b - compartiments et tansport ii 26dec
DESCRIPTION
ÂTRANSCRIPT

Compartiments et transport intracellulaires
A- Compartiments intracellulaires et tri des protéines
B-Transport vésiculaire dans les voies de sécrétion et d’endocytose







Réf 1: ALBERTS (B) ,



RETCULUM ENDOPLAIQUE
CYTOSOL
MITOCHONDRIES
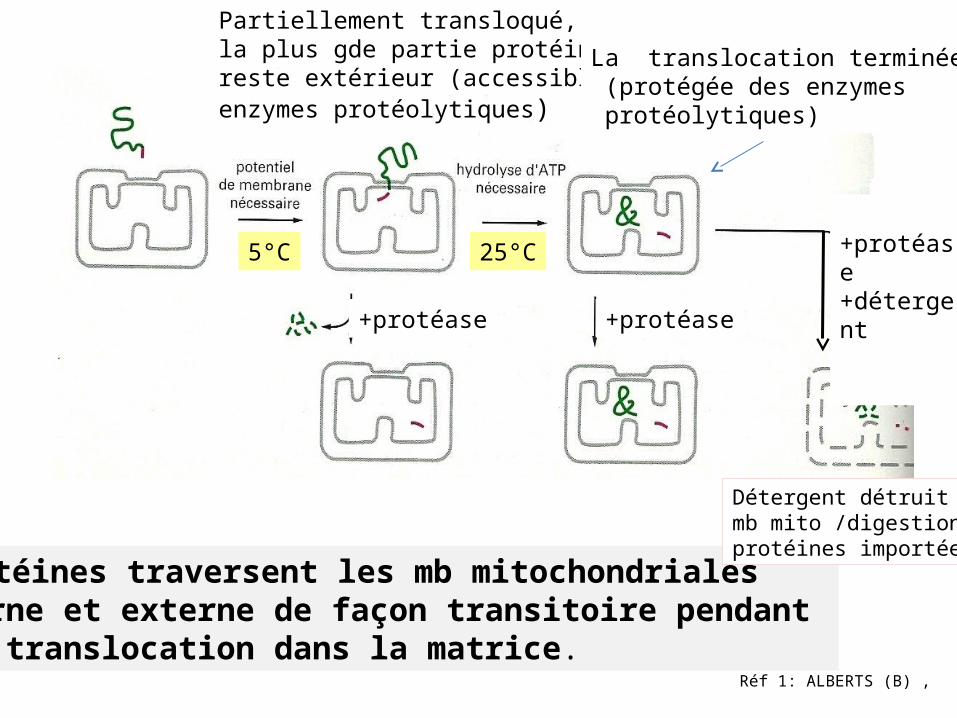
Transport des protéines vers les mitochondries

1-Fixation protéine sur récepteur spécifique
2-Translocation à travers mb externe
3 -Translocation à travers mb interne vers la matrice. - translocation dans la bicouche de la mb interne.

Réf 1: ALBERTS (B) ,
Peptide signal pour l’importationdes protéines mitochondriales .La cytochrome oxydase, un gros complexe multiprotéique dans mb interne mito.A. 12 premiers aa du précurseur de cette enzyme qui constituent un peptide signal pour l’importation de cette sous unité dans mito.B. Un peptide signal complet repliéen hélice α avec 3,6 résidus /tour. Les séquences mito des peptidessignal presque toujours capables de former une telle hélice amphiphile qui serait reconnue par un récepteurprotéique spécifique à la surface mito.Résidus chargés
+ en rouge groupéssur une face hélice
Résidus non polaires en vertface opposée
Réf 1: ALBERTS (B) ,
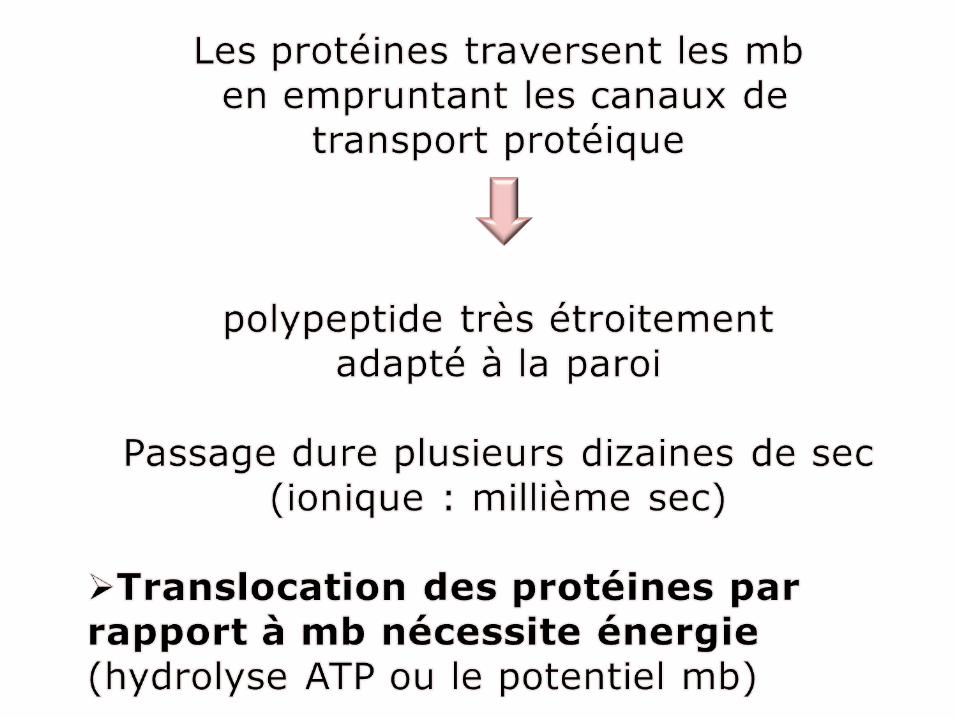
Protéines traversent les mb mitochondriales interne et externe de façon transitoire pendant leur translocation dans la matrice.
Partiellement transloqué, la plus gde partie protéinereste extérieur (accessibleenzymes protéolytiques)
La translocation terminée (protégée des enzymes protéolytiques)
+ protéase+ détergent
Détergent détruit mb mito /digestion protéines importées
5°C 25°C
+protéase +protéase
+protéase+détergent

Cette expérience montre que le processus d’importation a lieu en 2 étapes:
1. Pénétration , par l’intermédiaire du gradient électrochimique du peptide signal et des séquences avoisinantes à travers les 2 mb mito.
2. Déplacement du reste de la chaine polypeptidique vers la matrice qui nécessitel’hydrolyse de l’ATP et T° physiologique.
Récepteur
Peptide signal
1.Reconnaissance
2. Insertion dans mb couplée à gradient électrochimique
3. Translocation dans la matrice nécessitant de l’ATP Peptide
signal
4..
Importation des protéines par les mitochondries
Protéine mito mature
Précurseurprotéique
Réf 1: ALBERTS (B) ,

Réf 1: ALBERTS (B) ,
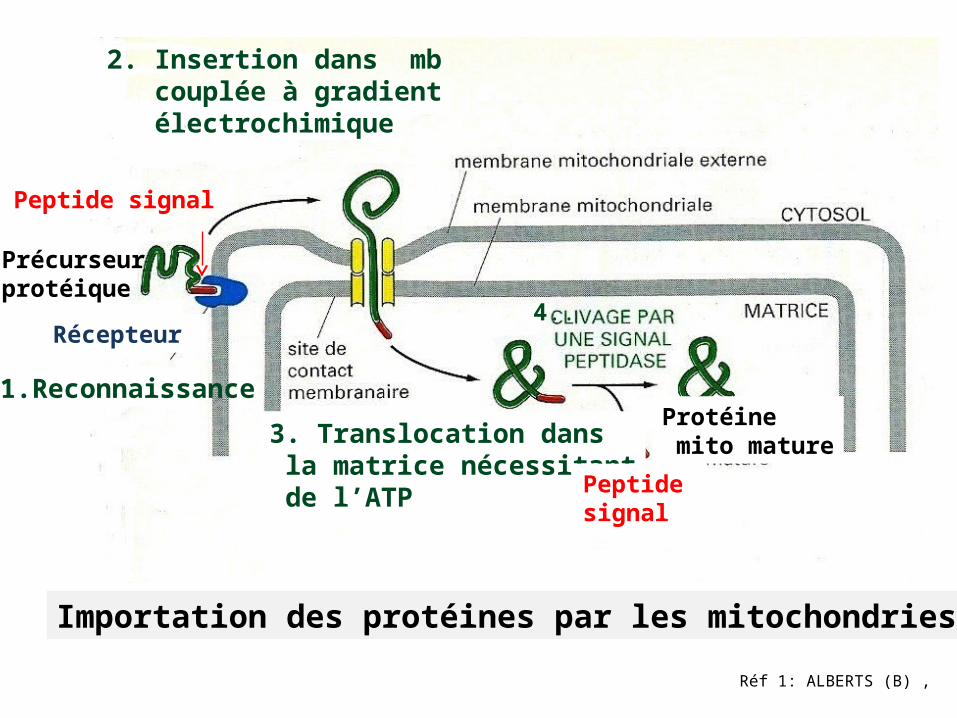
L’importation des protéines dans la matrice mitochondriale nécessite la présence de protéines Hsp70 des 2 côtés de la mb mitochondriale.

Importation protéines depuis le cytosol vers l’espace intermb ou la mb interne
des mitochondries
Quelques protéines se déplacent du cytosol vers
espace intermb -2 peptides signal
(rouge)/(orange)-2 événements de translocation
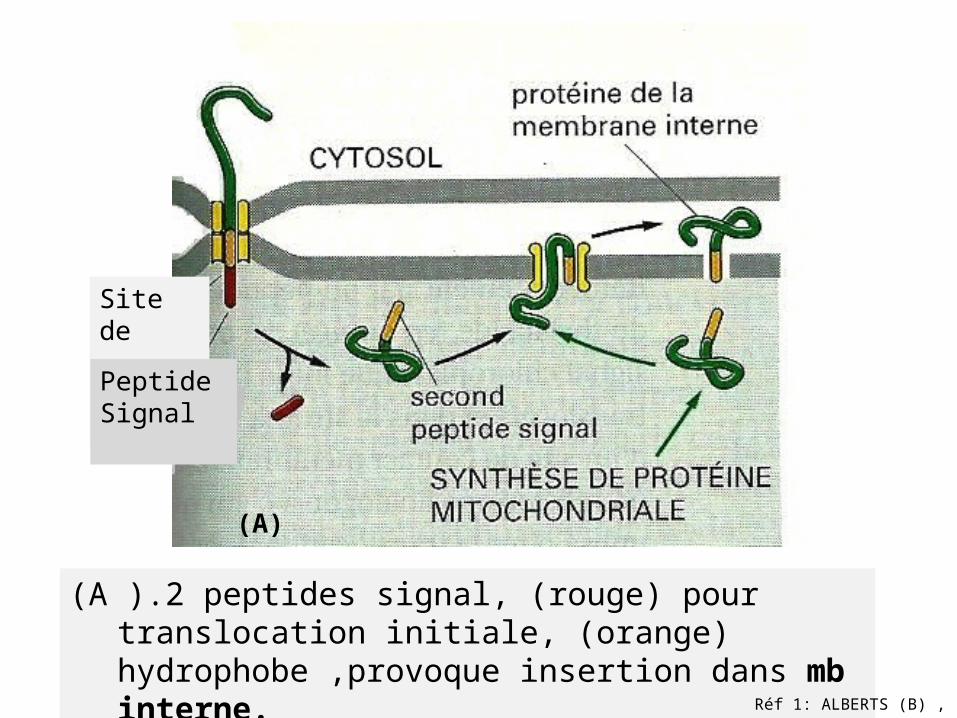
(A ).2 peptides signal, (rouge) pour translocation initiale, (orange) hydrophobe ,provoque insertion dans mb interne.
Site de clivage
Peptide Signal
(A)
Réf 1: ALBERTS (B) ,

(B) .Voie de terminaison de transfert . Après clivage du peptide signal (rouge) ,arrêt de translocation de protéine /mb interne, le reste de la protéines propulsé
dans l’espace intermb.Réf 1: ALBERTS (B) ,
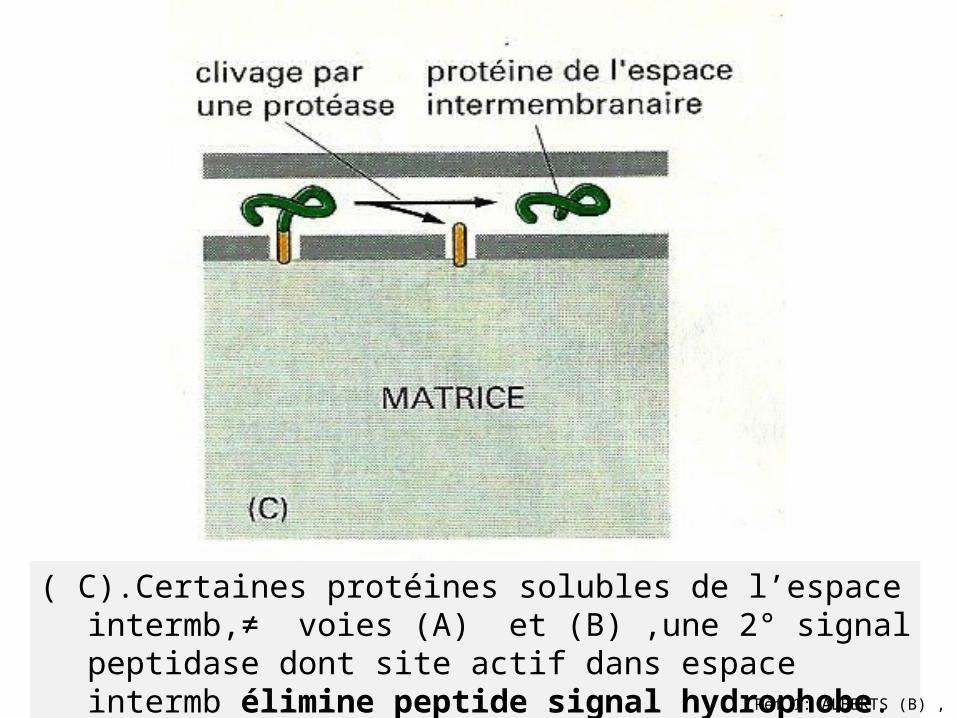
( C).Certaines protéines solubles de l’espace intermb,≠ voies (A) et (B) ,une 2° signal peptidase dont site actif dans espace intermb élimine peptide signal hydrophobe.
Réf 1: ALBERTS (B) ,

*TOM: translocase de la mb externe mito*TIM: ‘’ interne ‘’*N/C :extrémités terminale peptide
Eléments d’importation mitochondriale.
A- Image de microscopie électronique d’une coupe mince .B- Schéma de la machinerie d’importation avec le complexe *TOM de mb externe et *TIM de mb interne.C- reconstitution de tridimensionnelle du cœur du complexe TOM de la translocase de mb externe.D- Structure atomique d’une séquence signal liée à un domaine hydrophobe de TOM 20 Charges : - (rouge) et + (bleu)
Séquence signal: 2 tours α
Réf:14 Pollard (T-D)

Cytoplasme
A
Mb externe
Mb interne
Espace intermb
Matrice
Importation des protéines de la matrice mito. La barre blanche dans le pore d’une translocase indique sa fermeture.
Barre blanche
Réf:14 Pollard (T-D)

A. Une séquence interne de ciblage lie polypeptide à Tom70 qui le dirige vers canal Tom 40.
B. Dans espace intermb ,polypeptide capté par
Tim9/10 et Tim8/13 et dirigé vers complexe de translocation Tim 22/54 qui importe vers la matrice.
C. Tim 22/54 en association avec potentiel transmb ,favorise l’insertion des 6 hélices dans bicouche mb interne

RETCULUM ENDOPLASMIQUE
CYTOSOL
PLASTES

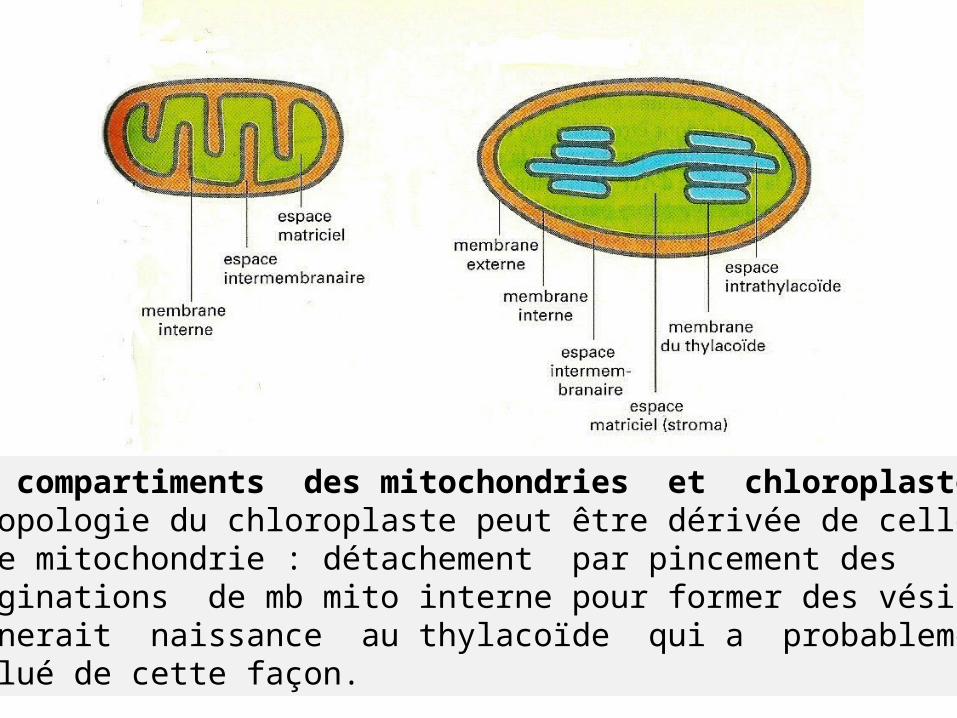
Sous compartiments des mitochondries et chloroplastes.La topologie du chloroplaste peut être dérivée de celle d’une mitochondrie : détachement par pincement des invaginations de mb mito interne pour former des vésicules, donnerait naissance au thylacoïde qui a probablement évolué de cette façon.
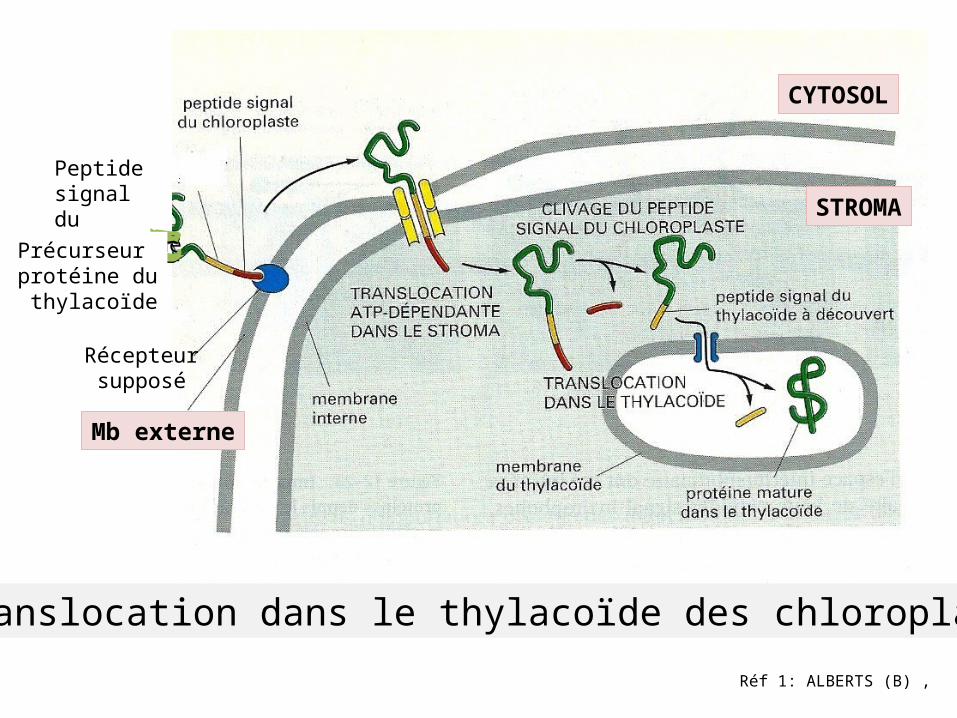
Réf 1: ALBERTS (B) ,
Peptide signal du thylacoïde
Précurseur protéine du thylacoïde
Récepteur supposé
Mb externe
Translocation dans le thylacoïde des chloroplastes
STROMA
CYTOSOL



Voie d’importation des protéines du chloroplaste par les complexes TOC et TIC
Réf:14 Pollard (T-D)

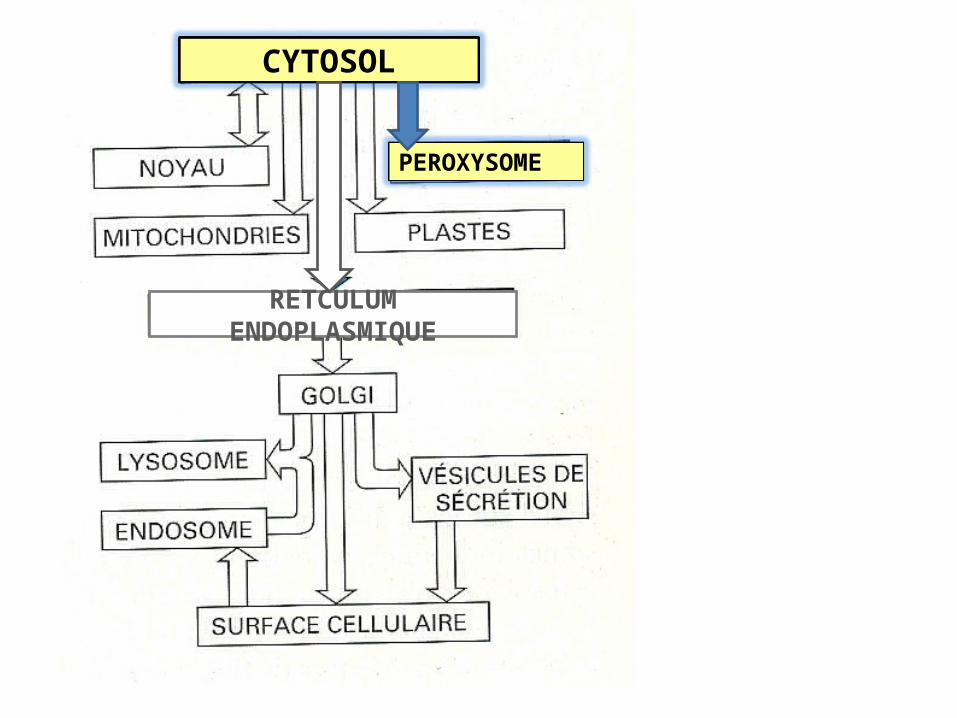
RETCULUM ENDOPLASMIQUE
CYTOSOL
PEROXYSOME

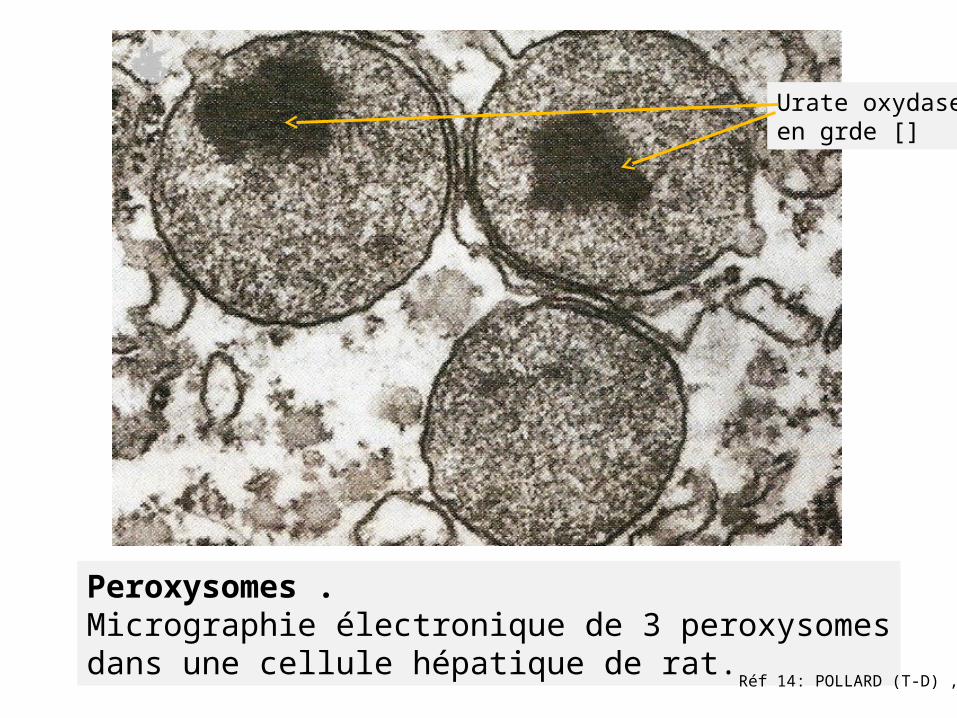
Peroxysomes . Micrographie électronique de 3 peroxysomesdans une cellule hépatique de rat.
Réf 14: POLLARD (T-D) ,
Urate oxydaseen grde []
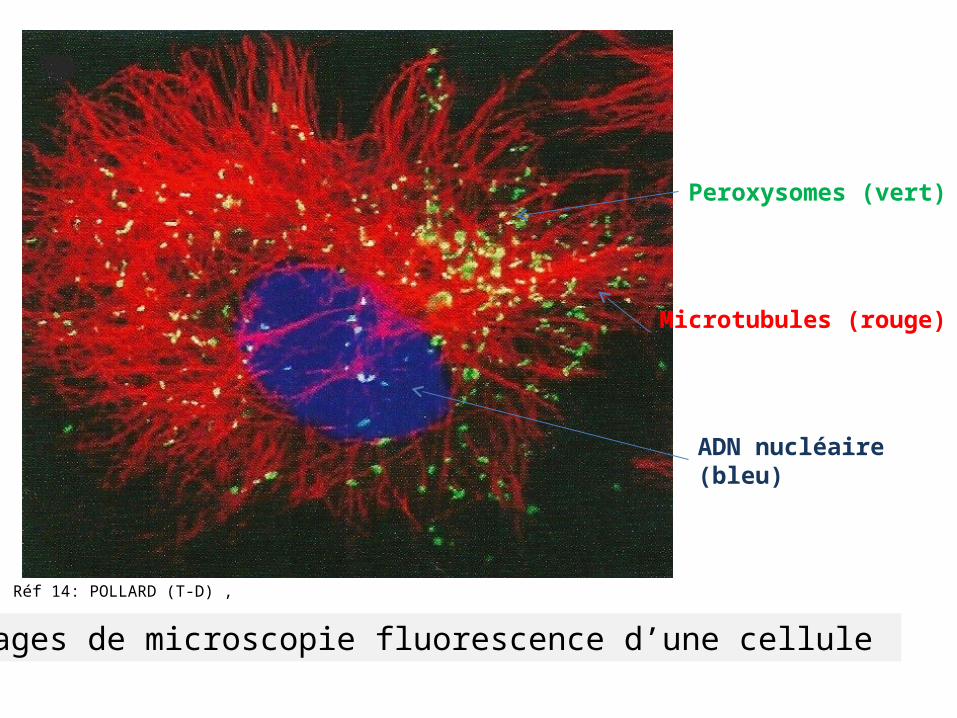
Réf 14: POLLARD (T-D) ,
Images de microscopie fluorescence d’une cellule
Peroxysomes (vert)
Microtubules (rouge)
ADN nucléaire (bleu)



L PTS1 /2:Peroxysomal Targeting Signal
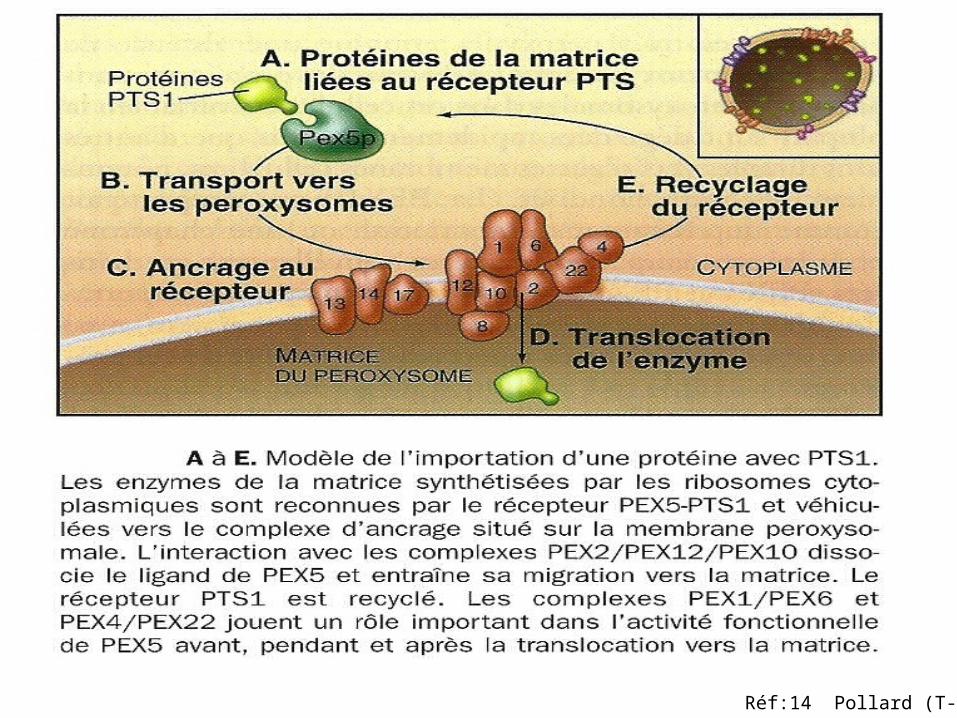
Réf:14 Pollard (T-D)





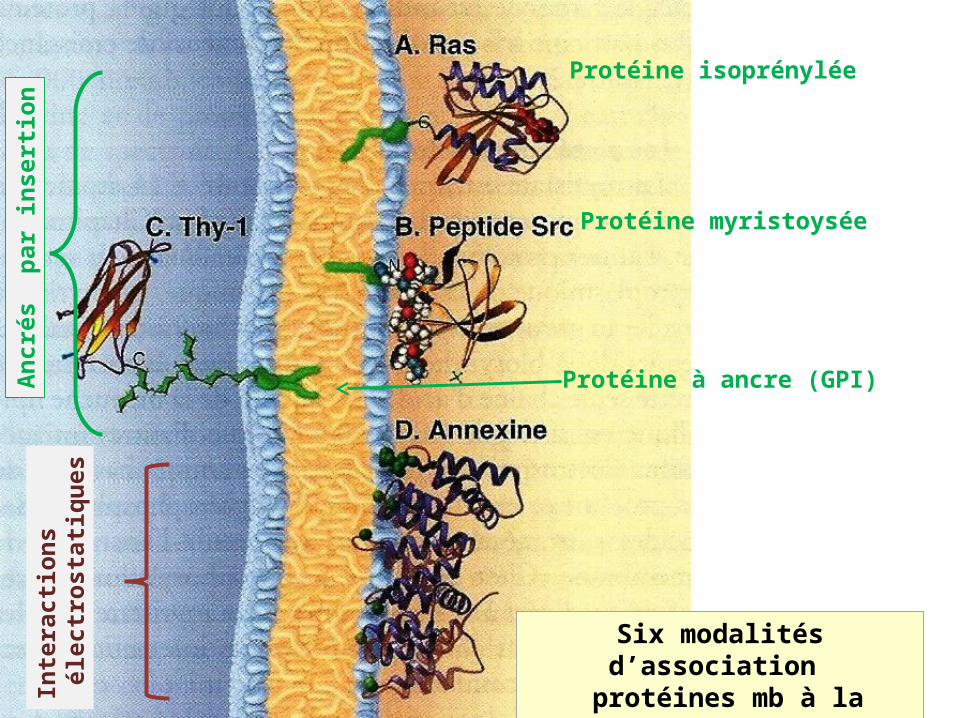
Six modalités d’association protéines mb à la bicouche
Lipidique (A-D)
Anc
rés
par
inse
rtio
nIn
tera
ctio
ns é
lect
rost
atiq
ues
Protéine isoprénylée
Protéine myristoysée
Protéine à ancre (GPI)

Six modalités d’association protéines mb à la bicouche
Lipidique (E-F)
Inse
rtio
n pa
rtie
lle
Caténine associée protéine transmb
Liaison aux protéines transmb
Hélices alpha hydrophobes pénètrent dans couche lipidique



Fabrique membranaire de la cellule
Rôle essentiel dans la biosynthèse des lipides et des protéines.(toutes les protéines transmb).
S’étend comme un filet sur tout le cytosol toutes les régions sont proches.
(Se limite en tout point par mb continue!)
[Réticulum endoplasmique]
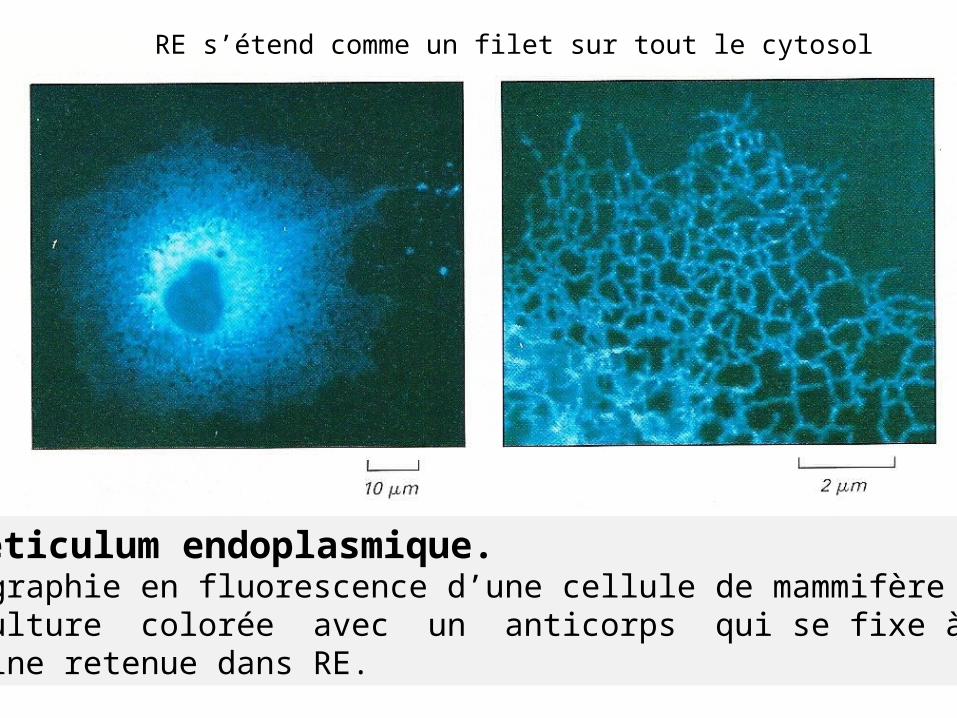
Le réticulum endoplasmique.Micrographie en fluorescence d’une cellule de mammifère en culture colorée avec un anticorps qui se fixe à une protéine retenue dans RE.
RE s’étend comme un filet sur tout le cytosol

RE
Rugueux LisseRER REL
La grande majorité des cellul contiennent peu de vrai REL, (plutôt petite région de transition , là ou *VT bourgeonnent) REL abondant dans certaines cellules spécialisées ,ex: hépatocyte.(*VT: vésicule de transport)
Sous forme de lames aplaties/avec ribosomes
Constitués de tubules contournés /sans ribosomes
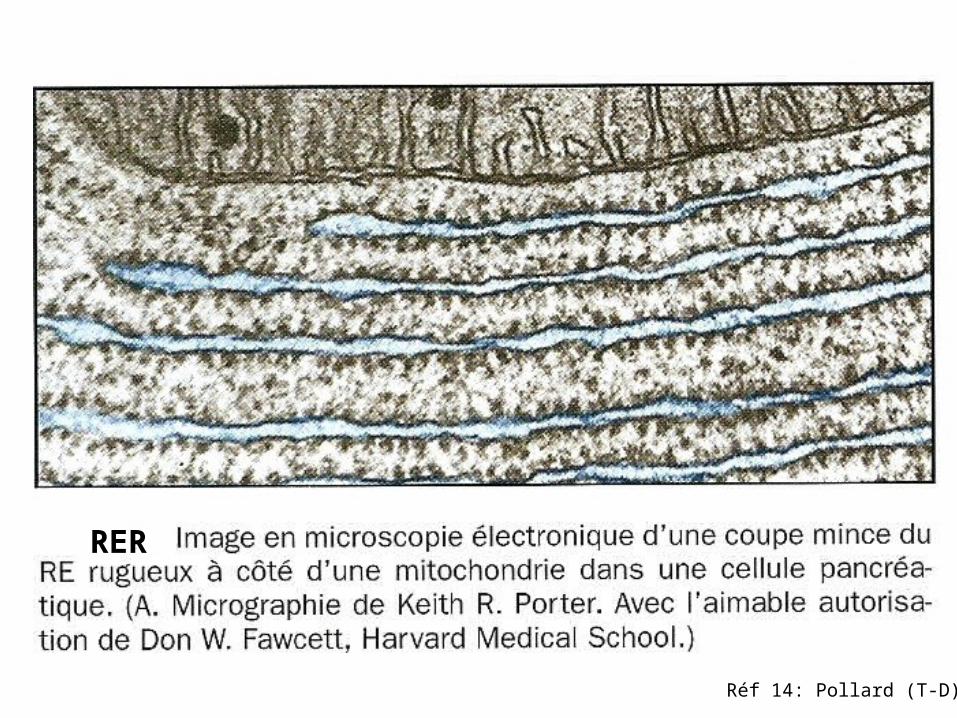
Réf 14: Pollard (T-D)
RER

Polyribosomes.Micrographie électronique d’une coupe fine de polyribosomes attachés à mb du RE .
Réf 1: ALBERTS (B) ,

Reconstitution tridimensionnelle d’une région des RE lisse et rugueux dans une cellule hépatique.
RE lisse: la mb du RE Lisse reliée aux saccules et forme un fin réseau de tubules de 30 à 60 nm de diam.
RE rugueux: forme des empilements orientés de 20 à 30 nm de large
Réf 1: ALBERTS (B) ,

Microsomes rugueux et lisses séparés en fonction de densité par centrifugation à l’équilibre/ gradient saccharose
Réf 1: ALBERTS (B) ,

Ribosomes du cytosol dirigés vers RE
Si protéine en train d’être fabriquée porte séquence signal, reconnue
par SRP situé dans cytosol
Complexe ribosome –SRP se lie à un récepteur de mb du RE
Processus de translocationSRP: Signal Recognition Particle

Ribosomes libres et ribosomes liés à la mbRéf 1: ALBERTS (B) ,


[Translocation dans le RE]
L’orientation d’une protéine dans la bicouche ou sa distribution vers la lumière , définie au cours du passage et préservée tout au long du transit par voie d’exocytose.
Le mécanisme de transport peut fonctionner dans le sens contraire: les protéines défectueuses sont expulsées du RE vers cytoplasme pour être dégradées.

Toutes les protéines destinées à:
la sécrétion RE lui-même AGLysosomesEndosomesMb plasmique
sont initialement importées dans RE à partir du cytosol

Il y a couplage de la synthèse et la translocation dans RE .
Les protéines destinés au RE :-Une séquence signal (N-terminale) dirige protéine vers RE;-un canal aqueux de la mb s’ouvre pour permettre translocation;-séquence signal éliminée après translocation dans lumière.
Séquence signal : variables , 15 à 21 aa , nécessaires et suffisantes pour ciblage du RE .
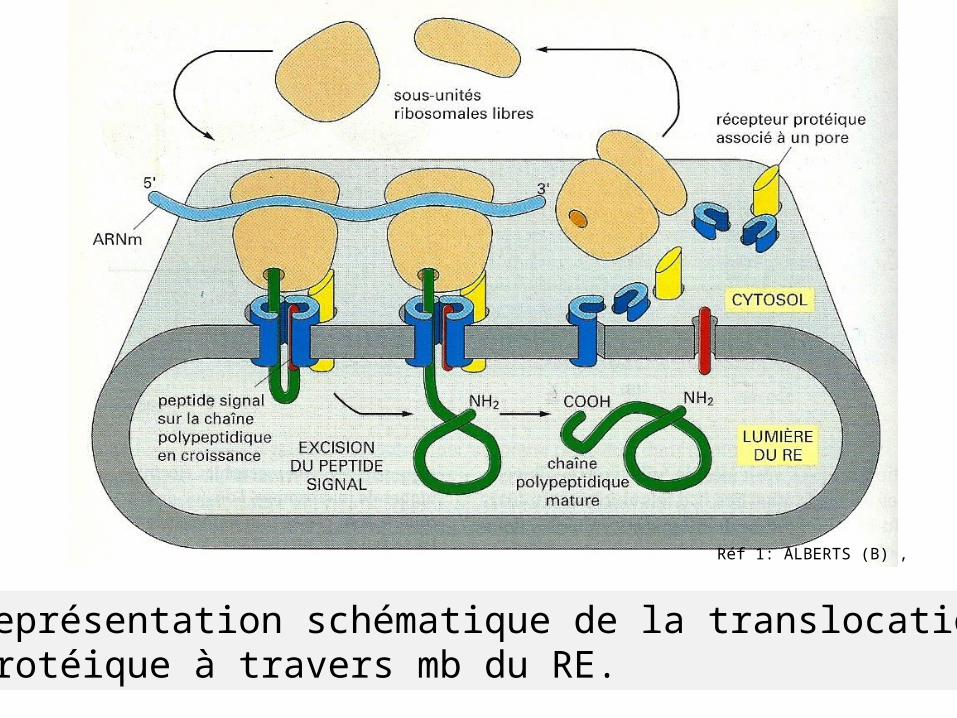
Réf 1: ALBERTS (B) ,
Représentation schématique de la translocation protéique à travers mb du RE.

*SRP se lie à la séquence signal et au ribosome
dirige ribosome vers récepteur (RER) le transfère vers un canal: complexe de translocation
dissociation de SRP et de son récepteur du ribosome
Reprise de synthèse protéique et passage du polypeptide
*SRP( signal recognision particle):
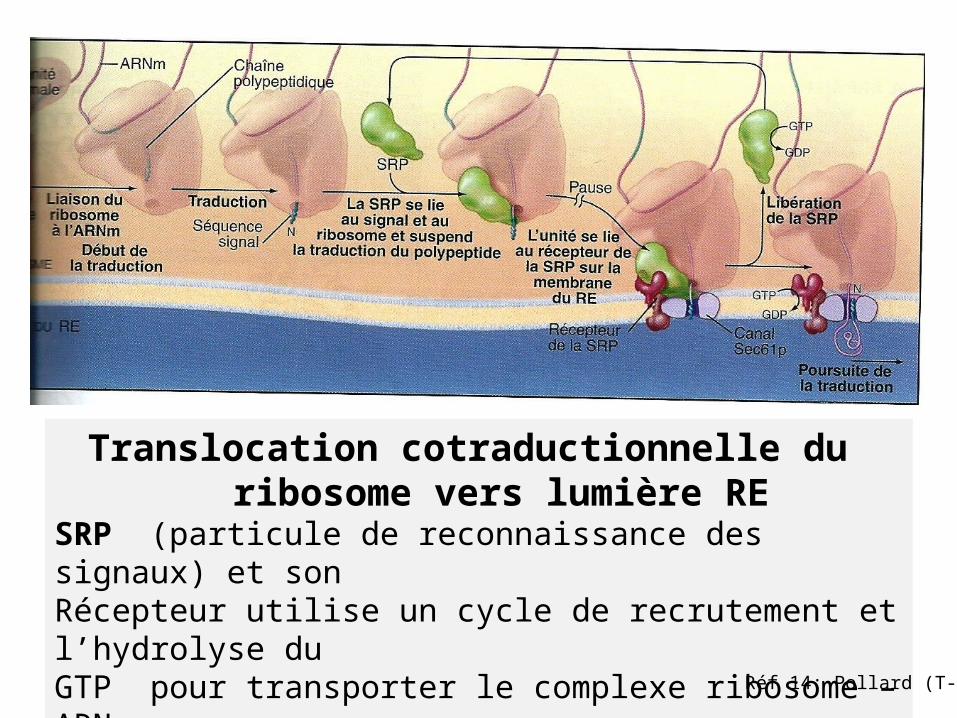
Translocation cotraductionnelle du ribosome vers lumière RE
SRP (particule de reconnaissance des signaux) et son Récepteur utilise un cycle de recrutement et l’hydrolyse du GTP pour transporter le complexe ribosome –ARNm vers le translocon du RE ;
Réf 14: Pollard (T-D)

Réf 2: ALBERTS (B) ,L’essentiel
Une séquence signal pour le RE et une RSP dirigent un ribosome vers la mb du RE

Les protéines traversent mb RE par un canal protéique étroitement ajusté :complexe de translocation.
Constituants du pore de translocation: 3 ou 4 copies du complexe Sec61p.
La liaison ribosome séquence signal à un complexe de translocation ouvre un pore transmb.

Structure du translocon Sec61p.A et B .Images de microscopies électroniques .C. canal Sec61 recombinant purifié .D. Canal natif isolé du RE. E. Ribosome lié à un complexe de translocation .F. Modèle de l’insertion d’un polypeptide dans un complexe de translocation et de son transfert vers bicouche lipidique.
Bicouche lipidique
Ribosome fixé par 3 à 4 minces supports
Hypothèse:dissociation partielle desSous unités
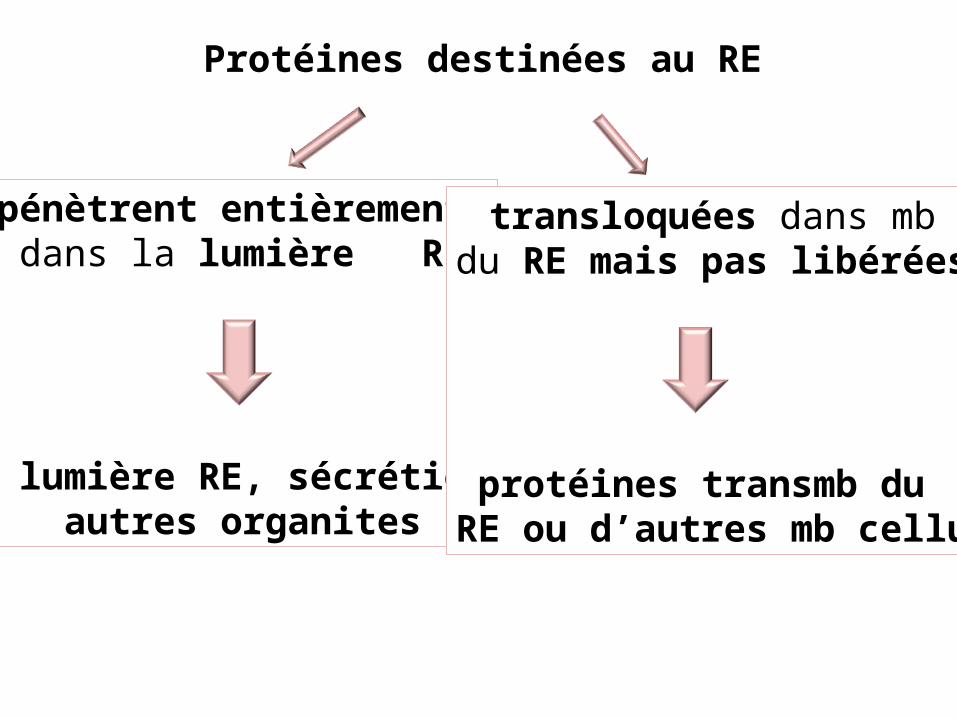
Protéines destinées au RE
pénètrent entièrement dans la lumière RE
lumière RE, sécrétionautres organites
transloquées dans mbdu RE mais pas libérées
protéines transmb du RE ou d’autres mb cellu

La sortie du RE
Etape importante de contrôle de qualité
protéines repliées incorrectement ou
assemblées à partenaires anormaux
retenues dans le RE et puis dégradées.

ALBERTS (B) ,L’essentiel
Translocation protéine soluble dans la lumière du RE , à travers mb du RE.

Intégration dans la mb du RE d’une protéine transmb.Une séquence signal amino-terminale pour le RE (rouge) initie le transfert ,une 2° séquence hydrophobe et une séquence de terminaison de transfert (orange)
Protéine ancrée/mb après clivage signal amino- terminal

Réf 2: ALBERTS (B) ,L’essentiel
Intégration dans la mb du RE d’une protéine à double passage transmb.

Ciblage des protéines dépendantes de Sec61p vers la lumière de la mb du RE.A. Libération de la protéine dans la lumière.B. Un seul segment transmb avec l’extrémité C- terminale dans le cytoplasme
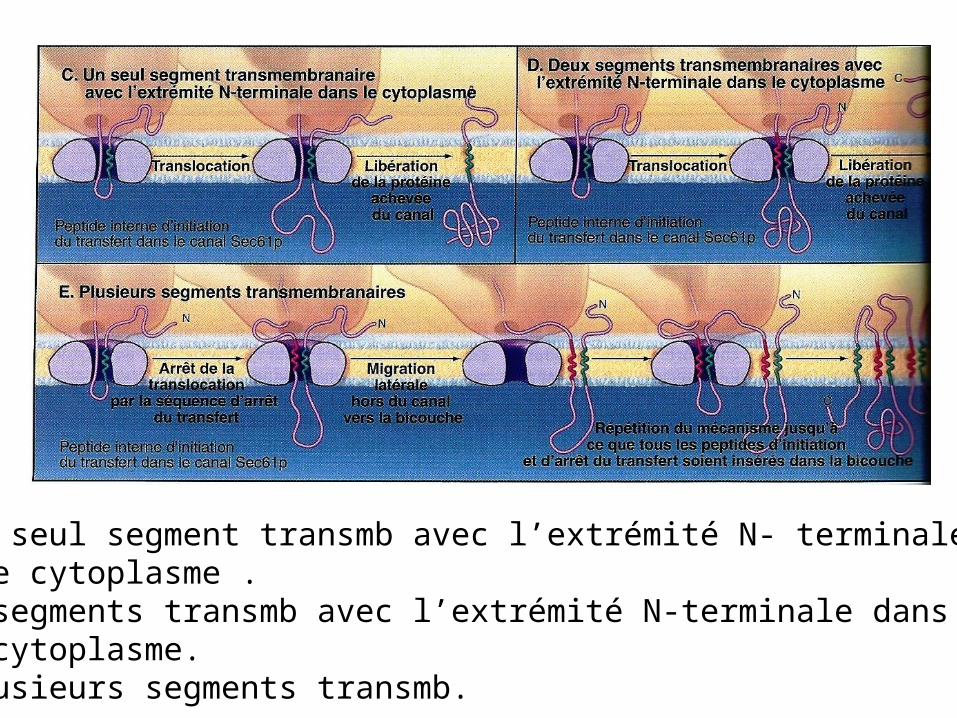
C. Un seul segment transmb avec l’extrémité N- terminale dans le cytoplasme .D. 2 segments transmb avec l’extrémité N-terminale dans le cytoplasme.E. Plusieurs segments transmb.

Protéines solubles : dans la plupart des cas , le peptide signal du RE est éliminé après la translocation.
Protéines qui traversent mb 1 seule fois: seul un peptide signal interne du RE reste dans la double couche lipidique sous forme α transmb.
Protéines transmb à traversées multiples: alternance de nombreux signaux d’initiation et de terminaison de transfert.


Le tunnel au sein de grande sous-unité ribosomique et le canal du complexe de translocation contiennent 70 aa .
Les chaines polypeptidiques longues émergent dans lumière du RE , elles :
-éliminent la séquence signal-(+) oligosaccharides-régulent le repliement- formation des ponts disulfures-constitution d’oligomères
Les facteurs de modification doivent être situés à proximité du versant luminal du complexe de translocation.
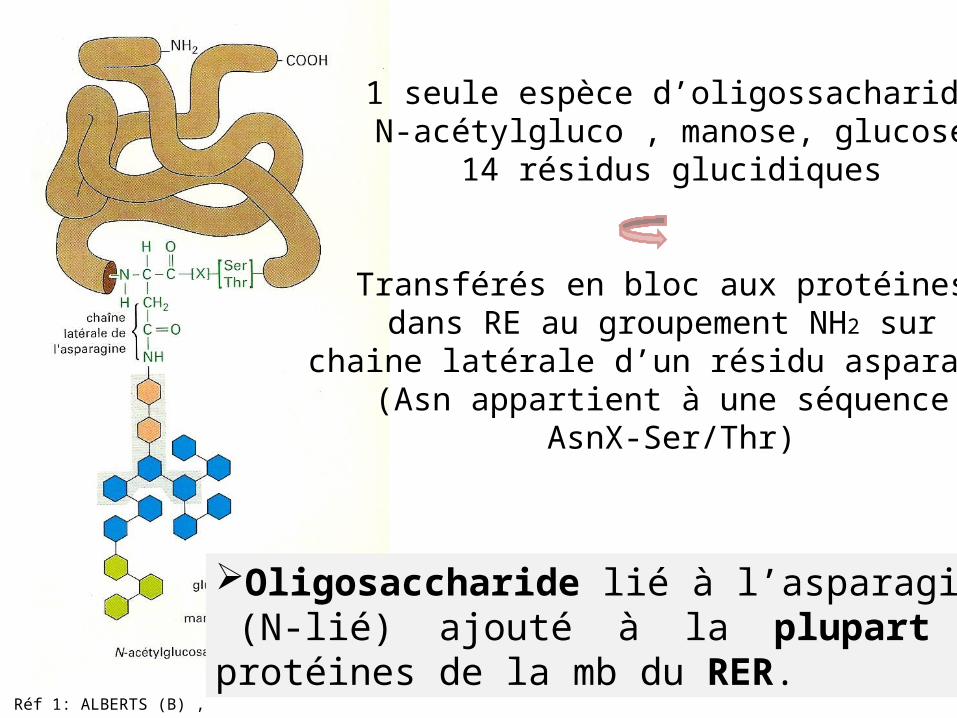
Réf 1: ALBERTS (B) ,
Oligosaccharide lié à l’asparagine (N-lié) ajouté à la plupart des protéines de la mb du RER.
1 seule espèce d’oligossacharideN-acétylgluco , manose, glucose
14 résidus glucidiques
Transférés en bloc aux protéines dans RE au groupement NH2 sur
chaine latérale d’un résidu asparagine(Asn appartient à une séquence
AsnX-Ser/Thr)

Glycosylation d’une protéine dans RER
oligosaccharide transféré en bloc
- risque d’agrégation
-rôle dans rôle dans repliement repliement
-rôle dans propriétés fonctionnelles rôle dans propriétés fonctionnelles dans milieu extracellu dans milieu extracellu

Réf 2: ALBERTS (B) ,L’essentiel
Glycosylation d’une protéine dans le RE rugueux
*ASN: asparagine
Copie de l’enzyme associée à chaque protéine de translocation dans mb du RE.

Le repliement complet et la formation d’oligomères ( assemblage sous unités) ne se produisent qu’après libération chaine polypeptidique/ribosome.
Chaque polypeptide a un mécanisme de repliement particulier , dictée par sa séquence.
Néanmoins ,les enzymes qui facilitent ces processus sont généralement les mêmes.

Les protéines qui entrent dans la voie d’exocytose
nombre élevé de ponts disulfures, stabilisent protéines dans conditions
oxydantes du milieu extracellu
(dans RE ces ponts disulfure grâce aux enzymes *PDI)
*PDI: protéine disulfure isomérase.

De nombreux polypeptides dans RE sont des sous unités de complexes oligomériques homologues ou hétérologues.
L’assemblage des oligomères ,généralement avant exportation et fait intervenir les chaperons.
Présence de chaperons favorise interactions entre les sous unités et empêche exportation ou dégradation prématurée des protéines.

RE doté d’un mécanisme régulé
Empêche l’exportation des protéines anormales
- Polypeptides mal repliés ,-sous unités en excès - oligomères mal assemblés
dégradés au sein du RE dans le protéasome

Dégradation dans la lumière du RE des protéines mal repliées.

Dégradation dans la lumière du RE des protéines mal repliées.
Mannosidase I identifie les glycoprotéines qui doivent être dégradés en synthétisant un
oligosaccharide à 8 mannoses qui est reglycolysé .
Ce substrat défavorable pour glucosidase IIreste lié à à calnexine
Transloqué hors RE pour être dégradédans le protéasome



Structure polarisée faite d’1 ou plurs empilements de saccules en forme de disque.


A
RE
Traitement du cœur des chaines N-osidiques dans le GolgiA à F . Les étapes successives comportent l’élimination de molécules de mannose (man) /glu (Glc) et + de N-acétylglucosamine (GlcNAc) , de galactose (gal) et d’acide Sialique (NANA) pour donner naissance à de nombreux oligosaccharides complexes.


[Synthèse lipidique]
La plupart des lipides cellulaires sont synthétisés dans RE et dans l’AG,
Cette synthèse est indispensable aux besoins de la voie d’exocytose et aux fonctions cellu qui dépendent des mb et des molécules de signalisation lipidique.
La plupart des glycérophopspholipides sont synthétisés dans RE ,les sphingolipides essentiellement dans AG.

[Stockage du calcium]
Une autre fonction du RE dans la plupart des cellu euc : séquestration des Ca++ cytosoliques .
Le cycle libération/ récupération des Ca++ du RE dans cytosol est responsable de la médiation d’un grand nombre de réponses rapides à des signaux cellu.

Le stockage de Ca++ dans lumière RE liée forte [ ] protéines liant Ca++.
Ex: cellul musculaires: REL spécialisé abondant = réticulum sarcoplasmique, pompe Ca++ par Ca++ATPase ,
libération et récupération par R sarco interviennent dans la contraction et le relâchement rapide des myofibrilles au cours de chaque cycle de contraction musculaire.