8R-1 96-97 cm, respectively). In addition,the δ13C value for α-tocopherol was measured in the latersample. The values for the hydrocarbons and ketones aredistinct, ranging from -28.4‰ to –30.6‰ and from –30.8‰ to -32.5‰, respectively (Table 7; Fig. 3), which suggests that theprincipal influence on the δ13C values may be compositionaldifferences between the OM of the two samples. Nevertheless,the values of hopanes and 2β-methylhopanes are similar, as arethose of sterones and alkenones. α-Tocopherol is less depletedwith a δ13C value of –26.4‰.
4. Discussion
4.1. Sources and Maturity of Organic Matter
4.1.1. Bulk characteristicsThe HI and OI values for the samples are generally similar
despite the vast differences in their organic carbon contents(Tables 1, 2). The high HI and extremely low OI values confirmthat Type I organic matter is dominant in all the samples(Espitalié et al., 1986) and indicates that they are composedpredominantly of organic matter of algal and bacterial origins.Similar characteristics were reported for specific intervals atSites 463 and 866 (Mid Pacific Mountains, and ResolutionGuyot, respectively; Dean et al., 1981; Mélières et al., 1981;Baudin et al., 1995; Bralower et al., 2002a) that correspond toOAE1a. Also, the HI values for the early Aptian at both Sites1207B and 1213B are comparable to the higher values (range167 - 400 mg HC/g TOC) for the organic-rich sediments (up to4.9 % Corg) corresponding to OAE1a (Selli unit) from theCismon core, Italy, where the OM is dominantly amorphous ofmarine origin (Hochuli et al., 1999).
4.1.2 C/N ratiosIn contemporary environments, high C/N ratios (>20–25) tend
to reflect land-derived organic matter (Meyers, 1994), but Corg-rich horizons are exceptions to this tenet. Many mid-Cretaceousblack shales (e.g., Meyers et al., 1984; Meyers, 1987) andNeogene or Quaternary sapropels (Meyers and Doose, 1999;Nijenhuis and de Lange, 2000) yield high C/N values that lie inthe range typically indicative of terrestrial organic matter. Yetthey appear to be dominantly composed of amorphous organicmatter derived from marine algae and bacteria and contain onlyminor contributions of material from land plants. Consequently,the C/N ratios for the organic-rich intervals corresponding toOAE1a (Table 1) seem somewhat anomalous, which suggeststhat either the characteristics of the source organic matter areunusual and/or nitrogen cycling in these horizons may beradically different from the norm. Exceptional preservation ofalgal organic matter may reduce recycling of its nitrogen
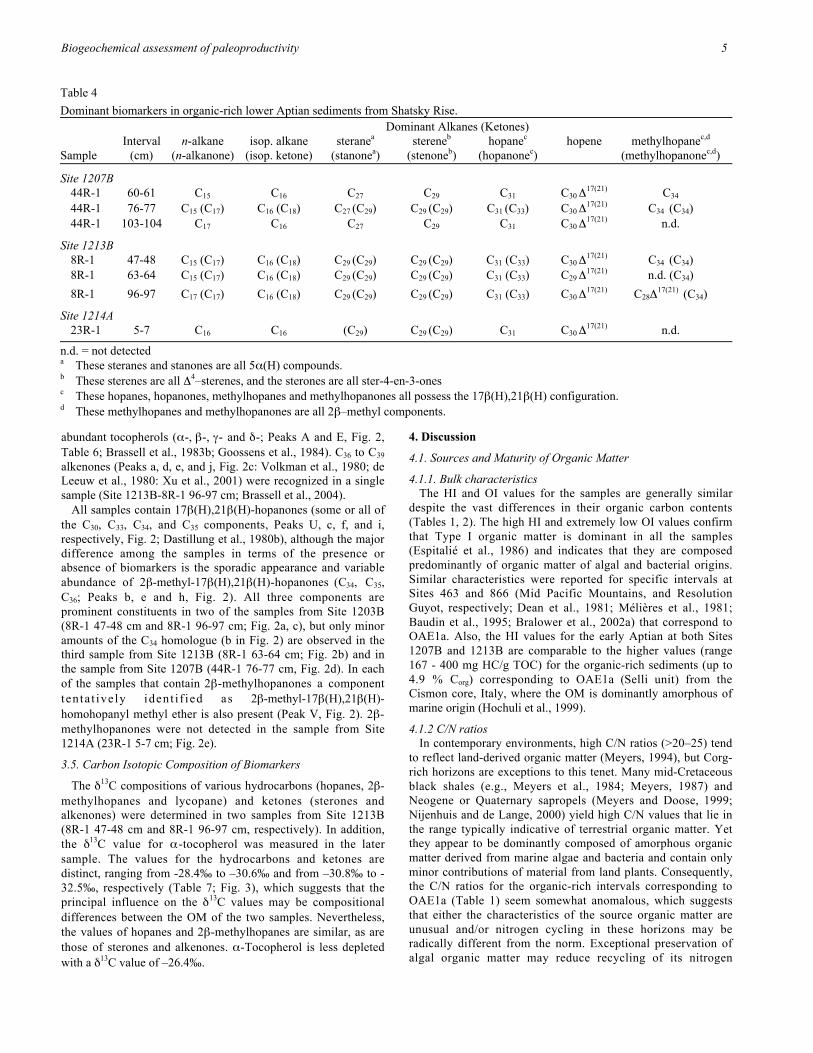
Table 4
Dominant biomarkers in organic-rich lower Aptian sediments from Shatsky Rise.Dominant Alkanes (Ketones)
Interval n-alkane isop. alkane steranea stereneb hopanec hopene methylhopanec,d
Sample (cm) (n-alkanone) (isop. ketone) (stanonea) (stenoneb) (hopanonec) (methylhopanonec,d)
Site 1207B44R-1 60-61 C15 C16 C27 C29 C31 C30 Δ
17(21) C34
44R-1 76-77 C15 (C17) C16 (C18) C27 (C29) C29 (C29) C31 (C33) C30 Δ17(21) C34 (C34)
44R-1 103-104 C17 C16 C27 C29 C31 C30 Δ
17(21) n.d.
Site 1213B8R-1 47-48 C15 (C17) C16 (C18) C29 (C29) C29 (C29)
C31 (C33) C30 Δ17(21) C34 (C34)
8R-1 63-64 C15 (C17) C16 (C18) C29 (C29) C29 (C29)
C31 (C33) C29 Δ17(21) n.d. (C34)
8R-1 96-97 C17 (C17) C16 (C18) C29 (C29) C29 (C29) C31 (C33) C30 Δ
17(21) C28Δ17(21)
(C34)
Site 1214A23R-1 5-7 C16 C16 (C29) C29
(C29) C31 C30 Δ17(21) n.d.
n.d. = not detecteda These steranes and stanones are all 5α(H) compounds.b These sterenes are all Δ4–sterenes, and the sterones are all ster-4-en-3-onesc These hopanes, hopanones, methylhopanes and methylhopanones all possess the 17β(H),21β(H) configuration.d These methylhopanes and methylhopanones are all 2β–methyl components.

6 Biogeochemical assessment of paleoproductivity
6
contents within the water column, thereby decreasing C/N ratios(Meyers and Doose, 1999). Perhaps the survival of cell wallsthat are rich in lipids and aliphatic biopolymers, like sheaths,significantly enhances Corg contents, without similarlyincreasing nitrogen content. C/N ratios would also be modifiedby preferential removal of nitrogen-rich organic compoundsduring anaerobic degradation of organic matter (Nijenhuis andde Lange, 2000). Nitrogen isotope profiles for Toarcian blackshales also provide evidence of partial denitrification inenvironments in which Corg-rich intervals are deposited(Jenkyns et al., 2001). Moreover, nitrogen-limiting conditionsmay produce higher C/N ratios. It seems likely that somecombination of these factors may account for the values oftenobserved for Cretaceous sediments deposited during OAE.
4.1.3. δ13C values for organic matterThe lower δ13C value for OM (–27.7‰; Table 1) at the base of
the organic-rich interval at Site 1207B corresponds (Authors’unpublished results) to the negative excursion associated withthe onset of OAE1a, which is well documented globally in thecarbon isotopic record of carbonates (δ13Ccarb; e.g. Menegatti etal., 1998; Luciani et al., 2001; de Gea et al., 2003; Wissler et al.,2003) including those from ODP Sites 866 and 463 in the Mid-Pacific Mountains (Jenkyns, 1995; Price, 2003), organic carbon(δ13Corg; Menegatti et al., 1998; Bralower et al., 1999; Jahren etal., 2001), and wood (Gröcke et al., 1999; Ando et al., 2002).However, the overall range of δ13Corg values in the early Aptian
at Site 1207B is significantly less (~2.5‰; Authors’ unpublishedresults) than those reported for δ13Corg in OM from Italy, Mexicoand Colombia (range of ~3.5‰ to ~6‰; Menegatti et al., 1998;Bralower et al., 1999; Jahren et al, 2001) and for wood fromEngland and Japan (range of ~3.5‰ to ~10‰; Gröcke et al.,1999; Ando et al., 2002). This smaller range for δ13Corg atShatsky Rise may reflect the fact that the core material did notsample intervals representative of the peak δ13Corg values,although a continuous record of the recovered section wasobtained. However, it seems likely that the differences in δ13Corg
ranges may reflect spatial variations in carbon reservoirs, in thedominant sources of organic matter (cf. Kuypers et al., 2001,2002 for OAE1b), in plankton productivity, or in upwellingintensity (cf. Price, 2003). Moreover, the pelagic nature of theSite 1207B sequence may record such effects differently fromterrestrial sections.
4.1.4. Preservation of organic matterThe combination of sediment laminations and exceptionally
high organic carbon contents in some sections of the organic-rich intervals (Table 1) indicates depositional conditions thatwere dysoxic or anoxic. The low pristane/phytane ratios – evenfor intervals less enriched in OM (Table 3) – are consistent withthat assessment (Didyk et al., 1978). The survival andabundance of functional biomarkers within the samples,especially at Site 1213 where the concentration of ketones
Table 5
Selected biomarker ratios for organic-rich lower Aptian sediments from Shatsky Rise.
Interval n-C17/ n-C17/ C31 ββ hopane/ C29 sterones C33 ββ hopanone/ C29 5α(H)-stanone/Sample (cm) n-C27 Pr Pr/Ph C29 Δ
4 sterene Δ4/5α(H) C29 Δ4 sterone 4-Me C29 stanone
Site 1207B44R-1 60-61 4.3 3.4 0.35 0.65 n.d. n.d. n.d.44R-1 76-77 2.6 3.3 0.56 0.77 1.3 0.14 n.d.44R-1 103-104 4.2 4.7 0.77 0.67 n.d. n.d. n.d.
Site 1213B8R-1 47-48 1.3 1.3 0.46 3.2 1.7 0.25 3.08R-1 63-64 3.7 3.6 0.27 2.0 3.7 0.05 4.28R-1 96-97 2.0 1.9 0.32 4.6 0.79 0.48 3.1
Site 1214A23R-1 5-7 6.1 2.3 1.2 1.6 0.92 0.05 3.6
Carbon No. Distributions of Steroids 2β-MethylHopanoid/Hopanoid RatiosInterval Δ4-sterenes stanones stenones hopanes hopanones
Sample (cm) 27:28:29 27:28:29 27:28:29 C32/C31 (%) C33/C32 (%) C34/C33 (%) C35/C34 (%)
Site 1207B44R-1 60-61 33:16:51 n.d. n.d. n.d. 0.49 (33)a n.d. n.d.44R-1 76-77 33:16:51 44:12:44 36:10:54 0.43 (30) n.d. 0.27 (21) n.d.44R-1 103-104 41:13:46 n.d. n.d. 0 0 n.d. n.d.
Site 1213B8R-1 47-48 30:18:52 41:11:48 41:11:48 0.64 (39) 0.61 (38) 1.2 (55) 1.3 (57)8R-1 63-64 27:18:55 37:9:54 35:9:56 0 0 0.32 (24) 08R-1 96-97 33:17:50 44:9:47 32:8:60 n.d. 0.27 (21) 2.1 (68) 1.9 (66)
Site 1214A23R-1 5-7 36:6:58 40:10:50 27:18:55 0 0 0 0
n.d. = not determineda Value for C34/C33 methylhopane/hopane ratio.

Biogeochemical assessment of paleoproductivity 7
7
significantly exceeds that of hydrocarbons, further echoes the
Table 6Identities of biomarkers within the ketone fractions, as labeled in Figure 2.
Peak Assignment Peak Assignment
A β-, γ-tocopherol T C30 triterpenoneB unknown U C30 hopanoneC C27 5β stanone [5β(H)-cholestan-3-one] V C32 ββ Me hopanyl Me ether?D C27 17β hopanone [17β(H)-22,29,30-trisnorhopane] [2β -methyl-17β(H),21β(H)-bishomohopanyl methyl ether]E α-tocopherol a C36:2 Et alkenone [hexatriaconta-16E,23E-dien-2-one]F C27 5α stanone [5α(H)-cholestan-3-one] b C34 ββ methylhopanoneG C28 Δ
22 stanone [24-methylcholest-22-en-3-one] [2β -methyl-17β(H),21β(H)-trishomohopan-32-one]H C28 4-Me 5α stanone [4-methyl-5α(H)-cholestan-3-one] c C33 ββ hopanone [17β(H),21β(H)-trishomohopan-32-one]I C27 Δ
4 stenone [cholesta-4-en-3-one] d C37:2 Me alkenone [heptatriaconta-15E,22E-dien-2-one]J C28 5α stanone [24-methyl-5α(H)-cholestan-3-one] e C35 ββ Me hopanoneK C28 Δ
4,22 stenone [24-methylcholesta-4,22-dien-3-one] [2β -methyl-17β(H),21β(H)-tetrakishomohopan-33-one]L C29 Δ
22 stanone [24-ethylcholest-22-en-3-one] f C34 ββ hopanone [17β(H),21β(H)-tetrakishomohopan-33-one]M C29 5β stanone [24-ethyl-5β(H)-cholestan-3-one] g C38:2 Et alkenone [octatriaconta-16E,23E-dien-3-one]N C29 4-Me 5α stanone [4,24-dimethyl-5α(H)-cholestan-3-one] h C36 ββ Me hopanoneO C30 Δ
22 4-Me 5α stanone [4,23,24-trimethyl-5α(H)-cholest-22-en-3-one] [2β -methyl-17β(H),21β(H)-pentakishomohopan-34-one]+ C28 Δ
4 stenone [24-methylcholest-4-en-3-one] i C35 ββ hopanoneP C29 5α stanone [24-ethyl-5α(H)-cholestan-3-one] [17β(H),21β(H)-pentakishomohopan-34-one]Q C29 Δ
4,22 stenone [24-ethylcholesta-4,22-dien-3-one] j C39:2 Me alkenone [nonatriaconta-15E,22E-dien-2-one]R C30 4-Me 5α stanone [4,24-dimethyl-5α(H)-cholestan-3-one]S C29 Δ
4 stenone [24-ethylcholest-4-en-3-one]
Fig. 2. Distributions of ketone fractionsand their assignments.

8 Biogeochemical assessment of paleoproductivity
8
significantly exceeds that of hydrocarbons, further echoes thewell-preserved character of organic matter and confirms thatvariations in source-specific biomarkers will reflect environmentchanges. In addition, the Rock-Eval Tmax values for the samples(range 401°C to 422°C; Table 2) attest to the immaturity of theOM (Espitalié et al., 1977).
4.1.5. Biomarker distributionsThe series of steroidal and hopanoidal hydrocarbons and
ketones that dominate all the samples provide evidence of algaland bacterial contributions to the sedimentary OM depositedduring OAE1a. The abundance of steroidal components,represented by sterenes in the hydrocarbon fraction (Fig. 1,Table 2-4) and by sterones in the ketone fraction (Fig. 2, Tables3-5), demonstrates that eukaryotic sources are majorcontributors to the OM within OAE1a at Shatsky Rise, andindicates that this environment experienced significantphytoplankton productivity. Evidence for dinoflagellates among
the phytoplankton is provided by the occurrence of 4-methylcomponents, including dinosterone, among the steroidal ketones(Robinson et al., 1984), although their relative abundance is lessthan their stanone counterparts (Table 5). The presence ofalkenones is diagnostic of OM derived from representatives ofhaptophyte algae among the calcareous nannoplankton (Brassellet al., 2004; Fig. 2), although their occurrence is confined to oneinterval at Site 1213B. The occurrence of several othercomponents in the sediments, such as acyclic isoprenoid alkanesand the tocopherols most likely derived from phototrophicorganisms, further reflect contributions of OM fromphytoplankton.
The variable abundances of 2β -methylhopanes and 2β-methylhopanones indicate fluctuating contributions of OM fromcyanobacteria at Sites 1207B and 1213B (Summons et al.,1999). Moreover, all cyanobacteria that synthesize 2-methylhopanoids also make hopanoids, which implies that aproportion of the latter compounds must also be ofcyanobacterial origin. This assessment is supported by thegeneral co-variance in their abundances: within the samples,consistent with derivation from a common biological source. Forsome samples, especially 1213B 8R-1 47-48 cm and 96-97 cm,the ratios of 2-methylhopanoids to hopanoids (Table 4) farexceed the average proportion of 2β -methyl-bacter io-hopanepolyols (BHPs) among total BHPs in cyanobacteria.Thus, cyanobacteria must account for a significant proportion ofhopanoids as well as 2β-methylhopanoids in the samples – anassessment supported by the consistency in the ratio of 2-methylhopanoids to hopanoids (Table 4) for differenthomologues (especially C34/C33, C35/C34, and C36/C35
hopanones). The prevalence of n-C17 is also consistent with OMcontributions from cyanobacteria (e.g. Blumer et al., 1971;Brassell et al., 1978; Shiea et al., 1990). It therefore appears that
Table 7
δ13C values and assignments of selected biomarkers.
No.a Compound Assignment Peakb δ13C (‰)
Hopanes and Lycopane in 1213B-8R-1 47-48 cm (Fig.1c)
1 C32 β,β hopane [17β(H),21β(H)-bishomohopane] t -29.04 ± 0.012 C33 β,β hopane [17β(H),21β(H)-trishomohopane] w -30.66 ± 0.403 C33 β,β 2β-Mehopane [2β-methyl-17β(H),21β(H)-bishomohopane] s -28.42 ± 0.194 C34 β,β 2β-Mehopane [2β-methyl-17β(H),21β(H)-trishomohopane] v -29.82 ± 1.175 lycopane [2,6,10,14,19,23,27,31-octamethyldocontane] u -30.18 ± 0.40
Steroidal Ketones and Tocopherol in 1213B-8R-1 96-97 cm (Fig.2c)
6 C37:2 methylalkenone [heptaconta-15,22-dien-2-one] d -31.29 ± 0.177 C38:2 ethylalkenone [octaconta-16,23-dien-3-one] g -31.878 C27 5α-stanone [5α(H)-cholestan-3-one] F -31.76 ± 0.049 C27 Δ
4 -stenone [cholesta-4-en-3-one] I -31.37 ± 0.2710 C28 Δ
4,22-stenone [24-methylcholesta-4,22-dien-3-one] K -32.47 ± 0.2411 C29 Δ
4,22-stenone [24-ethylcholesta-4,22-dien-3-one] Q -30.83 ± 0.2612 C29 Δ
4-stenone [24-ethylcholest-4-en-3-one] S -31.94 ± 0.4113 α-tocopherol E -26.45 ± 0.18
a Number of compounds given in Fig. 3.b Peak designation in Fig. 1c and Fig. 2c for aliphatic hydrocarbons and ketones, respectively.
Fig. 3. Carbon isotopic composition of various biomarkers.

Biogeochemical assessment of paleoproductivity 9
9
cyanobacteria were important members of the phytoplanktoncommunity during OAE1a.
Other constituents of the samples have microbial affinities.For example, the biological source of dammarenes is notconfirmed, but their structural affinities and occurrences areconsistent with a microbial source (Meunier-Christmann et al.,1991). Similarly, a bacterial origin for hop-17(21)-ene, neohop-13(18)-ene, and fern-8-ene seems likely (Brassell and Eglinton,1983) based on the occurrence of these hopenes or, in the case offern-8-ene, its precursor fern-7-ene in the anaerobicphotosynthetic bacterium Rhodomicrobium vannielli (Howard etal., 1984). Alternatively, hop-17(21)-ene may be a diageneticproduct from hop-22(29)-ene (diploptene; Ensminger et al.,1977; Brassell et al., 1980a), which occurs in cyanobacteria.(Sakata et al., 1997). The presence of 2β-methyl-22,29,30-trisnorhop-17(21)-ene suggests that 22,29,30-trisnorhop-17(21)-ene may also derive from cyanobacteria, presumably as adiagenetic product from a BHP precursor, or from hopan-22-ol(diplopterol). Both cyanobacteria and methanotrophic bacteriahave been reported to produce 2β-methylhopan-22-ol (2β-methyldiplopterol; Bisseret et al., 1985).
The low abundance of higher n -alkanes, especially oddhomologues (e.g. C29, C31) suggests little terrestrial plant input toShatsky Rise, implying perhaps trace amounts of aeolian dust.Thus, the domination of C29 steroidal components (range 44-60%, average 52%; Table 4) can be interpreted as a measure ofthe plankton sterol signature rather than any indication ofsignificant contributions from vascular plants. This assessmentconcurs with interpretations of the origin for C29 sterols inmodern settings (Namibian shelf and Peru Upwelling) thatreceive minimal terrestrial inputs (Lee et al., 1980; Gagosian etal., 1980, 1983).
4.1.6. δ13C compositions of biomarkersThe range of δ13C values of all of the biomarkers – hopanes,
methylhopanes, alkenones and sterones – is consistent with aplanktonic origin for the OM from Shatsky Rise (Table 7, Fig.3). The similarity of the δ13C values for hopanes and 2β-methylhopanes (-28.4‰ to -30.7‰) supports their commonorigin from cyanobacteria. Their comparability to the δ13Cvalues of steroidal components (Table 6) derived fromeukaryotes further confirms their origins within the photic zone(Sakata et al., 1997). The less negative values for the hopanes(Table 7) are consistent with evidence for less discriminationagainst 13C by cyanobacterial RUBISCO compared with algalRUBISCO (Farquhar et al., 1989a,b, Raven and Johnston, 1991,Raven, 1994). Similarly, the markedly higher δ13C value for α-tocopherol compared to the other biomarkers (Table 7) isconsistent with expectations for a component biosynthesizedfrom isoprene and tyrosine precursors (cf. Hayes, 1993).
4.1.7. Diagenetic AlterationThe ranges of Rock-Eval Tmax values for Sites 1207B and
1213B are similar, and therefore do not differentiate between thematurity of the sediments from these Sites. However, there areminor differences between the samples from the sites that can beattributed to different degrees of burial diagenesis related to thedepths of burial of sediments corresponding to OAE1a at Site1207B compared to Site 1213B, and Site 1214A. The marginallyhigher maturity of the sediments at Site 1207B is shown, forexample, by an absence of phytenes (Peak F, Fig. 1), thepresence of steranes (Peaks J, M, V, Y and c, Fig. 1; Table 3)
and the lower proportions of ketones relative to hydrocarbons(Fig. 2d). These differences directly reflect the depth of burial ofthe sequences.
The structural variety and stereochemical configurations of thebiomarkers within the samples (Tables 3, 6) all reflect theirimmaturity and are consistent with the data from Rock-Evalpyrolysis (Table 2). For example, Δ4- and Δ5-sterenes dominatetheir saturated counterparts; Δ4-sterones are accompanied by5α (H)- and 5β (H)-stanones, and the distributions ofboth sterenes and sterones include components withΔ22 unsaturation. Similarly, the dominant stereochemistry of thehopanes and hopanones, and their 2β-methyl homologues is the17β(H),21β(H) biological configuration. The occurrence ofsuites of hopenes, extended hopanoid alkanes and ketones andthe tentatively identified 2β-methylhomohopanyl methyl etherall help confirm that the samples show evidence of initial phasesof diagenetic alteration that transform functional lipids into theirimmediate hydrocarbon products (e.g. sterenes from sterols; e.g.Mackenzie et al., 1982).4.1.7.1. Origin of hopanones. The homologous series of hopanesand 2β-methylhopanes in all the OAE1a samples are dominatedby C31 and C32 components (Fig. 1; Table 3), respectively, whichfollows the pattern widespread in other immature Cretaceoussediments (e.g. Brassell et al., 1983b; Rullkötter et al., 1984,1987; Simoneit, 1986; Farrimond et al., 1989, 1990) andparallels the distribution expected for hydrocarbons derivedfrom BHP precursors (Ourisson and Rohmer, 1992; Innes et al.,1997, 1998; Farrimond et al., 2002). Similarly, it seemsplausible that the 2β-methylhomohopanyl methyl ether (Peak V,Fig. 2) tentatively identified in several samples is formed byd iagene t i c a l t e ra t ion o f a p recurso r 2β -methylbacteriohopanepentol comparable to BHPs known incyanobacteria (Ourisson and Rohmer, 1992; Zhao et al., 1996;Watson and Farrimond, 2000). By contrast, the carbon numberdistribution of the hopanones and 2β-methylhopanones (Fig. 2;Table 6), which are dominated by C33 and C34 compounds,respectively, raises questions about their precursors. In the vastmajority of immature sediments the major hopanoid acids,hopanols and hopanones are C32 or C31 constituents (Barnes etal., 1979; Dastillung et al., 1980a,b; Brassell and Eglinton, 1984;Jaffe et al., 1988; Grimalt et al., 1990; Ohkouchi et al., 1997;Innes et al., 1998; Farrimond et al., 2002) formed as productsfrom bacteriohopanetetrol or bacteriohopanepentol precursors(Ourisson et al., 1979, 1987; Rohmer, 1993; Innes et al., 1998).Enhanced preservation of the C35 hopane side chain can occurvia formation of organo-sulfur compounds (Valisolalao et al.,1984; Köster et al., 1997; Sinninghe Damsté et al., 1995), whichhelps to explain the prevalence of both C35 hopanes and hop-17(21)-enes in hypersaline environments (e.g. ten Haven et al.,1985; Brassell et al., 1988). Similarly, incorporation of BHPsinto kerogen provides a source for generation of varioushopanoid moieties (Richnow et al., 1992; Bishop et al., 1998;Farrimond et al., 2003). However, the occurrence of C33, C34,and C35 hopanones, and their 2β-methylhopanones homologues,with ω2 carbonyl groups, and with C33 and C34 components,respectively, as the dominant constituents of the series seemsinconsistent with known or likely diagenetic pathways fromprecursor BHPs. The predominance of C33 hopanes has beeninterpreted as an indication of contributions of OM from specificbacteria (Peters and Moldowan, 1991). Moreover, thesehopanone distributions are not unprecedented (Table 8); for

10 Biogeochemical assessment of paleoproductivity
10
example, 17β(H),21β(H)-trishomohopan-32-one (C33) wasfound to be the dominant hopanone in an Albian limestone fromHess Rise (Comet et al., 1981). Indeed it is often those sampleswhich possess high abundances of 2-methylhopanes in manyorganic-rich sediments, notably a number of Toarcian andCenomanian/Turonian black shales from the Atlantic andTethyean realms (Table 8), that also show a high relativeabundances of C33 hopanes and C34 2-methylhopanes. Thecomparability of these distributions with those of hopanones and2-methylhopanones from Shatsky Rise suggests that there maybe specific precursor hopanoids within cyanobacteria thatspecifically yield these homologues. Certainly the possibility ofhopanoid precursors other than C35 BHPs cannot be ignored(Farrimond et al., 1998, 2000), as suggested by the occurrenceof C31 to C34 hopane diols or triols in recent sediments (Rodier etal., 1999; Watson and Farrimond, 2000). However, the scarcityof dominant C33 hopanes within an extensive compilation of dataon hopanoid distributions (Bishop and Farrimond, 1995)suggests that this feature may be comparatively rare, or mayonly be manifest within a narrow maturity window prior to themajor phase of hopane generation from kerogen. In addition,there is an absence of 2-methylhopanoids in Albian samplescorresponding to OAE 1b (Table 8), perhaps reflecting thedominance of marine, non-thermophilic archaea (Kuypers et al.,2001) rather than cyanobacteria during this event.4.1.7.2. Organo-sulfur compounds. All the samples contain
minor amounts of phytenyl thiophenes (Peaks H, Fig.2) that arecommon constituents of immature OM, resulting fromdiagenetic incorporation of sulfur into chlorophyll-derivedphytol, or archaebacterial phytenes, or their diagenetic products(Brassell et al., 1986).
4.2.Paleoproductivity
4.2.1. Stratigraphic variabilityThe large range in organic carbon contents of the sediments
from OAE1a (Table 1) indicates that significant changesoccurred in either production and/or preservation of OM duringthis event. The depositional conditions, especially anoxia ordysoxia, were of crucial importance in preserving variableamounts of OM derived from planktonic sources. Themagnitude of organic carbon sequestration during OAE1a alsorequires that high levels of primary production were sustainedfor prolonged periods. At Site 1207B the mass accumulationrates for organic carbon range from 0.08 - 1.7 gCm-2y-1 based onthe Corg values (Table 1), the likely duration of OAE1a (0.5My;Leckie et al., 2002), shipboard measurements of physicalproperties (Bralower et al., 2002a), and the evidence from γ-raylogs that the total thickness of the organic-rich interval wasperhaps 1.2m (Bralower et al., 2002a,b). This accumulation ratefor Corg exceeds that of the early Aptian Selli unit in the Cismonsection, Italy (0.02 gCm-2y-1; Hochuli et al., 1999), but issignificantly less than the higher range of values calculated for
Table 8
2-Methylhopanoid ratios from selected Mesozoic organic-rich sediments
DSDP/ Number 2-Methylhopanoid IndicesODP Site of as % 2-MeH/(2-MeH + H)a Dominant Homologue(s)
Sample or Country Samples Hopanes (C no.)b Hopanones (C no.)
b Hopanes Hopanones Ref.c
Cenomanian/Turonian (OAE2)W. Africa 144 7 11 - 40d (C33) n.d. C33, C35 n.d. 1N.W. Africa 367 17 6 - 45d (C33) n.d. C33, C35 n.d. 1Western N. Atlantic 603B 11 2 - 33d (C33) n.d. C33, C35 n.d. 1Monte Massenza Italy 2 10, 13 (C29) n.d. n.r. n.d. 2,3Covelo Italy 2 11, 12 (C29) n.d. C31 n.d. 2,3Oued Bahloul Tunisia 2 15, 26 (C29) n.d. C33 n.d. 2,3
Albian (OAE 1b)Blake Nose 1049C 2 0 (C33) n.d. C29 n.d. 1Hess Rise 465 3 0 (C31) 0 (C33) C31 C33 4
Early Aptian (OAE1a)Cismon Italy 23 10 - 36 (C31) n.d. C31 n.d. 5Resolution Guyot 463 6 10 - 22 (C31) n.d. C31 n.d. 5Shatsky Rise 1207B 2 30, 33 (C31, C32) 21 (C33) C31 C33 6Shatsky Rise 1213B 3 21 - 39 (C31) 24 - 68 (C33) C31 C33 6Shatsky Rise 1214A 1 0 (C31) 0 (C33) C31 C33 6
ToarcianRizapol Italy 1 12 (C29) n.d. C32, C33 n.d. 2,7Monte Mangart Italy 1 14 (C29) n.d. n.r. n.d. 2,7Longarone Italy 1 18 (C29) n.d. C33 n.d. 2,7
4
n.d. = not determined; n.r. = not reported.a 2-MeH and H are abbreviations for 2-methylhopanoids and hopanoids, respectively.b The Cno. given in parentheses refers to the hopane homologue(s) for which the 2-methylhopanoid indices are calculated. Thus, C33 refers to
methyltrishomohopane and trishomohopane (i.e C34/C33)c Key to references cited: 1 = Kupyers, 2001; 2 = Farrimond, 1987; 3 = Farrimond et al., 1990; 4 = Comet et al., 1991; 5 = van Breugel et al.,
2002, 2003; 6 = This paper; 7 = Farrimond et al., 1989.d These 2-methylhopanoid indices were determined for sulfur-bound components. The average values for Sites 144, 367 and 603B, respectively,
are 23, 22 and 13 for C33 hopanes and 8, 9 and 10 for C35 hopanes.

Biogeochemical assessment of paleoproductivity 11
11
sediment sequences deposited during OAE2 (0.1 - 33 gCm-2y-1;Arthur et al., 1987; Kuypers et al., 2002) and in contemporaryregions of high primary production (0.8 - 39 gCm-2y-1; Kuyperset al., 2002). Thus, there appears to have been intermittent highproductivity during OAE1a at Shatsky Rise.
Despite these variations in productivity and associatedfluctuations in the abundance of Corg, the character of the OMcan be seen to be rather uniform, based the distribution patternsof hydrocarbons and ketones within these lower Aptian samples(Figs. 1, 2). Thus, the similarity in the profiles of steroidalketones and hydrocarbons derived from eukaryotes suggestsbroad uniformity in the algal primary producers contributingOM in the region of Shatsky Rise.
However, the biomarkers deposited within this environmentdo record fluctuations in the productivity of specific algae (e.g.haptophytes, dinoflagellates) based on the differences in therelative abundances of the biomarkers (i.e. alkenones, 4-methylsterones) that are diagnostic of contributions from suchphytoplankton. These stratigraphic variations suggest temporalfluctuations in phytoplankton populations and microbial activity,which are also illustrated by minor changes in the abundance orpresence of n-alkanes and dammarenes. Another series ofhydrocarbons, 2β-methylhopanes (Peaks m, s, and v in Fig. 1),and their ketone equivalents (2β-methylhopanones, Peaks b, e,and h in Fig. 2) that are both diagnostic of cyanobacteria,(Summons et al., 1999) show perhaps the greatest variationsamong the samples in terms of their occurrence and abundance(Table 5).
4.2.2. CyanobacteriaThe high proportions of 2β-methylhopanoids in several of the
samples yields methylhopane/hopane ratios (Table 5) that farexceed the range reported for other sedimentary rocks, includingnumerous samples from the Precambrian (cf. Farrimond et al.,1989, 1990; Summons et al., 1999; Kupyers, 2001). DuringOAE1a cyanobacteria therefore represented, at leastintermittently, a significant portion of the phytoplankton basedon these characteristics. This prevalence of OM contributionsfrom cyanobacteria is likely related to nutrient availability,especially nitrate- or iron-limited conditions within the photiczone (Bralower et al., 2002a). In addition, the proposedpronounced rise in atmospheric pCO2 during the Early Aptian(Retallack, 2001) would have inhibits biocalcification (Riebesellet al., 2000) through lowered carbonate saturation in surfacewaters (Wissler et al., 2003), which perhaps favored non-calcareous phytoplankton including cyanobacteria.
The relative abundance of 2β-methylhopanones relative tohopanes (Table 5) in the sediments from Site 1213B is inverselyproportional to Corg contents (Table 1). Thus, cyanobacterialcontributions of OM may be proportionately greater duringconditions of lower productivity, or their biosynthesis of 2-methylhopanone precursors may be enhanced during suchconditions. Indeed, earlier arguments that hopanes may also beproducts from cyanobacteria – an interpretation supported by theδ13C values determined for hopanes (Schoell et al., 1994; Sakataet al., 1997) – suggests that environmental or biological factorsmay control the biosynthesis of 2-methylhopanoids andhopanoids thereby affecting their relative proportions. Whateverthe precise cause for the elevated amounts of 2β -methylhopanoids at Shatsky Rise they exceed all previousreports (Summons et al., 1999; Table 8), including those ofblack shales assigned to OAE1a from the mid-Pacific (Site 463)
and Italy (Cismon section; van Breugel et al., 2002, 2003).Significant proportions of 2-methylhopanes have also beenfound in both Cenomanian/Turonian organic-rich sediments(Farrimond et al., 1990; Kuypers, 2001) and in Toarcian blackshales from Italy and Tunisia, where they have been interpretedto indicate dysoxic conditions (Farrimond et al., 1989).Collectively, these reports demonstrate that enhanced levels of2-methylhopanoids (Table 8), and by inference cyanobacteria,are often associated with organic-rich sequences.
4.3. Comparative biogeochemistry of OAE1a at Shatsky Rise
4.3.1. Hydrocarbons biomarkersThe distributions of biomarkers in the lower Aptian sediments
from ODP Leg 198 show many similarities to previous reportsof hydrocarbon distributions in other immature Cretaceousorganic-rich strata (cf. Barnes et al., 1979; Brassell et al., 1980b,1983; Comet et al., 1981; Brassell, 1984; Brassell andFarrimond, 1986; Farrimond et al., 1986a, 1989, 1990;Rullkötter et al., 1984, 1987; Simoneit et al., 1986; Kuypers etal., 2002). The suites of acyclic isoprenoid alkanes and thevarious series of steroidal and hopanoid alkanes and alkenes inthe Shatsky Rise samples (Fig. 1: Table 3) are all welldocumented in these previous reports. Even the 2-methylhopanoids that are prominent constituents of most of thesamples from Sites 1207B and 1213A have been observed inother Cretaceous sediments, albeit with 2α-methyl configuration(Table 8). Overall, the signatures of the hydrocarbon biomarkersshow that sediments deposited during OAE1a may becharacterized by different proportions of specific components,but provide no evidence of profound evolutionary changes inplankton populations akin to those recognized for Albian OAE1b in the Atlantic (Kuypers et al., 2001). Thus, enhancedsequestration of OM during OAE1a is best explained as anepisode of enhanced productivity and/or OM preservation. Thisinterpretation prompts consideration of the detailedcharacteristics of the sedimentary OM and biomarkerdistributions associated with OAE1a in efforts to identifyevidence for possible differences or changes in oceanographicconditions during the event.4.3.1.1. Organic matter in mid-Cretaceous sediments from thePacific. Assessment of the characteristics of OM by Rock-Evalpyrolysis revealed similarities between OAE1a samples fromLeg 198 (Sites 1207B and 1213B) and specific intervalscorresponding to OAE1a at Sites 463 and 866 (Mid PacificMountains, and Resolution Guyot, respectively; Bralower et al.,2002a). All these Pacific Sites include samples that possess highHI and low OI indices indicative of a dominance of OM ofalgal/bacterial origin. However, the character of the OM in thelower Aptian interval from Leg 198 appears more uniform thanthat from the other sites, a difference which perhaps reflectsdifferences in their depositional settings. The lower Aptiansediments from Shatsky Rise are truly pelagic (Bralower et al.,2002a, b), whereas those from Site 866 were deposited in ashallow water carbonate environment (Jenkyns, 1995), and someintervals from Site 463 contain a significant fraction of OMformed in a shallow water environment (Mélières et al., 1981;Dean et al., 1981). Similarly, the organic-rich Aptian sedimentrecovered from Manihiki Plateau was deposited in a lagoonalenvironment (Simoneit and Stuermer, 1982).
There are comparatively few investigations of biomarkers inmid-Cretaceous sediments from the Pacific, which limits thepossibility of inferring temporal changes in OM from their

12 Biogeochemical assessment of paleoproductivity
12
distributions. However, there are specific characteristics amongtheir dominant features that may reflect oceanic productivity.For example, the major biomarkers (n-alkanes, isoprenoidalkanes, steranes, sterenes, hopanes, hopenes) in Albiansediments from Sites 465 and 466 on Hess Rise are likelyderived from algal and bacterial phytoplankton (Comet et al.,1981; Simoneit, 1981). In addition, lycopane is the dominantalkane in an Albian limestone from the Hess Rise (Site 465;Comet et al., 1981; Brassell et al., 1983b). Archaebacteria wereinitially proposed as the likely biological source for lycopane(Brassell et al., 1981), but further studies indicate aphotoautotrophic source based on its occurrence and carbonisotopic composition (Sinninghe Damsté et al., 2003). Also, highrelative concentrations of lycopane have been related to anoxicconditions (Sinninghe Damsté et al., 2003), suggesting that itssedimentary abundance is dependent on both production andpreservation. Indeed, the general similarities among thehydrocarbon distributions for the Albian and lower Aptianorganic-rich sediments in the mid-Pacific that reflectscomparable biological sources of OM stands in marked contrastto the discrepancy in their concentrations of 2-methylhopanoids(Table 8). Moreover, investigations of biomarker compositionsin lower Aptian sediments from both Site 463 and the Cismonsection in Italy (van Breugal et al., 2002, 2003) confirm that theabundance of 2-methylhopanoids during this time interval isdistinctive (Table 8). Thus, enhanced cyanobacterial productionappears characteristic of OAE1a, differentiating this event fromOAE 1b, yet reappearing at some locations during OAE2(Cenomanian/Turonian; Kuypers, 2001; Farrimond et al., 1990),and resembling earlier intervals at some locations during EarlyToarcian OAE (Jenkyns, 1985, 1988; Farrimond et al., 1989;Table 8).
4.3.2. KetonesPublished investigations of ketones in Cretaceous sediments
are rare, although the suites of isoprenoidal, steroidal andhopanoidal ketones reported previously (Simoneit, 1979a,b,1981; Dastillung et al., 1980b; Comet et al., 1981; Grimalt et al.,1990) include most of the compounds, especially sterones,identified in the lower Aptian sediments from Shatsky Rise. Theseries of hopanones identified in these sediments were firstreported in shales (Dastillung et al., 1980b), and are documentedin other series of Cretaceous sediments (Wardroper, 1979;Simoneit et al., 1979b; Comet et al., 1981; Grimalt et al., 1990),except for the C34 to C36 2β-methylhopanones (Table 6), whichare novel compounds. Alkenones have previously been found inmid-Cretaceous strata (Albian and Cenomanian) from the Blake-Bahama basin (Farrimond et al., 1986b) where their presence, asat Shatsky Rise, can be attributed to the immaturity of thesediments (Brassell et al., 2004).
4.3.3. Organo-sulfur compounds in Cretaceous sedimentsOrgano-sulfur compounds (OSC), notably thiophenes, have
been reported in other Cretaceous marine sediments (e.g. vanGrass, 1982; Rullkötter et al., 1984; Brassell et al., 1986),although their abundance appears markedly higher inhypersaline settings, such as the Upper Cretaceous Jurf edDarawish oil shale (Kohnen et al., 1990). The occurrence ofphytenyl thiophenes is consistent with their prevalence amongOSC (Rullkötter et al., 1984; Brassell et al., 1986; SinningheDamsté and de Leeuw, 1987), and with the recognition that suchisoprenoid skeletons occur bound within OM by a diverse rangeof organo-sulfur bonds (Kohnen et al., 1990).
4.3.4. Distinctive biomarker compositions within OAE1aTwo features of the lower Aptian sediments from Shatsky Rise
are significant when compared with other organic-rich, mid-Cretaceous sediments. First, the effective absence of terrestrialOM contrasts with other sequences of organic-rich Aptiansediments from the Atlantic and Tethyean realms (Simoneit andStuermer, 1982; Simoneit, 1978, 1979a,b; Simoneit andMazurek, 1979a,b). This disparity primarily reflects thedifferences – continental margin versus open ocean – in thedepositional setting of these deposits.
Second, the relative abundance of 2β-methylhopanoidsrelative to hopanoids in the lower Aptian sediments fromShatsky Rise is extremely high (Table 5). Indeed, theproportions of hopanoid ketones in two samples from Site1213B represent the highest values ever reported (cf. Farrimondet al, 1989, 1990; Summons et al., 1999). They are comparableto, or exceed, the highest ratios for other Cretaceous sediments(Table 8) including sequences from Italy (Cismon section) andthe mid-Pacific mountains (DSDP Site 463) that correspond toOAE1a (van Breugel et al., 2003). Significantly lower values(12-18) for this index in Toarcian black shales from Italy (Table8) have been interpreted as indicative of dysoxic conditions(Farrimond et al., 1989). Yet the preponderance of 2-methylhopanes in lower Aptian sediments from both Pacific andTethyean realms (Table 8) suggests the critical factor controllingtheir abundance at this time was a change in globaloceanographic conditions that led to an increase ofcyanobacteria, rather than simply a difference in oxicity levels.Moreover, 2-methylhopanoids were not detected in the sample(Core 44R-1 103-104 cm) from the base of the organic-richinterval recovered at Site 1207B (Table 5). This sample recordsthe advent of enhanced sequestration of organic matter –although its Corg contents (1.7%) are far lower than subsequentvalues (Table 1) - and the negative δ13C excursion associatedwith OAE1a (Authors’ unpublished results). Thus, the evidencefor significant cyanobacterial production lags the initial phase ofcarbon burial consistent with the perspective that theseorganisms become more prevalent as the event proceeds,favored by temporal variations in nutrient levels, especiallydepletion of nitrate, that may reflect the profound changes in thepopulations, abundances and productivity of phytoplankton atthis time (e.g. Leckie et al., 2002), including the ‘nannoconidcrisis’ (Erba and Tremolada, 2004).
4.3.5. Carbon isotopic compositions and OAE1aThe δ13C values of individual biomarkers in the sediments
from Site 1213 corresponding to OAE1a are broadly comparableto values in mid-Cretaceous sequences but tend to besignificantly lower than those reported in organic-rich sedimentsfrom contemporary marine environments (Table 9). Forexample, the δ13C values of alkenones in OAE1a (-31.3‰ forC37:2 Me; -31.9‰ for C38:2 Et; Table 6) are less than thosereported for modern marine sediments, but similar to the rangeobserved elsewhere for pre-Neogene sediments (Table 9;Pagani, 2002). Other biomarkers show similar differences intheir carbon isotopic compositions (Table 9). The δ13C values ofsterols in modern sediments from the Peru upwelling region andArabian Sea are higher than those reported for steroidalcomponents in Miocene and Cretaceous sediments (Table 9),including sterones from OAE1a at Shatsky Rise (Table 7).Similarly, the δ13C values of lycopane in contemporarysediments, with the exception of the Black Sea, are higher than

Biogeochemical assessment of paleoproductivity 13
13
those reported in ancient sediments (Table 9). The δ13C valuesof hopanoids show a comparable trend if the low δ13C values fordiploptene (hop-22(29)-ene) in sediments from the CariacoTrench and Black Sea that reflect biosynthesis bymethanotrophic bacteria (Freeman et al., 1994) are excluded.The carbon isotopic compositions of hopanoids in Shatsky Risesediments (Table 7; Fig. 3) fall within the range of (δ13C = -27‰to -33‰) reported for hopanes (C31 hopane + C32 2-methylhopane) in lower Aptian sediments from the CismonValley and DSDP Site 463, which also match values for theCenomanian-Turonian (Table 9) and for Devonian sediments(Joachimski et al., 2001). They are markedly lower than thevalues for sediments corresponding to OAE1b from the NorthAtlantic Ocean (ODP Site 1049C), which range from -24‰ to -26‰ (Kuypers et al., 2002). Also, the δ13C values of hopanesshow minor, but consistent, differences relative to Corg (δ
13Chopane
- δ13CCorg) for separate OAE during the Cretaceous (Table 9).Specifically, hopanes in the lower Aptian (OAE1a) sequences
consistently show greater 13C fractionation relative to Corg (-3‰to -4.5‰) than their counterparts – and also steranes andlycopane where data are available – from theCenomanian/Turonian (OAE2) or Albian (OAE1b) intervals.Changes in the sources of biological OM, such as the dominanceof marine Archaea during OAE1b (Kuypers et al., 2001, 2002),could account for such variations in isotopic compositions(cf.Pagani et al., 2000b), although differences in the bacterialorigins of hopanes (photosynthetic phytoplankton versuschemoautotrophs) could create similar effects.
These low δ13C values for alkenones and other biomarkersassociated with early Aptian OAE1a are consistent with thoseexpected from biosynthetic isotopic fractionation duringepisodes of elevated pCO2 (e.g. Pagani, 2002; Brassell et al.,2004). This interpretation parallels evidence for high pC O2
during OAE1a based on leaf morphology (Retallack, 2001),except that this approach suggests even higher values for OAE2in contrast to isotopic studies of biomarkers assessing pCO2
Table 9
Comparisons of δ13C values for organic matter and specific lipid classes in organic-rich marine sediments.
δ13C Composition (‰) of Organic Carbon and Specific Lipid Classesa
Location Site(s)b Age TOC Alkenonesc Hopanes 2-Me Hopanes Steroids Lycopane Ref.d
Contemporary SettingsArabian Sea Several Recent -22 -22 to -24 -23 to -24 n.r. -23 to -25 ave. -24 1,2,3Peru Upwelling Several Recent (-17 to -24) n.d. n.d. n.r. -20 to -26 (-23) 3,4,5Cariaco Trench n.r. Recent n.d. n.d. ave. -43e n.r. n.d. ave. -24 3,4,6Black Sea Several Recent -24 -29 to -31 ave. -39e n.r. n.d. ave. -32 1,3,4,6,
Neogene and PaleogeneMonterey Naples Miocene -20 to -22 n.r. -25 to -31g n.r. -24 to -26g -27 to -33 3,8,9Monterey Pismo Miocene n.d. n.r. -23 to -28f n.r. -25 to -29g -23 to -26 3,10S.W. Pacific 588 Miocene -22 to -26 -17 to -27 n.d. n.r. n.r. n.r. 11-13N. Atlantic 608 Miocene -21 to -27 -18 to -29 n.d. n.r. n.r. n.r. 11-13Oman Margin 730 Miocene -20 to -23 -20 to -24 n.d. n.r. n.r. n.r. 11-13Southwest Atlantic 516 Miocene -21 to -27 -21 to -29 n.d. n.r. n.r. n.r. 12,13Southwest Atlantic 516 Eoc./Olig. n.d. -33 n.d. n.r. n.r. n.r. 12,13Campbell Plateau 277 Eoc./Olig. n.d. -34 to -38 n.d. n.r. n.r. n.r. 13
CretaceousCoast N.W. Africa 144 Cen./Tur. -25 to -28 n.r. -28 to -31g,h n.d. -28 3,14,15Coast N.W. Africa 367 Cen./Tur. -22 to -28 n.r. -19 to -30g.h n.d. -21 to -29 3,14-16Western N. Atlantic 603B Cen./Tur. -24 to -27 n.r. -24 to -29g,h n.d. n.r. 14Tafaya Basin S13 Cen./Tur. -24 to -28 n.r. n.d. n.r. -28 to -34 n.r. 14,15Blake Nose 1049 Albian -17 to -25 n.r. -24 to -27 n.r. -24 to -30 n.r. 14,17Cismon Italy Aptian -23 to -29 n.r. -27 to -32h n.d. n.r. 18Resolution Guyot 463 Aptian -24 to -28 n.r. -28 to -33h n.d. n.r. 18,19Shatsky Rise 1207, 1213 Aptian -25 to -28 ave. -32 ave. -30 ave. -30 -31 to -33 -30 20
PaleozoicHoly Cross, Poland Kowala Devonian -28 to -31 n.r. -28 to -33 n.d. n.r. n.r. 214
n.d. = not determined; n.r. = not reported; ave. = average value.a All values are rounded to whole numbers. Parentheses indicate values for suspended particulate organic matter rather than sedimentsb Sites that were DSDP and ODP Holes are shown in italics.c Values presented are those for C37 alkenones.d Key to references cited: 1 = Eglinton et al., 1997; 2 = Schouten et al., 2000a; 3 = Sinninghe Damste et al., 2003; 4 = Wakeham et al., 1993; 5
= Pancost et al., 1999; 6 = Freeman et al., 1994; 7 = Freeman and Wakeham, 1992; 8 = Schoell et al., 1994; 9 = Schouten et al., 2000b; 10 =Schouten et al., 1997; 11 = Pagani et al., 1999; 12 = Pagani et al., 2000a,b; 13 = Pagani, 2002; 14 = Kuypers, 2001; 15 = Kuypers et al.,2002; 16 = Kuypers et al., 1999; 17 = Kuypers et al., 2001; 18 = van Breugel et al., 2003; 19 = Dean et al., 1981; 20 = This paper, Tables 1and 7; 21 = Joachinski et al., 2001, 2002.
e Values for hop-22(29)-ene (diploptene) likely derived, at least in part, from chemoautotrophic bacteria (Freeman et al., 1994).f Range of values for free and sulfur-bound speciesg Values for sulfur-bound species.g Values for all hopanes (i.e. hopanes + 2-methylhopanes).

14 Biogeochemical assessment of paleoproductivity
14
during this later event (Freeman and Hayes, 1992, Kuypers etal., 1999).
5. Conclusions
The extraordinary sequestration of organic matter (up to34.7%) in lower Aptian sediments from Shatsky Rise, even bycomparison with other organic-rich intervals within the mid-Cretaceous, records a response to a radical change in theprocesses controlling biogeochemical cycling, which in turnreflects a perturbation of the ocean-climate system. Theelemental composition of the OM, and the distributions andisotopic compositions of biomarkers in these sediments all attestto major contributions of OM from phytoplankton. Source-specific biomarkers show contributions from eukaryotic algae,including haptophytes and dinoflagellates, accompanied bysignificant, yet variable proportions of 2β-methylhopanoids thatindicate an intermittent importance of cyanobacteria among thephytoplankton. Levels of primary production assessed from Corg
accumulation rates were high. However, biomarker distributionssuggest fluctuations in phytoplankton populations and microbialactivity especially changes in nutrient availability and nitrogencycling associated with nitrate depletion. Also, non-calcareousplankton may have been favored by lowered carbonatesaturation associated with high pCO2 as reflected by the lowδ13C values of biomarkers. These biogeochemical measuresattest to the complexity and dynamic character of OAE asreflected by evidence for significant fluctuations in pCO2, oceanproductivity, biota, and ocean temperatures (Hochuli et al.,1999; Luciani et al., 2001; Leckie et al., 2002; Wissler et al.,2003; Erba and Tremolada, 2004) are all associated with boththe early Aptian and other events (Norris and Wilson, 1998;Clarke and Jenkyns, 1999; Kuypers et al., 1999, 2002; Erbacheret al., 2001; Wilson and Norris, 2001; Huber et al., 2002; Norriset al., 2002; Bice and Norris, 2002; Bice et al., 2003).
Attempts to resolve and differentiate the influence of theseinterconnected variables on the biogeochemical carbon cyclerequire comprehensive analyses of the δ13C compositions ofsedimentary OM and biomarkers to define temporal trends insuch environmental variables, in addition to effects related tochanges in biosynthetic fractionation (εp; Freeman and Hayes,1992), phytoplankton growth rates and cell geometry (Francoiset al., 1993; Bidigare et al., 1997; Popp et al., 1998; Riebesell etal., 2000), sequestration of 13C-depleted OM (Arthur et al.,1988), episodes of enhanced production of volcanogenic CO2
(Erba and Tremolada, 2004) and CH4 release from hydratedissociation (Jahren et al, 2001; Beerling et al., 2002). Suchefforts are now in progress.
Acknowledgements
We should like to specially thank Anne Pimmel and TimBronk for their assistance with geochemical analyses in thechemistry laboratory onboard ODP Leg 198 and all R/VJOIDES Resolution marine technicians for their contributions tothe successful cruise. This research used samples and dataprovided by the Ocean Drilling Program (ODP). ODP issponsored by the U.S. National Science Foundation (NSF) andparticipating countries under management of JointOceanographic Institutions (JOI), Inc. NSF (EAR 9909857) andUSSAC are also thanked for research support to SCB.
References
Ando, A., Kakegawa, T., Takashima, R., Saito, T., 2002. Newperspective on Aptian carbon isotope stratigraphy: Data from δ13Crecords of terrestrial organic matter. Geology, 30, 227-230.
Arthur, M.A., Schlanger, S.O., Jenkyns, H.C., 1987. The Cenomanian-Turonian oceanic anoxic event, II. Palaeoceanographic controls onorganic-matter production and preservation. In Marine PetroleumSource Rocks (eds. Brooks, J., Fleet A.J.), 401-420, Blackwell,Oxford.
Arthur, M.A., Dean, W.E., Pratt, L.M., 1988. Geochemical andclimatic effects of increased marine organic carbon burial at theCenomanian/Turonian boundary. Nature, 335, 714-717.
Barnes, P.J., Brassell, S.C., Comet, P., Eglinton, G., McEvoy, J.,Maxwell, J.R., Wardroper, A. M. K., Volkman, J. K., 1979.Preliminary lipid analysis of core sections 18, 24 and 30 from Hole402A. In Initial Reports of the Deep Sea Drilling Project, 48 (eds.Montadert, L., Roberts D.G. et al.), 965-976, U.S. GovernmentPrinting Office, Washington D.C.
Baudin, F., Deconinck, J.-F., Sachsenhofer, R.F., Strasser, A., Arnaud,H., 1995. Organic geochemistry and clay mineralogy of LowerCretaceous sediments from Allison and Resolution guyots (Sites 865and 866), Mid-Pacific Mountains. In Proceedings of the OceanDrilling Program, Scientific Results, 143 (eds. Winterer, E.L., Sager,W.W., Firth, J.V., and Sinton, J.M.), 173-196.
Beerling, D.J., Lomas, M.R., Gröcke, D.R., 2002. On the nature ofmethane gas-hydrate dissociation during the Toarcian and Aptianoceanic anoxic events. American Journal of Science, 302, 28-49.
Bice, K.L., Norris, R.D., 2002. Possible atmospheric CO2 extremes ofthe Middle Cretaceous (late Albian – Turonian). Paleoceanography,17, 2002PA000778.
Bice, K.L., Huber, B.T., Norris, R.D., 2003. Extreme polar warmthduring the Cretaceous greenhouse? Paradox of the late Turonianδ18O record at Deep Sea Drilling Project Site 511.Paleoceanography, 18, 2002PA000848.
Bidigare, R.R., Fluegge, A., Freeman, K.H., Hanson, K.L., Hayes,J.M., Hollander, D., Jasper, J.P., King, L.L., Laws, E.A., Milder, J.,Millero, F.J., Pancost, R., Popp, B.N., Steinberg, P.A., Wakeham,S.G., 1997. Consistent fractionation of 13C in nature and in thelaboratory: Growth-rate effects in some haptophyte algae. GlobalBiogeochemical Cycles, 11, 279-292.
Bishop, A.N., Farrimond, P., 1995 A new method of comparingextended hopane distributions. Organic Geochemistry, 23, 987-990.
Bishop, A.N., Love, G.D., McCaulay, A.D., Snape, C.E., Farrimond,P., 1998. Release of kerogen-bound hopanoids by hydropyrolysis.Organic Geochemistry, 29, 989-1001.
Bisseret, P., Zundel, M., Rohmer, M., 1985. Prokaryotic triterpenoids.2. 2β-Methylhopanoids from Methylobacterium organophilum andNostoc muscorum, a new series of prokaryotic triterpenoids.European Journal of Biochemistry, 150, 29-34.
Blumer, M., Guillard, R.R.L., Chase, T., 1971. Hydrocarbons ofmarine phytoplankton. Marine Biology, 8, 183.
Bralower, T.J., CoBabe, E., Clement, B., Sliter, W.V., Osburn, C.L.,Longoria, J., 1999. The record of global change in mid-Cretaceous(Barremian-Albian) sections from the Sierra Madre, northeasternMexico. Journal of Foraminiferal Research, 29, 418-437.
Bralower, T.J., Premoli Silva, I., Malone, M.J., 2002a. Proceedings ofthe Ocean Drilling Program, Initial Reports, Volume 198: [online].College Station, Texas, Ocean Drilling Program (www-odp.tamu.edu/publications/198_IR/198ir.htm).
Bralower, T.J., Premoli Silva, I., Malone, M.J., and ScientificParticipants of Leg 198, 2002b. New evidence for abrupt climatechange in the Cretaceous and Paleogene: An Ocean DrillingProgram expedition to Shatsky Rise, northwest Pacific. GSA Today,12, 4-10.
Brassell, S.C., Eglinton, G., Maxwell, J.R., Philp, R.P., 1978. Naturalbackground of alkanes in the aquatic environment. In Aquatic

Biogeochemical assessment of paleoproductivity 15
15
pollutants: Transformation and Biological Effects (eds. HutzingerO., van Lelyveld L.H., Zoeteman B.C.J.), 69-86, Pergamon, Oxford.
Brassell S.C., Comet, P.A., Eglinton, G., Isaacson, P.J., McEvoy, J.,Maxwell, J.R., Thomson, I.D., Tibbetts, P.J.C., Volkman, J.K.,1980a. The origin and fate of lipids in the Japan Trench. In Advancesin Organic Geochemistry 1979 (eds. Douglas A.G., Maxwell J.R.),375-391, Pergamon, Oxford.
Brassell, S.C., Comet, P., Eglinton, G., McEvoy, J., Maxwell, J.R.,Quirke, J.M.E., Volkman, J.K. 1980b. Preliminary lipid Analyses ofCores 14,18, and 28 from Deep Sea Drilling Project Hole 416A. InInitial Reports of the Deep Sea Drilling Project, Vol. L (eds.Lancelot Y., Winterer E.L. et al.), pp. 647-664. U.S. GovernmentPrinting Office, Washington D.C.
Brassell, S.C., Wardroper, A.M.K., Thomson, I.D., Maxwell, J.R.,Eglinton, G., 1981. Specific acyclic isoprenoids as biologicalmarkers of methanogenic bacteria in marine sediments. Nature,290:693-696.
Brassell, S.C., Eglinton, G., 1983. Steroids and triterpenoids in deep-sea sediments as environmental and diagenetic indicators. InAdvances in Organic Geochemistry 1981 (Eds. Bjorøy, M., et al.),684-697, Wiley, Chichester.
Brassell, S.C., Howell, V.J., Gowar, A.P., Eglinton G., 1983a. Lipidgeochemistry of Cretaceous sediments recovered by the Deep SeaDrilling Project. In Advances in Organic Geochemistry 1981 (Eds.Bjorøy, M. et al.), 477-484. Wiley, Chichester.
Brassell, S.C., Eglinton, G., Maxwell, J.R., 1983b. The geochemistryof terpenoids and steroids. Biochemical Society Transactions 11,575-586.
Brassell, S.C., 1984. Aliphatic hydrocarbons of a Cretaceous blackshale and its adjacent green claystone from the southern AngolaBasin. Deep Sea Drilling Project, Leg 75. In Initial Reports of theDeep Sea Drilling Project, 75 (Pt. 2) (eds. Hay, W.W., Sibuet, J.C.,et al.), 1019-1030, U.S. Government Printing Office, WashingtonD.C.
Brassell, S.C., Eglinton, G., 1984. Lipid indicators of microbialactivity in marine sediments. In Heterotrophic Activity in the Sea(eds. Hobbie J. E. and Williams P. J.), 481-503, Plenum, New York.
Brassell, S.C., Farrimond, P., 1986. Fluctuations in biological markercompositions within a Cenomanian black shale from the Angolabasin. In Biogeochemistry of Black Shales (eds. Degens E.T., MeyersP.A., and Brassell S.C.) Mitt. Geol.-Palaontol. Inst. Univ. 60, 311-338.
Brassell, S.C., Lewis, C.A., de Leeuw, J.W., de Lange, F., SinningheDamste, J.S., 1986. Isoprenoid thiophenes: novel products ofsediment diagenesis?. Nature, 320, 160–162.
Brassell, S.C., Sheng Guoying, Fu Jiamo, Eglinton, G., 1988.Biological markers in lacustrine Chinese oil shales. In LacustrinePetroleum Source Rocks (eds. Fleet A.J., Kelts K., and Talbot M.R.),299-308, Blackwell, Oxford.
Brassell, S.C., Dumitrescu, M., and ODP Leg 198 Shipboard ScientificParty, (2004). Recognition of alkenones in a lower Aptianporcellanite from the west-central Pacific. Organic Geochemistry,35, 181-188.
Clarke, L.J., Jenkyns, H.C., 1999. New oxygen isotope evidence forlong-term Cretaceous climatic change in the Southern Hemisphere.Geology, 27, 699-702.
Comet, P.A., McEvoy, J., Brassell, S.C., Eglinton, G., Maxwell, J.R.,Thomson, I.D., 1981. Lipids of an Upper Albian limestone, DeepSea Drilling Project Site 465, Section 465A-38-3. In Initial Reportsof the Deep Sea Drilling Project, 62 (eds. Thiede J., Vallier T.L. etal.), 923-937, U.S. Government Printing Office, Washington D.C.
Dastillung, M., Albrecht, P., 1977. Δ 2-Sterenes as diageneticintermediates in sediments. Nature, 269, 678-679.
Dastillung, M., Albrecht, P., Ourisson, G., 1980a. Aliphatic andpolycyclic alcohols in sediments: Hydroxylated derivates of hopaneand of 3-methylhopane. Journal of Chemical Research (S), 168-169,(M) 2353-23.
Dastillung M., Albrecht, P., Ourisson, G., 1980b. Aliphatic andpolycyclic ketones in sediments: C27-C35 ketones and aldehydes ofthe hopane series. Journal of Chemical Research (S), 166-167, (M)2325-2352.
Dean, W.E., Claypool, G.E., Thiede, J., 1981 Origin of organic-carbon-rich mid-Cretaceous limestones, Mid-Pacific Mountains andSouthern Hess Rise. In Initial Reports of the Deep Sea DrillingProject, 62 (eds. Thiede J., Vallier T.L. et al.), 877-890, U.S.Government Printing Office, Washington D.C.
de Gea, G.A., Castro, J.M., Aguado, R., Ruiz-Ortiz, P.A., Company,M. 2003. Lower Aptian carbon isotope stratigraphy from a distalcarbonate shelf setting: the Cau section, Prebetic zone, SE Spain.Palaeogeography, Palaeoclimatology, Palaeoecology, 200, 207-219.
de Leeuw, J.W., van der Meer, F.W., Rijpstra, W.I.C., 1980. On theoccurrence and structural identification of long-chain unsaturatedketones and hydrocarbons in sediments. In Advances in OrganicGeochemistry 1979 (eds. Douglas A.G., Maxwell J.R.) 211-217,Pergamon Press, Oxford.
Didyk, B.M., Simoneit, B.R.T., Brassell, S.C., and Eglinton, G., 1978.Organic geochemical indicators of palaeoenvironmental conditionsof sedimentation: Nature, 272, 216-222.
Eglinton, T.I., Benitez-Nelson, B.C., Pearson, A., McNichol, A.P.,Bauer, J.E., Druffel, E.R.M., 1997. Variability in readiocarbon agesof individual organic compounds from marine sediments. Science,277, 796-799.
Ensminger, A., Albrecht, P., Ourisson, G., Tissot, B., 1977. Evolutionof polycyclic alkanes under the effect of burial (Early Toarcianshales, Paris Basin). In Advances in Organic Geochemistry 1975(eds. Campos R. and Gõni J.), 45-52, ENADIMSA, Madrid.
Erba, E., Tremolada, F., 2004. Nannofossil carbonate fluxes during theearly Cretaceous: phytoplankton response to nutrification episodes,atmospheric CO2, and anoxia. Paleoceanography, 19, PA1008.
Erbacher, J., Huber, B.T., Norris, R.D., Markey, M., 2001. Intensifiedthermohaline stratification as a possible cause for an ocean anoxicevent in the Cretaceous period. Nature, 409:325-327.
Espitalié, J., Laporte, J.L., Madec, M., Marquis, F., Leplat, P., Paulet,J., Boutefeu, A., 1977. Méthode rapide de caractérisation des rochesmères, de leur potential pétrolier et de leur degree d’évolution. Rev.Inst. Fr. Pet., 32, 23-42.
Espitalié, J., Deroo, G., Marquis, F., 1986. Rock-Eval pyrolysis and itsapplications: 3. Rev. Inst. Fr. Pet., 41, 73-89.
Farrimond, P., 1987. The Toarcian and Cenomanian/Turonian oceanicanoxic events: An organic geochemical study. Ph.D. Thesis,University of Bristol.
Farrimond, P., Eglinton, G., Brassell, S.C., 1986a. Geolipids of blackshales and claystones in Cretaceous and Jurassic sediment sequencesfrom the North American Basin. In North At lant icPalaeoceanography (eds. Summerhayes, C.P., Shackleton, N.J.), pp.347-360. Blackwell, Oxford.
Farrimond, P., Eglinton, G., and Brassell, S.C., 1986b, Alkenones inCretaceous black shales, Blake-Bahama Basin, western NorthAtlantic: Organic Geochemistry, 10, 897-903.
Farrimond, P., Eglinton, G., Brassell, S.C., Jenkyns, H.C., 1989.Toarcian anoxic event in Europe: an organic geochemical study.Marine and Petroleum Geology, 6, 136-147.
Farrimond, P., Eglinton, G., Brassell, S.C., Jenkyns, H.C., 1990. TheCenomanian/Turonian anoxic event in Europe: an organicgeochemical study. Marine and Petroleum Geology, 7, 75-89.
Farrimond P., Fox, P.A., Innes, H.E., Miskin, I.P., Head, I.M., 1998.Bacterial sources of hopanoids in recent sediments: Improving ourunderstanding of ancient hopane biomarkers. Ancient Biomolecules,2, 147-166.
Farrimond, P., Head, I.M., Innes, H.E., 2000. Environmental influenceon the biohopanoid composition of recent sediments. Geochimica etCosmochimica Acta, 64, 2985-2992.
Farrimond, P. Griffiths, T., Evdokiadis, E., 2002. Hopanoic acids inMesozoic sedimentary rocks: their origin and relationship withhopanes. Organic Geochemistry, 33, 965-977.

16 Biogeochemical assessment of paleoproductivity
16
Farrimond P., Love, G.D., Bishop, A.N., Innes, H.E., Watson D.F.,Snape, C.E., 2003. Evidence for the rapid incorporation of hopanoidsinto kerogen. Geochimica et Cosmochimica Acta, 67, 1383-1394.
Farquhar, G.D., Ehleringer, J.R. & Hubick, K.T., 1989a. Carbonisotope discrimination and photosynthesis. Annual Review of PlantPhysiology and Plant Molecular Biology, 40, 503-537.
Farquhar, D., Hubick, K.T., Condon, A.G., Richards, R.A., 1989b.Carbon Isotope Fractionation and Plant Water Use Efficiency. InStable Isotopes In Ecological Research. Ecological Studies, 68.Springer Berlin, pp. 525.
Francois, R., Altabet, M.A., Goericke, R., McCorkle, D.C., Brunet, C.,Poisson, A., 1993. Changes in the δ13C of surface water particulateorganic matter across the subtropical convergence in the SW IndianOcean. Global Biogeochemical Cycles, 7, 627-644.
Freeman, K.H., Wakeham, S.G., 1992. Variations in the distributionsand isotopic compositions of alkenones in Black-Sea particles andsediments. Organic Geochemistry, 19, 277-285.
Freeman K.H., Wakeham, S.G., Hayes, J.M., 1994. Predictive isotopicbiogeochemistry – hydrocarbons from anoxic marine basins.Organic Geochemistry, 21, 629-644.
Gagosian, R.B., Farrington, J.W., 1978. Sterenes in surface sedimentsfrom southwest African shelf and slope. Geochimica etCosmochimica Acta, 42, 1091-1101.
Gagosian, R.B., Smith, S.O., 1979. Steroids ketones in surfacesediments from the southwest African Shelf. Nature, 277, 287-289.
Gagosian, R.B., Smith, S.O., Lee, C., Farrington, J.W., Frew, N.M.,1980. Steroid transformations in Recent marine sediments. InAdvances in Organic Geochemistry 1979 (eds. Douglas A.G.,Maxwell J.R.) 407-419, Pergamon Press, Oxford.
Gagosian, R.B., Volkman, J.K., and Nigrelli, G.E., 1983. The use ofsediment traps to determine sterol sources in coastal sediments offPeru. In Bjoroy, M. (Ed.), Advances in Organic Geochemistry, 1981,369-379, Chichester, Wiley.
Goossens, H., de Leeuw, J.W., Schenck, P.A., Brassell, S.C., 1984.Tocopherols as likely precursors of pristane in ancient sediments andcrude oils. Nature 312, 440-442.
Grimalt, J.O., Angulo, L., López-Galindo A., Comas, M.C., andAlbaigés, J., 1990. Lipid and mineralogical composition of theCretaceous black shale deposits of the Fardes Formation (southernIberian Paleomargin, Betic Cordillera, south Spain). ChemicalGeology, 82, 341-363.
Gröcke, D.R., Hesselbo, S.P., Jenkyns, H.C., 1999. Carbon-isotopecomposition of Lower Cretaceous fossil wood: ocean-atmospherechemistry and relation to sea-level change. Geology, 27: 155-158.
Hayes, J.M., Freeman, K.H., Popp, B.N., and Hoham, C.H., 1990.Compound-specific isotopic analyses: A novel tool forreconstruction of ancient biogeochemical processes. OrganicGeochemistry, 16, 1115-1128.
Hayes, J.M., 1993. Factors controlling 13C contents of sedimentaryorganic compounds: Principles and evidence. Marine Geology, 113,111-125.
Hochuli, P.A., Menegatti, A.P., Weissert, H., Riva, A., Erba, E., andPremoli Silva, I., 1999. Episodes of high productivity and cooling inthe early Aptian Alpine Tethys. Geology, 27, 657-660.
Howard, D.L., Simoneit, B.R.T., and Chapman, D.J., 1984.Triterpenoids from lipids of Rhodomicrobium vannielli. Arch.Microbiol., 137, 200-204.
Huber, B.T., Norris, R.D., MacLeod, K.G., 2002. Deep-seapaleotemperature record of extreme warmth during the Cretaceous.Geology, 30, 123-126.
Innes, H., Bishop, A.N., Head, I.M., Farrimond, P., 1997. Preservationand diagenesis of hopanoids in Recent lacustrine sediments of PriestPot, England. Organic Geochemistry, 26, 565-576.
Innes, H.E., Bishop, A.N., Fox, P.A., Head, I.M., Farrimond, P., 1998.Early diagenesis of bacteriohopanoids in Recent sediments of LakePollen, Norway. Organic Geochemistry, 29, 1285-1295.
Jahren, A.H., Arens, N.C., Sarmiento, G., Guerrero, J., Amundson, R.,2001. Terrestrial record of methane hydrate dissociation in the EarlyCretaceous. Geology 29, 159-162.
Jaffe, R., Albrecht, P., Oudin, J.L., 1988. Carboxylic acids asindicators of oil migration- I. Occurrence and geochemicalsignificance of C-22 diastereoisomers of the (17αH,21βH) C30
hopanoic acid in geological samples. Organic Geochemistry, 13,483-488.
Jenkyns, H.C., 1995. Carbon-isotope stratigraphy andpaleoceanographic significance of the Lower Cretaceous shallow-water carbonates of Resolution Guyot, Mid-Pacific Mountains. InProc. ODP, Sci. Results, 143: (Eds. Winterer, E.L., Sager, W.W.,Firth, J.V., Sinton, J.M.), College Station, TX (Ocean DrillingProgram), 99-104.
Jenkyns, H.C., 1985. The early Toarcian and Cenomanian-Turoniananoxic events in Europe – comparisons and contrasts. GeologischeRundschau, 74, 505-518.
Jenkyns, H.C., 1988. The early Toarcian (Jurassic) anoxic event –stratigraphic, sedimenty, and geochemical evidence. AmericanJournal of Science, 288, 101-151.
Jenkyns, H.C., Gröcke, D.R., Hesselbo, S.P., 2001. Nitrogen isotopeevidence for water mass denitrification during the early Toarcian(Jurassic) oceanic anoxic event: Paleoceanography, 16, 593-603.
Joachimski, M.M., Ostertag-Henning, C., Pancost, R.D., Strauss, H.,Freeman, K.H., Littke, R., Sinninghe Damsté, J.S., Racki, G., 2001.Water column anoxia, enhanced productivity and concomitantchanges in δ13C and δ34S across the Frasnian-Famennian boundary(Kowala-Holy Cross Mountains/Poland). Chemical Geology, 175,109-131.
Joachimski, M.M., Pancost, R.D., Freeman, K.H., Ostertag-Henning,C., Buggisch, W., 2002. Carbon isotope geochemistry of theFrasnian-Famenian transition. Palaeogeography, Palaeoclimatology,Palaeoecology, 181, 91-109.
Kohnen, M.E.L., Sinninghe Damsté, J.S., Rijpstra, W.I.C., de Leeuw,J.W., 1990. Alkylthiophenes as sentsitive indiocators ofpalaeoenvironmental changes: a study of a Cretaceous oil shale fromJordan. In Geochemistry of Sulfur in Fossil Fuels (Eds. Orr, W.L.,White, C.M.) ACS Symposium Series 249, American ChemicalSociety, Washington, D.C., 444-485.
Köster, J., van Kaam-Peters, H.M.E., Koopmans, M.P., de Leeuw,J.W., Sinninghe Damste, J.S., 1997. Sulphurisation ofhomohopanoids: effects on carbon number distribution, speciation,and 22S/22R epimer ratios. Geochimica et Cosmochimica Acta, 61,2431-2452.
Kuypers, M.M.M., 2001. Mechanisms and biogeochemicalimplications of the mid-Cretaceous global organic carbon burialevents. Geologica Ultraiectina, 209, Universiteit Utrecht.
Kuypers M.M.M., Pancost, R.D., Sinninghe Damsté, J.S., 1999. Alarge and abupt fall in atmospheric CO2 concentrations duringCretacous times. Nature, 399, 342-345.
Kuypers M.M.M., Blokker, P., Erbacher, J., Kinkel, H., Pancost, R.D.,Schouten S., Sinninghe Damsté, J.S., 2001. Massive expansion ofmarine Archaea during a mid-Cretaceous oceanic anoxic event.Science, 293, 92-94.
Kuypers, M.M.M., Blokker, P., Hopmans, E.C., Kinkel, H., Pancost,R.D., Schouten S., Sinninghe Damste, J.S., 2002. Archaeal remainsdominate marine organic matter from the early Albian oceanicanoxic event 1b. Palaeogeography, Palaeoclimatology,Palaeoecology, 185, 211-234.
Leckie, R.M., Bralower, T.J., Cashman, R., 2002. Oceanic anoxicevents and plankton evolution: biotic response to tectonic forcingduring the mid-Cretaceous. Paleoceanography, 17, 1-29.
Lee, C., Gagosian, R.B., Farrington, J.W., 1980. geochemistry ofsterols in sediments from the Black Sea and the southwest Africanshelf and slope. Organic Geochemistry, 2, 103-113.
Luciani, V., Cobianchi, M., Jenkyns, H.C. 2001. Biotic andgeochemical response to anoxic events: the Aptian pelagic

Biogeochemical assessment of paleoproductivity 17
17
succession of the Gargano Promontory (southern Italy). Geol. Mag.138, 277-298.
Mackenzie, A.S., Brassell S.C., Eglinton G., Maxwell J.R., 1982.Chemical fossils: the geological fate of steroids. Science, 217, 491-504.
McEvoy, J., Giger, W., 1986. Origin of hydrocarbons in TriassicSerpiano oil shale - hopanoids. Organic Geochemistry, 10, 943-949.
McEvoy, J., Maxwell, J.R., 1983. Diagenesis of steroidal compoundsin sediments from the Southern California Bight (DSDP Leg 63, Site467). In Advances in Organic Geochemistry 1981 (Eds. Bjorøy, M.,et al.), 449-464, Wiley, Chichester.
Menegatti, A.P., Weissert, H., Brown, R.S., Tyson, R.V., Farrimond,P., Strasser, A., Caron, M., 1998. High resolution δ13C stratigraphythrough the early Aptian “Livello Selli” of the Alpine Tethys:Paleoceanography, 13, 530-545.
Mélières, F., Deroo, G., Herbin J.P., 1981. Organic-matter rich andhypersiliceous Aptian sediments from western Mid-PacificMountains, Deep Sea Dripping Project Leg 62. In Initial Reports ofthe Deep Sea Drilling Project, 62 (Pt. 2) (eds. Thiede J., Vallier T.L. et al.), 903-921, U.S. Government Printing Office, WashingtonD.C.
Meunier-Christmann, C., Albrecht, P., Brassell, S.C., ten Haven, H.L.,van der Linden, B, Rullkötter, J., Trendel, J.M., 1991. Occurrence ofdammar-13(17)-enes in sediments: indications for a yetunrecognized microbial constituent. Geochimica et CosmochimicaActa, 55, 3475-3483.
Meyers, P.A., Brassell, S.C., Huc, A.Y., 1984. Geochemistry oforganic carbon in South Atlantic sediments from Deep Sea DrillingProject Leg 75. In Initial Reports of the Deep Sea Drilling Project,75 (Pt. 2) (eds. Hay, W.W., Sibuet, J.C., et al.), 967-981, U.S.Government Printing Office, Washington D.C.
Meyers, P.A., 1987. Synthesis of organic geochemical studies, DSDPLeg 93, North American Continental Margin. In Initial Reports ofthe Deep Sea Drilling Project, 93, (eds. J.E. van Hinte, S.W. Wise etal.), 1333-1342, U.S. Government Printing Office, Washington D.C.
Meyers, P.A., 1994. Preservation of elemental and isotopic sourceidentification of sedimentary organic matter. Chemical Geology,114, 289-302.
Meyers, P.A., Doose, H., 1999. Sources, preservation, and thermalmaturity of organic matter in Pliocene-Pleistocene organic-carbon-rich sediments of the western Mediterranean Sea. In Proceedings,Ocean Drilling Program, Scientific Results, 161 (eds. R. Zahn, M.C.Comas, A. Kraus et al.), 383-390.
Nijenhuis, I.A., de Lange, G.J., 2000. Geochemical constraints onPliocene sapropel formation in the eastern Mediterranean. MarineGeology, 163, 41-63.
Norris, R.D., Wilson, P.A., 1998. Low-latitude sea-surfacetemperatures for the mid-Cretaceous and the evolution of plankticforaminifera. Geology, 26, 823-826.
Norris, R.D., Bice, K.L., Magno, E.A., Wilson, P.A., 2002. Jigglingthe tropical thermostat in the Cretaceous hothouse. Geology, 30,299-302.
Ohkouchi, N., Kawamura, K., Wada, E., Taira, A., 1997, Highabundances of hopanols and hopanoic acids in Cretaceous blackshale. Ancient Biomolecules, 1, 183-192.
Ourisson, G., Rohmer, M., 1992. The Hopanoids. Part 2: thebiohopanoids, a novel class of bacterial lipids. Accounts Chem. Res.25, 403-408.
Ourisson, G., Albrecht, P., Rohmer, M., 1979. The hopanoids:paleochemistry and biochemistry of a group of natural products.Pure Appl. Chem., 51:709-729.
Ourisson, G., Rohmer, M., Poralla, K., 1987. Microbial lipids betrayedby their fossils. Microbiol. Sci., 4:52-57.
Pagani, M., Arthur, M.A. Freeman, K.H., 1999. Miocene evolution ofatmospheric carbon dioxide. Paleoceanography 14, 273-292.
Pagani, M., Arthur, M.A., Freeman, K.H., 2000a. Variations inMiocene phytoplankton growth rates in the southwest Atlantic:Evidence for changes in ocean circulation. Paleoceanography, 15,
486-496.Pagani, M., Freeman, K.H., Arthur, M.A., 2000b. Isotope analyses of
molecular and total organic carbon from Miocene sediments.Geochimica et Cosmochimica Acta, 64, 37-49.
Pagani, M., 2002. The alkenone-CO2 proxy and ancient atmosphericcarbon dioxide. Philosophical Transactions of the Royal Society,London A., 360, 609-632.
Pancost, R.D., Freeman K.H., Wakeham, S.G., 1999. Controls on thecarbon-isotope compositions of compounds in Peru surface waters.Organic Geochemistry, 30, 319-340.
Peters, K.E., Moldowan, J.M., 1991. Effects of source, thermalmaturity and biodegradation on the distribution and isomerization ofhomohopanes in petroleum: Organic Geochemistry, 17, 47-61.
Popp, B.N., Laws, E.A., Bidigare, R.R., Dore, J.E., Hanson, K.L.,Wakeham, S.G., 1998. Effect of phytoplankton cell geometry oncarbon isotope fractionation. Geochimica et Cosmochimica Acta, 62,69-77.
Price, G.D., 2003. New constraints upon isotope variation during theearly Cretaceous (Barremian-Cenomanian) from the Pacific Ocean.Geological Magazine, 140, 513-522.
Raven, J.A., Johnston, A.M., 1991. Mechanisms of inorganic carbonacquisition in marine phytoplankton and their implications for theuse of other sources. Limnol. Oceanogr. 36, 1701-1714.
Raven, J.A., 1994. Carbon fixation and carbon availability in marinephytoplankton. Photosynthesis Research, 39, 259-273.
Retallack, G.J., 2001. A 300-million-year record of atmosphericcarbon dioxide from fossil plant cuticles, Nature, 411, 287-290.
Rhead, M.M., Eglinton, G., Draffen, G.H., 1971. Hydrocarbonsproduced by the thermal alteration of cholesterol under conditionssimulating the maturation of sediments. Chemical Geology, 8, 277-297.
Richnow, H.H., Jenisch, A., and Michaelis, W., 1992. Structuralinvestigations of sulphur-rich macromolecular oil fractions and akerogen by sequential chemical degradation. Organic Geochemistry,19, 351-370.
Riebesell, U., Burkhard, S., Dauelsberg, A., Kroon, B., 2000. Carbonisotope fractionation by a marine diatom: Dependence on the growthrate limiting resources. Marine Ecology Progress Series, 193, 295-303.
Robinson, N., Eglinton, G., Brassell, S.C., Cranwell, P.A., 1984.Dinoflagellate origin for sedimentary 4α-methylsteroids and 5α(H)-stanols: Nature, 308, 439-441.
Rodier, C., Llopiz P., Neunlist, S., 1999. C32 and C34 hopanoids inrecent sediments of European lakes: Novel intermediates in the earlydiagenesis of biohopanoids. Organic Geochemistry, 30, 713-716.
Rohmer, M., 1993. The biosynthesis of triterpenoids by the hopaneseries in Eubacteria: a mine of new enzyme reactions. Pure andApplied Chemistry 65, 1293-1298.
Rullkötter, J., Mukhopadhyay, P.K., Welte, D.H., 1984. Geochemistryand petrography of organic matter in sediments from Hole 530A,Angola Basin, and Hole 532, Walvis Ridge, Deep Sea DrillingProject. In Initial Reports of the Deep Sea Drilling Project, 75 (Pt. 2)(eds. Hay, W.W., Sibuet, J.-C., et al.), 1069-1087, Washington (U.S.Govt. Printing Office).
Rullkötter, J., Mukhopadhyay, P.K., Welte, D.H., 1987. Geochemistryand petrography of organic matter from Deep Sea Drilling ProjectSite 603, lower continental rise off Cape Hatteras. In Initial Reportsof the Deep Sea Drilling Project, 93 (Pt. 2) (eds. van Hinte, J.E.,Wise, S.W., Jr., et al.), 1163-1176, Washington (U.S. Govt. PrintingOffice).
Sakata, S., Hayes, J.M., McTaggart, A.R., Evans, R.A., Leckrone, K.J.,Togasaki, R.K., 1997. Carbon isotopic fractionation associated withlipid biosynthesis by a cyanobacterium: Relevance for interpretationof biomarker records. Geochimica et Cosmochimica Acta, 61, 5379-5389.
Schlanger, S.O., Jenkyns, H.C., 1976. Cretaceous oceanic anoxicevents: Causes and consequences. Geologie en Mijnbouw, 55,179–184.

18 Biogeochemical assessment of paleoproductivity
18
Schoell, M., Schouten S., Sinninghe Damsté, J.S., de Leeuw, J.W.,Summons, R.E., 1994. A molecular organic carbon isotope record ofMiocene climate change, Science, 263, 1122-1125.
Schouten, S., Schoell, M., Rijpstra, W.I.C., Sinninghe Damste, J.S., deLeeuw, J.W., 1997. A molecular and stable carbon isotope study oforganic matter in immature Miocene Monterey sediments (PismoBasin). Geochimica et Cosmochimica Acta, 61, 2065-2082.
Schouten, S., Hoefs, M.J.L., Sinninghe Damste, J.S., 2000a. Amolecular and stable carbon isotopic study of lipids in lateQuaternary sediments from the Arabian Sea. Organic Geochemistry,31, 509-521.
Schouten, S., Schoell, M., Summons, R.E., Sinninghe Damste, J.S., deLeeuw, J.W., 2000b. Molecular biogeochemistry of Montereysediments, Naples Beach, California, II: Stable carbon isotopiccompositions of free and sulfur-bound carbon skeletons. In TheMonterey Formation – From Rocks to Molecules (eds. Issacs, C.M.,Rullkötter, J.), 175-188, Columbia, New York.
Shiea, J., Brassell, S.C., Ward, D.M., 1990. Mid-chain branched mono-and dimethyl alkanes in hot spring cyanobacterial mats: A directbiogenic source for branched alkanes in ancient sediments? OrganicGeochemistry, 15, 223-231.
Simoneit, B.R.T., 1973. Identification of isoprenoidal ketones in DeepSea Drilling Project core samples and their geochemicalsignificance. In Initial Reports of the Deep Sea Drilling Project, 21(eds. Burns, R.E., Andrews, J.E., et al.), 909-923, U.S. GovernmentPrinting Office, Washington D.C.
Simoneit, B.R.T., 1978. Lipid analyses of sediments from Site 40-364in the Angola Basin. In Initial Reports of the Deep Sea DrillingProject, 40 Supplement (eds. Bolli, H., Ryan, W.B.F., et al.), 659-662, U.S. Government Printing Office, Washington D.C.
Simoneit, B.R.T., 1979a. Organic geochemistry of the shales from thenorthwestern Proto-Atlantic, DSDP Leg 43. In Initial Reports of theDeep Sea Drilling Project, 43 (eds. Tucholke, B.E., Vogt, P.R., etal.), 643-649, U.S. Government Printing Office, Washington D.C.
Simoneit, B.R.T., 1979b. Organic geochemistry of the Cretaceousblack shales from the Bay of Biscay, Site 402 and of Eocenemudstones from the Rockall Plateau, Site 404. In Initial Reports ofthe Deep Sea Drilling Project, 48 (eds. Montadert, L., Roberts, D.G.,et al.), 935-941, U.S. Government Printing Office, Washington D.C.
Simoneit, B.R.T., Mazurek, M.A., 1979a. Search for eolian lipids inthe Pleistocene off Cape Bojador and lipid geochemistry of aCretaceous mudstone, DSDP/IPOD Leg 47A. In Initial Reports ofthe Deep Sea Drilling Project, 47 Part A (eds. Ryan, W.B.F., vonRad, U., et al.), 541-545, U.S. Government Printing Office,Washington D.C.
Simoneit, B.R.T., Mazurek, M.A., 1979b. Lipid geochemistry ofCretaceous sediments from Vigo Seamount, DSDP/IPOD Leg 47B.In Initial Reports of the Deep Sea Drilling Project, 47 Part B (eds.Sibuet, J.-C., Ryan, W.B.F., et al.), 565-570, U.S. GovernmentPrinting Office, Washington D.C.
Simoneit B.R.T., 1981. Organic geochemistry of Albian sediment fromHess Rise, Deep Sea Drilling Project Hole 466. In Initial Reports ofthe Deep Sea Drilling Project, 62 (Pt. 2) (eds. Thiede J., Vallier T.L.et al.), 939-942, U.S. Government Printing Office, Washington D.C.
Simoneit, B.R.T., 1986. Biomarker geochemistry of black shales fromCretaceous oceans - An overview. Marine Geology, 70, 9-41.
Simoneit, B.R.T., and Stuermer, D.H., 1982. Organic geochemicalindicators for sources of organic matter and paleoenvironmentalconditions in Cretaceous oceans. In Nature and Origin of CretaceousCarbon-Rich Facies (eds. Schlanger, S.O., Cita, M.B.), 145-163,Academic, New York.
Simoneit, B.R., Grimalt, J.O., Wang, T.G., Cox, R.E., Hatcher, P.G.,Nissenbaum, A., 1986. Cyclic terpenoids of contemporary resinousplant detritus and of fossil woods, ambers and coals. OrganicGeochemistry, 10, 877-889.
Sinninghe Damste, J. S., de Leeuw, J. W., 1987. The origin and fate ofisoprenoid C20 and C15 sulphur compounds in sediments and crudeoils. International Journal of Environmental Analytical Chemistry,28, 1-19.
Sinninghe Damste, J. S., van Duin, A. C. T., Hollander, D., Kohnen,M. E. L., de Leeuw, J. W., 1995. Early diagenesis ofbacteriohopanepolyol derivatives: Formation of fossilhomohopanoids. Geochimica et Cosmochimica Acta, 59, 5141-5147.
Sinninghe Damste, J.S., Kuypers, M.M.M., Schouten S., Schulte, S.,Rullkötter, J., 2003. The lycopane/C31 n-alkane ratio as a proxy toassess paleoxicity during sediment deposition. Earth and PlanetaryScience Letters, 209, 215-226.
Sliter, W.V., 1989. Aptian anoxia in the Pacific Basin. Geology 17:909-912.
Summons, R.E., Jahnke, L.L., Hope, J. M., Logan G.A., 1999. 2-Methylhopanoids as biomarkers for cyanobacterial oxygenicphotosynthesis. Nature, 400, 554-557.
ten Haven, H.L., de Leeuw, J.W., Schenck, P.A., 1985. Organicgeochemical studies of a Messinian evaporitic basin, northernAppennines (Italy), I - Hydrocarbon biological markers for ahypersaline environment. Geochimica et Cosmochimica Acta, 49,2181-2191.
Valisolalao, J., Perakis, N., Chappe, B., Albrecht, P., 1984. A novelsulfur-containing C35 hopanoid in sediments. Tetrahedron Letters,25, 1183-1186.
van Breugel, Y., Schouten, S., Erba, E., Sinninghe Damsté, J.S., 2002.Biogeochemistry of the Aptian Selli event: Examination of thenegative isotope spike. Technical Program and Abstracts,Geochemistry Division, 223nd ACS National Meeting, Orlando,Florida (membership.acs.org/g/geoc/spring2002abstracts.pdf).
van Breugel, Y., Schouten, S., Erba, E., Price, G.D., SinningheDamsté, J.S., 2003. Molecular examination of the global negativecarbon isotope spike in Aptian sediments. 21st IMOG ConferenceAbstracts, Krakow, Poland.
van Graas, G., 1982. Organic geochemistry of Cretaceous black shaledeposits from Italy and France. Unpublished Thesis, TechnischeUniversiteit Delft, 95pp.
Volkman, J.K., Eglinton, G., Corner, E.D.S., Sargent, J.R., 1980.Novel unsaturated straight-chain C37-C39 methyl and ethyl ketones inmarine sediments and a coccolithophore Emiliania huxleyi. InAdvances in Organic Geochemistry 1979 (eds. Douglas A.G.,Maxwell J.R.) 219-227. Pergamon Press, Oxford.
Wakeham, S.G., Freeman, K.H., Pease, T.K., Hayes, J.M., 1993. Aphotoautotrophic source for lycopane in marine water columns.Geochimica et Cosmochimica Acta, 57, 159-165.
Wardroper, A.M.K., 1979. Aspects of the geochemistry of polycyclicisoprenoids. Ph.D. Thesis, University of Bristol.
Watson, D.F., and Farrimond, P., 2000. Novel polyfunctionalisedgeohopanoids in a recent lacustrine sediment (Priest Pot, UK).Organic Geochemistry, 31, 1247-1252.
Wilson, P.A., Norris, R.D., 1991. Warm tropical ocean surface andglobal anoxia during the mid-Cretaceous period. Nature 412, 425-429.
Wissler, L., Funk, H., Weissert, H., 2003. Response of EarlyCretaceous carbonate platforms to changes in atmospheric carbondioxide levels. P a l a e o g e o g r a p h y , Palaeoclimatology,Palaeoecology, 200, 187-205.
Xu, L., Reddy, C.M., Farrington, J.W., Frysinger, G.S., Gaines, R.B.,Johnson, C.G., Nelson, R.K., Eglinton, T.I., 2001. Identification of anovel alkenone in Black Sea sediments. Organic Geochemistry 32:633-645.
Zhao, N., Berova, N., Nakanishi, K., Rohmer, M., Mougenot, P.,Jürgens, U.J., 1996. Structure of two bacteriohopanoids with acyclicpentol side-chains from the cyanobacterium Nostoc PCC 6720.Tetrahedron, 52, 2777-2788.