becoming sedentary| the seasonality of food resource
TRANSCRIPT

103
Documenta Praehistorica XLIII (2016)
Becoming sedentary|The seasonality of food resource exploitation in the
Mesolithic-Neolithic Danube Gorges
Vesna Dimitrijevic 1, Ivana ?ivaljevic 2 and Sofija Stefanovic 2
1 Laboratory for Bioarchaeology, Department of Archaeology, Faculty of Philosophy, University of Belgrade, [email protected]
2 BioSense Institute, University of Novi Sad, [email protected]
Introduction
With their long temporal sequence, spanning be-tween c. 9500 to 5500 cal BC (corresponding to theregional Mesolithic, Mesolithic-Neolithic Transforma-tion phase and the Early/Middle Neolithic) (Bori≤,Dimitrijevi≤ 2009; Bori≤ 2011), open-air sites in theDanube Gorges (North-Central Balkans) provide greatopportunities for studying and understanding vari-ous aspects of Mesolithic-Neolithic transition. In par-ticular, intensified building activities in the Late Me-solithic and Transformational Mesolithic-Neolithicphases (cf. Bori≤, Dimitrijevi≤ 2009; Bori≤ 2011)have been interpreted as archaeological indicators ofincreased sedentism resulting from developing socialcomplexity of local hunter-gatherer-fisher communi-ties (cf. Voytek, Tringham 1989; Chapman 1993;Radovanovi≤ 1996; Radovanovi≤, Voytek 1997) andfrom merging local traditions with new economic
and social practices accompanying ‘Neolithic’ life-ways (cf. Bori≤ 2007; 2011; Jovanovi≤ 2008). Ob-viously, there is no universal mode of hunter-fisher-gatherer existence, as opposed to a presumed seden-tary lifestyle of food-producing communities, norcan a clear-cut change from mobility to sedentism beassumed. Moreover, degrees of sedentism are oftendifficult to assess and divide into neat categories dueto the complexity of human lifestyles and residen-tial practices (cf. Whittle 2001). One of the startingpoints, however, most commonly involves determin-ing whether the sites in question were occupied sea-sonally or year-round. This feature can be addressedby looking at subsistence patterns, i.e. the seasonsof exploitation of most important animal species, ashas been demonstrated by a number of studies,which range from simple observations of the pres-
ABSTRACT – In this paper, we investigate whether the Mesolithic-Neolithic sites in the Danube Gorgeswere occupied seasonally or all year round by looking at animal skeletal remains. The hunting sea-sons of most important game animals have been determined on the basis of antler and teethgrowth, supplemented by looking into the presence of migratory fish and birds. The patterns of foodresource exploitation seem to indicate year-round occupation of the settlements, and suggest that asignificant degree of sedentism existed in the Danube Gorges prior to, and independently of, theadoption of animal and plant husbandry.
IZVLE∞EK – V ≠lanku na podlagi analize ∫ivalskih ostankov raziskujemo, ali so bila mezolitsko-neo-litska najdi∏≠a na obmo≠ju soteske D∫erdap poseljena sezonsko ali ≠ez celo leto. Na podlagi rasti ro-govja in zob smo dolo≠ili lovno sezono za najpomembnej∏o divjad, podatke pa smo dopolnili z anali-zo selitve rib in ptic. Vzorci rabe raznih virov prehrane ka∫ejo na celoletno poselitev na naselbinah;sklepamo tudi, da se je na obmo≠ju D∫erdapa delno sedentaren na≠in ∫ivljenja pojavil lo≠eno od inpred pojavom prvih doma≠ih ∫ivali in kultiviranih rastlin.
KEY WORDS – seasonality; archaeozoology; sedentism; Danube Gorges; Mesolithic-Neolithic
DOI> 10.4312\dp.43.4

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
104
ence/absence of seasonally available fauna, to moresophisticated analytical procedures, including thedetermination of the individual ages and the seasonof death of exploited animals, skeletochronology, orgrowth-increment studies (cf. Pike-Tay et al. 2004),or stable isotope balances in teeth and bones (Blaise,Balasse 2011).
In this study, we present estimated seasons of cap-ture on the basis of the results of ageing of faunalskeletal elements (teeth and antler) of economical-ly important wild game species, supplemented byquantifications of fish and bird remains (some ofthem available seasonally and others more or lessall year round) from sites at Vlasac, Lepenski Vir,and Padina. By no means exhaustive, this study aimsto offer additional data relevant to understandingsubsistence and settlement patterns, and to shedmore light on the relationship between hunting, fish-ing and increased sedentism in the Danube Gorges.
Archaeological background
The Padina, Lepenski Vir, and Vlasac sites are situ-ated in the Upper Gorge (Gospo∂in Vir, or Lady’sWhirlpool) of the Danube Gorges (Fig. 1), on river-ine terraces sloping towards the Danube. The initialrescue excavations of the sites were undertaken inthe 1960s and 1970s (Srejovi≤ 1969; 1972; Srejo-vi≤, Letica 1978; Radovanovi≤ 1996; Jovanovi≤2008), prior to the raising of the water level due todam construction, and new revisory excavations ofthe preserved portion of the Vlasac terrace were car-ried out in 2006–2009 (Bori≤ 2006; Bori≤ et al.2008; 2014). The earliest traces of human presencein these locations date to the period of c. 9500–7400 cal BC (the Early Mesolithic) (Bori≤, Dimitrije-vi≤ 2009; Bori≤ 2011), and are manifested by occa-sional burials (some in a seated position), severalstone constructions (Bori≤, Miracle 2004; Jovanovi≤
2008), as well as caches of animal bones originatingpredominantly from wild game and fish (Bori≤, Di-mitrijevi≤ 2006), but also from canids displayingmixed wolf and dog features, suggesting that the ini-tial stages of dog domestication took place duringthis time (Dimitrijevi≤, Vukovi≤ 2015).
The few and fragmentary traces of Early Mesolithicoccupation are mostly due to their disturbance anddevastation by later (Late Mesolithic and Transfor-mation phase) building and occupational activities(Bori≤ 2011). The Late Mesolithic phase (c. 7400–6300/6200 cal BC) is particularly well documentedat Vlasac (Bori≤ 2011), where numerous burials, se-veral dug-out dwelling structures, stone-lined rectan-gular hearths and a large number of chipped stone,bone and antler artefacts have been discovered (Sre-jovi≤, Letica 1978; Nemeskéri 1978; Kozłowski,Kozłowski 1982; Roksandi≤ 1999; 2000; Bori≤ etal. 2008; 2014), as well as an enormous assemblageof animal bones (Bökönyi 1978). This phase alsoproduced evidence of large-scale exchange networks,witnessed by occasional finds of good-quality flint ofnon-local origin, and the occurrence of marine gas-tropods Cyclope neritea and Columbella rustica insome of the burials (Cristiani, Bori≤ 2012; Bori≤ etal. 2014; Srejovi≤, Letica 1978).
The Transformation/Early Neolithic phase (c. 6300/6200–5900 cal BC) (Bori≤, Dimitrijevi≤ 2007; Bori≤2011), coinciding with the emergence of the firstfarming communities in the Balkans, saw the estab-lishment of the first settlements with elaborate tra-pezoidal-base dwellings, distinctive sandstone sculpt-ed boulders and the introduction of pottery at Le-penski Vir and Padina (Srejovi≤ 1969; 1972; Rado-vanovi≤ 1996; Gara∏anin, Radovanovi≤ 2001; Jo-vanovi≤ 2008), but with unchanged subsistence stra-tegies dependent on hunting and fishing (Bori≤, Di-mitrijevi≤ 2006). The first domesticates (apart from
Fig. 1. The Danube Gor-ges area with relevantsites mentioned in thetext. The position of theDanube Gorges withina wider area – the Bal-kan peninsula shownin the lower left corner.Images from GoogleEarth.

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
105
dogs) make an appearance only after c. 6000/5900cal BC (Bori≤, Dimitrijevi≤ 2007), after most of thedwellings had been abandoned and new elements ofmaterial culture (domed ovens, ground stone tools,bone spoons), along with certain individuals of non-local origin appear in the area (Bori≤ 2011; Bori≤,Price 2013). Large-scale animal husbandry, however,was never fully introduced into the Upper Gorge;the sites continued to be in use until c. 5500 cal BC,but probably for specialised activities, most notablyfishing (Bonsall 2008; Bori≤ 2011).
The faunal assemblages
The assessment of the nature of animal exploitationand quantification of faunal remains from the DanubeGorges are hindered by several factors, namely theinadequate recovery techniques and selective collec-tion imposed by time constraints during the courseof the excavations, and the fragmentary state of fau-nal assemblages from some of the sites (Bori≤ 2001;2002; Bori≤, Dimitrijevi≤ 2006; Dimitrijevi≤ 2000;2008). The faunal assemblages from the old excava-tions of Lepenski Vir (1965–1970), Vlasac (1970–1971), and Padina (1968–1970) were collected en-tirely by hand, using collection grids consisting of4x4m quadrants. Furthermore, the enormous assem-blage from the original excavations of Vlasac, com-prising nearly 30 000 identified specimens was dis-carded after the initial analysis by Sándor Bökönyi(1978), with only a small sample of bird (Tab. 5)and fish remains preserved (Bori≤ 2002.App. 5; Di-mitrijevi≤ et al. submitted). Part of the faunal assem-blage from Lepenski Vir, mainly from first excavationseasons (up to 1969, cf. Bori≤, Dimitrijevi≤ 2006)was also discarded after the preliminary analysis byBökönyi (1969; 1970; 1972). The preserved part ofthe assemblage originated from later campaigns(1968–1970), and consisted of animal bones pri-marily related to the floors of trapezoidal-base dwel-lings, and areas between and underneath them (Di-mitrijevi≤ 2000; 2008). Only the faunal assemblagefrom the new (2006–2009) excavations of Vlasac, aswell as the one collected in 1992–1996 at the down-stream site of Schela Cladovei (Fig. 1) (Bartosiewiczet al. 1995; 2001; 2008), were collected by watersieving and flotation, in addition to hand collection(Bori≤ et al. 2014; Dimitrijevi≤ et al. submitted).
The environment of Padina, Lepenski Vir and Vla-sac, which were situated on riverine terraces in anarrow belt between the river and the forest, pro-vided both good hunting and fishing conditions. Thedense forests surrounding the sites were the natur-
al habitat of various wild game species which werehunted for meat, as well fur-bearing animals whichwere probably hunted for fur or as vermin. The vi-cinity of the river and its large whirlpools enabledfishing for both freshwater species (available mostof the year) and seasonally available anadromousspecies, as well as the exploitation of waterfowl.
In terms of wild game at all three sites, the most nu-merous remains are of red deer (Cervus elaphus)which was most commonly hunted, followed by wildboar (Sus scrofa), roe deer (Capreolus capreolus),and aurochs (Bos primigenius) (cf. Bökönyi 1969;1970; 1972; 1978; Clason 1980; Dimitrijevi≤ 2008).In terms of fishes, the remains of various cyprinids(Cyprinidae), Wels catfish (Silurus glanis), migrato-ry sturgeons (Acipenseridae), and huchen (Huchohucho) have been discovered at all three sites, butin varying quantities (Ωivaljevi≤ in prep.). As expect-ed, the hand-collected, water-sieved and flotatedsample from new excavations of Vlasac containedmany more remains of smaller cyprinids and exhib-ited the greatest species diversity in comparison tohand-collected assemblages from Lepenski Vir andPadina (Tab. 4). However, in spite of the same col-lection methods employed at the latter two sites,their faunal assemblages also exhibited significantdifferences, namely in the absolute predominance ofcatfish observed at Padina. The Lepenski Vir assem-blage contained a significant number of cyprinid, cat-fish and huchen remains, but also a much greaternumber of sturgeon remains in comparison to Pa-dina and Vlasac (Tab. 4).
The role of hunting vs. fishing in subsistence strate-gies is difficult to assess on the basis of a straight-forward comparison of quantified mammal and fishremains, given the differences in their anatomy andsize, and biases affecting their survival and recovery(cf. Wheeler, Jones 1989; Takács, Bartosiewicz1998; Bartosiewicz, Bonsall 2004; Bori≤ 2001). Theanalyses of carbon (δ13C) and nitrogen (δ15N) iso-tope ratios in human bone collagen have suggestedthat the Mesolithic inhabitants of the Danube Gorgesconsumed considerable amounts of fish, with a gra-dual broadening of the dietary spectrum to includemore terrestrial resources at the onset of the Neoli-thic (Bonsall et al. 1997; 2004; Grupe et al. 2003;Bori≤ et al. 2004), whereas the analysis of the sul-phur (δ34S) isotope ratio has suggested that therehave been significant inter- and intra-site variabili-ties in dietary practices (Nehlich et al. 2010). Onthe basis of faunal data, it can be hypothesised thatboth hunting and fishing made significant contribu-

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
106
tions to the diet; the former mainly oriented to-wards red deer and the latter exhibiting more intra-site (and possibly diachronic) variety.
Apart from a general emphasis on the importance ofannual cycles in shaping human lives in the DanubeGorges (cf. Srejovi≤ 1969; 1972), the subject of sea-sonality was first addressed by Anneke T. Clason(1980) in her study of Star≠evo and Padina animalremains. In the description of mammal remains, shespecified some specimens as seasonal markers, madesome remarks on the seasonal availability of birdand fish, and hypothesised that “Padina was perma-nently inhabited during all seasons of the year”,and furthermore, that this was probably also truefor Vlasac and Lepenski Vir. A discussion on the in-fluences of seasonal cycles on life in the MesolithicDanube Gorges is given by Bori≤ in his thesis (Bo-ri≤ 2002). Here, and in several later references, sea-sonal estimates determined on the basis of particu-lar specimens of mammal remains were also noted(Dimitrijevi≤ 2000; 2008; Bori≤, Dimitrijevi≤ 2006;2009).
Materials and methods
The seasonal schedule of food resource acquisitionin the Danube Gorges has been determined on thebasis of skeletal elements which are sensitive to thequestion of seasonality. This included determiningthe kill season of red deer and roe deer on the basisof antler growth (cf. Schmid 1972), and ageing ofred deer, roe deer, fox and wild boar on the basisof tooth eruption and wear (cf. Habermehl 1985;Brown, Chapman 1991; Carter 2006; Carter, Mag-nell 2007) (Tabs. 1–3). Although radiographs werenot made, the fragmentary state of some of the man-dibles and loose teeth enabled ageing on the basisof root formation stages (after Brown, Chapman1991; Carter 2006; Carter, Magnell 2007). Othermammal species were not taken into account, eitherbecause they were not represented by juvenile ani-mals, are not seasonal breeders, or they give birthseveral times a year. The breeding and reproductionpatterns of dogs in particular could have changedsignificantly as a result of domestication, and couldhave extended throughout several seasons.
In this respect, the most reliable season indicatorsare milk and erupting permanent teeth of juvenileanimals, given that they follow a fairly standardisedschedule, whereas tooth wear does not occur at aneven pace (Bartosiewicz 1989; Pike-Tay et al. 2004).Less reliable season indicators are shed antlers; even
if they are dropped during limited periods of theyear (February-March in the case of red deer and Oc-tober-November for roe deer, cf. Schmid 1972), theycould have been collected at any time and/or curat-ed, especially since they were extremely valued asraw material and often structurally deposited.
We assumed that red deer give birth at the begin-ning of June (cf. Carter 2006); similarly, the birth-ing season of recent red deer populations in the re-gion is described as taking place “at the end of May– beginning of June” (Grupa autora 1991). Roedeer give birth in May (Grupa autora 1991; Haber-mehl 1985; Carter 2006). The farrowing season ofwild swine is seasonally less restricted, occuring inMarch-April in recent wild pig populations in theBalkans (Grupa autora 1991), whereas in Swedenand Germany (Carter, Magnell 2007), as well as inTurkey (Bull, Payne 1982) it occurs between Marchand May. Wild sows may breed twice a year, thussubstantially widening the farrowing season; how-ever, 65–90% of births occur between March andMay, mostly March and April in Sweden and Ger-many (Carter, Magnell 2007 and references there-in). Similarly, in the Balkans, 85% of births takeplace in March and April (Grupa autora 1991). Foxcubs are born in March (Habermehl 1985).
The observations on the hunting seasons of mostimportant game animals were supplemented bylooking into the seasonal availability of migratoryfish and birds and quantifying their remains (Tabs.4–5). As noted by László Bartosiewicz and CliveBonsall (2004) and Bartosiewicz et alii (2008), thepresence/absence criteria is highly probabilistic,given that the presence of a certain species mightsignal occupation during a particular season, but itsabsence does not imply the lack of occupation atother times. For example, prior to the Iron Gates’dams construction, anadromous sturgeons – beluga(Huso huso), Russian sturgeon (Acipenser guelden-staedtii), ship sturgeon (Acipenser nudiventris), andstellate sturgeon (Acipenser stellatus), undertooktheir bi-annual spawning migrations from the BlackSea at particular times of the year (in early springand autumn), but with inter-specific and individualdifferences in the start and peak dates depending onwater temperature. Also, the populations migratingin autumn were known to overwinter in the river(Risti≤ 1977). It should be noted, however, that themain sturgeon fishing seasons in the Danube Gorgesin more recent times corresponded with their migra-tion patterns, with most catches occurring in spring,followed by autumn (cf. Petrovi≤ 1998. [1941]; for

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
107
different data from Hungary, see Bartosiewicz etal. 2008).
The investigation of the presence of migratory birdsas seasonal indicators is also based on the assump-tion that their behaviour in the past roughly mir-rored their present migration patterns. In addition,it should be noted that bird remains at sites couldhave been accumulated not only by humans anddogs, but wild predators as well, having in mind thatno bird bones bore butchering marks. This hypoth-esis does not seem likely for certain species – parti-cularly birds of prey, which nested in the high cliffsof the Gorges – but cannot be completely ruled out.
Therefore, the data we present here includes the sug-gested seasons of kill of the most important gameanimals, which could vary to some extent due toslight or greater variations in the birthing seasons ofthe species and their ontogeny, as well as the mostprobable seasons of exploitation of migratory spe-cies, assuming their past migration patterns broadlycorrespond to those observable in more recent times.
Results
VlasacAs opposed to the 640m2 area excavated in 1970–1971 (Srejovi≤, Letica 1978), the preserved portionof the Vlasac terrace encompassed by the 2006–2009 excavations (326m2) covered the southern-most peripheral area of the original settlement (Bo-ri≤ et al. 2014), with much less evidence of domes-tic activities or food preparation and consumption(Dimitrijevi≤ et al. submitted). This feature is cer-tainly manifested in the quantity and preservationof animal remains from the two areas of the site, inparticular concerning mammal remains, which areleast likely to suffer from bias in recovery techni-ques. The total number of mammal bone specimensidentified to the species/genus/family level from the1970–1971 excavations equaled 11188 (Bökönyi1978), as opposed to 801 specimen identified to thespecies/genus/family level and 10955 unidentifiedmammal remains from the 2006–2009 excavations(Dimitrijevi≤ et al. submitted). In terms of the mostimportant game animals, the NISP (number of iden-tified specimens) of red deer, wild boar and roe deerin the 1970–1971 assemblage equaled 6732, 1185and 510, respectively (Bökönyi 1978), as opposedto their NISP in the 2006–2009 assemblage (209, 86and 47, respectively) (Dimitrijevi≤ et al. submitted).However, due to more sophisticated collection tech-niques, the new excavations of Vlasac enabled the
recovery of even the most fragile skeletal elements,including the deciduous teeth which were used inthis study (Tab. 1), as well as a plentitude of fish re-mains (Tab. 4).
Most of the mammal remains used in this study origi-nated from the 2006–2009 excavations of Vlasac,and included a fragmented red deer skull, a fragmentof a roe deer skull with an antler, as well as decidu-ous and permanent teeth of red deer, roe deer, wildboar and fox. Given that the largest part of the fau-nal assemblage from 1970–1971 was discarded, inthis study we were able to include only 9 unshedand 31 shed antlers of red deer, and 3 unshed ant-lers of roe deer, which were originally packed withbone and antler artefacts and thus saved (Tab. 1).
The fragmented red deer skull VL 29/1 (Fig. 2) ori-ginated from a young male, and on the basis of par-tially preserved antlers still on pedicles, it was deter-mined that it was captured between September andFebruary. The skull had been used (possibly, for asignificant amount of time), most likely in a series ofritualistic activities, given that it exhibited a num-ber of modifications (numerous cuts and damage tothe cranial roof and flattened occipital condyles)which could not be explained by standard butcher-ing practices (Dimitrijevi≤ et al. submitted), andthat it had been deposited at the top of a sequenceof several inhumations and cremations (Bori≤ 2010;Bori≤ et al. 2014.Tab. 1: the skull is described as ori-ginating from context 19). Unshed antlers collectedover the course of previous excavations indicate thesame broad (autumn-winter) time span, whereas, as
Fig. 2. Fragmented male red deer skull (VL 29/1)from Vlasac, with fragmented antlers attached topedicles, captured between September and Febru-ary.

already noted, the use of shed antlers (even thoughshedding occures within a short time span) as sea-sonal indicators should be taken with caution, as itis possible they were collected later.
The stages of teeth eruption and wear indicate sev-eral seasons of capture of red deer, some with moreprecision than others. In particular, a 3rd and 4th
decidious premolar from a fragmented left maxilla(VL 216/15) (Fig. 3) pointed to an age of 7–8 months,indicative of a December-January capture; whereas aleft 3rd permanent incisor with an open root andmilk incisors (left and right 2nd and 3rd) and rightcanine (VL 237/4) (Fig. 4) from a mandible that hadnot been preserved pointed to an approximat eageof 15–18 months or a kill that occurred between Au-gust and November.
A fragment of a roe deer skull with an antler frag-ment on a pedicle (VL 314 x. 23), as well as threeunshed antlers collected in previous excavations in-dicated that the capture seasons occurred betweenMay and October (cf. Schmidt 1972). Assuming theend of May as the season when roe deer give birth(cf. Habermehl 1985), one decidious premolar (3/4)(VL 51/1) indicated an age of one month, i.e. a Junecapture.
As already noted, the farrowing season for wildswine is generally from March to April (cf. Grupaautora 1991; Carter, Magnell 2007); in our study,the ageing and the estimation of season is givencounting from April (Tab. 1). Two teeth – an upper3rd deciduous premolar (VL 237/5) and an upper 2nd
deciduous premolar (VL 282/3) – produced fairly re-liable season estimates, an April-May and September-October kill respectively.
Fox teeth (an upper deciduous canine and a germ ofthe first molar, VL 222/22–23) were included in thisstudy, as they provided a fairly reliable kill season
(May), given that this stage of teeth eruption is in-dicative of a three-month-old individual (cf. Haber-mehl 1985).
Although most of the elements utilised in ageingoffered very broad season estimates spanning overseveral months (Tab. 1), the aforementioned spec-imens provided fairly accurate and short seasons ofdeath. They signal particular events which took placein December-January, April-May, May, June and Sep-tember-October, i.e. are indicative that hunting didnot take place during a limited period of the year.Both the fragment of a roe deer skull with an antler(VL 314 x. 23) and the red deer skull (VL 29/1) havebeen directly dated, producing a date of 7131–6823cal BC at 95% probability (OxA-21962) and 6006–5838 cal BC at 95% probability (OxA-16544) respec-tively (Bori≤ et al. 2014). At the same time, thesetwo dates are currently the oldest and the youngestobtained from the new excavations of Vlasac, thusdating the span of activities in the peripheral zoneof the settlement, with Transformation phase activ-ity (post c. 6200 cal BC) mainly related to the inter-ment of the latest burials in the successive burialzone closed by the red deer skull (Bori≤ et al. 2008;2014). A wild boar tooth (VL 282/3) was found ina context (a concentration of artefacts beneath thefloor of a dug-out dwelling) dated with two absolutedates: 7034–6692 cal BC at 95% probability (OxA-24809) and 7035–6698 cal BC at 95% probability(OxA-24810) (Bori≤ et al. 2014). From the previous1970–1971 excavations, encompassing the centralarea of the settlement, a roe deer skull with antlersfound on the floor of Building 2 (Srejovi≤, Letica1978) produced a date of 7047–6699 cal BC at 95%probability (OxA-16216) (Bori≤ et al. 2008). There-fore, it can be concluded that the majority of theseevents occurred during the last century of the 8th
millennium and the first half of 7th millennium BC,and could be taken as representative of Late Mesoli-thic resource exploitation patterns at the site.
Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
108
Fig. 3. 3rd and 4th decidious premolar from a fragmented red deer left maxilla (VL 216/15) from Vlasac,indicating an age of 7–8 months or a December-January capture: a lingual view; b buccal view; c roots,apical view.

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
109
Similarly to mammal remains, the fish faunal assem-blage from 2006–2009 displayed significant differ-ences in comparison to the one collected in 1970–1971. Only common carp (Cyprinus carpio), catfish,pike (Esox lucius), unidentified cyprinids and a greatnumber of unidentified fish bones were reported byBökönyi (1978). The absence of sturgeon was clear-ly a result of author-related bias, given that sturgeonremains were in fact identified in the preserved fau-nal sample from 1970–1971 (Bori≤ 2002.App. 5), aswell as in the sample from new excavations of Vla-sac (Ωivaljevi≤ in prep.) (Tab. 4). Albeit few, the re-mains of migratory beluga, Russian sturgeon andstellate sturgeon are indicative of spring and autumnfishing at the site, given that sturgeon remains as arule tend to be underrepresented due to their large-ly cartilaginous skeleton (Brinkhuizen 1986; Barto-siewicz et al. 2008). The exploitation of anadro-mous Black Sea shad (Alosa immaculata) wouldhave been even more seasonally restricted, as thespecies is present in the Danube only between theend of March and the beginning of June (Tab. 4).However, the vast majority of fish remains from thenew excavations of Vlasac originated from yet anoth-er anadromous species – the vyrezub (Rutilusfrisii) of the cyprinid family, mainly due to the largenumber of its pharyngeal teeth found in burials, butalso in ‘loose’ contexts. Burials with pharyngealteeth ornaments, from both the old and new excava-tions of Vlasac have mainly been related to the LateMesolithic phase, but have also been found in aTransformation phase burial at Vlasac and settle-ment deposits at Lepenski Vir (Cristiani, Bori≤ 2012;Cristiani et al. 2014; Bori≤ et al. 2014; Ωivaljevi≤ inprep.), whereas unmodified vyrezub teeth and bonesoccur as early as mid-10th millenium BC (Early Me-solithic) at Lepenski Vir (Ωivaljevi≤ in prep.). Thespecies has not been reported in the Middle/Lower
Danube previously, but it was probably migratinginto the river from the Black Sea at least up to theMiddle Holocene (Ωivaljevi≤ et al. submitted). Mo-dern Black Sea populations enter the rivers in Octo-ber, overwinter in freshwater, and return to estuar-ies after spawning, which takes place in April andMay (Kottelat, Freyhof 2007). Therefore, it can behypothesised that the main fishing seasons at Vla-sac corresponded with their autumn migration andspring spawning season, although the possibilitiesof winter catches should not be excluded. The possi-bility of opportunistic winter exploitation of fresh-water fish should also be allowed, although fishingduring this time would have been difficult, as mostspecies take refuge on the river bottom and hiber-nate until spring. The remains of pike, one of thefew species most succesfully caught from January toApril (Tab. 4), could be indicative of winter fishingat Vlasac.
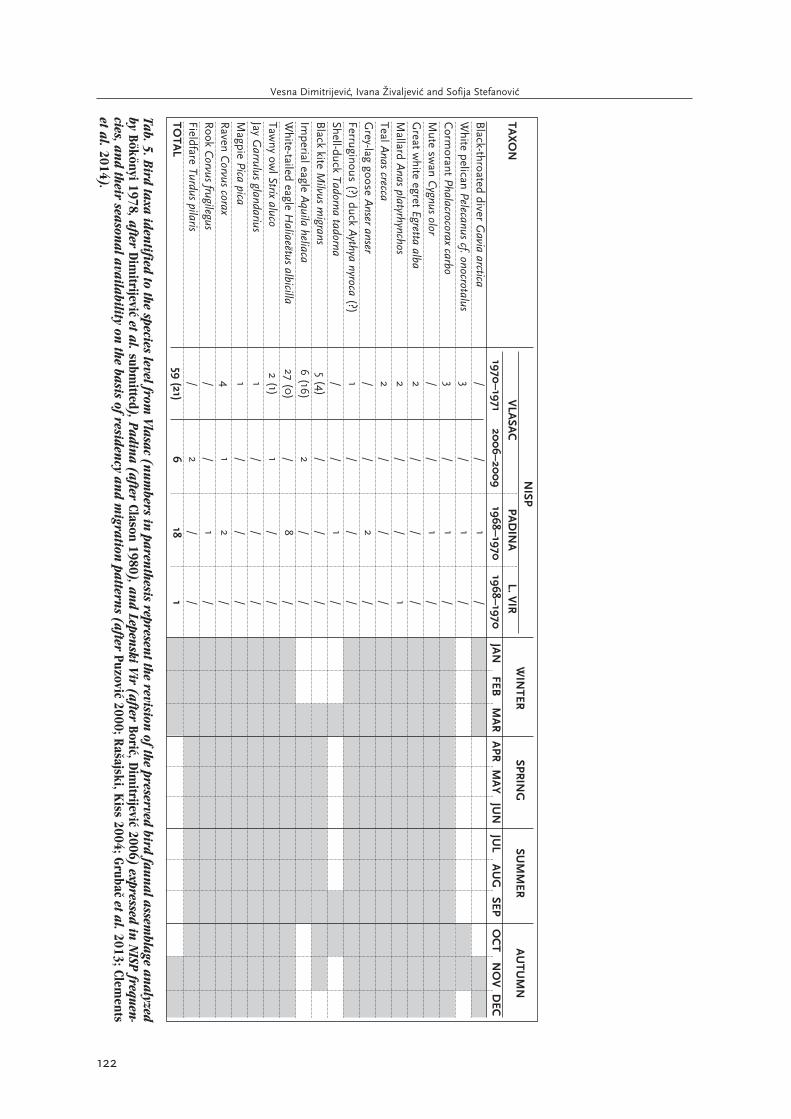
The bird faunal assemblage from both new and oldexcavations of Vlasac contained the remains of aqua-tic birds and waterfowl, as well as birds of prey andsongbirds. Given that bird remains were few evenin the hand-collected, water-sieved and flotated sam-ple from the new excavations (Tab. 5), it may be hy-pothesised that their exploitation was opportunistic.A somewhat greater frequency of imperial eagle(Aquila heliaca) and white-tailed eagle (Haliaeëtusalbicilla) remains (Bökönyi 1978; Dimitrijevi≤ etal. submitted), and the spatial clustering of some oftheir remains around the area of Late MesolithicBuilding 2 (Bori≤ 2002.190) could suggest that thesebirds of prey were possibly held in special regard.Most of the species represented in the 1970–1971and 2006–2009 assemblage are still resident in theDanube Gorges, and therefore their presence cannotbe used as a seasonal indicator. The only exceptions
Fig. 4. Red deer permanent and milk front teeth (VL 237/4) from a mandible that had not been preserv-ed from Vlasac, indicating an age of 15–18 months or an August-November kill: a right milk 2nd incisor;b right milk 3rd incisor; c right milk canine; d a germ of the left 3rd permanent incisor; e left 2nd milk in-cisor; f left 3rd milk incisor.

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
110
stuck to the crushed skull bones. The age of the ani-mal has been determined fairly accurately on thebasis of its upper teeth, which were in the last stagesof decidious/ permanent dentition replacement. Inboth right and left maxilla (LV 273/4), 3rd decidiousmolars were still holding above the crowns of per-manent fourth premolars, and it would have been amatter of days before they fell out. The 2nd and 3rd
premolars, and the left 3rd molar were erupting. Thisstage of dental development is related to an age of27 months, which would signal that this individualwas caught in October (Dimitrijevi≤ 2000; 2008; cf.Brown, Chapman 1991).
Another example is a red deer skull (LV 261/1–2)deposited on the floor of House 22 (Dimitrijevi≤2008.Fig. 3). At present, only antlers have been pre-served, one of which produced a date of 6084–5926cal BC at 95% probability (OxA-16075) (Bori≤, Di-mitrijevi≤ 2009), but both the skull (placed upsidedown) and antlers can be seen in a photographtaken during excavation (Fig. 5). The animal wasjust about to shed its antlers at the moment of cap-ture; as a result, they parted from the skull when ex-cavated. This occurrence is a fairly reliable indica-tor that the kill took place in February-March, andthe deposition of the skull shortly after. Given thatthe abandonment of aforementioned House 28 wasmarked by a red deer skull from an individual caughtin October, these two instances indicate that the ‘lifecycles’ of each building were probably unique, end-ing with similar preparations but at different timesthroughout the year. It would seem that certain buil-dings were in use after others fell into disuse; thispractice is also suggested by the deposition of sculpt-ed boulders. Most boulders were initially placed inthe rear area of rectangular hearths; however, cer-tain dwellings had empty sockets in the floor behindthe hearth area, or merely a fragment of an origi-nally larger boulder inserted in the socket, whichsuggests that some of the boulders were moved fromabandoned buildings and inserted at new locations(Srejovi≤ 1969; Bori≤ 2005). Consequently, the dataon seasons of abandonment enables a better under-standing of the individual histories of the housesand the occupation of the settlement as a whole.
Another fairly accurate season indicator comes froma context underneath of the floor of House 31, wherea cache of animal bones, including a large part of ared deer and a brown bear skeleton, as well as re-mains of chamois, wild boar, beluga, Russian stur-geon, common carp, vyrezub, catfish, huchen and anunidentified bird bone have been found (Dimitrije-
include the remains of white pelican (Pelecanus cf.onocrotalus) which migrates to the area in October-November, black kite (Milvus migrans) and imperi-al eagle which are passage visitors, present fromspring to autumn, and fieldfare (Turdus pilaris)which overwinters in the area (from November toMarch) (Tab. 5). In the case of the resident cormo-rant (Phalacrocorax carbo), teal (Anas crecca),and mallard (Anas platyrhynchos), it should benoted that their populations increase in winter,joined by visitors from Northwestern Europe (cf. Ra-∏ajski, Kiss 2004; Clason 1980.164). According toAnne Pike-Tay et alii (2004), teal and mallard aremigratory, and are present in neighbouring Hungaryin early spring and in autumn (the former used tobreed in Hungary), whereas the great white egret(Egretta alba) and ferruginous duck (Aythya nyro-ca) leave the area in winter. The data on present po-pulations in Serbia and the Danube Gorges (Ra∏aj-ski, Kiss 2004; Gruba≠ et al. 2013) suggest that theseare resident species, so their past migratory status isless clear.
Lepenski VirThe hand-collected and partially preserved faunalassemblage from Lepenski Vir contained fewer ele-ments sensitive to seasonality (Tab. 2), but their con-textual provenance enables a better understandingof the use of trapezoidal-base dwellings and the siteas a whole. At least 13 buildings contained red deerskulls with antlers (Bökönyi 1969; 1970; 1972),which were probably deposited on the floors, mark-ing the event of house abandonment (Dimitrijevi≤2000; 2008). Similarly to the structurally depositedred deer skull from Vlasac, these could have beencurated over longer periods of time, but in a coupleof cases provided a fairly reliable indication of thekill season.
One such example comes from House 28 in the west-ernmost (upstream) periphery of the settlement,where a red deer skull with unshed antlers (verythin and quite unusual in their asymmetry) hadbeen deposited on the floor (Dimitrijevi≤ 2000.Fig.16; 2008.Fig. 7). The skull has been directly datedto a range 6206–5989 cal BC at 95% probability(OxA-16078), which corresponds with the construc-tion and use of the trapezoidal buildings, i.e. thephase identified as the Mesolithic-Neolithic Transfor-mation (Bori≤, Dimitrijevi≤ 2009). The skull wasdamaged during excavation, with only the antlersand frontal, temporal and occipital parts of the skulland the upper jaws remaining (Dimitrijevi≤ 2000.Fig. 17), while fish and gastropod remains were

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
111
vi≤ 2000; Bori≤, Dimitrijevi≤ 2006; Ωivaljevi≤ inprep.). The context has not been AMS dated, butgiven that the deposition of faunal remains predat-ed the house construction, it can be concluded thatthe event(s) took place during an early settlementphase. The unity of the assemblage is seen in thelarge portions of the red deer and brown bear car-casses, which were probably butchered in situ. Thered deer remains originated from a young individ-ual, given its milk dentition, unfused proximal anddistal epiphyses of long bones and porous structureof the short bones. Its age has been determined onthe basis of a left mandible (LV bb-30/2) with a 2nd
and 3rd decidious molar, alveole for the 4th decidi-ous molar and an erupting 1st permanent molar (Di-mitrijevi≤ 2000.Fig. 12). This stage of dental devel-opment and wear indicate a four-month-old indivi-dual, suggesting that the kill season was in Septem-
ber (Dimitrijevi≤ 2000; cf. Brown, Chap-man 1991; Carter 2006). If we assume thatthe whole faunal assemblage underneathHouse 31 accumulated over a short period,September would correspond with the startof the autumn migration of beluga and Rus-sian sturgeon (Tab. 4), slightly precedingthe start of vyrezub migration, if migratorypatterns of modern populations are takeninto account.
Another red deer mandible (LV bb-14/3)(Fig. 6) from an unspecified context origi-nated from a 13- to 15-month-old individ-ual, judging from the presence of 2nd, 3rd
and 4th decidious molars with formed butunresorbed roots (cf. Brown, Chapman1991). This specimen is indicative of a sum-mer kill event that took place between Julyand September.
An autumn/winter kill is suggested in thecase of a red deer left maxilla (LV 1074/5)found in the eastern part of the settlement
(sq. a/11-14). The presence of both decidious 4th
molar and the 1st permanent molar indicates a 12-to 16-month-old individual (cf. Habermehl 1985),captured between October and February. The sameseason has been determined in the case of red deermaxillae (LV 1296/1) found beneath the floor ofHouse 19. Both left and right maxilla had mixed de-cidious/permanent dentition (D4, M1; and D3-D4,M1, respectively) (Fig. 7), suggesting they also orig-inated from a 12- to 16-month-old individual (cf.Habermehl 1985). In addition, vyrezub pharyngealteeth were stuck to the right maxilla. The October-February time span corresponds well with the sea-son during which vyrezub migrate to rivers and over-winters, but precedes its spawning season (Tab. 4).
The red deer maxilla with vyrezub teeth, as well asthe aforementioned red deer skull from House 28
Fig. 5. Red deer skull with antlers (LV 261/1–2) depositedfacing the floor of House 22 at Lepenski Vir; the kill tookplace in February-March. Photo from the archive of the Com-puter Documentation Centre, Faculty of Philosophy, Belgrade.
Fig. 6. Red deer right mandible with milk molars (LV bb-14/3) from Lepenski Vir, originating from a 13–15 month old individual, killed between July and September: a buccal; b lingual.

with small fish bones stuck to it seem to embodyquite literally the merging role of wild game andfish. The fish faunal assemblage from Lepenski Virconsisted mainly of cyprinid remains (of freshwaterspecies and anadromous vyrezub), as well as catfish,huchen and a small number of Black Sea shad, pikeand pikeperch (Sander lucioperca) bones, but alsoa significant amount of remains of migratory stur-geon. Having in mind their largely cartilaginous ske-leton and preservation bias affecting their remains(Brinkhuizen 1986; Bartosiewicz et al. 2008), it isremarkable that sturgeon constitute nearly 20% (interms of NISP) of identified fish remains at Lepen-ski Vir (Ωivaljevi≤ in prep.); it is also highly signifi-cant that some of the sculpted boulders from thissite bore a resemblance to sturgeon features (Rado-vanovi≤ 1997; Bori≤ 2005). At the same time, theLepenski Vir assemblage exhibited the greatest di-versity of sturgeon species, including the remains ofmigratory beluga, Russian sturgeon, ship sturgeon,and stellate sturgeon, as well as those of freshwatersterlet (Acipenser ruthenus). The first species toenter the Danube is the large beluga, whose springmigration commences in late winter/early spring,reaching a peak in April and ending in late May/June. Its autumn migration starts by late August,peaking in October/November. The Russian sturgeonstarts its spring migration somewhat later than bel-uga (end of February/March), which peaks in April,and ends mid-May. The second migration starts inAugust/September, peaks in late September/mid-Oc-tober and ends by mid-November. The ship sturgeonmigrates in spring and autumn, with autumn popu-lations remaining in the river until spawning, whichtakes place from March/April until June. The stellatesturgeon, the last species to enter the Danube, startsits spring migration in late April/ March, reaching
a peak in May. Its autumn run begins by August andends by September/mid-October (Tab. 4) (cf. Dinu2010; Petrovi≤ 1998[1941]; Risti≤ 1977; Simonovi≤2001; Kottelat, Freyhof 2007; Bartosiewicz et al.2008). Judging from interspecific variations in themigration start and peak dates of sturgeon species,migration pattern of vyrezub, as well as availabili-ty of freshwater fish, it might be hypothesised thatfishing could have successfully taken place betweenlate winter and late autumn, with particular inten-sity in spring and autumn. The remains of speciesactive in winter (namely pike) are few, but the pos-sibility of smoking and drying fish (the hearths inparticular could have been used for this purpose)and storing it for winter months should not be ex-cluded (cf. Bonsall 2008; Bori≤ 2002; 2007). On thebasis of fish remains from AMS dated contexts fromLepenski Vir (cf. Whittle et al. 2002; Bonsall et al.1997; 2008; Bori≤, Dimitrijevi≤ 2009; Bori≤ 2011),it was determined that fishing patterns remainedrelatively unchanged throughout the temporal se-quence, i.e. that the distribution and proportion ofspecies is more or less the same in both Mesolithicand Transformation phase contexts at the site (Ωi-valjevi≤ in prep.).
Although bird remains have been uncovered at Le-penski Vir (Bökönyi 1969; 1970; 1972; Dimitrijevi≤2008), only a single bird bone found between thesuperimposed floors of Houses 20 and 33 (contextdated to 6231–6056 cal BC at 95% probability (OxA-15998) (Bori≤, Dimitrijevi≤ 2009) has been iden-tified to the species level, originating from mallard(Bori≤, Dimitrijevi≤ 2006). As already noted, AnnePike-Tay et alii (2004) state that the species is mi-gratory, but used to breed in Hungary; whereas Ja-vor Ra∏ajski and Andrei Kiss (2004) note that it is
Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
112
Fig. 7. Red deer left andright fragmented maxil-lae (LV 1296/1) found be-neath the floor of House19 at Lepenski Vir, origi-nating from a 12–16month old individual,captured between Octo-ber and February. A frag-ment of vyrezub left pha-ryngeal bone with a rowof 4 preserved teeth isattached to the right max-illa. a a fragment of theright maxilla with thetwo milk molars (D3 andD4) and the first molar (M1) in eruption, lingual, attached vyrezub pharyngeal bone at the left; bsame, buccal view, vyrezub teeth seen at the right side; c a fragment of the left maxilla with the lastmilk (D4) and the first permanent molar (M1), lingual; d same, buccal view.

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
113
resident in Serbia, with a winter population increasedue to migrant populations from the North of Europe.
PadinaThe faunal assemblage from Padina, albeit entirelypreserved, contained the fewest elements for a reli-able estimate of occupational seasons (Tab. 3). Si-milarly to Lepenski Vir, this was mainly due to thehand-collection of animal bone, particularly biasedtowards fragile deciduous teeth and fish and birdremains. The majority of elements indicative of sea-sonality included shed and unshed red deer and roedeer antlers from both Mesolithic and Transforma-tion phase contexts. These indicate kill seasons whichoccurred between September and February, deter-mined on the basis of unshed red deer antlers, andMay and October, on the basis of unshed roe deerantlers. The occurrence of shed antlers of red androe deer indicate occupation in February-March andOctober-November, respectively, assuming they werecollected shortly after shedding. The only mandiblefrom a juvenile animal (PAD 28.8.69/1) originatedfrom a roe deer, and was found in a Mesolithic occu-pational context (cf. Bori≤ 2002.App. 3). Based onthe presence of alveoli for D2-D3, D4 and a fragmentof M1 (Fig. 8), it was determined that the animalwas between four and eight months old (cf. Carter2006), which suggested that it was captured betweenSeptember and January.
In terms of fish remains, a specific feature of thefaunal assemblage from Padina (even in comparisonwith other assemblages collected by hand) is theabsolute predominance of catfish (69.9% of all iden-tified fish remains, and 77.1% of all fish remainsidentified to species) and least species diversity(Tab. 4) (Ωivaljevi≤ in prep.). Moreover, judgingfrom the species distribution in AMS dated contexts(cf. Whittle et al. 2002; Bori≤, Miracle 2004; Bori≤2011) and in contexts attributed to particular phas-es on the basis of stratigraphy (cf. Bori≤ 2002.App.3), it was determined that catfish remains becomeexceptionally abundant in the Transformation phaseof the settlement. This could suggest that Padinaestablished itself as a specialised centre oriented to-wards catching catfish during this time (Ωivaljevi≤in prep.). Catfish is most successfully caught duringits spawning season (from May to July) and duringsummer months, when it feeds most intensely (Ri-sti≤ 1977; Simonovi≤ 2001; Kottelat, Freyhof 2007;Dinu 2010); however, given that the species is sta-tionary, and hardly moves from its respective habi-tat (Risti≤ 1977), it can be hypothesised that it wasavailable for most of the year. During cold seasons,
however, catfish populations would decrease innumber, as the species requires a water temperatureof at least 18°C to reproduce (cf. Bartosiewicz, Bon-sall 2004.Tab. 7). As already suggested, the harshwinter seasons and periods when the river was co-vered in ice could be overcome by storing fish bysmoking and drying (cf. Bonsall 2008; Bori≤ 2002;2007). In addition, the presence of anadromoussturgeons and vyrezub in the faunal assemblage sug-gests that freshwater fish exploitation at Padina wassupplemented with fishing of seasonally availablespecies, most notably in spring and autumn.
Bird remains are few in the faunal assemblage fromPadina, which is suggestive of opportunistic exploi-tation; however, it is probably also a consequence ofthe sampling bias. The faunal assemblage consistedof the remains of six resident and three migratoryspecies, some of them available in the Danube Gor-ges area for fairly short and seasonally limited pe-riods of time (Tab. 5). Such is the case with the shell-duck (Tadorna tadorna), which is present only inMarch and September-October, and white pelican,present in October-November. The black-throateddiver (Gavia arctica) is present in the area from No-vember to March (Ra∏ajski, Kiss 2004), whereas inCentral Europe it migrates in spring and autumn (Cla-son 1980). As already noted, the numbers of someof the resident populations (such as cormorant) in-crease in winter (cf. Clason 1980; Ra∏ajski, Kiss2004), which tentatively suggests they could havebeen exploited more frequently during this time.
Discussion
An important issue to be addressed here is whetherpermanent, year-round occupation can be distingui-shed from a series of repeated visits, of varying du-
Fig. 8. Roe deer mandible with alveoli for milk mo-lars (D2-D3), last milk molar (D4) and a fragmentof the first permanent molar (PAD 28.8. 69/1) froma Mesolithic occupational context at Padina. Theanimal was between 4–8 months old, suggestingthe capture occurred between September and Janu-ary.

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
114
ration, occurring at different seasons throughoutthe year? (cf. Whittle 2001; Conneller et al. 2012).This question, of course, remains very difficult toanswer archaeologically. The patterns of food re-source exploitation at Vlasac, corroborated witharchaeological evidence, could be indications of thefirst scenario. The intensity of occupation in the LateMesolithic phase (c. 7400–6300/6200 cal BC) is evi-denced by several dug-out dwellings vaguely resem-bling the trapezoidal shape in plan, numerous stone-lined hearths which were constructed in the open orwere originally incorporated into dwellings that leftno trace in the archaeological record, over hundredburied individuals and an enormous assemblage oftools and animal bones (cf. Srejovi≤, Letica 1978;Nemeskéri 1978; Bökönyi 1978; Kozłowski, Koz-łowski 1982; Roksandi≤ 1999; 2000; Bori≤ et al.2008; 2014). The faunal assemblage from the new2006–2009 excavations of the site, which containedmost elements suggestive of kill seasons (namely,erupting teeth of young animals), seems to indicateyear-round occupation of the settlement. The exploi-tation patterns of red deer, the most important gameanimal, do not suggest any significant seasonal hia-tus. Similarly, wild boar and roe deer hunting doesnot seem restricted to a particular season. Althoughmost elements utilised in this study indicated a rel-atively broad time span, those that provided mostprecise and temporally restricted seasonal estimatespoint to hunting events which took place in all sea-sons. The fishing activity is indicative of spring andautumn months, judging from the presence of ana-dromous vyrezub and various sturgeon species, butfishing at other times was certainly possible, giventhat freshwater species represented in the samplewould have been available for longer periods oftime. Moreover, the fish could have been stored forwinter by smoking and drying.
The hand-collected faunal assemblages from Lepen-ski Vir and Padina offered much less information forprecise season determination, but tentative estimatesdo not point to a particular season of hunting. Fur-thermore, it can be suggested that resource exploita-tion patterns observed in the Late Mesolithic wouldhave continued in the Transformation phase settle-ments (c. 6200–5900 cal BC). Several fairly preciseseason estimates obtained from specimens relatedto the construction and use of elaborate trapezoidal-base buildings at Lepenski Vir offer important in-sights into the life-cycles of particular buildings and,consequently, of the settlement as a whole. It is evi-dent that some of dwellings were abandoned at dif-ferent times of the year, while others remained in
use; this feature is also suggested by the secondaryposition of some of the sculpted boulders. The signi-ficance of both hunting and fishing activities, whichtook place simultaneously for most of the year, isparticularly illustrated by caches of mammal andfish bones stuck together and formed over a shortperiod of time. It remains unclear to what extent theobserved differences in preferences for fish speciesat Lepenski Vir and Padina would influence the na-ture of occupation of these sites; the latter orientedmostly towards catfish available for most of the year,and the former exhibiting greater species diversityand a significant amount of seasonably availablesturgeon. Whereas the environment of Lepenski Virand other sites would have provided substantialmeans for occupation lasting throughout the year,the exploitation of seasonally available fish couldhave been vital in (re)creating memories, eventsand specific temporal perspectives (Bori≤ 2002).
Conclusion
This study offers additional data supporting the hy-pothesis that a significant degree of sedentism inthe Danube Gorges existed already in the Late Me-solithic, prior to, and independently of, the adoptionof animal and plant husbandry. Furthermore, recentstudies of other Mesolithic contexts in Europe pro-vide evidence of practices involving prolonged staysof human groups in certain locations; for example,the construction (involving a significant amount oftime and effort) of substantial timber structures atStarr Car in Britain (Conneller et al. 2012) and large-scale food storage and fish fermentation, implyinga delayed-return subsistence strategy, at Norje-Sun-nansund in Southern Scandinavia (Boethius 2016).Moreover, ‘full-scale’ sedentism commonly associatedwith food-producing communities could have alsotaken many different forms, including shifting culti-vation crops or herds, and varying levels of mobili-ty of individuals involved in such practices (Whittle2001). These studies further problematise the rela-tionship of sedentism and subsistence strategies, andadd to our understanding of the complexity anddiversity of forager and food-producing lifestyle andresidential practices. In the Danube Gorges, evidenceof Early Mesolithic occupation is relatively scarce,but the presence of several burials and stone con-structions indicate a prolonged stay at riverine ter-races. The higher resolution of dates, architecturalfeatures and artefacts related to the Late Mesolithicoccupancy signal that by this time these locationswere a home to fairly sedentary populations prac-ticing an exclusively hunter-gatherer-fisher lifestyle.

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
115
Their environment enabled the successful exploita-tion of wild game and fish for most, if not the whole,of the year.
Therefore, it can be argued that the construction oflater, Mesolithic-Neolithic Transformation phase set-tlements at Lepenski Vir and Padina took place ina context where continuous human occupation andrelationship with particular locales had been longestablished. It can also be hypothesised that thesesettlements, with more elaborate and permanentdwellings, would have followed similar patterns ofoccupation. Although the changes observed in theTransformation phase largely coincide with theemergence of the first food-producing communitiesin the region (and contacts with distant communi-ties are evident already in the Late Mesolithic), cer-tain (but not all) elements of Neolithic lifestyles wereincorporated only after “they made sense in localterms” (Bori≤ 2007). Most notably, animal husban-dry had fairly little impact on the economy and life-style of Upper Gorge communities, with the remainsof the first domesticates appearing only after mostof the trapezoidal buildings had been abandoned.
It can be assumed then, that local traditions, estab-lished in a particular environment and involvinglocally available resources, merged with new cultur-al elements in the later parts of the sequence, playedsubstantial roles in settlement practices in the Da-nube Gorges context.
This paper is a result of the Project “BIRTH: Births,mothers and babies: prehistoric fertility in the Balkansbetween 10 000–5000 BC”, funded by the EuropeanResearch Council (ERC) under the European Union’sHorizon 2020 research and innovation programme(Grant Agreement No. 640 557). We thank Du∏an Bo-ri≤ for the opportunity to study the faunal remainsfrom the 2006–2009 excavations of Vlasac, Igor Aske-yev for his help with determining some of the fishbones, and Mihael Budja for his kind invitation toparticipate in the 22nd Neolithic Seminar and for thesuperb organisation of the conference. We would alsolike to thank two anonymous reviewers for their in-sightful and constructive comments on earlier draftsof this paper.
ACKNOWLEDGEMENTS
References
Bartosiewicz L. 1989. Variability of tooth eruption in cat-tle. Acta Veterinaria Hungarica 37(4): 303–317.
Bartosiewicz L., Bonsall C. 2004. Prehistoric fishing alongthe Danube. Antaeus 27: 253–272.
Bartosiewicz L., Bonsall C. and Sisu V. 2008. Sturgeon fish-ing along the Middle and Lower Danube. In C. Bonsall,V. Boroneant and I. Radovanovi≤ (eds.), The Iron Gatesin Prehistory. New perspectives. British ArchaeologicalReports IS 1893. Archaeopress. Oxford: 39–54.
Bartosiewicz L., Boroneant V., Bonsall C. and Stallibrass S.1995. Schela Cladovei: a preliminary review of the prehi-storic fauna. Mesolithic Miscellany 16(2): 2–19.
2001. New data on the prehistoric fauna of the IronGates: a case study from Schela Cladovei, Romania. InR. Kertész, J. Makkay (eds.), From the Mesolithic to theNeolithic. Proceedings of the International Archaeologi-cal Conference held in the Damjanich Museum of Szol-nok, September 22–27, 1996. Archaeolingua. Budapest:15–21.
Blaise E., Balasse M. 2011. Seasonality and season of birthof modern and late Neolithic sheep from south-eastern
France using tooth enamel δ18O analysis. Journal ofArchaeological Science 38(11): 3085–3093.
Boethius A. 2016. Something rotten in Scandinavia: Theworld’s earliest evidence of fermentation. Journal of Ar-chaeological Science 66: 169–180.
Bökönyi S. 1969. Ki≠menjaci (prethodni izve∏taj). In D.Srejovi≤ (ed.), Lepenski Vir. Nova praistorijska kultura uPodunavlju. Srpska knji∫evna zadruga. Beograd: 224–228.
1970. Animal Remains from Lepenski Vir. Science 167(926): 1702–1704.
1972. The vertebrate fauna. In D. Srejovi≤ (ed.), Eu-rope’s first monumental sculpture: new discoveries atLepenski Vir. Thames and Hudson. London: 186–189.
1978. The vertebrate fauna of Vlasac. In M. Gara∏anin(ed.), Vlasac. A Mesolithic Settlement in the Iron Gates.Volume II. Geology – Biology – Anthropology. SerbianAcademy of Sciences and Arts Monographies. Belgrade:35–65.
Bonsall C. 2008. The Mesolithic of the Iron Gates. In G. N.Bailey, P. Spikins (eds.), Mesolithic Europe. CambridgeUniversity Press. Cambridge: 238–279.
∴∴

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
116
Bonsall C., Cook G. T., Hedges R. E. M., Higham T. F. G.,Pickard C. and Radovanovi≤ I. 2004. Radiocarbon and sta-ble isotope evidence of the dietary change from the Me-solithic to the Middle Ages in the Iron Gates: new resultsfrom Lepenski Vir. Radiocarbon 46: 293–300.
Bonsall C., Lennon R., McSweeney K., Stewart C., Hark-ness D., Boroneant V., Bartosiewicz L., Payton R. and Chap-man J. 1997. Mesolithic and Early Neolithic in the IronGates: a palaeodietary perspective. Journal of EuropeanArchaeology 5(1): 50–92.
Bonsall C., Radovanovi≤ I., Roksandi≤ M., Cook G., Hig-ham T. and Pickard C. 2008. Dating burials and architec-ture at Lepenski Vir. In C. Bonsall, V. Boroneant and I. Ra-dovanovi≤ (eds.), The Iron Gates in Prehistory. New per-spectives. British Archaeological Reports IS 1893. Archaeo-press. Oxford: 175–204.
Bori≤ D. 2001. Mesolithic and early Neolithic hunters andfishers in the Danube Gorges: an analysis of archaeozo-ological data. In R. Kertész, J. Makkay (eds.), From theMesolithic to the Neolithic. Proceedings of the Internatio-nal Archaeological Conference held in the Damjanich Mu-seum of Szolnok, September 22–27, 1996. Archaeolingua.Budapest: 101–124.
2002. Seasons, Life Cycles and Memory in the DanubeGorges, c. 10,000–5500 BC. Unpublished PhD thesis.University of Cambridge. Cambridge.
2005. Body metamorphosis and animality: volatile bod-ies and boulder artworks from Lepenski Vir. CambridgeArchaeological Journal 15(1): 35–69.
2006. New discoveries at the Mesolithic-Early Neolithicsite of Vlasac: preliminary notes. Mesolithic Miscellany18(1): 7–14.
2007. The House between Grand Narrative and Micro-history: a house society in the Balkans. In R. A. Beck(ed.), The Durable House. House Society Models inArchaeology. South Illinois University, Center for Ar-chaeological Investigations, Occasional Paper No. 35.Carbondale: 97–129.
2010. Happy forgetting? Remembering and dismem-bering dead bodies at Vlasac. In D. Bori≤ (ed.), Archaeo-logy and Memory. Oxbow Books. Oxford: 48–67.
2011. Adaptations and transformations of the DanubeGorges foragers (c. 13,000–5500 BC): an overview. InR. Krauß (ed.), Beginnings – New Research in the Ap-pearance of the Neolithic between Northwest Anato-lia and the Carpathian Basin. Verlag Marie LeidorfGmbh. Rahden: 157–203.
Bori≤ D., Dimitrijevi≤ V. 2006. Continuity of foraging stra-tegies in the Mesolithic-Neolithic transformations: dating
faunal patterns at Lepenski Vir (Serbia). Atti della Socie-tà per la preistoria e protoistoria della regione Friuli-Venezia Giulia XV (2004–05): 33–107.
2007. When did the ‘Neolithic package’ reach LepenskiVir? Radiometric and faunal evidence. DocumentaPraehistorica 34: 52–71.
2009. Apsolutna hronologija i stratigrafija LepenskogVira. Starinar LVII/2007: 9–55.
Bori≤ D., French C. and Dimitrijevi≤ V. 2008. Vlasac re-visited: formation processes, stratigraphy and dating. Do-cumenta Praehistorica 35: 261–287.
Bori≤ D., French C. A. I., Stefanovi≤ S., Dimitrijevi≤ V., Cri-stiani E., Gurova M., Antonovi≤ D., Allué E. and Filipovi≤D. 2014. Late Mesolithic lifeways and deathways at Vla-sac (Serbia). Journal of Field Archaeology 39(1): 4–31.
Bori≤ D., Grupe G., Peters J. and Miki≤ Ω. 2004. Is the Me-solithic-Neolithic subsistence dichotomy real? New stableisotope evidence from the Danube Gorges. EuropeanJournal of Archaeology 7(3): 221–248.
Bori≤ D., Miracle P. 2004. Mesolithic and Neolithic (dis)continuities in the Danube Gorges: new AMS dates fromPadina and Hajdu≠ka Vodenica (Serbia). Oxford Journalof Archaeology 23(4): 341–371.
Bori≤ D., Price T. D. 2013. Strontium isotopes documentgreater human mobility at the start of the Balkan Neoli-thic. Proceedings of the National Academy of Sciencesof USA 110(9): 3298–3303.
Brinkhuizen D. C. 1986. Features observed on the skele-tons of some recent European Acipenseridae: their impor-tance for the study of excavated remains of sturgeon. InD. C. Brinkhuizen, A. T. Clason (eds.), Fish and Archaeo-logy. Studies in osteometry, taphonomy, seasonality andfishing methods. British Archaeology Reports IS 294.Archaeopress. Oxford: 18–33.
Brown W. A. B., Chapman N. G. 1991. The dentition ofred deer (Cervus elaphus): a scoring scheme to assess agefrom wear of the permanent molariform teeth. Journalof Zoology 224(4): 519–536.
Bull G., Payne S. 1982. Tooth eruption and epiphysealfusion in pigs and wild boar. In B. Wilson, C. Grigson andS. Payne (eds.), Ageing and sexing animal bones fromarchaeological sites. British Archaeology Reports IS 109.Archaeopress. Oxford: 55–71.
Carter R. J. 2006. A method to estimate the ages at deathof red deer (Cervus elaphus) and roe deer (Capreolus ca-preolus) from developing mandibular dentition and itsapplication to Mesolithic NW Europe. In D. Ruscillo (ed.),Recent advances in ageing and sexing animal bones.

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
117
Proceedings of the 9th ICAZ conference. Oxbow Books.Oxford: 40–61.
Carter R. J., Magnell O. 2007. Age estimation of wild boarbased on molariform mandibular tooth development andits application to seasonality at the Mesolithic site of Ring-kloster, Denmark. In U. Albarella, K. Dobney, A. Ervynckand P. Rowley-Konwy (eds.), Pigs and humans: 10,000years of interaction. Oxford University Press. Oxford:197–217.
Chapman J. 1993. Social power in the Iron Gates Mesoli-thic. In J. Chapman, M. P. Dolukhanov (eds.), CulturalTransformations and Interactions in Eastern Europe.Aldershot. Avebury: 71–121.
Clason A. T. 1980. Padina and Star≠evo: game, fish andcattle. Palaeohistoria 22: 141–173.
Clements J. F., Schulenberg T. S., Iliff M. J., Roberson D.,Fredericks T. A., Sullivan B. L. and Wood C. L. 2014. TheeBird/Clements checklist of birds of the world. Version6.9. http://www.birds.cornell.edu/clementschecklist/download/
Conneller C., Milner N., Taylor B. and Taylor M. 2012.Substantial settlement in the European Early Mesolithic:new research at Star Carr. Antiquity 86(2012): 1004–1020.
Cristiani E., Bori≤ D. 2012. 8500-year-old Late Mesolithicgarment embroidery from Vlasac (Serbia): technological,use-wear and residue analyses. Journal of Archaeologi-cal Science 39(11): 3450–3469.
Cristiani E., Ωivaljevi≤ I. and Bori≤ D. 2014. Residue ana-lysis and ornament suspension techniques in prehistory:Cyprinid pharyngeal teeth beads from Late Mesolithic bu-rials at Vlasac (Serbia). Journal of Archaeological Science46: 292–310.
Dimitrijevi≤ V. 2000. The Lepenski Vir fauna: bones inhouses and between houses. Documenta Praehistorica27: 101–117.
2008. Lepenski Vir animal bones: what was left in thehouses? In C. Bonsall, V. Boroneant and I. Radovanovi≤(eds.), The Iron Gates in Prehistory. New perspectives.British Archaeological Reports IS 1893. Archaeopress.Oxford: 117–130.
Dimitrijevi≤ V., Vukovi≤ S. 2015. Was the dog locally do-mesticated in the Danube Gorges? Morphometric study ofdog cranial remains from four Mesolithic–Early Neolithicarchaeological sites by comparison with contemporarywolves. International Journal of Osteoarchaeology 25(1): 1–30.
Dimitrijevi≤ V., Ωivaljevi≤ I. and Bla∫i≤ S. Submitted. Fau-nal remains. In D. Bori≤ (ed.), Vlasac: a Mesolithic site inthe Danube Gorges revisited.
Dinu A. 2010. Mesolithic fish and fishermen of the lowerDanube (Iron Gates). Documenta Praehistorica 37: 299–310.
Gara∏anin M., Radovanovi≤ I. 2001. A pot in House 54at Lepenski Vir I. Antiquity 75: 118–125.
Gruba≠ B., Milovanovi≤ Z. and πekler M. 2013. Ptice ∑er-dapa. Princip pres: Zavod za za∏titu prirode Srbije – Na-cionalni park “∑erdap” – Veterinarski specijalisti≠ki in-stitut “Kraljevo”. Beograd – Donji Milanovac – Kraljevo.
Grupa autora 1991. Velika ilustrovana enciklopedija lov-stva. DIP, gra∂evinska knjiga – DNEVNIK. Beograd – NoviSad.
Grupe G., Miki≤ Ω., Peters J. and Manhart H. 2003. Verte-brate food webs and subsistance strategies of Meso andNeolithic populations of Central Europe. In G. Gruppe, J.Peters (eds.), Documenta Archaeobiologiae 1. Jahrbuchder Staatssammlung für Anthropologie und Paläoana-tomie. Verlag Marie Leidorf, Rahden/Westf. München:193–213.
Habermehl K.-H. 1985. Altersbestimmung bei Wild-undPelztieren. Verlag Paul Parey. Hamburg, Berlin.
Jovanovi≤ B. 2008. Micro-regions of the Lepenski Vir cul-ture: Padina in the Upper Gorge and Hajdu≠ka Vodenicain the Lower Gorge of the Danube. Documenta Praehisto-rica 35: 289–324.
Kottelat M., Freyhof J. 2007. Handbook of EuropeanFreshwater Fishes. Publications Kottelat. Cornol.
Kozłowski J. K., Kozłowski S. K. 1982. Lithic industriesfrom the multi-layered Mesolithic site Vlasac, in Yugosla-via. In J. K. Kozłowski (ed.), Origin of the Chipped StoneIndustries of the Early Farming Cultures in the Balkans.Prace archeologiczne 33. Panstwowe Wydawnictwo Nau-kowe. Warszawa-Krakow: 11–109.
Nehlich O., Bori≤ D., Stefanovi≤ S. and Richards M. P.2010. Sulphur isotope evidence for freshwater fish con-sumption: a case study from the Danube Gorges, SE Eu-rope. Journal of Archaeological Science 37: 1131–1139.
Nemeskéri J. 1978. Demographic structure of the VlasacEpipalaeolithic population. In M. Gara∏anin (ed.), Vlasac.A Mesolithic Settlement in the Iron Gates. Volume II.Geology – Biology – Anthropology. Serbian Academy ofSciences and Arts Monographies. Belgrade: 97–133.
Petrovi≤ M. 1998. [orig. 1941]. ∑erdapski ribolovi u pro-∏losti i sada∏njosti. In D. Trifunovi≤ (ed.), Ribarstvo. Sa-

Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
118
brana dela Mihaila Petrovi≤a. Zavod za ud∫benike i nas-tavna sredstva. Beograd: 175–270.
Pike-Tay A., Bartosiewicz L., Gál E. and Whittle A. 2004.Body part representation and seasonality: sheep/goat,bird and fish remains from Early Neolithic Ecsegfalva23, SE Hungary. Journal of Taphonomy 2(4): 221–246.
Puzovi≤ S. 2000. Atlas ptica grabljivica Srbije – maperasprostranjenosti i procene brojnosti 1977–1996. Za-vod za za∏titu prirode Srbije. Beograd.
Radovanovi≤ I. 1996. The Iron Gates Mesolithic. Interna-tional Monographs in Prehistory. Ann Arbor.
1997. The Lepenski Vir culture: a contri-bution to in-terpretation of its ideological aspects. In D. Srejovi≤, M.Lazi≤ (eds.), Antidoron Dragoslavo Srejovi≤ completisLXV annis ab amicis, collegis, discipulis oblatum. Cen-tar za arheolo∏ka istra∫ivanja. Beograd: 87–93.
Radovanovi≤ I., Voytek B. 1997. Hunters, fishers or far-mers: sedentism, subsistence and social complexity in the∑erdap Mesolithic. In A. van Gijn, C. Bakels and M. Zvele-bil (eds.), Ideology and Social Structure of Stone AgeCommunities in Europe. Analecta Praehistorica Leiden-sia 29: 19–31.
Ra∏ajski J., Kiss A. 2004. Ptice Banata. Gradski muzej Vr-∏ac. Vr∏ac.
Risti≤ M. 1977. Ribe i ribolov u slatkim vodama. Nolit.Beograd.
Roksandi≤ M. 1999. Transition from Mesolithic to Neoli-thic in the Iron Gates Gorge: Physical Anthropology Per-spective. Unpublished PhD thesis. Department of Anthro-pology. Simon Fraser University. Vancouver.
Roksandi≤ M. 2000. Between foragers and farmers in theIron Gates Gorge: physical anthropology perspective. ∑er-dap population in transition from Mesolithic to Neolithic.Documenta Praehistorica 27: 1–100.
Schmid E. 1972. Atlas of Animal Bones: For Prehisto-rians, Archaeologists and Quaternary Geologists. Else-vier Publishing Company. Amsterdam, London, New York.
Simonovi≤ P. 2001. Ribe Srbije. NNK Internacional. Bio-lo∏ki fakultet Univerziteta u Beogradu. Zavod za za∏tituprirode Srbije. Beograd.
Srejovi≤ D. 1969. Lepenski Vir: Nova praistorijska kultu-ra u Podunavlju. Srpska Knji∫evna Zadruga. Beograd.
1972. Europe’s first monumental sculpture: new disco-veries at Lepenski Vir. Thames and Hudson. London.
Srejovi≤ D., Letica, Z. 1978. Vlasac. Mezolitsko naselje u∑erdapu (I arheologija). Srpska akademija nauka i umet-nosti. Beograd.
Takács I., Bartosiewicz L. 1998. Data on the history of fishexploitation in Hungary. Internet Archaeology 4. http://intarch.ac.uk/journal/issue4/takacs_index.html
Voytek B. A., Tringham R. 1989. Rethinking the Mesoli-thic: the case of South-East Europe. In C. Bonsall (ed.),The Mesolithic in Europe. Papers Presented at the ThirdInternational Symposium Edinburgh 1985. John DonaldPublishers Ltd. Edinburgh: 492–499.
Wheeler A., Jones A. K. G. 1989. Fishes. Cambridge Ma-nuals in Archaeology. Cambridge University Press. Cam-bridge.
Whittle A. 2001. From mobility to sedentism: change bydegrees. In R. Kertész, J. Makkay (eds.), From the Mesoli-thic to the Neolithic. Proceedings of the International Ar-chaeological Conference held in the Damjanich Museumof Szolnok, September 22–27, 1996. Archaeolingua. Buda-pest: 447–461.
Whittle A., Bartosiewicz L., Bori≤ D., Pettitt P. and RichardsM. 2002. In the beginning: new radiocarbon dates for theEarly Neolithic in Northern Serbia and South-East Hun-gary. Antaeus 25(2002): 63–117.
Ωivaljevi≤ I. in prep. Ribolov na ∑erdapu u ranom holo-cenu (10. – 6. milenijum pre n. e.). Unpublished PhD the-sis. Department of Archaeology. Faculty of Philosophy.University of Belgrade. Belgrade.
Ωivaljevi≤ I., Popovi≤ D., Snoj A. and Mari≤ S. submitted.Ancient DNA analysis of cyprinid remains from the Meso-lithic-Neolithic Danube Gorges reveals an extirpated fishspecies Rutilus frisii (Nordmann, 1840).

Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
119
TAX
ON
INV
. NO
.ELEM
ENT
WIN
TERSPR
ING
SUM
MER
AU
TUM
NR
eference where the age
JAN
FEBM
AR
APR
MA
YJU
NJU
LA
UG
SEPO
CT
NO
VD
ECstage has been defined
VL 29\1
skull fragment w
ith antlersSchm
idt 1972\
unshed antler (9)*Schm
idt 1972\
shed antler (31)*Schm
idt 1972V
L 216\15m
axilla sin. (D3, 4)
Carter 2006
VL 145 x.5
P4 inf. dext.C
arter 2006V
L 204\5
D3 sup.sin.
Carter 2006
Red deer
VL 20
9 x.1P4 inf. dext.sin.
Carter 2006
Cervus elaphus
VL 224\6
D2 inf.dext.
Carter 2006
VL 237\4
I3, Di2, D
i3 sin. & D
i2, Di3, D
c dext.H
abermehl 1985
VL 264 x.2
D3 sup.dext.
Carter 2006
VL 286\3
M2, M
3 inf. dext.C
arter 2006V
L 306\24
P2 inf.sin.C
arter 2006V
L 35\2P4 inf.sin.
Carter 2006
VL 66 x.2
P4 inf. dext.C
arter 2006V
L 314 x.23skull fragm
ent with left antler
Schmidt 1972
Roe deer
\unshed antler (3)*
Schmidt 1972
Capreolus capreolus
VL 51\1
D3\4 sup.sin.
Carter 2006
VL 285\7
D2 sup.sin.
Carter 2006
Fox Vulpes vulpes
VL 222\22-23
Dc sup.dext. &
M1 sup.dext. (germ
)H
abermehl 1985
VL 133 x.2
max. sin. (D
4)C
arter, Magnell 2007**
Wild boar
VL 237\5
D3 sup. dext.
Carter, M
agnell 2007** Sus scrofa
VL 282\3
D2 sup.dext.
Carter, M
agnell 2007** V
L 308 x.9
D4 inf.sin.
Carter, M
agnell 2007V
L 242\1M
2 inf.dext.C
arter, Magnell 2007
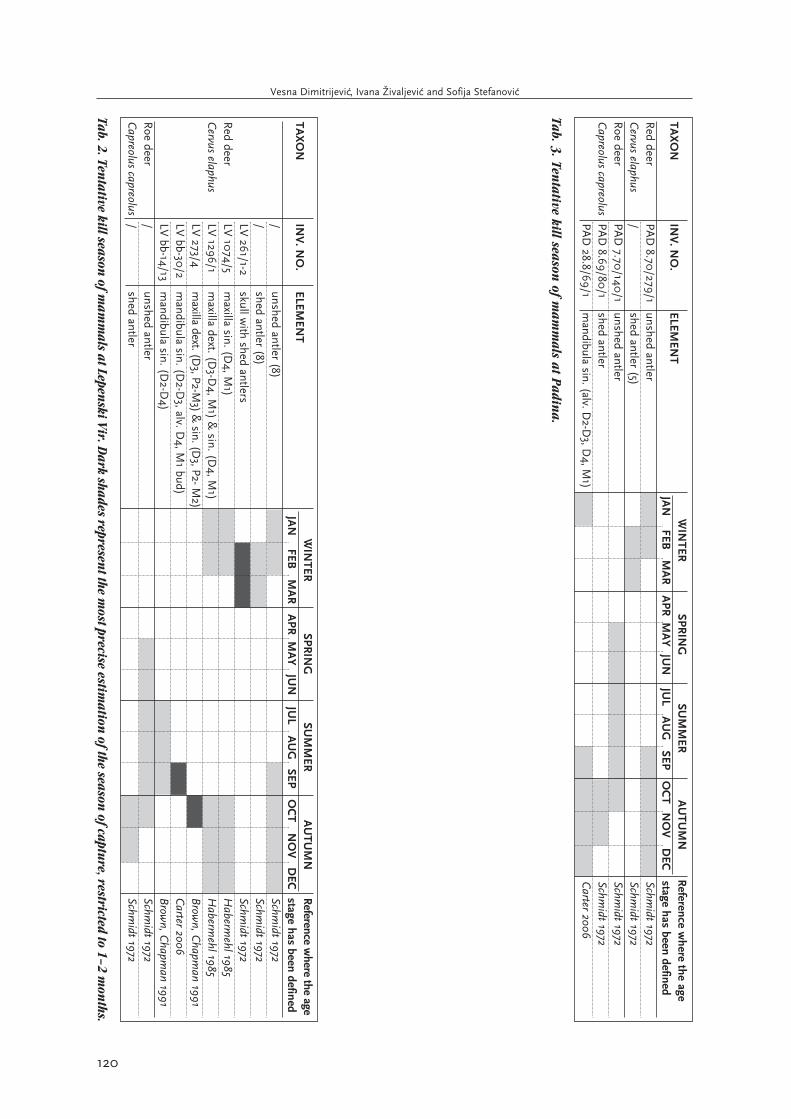
Tab. 1. Tentative kill season
of mam
mals at V
lasac. Dark shades represen
t the most precise estim
ation of the season
of capture, restricted to 1–2 mon
ths. All
elemen
ts used for estimation
s of seasonality origin
ate from the faun
al assemblage collected in
2006–2009, apart from elem
ents m
arked with an
asterisk (*)w
hich were collected in
1970–1971. **The ageing of w
ild boar maxillary teeth assum
ing they follow
the same developm
ent schedule as m
andibular teeth.
Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
120
TAX
ON
INV
. NO
.ELEM
ENT
WIN
TERSPR
ING
SUM
MER
AU
TUM
NReference w
here the ageJA
NFEB
MA
RA
PRM
AY
JUN
JUL
AU
GSEP
OC
TN
OV
DEC
stage has been defined\
unshed antler (8)Schm
idt 1972\
shed antler (8)Schm
idt 1972LV
261\1-2skull w
ith shed antlersSchm
idt 1972R
ed deerLV
1074\5
maxilla sin. (D
4, M1)
Haberm
ehl 1985C
ervus elaphusLV
1296\1m
axilla dext. (D3-D
4, M1) &
sin. (D4, M
1)H
abermehl 1985
LV 273\4
maxilla dext. (D
3, P2-M3) &
sin. (D3, P2- M
2)B
rown, C
hapman 1991
LV bb-30
\2m
andibula sin. (D2-D
3, alv. D4, M
1 bud)C
arter 2006LV
bb-14\13m
andibula sin. (D2-D
4)B
rown, C
hapman 1991
Roe deer
\unshed antler
Schmidt 1972
Capreolus capreolus
\shed antler
Schmidt 1972
Tab. 2. Tentative kill season
of mam
mals at Lepen
ski Vir. D
ark shades represent the m
ost precise estimation
of the season of capture, restricted to 1–2 m
onths.
TAX
ON
INV
. NO
.ELEM
ENT
WIN
TERSPR
ING
SUM
MER
AU
TUM
NR
eference where the age
JAN
FEBM
AR
APR
MA
YJU
NJU
LA
UG
SEPO
CT
NO
VD
ECstage has been defined
Red deer
PAD
8.70\279\1
unshed antlerSchm
idt 1972C
ervus elaphus\
shed antler (5)Schm
idt 1972R
oe deerPA
D 7.70
\140\1
unshed antlerSchm
idt 1972C
apreolus capreolusPA
D 8.69\80
\1 shed antler
Schmidt 1972
PAD
28.8\69\1m
andibula sin. (alv. D2-D
3, D4, M
1)C
arter 2006
Tab. 3. Tentative kill season
of mam
mals at Padin
a.
Becoming sedentary| The seasonality of food resource exploitation in the Mesolithic-Neolithic Danube Gorges
121
TAX
ON
VL 20
06–20
09
LV 1968–1970
PA 1968–1970
WIN
TERSPR
ING
SUM
MER
AU
TUM
NN
ISP%
NISP
%N
ISP%
JAN
FEBM
AR
APR
MA
YJU
NJU
LA
UG
SEPO
CT
NO
VD
ECR
ussian sturgeon Acipenser gueldenstaedtii
90
,154
6,413
0,6
Ship sturgeon Acipenser nudiventris
\\
40
,5\
\Sterlet A
cipenser ruthenus\
\9
1,11
0,0
Stellate sturgeon Acipenser stellatus
50
,17
0,8
160
,7B
eluga Huso huso
270
,383
9,972
3,1B
lack Sea shad Alosa im
maculata
921,1
10
,1\
\C
omm
on carp Cyprinus carpio
803
9,592
10,9
208
8,9B
ream A
bramis bram
a22
0,3
\\
\\
Danube bleak A
lburnus chalcoides2
0,0
\\
\\
Asp A
spius aspius8
0,1
20
,2\
\B
elica Leucaspius delineatus1
0,0
\\
\\
Chub Leuciscus cephalus
\\
30
,4\
\Ide Leuciscus idus
40
,02
0,2
\\
Razor fish Pelecus cultratus
210
,21
0,1
\\
Vyrezub R
utilus frisii7320
86,4257
30,5
1626,9
Pigo Rutilus pigus
20
,01
0,1
\\
Roach R
utilus rutilus1
0,0
\\
\\
Rudd Scardinius erythrophthalm
us7
0,1
\\
\\
Vim
ba bream V
imba vim
ba4
0,0
10
,1\
\W
els catfish Silurus glanis61
0,7
16820
,0180
677,1
Pike Esox lucius44
0,5
10
,1\
\H
uchen Hucho hucho
290
,3154
18,365
2,8B
lack Sea trout Salmo trutta labrax
\\
10
,1\
\Pikepearch Sander lucioperca
80
,11
0,1
\\
TOTA
L8470
100
84210
02343
100
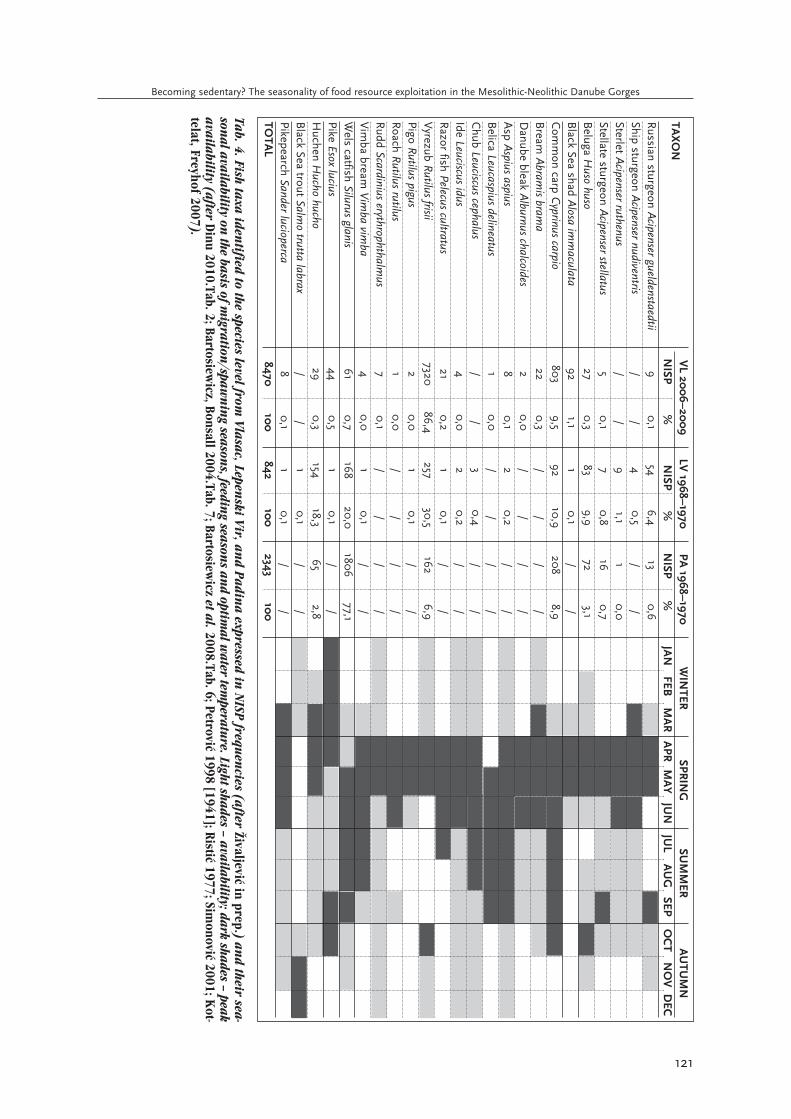
Tab. 4. Fish taxa identified to the species level from
Vlasac, Lepen
ski Vir, an
d Padina expressed in
NISP frequen
cies (after Ωivaljevi≤ in prep.) and their sea-
sonal availability on
the basis of migration
/spawn
ing season
s, feeding season
s and optim
al water tem
perature. Light shades – availability; dark shades – peakavailability (after D
inu 2010.Tab. 2; Bartosiewicz, Bonsall 2004.Tab. 7; Bartosiew
icz et al.2008.Tab. 6; Petrovi≤ 1998 [1941]; Risti≤ 1977; Sim
onovi≤ 2001; Kot-
telat, Freyhof 2007).
Vesna Dimitrijevic, Ivana ?ivaljevic and Sofija Stefanovic
122
NISP
WIN
TERSPR
ING
SUM
MER
AU
TUM
NTA
XO
NV
LASA
CPA
DIN
AL. V
IR1970
–197120
06–20
09
1968–19701968–1970
JAN
FEBM
AR
APR
MA
YJU
NJU
LA
UG
SEPO
CT
NO
VD
ECB
lack-throated diver Gavia arctica
\\
1\
White pelican Pelecanus cf. onocrotalus
3\
1\
Corm
orant Phalacrocorax carbo3
\1
\M
ute swan C
ygnus olor\
\1
\G
reat white egret Egretta alba
2\
\\
Mallard A
nas platyrhynchos2
\\
1Teal A
nas crecca2
\\
\G
rey-lag goose Anser anser
\\
2\
Ferruginous (|) duck Aythya nyroca
(|)1
\\
\Shell-duck Tadorna tadorna
\\
1\
Black kite M
ilvus migrans
5 (4)\
\\
Imperial eagle A
quila heliaca6 (16)
2\
\W
hite-tailed eagle Haliaeëtus albicilla
27 (0)
\8
\Taw
ny owl Strix aluco
2 (1)1
\\
Jay Garrulus glandarius
1\
\\
Magpie Pica pica
1\
\\
Raven C
orvus corax4
12
\R
ook Corvus frugilegus
\\
1\
Fieldfare Turdus pilaris\
2\
\TO
TAL
59 (21)6
181
Tab. 5. Bird taxa iden
tified to the species level from V
lasac (num
bers in paren
thesis represent the revision
of the preserved bird faunal assem
blage analyzed
by Bökönyi 1978, after Dim
itrijevi≤et al. subm
itted), Padina (after Clason 1980), an
d Lepenski Vir (after Bori≤, D
imitrijevi≤ 2006) expressed in
NISP frequen
-cies,an
d their seasonal availability on
the basis of residency an
d migration
patterns (after Puzovi≤ 2000; R
a∏ajski, Kiss 2004; G
ruba≠et al. 2013; Clem
entset al. 2014).