binding of neomycin-class aminoglycoside antibiotics to the a …€¦ · between aminoglycoside...
TRANSCRIPT

Binding of Neomycin-class AminoglycosideAntibiotics to the A-site of 16 S rRNA
Dominique Fourmy, Michael I. Recht and Joseph D. Puglisi*
Center for Molecular Biology ofRNA, University of CaliforniaSanta Cruz, CA, 95064, USA
Aminoglycoside antibiotics that bind to ribosomal RNA in the aminoa-cyl-tRNA site (A-site) cause misreading of the genetic code and inhibittranslocation. We have recently solved the structure of an A-site RNA-paromomycin complex. The structure suggested that rings I and II, com-mon to all aminoglycosides that bind to the A-site, are the minimummotif for speciÆc ribosome binding to affect translation. This hypothesiswas tested biochemically and with a detailed comparative NMR study ofinteraction of the aminoglycosides paromomycin, neomycin, ribostamy-cin, and neamine with the A-site RNA. Our NMR data show that rings Iand II of neomycin-class aminoglycosides are sufÆcient to confer speci-Æcity to the binding of the antibiotics to the model A-site RNA. Neomy-cin, paromomycin, ribostamycin and neamine bind in the major grooveof the A-site RNA in a unique binding pocket formed by non-canonicalbase pairs and a bulged nucleotide. Similar NMR properties of the RNAand the diverse antibiotics within the different complexes formed withneomycin, paromomycin, ribostamycin and neamine suggest similarstructures for these complexes.
# 1998 Academic Press Limited
Keywords: aminoglycoside antibiotics; NMR; decoding region;RNA±ligand interaction; ribosomal RNA*Corresponding author
Introduction
The ribosome is the target of a class of structu-rally related antibiotics, the aminoglycosides (Galeet al., 1981). These antibiotics affect protein syn-thesis by inducing codon misreading and inhibit-ing translocation (Davies & Davis, 1968; Davieset al., 1965). Neomycin and related aminoglyco-sides bind directly to phylogenetically conservedsequences of 16 S ribosomal RNA (rRNA), whichpartially form the aminoacyl-tRNA (A site) andpeptidyl-tRNA (P site) binding sites in the 30 Sribosomal subunit (Moazed & Noller, 1986, 1990).The A site in the 30 S subunit is where aminogly-cosides bind to the ribosome, as determined bybiochemical and genetic methods. In particular,chemical modiÆcation experiments on the ribo-
some indicate that neomycin-class aminoglycosidesprotect nucleotides in the A-site from reaction withchemical probes (Moazed & Noller, 1987;Woodcock et al., 1991).Aminoglycoside antibiotics are composed of
amino sugars linked to a deoxystreptamine ring.The 2-deoxystreptamine-containing aminoglyco-sides, which have common functional groups onrings I and II (Figure 1), cause a characteristic mis-coding pattern, whereas streptomycin, which doesnot contain these conserved functional groups,induces a pattern of miscoding that is distinct fromthat of the neomycins (Davies et al., 1966). Mostaminoglycosides have a disubstituted 2-deoxy-streptamine (ring II). Ribostamycin, paromomycinand neomycin (Figure 1) contain additional ringson the 5 position of ring II, whereas gentamicinsand kanamycins contain an additional ring at pos-ition 6 of ring II. Despite a large number of amino-glycoside variants, there has been no structuralrationale for chemical synthesis of novel aminogly-cosides.The interaction between aminoglycosides and
16 S rRNA was recently characterized using anRNA fragment (27mer) containing the target sitefor these antibiotics (Figure 2) (Recht et al., 1996).
Present addresses: D. Fourmy, M. I. Recht and J. D.Puglisi, Department of Structural Biology, StanfordUniversity, School of Medicine, Stanford, CA 94305-5400, USA.Abbreviations used: A site, aminoacyl-tRNA site; P
site, peptidyl-tRNA site; NOE, Nuclear Overhauserenhancement; NOESY, NOE spectroscopy; COSY,Correlated spectroscopy.
J. Mol. Biol. (1998) 277, 347±362
0022±2836/98/120347±16 $25.00/0/mb971552 # 1998 Academic Press Limited

Aminoglycoside antibiotics bind to this modelRNA with similar afÆnity and speciÆcity as to 30 Ssubunits, indicating that the oligonucleotide con-tains the intact ribosomal binding site for amino-glycosides. The solution structure of this RNAfragment complexed to one of the aminoglycosides
(paromomycin) was solved by NMR (Fourmy et al.,1996). Paromomycin binds in the major groove ofthe RNA within a pocket created by anA1408 �A1493 base-pair and a single bulged adeno-sine (A1492). SpeciÆc interactions were observedbetween aminoglycoside chemical groups import-ant for antibiotic activity and conserved nucleo-tides in the RNA. Common chemical groupsamong A-site binding aminoglycosides on rings Iand II of paromomycin make sequence speciÆccontacts with the A-site RNA (Figure 3(a)). RingsIII and IV contact RNA non-speciÆcally by electro-static interactions with the phosphodiester back-bone (Figure 3(b) and Table 1). The structuresuggests that rings I and II are essential for speciÆcbinding of aminoglycosides to rRNA and that thecontribution of rings III and IV to the speciÆcity ofbinding is minor. This prediction is supported byexperimental data: neamine, which lacks rings IIIand IV (Figure 1), binds to 16 S rRNA and causesmiscoding (Woodcock et al., 1991). This suggeststhat the conserved rings, I and II, are the minimummotif for speciÆc binding of aminoglycosides tothe A site of the ribosome.To test the roles of rings I and II for speciÆc ami-
noglycoside binding to rRNA, we present here acomparative NMR and biochemical study of theinteraction of the aminoglycosides paromomycin,neomycin, ribostamycin, and neamine with the A-site RNA. The NMR properties and solution struc-ture of the paromomycin-RNA complex were usedas a reference. The data demonstrate that rings Iand II of neomycin-class aminoglycosides are sufÆ-cient for speciÆc binding of neomycin-class amino-glycosides to A-site RNA.
Figure 1. (a) Structures of the aminoglycoside anti-biotics. The A-site binding aminoglycosides all containrings I and II. The neomycin group of aminoglycosidesincludes paromomycin, neomycin, ribostamycin, andneamine. Ribostamycin contains all rings except ring IV,whereas neamine lacks both rings III and IV. (b) Thecommon structural elements among all aminoglycosideantibiotics that bind to the ribosomal A-site are in bold
Figure 2. Secondary structure of the A-site RNA asdetermined from NMR data (Recht et al., 1996). Thedashed lines represent canonical base-pairs and thecircles the non-canonical base-pairs.
348 Aminoglycoside Binding to A-site Oligonucleotide

Results
The A-site RNA-paromomycin complexstruc ture and its NMR charac teristics
The complete assignment by heteronuclear NMRof the RNA 1H, 13C and 31P resonances and theparomomycin 1H resonances (Fourmy et al., 1998)allowed collection of a set of NMR distance anddihedral restraints for the determination of the sol-ution structure of the 1 : 1 paromomycin-RNA com-plex (Fourmy et al., 1996). Sequential connectivities
between nucleotides in RNA were obtainedthrough the phosphodiester backbone by perform-ing HP-COSY and HCP experiments.Paromomycin binds in the major groove of the
A-site RNA, as indicated by intermolecular NOEs(Table 2). As shown in Figure 4(a), intermolecularNOEs are observed between U1495(H5) and paro-momycin ring II axial and equatorial protons andbetween U1490(H5) and the paromomycin ring IVprotons. These intermolecular NOEs orient ring IIclose to U1495 in the upper stem and ring IV in thevicinity of U1490 from the lower stem.Paromomycin contacts the bases and backbone
of the non-canonical base pairs within the internalloop (Figure 3(a)). The geometry of theU1406 �U1495 base-pair was deÆned by both intra-molecular RNA-RNA and intermolecular RNA-paromomycin NOEs: U1406(H3)-U1495(H3), U1406and U1495(H3)-paromomycin ring II (proton 6),and U1406 and U1495(H5/H6) to base or riboseprotons of the two adjacent base-pairsG1405 �C1496 and C1407 �G1494.The A1408 �1493 pair geometry was deÆned
using distance restraints between the H8/H2 pro-tons of A1408 and A1493 to base or ribose protons(H10) of the two adjacent base-pairs (C1407 �G1494and C1409 �G1491). The following distancerestraints were also used to deÆne the confor-mation of the A �A pair: A1493(H8)-protons of ringI (30,40,50), A1493(H8)-ring II(2-axial), A1493(H20)-paromomycin ring II(3), and A1493(H30)-ring II(2-equatorial, 3). In addition, NOE restraints derivedfrom 15N-correlated NOESY experiments forexchange-broadened protons (Mueller et al., 1995)were essential to deÆne the A �A pair geometry:A1408(NH2)-A1493(H8), A1408(NH2)-A1493(NH2),A1408(N6)-ring I(10) and A1408(N6)-ring II(6). TheNOESY experiments were performed on the com-plex containing speciÆcally 15N, 13C-labeled adeno-sines at 5 and 15�C with different mixing times(180 and 250 ms).Paromomycin bound to the A-site RNA adopts
an L-shaped conformation. Rings II, III, and IVform a linear array that lines the major groovefrom the U1406 �U1495 base-pair to theA1410 �U1490 base-pair. Ring I is positioned nearthe A1408 �A1493 pair and A1492 at approximatelya 90� angle to rings II, III, and IV. Rings I and IIadopt chair conformations, with the amino andhydroxyl substituents in equatorial positions, andthese exocyclic groups make speciÆc contacts thatstabilize the antibiotic-RNA complex (Figure 3(a)).Antibiotic-RNA interaction is extended along the
major groove by rings III and IV. The amino andhydroxyl groups of ring IV make probable electro-static contacts with the phosphate backbone atU1406, C1407, and U1490 (Figure 3(b)). The Ænalstructure statistics for the ensemble of 20 structures(see Table 1 in Fourmy et al. (1998)) illustrate therelative disorder of rings III and IV. This disorderarises from qualitatively different dynamic proper-ties of rings I and II and rings III and IV: the reson-ances of rings III and IV have narrower line widths
Figure 3. (a) SpeciÆc contacts made between rings I andII of paromomycin and the A-site RNA. The view isinto the major groove of the RNA core. The RNA is inblue, paromomycin is tan. Important chemical groupsare shown explicitly. The U1406 �U1495, A1408 �A1493base-pairs, as well as G1491 and G1494 are highlightedin the structure. Possible hydrogen bonding contacts areindicated by dashed lines. (b) Contacts made betweenrings III and IV of paromomycin and the A-site RNA.The view is looking down the helix axis of the upperstem into the RNA core. Possible hydrogen bondingcontacts are indicated by dashed lines.
Aminoglycoside Binding to A-site Oligonucleotide 349

Table 1. Statistics of intermolecular contacts identiÆed in the Ænal 20 simulated annealing structures of the A-site RNA-paromomycin complex
Intermolecular contacts:U1495(O4)-ring II (N1)
G1494(N7)-ring II (N6)
A1493(OP)-ring I (C6)a
A1493(OP)-ring I (O40)
A1493(OP)-ring II (N1)
A1492(OP)-ring I (O30)
G1491(N7)-ring III (O500)
U1490(OP)-ring IV (N2000)
U1406(OP)-ring IV (O3000)
U1406(OP)-ring IV (N2000)
C1407(OP)-ring IV (O3000)
Average distance (A ) 3.07 3.40 4.45 3.74 3.39 3.94 3.51 3.62 3.13 3.69 3.65Standard deviation 0.15 0.06 0.48 0.24 0.30 1.07 0.56 1.35 0.15 0.50 0.78
a The orientation of the 60 hydrogen bond donor with respect to the RNA is determined by the dihedral angle about the ring I C50-C60 bond, but is not well-deÆned in the solution structure.Nonetheless, the distance between the ring I C60, which is well positioned in the structure, and the pro-R oxygen of A1493 phosphate (4.4(⌃0.5) A ) is consistent with a direct contact between the60 hydroxyl group of ring I and the phosphate group of A1493.

than those of rings I and II. Rings III and IV arethought to contribute weakly to the speciÆcity ofantibiotic binding and function (Benveniste &Davies, 1973).To characterize speciÆc binding of paromomycin
to the A-site RNA, the chemical shifts of RNA res-onances were monitored as a function of antibioticconcentration (Figure 5(a)). A 1 : 1 complex isformed between the A-site RNA and paromomycin(Figure 5(a)), as described and discussed (Rechtet al., 1996). The proton chemical shift assignmentsof the A-site RNA complexed to paromomycinwere already presented (Fourmy et al., 1998). Uponbinding of paromomycin, the aromatic protonchemical shifts of residues U1406 to A1408 andG1491 to U1495 change at least 0.1 p.p.m. betweenthe free and bound form. On formation of the 1:1paromomycin-RNA complex, the A1408(H2) reson-ance shifts upÆeld 1 p.p.m. and broadens and theA1492 31P resonance shifts downÆeld at least0.8 p.p.m. (Fourmy et al., 1998). The G1494(H8)proton becomes extremely broad. In contrast to thedata from the free RNA, no internucleotide NOEconnectivities are observed from A1492(H8) andA1493(H8).Detailed heteronuclear NMR study indicated a
small change in RNA structure on paromomycinbinding and provided evidence for the stabilizationof the conformation of the A1492 and A1493 resi-dues upon binding of paromomycin. This wasclearly demonstrated by resolution of the three-dimensional structure of the free form of the A-siteRNA and its comparison to the bound form(Fourmy et al., 1998).The conformation of paromomycin is likewise
affected by binding to the RNA. The non-exchangeable protons of paromomycin free in sol-ution and bound to the A-site RNA were assigned(Table 3); the antibiotic was not isotopicallylabeled. Once paromomycin is bound to the A-siteRNA, proton resonances of rings I and II becomebroad, indicating conformational exchange on theNMR experimental time scale (⇠103 seconds�1).The 20 and 4 protons of rings I and II, respectively,were particularly broad and only detected in shortmixing time TOCSY (12 ms) and NOESY (50 ms)
experiments. In contrast, the sharp proton reson-ances of rings III and IV indicate more rapiddynamic processes for these two rings. Upon bind-ing to the RNA, 20 30 and 50 protons of ring I wereshifted upÆeld 0.8, 0.22, and 0.63 p.p.m., respect-ively (Table 3). Proton chemical shifts of ring IIwere not strongly affected upon binding to theRNA, except a 0.67 p.p.m. downÆeld shift for pro-ton 4 (Table 3). Proton chemical shifts of ring IIIand IV were weakly modiÆed by binding to theRNA. The 3000 proton was shifted downÆeld by0.43 p.p.m. and the 4000 proton were shifted upÆeldby 0.39 p.p.m. (Table 3).The paromomycin-A-site RNA structure with its
NMR characteristics is a reference for comparisonwith other neomycin-class aminoglycoside-A-siteRNA complexes. SpeciÆc interactions wereobserved between chemical groups of rings I andII important for antibiotic activity and conservednucleotides in the RNA (Figure 3(a)). The role ofaminoglycoside rings in RNA binding was testedbiochemically and by NMR.
Binding of neomy cin-class aminogl ycoside s tothe A-site oligonucleo tide
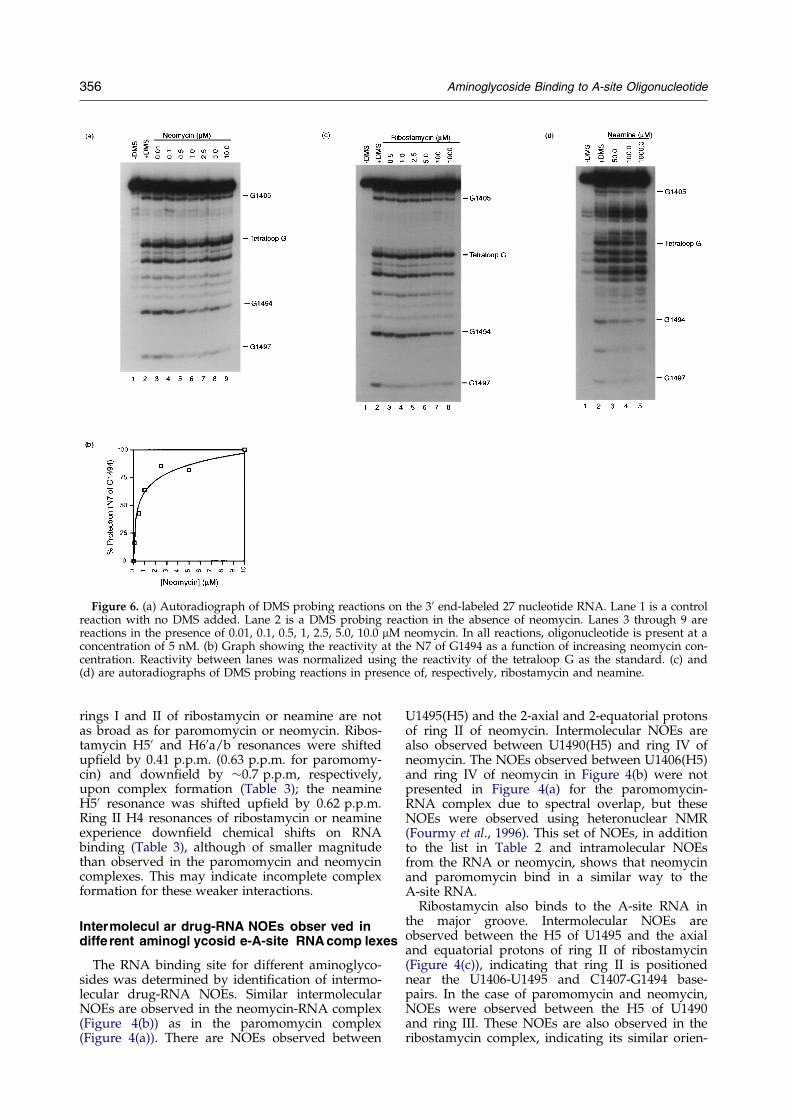
Binding of neomycin, ribostamycin and neamineto the A-site oligonucleotide was assayed bychemical probing with DMS at pH 7.0. It wasshown previously that in the presence of boundparomomycin, residues G1405, A1408 and G1494are strongly protected from chemical modiÆcationby DMS, whereas G1491 and G1497 are weaklyprotected and a Kd of 0.2 mM was measured byquantitative footprinting experiments (Recht et al.,1996). A similar footprint pattern for neomycinwas observed for G1494 and G1497 in the A-siteoligonucleotide (Figure 6(a)) and a Kd of 0.5 mMwas observed by quantitative footprinting exper-iments (Figure 6(b)). Neomycin binds with similarafÆnity to 30 S subunits (Moazed & Noller, 1987).Chemical probing experiments were performed
on the 27 nucleotide A-site RNA in the presence ofribostamycin (Figure 6(c)). A weak footprint at theN7 of G1494 could be observed on the A-site oligo-nucleotide at 100 mM or 1 mM ribostamycin, con-
Table 2. List of intermolecular NOEs observed for different complexes between the A-site RNA and paromomycin,neomycin, ribostamycin and neamine
Paromomycin-RNA NOEs used forstructure calculations
U1406(H5)-ring IV(1000,2000), U1406(H3)-ring II(6), U1406(H6)-ring IV(4000), C1407(H4)-ringII(6), ring III(200), C1407(H5)-ring IV(1000,2000), A1408(H6)-ring I(10) ring II(6), G1489(H8)-ring IV(4000), U1490(H5)-ring III(300, 400, 500) ring IV(3000, 4000, 5000, 6000), U1490(H6)-ring II(500),G1491(H1)-ring I(10), G1491(H20)-ring I(30, 40), G1491(H30)-ring I(20, 30), A1492(H30)-ringI(30, 40, 50), A1492(H50)-ring I(30), A1493(H20)-ringII(3), A1493(H30)- ringII(2 equatorial, 3),A1493(H50)-ring II(3), A1493(H8)-ring I(30, 40, 50) ring II(2 axial), G1494(H1)-ring II(2axial, 2 equatorial, 6), G1494(H30)-ring II(2 axial, 2 equatorial), U1495(H5)-ring II(1, 2axial, 2 equatorial, 3,6), U1495(H3)-ringII(6).
Neomycin-RNA NOEs U1406(H5)-ring III(5000), U1406(H5)-ring IV(2000, 4000, 6000), U1490(H5)-ring III(500) ringIV(4000, 6000), U1495(H5)-ringII(2 axial, 2 equatorial).
Ribostamycin-RNA NOEs U1490(H5-ring III(500), U1495(H5)-ringII(2 axial, 2 equatorial, 3)Neamine-RNA NOEs U1406(H5)- ringI(30, 40), U1406(H5)- ring II(2 axial, 2 equatorial (at pH 5.5), 1 or 3),
U1495(H5)- ringII(2 axial, 2 equatorial (at pH 5.5), 1, or 3, 5)
Intermolecular NOEs common between paromomycin and neomycin, ribostamycin or neamine are highlighted in bold.
Aminoglycoside Binding to A-site Oligonucleotide 351

sistent with a weaker afÆnity for the RNA. Thesame level of protection of G1494 was observedat 100 mM or 1 mM ribostamycin. This weakintensity of protection prevents any precise deter-mination of the Kd of ribostamycin for the A-siteRNA. Nevertheless, saturation of the signal for
100 mM and 1 mM ribostamycin indicates that theKd of ribostamycin can be estimated at 5 to 10 mM,which is a factor of ten above the value measured forparomomycin and neomycin. There are no literaturereports of ribostamycin footprint on 30 S subunits.For neamine, a weak footprint at position G1494 was
Figure 4(a)±(d) (legend opposite)
352 Aminoglycoside Binding to A-site Oligonucleotide

observed at 100 mM at 4�C (Figure 6(d)). Woodcocket al. (1991) observed a weak protection of G1494upon binding of neamine to the ribosome at a con-centration of 100 mM.The binding of ribostamycin to 30 S subunits was
assayed by chemical probing with DMS at pH 7.2.A weak footprint in the A-site of 16 S rRNA wasobserved at G1494(N7) and G1491(N7) in the pre-sence of 100 mM ribostamycin (Figure 7(a)). Thefootprint is similar to that observed for neamine(Woodcock et al., 1991), but is much weaker thanthat observed for the binding of neomycin or paro-momycin to 30 S subunits (Moazed & Noller, 1987).The protection of 16 S rRNA from reaction withDMS in the presence of ribostamycin and neamineis weaker, and the concentration of antibioticrequired to see the footprint is higher, than for theaminoglycosides that contain four rings. The resultson 30 S subunits and 27 nt RNA agree.The speciÆcity of ribostamycin and neamine
binding to the A site was examined by performingchemical modiÆcation experiments on A-site oligo-nucleotides (Recht et al., 1996) corresponding to thewild-type, U1406! A, and U1495! A sequencevariants (Figures 7(b±d) and 8(a±c)). Paromomycinbinds with wild-type afÆnity to the A1406 variant,but the A1495 mutation signiÆcantly decreases par-omomycin afÆnity. A weak footprint with ribosta-mycin, similar to that observed for 30 S subunits,is seen on the wild-type (Figure 7(b)) andU1406! A (Figure 7(c)) oligonucleotides at 10 mMribostamycin. As was observed for the 30 S sub-unit, the footprint at the N7 of G1494 in the pre-sence of ribostamycin is not as strong as in the caseof either paromomycin or neomycin. A weak foot-print is observed at the N7 of G1489 upon additionof 10 mM ribostamycin, but this footprint is notobserved in the 30 S subunit (Figure 8(a±c)). Ribos-
tamycin produces only a very weak footprint onthe U1495! A oligonucleotide at 1 mM antibiotic(Figure 7(d)). The same speciÆcity was also foundfor neamine (Figure 8(a±c)).
Complex forma tion betwe en the A-site anddifferent neom ycin-clas s aminoglyc osides
To characterize the speciÆc binding of aminogly-cosides to the A-site RNA, chemical shifts of iminoproton RNA resonances were monitored as a func-tion of antibiotic concentration (Recht et al., 1996)(Figure 5(a±d)). Neomycin forms a 1 : 1 complexwith the A-site RNA oligonucleotide. The iminoprotons of U1490 and G1491 are shifted downÆeld(0.2 p.p.m and 0.5 p.p.m, respectively) to a similarextent as in the paromomycin complex(Figure 5(b)). The slow exchange regime observedbetween the free and bound forms of the RNA isconsistent with a Kd of neomycin for the RNA oli-gonucleotide similar to the one found for paromo-mycin (Recht et al., 1996).For ribostamycin and neamine, the U1490 and
G1491 imino resonances were shifted downÆeld by0.2 p.p.m. These two resonances are increasinglyshifted upon addition of ribostamycin (Figure 5(c))or neamine (Figure 5(d)) until a 1 : 1 drug : RNAratio was reached. This is characteristic of a fastexchange regime, consistent with a lower afÆnity ofribostamycin or neamine for the RNA oligonucleo-tide.The same secondary structural elements of the
A-site RNA were affected by binding of either par-omomycin, neomycin, ribostamycin or neamine.A detailed NMR study of these different complexeswas then performed.
Figure 4. Portion of two-dimen-sional NOESY spectra in D2Oshowing RNA-antibiotic NOEs for:(a) the paromomycin-RNA com-plex; (b) the neomycin-RNA com-plex; (c) the ribostamycin-RNAcomplex; and (d) the neamine-RNAcomplex. The RNA:aminoglycosidestoichiometry is 1 : 1 for all fourcases. The intermolecular NOEsbetween the H5 of U1490 and thering IV of paromomycin or neomy-cin or ring III of ribostamycin areboxed with a solid line. The inter-molecular NOEs between the H5 ofU1495 and the ring II of paromo-mycin, neomycin, ribostamycin andneamine are also boxed with asolid line. In the complex with nea-mine, the intermolecular NOEs
between the H5 of U1406 and the ring II of neamine are boxed. For the neamine-RNA complex, NOEs betweenU1406(H5) and neamine ring I are also indicated by solid boxes. Antibiotic-intramolecular NOEs are indicated with abroken line. NOESY experiments were performed at 35�C, in 10 mM Na phosphate at pH 6.4 with a mixing time of250 ms.
Aminoglycoside Binding to A-site Oligonucleotide 353

RNA chemical shift changes in RNA onbinding of the aminoglycosides
The non-exchangeable and exchangeable protonsof the RNA complexed to neomycin, ribostamycinand neamine were assigned (data not shown).Chemical shift modiÆcations occur in the sameregions of the RNA upon formation of complexeswith all the aminoglycosides tested with someslight differences.The RNA chemical shift differences between the
free and bound forms of the neomycin-RNA com-plex are similar to what was observed with paro-momycin. The G1494(H8) resonance becomes
broad, like in the case of paromomycin. TheA1492(31P) resonance shifts 0.3 p.p.m downÆeld(data not shown). The A1408(H2) resonance isprobably very broad and could not be observed.This resonance was broad in the paromomycin-A-site RNA complex, but the A1408(H2) proton wasdetected at 45�C. At 45�C, the A1408(H2) reson-ance in the neomycin-A-site RNA complex was notdetected.The U1406! A variant forms a high afÆnity
complex with paromomycin. A1408(H2) is shiftedupÆeld in the paromomycin-A1406 RNA complex,and the resonance is sharper than in the wild-typecomplex. Neomycin also binds with wild-type afÆ-
Figure 5. (a) Titration of A-site oligonucleotide with paromomycin yields a speciÆc 1 : 1 complex. Imino proton spec-tra for the free RNA, 0.5 : 1 paromomycin : RNA and for the 1 : 1 complex. Imino proton resonances that change sig-niÆcantly upon binding of paromomycin are indicated (Recht et al., 1996). The same titration is shown with theantibiotics neomycin (b), ribostamycin (c) and neamine (d). Titration experiments were performed at 25�C at pH 6.4in 10 mM sodium phosphate.
354 Aminoglycoside Binding to A-site Oligonucleotide

nity to the A1406 variant, and a 1 : 1 complex wasformed for NMR characterization. Indeed, A1408(H2) is sharp and shifted by 0.9 p.p.m upÆeld,indicating a similar behavior of the paromomycinand neomycin complexes.Upon formation of the ribostamycin-RNA com-
plex, aromatic 1H chemical shifts are observed forthe same nucleotides affected by paromomycinbinding. However, smaller chemical shift changeswere detected; for instance, A1408(H2) resonanceshifts only 0.06 p.p.m. upÆeld, but still becomesvery broad. G1494(H8) broadens, as upon paromo-mycin binding. The A1492(31P) resonance does notshift signiÆcantly downÆeld from the helical phos-phate region (data not shown).For the neamine-RNA complex, the aromatic 1H
chemical shift differences between the free andbound form are observed in the same region of theRNA as for paromomycin, but are smaller or evenabsent; the A1408(H2) resonance does not shift, butG1494(H8) becomes broad as in the paromomycincomplex. The A1492(31P) resonance does not shiftsigniÆcantly downÆeld from the helical phosphateregion, similar to its behavior in the ribostamycin-RNA complex, (data not shown).In summary, 1H chemical shift changes in the A-
site RNA indicate a qualitatively similar bindingsite for paromomycin, neomycin, ribostamycin andneamine. For ribostamycin and neamine, thechemical shift modiÆcations are weaker. Since the
same region of the RNA is affected by binding ofthe neomycin-class aminoglycosides, this suggestsa common binding site of the drugs to the A-siteRNA albeit with different afÆnities.
Chemical shift changes in neom ycin-cla ssaminog lycosid es on binding to the RNA
The non-exchangeable protons of neomycin,ribostamycin, and neamine free and bound to theRNA were assigned (Table 3).The 1H chemical shifts of neomycin bound to the
A-site RNA are very similar to those of paromomy-cin. The proton resonances of rings I and II arevery broad, whereas resonances from rings III andIV are sharp. In particular, H20 and H4 resonancesare very broad, as in the paromomycin complex.Upon binding to the RNA, the H20 and H30 reson-ances of ring I were shifted upÆeld 0.17 and0.48 p.p.m. respectively, which is a smaller shiftthan the one observed for paromomycin H20
(0.82 p.p.m.). The H4 proton of ring II shifts down-Æeld 0.70 p.p.m. (0.67 for paromomycin) uponRNA binding (Table 1). Similar changes in ring IVH3000 and H4000 resonances were observed in theneomycin and paromomycin complexes (Table 3).Qualitatively similar chemical shift changes were
observed for ribostamycin and neamine resonancesupon RNA binding as for aminoglycosides withfour rings. In the bound form, resonances from
Table 3. Non-exchangeable proton chemical shifts (p.p.m.) at 35�C for free forms of paromomycin, ribostamycin andneamine and when they are complexed to the A-site oligonucleotide
Paromomycin Ribostamycin Neamine
Free Bound(free-bound)
Neomycinbound
(free-bound) Free Bound
(free-bound) Free Bound
(free-bound)
Ring I10 6.0 5.95 ⇠0 6.12 �0.12 5.97 6.02 ⇠0 5.80 5.91 �0.1120 3.42 2.6 0.82 3.25 0.17 3.38 3.38 0 3.33 3.47 �0.1430 4.00 3.78 0.22 (3.52) 0.48 3.98 4.09 0.11 3.90 4.06 �0.1640 3.25 3.25 0 n.d. ± 3.48 3.17 0.31 3.46 3.25 0.2150 4.08 3.45 0.63 n.d. ± 4.06 3.65 0.41 4.07 3.45 0.6260 ab 3.25/3.5 (3.25) ⇠0 n.d. ± 3.26/3.48 4.09 �0.79 3.21/3.26 n.d. ±Ring II1 3.32 3.34 ⇠0 3.35 ⇠0 3.28 3.34 ⇠0 3.28 3.35 ⇠02eq 2.36 2.37 ⇠0 2.32 ⇠0 2.32 2.42 �0.1 2.32 2.47 �0.162ax 1.79 2.07 �0.28 1.87 ⇠0 1.70 1.86 �0.16 1.64 1.94 �0.343 3.30 3.44 �0.14 3.44 �0.14 3.23 3.4 �0.17 3.21 3.49 �0.284 3.95 4.62 �0.67 (4.65) �0.7 3.81 4.16 �0.35 3.62 3.96 �0.345 3.95 3.9 ⇠0 3.96 ⇠0 3.86 3.95 ⇠0 3.53 3.71 �0.186 3.74 3.77 ⇠0 3.78 ⇠0 3.66 3.71 ⇠0 3.52 3.61 ⇠0Ring III100 5.43 5.53 �0.1 5.57 �0.14 5.40 5.40 0 ± ± ±200 4.48 4.24 0.24 4.32 0.16 4.25 4.18 ⇠0 ± ± ±300 4.55 4.38 0.17 4.38 0.17 4.21 4.14 ⇠0 ± ± ±400 4.25 4.42 �0.17 (4.40) �0.15 4.06 4.04 ⇠0 ± ± ±500 ab 3.95 3.85 0.1 3.78 0.17 3.71/3.93 3.87/3.92 ⇠0 ± ± ±Ring IV1000 5.30 5.3 ⇠0 5.3 ⇠0 ± ± ± ± ± ±2000 3.60 3.64 ⇠0 3.61 ⇠0 ± ± ± ± ± ±3000 3.85 4.28 �0.43 4.30 �0.45 ± ± ± ± ± ±4000 4.25 3.86 0.39 3.86 0.39 ± ± ± ± ± ±5000 4.38 4.32 ⇠0 4.39 ⇠0 ± ± ± ± ± ±6000 ab 3.45 3.36/3.43 ⇠0 3.37/3.44 ⇠0 ± ± ± ± ± ±
For neomycin, only the bound form is indicated. () indicates tentative assignment. N. D.: not determined. For neomycin, the chemi-cal shifts of the free paromomycin were used as reference for the free antibiotic form. The large modiÆcations are in bold.
Aminoglycoside Binding to A-site Oligonucleotide 355
rings I and II of ribostamycin or neamine are notas broad as for paromomycin or neomycin. Ribos-tamycin H50 and H60a/b resonances were shiftedupÆeld by 0.41 p.p.m. (0.63 p.p.m. for paromomy-cin) and downÆeld by ⇠0.7 p.p.m, respectively,upon complex formation (Table 3); the neamineH50 resonance was shifted upÆeld by 0.62 p.p.m.Ring II H4 resonances of ribostamycin or neamineexperience downÆeld chemical shifts on RNAbinding (Table 3), although of smaller magnitudethan observed in the paromomycin and neomycincomplexes. This may indicate incomplete complexformation for these weaker interactions.
Intermolecul ar drug-RNA NOEs obser ved indiffe rent aminogl ycosid e-A-site RNAcomp lexes
The RNA binding site for different aminoglyco-sides was determined by identiÆcation of intermo-lecular drug-RNA NOEs. Similar intermolecularNOEs are observed in the neomycin-RNA complex(Figure 4(b)) as in the paromomycin complex(Figure 4(a)). There are NOEs observed between
U1495(H5) and the 2-axial and 2-equatorial protonsof ring II of neomycin. Intermolecular NOEs arealso observed between U1490(H5) and ring IV ofneomycin. The NOEs observed between U1406(H5)and ring IV of neomycin in Figure 4(b) were notpresented in Figure 4(a) for the paromomycin-RNA complex due to spectral overlap, but theseNOEs were observed using heteronuclear NMR(Fourmy et al., 1996). This set of NOEs, in additionto the list in Table 2 and intramolecular NOEsfrom the RNA or neomycin, shows that neomycinand paromomycin bind in a similar way to theA-site RNA.Ribostamycin also binds to the A-site RNA in
the major groove. Intermolecular NOEs areobserved between the H5 of U1495 and the axialand equatorial protons of ring II of ribostamycin(Figure 4(c)), indicating that ring II is positionednear the U1406-U1495 and C1407-G1494 base-pairs. In the case of paromomycin and neomycin,NOEs were observed between the H5 of U1490and ring III. These NOEs are also observed in theribostamycin complex, indicating its similar orien-
Figure 6. (a) Autoradiograph of DMS probing reactions on the 30 end-labeled 27 nucleotide RNA. Lane 1 is a controlreaction with no DMS added. Lane 2 is a DMS probing reaction in the absence of neomycin. Lanes 3 through 9 arereactions in the presence of 0.01, 0.1, 0.5, 1, 2.5, 5.0, 10.0 mM neomycin. In all reactions, oligonucleotide is present at aconcentration of 5 nM. (b) Graph showing the reactivity at the N7 of G1494 as a function of increasing neomycin con-centration. Reactivity between lanes was normalized using the reactivity of the tetraloop G as the standard. (c) and(d) are autoradiographs of DMS probing reactions in presence of, respectively, ribostamycin and neamine.
356 Aminoglycoside Binding to A-site Oligonucleotide

tation within the major groove with ring III nearthe A1410-U1490 base-pair.Neamine binds more weakly to the A-site RNA,
and the complex is in fast exchange with the freeform; nonetheless, intermolecular NOEs areobserved (Figure 4(d)). NOEs between the H5 ofU1406 and U1495 and the 2-axial proton of ring IIof neamine were observed at 35�C, pH 6.4. At
pH 5.5, the NOE between U1495(H5) and the 2-equatorial proton of ring II was detected. InFigure 4(d) are also displayed NOEs between theH5 of U1406 and U1495 and proton 3 of ring II ofneamine. Intermolecular NOEs of ring II protons ofneamine are observed with U1495(H5) andU1406(H5), whereas only NOEs to U1495(H5)were observed with paromomycin. This suggests
Figure 8. (a) Autoradiograph ofDMS probing reactions on thewild-type 44 nucleotide A-siteRNA in the presence of 0, 1, 10,100 and 1000 mM neamine. (b) and(c) are the same reactions, respect-ively, on the U1406! A variantoligonucleotide and on theU1495! A variant oligonucleotide.Reaction conditions were the sameas described in Figure 7.
Figure 7. (a) Autoradiograph of DMS probing reactions on 30 S ribosomal subunits. As indicated on the Ægure, ribos-tamycin was present at concentrations of 1, 10, 100, and 1000 mM. Subunits were present at a concentration of100 nM in all reactions. Bands corresponding to nucleotides G1489, G1491 and G1494 are indicated. (b) Autoradio-graph of DMS probing reactions on the wild-type 44 nucleotide A-site RNA in the presence of 0, 1, 10, 100 and1000 mM ribostamycin. The oligonucleotide was present at a concentration of 75 nM in all reactions. Lanes G and Aare dideoxy sequencing reactions. Nucleotide positions that show changes in reactivity upon addition of 10 mM ribos-tamycin are indicated. (c) Autoradiograph of DMS probing reactions on the U1406! A variant oligonucleotide in thepresence of 0, 1, 10, 100 and 1000 mM ribostamycin. The oligonucleotide was present at a concentration of 75 nM inall reactions. Nucleotide positions that show changes in reactivity upon addition of 10 mM ribostamycin are indicated.(d) Autoradiograph of DMS probing reactions on the U1495! A variant oligonucleotide in the presence of 0, 1, 10,100 and 1000 mM ribostamycin. The oligonucleotide was present at a concentration of 75 nM in all reactions. Nucleo-tide positions that show changes in reactivity upon addition of 1000 mM ribostamycin are indicated.
Aminoglycoside Binding to A-site Oligonucleotide 357

that neamine may be sampling multiple bindingsites within the A-site major groove. At pH 6.4 and35�C, NOEs between U1406(H5) and the protons 30
and 40 of ring I of neamine were detected, butthese were not detected at pH 5.5.These NMR results indicate speciÆc binding of
neomycin, ribostamycin and neamine in the majorgroove of the A-site RNA, similar to what wasobserved for paromomycin. The orientations ofparomomycin, neomycin, and ribostamycin arequalitatively similar with ring II contacting theU1406 �U1495 base-pair and rings III and IV (whenpresent) close to the A1410 �U1490 base-pair of thelower stem.
A common conform ation of the complexesbetween the A-site RNA and aminogl ycosidescont aining rings I and II
Binding of paromomycin stabilizes a speciÆcconformation for A1408, A1492 and A1493(Fourmy et al., 1998). A1492 and A1493 are dis-placed towards the minor groove by interaction ofring I of paromomycin within a speciÆc bindingpocket. NMR data indicate the same conformation-al change with neomycin. Although the overallconformations of the ribostamycin and neamine-A-site RNA complexes are similar, the NMR datasuggest that the RNA conformational equilibriumis shifted towards the free conformation.Interresidue NOEs within paromomycin were
crucial to deÆne the geometry of the antibioticbound to the RNA (Figure 3) (Fourmy et al., 1996).Two strong NOEs (in the 1.8 to 2.5 A range) werevery important to deÆne the position of rings I andIII relative to ring II: ring I(10)-ring II(4) and ringIII(100)-ring II(5). For neomycin in the RNA-neomy-cin complex, the two same strong NOEs wereobserved. For neamine in the RNA-neamine com-plex, a strong ring I(10)-ring II(4) NOE wasobserved. In the RNA-drug complex, the rings Iand II of paromomycin have a chair conformation(Fourmy et al., 1996). The same chair conformationwas found for rings I and II of ribostamycin andring II of neomycin (data not shown).In summary, the conformation of the A-site
RNA-neomycin-class aminoglycosides are verysimilar with only small differences in the RNA con-formation observed with ribostamycin and nea-mine. The solution structures of the A-site RNAcomplexed to neomycin, ribostamycin and nea-mine were not determined, since it would requireisotopic labeling and extensive heteronuclear NMRanalysis.
Discus sion
The different neom ycin-cla ss aminogly cosidescan be used to probe the role of each ring inthe specifi c binding to the A-site RNA
In the solution structure of the A-site RNA com-plexed to paromomycin (Fourmy et al., 1996),
speciÆc interactions were observed between chemi-cal groups of rings I and II of aminoglycosides andconserved nucleotides in the RNA (Figure 3(a)).Rings III and IV make non-sequence-speciÆc elec-trostatic interactions with the phosphate backboneof the RNA (Figure 3(b)). Neomycin, ribostamycinand neamine represent a family of aminoglycosideswith four, three and two rings, respectively, whichall contain the critical rings I and II for RNA bind-ing (Figure 1). The current work tested the role ofindividual rings of paromomycin in RNA bindingafÆnity and speciÆcity.
Different affinities of the neomyc in-classaminog lycosid es for the A-site RNA
A greater number of rings in the aminoglycosideantibiotics increases their afÆnity to the A-siteRNA. The dissociation constant (Kd) for the neo-mycin-A-site RNA complex, measured by quanti-tative footprinting, was 0.5 mM, which is similar tothe value found for paromomycin (Recht et al.,1996). The Kd was estimated to be 5 to 10 mM forribostamycin and approximately 100 mM for nea-mine interaction with the A-site RNA. NMRtitrations of the A-site RNA with ribostamycin andneamine conÆrmed that the afÆnity of ribostamy-cin and neamine for the A-site RNA is reduced,based on different exchange behavior. Footprintingon mutant oligonucleotides demonstrated thespeciÆcity of ribostamycin and neamine interactionwith A-site RNA. The U1495! A mutation dis-rupts the ring II hydrogen bond from paromomy-cin, and signiÆcantly decreases binding afÆnity.A similar effect was observed for ribostamycin andneamine. These results indicate speciÆc targeting ofat least ring II of these antibiotics to the A-site.The results with the oligonucleotide agree with
the approximate dissociation constants for amino-glycoside±ribosome interactions determined byfootprinting (Moazed & Noller, 1987). A completeprotection was observed for 5mM paromomycin orneomycin and 100 mM ribostamycin (this work).Woodcock et al. (1991) used a concentration of100 mM neamine to observe the protections on the16 S rRNA. A factor of about Æve- to tenfold wasobserved between the minimal inhibitory concen-tration of neamine (40 mg/ml) and neomycin(5 mg/ml), paromomycin (10 mg/ml) or ribostamy-cin (10 mg/ml) (DeStasio & Dahlberg, 1990). RingsIII and IV of paromomycin or neomycin accountfor a small part of the afÆnity of the neomycin-class aminoglycosides for the ribosome.
Cons erved rings I and II of neomyc in-classaminog lycosid es are essen tial for speci ficbinding to the A-site RNA
Qualitative NMR data conÆrm the differentbinding afÆnities of the neomycin family of amino-glycosides for A-site RNA. Neomycin binds with
358 Aminoglycoside Binding to A-site Oligonucleotide

similar afÆnity as paromomycin, with the RNA-antibiotic complex in slow exchange with the freeform. Very similar changes in both RNA and ami-noglycoside chemical shifts were observed in theparomomycin and neomycin complex. Most ami-noglycoside antibiotics have a 60-NH2 group onring I, and the change of the 60-OH group to a60-NH2 group has little effect on aminoglycosidebinding afÆnity. Ribostamycin binds more weaklyto the model oligonucleotide, with the complex infast exchange with the free form. Although similarchemical shift changes were observed as in theneomycin and paromomycin complexes, their mag-nitudes were smaller. This may indicate incompletespeciÆc complex formation at 1 : 1 stoichiometry inthe NMR experiment. Finally, neamine binds mostweakly to the RNA, with complex and free RNAin fast exchange. As with the ribostamycin com-plex, the magnitudes of NMR chemical shiftchanges on neamine binding were smaller than forneomycin or paromomycin binding.Distance and torsion angle restraints collected
for the A-site RNA-paromomycin complex werecompared to the NMR data for the neomycin,ribostamycin and neamine complexes with A-siteRNA. Intermolecular NOEs between each of theseaminoglycosides and the RNA demonstrate thatthey speciÆcally bind in the major groove of theRNA in the same site as paromomycin. The orien-tation of ribostamycin, paromomycin and neomy-cin within the major groove was found to be thesame. Rings III and IV of neomycin are close to theA1410-U1490 base-pair, as observed with the samerings of paromomycin. Ring III of ribostamycin isalso close to this A1410-U1490 base-pair. The con-served ring II of paromomycin, neomycin, ribosta-mycin and neamine is close to the unusual U1406-U1495 base-pair. U1495 is required for speciÆcinteraction with these aminoglycosides, asmutation to A1495 disrupts interaction with ami-noglycosides. In the paromomycin-A-site RNAcomplex, the 1-amino group on ring II forms ahydrogen bond with the O4 of U1495.The lower afÆnity interaction of neamine with
the A-site RNA results in a more dynamic RNA-drug complex. Several intermolecular NOEs wereobserved between ring I of neamine and
U1406(H5). This is different from the complex withparomomycin where it is ring III of paromomycinwhich is in proximity of U1406(H5). These NOEsin the neamine complex may arise from rotation ofring I around the glycosidic linkage to ring II orfrom a reversed binding mode of rings I and II,which places ring I close to U1406 instead ofA1493 and A1492 (Figure 9). The amino groups 1and 3 in ring II are approximately symmetricallyrelated to the 2-axial and equatorial positions andare hydrogen bonded to U1495(O4) andG1494(N7), respectively, in the paromomycin-RNAcomplex (Figure 3(a)). Rings III and IV of the neo-mycin class aminoglycosides confer a directionalspeciÆcity to antibiotic binding, assuring a correctinteraction of rings I and II with the RNA. The pre-ponderance of aminoglycoside antibiotics withthree rings may reØect this aspect of RNA inter-action.Rings I and II of neomycin-class aminoglyco-
sides are sufÆcient to direct their binding to aunique site on the model A-site RNA. The presenceof a 60-OH or NH2 group on ring I does not affectthe interaction of aminoglycosides within theirRNA binding pocket. Deletion of ring IV decreasesby about tenfold the afÆnity of aminoglycoside-RNA complex formation, by eliminating possibleelectrostatic contacts between ring IV and the heli-cal stem of the RNA. Further deletion of ring IIImaintains binding, but afÆnity is further decreasedÆve- to tenfold. For the aminoglycosides such asneamine that bind with weaker afÆnity, NMR datasuggest a major population of speciÆc complexwithin the paromomycin binding site, but withpossible alternate conformations that representnon-speciÆc complexes.Binding of paromomycin or neomycin to A-site
RNA stabilizes the conformations of A1408, A1492and A1493 within the asymmetric internal loop.Interaction of ring I in the aminoglycoside bindingpocket displaces A1492 and A1493 towards theminor groove. In the ribostamycin and neamine-A-site RNA complexes, the conformational equili-brium is shifted towards the free A-site confor-mation, and there is evidence for multiple bindingmodes for neamine. Additional rings of ribostamy-cin, neomycin and paromomycin slightly increase
Figure 9. (a) Schematic of the con-formation of neamine within thebinding pocket. This conformationcorresponds to the structure of ringI and II in the A-site RNA-paromo-mycin complex (Fourmy et al.,1996). The amino groups at pos-itions 1 and 3 of ring II makehydrogen bonds to the O4 ofU1495 and the N7 of G1494,respectively. (b) Alternate bindingmode of neamine in the binding
pocket as indicated by the NMR data (NOEs of ring I of neamine with U1406(H5)). In this conformation, the N1 andN3 positions of ring II are hydrogen bonded to G1494(N7) and U1495(O4) respectively, in a reversed orientation com-pared to (a).
Aminoglycoside Binding to A-site Oligonucleotide 359

the afÆnity of the antibiotic for the A-site of theribosome. This correlates with their in vivo activity.They might also increase the speciÆcity of inter-action of rings I and II with the RNA. Additionalrings could also help in the correct orientation ofrings I and II by creating additional drug-RNA ordrug-drug contacts (Fourmy et al., 1996). Betterunderstanding of the role of aminoglycoside ringsin RNA binding is helpful in the development ofnew antibiotics.
Materials and Methods
Preparation of the A-site oligonucleotide RNAfor NMR
Milligram quantities of the A-site RNA (27 nucleo-tides) were prepared unlabeled, uniformly 13C labeled orwith adenosines speciÆcally 13C and 15N labeled byin vitro transcription from an oligonucleotide templateand puriÆed as described (Puglisi & Wyatt, 1995). Afterelectroelution and ethanol precipitation, the resuspendedRNA was then dialyzed against the buffer used for theNMR experiment in a microdialysis apparatus with a3500 MW cut-off membrane.
Proton and heteronuclear NMR for the A-site RNA
All the NMR experiments were recorded on a VarianUnityá 500 MHz spectrometer equipped with triple res-onance, z-gradient probes. NMR data were processedusing Varian or Felix software. NMR experiments wereperformed in 10 mM sodium phosphate (pH 6.4 or 5.5)at 25, 35 and 45�C, unless otherwise indicated. The con-centrations of unlabeled RNA, uniformly13C-labeledRNA, and speciÆcally 15N, 13C labeled RNA were4.5 mM, 3 mM and 3 mM, respectively. 15N chemicalshifts were indirectly referenced compared to the nitro-gen chemical shift of 15NH4
áC1� in HCl (Levy & Lichter,1979). 13C chemical shifts were referenced to the knownchemical shifts of the ribose carbons from the tetraloop(Varani & Tinoco, 1991).
For the unbound form of the RNA (without antibiotic)and for the paromomycin-RNA complex, exchangeableand non-exchangeable protons of the unlabeled RNAand non-exchangeable protons of the paromomycin wereÆrst partially assigned using standard NMR exper-iments. Temperature was varied to help the protonassignment. Rings I and II of paromomycin and neomy-cin gave broad resonances and, therefore, their protonswere assigned using different mixing times TOCSY (10,25, 70 and 100 ms) combined with various temperatures(25, 35 and 45�C). The assignment of the non-exchange-able protons of the labeled RNA and paromomycin wascompleted using constant-time HSQC (Santoro & King,1992), three-dimensional HCCH-TOCSY, three-dimen-sional HMQC-NOESY (Clore et al., 1990) and 13C ÆlteredNOESY and TOCSY experiments (Lee et al., 1994; Otting& Wu» thrich, 1990). Sequential connectivities betweennucleotides in RNA were obtained by HP-COSY (Skle¬naret al., 1986) and HCP experiments (Marino et al., 1994b).The H2 protons of the four adenines were unambigu-ously assigned for the labeled RNA by correlation of theH2/H8 resonances in a two-dimensional HCCH-TOCSYexperiment (Marino et al., 1994a). The hydrogen bondingpatterns of the base-pairs were determined from analysis
of SSNOESY spectra in H2O at different mixing times(75, 150 and 300 ms) (Smallcombe, 1993).
RNA titration by aminoglycosides
The free form of the A-site RNA (3 mM) was titratedby adding 5 ml of a concentrated solution of paromomy-cin, neomycin, ribostamycin or neamine. The antibioticwas dissolved in 10 mM sodium phosphate (pH 6.4), thesame buffer as used for the RNA sample. The titrationswere performed at 25�C.
Exchangeable proton NMRspectra of the antibiotic-RNA complexes studied
One-dimensional NMR spectra of exchangeable pro-tons from the RNA were obtained in 9/1 H2O/
2H2O at5, 10 and 25�C, and water suppression was achievedusing a binomial (1331) pulse sequence. Two-dimen-sional NMR experiments (SSNOSEY) (Smallcombe, 1993)were recorded at 5�C with 75 and 300 ms mixing times.
Non-exchangeable proton NMRof the freeform antibiotics
Standard NOESY, DQF-COSY and TOCSY exper-iments in 99.996% 2H2O were acquired. NOESY exper-iments were acquired with mixing times of 50 and250 ms. The same solution conditions as for the RNA-antibiotic complexes were used.
Non-exchangeable proton NMRspectra ofantibiotic ±RNA complexes
Most of the NMR experiments were recorded on aVarian Unityá 500 MHz spectrometer. Some exper-iments performed on the neomycin-RNA complex wereacquired on a Bruker 750 MHz spectrometer at theNuclear Magnetic Resonance Facility at Madison. Stan-dard NOESY, 31P decoupled DQF-COSY and TOCSYexperiments in 99.996% D2O were acquired. NOESYexperiments were acquired with mixing times of 50, 100,150, 200 and 400 ms. NMR experiments were performedin 10 mM sodium phosphate (pH 6.4 and pH 5.5) at anRNA concentration of 4.5 mM at 25, 35 and 45�C unlessotherwise indicated.
For the neomycin±RNA, ribostamycin±RNA, andneamine±RNA complexes, the exchangeable and non-exchangeable protons of the RNA and non-exchangeableprotons of the aminoglycosides were partially or totallyassigned using standard proton NMR experiments withno use of isotopic labeling. When necessary for theassignment, natural abundance 13C HSQC were acquiredon these antibiotic±RNA complexes.
Chemical modification
ModiÆcation reactions used 30 end-labeled 27 nt A-siteRNA. Chemical modiÆcation reactions (300 ml) wereperformed in 80 mM potassium cacodylate, pH 7.0,with 5 nM RNA oligonucleotide. Antibiotics (Sigma)were added and modiÆcation was performed byaddition of DMS (15 ml of a 1/10 dilution in ethanol)followed by incubation at room temperature for Æveminutes. Reactions were stopped by ethanol precipi-tation. Sodium borohydride reduction and aniline-induced strand scission was performed as described(Recht et al., 1996). ModiÆed RNA was resuspended in
360 Aminoglycoside Binding to A-site Oligonucleotide

10 ml 1 M Tris-HCl (pH 8.2). Upon addition of 10 ml offreshly prepared 0.2 M NaBH4, the samples were incu-bated on ice in the dark for 30 minutes. The reactionwas quenched by addition of 100 ml 0.4 M NaOAc fol-lowed by ethanol precipitation. Pellets were dissolvedin 20 ml 1.0 M aniline/acetate (pH 4.5), followed byincubation in the dark for 20 minutes at 60�C. Thereaction was terminated by addition of 100 ml 0.4 MNaOAc and 100 ml phenol/chloroform/isoamyl alcohol(25 : 24 : 1) followed by vigorous mixing and centrifu-gation. The RNA was concentrated by ethanol precipi-tation of the aqueous phase and pellets were washedwith 100 ml cold 70% ethanol.
ModiÆcation reactions (100 ml) with 30 S subunits(10 pmol) were performed in a buffer containing80 mM potassium cacodylate (pH 7.2), 100 mMammonium chloride, 20 mM magnesium chloride,1 mM dithiothreitol, and 0.5 mM EDTA. Followingactivation of the subunits by incubation at 42�C for 20minutes in the buffer described, ribostamycin wasadded. The subunits and antibiotic were incubated at37�C for 30 minutes followed by incubation for anotherten minutes on ice. DMS (2 ml, 1 : 6 in ethanol) wasadded, and the reaction was incubated at 37�C for tenminutes. Following addition of 25 ml DMS stop sol-ution (1 M Tris acetate, pH 7.5, 1.5 M sodium acetate,1 M b-mercaptoethanol), the subunits were precipitatedwith ethanol. The subunits were suspended in 200 ml0.3 M sodium acetate (pH 5.5). ModiÆed RNA wasobtained by extraction of 30 S subunits three timeswith phenol and twice with chloroform. The RNA wasprecipitated with ethanol and sodium borohydridereduction and aniline-induced strand scission were per-formed as described above. Primer extension of the16 S rRNA was performed as described using a DNAprimer that is complementary to nucleotides 1530 to1509 of 16 S rRNA (Moazed et al., 1986; Stern et al.,1988).
ModiÆcation reactions (40 ml) with the 44-nucleotideA-site RNA were performed as described (Recht et al.,1996) in 80 mM potassium cacodylate (pH 7.2) with3 pmol RNA oligonucleotide. After heating the RNAat 90�C for one minute and immediately cooling onice for Æve minutes, ribostamycin or neamine wasadded. The RNA and antibiotic were incubated atroom temperature (ribostamycin) or on ice (neamine)for Æve minutes. DMS (2 ml, 1 : 10 in ethanol) wasadded and the reaction incubated at room temperaturefor Æve minutes (ribostamycin) or 75 minutes on ice(neamine). DMS stop solution (10 ml) was added andsodium borohydride reduction and aniline-inducedstrand scission were performed. Primer extension wasperformed with the modiÆcations described (Rechtet al., 1996).
Modification data analysis
Gels were typically exposed on Ælm for 12 to 24hours. ModiÆcation data was quantiÆed by exposingthe gel on a Molecular Dynamics PhosphorImagerscreen and analyzed using Molecular DynamicsImageQuant software.
For quantiÆcation of the data, the total number of cpmloaded per lane was normalized. Additionally, to nor-malize for the efÆciency of strand scission, the cleavageat G (N7) of the tetraloop was used as the standard.
Acknowledgments
The authors thank Professor H. Noller and the mem-bers of his laboratory for support and discussion. Thiswork was supported by grants from N.I.H. (GM51266-01A1), Packard Foundation, Deafness Research Foun-dation, and Lucille P. Markey Charitable Trust. D.F. wassupported by a grant from INSERM (Institut National dela Sante¬ et de la Recherche Meƒdicale). The NMR facilityat UCSC is supported by a grant from Lucille P. MarkeyCharitable Trust. We also thank the Nuclear MagneticResonance Facility at Madison for use of their Bruker750 MHz spectrometer.
References
Benveniste, R. & Davies, J. (1973). Structure-activityrelationships among the aminoglycoside antibiotics:role of hydroxyl and amino groups. Antimicrob. Ag.Chemother. 4, 402±409.
Clore, G. M., Bax, A., Driscoll, P. C., WingÆeld, P. T. &Gronenborn, A. M. (1990). Assignment of the side-chain 1H and 13C resonances of Interleukin-1b usingdouble and triple-resonance heteronuclear three-dimensional NMR spectroscopy. Biochemistry, 29,8172±8184.
Davies, J. & Davis, B. D. (1968). Misreading of ribonu-cleic acid code words induced by aminoglycosideantibiotics. J. Biol. Chem. 243, 3312±3316.
Davies, J., Gorini, L. & Davis, B. D. (1965). Misreadingof RNA codewords induced by aminoglycosideantibiotics. Mol. Pharmacol. 1, 93±106.
Davies, J., Jones, D. S. & Khorana, H. G. (1966).A further study of misreading of codons inducedby streptomycin and neomycin using ribopolynu-cleotides containing two nucleotides in alternatingsequence as templates. J. Mol. Biol. 18, 48±57.
DeStasio, E. A. & Dahlberg, A. E. (1990). Effects ofmutagenesis of a conserved base-paired site nearthe decoding region of Escherichia coli 16 S riboso-mal RNA. J. Mol. Biol. 212, 127±133.
Fourmy, D., Recht, M. I., Blanchard, S. C. & Puglisi, J. D.(1996). Structure of the A site of E. coli 16 S rRNAcomplexed with an aminoglycoside antibiotic.Science, 274, 1367±1371.
Fourmy, D., Yoshizawa, S. & Puglisi, J. D. (1998).Paromomycin binding induces a local conformation-al change in the A-site of 16 S rRNA. J. Mol. Biol.277, 333±345.
Gale, E. F., Cundliffe, E., Reynolds, P. E., Richmond,M. H. & Waring, M. J. (1981). The Molecular Basis ofAntibiotic Action, Wiley, London.
Lee, W., Revington, M. J., Arrowsmith, C. & Kay, L. E.(1994). A pulsed Æeld gradient isotope-Æltered 3D13C HMQC-NOESY experiment for extracting inter-molecular NOE contacts in molecular complexes.FEBS Letters, 350, 87±90.
Levy, G. C. & Lichter, R. L. (1979). Nitrogen-15 nuclearmagnetic resonance spectroscopy. In Nitrogen-15Nuclear Magnetic Resonance Spectroscopy, Wiley, NewYork.
Marino, J. P., Prestegard, J. H. & Crothers, D. M.(1994a). Correlation of adenine H2/H8 resonancesin uniformly 13C labeled RNAs by 2D HCCH-TOCSY: a new tool for 1H assignment. J. Am. Chem.Soc. 116, 2205±2206.
Marino, J. P., Schwalbe, H., Anklin, C., Bermel, W.,Crothers, D. M. & Griesinger, C. (1994b). A three-
Aminoglycoside Binding to A-site Oligonucleotide 361

dimensional triple-resonance 1H, 13C, 31P exper-iment: sequential through-bond correlation of riboseprotons and intervening phosphorus along theRNA oligonucleotide backbone. J. Am. Chem. Soc.116, 6472±6473.
Moazed, D. & Noller, H. F. (1986). Transfer RNA shieldsspeciÆc nucleotides in 16 S ribosomal RNA fromattack by chemical probes. Cell, 47, 985±994.
Moazed, D. & Noller, H. F. (1987). Interaction of anti-biotics with functional sites in 16 S ribosomal RNA.Nature, 327, 389±394.
Moazed, D. & Noller, H. F. (1990). Binding of tRNA tothe ribosomal A and P sites protects two distinctsets of nucleotides in 16 S rRNA. J. Mol. Biol. 211,135±145.
Moazed, D., Stern, S. & Noller, H. F. (1986). Rapidchemical probing of conformation in 16 S ribosomalRNA and 30 S ribosomal subunits using primerextension. J. Mol. Biol. 187, 399±416.
Mueller, L., Legault, P. & Pardi, A. (1995). ImprovedRNA structure determination by detection of NOEcontacts to exchange-broadened amino protons.J. Am. Chem. Soc. 117, 11043±11048.
Otting, G. & Wu» thrich, K. (1990). Heteronuclear Æltersin two-dimensional [1H-1H]-NMR spectroscopy:combined use with isotope labelling for studies ofmacromolecular conformation and intermolecularinteractions. Quart. Rev. Biophys. 23, 39±96.
Puglisi, J. D. & Wyatt, J. R. (1995). Biochemical andNMR studies of RNA conformation with an empha-
sis on RNA pseudoknots. Methods Enzymol. 261,323±350.
Recht, M. I., Fourmy, D., Blanchard, S. C., Dahlquist,K. D. & Puglisi, J. D. (1996). RNA sequence deter-minants for aminoglycoside binding to an A-siterRNA model oligonucleotide. J. Mol. Biol. 262, 421±436.
Santoro, J. & King, G. (1992). A constant-time 2D over-bodenhausen experiment for inverse correlation ofisotopically enriched species. J. Magn. Reson. 97,202±207.
Skle¬nar, V., Miyashiro, H., Zon, G. & Bax, A. (1986).Assignment of the 31P and 1H resonances in oligo-nucleotides by two-dimensional NMR spectroscopy.FEBS Letters, 208, 94±98.
Smallcombe, S. H. (1993). Solvent suppression with sym-metrically-shifted pulses. J. Am. Chem. Soc. 115,4776±4785.
Stern, S., Moazed, D. & Noller, H. F. (1988). Structuralanalysis of RNA using chemical and enzymaticprobing monitored by primer extension. MethodsEnzymol. 164, 481±489.
Varani, G. & Tinoco, I. J. (1991). Carbon assignments andheteronuclear coupling constants for an RNA oligo-nucleotide form natural abundance 13C-1H correlatedexperiments. J. Am. Chem. Soc. 113, 9349±9354.
Woodcock, J., Moazed, D., Cannon, M., Davies, J. &Noller, H. F. (1991). Interaction of antibiotics withA and P-site-speciÆc bases in 16 S ribosomal RNA.EMBO J. 10, 3099±3103.
Edited by I. Tinoco
(Received 22 August 1997; received in revised form 17 November 1997; accepted 17 November 1997)
362 Aminoglycoside Binding to A-site Oligonucleotide