bacterial genetics. classification and taxonomy phylogeny the most recent model for the basic...
Post on 21-Dec-2015
216 views
TRANSCRIPT

Bacterial Genetics

Classification and Taxonomy

Phylogeny• The most recent model for the basic divisions of life is the
“three domain model”, first put forth by Carl Woese in the 1970’s.
• He compared the sequences of 16S ribosomal RNA genes, which are fundamentally important for protein synthesis and found in all known living organisms.
• He discovered that “bacteria” could be divided into 2 very different groups, the Eubacteria (often just called Bacteria) and the Archaea
• The third group is the eukaryotes, organisms in which the DNA is contained within a membrane-bound nucleus.
• Eubacteria and Archaea are the two type of prokaryote, organisms in which the DNA is loose within the cytoplasm and not contained within a nucleus.
• Archaea usually live in extreme environments: very hot, acidic, salty, etc. They use quite different information processing machinery than the bacteria. We are going to mostly ignore them.

Classifying Bacteria
• Classically, bacteria have been characterized by their staining pattern, shape, reaction to oxygen, pH, temperature, and salt optima, and their ability to metabolize various compounds.– good functional classification: what they look like, and
where they live, but often evolutionary relationships are not accurate
• More recent classification schemes are based in 16S ribosomal RNA., which is found in all (known) cells. Also, the percentage of G and C (G+C content) is used for classification.

Gram Stain• A major distinction between groups
of bacteria is based on the Gram stain. In this method, bacteria are treated with the dye “crystal violet”, then washed. Often a second stain, “safranin” is applies to make the unstained bacteria visible.
• Gram stain causes bacteria with a lot of peptidoglycan and very little lipid in their cells walls to stain purple. The presence or absence of peptidoglycan is a fundamental biochemical difference between groups of bacteria
• Another stain, the “acid-fast stain” is used to identify Mycobacteria, such as the tuberculosis agent Mycobacterium tuberculosis.

Bacterial Morphology
• Bacteria only take a few basic shapes, which are found in many different groups. Bacterial cells don’t have internal cytoskeletons, so their shapes can’t be very elaborate.
• Shape: coccus (spheres) and bacillus (rods). Spirillum (spiral) is less common.– note: “bacillus” is a shape, but “Bacillus” or
(better) Bacillus is a taxonomic group, a genus containing such species as Bacillus subtilis, Bacillus anthracis, and Bacillus megaterium. The bacillus shape is NOT limited to the Bacillus genus.
• Aggregation of cells: single cells, pairs (diplo), chains (strepto), clusters (staphylo).
• Thus we have types such as diplococcus (pair of spheres) and streptobacillus (chain of rods).

Relationship to Oxygen
• For more than half of Earth’s history, oxygen wasn’t present in the atmosphere. Many bacteria evolved under anaerobic conditions.
• Classification:– strict aerobes (need oxygen to survive)– microaerobes need oxygen, but at reduced
concentration (such as in cow guts)– strict anaerobes (killed by oxygen)– aerotolerant (don’t use oxygen, but survive it).– facultative anaerobes (use oxygen when it is present,
but live anaerobically when oxygen is absent).

Temperature
• thermophiles have an optimum growth temperature above 50oC
• hyperthermophiles have an optimum growth temperature above 80oC. Many of these are Archaea, not Bacteria
• psychrophiles (cryophiles) have an optimum growth temperature below 15oC
• mesophiles are those with optima between 15oC and 50oC.

Metabolic Classification• All living organisms need to obtain energy from the environment, and they need to
obtain or make reduced, organic carbon compounds.– CO2 (carbon dioxide) is the most oxidized form of carbon, and it is not considered “organic”
• Energy comes from 2 sources, sunlight or chemical bonds– an organism that uses light for energy is a phototroph
– an organism that uses chemical bonds for energy is a chemotroph
– chemotrophs are sub-divided:
• if the chemical bonds used for energy come from organic molecules, it is a chemoorganotroph.
• If inorganic compounds are used, it is a chemolithotroph (litho = rock)
• Organic carbon compounds are often obtained from other organisms: heterotroph.
• Or, organic compounds can be made by reducing carbon dioxide: autotroph.
• Humans are thus chemoorganoheterotrophs. Plants are photoautotrophs. Various bacteria are found in all 6 roles.

Tree of Life: Bacterial Phylahttp://tolweb.org/tree?group=Eubacteria&contgroup=Life_on_Earth

Bacterial Structure
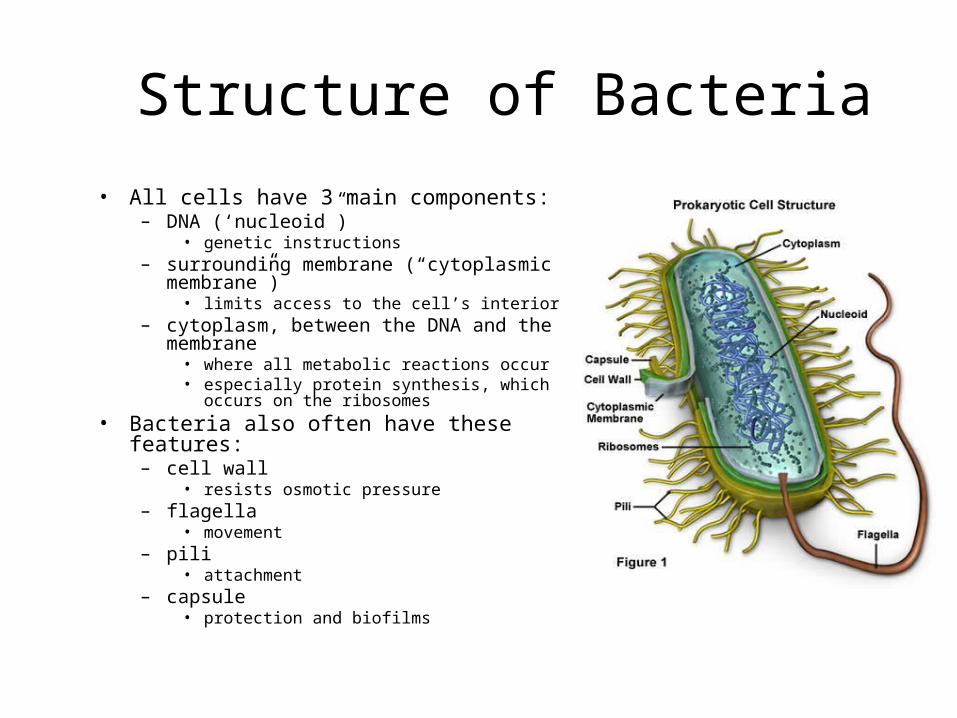
Structure of Bacteria
• All cells have 3 main components:– DNA (‘nucleoid”)
• genetic instructions– surrounding membrane (“cytoplasmic
membrane”)• limits access to the cell’s interior
– cytoplasm, between the DNA and the membrane
• where all metabolic reactions occur• especially protein synthesis, which occurs on
the ribosomes
• Bacteria also often have these features:– cell wall
• resists osmotic pressure– flagella
• movement– pili
• attachment– capsule
• protection and biofilms

Cell Envelope
• The cell envelope is all the layers from the cell membrane outward, including the cell wall, the periplasmic space, the outer membrane, and the capsule.
– All free-living bacteria have a cell wall
– periplasmic space and outer membrane are found in Gram-negatives
– the capsule is only found in some strains

Cell Membrane
• The cell membrane (often called the plasma membrane) is composed of 2 layers of phospholipids.
• Phospholipids have polar heads and non-polar tails.
– “Polar” implies that the heads are hydrophilic: they like to stay in an aqueous environment: facing the outside world and the inside of the cell.
– “non-polar” means that the tails are hydrophobic: they want to be away from water, in an oily environment. The tails are in the center of the membrane
• A pure phospholipid membrane only allows water, gasses, and a few small molecules to move freely through it.

Membrane Proteins• Proteins float in the membrane like ships
on the surface of the sea: the fluid-mosaic model.
• Peripheral membrane proteins are bound to one surface of the membrane.
– Some attached to the cell membrane by a fatty acid covalently attached to one of the protein’s amino acids
– Others are attached by stretches of hydrophobic amino acids of the protein’s surface
• Integral membrane proteins are embedded in the membrane by one or more stretches of hydrophobic amino acids. Many of these proteins transport molecules in and out of the cell. The transport proteins are very selective: each type of molecule needs its own transporter.

Transport Across the Cell Membrane
• Basic rule: things spontaneously move from high concentration to low concentration (downhill). This process is called diffusion.
– Getting many molecules into the cell is simply a matter of opening up a protein channel of the proper size and shape. The molecules then move into the cell by diffusing down the concentration gradient. Passive transport, or facilitated diffusion.
• To get things to move from low to high (uphill), you need to add energy: the molecules must be pumped into the cell. Pumps are driven by ATP energy. Active transport.

More Membrane Transport• Often2 molecules are transported
together, with one moving by diffusion down its concentration gradient and the other carried along up its concentration gradient.
– If the two molecules move in the same direction, the protein channel is a symporter. See the diagram of the sodium-glucose symport mechanism
– If the two molecules move in opposite directions, the channel is an antiporter.
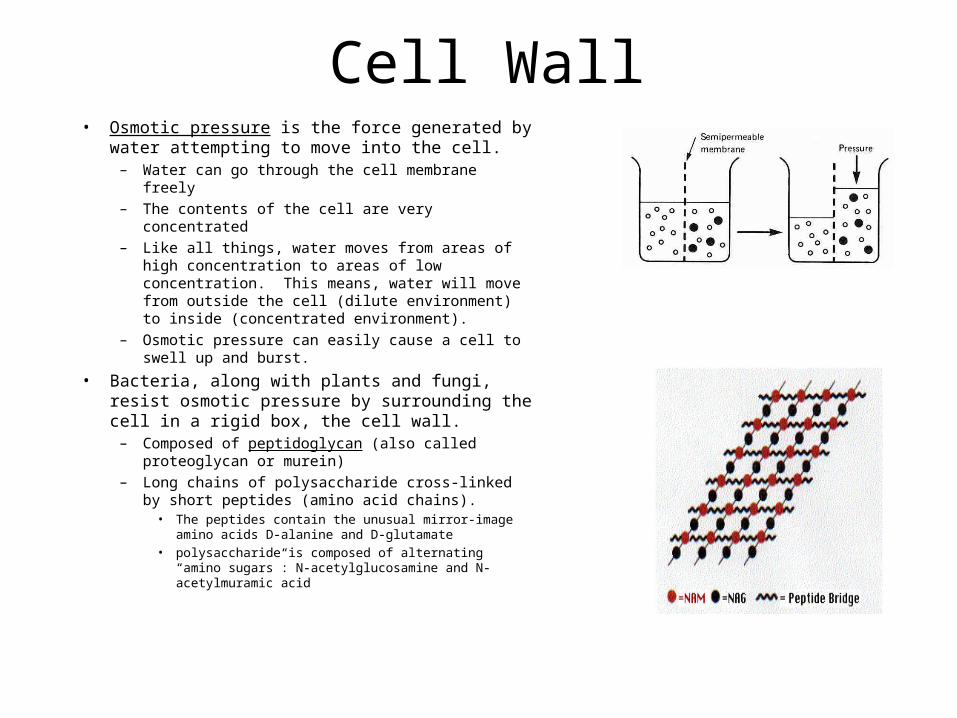
Cell Wall• Osmotic pressure is the force generated by water
attempting to move into the cell.– Water can go through the cell membrane freely
– The contents of the cell are very concentrated
– Like all things, water moves from areas of high concentration to areas of low concentration. This means, water will move from outside the cell (dilute environment) to inside (concentrated environment).
– Osmotic pressure can easily cause a cell to swell up and burst.
• Bacteria, along with plants and fungi, resist osmotic pressure by surrounding the cell in a rigid box, the cell wall.
– Composed of peptidoglycan (also called proteoglycan or murein)
– Long chains of polysaccharide cross-linked by short peptides (amino acid chains).
• The peptides contain the unusual mirror-image amino acids D-alanine and D-glutamate
• polysaccharide is composed of alternating “amino sugars”: N-acetylglucosamine and N-acetylmuramic acid

More Cell Wall• Gram-positive vs Gram-negative are defined by the
structure of the cell wall– the Gram stain binds to peptidoglycan
• Gram-positive: many layers of peptidoglycan, which is anchored to the cell membrane by teichoic acid.
• Gram-negative: 1-2 layers of peptidoglycan = thin– The periplasmic space is between the cell membrane
and the cell wall. It contains enzymes and other proteins, such as chemoreceptors for sensing the environment.
– Outside the peptidglycan layer is the “outer membrane”. It is pierced by porins: protein channels, and its out surface is covered with lipopolysaccharides (sugars linked to membrane lipids), which are often antigenic and or toxic.

Capsule
Some bacteria (often pathogens) are surrounded by a thick polysaccharide capsule. This is a loose jelly-like or mucus-like layer. It helps prevent immune system cells from reaching the bacteria, and it forms part of biofilms.

Membrane Structures• Pili (singular = pilus) are hairs projecting from the
surface. They are composed of pilin protein. There are several types:
– DNA can be transferred between bacteria by conjugation, which is initiated when sex pili on the donor cell attach to and draw in the recipient cell.
– Fimbriae (singular = fimbria) are pili used to attach the bacteria to target cells ( in infection) or to surfaces, where they form a biofilm.
• Flagella are long hairs used to propel the cells. They are composed of flagellin protein.
– At the base of each flagellum is a motor embedded in the membrane and cell wall. It turns in a rotary motion, driven by proton-motive force (the flow of protons i.e. H+ ions across the cell membrane).
– The suffix “-trichous” is used to describe the placement of flagella: e.g. lophotrichous = several flagella all clustered at one end.

Chemotaxis
• The flagellar motor is reversible:– Counterclockwise rotation: bacterium
moves in a straight line
– clockwise rotation: bacterium tumbles randomly
– the motor periodically reverses, causing a random change in direction: bacteria move in a random walk.
• chemotaxis: bacteria move toward the source of nutrients by swimming up the chemical gradient. Or, away from a toxin.
– When moving up the gradient, bacteria rarely tumble, but when moving across it, or in the opposite direction, tumbling is frequent.
– This produces a net motion in the proper direction

Spores• Some bacteria can form very tough spores, which are
metabolically inactive and can survive a long time under very harsh conditions.
– Allegedly, some bacterial spores that were embedded in amber or salt deposits for 25 million years have been revived. These experiments are viewed skeptically by many scientists.
– Panspermia: the idea that life got started on Earth due to bacterial spores that drifted in from another solar system. (However, it still had to start somewhere!).
• “Extraordinary claims demand extraordinary proof”
• Spores can also survive very high or low temperatures and high UV radiation for extended periods. This makes them difficult to kill during sterilization.
– Anthrax• Spores are produced only by a few genera in the
Firmicutes: – Bacillus species including anthracis (anthrax) and
cereus (endotoxin causes ~5% of food poisoning)– Clostridium species including tetani (tetanus),
perfringens (gangrene), and botulinum (botulism: food poisoning from improperly canned food)

Metabolism

Oxidation-Reduction• Two truisms:
– Chemistry is the study of the movement of electrons between atoms
– Life is applied chemistry
• Oxidation: a molecule loses an electron– LEO: Lose Electron Oxidation
• Reduction: a molecule gains an electron– GER: Gain Electron Reduction
• In living cells, there are no free electrons: every time an electron leaves one molecule, it goes to another one.
– Thus, all oxidation reactions are coupled with reduction reactions: one compound is oxidized while the other is reduced. “Redox” reactions
• Electrons are often accompanied by H+ ions. Thus, FAD is the oxidized form, and FADH2 is the reduced form: it has 2 more electrons (and H’s) than FAD.
– For this reason, enzymes that perform oxidations are usually called dehydrogenases.

Redox Potential
• Redox potential is a measure of the affinity of compounds for electrons. The more positive a compound’s redox potential is, the greater its tendency to acquire electrons.– Redox potential is measured in millivolts (mV), relative to hydrogen at 1
atm pressure. Compounds are at 1 M concentration.• H2 2 H+ + 2 e- • The idea is, if your compound was mixed with hydrogen gas, would electrons
flow from your compound to the hydrogen (compound has a negative redox potenitial), or from the hydrogen to your compound (compound has a positive redox potenital)?
– Redox potential is affected by the concentration of the reactants and also by the redox potential of the environment
• When electrons in compounds with lower (more negative) redox potentials are moved to compounds with higher potentials, energy is released. Organisms capture this energy to live on.

Some Redox Reactions Used in Bacteria

Some Lithotrophic Reactions

Fermentations•Fermentation is defined as a process where organic molecules are both the electrondonor and the electron acceptor.
•Since the complete oxidation of organic molecules ends at carbon dioxide, fermentations are by definition incomplete oxidations: there is always some potential energy left in the products of a fermentation.
•And thus the fermentation products excreted by one species are often used as food sources for another species.
•The best known fermentations involve the products of glycolysis.
•Glucose is oxidized to pyruvate, but the electrons from glucose are used to convert NAD+ to NADH. •To get rid of these electrons, NADH is used to reduce pyruvate to lactate (as in anaerobic human muscle) or to ethanol (as in yeast). Both of these pathways are used in various bacteria.

Other Fermentations

Aerobic Respiration
• The most efficient way of producing energy is by oxidizing organic compounds to carbon dioxide = respiration. This is the process used by most eukaryotes and aerobic bacteria.

Energy Generation
• Energy in the cell is generated and used in the form of ATP.
• Two basic way s of generating ATP:– substrate-level
phosphorylation. The simplest form: transfering a phosphate group from another molecule to ADP, creating ATP.
– chemiosmotic: generation of a proton (H+) gradient across a membrane. This gradient is called “proton-motive force”.

Chemiosmotic Theory
•The same basic process in the mitochondria as in many bacteria.•High energy electrons from an electron donor are used to pump H+ ions out of the cell, into the periplasmic space
•This drains energy from the electrons•Electron transport
•There are thus more H+ ions outside than inside: the pH outside is lower than inside.
•The H+ ions are then allowed back into the cell by passing them through the ATP synthase protein, which uses the energy of the H+ ions flowing down the gradient to attach phosphate (Pi) to ADP, creating ATP.
•the gradient is both chemical: more H+ outside than inside, and electrical: more + charge outside than inside

Carbon Assimilation• Heterotrophic organisms obtain
organic carbon compounds from pre-existing organic molecules.– often the same molecules they are
using for energy: glucose for example.
• Autotrophs “fix” carbon dioxide into organic carbon.– 4 pathways:1. Calvin cycle (ribulose bis-phosphate
pathway. Used in plants and many bacteria: the most common pathway
2. Reductive TCA cycle: run the Krebs cycle backwards
3. Reductive acetyl CoA pathway, which requires hydrogen gas and produces carbon monoxide as an intermediate.
4. 3-hydroxypropiuonate cycle. Seems to mostly be in Archaea

Carbon Assimilation Pathways

Nitrogen Assimilation• Most of the nitrogen on Earth is
nitrogen gas, N2, which is strongly held together by a triple bond.
– nitrogen fixation, converting nitrogen to ammonia, is very energy-intensive and carried out by a small group of bacteria, including some Clostridium.
– some nitrogen is also fixed by lightning.
• Nitrogen’s major use in the cell is as a component of amino acids, in the ammonium form: -NH2
– Many organisms get their nitrogen from organic nitrogen compounds
– some organisms perform ammonification, which means splitting the amino group off organic compounds, releasing ammonium ions.
• Nitrogen is also found as nitrate. Most bacteria can reduce nitrate (NO3
-) to nitrite (NO2
-) and then to ammonia: assimilatory nitrate reduction.
– nitrate is also degraded back into nitrogen gas by other bacteria: denitrification.
– the reverse process, converting ammonia into nitrate and nitrite, is used as an energy source by some lithotrophs. It is called nitrification.

More Nitrogen Assimilation
• When nitrogen is taken into the cell in the form of ammonium ions, it is attached to glutamate, forming glutamine, using the enzyme glutamine synthetase.
• Alpha-ketoglutarate, glutamate, and glutamine can all be interconverted.– this is the source of the amino group
of amino acids and amino sugars.

Assimilation of Other Elements
• Cells use several other elements: C, H, O, and N are the major ones
– also covalently bound: P, S, Se– ions: Na, K, Mg, Ca, Cl– trace elements (mostly as enzyme
co-factors): Fe, Mn, Co, Cu, Ni, Zn, others....
• Sulfur can be incorporated from organic sources, but it is often taken into the cell as sulfate (SO4
-
2). Getting into the cell requires attaching it to the ATP derivative APS, after which it is reduced to sulfide (S-2) and then attached to serine, converting it to cysteine.
• phosphate (PO4-3) is generally
found in the same form as it is used. It just needs to be transported into the cell.

Intermediary Metabolism
• Lots of interconversions. • it is necessary to make:
– amino acids (proteins)– nucleotides (DNA, RNA, and ATP)– sugars (part of nucleotides, food, structure)– lipids: food storage and membrane– several co-factors for enzymes: biotin, cytochromes, panthothenic
acid, NAD, riboflavin, cobalamin, ubiquinone, etc.
• The central metabolic pathways of glycolysis and the Krebs cycle have several side branches that feed these biosynthetic pathways.


Enzyme Nomenclature• Every chemical interconversion requires an enzyme to catalyze it.• Nearly all enzyme names end in –ase• Enzyme functions: which reactants are converted to which products
– Across many species, the enzymes that perform a specific function are usually evolutionarily related. However, this isn’t necessarily true. There are cases of two entirely different enzymes evolving similar functions.
• Enzyme functions are given unique numbers by the Enzyme Commission.– E.C. numbers are four integers separated by dots. The left-most number is the least specific– For example, the tripeptide aminopeptidases have the code "EC 3.4.11.4", whose
components indicate the following groups of enzymes:• EC 3 enzymes are hydrolases (enzymes that use water to break up some other molecule) • EC 3.4 are hydrolases that act on peptide bonds • EC 3.4.11 are those hydrolases that cleave off the amino-terminal amino acid from a polypeptide • EC 3.4.11.4 are those that cleave off the amino-terminal end from a tripeptide
• Top level E.C. numbers: – E.C. 1: oxidoreductases (often dehydrogenases): electron transfer– E.C. 2: transferases: transfer of functional groups (e.g. phosphate) between molecules.– E.C. 3: hydrolases: splitting a molecule by adding water to a bond.– E.C. 4: lyases: non-hydrolytic addition or removal of groups from a molecule– E.C. 5: isomerases: rearrangements of atoms within a molecule– E.C. 6: ligases: joining two molecules using energy from ATP

Detailed Pathways
• For many compounds, there can be more than one way to produce it. Some organisms have more than one pathway to a given compound, and sometimes different organisms produce it by different mechanisms.
• KEGG (Kyoto Encyclopedia of Genes and Genomes) has a comprehensive set of pathway maps, with individual species differences noted.– KEGG also has information about individual enzymes and ligands.
You can get there by clicking the elements of the map.– http://www.genome.ad.jp/kegg/pathway.html


Genes and Gene Expression

Polypeptides and Proteins
• All of the work in the cell: energy generation, synthesis of new components, response to environmental stimuli, etc., is performed by proteins.
• Proteins are primarily composed of polypeptides: linear chains of amino acids.
– each polypeptide is called a “subunit”. • each gene produces one type of polypeptide
– some proteins are composed of a single polypeptide, while others have 2 or more (up to maybe 20 in very complex proteins) subunits
• Proteins, especially enzymes, also often have co-factors bound to them.
– some co-factors are just metal ions such as Zn+2
– others are more complex: most of the human “vitamins” are enzyme co-factors
• in many species of bacteria, co-factors are synthesized by complex pathways.
• Some proteins have sugars, lipids, or other small molecules attached to them.

Gene Expression
• Most genes code for polypeptides– maybe 5% of genes produce
special RNA molecules only
• The general process of gene expression is to transcribe an RNA copy of the gene, called messenger RNA (mRNA). The mRNA is then translated into the polypeptide by the action of ribosomes.– this is sometimes referred to as
the Central Dogma of Molecular Biology

Genes on the Chromosome• The bacterial chromosome is a very long
molecule of DNA. – The genes are short regions of this molecule.– A typical bacterial genome has 2000-5000
genes.• the position of each gene on the
chromosome is the same in all members of a species
– but not necessarily conserved across species lines
• there is some local clustering of genes in the same biochemical pathway, but in general position on the chromosome cannot be correlated with gene function.
• Genes have a particular orientation on the DNA strand: they are written with the 5’ end on the left and the 3’ end on the right.
– However, either strand of the DNA can encode a gene. The result of this is that genes on one strand face in one direction, and genes one the other strand face the other direction

Transcription• Gene expression begins when the enzyme RNA
polymerase binds to the promoter region just upstream from the gene.
– the promoter consists of 2 segments of important nucleotides, with spacers (whose sequence doesn’t matter) in between.
– positioned -10 and -35 bp upstream from RNA start– it’s a consensus sequence: variations on a common theme.
• Transcription is said to start at the 5’ end of the mRNA and end at the 3’ end. This refers to the free ends of the ribose sugar in the RNA molecule.
• The RNA polymerase then moves down the DNA, using one DNA strand as a template to synthesize an RNA copy of the gene.
– the raw materials are “NTPs”: nucleoside triphosphates. The energy needed to do the synthesis come from removing the 2 terminal phosphate groups, the same process as in using ATP energy.
• Transcription ends at a terminator sequence. The RNA polymerase falls off the DNA, releasing the new mRNA.
• In eukaryotes, the mRNA is processed by splicing out introns and protecting the ends. These events do not occur in prokaryotes.

Transcription Control• The key event is binding of RNA polymerase to the
promoter. It is affected by several factors.– promoters vary slightly in sequence, and these
variations affect the strength of binding.– RNA polymerase has a subunit called sigma. There
are several different sigma factors in the cell, each of which is specific to a different class of promoter. This provides large-scale gene regulation.
– genes are controlled individually by the binding of regulatory proteins, called transcription factors, to regions near the promoter.
• Some transcription factors block transcription and others encourage it
• Some transcription factors regulate whole groups of genes scattered throughout the chromosome: a regulon
• Lac operon model (Jacob and Monod). – Gene used to convert the sugar lactose into glucose,
which is used as food. – The repressor protein (transcription factor) binds to
DNA (the operator) near the promoter and physically blocks transcription.
– when lactose is present, the repressor binds to it, changes its conformation, and falls off the DNA. This allows RNA polymerase to bind and transcription to proceed.

Transcription Termination
• Two basic types: rho-dependent and rho-independent.– rho is a protein
• Rho-dependent termination involves rho protein binding to the RNA and moving along it until it catches up with the RNA polymerase and knocks it off the DNA
• Rho-independent termination involves the newly synthesized RNA folding up into a hairpin loop, due to complementary bases. This sudden folding knocks the RNA polymerase off the DNA.
– formation of stem-loops in RNA also affects transcription initiation in some genes

mRNA, Translation, Operons
•Translation is the process of converting the information on messenger RN.A into protein.
•Note that not all of the mRNA is translated: there is an untranslated region of variable length at both ends, called 5’-UTR and 3’-UTR
•In bacteria, some adjacent genes with related functions are transcribed onto the same mRNA. This is called an operon.
•the proteins are translated individually
•this allows a fixed ratio of the proteins to be
produced.

Genetic Code• There are only 4 bases in DNA and RNA, but there
are 20 different amino acids that go into proteins. How can DNA code for the amino acid sequence of a protein?
• Each amino acid is coded for by a group of 3 bases, a codon. 3 bases of DNA or RNA = 1 codon.
– Since there are 4 bases and 3 positions in each codon, there are 4 x 4 x 4 = 64 possible codons.
– This is far more than is necessary, so most amino acids use more than 1 codon.
• 3 of the 64 codons are used as STOP signals; they are found at the end of every gene and mark the end of the protein.
• In bacteria, translation usually begins at an ATG codon, but GTG and TTG are also common start codons.
– and, a few others are occasionally used• In all cases, the first amino acid is N-formyl
methionine, a derivative of the normal methionine used within the protein.
– note that “start” codons are also used within the protein
– this makes is difficult to be sure where the protein actually starts

Transfer RNA• Transfer RNA molecules act as adapters between the
codons on messenger RNA and the amino acids. Transfer RNA is the physical manifestation of the genetic code.
• Each transfer RNA molecule is twisted into a knot that has 2 ends.
• At one end is the “anticodon”, 3 RNA bases that matches the 3 bases of the codon. This is the end that attaches to messenger RNA.
• At the other end is an attachment site for the proper amino acid.
• A special group of enzymes (aminoacyl tRNA synthetases, which are highly conserved in evolution) pairs up the proper transfer RNA molecules with their corresponding amino acids.
• Transfer RNA brings the amino acids to the ribosomes, which are RNA/protein hybrids that move along the messenger RNA, translating the codons into the amino acid sequence of the polypeptide.

Translation
• Three main players here: messenger RNA, the ribosome, and the transfer RNAs with attached amino acids.
• First step: initiation. The messenger RNA binds to a ribosome, and the transfer RNA corresponding to the START codon binds to this complex. Ribosomes are composed of 2 subunits (large and small), which come together when the messenger RNA attaches during the initiation process.
– there are also several protein “initiation factors” that assist in this process

More Translation• Step 2 is elongation: the ribosome
moves down the messenger RNA, adding new amino acids to the growing polypeptide chain.
• The ribosome has 2 sites for binding transfer RNA. The first RNA with its attached amino acid binds to the first site, and then the transfer RNA corresponding to the second codon bind to the second site.
• The ribosome then removes the amino acid from the first transfer RNA and attaches it to the second amino acid.
• At this point, the first transfer RNA is empty: no attached amino acid, and the second transfer RNA has a chain of 2 amino acids attached to it.

Translation, part 3
• The ribosome then slides down the messenger RNA 1 codon (3 bases).
• The first transfer RNA is pushed off, and the second transfer RNA, with 2 attached amino acids, moves to the first position on the ribosome.

Translation, part 4
• The elongation cycle repeats as the ribosome moves down the messenger RNA, translating it one codon and one amino acid at a time.
• Repeat until a STOP codon is reached.

Translation, end
• The final step in translation is termination. When the ribosome reaches a STOP codon, there is no corresponding transfer RNA.
• Instead, a small protein called a “release factor” attaches to the stop codon.
• The release factor causes the whole complex to fall apart: messenger RNA, the two ribosome subunits, the new polypeptide.
• The messenger RNA can be translated many times, to produce many protein copies.

Post-translation
• The new polypeptide is now floating loose in the cytoplasm. It might also be inserted into a membrane, if the ribosome it was translated on was attached to the membrane by a special RNA/protein hybrid molecule.
• Polypeptides fold spontaneously into their active configuration, and they spontaneously join with other polypeptides to form the final proteins.
• Sometimes other molecules are also attached to the polypeptides: sugars, lipids, phosphates, etc. All of these have special purposes for protein function.