bidirectional small-intestinal permeability in the rat to some common marker molecules in vitro
TRANSCRIPT

Bidirectional Small-Intestinal Permeability in the Rat to Some Common Marker Molecules in vitro N. PANTZAR, S. LUNDIN, L. WESTER & B. R. WESTROM Depts. of Animal Physiology and Clinical Pharmacology, University of Lund, Lund, Sweden
Pantzar N, Lundin S , Wester L, Westrom BR. Bidirectional small-intestinal permeability in the rat to some common marker molecules in vitro. Scand J Gastroenterol 1994;29:703-709.
Background: The barrier properties of the small intestine were investigated by studying the bidirectional permeability to five commonly used marker molecules. Methods: Proximal and distal small-intestinal segments from rats were mounted in diffusion chambers, and the permeation of the markers 3H-mannitol (Mw 182), 51Cr-ethylenediaminetetraacetic acid (Mw 341), [mercaptopropionic acid', ~-arginine*]- vasopressin (Mw 1069), fluorescein isothiocyanate (F1TC)-dextran (mean Mw 3000), and inulin (Mw 5200) was measured across the mucosa in both directions. Results: A generally increased inward (mucosa to serosa) and a decreased outward (serosa to mucosa) permeation of the markers was found in the proximal to distal direction. The inward permeability showed increasing regional differences with decreasing sue of the markers. In the absence of the villous epithelium, removed by scraping the intestinal wall, 86% to 62% of the proximal and distal barrier was lost in the inward direction but only 14% to 26% in the outward direction. Conclusions: The intestinal epithelial barrier is more permeable in the outward than in the inward direction, and regional permeability differences exist in a size- dependent fashion. The results suggest two passage routes, one for the smallest molecule, mannitol, and a second for the larger markers in the present size range, both apparently different from the route for macromolecules such as intact proteins. Key words: SICr-labelled ethylenediaminetetraacetic acid; 1-deamino, 8-D-arginine vasopressin; dextran; inulin; mannitol; permeability; rat; small intestine
Niclas Pantzar, University of Lund, Dept. of Animal Physiology, Helgonavagen 3b, S-223 62 Lund, Sweden (fax: +46 46 104539)
The barrier function of the gastrointestinal tract is incom- plete, and luminal macromolecules may traverse its epi- thelium into the internal milieu. Intestinal barrier functions are affected during both normal physiologic conditions, such as ontogenetic maturation (l) , and during several pathologic conditions (2,3). To assess the intestinal permeability and its changes, several different marker molecules have been used, including di-mono-saccharides such as lactulosel mannitol (4), 51Cr-ethylenediaminetetraacetic acid (%r- EDTA) (5 ) , polyethylene glycols (PEG) (6), inulin (7,8), and dextrans (9). The permeability to these different markers has been assessed mainly in vivo after oral feeding, when factors both before and after the passage of the intestinal epithelium may obscure the interpretation, but has not been compared under more simplified and controlled exper- imental conditions in vitro. Moreover, little attention has been paid to the regional aspects of the intestinal permeability; although regional permeability differences along the intestine have been observed for the enzymatically stable nona-peptide 1-deamino, 8-D-arginine vasopressin (dDAVP) by using isolated intestinal segments in rabbits and rats (l@-12). Furthermore, it is likely that smaller mol- ecules use a passage route through the intestinal epi- thelium different from that for larger ones (12-14) and that lipophilicity is an important property for transmembranous passage.
The purpose of the present investigation was to charac- terize the small-intestinal permeability in the rat to several commonly used marker molecules differing in size and physicochemical nature-for example, molecular class and lipophilicity. To achieve this, intestinal segments were incu- bated in diffusion chambers (13) together with the markers 3H-mannitol (Mw 182), S*Cr-EDTA (Mw 341), [mer- captopropionic acid1, D-arginine8]-vasopressin (dDAVP) (Mw 1069), fluorescein isothiocyanate (FITCfiextran 3000 (mean Mw 3000), and 3H-inulin (Mw 5200). The bidi- rectional passage-that is, both from the mucosal to the serosal side and from the serosal to the mucosal side-of these marker molecules was compared, to obtain further knowledge about the barrier mechanisms of the intestine. To investigate the existence of any regional passage differences within the small intestine, segments from both the proximal jejunum and the distal ileum were used.
MATERIALS AND METHODS
Animals Male rats (n = 25) of the Sprague-Dawley strain (Mdle-
gaard, Skensved, Denmark), weighing 400-500 g, were kept on chopped wood bedding in polycarbonate cages under a 12-h day-night rhythm at 2Ok2"C and with a relative humidity of 50 * 10%. The rats had free access to rat chow
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.
704 N. Ponrzor er 01.
n 20 40 60 80 100 120
l ime (min)
c Bidirectional passage of dDAVP d Bidirectional passage of FITC-dextran 3000
+ proximaim-sm
distals-mm
h 1.0 55 0 LI m m a
a n 0.5
0.0 0 20 40 60 80 100 120
Time (min)
e Bidirectional passage of 3H-inulin
0 20 40 60 80 100 120
0.6
+ proximalm-sm
0.4 z 0 0)
m a m a a 0.2
0.0 0 20 40 60 80 100 120
Time (min)
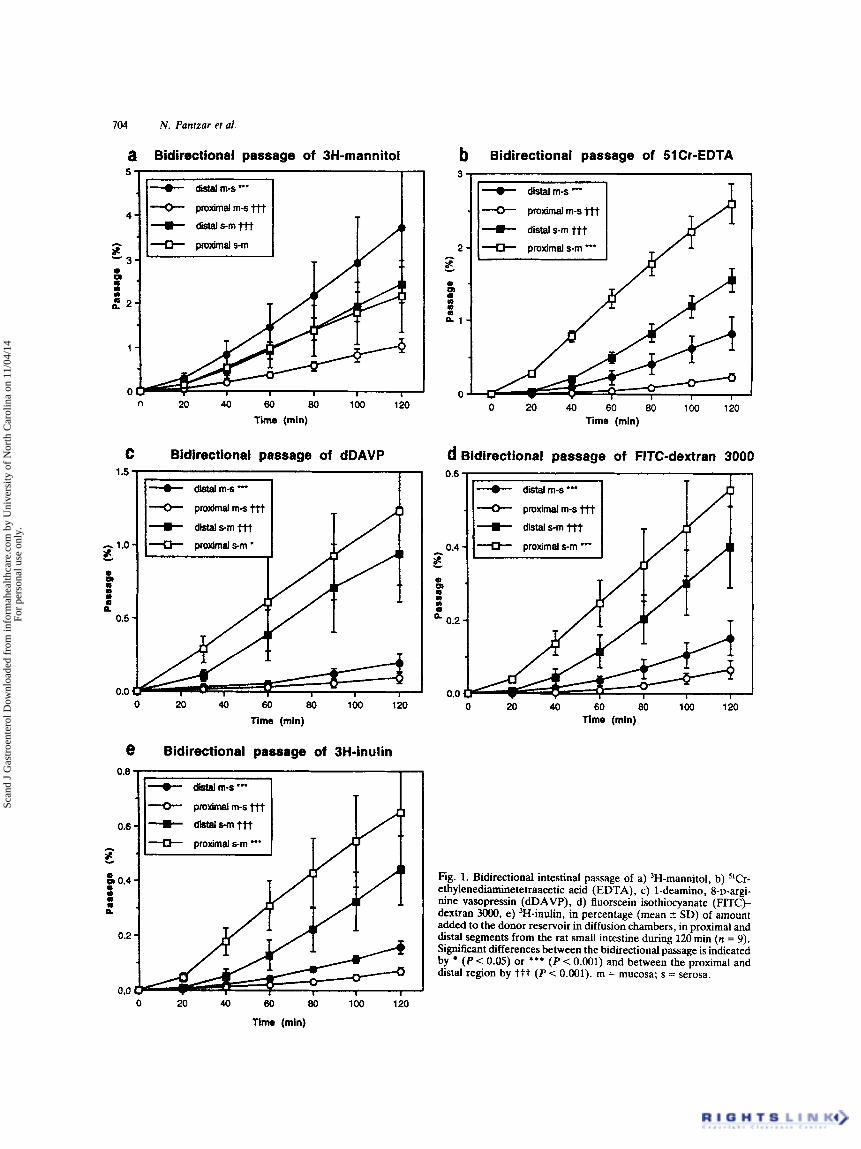
Fig. 1. Bidirectional intestinal passage of a) 'H-mannitol, b) "Cr- ethylenediaminetetraacetic acid (EDTA), c) 1-deamino, 8-D-argi- nine vasopressin (dDAVP), d) fluorscein isothiocyanate (FITC)- dextran 3o00, e) 'H-inulin, in percentage (mean 2 SD) of amount added to the donor reservoir in diffusion chambers, in proximal and distal segments from the rat small intestine during 120 min (n = 9). Significant differences between the bidirectional passage is indicated by (P < 0.05) or *** (P < 0.001) and between the proximal and distal region by ttt (P < 0.001). m = mucosa; s = serosa.
'
TIM (min)
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.

Bidirectional Small-Intestinal Permeability 705
(R3, Ewos, Sodertalje, Sweden) and tap water until the day of experiment.
Experimental procedures The animals were ether anaesthetized at lo00 h k 1 h.
An abdominal incision was performed, and two 15 cm-long segments of the small intestine were removed: one proximal segment, beginning 5 cm distal to the pylorus, and one distal segment, beginning 5 cm proximal to the caecum. These were immediately immersed in room-tempered modified Krebs-Ringer buffer, pH 7.4 (NaCI, 110.0 mmol.1-l; CaCI2, 3.0 mmol.l-'; KCI, 5.5 mmol.l-'; KH2P04, 1.4 mmol.l-'; NaHC03, 29.0 mmol.1-'; Na-pyruvate, 5.7 mrnol.l-'; Na- fumarate, 7.0 mmol.1-I; Na-glutamate, 5.7 mmol.l-'; and glucose, 13.4 mmol.1-l) oxygenated with carbogen (95% O2 and 5% C02). Both segments were then divided into three pieces, providing three proximal and three distal small- intestinal pieces from the same rat in each experiment. Each piece was cut along the mesenteric border and mounted in modified Ussing diffusion chambers (Precision Instrument Design, Los Altos, Calif., USA) (13), with an exposed intestinal area of 1.78cm2. The serosal and mucosal res- ervoirs were immediately filled with 5 ml buffer that was continuously oxygenated and circulated by gas lift at a tem- perature of 37°C. Under these conditions the tissue prep- arations were considered viable for 120min (12). At the beginning of the incubations (t = 0), within 25 minutes of induction of anaesthesia, the buffer at the mucosal or serosal reservoir was exchanged with 5 ml buffer containing the different marker molecules. Thereafter, 1-ml samples were taken from the corresponding receiver reservoirs at 20, 40, 60, 80, 100, and 120min and were replaced by the same amount of fresh buffer. In some experiments the mucosa had been scraped with the edge of a glass slide. The intestine was opened along the mesenteric border and placed on a plate, and the mucosa was scraped by gently letting the glass edge slide on the mucosa. Routine histology showed that the scraping procedure removed and disrupted the villous structure but left the crypts and submucosal and muscular layers intact.
Marker molecules The different marker molecules introduced on the mucosal
or serosal side were 3H-mannitol (Du Pont, Dreieich, Ger- many), with a specific activity of 30.0Ci/mmol and. in a concentration of 0.14 pM; 52Cr-EDTA (Du Pont), with a specific activity of 126Ci/mmol and in a concentration of 0.13 pM; 3H-inulin (Amersham, Buckinghamshire, England), with a specific activity of 1.7 Ci/mmol and in a Concentration of 4.4 pM; dDAVP (Ferring AB, Malmo, Sweden) in a concentration of 9.4 pM; and FITC-dextran 3000 (mean Mw, 3.0kDa; Pharmacia Fine Chemicals, Uppsala, Sweden) in a concentration of 140 pM.
Estimation of marker lipophilicity The different marker molecules were phase-partitioned
between n-octanol and 50 mM phosphate buffer, pH 7.4, by shaking for 10 min. After separation, the two phases were analysed for their marker content. The ratio (D) of n- octanol/buffer concentrations of each marker was calcu- lated, and log D was used a measure of the marker lipo- philicity.
Analyses The samples from the receiver reservoirs were analysed
for mannitol and inulin content by placing 1-ml samples in vials containing 10 ml scintillation fluid (Ready Safe, Beckman, Fullerton, Calif., USA) for beta-counting. The radioactivity of the vials was then counted for 600 sec in a liquid scintillation counter (1217 Rackbeta, LKB, Bromma, Sweden). %2r-EDTA was analysed by counting in 1-ml samples for 600sec in a gamma-counter (1282 Compu- gamma, LKB, Bromma, Sweden). Analysis of dDAVP was performed by radioimmunoassay, as previously described (16), within 2 weeks after the experiments. Cross-reactivity with oxytocin and vasopressin was less than 0.1%. Intra- and inter-coefficients of variation of the analysis were 8.1% and 18%, respectively. FITC-dextran 3000 concentrations were determined by spectrophotofluorometry (Cytofiuor 2300, Millipore, Bedford, Mass., USA) at an excitation wavelength of 480 nm and an emission wavelength of 520 nm, and with FITC-dextran 3000 dissolved in modified Krebs- Ringer buffer as a standard.
Calculations
marker was calculated by means of the formula:
PaPp(cm.sec-l) = (dc/dt). V.( l/co-A),
where dc/dt represents the change of the marker con- centration in the receiver reservoirs between 60 and 120 min (rnol.l-'.sec-'), V the volume in the reservoir (I), co the initial marker concentration in the donor reservoir (mol.l-'), andA the area of exposed intestine in the diffusion chambers (cm2).
To compensate for the molecular size-dependent diffusion rates in aqueous solutions the Papp was plotted against the reciprocal molecular radius in accordance with the Stokes- Einstein relation:
D = kT-(6 my-',
stating that the diffusion rate (D) is inversely proportional to the radius (r) of the molecule if the temperature (7') and the viscosity ( r ] ) are constant.
To calculate the permeability in the mucosa layer (Pcalc), after scraping the intestinal wall, the following formula was used:
(Papp intact intestinal wall)-' =
The apparent permeability coefficient (PaPp) for each
(PCarc)-' + (Papp scraped intestinal waIl)-l.
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.
706 N. Pantzar ei al.
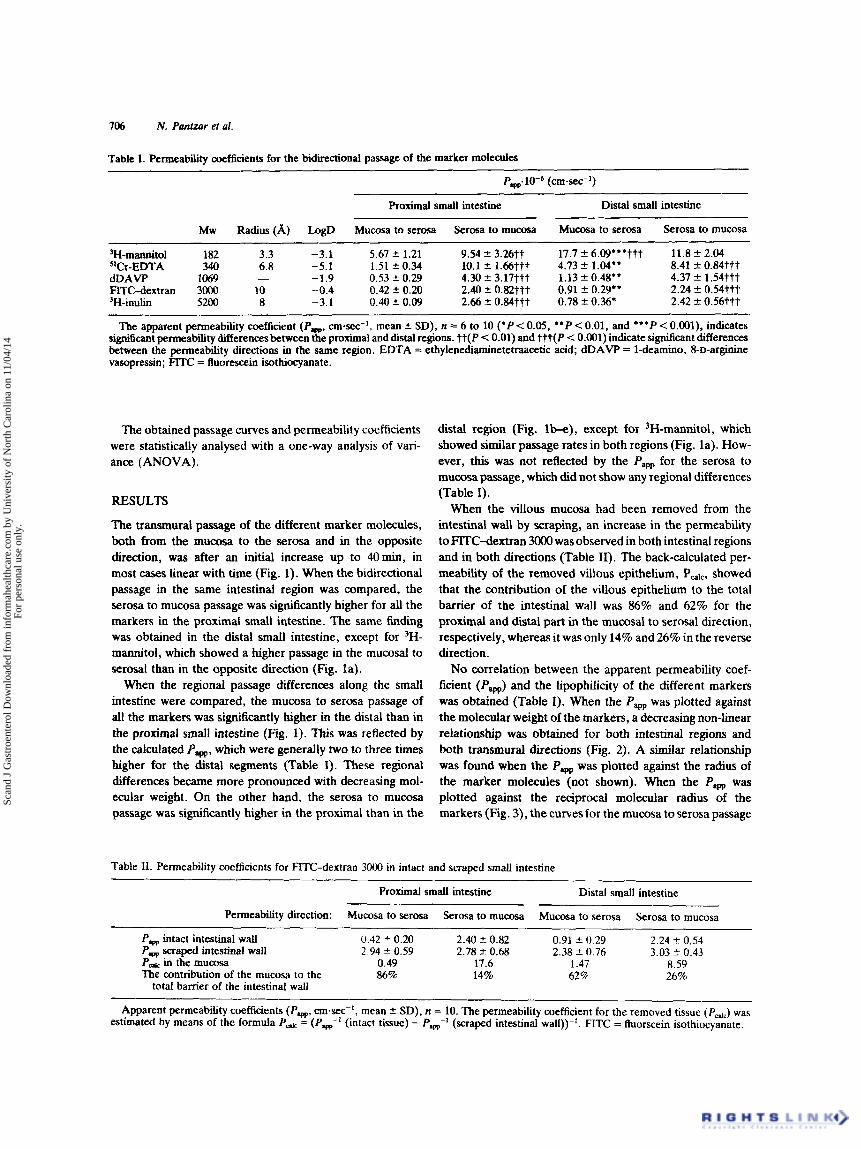
Table I. Permeability coefficients for the bidirectional passage of the marker molecules
Pw.10-6 (cm-sec-')
Proximal small intestine Distal small intestine
Mw Radius (A) LogD Mucosa to serosa Serosa to m u m a Mucosa to serosa Serosa to mucosa
B-rnannitol 182 3.3 -3.1 5.67 f 1.21 9.54 2 3.26tt 17.7 f 6.09"'ttt 11.8 2 2.04 -5.1 1.51 f 0.34 10.1 f 1.66ttt 4.73 f 1.04" 8.41 f 0.84ttt slCr-EDTA 340 6.8
dDAVP 1069 - -1.9 0.53 f 0.29 4.30 2 3.17777 1.13 f 0.48" 4.37 f 1.54ttt 2.24 f 0.54ttt FlTC-dextran 3000 10 -0.4 0.42 f 0.20 2.40 f 0.82ttt 0.91 f 0.29"
'H-inulin 5200 8 -3.1 0.40 f 0.09 2.66 2 0.84ttt 0.78 f 0.36' 2.42 2 0.56ttt
The apparent permeability coefficient (P-. cm-sec-', mean f SD), n = 6 to 10 ('P < 0.05, * * P < 0.01, and ***P < 0.001), indicates signi6cant permeablty differencesbetween the proximal and distal regions. tt(P < 0.01) and t t t ( P < 0.001) indicate significant differences between the permeability directions in the same region. EDTA = ethylenediaminetetraetic acid; dDAVP = 1-deamino, 8-D-arginine vasopressin; FITC = fluorescein isothiocyanate.
The obtained passage curves and permeability coefficients were statistically analysed with a one-way analysis of vari- ance (ANOVA).
RESULTS
The transmural passage of the different marker molecules, both from the muam to the serosa and in the opposite direction, was after an initial increase up to 40min, in most cases linear with time (Fig. 1). When the bidirectional passage in the same intestinal region was compared, the serosa to mucosa passage was significantly higher for all the markers in the proximal small intestine. The same finding was obtained in the distal small intestine, except for 3H- mannitol, which showed a higher passage in the mucosal to serosal than in the opposite direction (Fig. la).
When the regional passage differences along the small intestine were compared, the mucosa to serosa passage of all the markers was significantly higher in the distal than in the proximal small intestine (Fig. 1). This was reflected by the calculated Pw, which were generally two to three times higher for the distal segments (Table I). These regional differences became more pronounced with decreasing mol- ecular weigh!. On the other hand, the serosa to mumsa passage was significantly higher in the proximal than in the
distal region (Fig. lb-e), except for 3H-mannitol, which showed similar passage rates in both regions (Fig. la). How- ever, this was not reflected by the Paw for the serosa to muwsa passage, which did not show any regional differences (Table I).
When the villous mucosa had been removed from the intestinal wall by scraping, an increase in the permeability to FITC-dextran 3OOO was observed in both intestinal regions and in both directions (Table 11). The back-calculated per- meability of the removed villous epithelium, Palc, showed that the contribution of the villous epithelium to the total barrier of the intestinal wall was 86% and 62% for the proximal and distal part in the mucosal to serosal direction, respectively, whereas it was only 14% and 26% in the reverse direction.
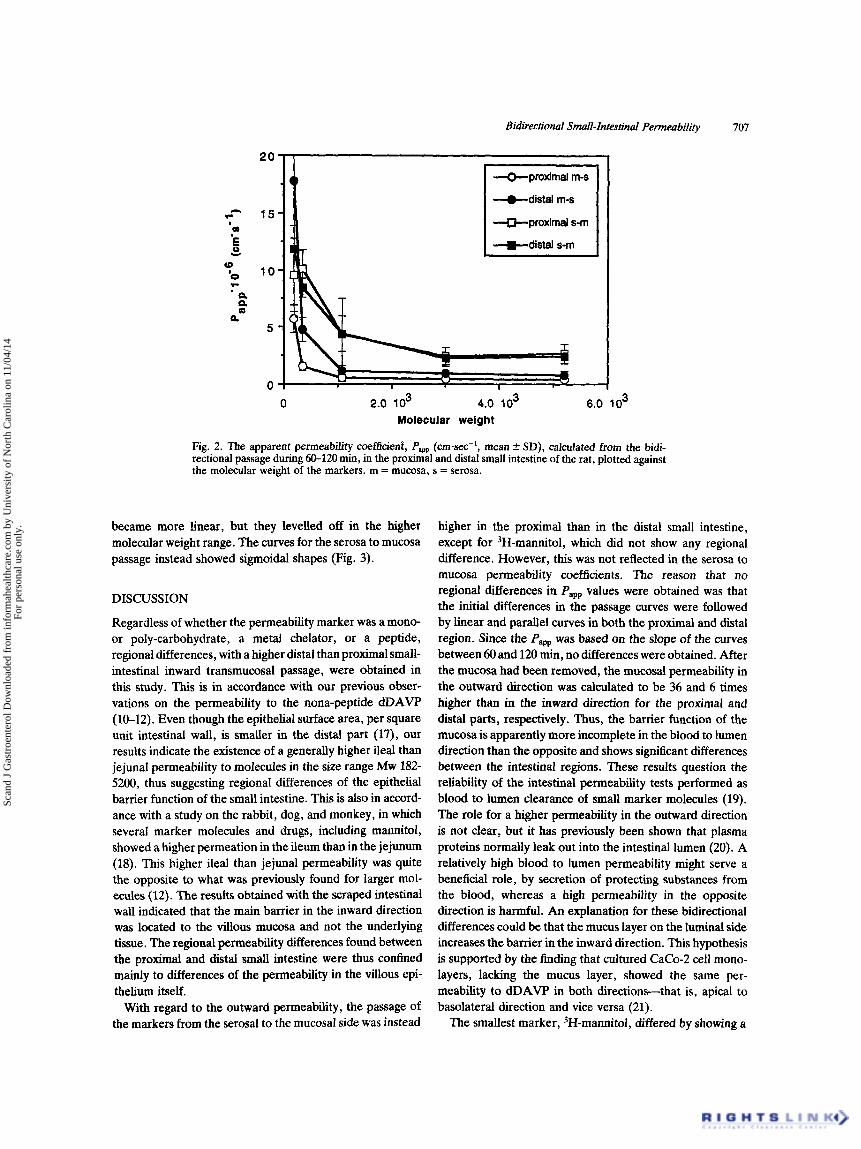
No correlation between the apparent permeability coef- ficient (Paw) and the lipophilicity of the different markers was obtained (Table I). When the Papp was plotted against the molecular weight of the markers, a decreasing non-linear relationship was obtained for both intestinal regions and both transmural directions (Fig. 2). A similar relationship was found when the Pw was plotted against the radius of the marker molecules (not shown). When the Pam was plotted against the reciprocal molecular radius of the markers (Fig. 3), the curvesfor the mumsa to serosa passage
Table 11. Permeability coefficients for FITC-dextran 3000 in intact and scraped small intestine
Proximal small intestine Distal small intestine
Permeability direction: Mucosa to serosa Serosa to m u m a Mucosa to serosa Serosa to mucosa
Pw intact intestinal wall 0.42 4- 0.20 2.40 5 0.82 0.91 * 0.29 2.24 2 0.54 Pw scraped intestinal wall 2.94 * 0.59 2.78 % 0.68 2.38 2 0.76 3.03 ? 0.43
in the m u a m 0.49 17.6 1.47 8.59 The contribution of the mucosa lo the 86% 14% 62% 26%
total barrier of the intestinal wall
Apparent permeability coefficients (Pw, cmsec-', mean f SD), n = 10. The permeability coefficient for the removed tissue (Pcdc) was estimated by means of the formula Pd = (P,-' (intact tissue) - P&' (scraped intestinal wall))-'. FITC = fluorscein isothiocyanate.
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.
Bidirectional Small-Intestinal Permeability 707
0 2.0 lo3 4.0 lo3 6.0 lo3 Molecular weight
Fig. 2. The apparent permeability ooefficienf, Papp (cmsec-', mean f SD), calculated from the bidi- rectional passage during 60-120 min, in the proximal and distal small intestine of the rat, plotted against the molecular weight of the markers. m = mucosa, s = serosa.
became more linear, but they levelled off in the higher molecular weight range. The curves for the serosa to mucosa passage instead showed sigmoidal shapes (Fig. 3).
DISCUSSION
Regardless of whether the permeability marker was a mono- or poly-carbohydrate, a metal chelator, or a peptide, regional differences, with a higher distal than proximal small- intestinal inward transmucosal passage, were obtained in this study. This is in accordance with our previous obser- vations on the permeability to the nona-peptide dDAVP (10-12). Even though the epithelial surface area, per square unit intestinal wall, is smaller in the distal part (17), OUT
results indicate the existence of a generally higher ileal than jejunal permeability to molecules in the sue range Mw 182- 5200, thus suggesting regional differences of the epithelial barrier function of the small intestine. This is also in accord- ance with a study on the rabbit, dog, and monkey, in which several marker molecules and drugs, including mannitol, showed a higher permeation in the ileum than in the jejunum (18). This higher ileal than jejunal permeability was quite the opposite to what was previously found for larger mol- ecules (12). The results obtained with the scraped intestinal wall indicated that the main barrier in the inward direction was located to the villous mucosa and not the underlying tissue. The regional permeability differences found between the proximal and distal small intestine were thus confined mainly to differences of the permeability in the villous epi- thelium itself.
With regard to the outward permeability, the passage of the markers from the serosal to the mucosal side was instead
higher in the proximal than in the distal small intestine, except for 3H-mannitol, which did not show any regional difference. However, this was not reflected in the serosa to mucosa permeability coefficients. The reason that no regional differences in Papp values were obtained was that the initial differences in the passage curves were followed by linear and parallel curves in both the proximal and distal region. Since the Paw was based on the slope of the curves between 60 and 120 min, no differences were obtained. After the mucosa had been removed, the mucosal permeability in the outward direction was calculated to be 36 and 6 times higher than in the inward direction for the proximal and distal parts, respectively. Thus, the barrier function of the mucosa is apparently more incomplete in the blood to lumen direction than the opposite and shows significant differences between the intestinal regions. These results question the reliability of the intestinal permeability tests performed as blood to lumen clearance of small marker molecules (19). The role for a higher permeability in the outward direction is not clear, but it has previously been shown that plasma proteins normally leak out into the intestinal lumen (20). A relatively high blood to lumen permeability might serve a beneficial role, by secretion of protecting substances from the blood, whereas a high permeability in the opposite direction is harmful. An explanation for these bidirectional differences could be that the mucus layer on the luminal side increases the barrier in the inward direction. This hypothesis is supported by the finding that cultured CaCo-2 cell mono- layers, lacking the mucus layer, showed the same per- meability to dDAVP in both directions-that is, apical to basolateral direction and vice versa (21).
The smallest marker, 3H-ma~itol , differed by showing a
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.

708 N. Pantzar et al.
20
15
10
5
0
0.05 0.1 0.15 0.2 0.25 0.3 0.35
reciprocal molecular radius (Am')
Fig. 3. The apparent permeability coefficient, Pppp (cm-sec-', mean ? SD), calcuiated from the bidi- rectional passage during 60-120 min. in the proximal and distal small intestine of the rat, plotted against the reciprocal molecular radius of the markers. m = mucosa; s = serosa.
higher permeability in the mucosa to serosa direction than in the reverse direction in the distal ileum. This could be due to the larger surface area on the luminal side of the enterocytes than on the basolateral side, This, of course, is on the condition that the main route for mannitol, when passing the epithelium, is through aqueous pores in the cell membrane of the enterocytes (see below). However, the possible degradation of 3H-mannitol to tritium-labelled water (22,23), which could be readily absorbed and thus give an incorrect view of the 3H-mannitol permeability, should not be excluded. If the discrepancy observed for mannitol is due to regional differences of degradation, the use of 3H-mannitol as a permeability marker could be ques- tioned, especially in vivo, where the site of absorption usually is unknown. It is therefore preferable to use 14C-mannitol, producing radiolabelled C 0 2 instead of radiolabelled water when degraded. In conclusion, our results indicate regional differences with an increasing absorptive and a decreasing exudative capacity in the proximal to distal direction of the small intestine.
Since there was no correlation between the Pa, and the hydrophilicity of the different markers, a passage pathway involving penetration of the iipid membranes of the enterocytes is unlikely for these molecules. When the Papp for the mucosal to serosal passage instead was plotted against the molecular weight or the radius, an apparently non- linear relationship was obtained, with a shift of the slope at approximately the size of 51Cr-EDTA and with the curves
levelling off with increasing molecular size, indicating the presence of two different routes in the present size range. Thus, the linear relationship between the in vivo absorption and the molecular radius for some permeability probes, found by Hollander et al. (24), is probably valid only in a narrow size range. We have previously shown; by plotting the Papp against (3uMw)-1 (an approximation of the recipro- cal radius) for different-sized proteins and FITC-dextrans, a shift of the curves in the range between 10 and 30 kDa (12). This shift indicated a changeover between two different pathways, one of low permeability for molecules >10 kDa and another of relatively higher permeability for molecules <lo kDa. These results in conjunction with the present, indicate the possible presence of three different size-depen- dent inward routes through the epithelium: one for mol- ecules with a cross-sectional diameter smaller than about 10 A (for example, mannitol), probably through aqueous pores in the apical membrane of the enterocytes (25), a second from l 0 A to about 30& probably a paracellular route through the tight junctions (26), and thirdly, apathway of low permeability, the main route for molecules larger than about 30 A (such as proteins), perhaps a transcellular one involving transcytosis (27,28). The marker FITC-dex- tran 3000 showed absorption properties quite different from those of the larger macromolecules in the 30- to 70-kDa range in a previous study (12). Hence, since FITC-dextran 3OOO fitted nicely into the group of low molecular weight markers used in this study, it is likely that the common use
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.

Bidirectional Small-Intestinal Permeability 709
of low molecular weight permeability probes is of minor relevance for estimating the intestinal permeability to larger macromolecules, including antigens.
ACKNOWLEDGEMENTS
This study was supported by grants from The Swedish Nat- ural Science Research Council, The Royal Swedish Academy of Sciences, and the Royal Physiographical Society of Sweden. Stefan Lundin was supported by the Medical Faculty, University of Lund.
REFERENCES 1. Baintner K. Intestinal absorbtion of macromolecules and
immune transmission from mother to young. Boca Raton (FL): CRC Press, 1986.
2. Gardner MLG. Gastrointestinal absorption of intact proteins. Ann Rev Nutr 1988;8:329-50.
3. Travis S, Menzies I. Intestinal permeability: functional assess- ment and significance. Clin Sci 1992;82:471-88.
4. Menzies IS, Laker MF, Pounder RE, et at. Abnormal intestinal permeability to sugars in villous atrophy. Lancet 1979;2:1107- 9.
5. Bjarnason I, Peters TJ, Veal1 N. A persistent defect of intestinal permeability in coeliac disease as demonstrated by a Wr-EDTA absorption test. Lancet 1983;1:323-5.
6. Chadwick VS, Phillips SF, Hofmann AF. Measurements of intestinal permeability using low molecular weight polyethylene glycols (PEG 400). 11. Application to normal and abnormal permeability states in man and animals. Gastroenterology
7. Pappenheimer JR, Reiss KZ. Contribution of solvent drag through intracellular junctions to absorption of nutrients by the small intestine of the rat. J Membr Biol 1987;100:123-36.
8. Ma TY, Hollander D, Erickson RA, Truong H, Krugliak P. Is the small intestinal epithelium truly ‘tight’ to inulin permeation? Am J Physiol 1991;260:0669-76.
9. Tagesson C, Sjodahl R, Thorkn B. Passage of molecules through the wall of the gastrointestinal tract. I. A simple experimental model. Scand J Gastroenterol 1978;13:519-24.
10. Lundin S, Pantzar N, Hedin L, Westrom BR. Intestinal absorp- tion enhancement by sodium taurodihydrofusidate of a peptide hormone analogue (dDAVP) and a macromolecule (BSA) in vitro and in vivo. Int J Pharmaceut 1990;59:263-9.
11. Lundin S, Pantzar N, Broeders A, Ohlin M, Westrom BR. Differences in transport rate of oxytocin and vasopressin ana- logues across proximal and distal isolated segments of the small intestine of the rat. Pharm Res 1991;8:1274-80.
1977;73:247-51.
12. Pantzar N, Lundin S, Luts A, Westrom BR. Regional small- intestinal permeability in vitro to different-sized dextrans and proteins in the rat. Scand J Gastroenterol 1993;28:205-11.
13. Weaver LT, Coombs RRA. Does ‘sugar’ permeability reflect macromolecular absorption? A comparison of the gastro-intes- tinal uptake of lactulose and beta-lactoglobulin in the neonatal guinea pig. Int Arch Allergy Appl Immun 1988;85:133-5.
14. Turner Mw, Boulton P, Shields JG, Strobe1 S, Gibson S, Miller HRP, et al. Intestinal hypersensitivity reactions in the rat. I. Uptake of intact protein, permeability to sugars and their correlation with mucosal mast-cell activation. Immunology 1987; 63: 119-24.
15. Grass GM, Sweetana SA. In vitro measurements oi gastro- intestinal tissue permeability using a new diffusion cell, Pharma- ceut Res 1988;6:372-6.
16. Lundin S, Melin P, Vilhardt H. Plasma concentrations of 1- deamino-8-D-arginine vasopressin after intragastric adminis- tration in the rat. Acta Endocrinol 1985;108:179-83.
17. Fisher RB, Parsons DS. The gradient of mucosal area in the small intestine of the rat. J Anat 1950;84:272-82.
18. Jezyk N, Rubas W, Grass GM. Permeability characteristics of various intestinal regions of rabbit, dog and monkey. Pharm Res 1992;9:1580-6.
19. Crissinger KD, Kvietys PR, Granger DN. Pathophysiology of gastrointestinal mucosal permeability. J Intern Med 1990;228: 145-54.
20. Andersen V, Hegnhc4j JH. Identification and quantification of serum proteins secreted into the normal human jejunum. Scand J Clin Lab Invest 1990;50:857-64.
21. Lundin S, Artursson P. Absorption of a vasopressin analogue, 1-deamino-8-D-arginine-vasopressin (dDAVP), in a human intestinal epithelial cell line, CaCO-2. Int J Pharmaceut 1990; 64:181-6.
22. Nasrallah SM, Iber FL. Mannitol absorption and metabolism in man. Am J Med Sci 1969;258:80-8.
23. Pappenheimer JR. Paracellular intestinal absorption of glucose creatinine and mannitol in normal animals: relation to body size. Am J Physiol 1990;259:6290-9.
24. Hollander D, Ricketts D, Boyd CAR. Importance of ‘probe’ molecular geometry in determining intestinal permeability. Can J Gastroenterol 1988;2:35A-8A.
25. Hamilton I, Rothwell J, Archer D, Axon ATR. Permeability of the rat small intestine to carbohydrate probe molecules. Clin Sci 1987;73:189-96.
26. Madara JL, Barenberg D, Carlson S. Effects of cytochalasin D on occluding junctions of intestinal absorptive cells: further evidence that the cytoskeleton may influence paracellular per- meability and junctional charge selectivity. J Cell Biol 1986; 102:2125-36.
27. Walker WA. Antigen handling by the small intestine. Clin Gastroenterol 1986;15:1-20.
28. Heyman M, Ducroc R, Desjew J-F, Morgan JL. Horseradish peroxidase transport across adult rabbit jejunum in vitro. Am J Physiol 1982;242:G558-64.
Received 27 July 1993 Accepted 8 December 1993
Scan
d J
Gas
troe
nter
ol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Nor
th C
arol
ina
on 1
1/04
/14
For
pers
onal
use
onl
y.