atividade tripanocida e anti-inflamatória de compostos
TRANSCRIPT

UNIVERSIDADE FEDERAL DE OURO PRETO
Núcleo de Pesquisas em Ciências Biológicas - NUPEB
Programa de Pós-Graduação em Ciências Biológicas – CBIOL
Laboratório de Imunobiologia da Inflamação – LABIIN
Atividade tripanocida e anti-inflamatória de compostos derivados de
1,8-dioxo-octa-hidroxantenos.
ANA PAULA DE JESUS MENEZES
OURO PRETO- MG
2019

II
ANA PAULA DE JESUS MENEZES
Atividade tripanocida e anti-inflamatória de compostos
derivados de 1,8-dioxo-octa-hidroxantenos
Orientador: Prof. André Talvani
Co-orientador: Guilherme de Paula Costa
OURO PRETO
ABRIL DE 2019
Tese apresentada ao Programa de Pós-graduação
em Ciências Biológicas do Núcleo de Pesquisas
em Ciências Biológicas da Universidade Federal
de Ouro Preto, como parte integrante dos
requisitos para obtenção do grau de Doutora em
Ciências Biológicas.
Área de concentração: Imunobiologia de
Protozoários

III

IV

V
AGRADECIMENTOS
Ao professor André Talvani pela orientação e amizade;
Ao agora professor Guilherme de Paula Costa, pela co-orientação;
Ao professor Dr. Robson Teixeira da Universidade Federal de Viçosa e a sua aluna Milene
Lopes, que gentilmente cederam as Xantenodionas sempre foram solícitos e dispostos a
contribuir de maneira efetiva para a realização deste trabalho;
Ao professor Dr. Rick Tarleton (UGA) e toda sua equipe pela orientação no Programa de
Doutorado Sanduíche e pelo auxílio não só pela construção do conhecimento científico e
tórico, mas também pelo crescimento pessoal;
Á Ludmila Walter dos Reis e Luíza Perucci por compartilharem toda experiência acadêmica
e pela incansável determinação em nos ajudar na rotina do LABIIN;
À todos os integrates do Laboratório de Imunobiologia da Inflamação pela ajuda na execução
deste trabalho, pela convivência diária e por serem a família que eu escolhi na UFOP;
À Universidade Federal de Ouro Preto pelo ensino público e gratuito de qualidade;
Aos órgãos de fomento, Capes, Fapemig e CNPQ.
Um agradecimento especial a Glenda, Paula e Priscila, amigas presentes desde o início desta
jornada. A República Mistura Perfeita pela amizade e por me permitir viver tudo que Ouro
Preto tem de melhor;
Aos amigos de Athens por estarem ao meu lado todos os dias e por me ajudarem em todas
as situações! Vocês sabem o quão são importantes para mim!
Á minha mãe Vitoria, por ser exemplo de determinação e coragem e por me incentivar a
sempre ir além! Ao meu irmão Daniel e a Ludmila pelo companheirismo! À minha avó
Mundica, zelosa pela minha educação desde criança e certamente estará torcendo por mim
no plano espiritual. Obrigada por serem a melhor família, por me permitirem embarcar em
busca dos meus sonhos, me distanciando de vocês que mais amo. Hoje, percebo que não
estou livre do medo e de tantas outras fraquezas, mas chegar até aqui me deixa livre do medo
de nunca ter tentado!

VI
RESUMO
A infecção pelo Trypanosoma cruzi acomete milhares de pessoas pelo mundo e a eficácia dos
fármacos disponíveis para eliminar este parasito é limitada à fase aguda da doença. A busca por
moléculas mais efetivas contra o T. cruzi e de menor toxicidade para o hospedeiro é, portanto, uma
prioridade para a saúde pública mundial. Neste contexto surgem as 1,8-dioxo-octahidroxanthenos ou
xanthenodionas, compostos sintéticos derivados das xantonas que possuem inúmeras aplicações
terapêuticas descritas, dentre elas atividades anti-inflamatória, tripanocida e cardioprotetora. Assim,
o objetivo deste trabalho foi avaliar a ação tripanocida, in vitro, de compostos derivados das 1,8-
dioxo-octahydroxantenos e sua influência, in vivo, na parasitemia e na alteração dos parâmetros
inflamatórios durante a infecção murina. Para avaliar a toxicidade dos compostos in vitro utilizou-se
testes colorimétricos (MTT) em formas tripomastigotas (cepa Y) e em macrófagos J774A.1, além de
ensaios fluorimétricos (AlamarBlue) em células Vero com parasitos da cepa CL expressando uma
proteína vermelho fluorescente (CLtdTomato) para determinação da CC50, IC50 e IS. Nesta etapa,
doze compostos foram avaliados (MI17, MI18, MI24, MI25, MI39, MI45, MI68, MI73, MI78, MI79,
MI80 e MI81) foram avaliados nesta etapa. O compostos MI80 (IC50=30.65) destacou-se por
apresentar atividade anti-T. cruzi, com o melhor IS (170). Para avaliar in vivo o desempenho
tripanocida e imunológico do composto MI80, camundongos Swiss foram infectados com as cepas
Y (n = 10) e CL (n = 10) de T. cruzi utilizando um protocolo de tratamento por 10 dias com 50mg /
kg de MI80. O MI80 reduziu 40% do pico de parasitemia em animais infectados com a cepa Y,
enquanto reduziu, em ambas as cepas, o infiltrado inflamatório cardíaco. RankL, CCL2 e IFN-g
foram mensurados no tecido cardíaco, mas o MI80 não foi capaz de modificar seu padrão em animais
infectados com a cepa Y. No entanto, o MI80 aumentou o RankL e IL-10 em camundongos
infectados com CL. Não foi observada toxicidade hepática e renal nos animais advindas do
tratamento com o composto MI80. . Nossos achados mostraram que o 1,8-dioxo-octa-hidroxanteno
tem um efeito anti-parasitário limitado e foi capaz de melhorar os parâmetros inflamatórios cardíacos
relacionados à infecção por T. cruzi.

VII
ABSTRAT
Trypanosoma cruzi infection affects thousands of people worldwide and the efficacy of drugs
available to eliminate this parasite is limited. The1,8-dioxo-octahydroxanthenos or xanthenediones
are derivatives synthetic compounds from xanthones presenting considerable biological applications.
The aim of this study was to assess the trypanocidal and inflammatory activities of 1,8-dioxo-
octahydroxanthenes derivatives using the CL strain of T. cruzi expressing the tandem red fluorescent
protein (tdTomato) to test in vitro 12 xanthenediones derivatives (MI17- 18, 24, 25, 39, 45, 68, 73,
78, 79, 80, 81). The killing assay showed that three compounds were able to eliminate this parasite:
MI24 (Ic50=26.43µM), MI25 (Ic50= 14.11µM) and MI80 (Ic50=30.65). However, only MI80 was
not toxic for Vero cells after the viability assay. To evaluate in vivo the tripanocidal and the
immunological performance of the MI80, Swiss mice were infected with the Y (n=10) and CL (n=10)
strains of T. cruzi using a treatment protocol for 10 days with 50mg/kg of MI80. MI80 reduced 40%
of the parasitemia peak in animals infected with Y strain while reduced, in both strains, the cardiac
inflammatory infiltration. RankL, CCL2 and IFN-g were measured in cardiac tissue, but the MI80
was not able to modify their pattern in Y-strain infected animals. However, MI80 increased the
RankL and IL-10 in CL-infected mice. No hepatic and renal toxicity was observed with the MI80.
Our findings showed that 1,8-dioxooctahydroxanthene has a limited anti-parasitic effect and it was
capable to ameliorate the cardiac inflammatory parameters related to T. cruzi infection.

VIII
LISTA DE ABREVIATUTAS
ABTS – Ácido 2,2'-azino-bis (3-etilbenztiazolina-6-sulfonico)
AMPc – Monofosfato cíclico de adenosina
Bz – Benznidazol
CC50 - Concentração citotóxica para 50% das células
CCA – Centro de ciência animal
CCL2 - Quimiocina ligante de monocito
CEUA – Comissão de experimentação em uso animal
COX – Ciclo-oxigenase
DMSO - Dimetilsulfóxido
DMXAA - Ácido dimetilxantenona-4-acético
DNA – Ácido dexiribonucleico
DPI – Dias após a infecção
GAL - D-galactosidase
IC50 – Concentração inibitória para 50% dos parasitos
IFN - Interferon
IL-10 - Interleucina 10
IL4 – Interleucina 4
IL6 – Interleucina 6
iNOS – Enzima óxido-nitrico sintase
IS - Índice de seletividade
LDNT – Triptose neutralizada e digerida de fígado
MTT – 3-(4,5-di-metilazol-2-il) -2,5-difeniltetrazolium brometo tetrazol
NFAT – Fator nuclear de células T ativadas
NF-kb – Fator nuclear kappa-Β
NI - Grupo de animais não infectados
NIT - Grupo de animais não infectados e tratados com 50mg/kg da XtdMI80
NO – Óxido nítrico
Nxt – Nifurtimox
PBS – Tampão fosfato salino
PCR – Reação em cadeia da polimerase
PGE – Prostaglandina E

IX
RANK – Ligante ativador do receptor do fator nuclear kappa-Β
RNA – Ácido ribonucleico
ROS – Espécies reativas de oxigênio
rRNA – Ácido ribonucleico ribossomal
SE – Erro padrão
SFB – Soro fetal bovino
AST/TGO – Transaminase oxalacética
ALT / TGP– Transaminase pirúvica
Th1 – Células T auxiliares com perfil do tipo 1
TNF - Fator de necrose tumoral
Treg – Células T reguladoras
Xtd - Xantenodiona
MI80 - Derivado de xantenodiona MI80

X
SUMÁRIO
1. INTRODUÇÃO ...................................................................................................... 12
1.1 Xantenodionas: histórico e relevância ............................................................................... 12
1.2 Potencial anti-inflamatório ................................................................................................ 16
1.3 Ação cardioprotetora das xantonas.................................................................................... 19
1.4 - Atividade antiparasitária das xantonas ............................................................................ 20
1.5 O T.cruzi como um indutor do processo inflamatório ...................................................... 22
1.6 Estratégias terapêuticas na doença de Chagas ................................................................... 23
2. OBJETIVOS............................................................................................................ 25
2.1 Objetivo geral .................................................................................................................... 25
2.2 Objetivos específicos......................................................................................................... 25
3. MATERIAL E MÉTODOS .................................................................................... 26
a. Representação esquemática do delineamento experimental ............................................ 26
3.1 Compostos ......................................................................................................................... 26
3.2 Parasito .............................................................................................................................. 28
3.3 Culturas Celulares ............................................................................................................. 29
3.4 Obtenção das formas tripomastigotas de cultura celular ................................................... 29
3.5 Modelo animal .................................................................................................................. 30
3.6 Experimentos In vitro ........................................................................................................ 30
3.6.1 Verificação da atividade tripanocida - Ensaio de citotoxidade em formas tripomastigotas
derivadas de cultura celular ..................................................................................................... 30
3.6.2 Avaliação do efeito tripanocida em formas amastigotas ................................................ 31
3.6.2.1 Geração de parasitos modificados – T. cruzi fluorescentes (tdTomato)...................... 31
3.6.2.2 Avaliação do potencial tripanocida em amastigotas ................................................... 33
3.6.3 Ensaios de citotoxidade em culturas de células – determinação da CC50....................... 33
3.6.3.1 Macrófagos murinos-MTT® ....................................................................................... 34
3.6.3.2 Células Vero- AlamarBlue® ....................................................................................... 34
3.7 Experimentos In vivo ......................................................................................................... 36
3.7.1 Padronização – Experimento de dose-resposta .............................................................. 36

XI
3.7.2 Infecção e esquema de tratamento ................................................................................. 36
3.7.3 Exame de sangue a fresco - parasitemia ......................................................................... 36
3.7.4 Mortalidade .................................................................................................................... 37
3.7.5 Histopatologia do coração .............................................................................................. 37
3.7.6 Estereologia do coração ................................................................................................. 38
3.7.7 Produção de citocinas ..................................................................................................... 38
3.7.8.1 Transaminase oxalacética (AST/TGO) ....................................................................... 39
3.7.8.2 Transaminsae pirúvica (ALT/TGP) ............................................................................ 40
3.7.8.3 Creatinina .................................................................................................................... 41
3.7.8.4 Ureia enzimática .......................................................................................................... 41
3.8 Análise estatística .............................................................................................................. 42
4. RESULTADOS ....................................................................................................... 43
4.1 - Resultados in vitro .......................................................................................................... 43
4.1.1 Avaliação da toxicidade em formas tripomastigotas do T.cruzi e determinação do IC50
................................................................................................................................................. 43
4.1.2 Efeito dos compostos na viabilidade celular demacrófagos murinos J774A.1)avaliados
por ensaios colorimétricos ....................................................................................................... 45
4.1.3 Atividade tripanocida em formas amastigotas do T.cruzi (cepa CLtdTomato) .............. 47
4.1.4 Efeito sobre a viabilidade celular em células Vero por ensaios fluorimétricos e seu
respectivo índice de seletividade ............................................................................................. 50
4.2 Resultados In vivo ............................................................................................................. 55
4.2.1 Padronização da dose e via de tratamento ...................................................................... 55
4.3 Avaliação do tratamento com o composto MI80 na dose de 50mg/kg por via oral .......... 57
4.3.1 Parasitemia e mortalidade .............................................................................................. 57
4.3.2 Massa corporal e massa relativa do coração, baço e fígado ........................................... 59
4.3.4 Efeito do tratamento com MI80 nos parâmetros inflamatórios na infecção experimental
pelo T.cruzi .............................................................................................................................. 60
4.3.4.1 Histopatologia e estereologia do miocárdio ................................................................ 60
4.3.4.2 Mediadores inflamatórios ............................................................................................ 64
4.3.6 Tratamento com a xantenodiona MI80 e avaliação da toxicidade renal e hepática ....... 65

XII
5. DISCUSSÃO ........................................................................................................... 67
6. CONCLUSÃO ........................................................................................................ 76
7. REFERÊNCIAS ...................................................................................................... 77

XIII
ÍNDICE DE FIGURAS
Figura 1 – Representação molecular dos compostos. Em (A) Xanteno (9H-Xanthene ou
Dibenzo pirano) e em (B) Xantona (9H-xanten-9-ona). ............................................................. 13
Figura 2 – Estrutura do núcleo das xantenodionas e seu padrão de distribuição ................ 15
Figura 3 – Representação esquemática dos métodos realizados para estudo da atividade das
xanthenodionas. (A) Experimentos in vitro. (B) Ensaios in vivo de padronização da dose e via
de tratamento(n=5). (C) Avaliação do tratamento com a MI80 na redução da parasitemia e do
potencial anti-inflamatório (n=10). CC50, concentração citotóxica de 50%; IC50, concentração
inibitória de 50%, DPI, dias após a infecção. .............................................................................. 26
Figura 4 - Formula estrutural do núcleo xantenodiona. Esqueleto utilizado para construção das
respectivas séries químicas.......................................................................................................... 27
Figura 5 - Plasmídeo utilizados na geração de T. cruzi fluorescente. Representação
esquemática do plasmídeo pTrex-Neo-tdTomato. ...................................................................... 32
Figura 6 - Avaliação da fluorescência do T. cruzi (cepa CL) tdtomato. (A) Imagem
microscópica dos parasitas expressando tdTomato em todas as fases evolutivas. (B) Intensidade
de fluorescência em epimastigotas em citômetro de fluxo CyAn (DakoCytomation) e analisada
com o software FlowJo (Tree Star). A fluorescência de fundo dos parasitas selvagens (verde)
também pode ser observada. Fonte: Canavaci et al, 2010 ........................................................... 32
Figura 7 – Análise do efeito citotóxico em formas tripomastigotas T. cruzi (cepaY), pelo
método do MTT. 1x106 foram adicionados por poço, em placa de 96 poços e os compostos
MI17, MI24, MI25, MI79, MI80 ou MI81 foram nas concentrações de 5 a 300µM/mL e incubados
por 24h. Os resultados foram expressos como valores em porcentagem do número de parasitos
viáveis após 24 horas de incubação com os derivados de xantenodionas. Análise de variância
(ANOVA) e pos-teste de Tukey foram aplicadas para determinar a diferença estatística entre os
tratamentos em relação ao controle DMSO. (*) representa diferença estatística determinada por
p<0,05. ........................................................................................................................................ 44
Figura 8 – Análise de viabilidade celular de culturas de macrófagos murinos J774A.1
avaliadas pelo método do MTT frente aos derivados de xantenodionas. 1x105 celulas foram
adicionada aos poços em placa de 96 poços.Após a formação da monocamada os compostos
MI17, MI24, MI25, MI79, MI80 ou MI81 foram adicionados em concentrações decrescentes de
5 a 300µM/mL e as placas incubadas por 48h em estufa 37ºC 5% CO2.Após 48h foi realizado o
ensaio de MTT e os resultados foram expressos em porcentagem do número de células viáveis
após 48 horas de incubação com os derivados de xantenodionas . ............................................. 46
Figura 9 - Atividade das xantenodionas MI24, MI25 e MI80 sobre a infecção de células Vero
pela cepaCLtdtomato do T. cruzi. 2,5x104 células foram infectadas pela cepa CLtdtomato e,
posteriormente, incubadas na presença ou ausência de concentrações decrescentes de

XIV
xantenodionas MI24, MI25 e MI80, por 72 horas. Cada ponto das curvas de dose-resposta
corresponde à média de 2 experimentos independentes, em triplicata. O cálculo de IC50 foi
realizado utilizando-se o software GraphPadPrism6. .................................................................. 48
Figura 10 - Fotomicrografias representativas dos efeitos anti-T.cruzi dos compostos MI24,
MI25 e MI80. Estão representadas fotomicrografias de células Vero infectadas com cepa
CLtdTomato sem tratamento e tratadas com as xantenodiona MI24, MI25 e MI80 ou com o
fármaco benznidazol (Bz) ao terceiro dia após a infecção in vitro. ............................................ 49
Figura 11 – Análise da viabilidade celular em culturas de células Vero pelo método do
AlamarBlue. 2,5x104 foram plaqueadas e incubadas em estufa 37ºC, 5%CO2 por 72h com os
derivados de xantenodionas MI79, MI80, MI81, MI73 e MI39 em concentrações decrescentes de
100 a 0,39µM/mL. As barras representam os valores em porcentagem do número de células
viáveis de três experimentos independentes em triplicata e erro padrão (SE). ........................... 51
Figura 12 - Viabilidade celular em culturas de células Vero pelo método do AlamarBlue,
determinado após 72 horas de incubação com os derivados de xantenodionas MI17, MI24, MI25,
MI45 e MI49 em diferentes concentrações. As barras representam os valores em porcentagem do
número de células viáveis de três experimentos independentes em triplicata e erro padrão (SE).
..................................................................................................................................................... 52
Figura 13 - Viabilidade celular em culturas de células Vero pelo método do AlamarBlue,
determinado após 72 horas de incubação com os derivados de xantenodionas MI18, MI68 e MI78
em diferentes concentrações. As barras representam os valores em porcentagem do número de
células viáveis de três experimentos independentes em triplicata e erro padrão (SE). ............... 53
Figura 14 - Curva de parasitemia. Camundongos Swiss foram infectados com 5.000 formas
tripomastigotas sanguíneas da cepa Y do T. cruzi (n=5) e tratados ou não com a xantenodiona
MI80 ou Benznidazol (Bz). Os dados em cada ponto da curva equivalem à média da
parasitemia/dia referente a cada grupo. O símbolo (*) indica p<0,05 em relação ao grupo controle
infectado e não tratado.O gráfico superior indica a parasitemia durante o tratamento por via oral
e o gráfico inferior por via intraperitoneal. ................................................................................. 56
Figura 15 - Curvas de letalidade dos animais infectados e tratados com a MI80 por via oral
ou intraperitoneal. Animais Swiss fêmeas foram infectados com a cepa Y do T. cruzi, tratados
ou não com MI80 em doses decrescentes de 800 a 50mg/kg ou BZ (100mg/kg) (n=5). Curvas
referentes ao período compreendido entre o 1º e o 15 º dia de infecção. O gráfico a direita
representa a taxa de mortalidade dos animais tratados por via oral e o gráfico da esquerda a
mortalidade dos camundongos tratados intraperitonealmente. ................................................... 57
Figura 16 - Curva de parasitemia em camundongos Swiss infectados pelo T. cruzi. Os animais
foram infectados com 5x103 formas tripomastigotas sanguíneas da cepa Y (A) ou CL (B) do T.
cruzi (n=10), tratados ou não com a xantenodiona MI80 ou Bz. Os dados em cada ponto da curva

XV
equivalem à média da parasitemia/dia referente a cada grupo. (*) representa p<0,05 em relação
ao grupo infectado e tratado com Bz. .......................................................................................... 58
Figura 17 - Curvas de letalidade de camundongos Swiss infectados com 5x103 formas
tripomastigotas sanguíneas da cepa Y ou CL do T. cruzi, tratados ou não com a MI80 ou
Bz. Animais Swiss fêmeas foram infectados com a cepa Y ou CL do T. cruzi, tratados ou não
com MI80(50mg/kg) ou BZ (100mg/kg) (n=10). Curvas referentes ao período compreendido
entre o 1º e o 15 º dia de infecção para cepa Y e 1º ao 18º para a cepa CL. O gráfico a direita
representa a taxa de mortalidade dos animais tratados para cepa CL o gráfico da esquerda a
mortalidade dos camundongos infectados coma cepa Y.Os resultados representam porcentagem
de animais vivos. ......................................................................................................................... 59
Figura 18 - Massa corporal e massa relativa do coração, baço e fígado. Camundongos Swiss
infectados e não infectados com a cepa Y ou CL do T. cruzi, tratados ou não com a MI80 ou Bz.
A massa corporal (A) dos animais foi avaliada após a eutanásia e, em seguida, o peso relativo do
coração (B), baço (C) e fígado (D), obtido a partir relação entre a massa do órgão dividido pela
massa corporal do animal. As caixas representam a mediana, máximo e mínimo. (*) representa
p<0,05 em relação aos controles NI e NIT. (#) mostra também diferença estatística em relação ao
grupo infectado e tratado com Bz. (Xtd) Xantenodiona MI80, (Bz) Benznidazol, (NI) Não
infectado e (NIT) Não infectado tratado com a MI80. ................................................................ 60
Figura 19 - Quantificação do processo inflamatório no músculo cardíaco. Camundongos
Swiss infectados com 5.000 formas tripomastigotas sanguíneas da cepa Y ou CL do T. cruzi foram
tratados ou não com MI80 ou Benznidazol (Bz) na fase aguda. Análise do infiltrado inflamatório
contendo células mononucleares (A) e polimorfonucleares (B) no miocárdio dos animais. (C)
Diagrama de campo octogonal organizado em domínios que representa a celularidade tecidual da
seguinte forma: (-) celularidade mínima / (+ ) leve, (+ +) moderada ou (+ + +) intensa / infiltrado
inflamatório. * p< 0,05 comparado com o grupo controle infectado e não tratado; # p<0,05 em
relação ao grupo tratado com Bz e & p<0,05 comparado ao grupo tratado com MI80. (NI) Não
infectado e (NIT) Não infectado tratado com a MI80. ................................................................ 61
Figura 20 - Fotomicrografia de cortes histológicos do tecido muscular cardíaco.
Camundongos Swiss foram infectados com 5.000 formas tripomastigotas sanguíneas da cepa Y
ou CL e tratados ou não com MI80 ou Bz, na fase aguda. As figuras superiores representam os
animais infectados pela cepa Y, além do controle não infectado. As imagens inferiores mostram
os animais infectados pela cepa CL e o controle não infectado e tratado com a MI80.Nos animais
infectados e não tratados (Y) ou (CL), o infiltrado inflamatório apresentou-se de maneira
moderada e foi possível identificar ninhos de amastigotas. Para o grupo de animais infectados e
tratados (Y+MI80) e (CL+MI80) verificou-se processo inflamatório discreto e focal. Nos animais
infectados e tratados com as doses 100 mg/Kg de Bz não houve detecção de infiltrado. Os
controles NI e NI+MI80 apresentaram aspecto histológico cardíaco normal. Barra 60μm2. (MI80)

XVI
Xantenodiona MI80, (Bz) Benznidazol, (NI) Não infectado e (NIT) Não infectado tratado com a
MI80. ........................................................................................................................................... 62
Figura 21 – Aspectos estereológicos do miocárdio. Camundongos Swiss foram infectados com
5.000 formas tripomastigotas sanguíneas da cepa Y ou CL e tratados ou não com MI80 ou Bz, na
fase aguda. Confirmada a infecção, os animais foram tratados por 10 dias e o coração foi removido
e fixado em formol tamponado (10%). (A) Densidade volumétrica do caridiomiócito. (B)
Densidade volumétrica do tecido conjuntivo. (C) Densidade volumétrica dos vasos sanguíneos.
(D)Vascularização do miocárdio. (E) Compartimentos estruturais e funcionais. * p< 0,05
comparado com o grupo controle infectado e não tratado; # p<0,05 em relação ao grupo tratado
com Bz e & p<0,05 comparado ao grupo tratado com MI80. (NI) Não infectado e (NIT) Não
infectado tratado com a MI80. (Vv[cmy]) = volume total dos cardiomiócitos, (Vv [cnt])= volume
intersticial e (Vv [bvs])= volume de vasos sanguíneos. .............................................................. 63
Figura 22 - Perfil de marcadores inflamatórios no sobrenadante do macerado de miocárdio
de camundongos infectados ou não com a cepa Y ou CL do T.cruzi e tratados com a MI80
ou Bz. (A) Concentração de RankL. (B) níveis de CCL2, (C) IFN-gama no miocárdio e (D) IL-
10. O símbolo (#) representa p<0,05 em relação ao grupo infectado e tratado com Bz e (*) em
relação aos grupos NI e NIT. Os resultados estão expressos em caixas contendo a mediana,
máximo e mínimo. (Xtd) Xantenodiona MI80, (Bz) Benznidazol, (NI) Não infectado e (NIT) Não
infectado tratado com a MI80. .................................................................................................... 65
Figura 23 - Concentrações plasmáticas de TGO (A), TGP (B), ureia (C) e creatinina (D) de
camundongos infectados com as cepas Y e CL do T. cruzi, tratados com 50mg/kg durante
10 dias por via oral. O símbolo (*) representa diferença estatística (p<0,05) em relação aos
controles NI e NIT (#) representa p<0.05 em relação ao grupo infectado e tratado com Bz e (&)
indica diferença entre o grupo infectado e não tratado e o grupo tratado com a MI80. (, (Bz)
Benznidazol, (NI) não infectado e (NIT) não infectado tratado com a MI80 ............................. 66

XVII
ÍNDICE DE TABELAS
Tabela 1 – Atividade anti-inflamatória in vivo de xantonas e seus derivados ............................ 18
Tabela 2 - Estrutura molecular representativa das xantenodionas. Fórmula estrutural evidenciando
radicais e suas posições no núcleo das xantenodionas avaliadas neste estudo. ........................... 28
Tabela 3 – Citotoxidade em macrófagos murinos, atividade tripanocida em tripomastigotas e
índice de seletividade de compostos derivados de xantenodionas .............................................. 47
Tabela 4 - Citotoxidade em células Vero, atividade tripanocida em formas amastigotas do T. cruzi
e o índice de seletividade de compostos derivados de xantenodionas ........................................ 54

12
1. INTRODUÇÃO
1.1 Xantenodionas: histórico e relevância
Na química orgânica, os compostos heterocíclicos constituídos principalmente por
oxigênio e nitrogênio possuem grande relevância por apresentarem, dentre outras
vantagens, propriedades biológicas (Dua et al., 2009; Quin, 2010). Dessa forma, os
xantenos (Fig.1A) e seus derivados têm despertado interesse científico por suas atividades
antivirais, antibacterianas, anti-inflamatórias e antiparasitárias (Poupelin, 1978;
Vennerstrom et al., 1995; Reddi Mohan Naidu et al., 2012; Kaya et al., 2013). Além do
uso como precursor sintético para muitos compostos orgânicos (Shchekotikhin, 2002) e
corantes (Hilderbrand e Weissleder, 2007), os xantenos também foram investigados em
aplicações com tecnologias de laser (Pohlers et al., 1997; Hilderbrand e Weissleder,
2007) e com materiais fluorescentes para visualização de biomoléculas (Knight Cg,
1989).
Os xantenos isolados a partir de fontes naturais são denominados xantonas (9H-
xanten-9-ona) (Fig.1B). Esta classe de moléculas apresenta em sua estrutura dois anéis
aromáticos fundidos com uma y-piranona. Podem ser subdivididas, dependendo da
natureza dos substituintes na estrutura do dibenzo-γ-pirano, em xantonas oxigenadas
simples, xantonas glicosiladas, xantonas preniladas e seus derivados, dímeros de
xantonas, xantonolignóides e xantonas miscelâneas (Pinto et al., 2005). Em contrapartida,
as xantonas de origem sintética podem ter grupos simples, como hidroxila, metoxila,
metila, carboxila, bem como substituintes mais complexos, como epóxido, azol,
metilidenobutirolactona, aminoálcool, sulfamoil, ácido metiltiocarboxílico e
diidropiridina (Vieira e Kijjoa, 2005; Yang Chun-Hui, 2012).

13
Figura 1 – Representação molecular dos compostos. Em (A) Xanteno (9H-Xanthene ou
Dibenzo pirano) e em (B) Xantona (9H-xanten-9-ona).
Historicamente, a primeira xantona foi isolada em 1855 pelo químico alemão Dr.
W. Schmid que a extraiu a partir da casca de mangostão (Garcinia mangostana), a
primeira xantona natural denominada α-mangostina (Kuan, 2012). Devido a cor amarela
brilhante da α-mangostina, Schmid nomeou a nova classe química como xantona, termo
baseado na palavra grega xantos (amarelo) (Hostettman, 1989). Entretanto, as primeiras
investigações do potencial farmacológico destes compostos datam de cem anos após a
sua descoberta. Em 1968, o grupo de Bhattacharya isolou e caracterizou a mangiferina,
uma xantona da raíz das Malpighiacea com propriedades diuréticas e cardiotônicas
(Finnegan, 1968). Mais tarde, Da Re P (1970) foi o pioneiro a descrever as atividades
estimulantes e analépticas centrais de derivados sintéticos de aminoalquilxantonas. Uma
análise profunda desta atividade levou ao relato de um notável efeito estimulante sobre o
sistema nervoso central (SNC) e a inibição da enzima monoamina oxidase (MAO), uma
das mais importantes e açoes descritas destas classe de compostos. (Bhattacharya et al.,
1972; Suzuki et al., 1981; Deeb et al., 2007; Zuo et al., 2014). Outros estudos foram
descritos em 1972 evidenciando as xantonas como agentes bloqueadores β-adrenérgicos
(Dare, 1972), com atividade antitumoral in vivo (Schneider et al., 1972) e com
propriedades anti-alérgicas (Pfister et al., 1972). Ao final da década de 70, muito foi
alcançado para o reconhecimento das atividades biológicas destes compostos, com
estudos farmacológicos sendo realizados para as xantonas naturais e sintéticas.
Com o avanço da ciência e a melhoria das técnicas empregadas para extração de
produtos naturais e, também o aprimoramento da síntese de novos compostos, as
pesquisas envolvendo as atividades biológicas das xantonas expandiram (Pinto et al.,
2005). Em meados da primeira década do século XXI, o ácido dimetilxantenona-4-acético
ou DMXAA, análogo de xantona, foi testado clinicamente como agente antitumoral,
(A) (B)

14
apresentando-se como um potente indutor de interferon-β (IFN-β) (Roberts et al., 2007).
A atividade antitumoral de DMXAA foi associada à sua capacidade de induzir uma
variedade de mediadores inflamatórios, incluindo o fator de necrose tumoral (TNF), a IL-
10, a IL-6 e CCL5 (Jassar et al., 2005; Head, 2011). Apesar da promessa clínica, o
composto falhou nos testes em seres humanos, uma vez que o composto atuava
diretamente na via de sinalização celular STING (Baguley e Siemann, 2010; Prantner D,
2012). Após estes estudos, foi descoberto que esta via é diferente em humanos e
camundongos, não havendo correlação entre os resultados encontrados em modelo animal
e os testes clinicos em humanos. Entretanto a molécula é amplamente utilizada para
experimentação em modelo murino, uma vez que se conhece seu mecanismo de atuação
e (Daei Farshchi Adli et al., 2018; Conlon J e Monks B, 2103).
Comercialmente dois produtos contendo majoritariamente xantonas em sua
composição ganham destaque, o Vimang® e o Xango® (Garrido et al., 2001; Pardo-
Andreu et al., 2006). O Vimag®, extrato aquoso do caule de Mangifera indica L., planta
popularmente conhecida como mangueira, é constituído de mangiferina, polifenóis,
triterpenos, fitoesteróis, ácidos graxos e outros microelementos (Nunez A, 2002; Rivera
et al., 2011). Em Cuba, a suplementação com Vimang® é indicada para a redução do
stress oxidativo e melhoria da qualidade de vida dos pacientes (Pardo-Andreu et al.,
2006). Além do potencial antioxidante, o trabalho de Garrido et al. (2001), em modelo de
edema de pata em camundongos, demostrou que a injestão do Vimang® foi capaz de
reduzir a dor e a inflamação nestes animais, demostrando seu papel anti-inflamatório e
analgésico. O Xango® já é um produto que movimenta o mercado de bebidas funcionais
nos EUA e possui em sua composição a α-magostina. Contrariamente ao Vimag®,
estudos científicos não demonstraram sua eficacia em reduzir os parâmetros inflamatórios
em modelo animal e humano (Gutierrez-Orozco e Failla, 2013).
Os resultados emergentes da atividade farmacológica de compostos naturais
resultaram numa busca crescente pela otimização dos compostos líderes (Pinto et al.,
2005; Gong et al., 2017). Os métodos de extração tradicionais utilizados para obtenção
dos produtos naturais apresentam algumas desvantagens como o elevado tempo para a
extração com técnicas complexas e laboriosas, baixa seletividade e/ou baixos
rendimentos, além de empregar grandes quantidades de solventes tóxicos (Menendez et
al., 2017). Dessa forma, devido às importantes aplicações biológicas das xantonas, novas
estratégias sintéticas foram propostas e originaram distintos derivados com base nas

15
diferentes posições dos constituintes no esqueleto xantônico (Yang Chun-Hui, 2012;
Shagufta e Ahmad, 2016; Cidade et al., 2017). Isso permitiu aos pesquisadores
racionalizar e determinar as características estruturais essenciais para sua bioatividade
(Pedro et al., 2002; Shagufta e Ahmad, 2016). Shagufta e Ahmad (2016), em uma revisão
sobre as aplicações biológicas das xantonas sintéticas, enfatizaram que as modificações
moleculares como a inserção de grupos polares ou hidrofóbicos, além de outros
componentes, podem aumentar a eficiência dos compostos para suas atividades
antiproliferativas, antimicrobianas, antimaláricas, anti-HIV, anticonvulsivantes,
anticolinesterásicas, anti-oxidantes, anti-inflamatórias, além de aumenta a atividade
inibitória em diferentes enzimas.
Neste contexto surgem as xantenodionas, compostos heterocíclicos também
conhecidos como 1,8-dioxo-octa-hidroxantenos contendo um núcleo pirano fundido em
ambos os lados com anéis ciclohex-2-enona (Fig.2). A metodologia usada para gerar esta
molécula emprega técnicas caracterizadas como “green chemisty”, que permitem realizar
reações sob condições que não sejam prejudiciais ao ambiente, em um processo
ambientalmente aceitável (Sauer, 2000; Wender, 1997).
Além das vantagens de uma produção sustentável, as xantenodionas apresentam
distintas atividades biológicas descritas na literatura, que incluem propriedades
antibacterianas (Narasimha Murthy et al., 2014), antifungicas (Rahimifard et al., 2015),
leishmanicida (Muhammad Nisar, 2013), antimalárica (Winter et al., 1997), anti-
inflamatórias(Cidade et al., 2017) e antiproliferativa (Mulakayala et al., 2012). E, foi
pensando no viés inflamatório e em sua atividade em tripanosomatídeos (anti-
Leishmania), que elegemos as xantenodionas como a molécula alvo do nosso estudo.
Núcleo da Xantenodiona
Padrão de substituição comum
Figura 2 – Estrutura do núcleo das xantenodionas e seu padrão de distribuição

16
1.2 Potencial anti-inflamatório
O corpo humano é submetido a constantes danos que estimulam a resposta
inflamatória. Uma vez que a homeostase é perturbada, e em resposta a certos estímulos,
nosso corpo pode lançar um processo que define a inflamação, manifestada com sinais
macroscópicos cardinais (isto é, vermelhidão, inchaço, febre e dor) e sublinhado por eventos
microvasculares, celulares e moleculares que culminam no acúmulo de leucócitos (por
exemplo, granulócitos) nas áreas teciduais afetadas (Sugimoto et al., 2019)
O processo inflamatório é desencadeado através da ativação celular com consequente
liberação mediadores inflamatórios com efeitos autócrinos e parácrinos como o TNF, o IFN-
g, a IL-1b, IL-2, IL-6, dentre outros. Estes mediadores inflamatórios promovem a ativação
local de macrófagos, neutrófilos, células endoteliais e, com isso, aumentam a
permeabilidade vascular, facilitam a migração de células da resposta adaptativa para o local
do estímulo antigênico e possibilitam a ação resolutiva do processo inflamatório (Dinarello,
1996; 2007; Griffith et al., 2014). As quimiocinas são citocinas quimiotáticas que controlam
os padrões migratórios e o posicionamento das células imunes. A função da quimiocina é
fundamental para todo o movimento celular imunológico, desde a migração necessária para
o desenvolvimento e homeostase da célula imune, até a geração das respostas imunes celular
e humoral(Allavena et al., 1999; Bonecchi et al., 1999; Murdoch e Finn, 2000).
Mecanismos que regulam a resposta inflamatória são de suma importância para o
retorno à homeostase (Netea et al., 2017). A resolução da inflamação não é simplesmente a
eliminação do agente patogênico ou estressor, mas sim um processo ativo e autorregulado
que envolve reprogramação funcional de células por meio da produção de mediadores ad
hoc (Ouyang et al., 2011; Headland e Norling, 2015). Vários mediadores são capazes de
inibir a inflamação, dentre eles a IL-10, que suprime a produção de mediadores pró-
inflamatórios e é derivada principalmente de células T reguladoras (Treg) (Libby, 2007;
Dinarello et al., 2016). Os mediadores inflamatórios também podem ser neutralizados pela
interação célula-célula, desensibilização de receptores GPCRs e/ou por vias de sinalização
intracelular que dirigem a inflamação para uma regulação negativa (Headland e Norling,
2015). O fator de transcrição nuclear κB (NF-κB) regula muitos processos celulares,
incluindo produção de mediadores inflamatórios, proliferação e sobrevivência celular,
diferenciação de células T efetoras e reguladoras e maturação de células dendríticas (Arthur
e Ley, 2013; Afonina et al., 2017; Zhang et al., 2017).

17
Xantonas naturais e sintéticas são amplamente descritas na literatura por
apresentarem atividade anti-inflamatória induzindo ou bloqueando a expressão/produção de
mediadores inflamatórios. Sua ação é descrita em distintos tipos celulares in vitro e in vivo,
por diferentes mecanismos de ação (Pinto et al., 2005; Vieira e Kijjoa, 2005; Cidade et al.,
2017). Além disso podem alterar vias de sinalização celular relacionadas a atividade
inflamatória, a exemplo da sinalização das vias cinases ativadas por mitógenos, MAPK,
JNK, ERK, p38 e NF-kB (Pal et al., 2013; Jeong et al., 2014; Saha et al., 2016)
Nos modelos in vitro de inflamação induzida por lipopolissacarídeos (LPS) xantonas
e seus derivados foram capazes de alterar a expressão e a produção de citocinas(Deshpande
et al., 1997; Jeong et al., 2009; Tewtrakul et al., 2009; Bumrungpert et al., 2010) . A
mangostina, em particular, já foi descrita por regular negativamente a expressão de TNF, IL-
6 e IL-4, em macrófagos e adipócitos humanos (Bumrungpert et al., 2010; Liu et al., 2012;
Ding et al., 2018), além de atuar nas vias de sinalização celular das MAPK, STAT1, c-Fos,
c-Jun, EIK-1 e NF-kb (Lopaczynski e Zeisel, 2001; Liu et al., 2012; Ding et al., 2018). As
xantonas também possuem atividades antioxidantes, pois são unidades importantes de
doação de elétrons (Boots et al., 2002). Esta ação antioxidante desempenha um papel
preventivo em algumas patologias por meio da remoção de espécies reativas de oxigênio
(ROS), responsável por danos irreversíveis aos componentes celulares (Nakatani et al.,
2002; Chen et al., 2008; Tewtrakul et al., 2009).
O ativador do receptor do ligante nuclear fator kappa (RANKL) é a principal citocina
para a formação e ativação de osteoclastos (Lacey et al., 1998). A interação entre RankL e
o seu receptor é responsável pela sinalização intracelular envolvendo proteínas distintas
como o NF-kB, as proteínas quinases ativadas por mitógeno AKT, o fator nuclear de células
T ativadas (NFAT) e a cinase dependente de calmodulina (Ang et al., 2011). Estudos
mecanísticos revelaram que a mangiferina inibe a ativação do NF-kB induzida por RankL,
concomitante com a inibição da degradação de IkB-a e a translocação nuclear de p65. Além
disso, a mangiferina também inibe a fosforilação de ERK e JNK (quinase N-terminal c-Jun),
p38 e fosforilação de Akt (Ma et al., 2015; Fu et al., 2018).
A tabela 1, sumariza os resultados da experimentação in vivo, onde reforçam a
atividade anti-inflamatória das xantonas em diferentes modelos animais (Imran et al., 2017).

18
Tabela 1 – Atividade anti-inflamatória in vivo de xantonas e seus derivados
Modelo
Resultados Referência
Edema de pata induzido por
carregenina
Redução do diâmetro do edema de pata
(Lin et al., 1996;
Librowski et al.,
2005)
Colite induzida por TNBS
(2,3,4-trinitrobenzeno
sulfônico)
Redução do infiltrado inflamatório e
aumento da expressão de IL-10 (Jeong et al., 2014)
Asma induzida por albumina
(de ovo)
Redução da inflamação nos vasos e
brônquios, inibição das citocinas com perfil
Th2 e proliferação de linfócitos
(Rivera et al., 2011)
Artrite induzida por
adjuvante
Redução da produção de TNF, IL1-b, IFN e
IL-10 (Pal et al., 2013)
Toxicidade hepática induzida
por D-galactosamina
Inibição de NfKb em macrófagos
Redução de TGO/AST e TGP/ALT (Dar et al., 2005)
Em seres humanos, Rassameemasmaung et al. (2008) mostraram que a aplicação
tópica de um gel contendo mangostina reduziu a inflamação periodontal enquanto Udani et
al. (2009) e Tang et al. (2009) demonstraram que a ingestão de suco de mangostão diminuiu
os níveis séricos da proteína C reativa e reduziu os marcadores inflamatórios (CXCL10 e
CCL4) em indivíduos obesos. As xantonas também são capazes de inibir a formação de
radicais livres e das citocinas envolvidas no processo inflamatório da acne juvenil
(Chomnawang et al., 2007). Além disso, vale ressaltar a importância do DMXAA, derivado
xantônico, já testado clinicamente, com potencial antiproliferativo para células tumorais
(Roberts et al., 2007; Head, 2011; Prantner D, 2012; Daei Farshchi Adli et al., 2018).

19
Apesar dos estudos relatados anteriormente, a aplicação das xantonas naturais ou
sintéticas em doenças com alto índice de morbidade e/mortalidade despertaria interesse das
indústrias farmacêuticas e dos órgãos gerenciadores de saúde pública. As doenças
cardiovasculares estariam na lista alvo para as xantonas uma vez que previamente foi
demonstrado o papel cardioprotetor advindo do efeito anti-inflamatório destes compostos
(Jiang, D.-J. et al., 2004). Neste contexto, o próximo tema desta revisão enfatizará o papel
cardioprotetor de xantonas naturais e sintéticas.
1.3 Ação cardioprotetora das xantonas
A resposta inflamatória apresenta papel crucial na formação e perpetuação das lesões
cardíacas de diferentes etiologias. Estas inflamações nas células cardíacas têm sido
associadas à ativação coordenada do sistema complemento, do aumento da expressão de
citocinas e moléculas de adesão, recrutamento de neutrófilos e consequentemente aumento
da permeabilidade vascular (Frangogiannis, 2014). O efeito protetor das xantonas no sistema
cardiovascular pode ser devido a uma combinação de fatores como atividades antioxidante,
anti-inflamatória, inibitória da agregação plaquetária, antitrombótica e / ou vasodilatadora
(Jiang, D.-J. et al., 2004).
Foi demonstrado que a norathyriol, uma xantona isolada de Tripterospermum
lanceolatum, atenua o dano ao miocárdio aumentando a permeabilidade das células
endoteliais cardíacas e a “explosão respiratória” em neutrófilos. Esse fenômeno é induzido
por agonistas inflamatórios que inibem a ativação da proteína quinase C e da fosfolipase C
(Hsu et al., 1997; Lee et al., 1998). Além disso, estes compostos possuem ação direta na
liberação de creatina kinase (CK) e na redução dos níveis de TNF no miocárdio, em modelos
in vitro e in vivo, sugerindo que os efeitos protetores estão relacionados a redução da
atividade anti-inflamatória (Madan et al., 2002; Dai et al., 2004).
Estes compostos são também capazes de inibir o influxo de Ca+ por intermédio dos
canais de Ca+ receptores-operativos e voltagem-dependentes, ou por aumento nos níveis de
monofosfato cíclico de adenosina (AMPc). Além disso, as xantonas aminoalcanólicas ainda
demostraram atividade hipotensora por ação dependente de canal de cálcio e pelo bloqueio
dos receptores b-adrenérgicos (Rapacz et al., 2014). Estes efeitos das xantonas e de seus
derivados em receptores adrenérgicos podem prevenir arritmias de forma mais eficiente que
o medicamento de referência Urapidil® (Marona et al., 2008; Szkaradek et al., 2015).

20
A busca por xantenodionas que atuem com efeito cardioprotetor em função da
atividade anti-inflamatória despertou o interesse do nosso grupo de pesquisa. O T. cruzi,
causador da doença de Chagas, induz lesões no miocárdio, principalmente, pelo
desequilíbrio da resposta inflamatória do hospedeiro. A seguir compreenderemos os aspectos
inter-relacionados entre o efeito antiparasitário das xantonas, sua ação anti-inflamatória
conectada ao seu efeito cardioprotetor na infecção pelo T. cruzi.
1.4 - Atividade antiparasitária das xantonas
Infecções por protozoários são problemas emergentes de saúde pública mundial.
Suramina, pentamidina, melarsoprol, efloritina e benznidazol constituem drogas utilizadas
no tratamento contra alguns agentes infecciosos, principalmente em países de clima tropical
(Theel e Pritt, 2016). No entanto, a toxicidade e o potencial de desenvolvimento de
resistência a estes compostos enfatizam a necessidade de medicamentos mais eficazes e
seguros para a população (Dua et al., 2009; Scarim et al., 2018). Recentemente, estudos
destacaram as atividades antiparasitárias das xantonas naturais ou sintéticas, em protozoários
e helmintos, atuando como principal agente parasiticida ou em sinergia com outros
medicamentos já utilizados na clínica, a exemplo da malária (Mauss, 1948; Nabih, 1966;
Dua et al., 2009; Aberham et al., 2011; Keiser et al., 2012)
É notória a ação das xantonas contra parasitos do gênero Plasmodium sp, agente
etiológico da malária (Pontius et al., 2008; Al-Massarani et al., 2013). Os compostos ativos
são extraídos de diversas fonte naturais, como espécies de fungos (Pontius et al., 2008) e
vegetais (Al-Massarani et al., 2013) além de compostos sintéticos (Winter et al., 1997;
Ignatushchenko Mv, 2000; Xu Kelly et al., 2001; Kelly et al., 2002). Sua ação malaricida
deve-se, possivelmente, a ação antioxidante das xantonas que, ao reduzir a produção de
radicais livres inibe a polimerização do grupo heme, fundamental para a sobrevivência deste
parasito (Ignatushchenko Mv, 2000; Xu Kelly et al., 2001; Zarena e Sankar, 2009). Sugere-
se, ainda, que modificações no posicionamento de grupos aminas protonáveis das
hidroxixantonas são capazes de aumentar a afinidade para o grupo heme livre e melhorar,
assim, as propriedades antimaláricas do composto (Winter et al., 1997). Estes compostos
apresentam ação sinérgica em associação com a artemisina, fármaco de escolha no
tratamento da malária (Tjahjani, 2017).

21
Existem poucos relatos da ação de xantonas, xantenos e xantenodionas com
propriedades anti-tripanosomastídeos e, os disponíveis, advêm principalmente de modelos
in vitro. Em geral, essa moléculas apresentam razoável atividade citotóxica e podem ganhar
ou perder radicais afim de aumentar sua eficácia antiparasitária (Abe et al., 2003; Abe et al.,
2004; Dua et al., 2009). Esta estratégia foi usada por Nisar et al. (2013) mostrando que a
natureza não citotóxica de análogos de 1,8-dioxo-octa-hidroxanteno fornece uma rota mais
segura para estudos biológicos com atividade leishmanicida contra promastigotas de
Leishmania major. Xantonas naturais extraídas de Andrographis paniculata (Dua et al.,
2009) e G.mangostana também apresentaram ação anti-leishmania (Al-Massarani et al.,
2013).
Apenas xantonas oriundas de fontes naturais foram testadas contra o T. cruzi e a
maior parte dos compostos testados possuem baixos índices de seletividade (Al-Massarani
et al., 2013). Estes compostos apresentaram atividade contra epimastigotas axênicos,
tripomastigotas de cultura celular e formas amastigotas, intracelulares (Abe et al., 2003; Abe
et al., 2004; Pontius et al., 2008)
Os trabalhos descritos anteriormente relatam apenas a atividade dos compostos
contra o T.cruzi, sem revelar quais são os eventos responsáveis por esta atividade. Entretanto,
em 2013, um grupo de pesquisadores brasileiros demonstrou o potencial anti-T. cruzi da
1,3,7-tri-hidroxi-2-(3-metilbut-2-enil)-xantona, isolada da Kielmeyera coriácea, árvore
nativa do cerrado e popularmente conhecida como “pau santo”. Este composto foi capaz de
gerar alterações ultraestruturais e morfológicas no parasito, sendo a mitocôndria seu alvo
principal (De Oliveira Caleare et al., 2013).
As perspectivas sobre o conhecimento e a identificação de novas moléculas é uma
área que proporciona grande avanço econômico e farmacológico (Scarim et al., 2018;
Osorio-Méndez e Cevallos, 2019). O impacto causado por doenças infecciosas assume
relevância mundial porque atinge a população, provocando graves problemas à saúde
pública (Rodriques Coura e De Castro, 2002; Dias et al., 2009). Dessa forma, existe a
necessidade de novas alternativas terapêuticas para o tratamento, principalmente para
doenças que são poucos estudadas ou mesmo negligenciadas farmacologicamente, como é
caso da doença de Chagas, impulsionando a busca por novos compostos com atividade
biológica (Osorio-Méndez e Cevallos, 2019).

22
1.5 O T.cruzi como um indutor do processo inflamatório
A presença do T. cruzi no organismo de mamíferos induz a uma complexa interação
entre suas moléculas e células de defesa do hospedeiro e, desta interação, emerge uma
resposta inflamatória sistêmica e focal no intuito de contenção da replicação parasitária
(Rocha E Silva, 1978; Marin-Neto et al., 2007; Talvani e Teixeira, 2011). No entanto, a não
eliminação destes protozoários promove a cronificação e a persistência da resposta
inflamatória podendo gerar lesões teciduais, perdas funcionais de alguns órgãos e até a morte
do hospedeiro (Teixeira et al., 2002; Gazzinelli, 2004). A intensa replicação deste protozoário
durante a fase aguda da infecção ativa uma resposta inflamatória inicial coordenada por células do
sistema mononuclear fagocitário (ex. macrófagos) que respondem à presença de antígenos do T. cruzi
através de uma via dependente de receptores do tipo Toll (TLR)-Myd88 (Bafica et al., 2006). Estes
receptores, ao se ligarem a proteínas (glicosilfosfadilinositol) presentes na superfície do parasito,
formam âncoras, conhecidas por GPI-mucinas (Cardoso et al., 2015), conduzindo, principalmente, à
ativação de uma resposta imune do tipo TH1, secretando mediadores inflamatórios como citocinas
(IL-12, TNF, IFN- gama), quimiocinas (CCL2, CCL5), óxido nítrico (NO), dentre outros, que além
de eliminarem o agente agressor, e modularem o curso da doença, atuam como indicadores
quantitativos de processos biológicos ou patológicos empregados para fins de diagnóstico (Pinazo et
al., 2015; Guedes et al., 2016). A produção do IFN-gama, é realizada tanto por células da resposta
imune inata (ex. células NK, Natural killer cells) quanto por células T, que em conjunto com o TNF,
ativam células do sistema fagocitário mononuclear, a produzirem o óxido nítrico (NO), molécula
efetora, responsável por controlar a replicação intracelular desse protozoário (Gutierrez et al., 2009);
além de acionarem quimiocinas (CCL2-MCP1, CCL5-RANTES), que ativam e recrutam novas
células para o local da infecção (Teixeira et al., 2002). Uma vez que o excesso de células pró-
inflamatórias em determinado tecido por um tempo prolongado pode ser prejudicial ao hospedeiro,
torna-se essencial a participação de citocinas regulatórias, como a IL-10, responsáveis por inibir e
controlar a resposta imune mediante a infecção por esse protozoário (Dutra et al., 1997). A constância
desses eventos inflamatórios, ao longo do tempo, conduz à prevalência desse parasito em diferentes
tecidos do hospedeiro mamífero, especialmente em tecidos musculares. Em particular, o tecido
muscular cardíaco representa um dos sítios mais importantes desta interação parasito-hospedeiro.
Dito isso, a infiltração maciça de células inflamatórias, seja devido à permanência do estímulo ou à
ausência de mecanismos regulatórios, embora priorize a eliminação do protozoário, inevitavelmente,
conduz à destruição de células hospedeiras e de plexos simpáticos e parassimpáticos, essenciais para
o funcionamento autonômico cardíaco. Estas ações em conjunto, culminam em uma nova
reorganização estrutural do coração, afetando negativamente sua funcionalidade, e assim,

23
conduzindo cerca de 20- 30% dos pacientes sintomáticos nesta condição, a um quadro de cardiopatia
chagásica crônica (CCC)(Prata, 2001; Marin-Neto et al., 2007; Talvani e Teixeira, 2011; Malik et
al., 2015). Nestes indivíduos, já foi demonstrada maior produção/expressão de citocinas e
quimiocinas pró- inflamatórias como TNF e CCL2, e a presença dessas, estão correlacionadas com
o comprometimento do coração (Dutra et.al., 1997 Cunha-Neto et al., 2009). Por outro lado, em
indivíduos crônicos, com ausência de manifestações cardíacas e digestórias, já foi demonstrado uma
prevalência de citocinas regulatórias circulantes, como a IL-10, mostrando que, em parte, o perfil
dos mediadores inflamatórios parece definir o quadro clínico na fase crônica dos indivíduos
infectados pelo T. cruzi (Dutra et al., 2014). No entanto, a intensidade da resposta inflamatória
mediante a infecção pelo T. cruzi, pode decorrer de diferentes fatores como, por exemplo, a cepa do
parasito, a via de infecção, além da genética e da resposta imune do hospedeiro vertebrado, capaz de
direcionar o curso de evolução da doença (Machado, 2013; Magalhães et al., 2015). Dito isso, torna-
se importante o desenvolvimento/utilização de fármacos, visando não apenas a destruição do
parasito, mas também o equilíbrio entre a interação parasito/hospedeiro.
1.6 Estratégias terapêuticas na doença de Chagas
Atualmente há dois fármacos disponíveis para o tratamento da doença de Chagas: o
nitrofurano Nifurtimox (Nfx; Lampit®, Bayer) e o nitroimidazol Benznidazol (Bz;
Lafepe®). Esses fármacos são compostos nitroheterocíclicos, pontuados pela existência de
um grupo nitro unido a um anel aromático. São incluídos em grupo de nitrofuranos e
antibióticos nitroimidazólicos que apresentam eficiência frente a um grande número de
infecções bacterianas e parasitárias (Wilkinson et al., 2008; Romanha et al., 2010).
No Brasil, o fármaco utilizado para o tratamento desta doença parasitária é o Bz desde
1978. A atuação do Bz parece ocorrer por meio de um mecanismo envolvendo a modificação
covalente de macromoléculas do parasito (como por exemplo, DNA, RNA, citocromo P450
e outras proteínas) provocada por radicais aniônicos gerados por nitroredução (Urbina e
Docampo, 2003) e resultando na inibição da síntese proteica. Este fármaco possui ação em
formas circulantes (tripomastigotas) e teciduais (amastigotas) do T. cruzi. Entretanto,
ocasiona indesejáveis reações aos usuários como reações de hipersensibilidade, dermatites,
toxicidade hepática, intolerância digestiva, edemas e depressão da medula óssea que leva,
frequentemente, ao abandono do tratamento (Salomon, 2012)

24
A eficácia dos dois agentes quimioterápicos de uso atual (Nfx e Bz) para o tratamento
da doença de Chagas é altamente variável e muitas vezes limitada, especialmente durante a
fase crônica da infecção (Viotti et al., 1994; Tarleton, Rick L., 2003; Urbina, 2010). Ambas
requerem longos períodos de administração e têm efeitos colaterais significativos (Rodriques
Coura e De Castro, 2002). Além disso, cepas resistentes também são responsáveis pelas
falhas terapêuticas (Filardi e Brener, 1987; Chatelain, 2015) Recentemente, formas do
parasita resistentes ao tratamento farmacológico prolongado in vivo e in vitro foram
identificadas e permanecem capazes de restabelecer a infecção após um dia de exposição à
droga (Sanchez-Valdez et al., 2018). Assim, há uma necessidade urgente de
descobrir/desenvolver novos alvos terapêuticos bem como drogas mais seguras, eficazes e
de baixo custo (Lombardino e Lowe Iii, 2004; Chatelain e Konar, 2015).
Existe uma demanda contínua por novas terapias para doenças tropicais, incluindo a
doença de Chagas. A falta de consenso em relação aos protocolos de triagem in vitro e in
vivo tem dificultado a descoberta de novas drogas para o tratamento da doença de Chagas
(Romanha et al., 2010).
Apesar do potencial dos fármacos utilizados na rotina clínica contra o T. cruzi, o
desenvolvimento de novos compostos com atividade tripanocida ainda é incipiente.
Atualmente, o tratamento com Bz (LAFEPE®) possui eficácia questionável, além de ter
atividade mutagênica e carcinogênica(Caldas et al., 2008). Por este motivo, novos
compostos vêm sendo testadas para o tratamento da tripanossomíase americana. Dessa
forma, a identificação de um composto derivado de xantenodionas que elimine o parasito
circulante e atue modulando a resposta inflamatória no hospedeiro, forneceria uma estrutura
química com potencial capacidade de tornar-se um fármaco. Além disso, a geração de
produtos com subsídios para contribuir nas ações públicas voltadas para o tratamento e
profilaxia de doenças tropicais, traria enorme benefício para uma parte da população latino-
americana, ainda parcialmente negligenciada farmacologicamente. Por estas razões as
xantenodionas tornaram-se alvo de investigação nacional e internacional nas últimas
décadas, por suas propriedades-alvo microbiocidas e anti-inflamatórias. Dessa forma, o
objetivo deste trabalho foi avaliar o efeito anti-inflamatório e anti-T.cruzi de derivados da
1,8-dioxo-octa-hidroxantenos.

25
2. OBJETIVOS
2.1 Objetivo geral
Avaliar as atividades tripanocida e anti-inflamatória de compostos derivados dos
1,8-dioxo-octa-hidroxantenos.
2.2 Objetivos específicos
In vitro
Avaliar a ação tripanocida das xantenodionas em formas triposmatigotas e
amastigotas;
Verificar a citotoxicidade dos compostos em macrófagos J774A.1 e células Vero;
Determinar o índice de seletividade;
Selecionar, dentre os 12 compostos avaliados, o mais promissor para prosseguir na
etapa in vivo.
In vivo
Padronizar a dose a ser administrada capaz de reduzir a parasitemia sem gerar a
mortalidade dos animais;
Avaliar a capacidade do composto em atuar na parasitemia em camundongos Swiss
infectados pela cepa Y e CL do T. cruzi;
Avaliar a influência do tratamento com o composto mencionado nos parâmetros
biológicos dos animais;
Quantificar a resposta inflamatória no tecido muscular cardíaco, renal e hepático dos
camundongos infectados pelo T. cruzi, após o tratamento com o composto.
Avaliar a produção de marcadores regulatórios e inflamatórios no miocárdio dos
animais;
Verificar a toxicidade hepática e renal nos animais tratados com a xantenodiona
selecionada .

26
3. MATERIAL E MÉTODOS
a. Representação esquemática do delineamento experimental
Figura 3 – Representação esquemática dos métodos realizados para estudo da atividade das
xanthenodionas. (A) Experimentos in vitro. (B) Ensaios in vivo de padronização da dose e via
de tratamento(n=5). (C) Avaliação do tratamento com a MI80 na redução da parasitemia e do
potencial anti-inflamatório (n=10). CC50, concentração citotóxica de 50%; IC50, concentração
inibitória de 50%, DPI, dias após a infecção.
3.1 Compostos
Os compostos utilizados neste trabalho foram cedidos pelo pesquisador Prof. Dr. Róbson
Ricardo Teixeira, do departamento de Química da Universidade Federal de Viçosa. A
estratégia geral de síntese para construção dessa biblioteca foi a derivação química das
xantonas, composto natural supracitados, utilizando-se o núcleo1,2,3-triazólico como
espaçador entre a molécula base e um grupamento aromático substituído, da qual deriva a

27
diversidade química da biblioteca de compostos da UFV. A estrutura do núcleo da
xantenodiona utilizado como grupo substituinte é mostrado na figura 4. A estrutura dos
compostos avaliadas neste estudo, está representada na tabela 2. Cada composto recebeu um
código aleatório, sendo eles: MI17, MI18, MI24, MI25, MI39, MI45, MI68, MI73, MI78,
MI79, MI80 e MI81.
Figura 4 - Formula estrutural do núcleo xantenodiona. Esqueleto utilizado para construção das
respectivas séries químicas

28
Tabela 2 - Estrutura molecular representativa das xantenodionas. Fórmula estrutural evidenciando
radicais e suas posições no núcleo das xantenodionas avaliadas neste estudo.
3.2 Parasito
Duas cepas do T. cruzi com perfis variáveis de susceptibilidade ao Bz foram
utilizadas neste estudo. A cepa Y classificada como Discrete Typing Units II (DTU II) sendo
parcialmente resistente ao fármaco e a cepa CL, DTU VI, susceptível ao Bz em modelo
murino (Zingales et al., 2009). A cepa CL foi escolhida nos experimento in vitro por permitir
a inserção de plasmídeos fluorescentes em seu material genético e ser descrita como
prioritária no protocolo de screnning de novos fármacos para a Doença de Chagas in vivo

29
(Xu et al., 2009; Canavaci et al., 2010; Chatelain e Konar, 2015). Já cepa Y foi selecionada
por apresentar características biológicas como a indução de alta parasitemia e 100% de
mortalidade na infecção murina, facilitando, neste caso, a avaliação da atividade tripanocida
com diferentes doses do composto (Filardi e Brener, 1987).
3.3 Culturas Celulares
Os macrófagos J774 foram cultivados em meio DMEM 1640 (GIBCO®)
suplementado com 10% de soro fetal bovino (SBF), L-glutamina 2mM,
Penicilina/Streptomicina 100U, 1mM de Piruvato de sódio (Sigma), aminoácidos não
essenciais, pH7.2 e mantidos em estufa 5% CO2, a 37 °C. Os repiques foram realizados a
cada 72h, quando a cultura apresentava 80% de confluência. As células foram ressuspensas
por raspagem suave utilizando “cell scraper”. Em seguida, as células foram coletadas,
quantificadas e utilizadas para novos repiques e ensaios biológicos.
As células da linhagem Vero foram cultivadas em meio RPMI suplementado com
5% de SFB, 2mM de L-glutamina, 100U de Penicilina/Streptomicina, pH 7.2 e mantidas em
estufa 5% CO2 a 37ºC. Os repiques foram realizados a cada 72h, quando a cultura
apresentava 80% de confluência. As células foram ressuspensas por tripsinização e, em
seguida, coletadas, quantificadas e utilizadas para novos repiques e ensaios biológicos.
3.4 Obtenção das formas tripomastigotas de cultura celular
As formas tripomastigotas da cepa Y utilizadas para infectar células foram obtidas a
partir do sobrenadante de cultura de células infectadas. Para isso, células Vero foram
cultivadas em meio RPMI suplementado com 5% de SFB. Ao atingir cerca de 70% de
confluência, a cultura foi infectada, na proporção de 10 parasitos por célula, com
tripomastigotas sanguíneos oriundos do sangue de camundongos infectados pela cepa Y no
pico de parasitemia, que corresponde ao 8o dia de infecção. Após 24 horas de incubação a
37º C e 5% CO2, o meio de cultura foi removido e os parasitos não internalizados, bem como
detritos celulares, foram retirados através de sucessivas lavagens com meio. Às células, foi
adicionando meio RPMI suplementado com 1% de SFB e, novamente, incubadas a 37º C
por sete dias, quando o sobrenadante foi coletado e centrifugado, a 1100rpm, 24º C, por 10
minutos para remoção dos debris celulares. Posteriormente, o sobrenadante rico em parasitos
foi centrifugado a 3000 rpm a 4º C por10 minutos e o sedimento ressuspendido em meio de
cultura para posterior utilização nos experimentos. Apenas parasitos recém liberados das

30
células foram utilizados nos experimentos, uma vez que o sobrenadante era periodicamente
coletado e um novo meio de cultura adicionado.
3.5 Modelo animal
Os animais foram fornecidos e mantidos pelo Centro de Controle Animal (CCA) da
Universidade Federal de Ouro Preto. Este trabalho foi aprovado pelo Comitê de Ética de Uso
em Experimentação Animal com o número 2016-34. Foram utilizados camundongos Swiss,
fêmeas, com idade variando entre 30 a 35 dias, pesando entre 25 e 30 g.
3.6 Experimentos In vitro
3.6.1 Verificação da atividade tripanocida - Ensaio de citotoxidade
em formas tripomastigotas derivadas de cultura celular
Para avaliar a citotoxidade dos compostos em formas tripomastigotas de cultura
celular do T. cruzi (cepa Y) foi adicionado uma suspensão 1x106 parasitos a cada poço (placa
de 96 poços) e incubados em oito concentrações diferentes (5, 10, 20, 50, 80, 100, 200 e 300
μM/mL) dos compostos MI17, MI24, MI25, MI79, MI80 e MI81 por 24 horas. Após este
período, foram adicionados aos poços 20 μL (2mg/mL) de MTT 3-(4,5-di-metilazol-2-il) -
2,5-difeniltetrazolium brometo tetrazol (Sigma-Aldrich) por 2 horas nas mesmas condições
de cultivo conforme descrito por Mosmann (1983). Após a reação, os parasitos foram
centrifugados a 1000 g, por 10 minutos e o sobrenadante foi descartado sendo o sedimento
resuspenso em DMSO (Sigma-Aldrich) para completa dissolução dos cristais. Em seguida,
a absorbância foi determinada em 530 nm em leitor de placas VICTOR™ X3 (Perkin Elmer).
O percentual de parasitos viáveis foi calculado por meio da seguinte fórmula:
% Viabilidade = (Absorbância do tratado/Absorbância do controle) x 100
Os valores obtidos com esses cálculos foram plotados no programa GraphPadPrism
versão 6.0 para a determinação da IC50.

31
3.6.2 Avaliação do efeito tripanocida em formas amastigotas
3.6.2.1 Geração de parasitos modificados – T. cruzi fluorescentes
(tdTomato)
Os experimentos para geração de parasitos modificados, bem como os ensaios de
citotoxidade em células Vero foram realizados Na Universidade da Geórgia(EUA) sob
orientação do Dr.Rick L Tarleton durante o período de doutorado sanduiche. Para gerar
parasitas expressando em tandem uma proteína dimérica tomate vermelho fluorescente foi
construído um plasmídeo usando o vetor de expressão pTrex como suporte. O gene
tdTomato (1464 pb) foi gerado por amplificação via PCR usando o conjunto de primers 5’-
AGAATTCATGGCGCCTAGGGTGAGC-3’ (senso) e 5’-
TACGTCGACTTAGAGCTCGATATCGACG -3’ (anti-senso), e o vetor pCTR2t como
modelo. O primer senso possui um sitio EcoRI e o primer anti-senso um sitio SalI. O
fragmento de PCR foi digerido e clonado a jusante do promotor do gene rRNA e da fração
HX1 dentro dos locais de multi-clonagem do plasmídeo pTREX, gerando o plasmídeo
pTREX-tdTomato (Fig.5). Os parasitos transgênicos foram gerados por knockout como
descrito previamente por Xu et al. (2009) e Canavaci et al. (2010). Culturas de epimastigotas
da cepa CL na fase log de crescimento foram centrifugadas a 1,620 g por 15 min e suspensos
em 100 µl de solução de Nucleofector TM Solution de células T humanas (Lonza, Alemanha)
à temperatura ambiente. Os parasitos foram ressuspensos e, então, misturados com 10 µg de
DNA oriundos do produto da pcr descrita acima, em volume total de 10 µl e eletroporados
usando o programa "U33" no dispositivo Nucleofector AMAXA (Lonza, Alemanha). Os
parasitos eletroporados foram então cultivados em garrafas de cultura de 25 cm2 (Corning
Incorporated, Lowell, MA) com 10 ml de meio LDNT (Triptose neutralizada e digerida de
fígado) e adicionou-se 300 mg / ml de antibiótico G418 24 h após a transfecção. As culturas
de parasitos foram monitoradas por citometria de fluxo frequentemente para verificar a
intensidade de fluorescência. Quando necessário, as culturas eram “sorteadas” para remover
contaminantes não fluorescentes usando o citómetro MoFlo (DakoCytomation, Carpinteria,
CA) com um laser Enterprise 631 488 nm para excitação e um filtro de emissão com uma
banda de passagem 570/40 nm (Fig.6) Este protocolo garantiu a geração da cepa CL
tdTomato utilizada nos experimentos de “drug screnning”.

32
Figura 5 - Plasmídeo utilizados na geração de T. cruzi fluorescente. Representação
esquemática do plasmídeo pTrex-Neo-tdTomato.
Figura 6 - Avaliação da fluorescência do T. cruzi (cepa CL) tdtomato. (A) Imagem microscópica
dos parasitas expressando tdTomato em todas as fases evolutivas. (B) Intensidade de fluorescência
em epimastigotas em citômetro de fluxo CyAn (DakoCytomation) e analisada com o software
FlowJo (Tree Star). A fluorescência de fundo dos parasitas selvagens (verde) também pode ser
observada. Fonte: Canavaci et al, 2010

33
3.6.2.2 Avaliação do potencial tripanocida em amastigotas
Os parasitos fluorescentes descritos no item 3.6.2.1 foram utilizados neste
experimento para avaliar a capacidade das xantenodionas (MI17, MI18, MI24, MI25, MI39,
MI45, MI68, MI73, MI78, MI79, MI80 e MI81) em eliminar o T. cruzi. As células Vero
foram expostas a 2000 rad de radiação gama durante dez minutos (Yan e Moreno, 1998) e
2,5x104 células foram adicionadas em placas de 96 poços Black Greiner (Bio-One). Após a
confirmação da aderência e formação da monocamada, formas tripomastigotas da cepa
CLtdTomato do T. cruzi foram adicionas durante aproximadamente 5 h na proporção de 10
parasitos para 1 célula. Após a infecção, as culturas celulares foram lavadas para remover
parasitas não internalizados e meios frescos contendo os compostos ou Bz (como o fármaco
tripanocida de referência) em concentrações decrescentes (50 a 0,25µM/mL) foram
adicionados. A mudança na intensidade de fluorescência foi determinada como uma medida
de crescimento ao longo de 3 dias de cultura. Os valores de concentração inibitória de 50%
(IC50) foram determinados por análise de regressão linear no dia 3. Esse percentual de
inibição foi utilizado para a construção de curvas de dose-efeito e cálculo da IC50 com
auxílio dos software GraphPadPrism6. Todos os experimentos realizados com as formas
amastigotas foram feitos em duplicatas, em no mínimo 3 repetições,
3.6.3 Ensaios de citotoxidade em culturas de células – determinação
da CC50
Os compostos foram diluídos em dimetilsulfoxido (DMSO-Sigma-Aldrich) a 0,01%
e posteriormente preparada uma solução estoque a 1000µM/mL que, em seguida, foi
sonicada em Cuba Ultrassônica CD-4820 (Kondentech) por dois minutos. Logo após, a
solução foi adicionada ao meio de cultura utilizado para os experimentos in vitro.
Duas linhagens celulares foram testadas nesta etapa, os macrófagos J774A.1 e as
células Vero. Também foram testadas duas metodologias para verificação da viabilidade
celular, a do MTT® e a do AlamarBue®.

34
3.6.3.1 Macrófagos murinos-MTT®
O ensaio de MTT® foi utilizado nos experimentos com macrófagos J774A.1. As
células foram cultivadas em meio DMEM completo suplementado e ajustadas para um
volume de 1x105 células por poço em um volume final de 150µL. As placas foram incubadas
24h em estufa a 37°C e atmosfera de 5% de CO2. Após o período de incubação as células
foram observadas em microscópio invertido para verificação do aspecto e aderência celular.
Os poços foram lavados com meio para retiradas de células mortas e, posteriormente, os
compostos adicionados em oito concentrações 5, 10, 20, 50, 80, 100, 200 e 300 µM/mL. As
culturas permaneceram expostas aos compostos durante 48h horas, incubadas a 37ºC com
atmosfera de 5% de CO2.
Após os períodos de incubação, foram adicionados aos poços 20μL da solução MTT
a 20mg/mL (50μL/poço). As placas foram novamente incubadas por 2 horas a 37º C com
atmosfera de 5% de CO2. Após incubação, o sobrenadante foi removido e adicionou-se
150μL de DMSO para solubilizar os cristais de formazan formados. Posteriormente, as
placas foram submetidas à leitura em espectrofotômetro em comprimento de onda de 490nm
em leitor de placas VICTOR™ X3 (Perkin Elmer). O valor final da densidade ótica de cada
poço foi subtraído do valor da densidade ótica de uma triplicata de poços contendo apenas
DMSO. Os resultados foram expressos em gráficos de porcentagem, sendo os valores do
eixo y relativos ao controle, considerado como apresentando 100% de viabilidade. A
viabilidade das linhagens celulares com diferentes concentrações dos compostos foi
conduzida de acordo com Mosmann (1983).
Com os resultados obtidos calculou-se a percentagem de inibição através da fórmula
citada no item 3.6.2 e os valores plotados no programa GraphPadPrism versão 6.0 para a
determinação da CC50 e posterior cálculo do índice de seletividade.
3.6.3.2 Células Vero- AlamarBlue®
Para excluir os efeitos tóxicos das xantenodionas sobre as células hospedeiras,
células Vero não infectadas por T. cruzi, foram tratadas com oito concentrações decrescentes
(100-0.39μM/μL) dos compostos. O experimento foi realizado utilizando o corante Alamar
Blue® (Bio-Rad), um indicador colorimétrico de proliferação celular. Esse método se baseia

35
na redução química do reagente devido à atividade metabólica celular. Através de reações
de oxirredução, o corante passa da cor azul (forma oxidada e não fluorescente) para rosa
(forma reduzida, fluorescente) e esta mudança de cor pode ser medida pela leitura
colorimétrica ou fluorimétrica (Rampersad, 2012)
As células Vero foram expostas a 2000 rad de radiação gama durante dez minutos
(Yan e Moreno, 1998) e posteriormente, cem microlitros destas células, na concentração de
2.5x104/poço, foram adicionadas às placas Black Greiner (Bio-One) e mantidas em estufa
CO2 a 37° C por pelo menos uma hora para completa adesão. Os compostos foram diluídos
em DMSO, a partir de uma solução estoque de 5mM/μL, em concentrações crescentes (8
diluições 1:2 iniciando em 100μM/μL). Ressalta-se que a concentração final de DMSO no
poço foi inferior a 0.01%. Após a adesão das células e formação da monocamada 100μL de
cada concentração foi adicionada aos poços em triplicata e a placa novamente incubada em
estufa CO2 a 37°C por 72h. Como controle de morte foi utilizado o fármaco
ANACOR14353® nas mesmas concentrações testadas para os compostos, sabidamente
tóxico para esta linhagem celular (dados não publicados). Cada placa continha controles
negativos (meio) e controles positivos (meio+célula). Após 72h de incubação, 50 μL do meio
de cultura foi removido para que 50 μL de Alamar Blue® 10% fosse adicionado. As placas
foram cobertas com papel alumínio e novamente incubadas em estufa CO2 a 37°C por 4h.
Após este período a fluorescência foi mensurada no leitor de placas SynergyH4 (BioTek) em
comprimento de 560nm e 590nm. A porcentagem de redução do corante induzida pelos
compostos foi calculada considerando o percentual de redução das células incubadas na
ausência de drogas, utilizando-se a fórmula abaixo:
% redução = [A560-(A590xR0) Tratado / A560-(A590xR0) Controle+]
Nessa fórmula, A560= fluorescência a 560nm, A590= fluorescência a 590nm.
Controle + é o poço contendo células, meio e Alamar Blue® na ausência do composto. R0
é o fator de correção, calculado a partir dos valores de emissão de fluorescência do controle
negativo (C-), ou seja, apenas meio de cultura e Alamar Blue® na ausência de células [Ro =
(A560 /A590)C-].
O composto mais eficiente (<toxicidade >atividade tripanocida) na fase in vitro foi
selecionado para os ensaios em modelo animal, conforme descritos nos itens metodológicos
a seguir.

36
3.7 Experimentos In vivo
3.7.1 Padronização – Experimento de dose-resposta
Para avaliar a via de tratamento e dosagem máxima do composto (mais eficiente nos
experimentos in vitro) e capaz de ser tolerado pelos animais e potencialmente reduzir ou
suprimir a parasitemia, foi realizado um experimento de dose-resposta. Os animais foram
inoculados intraperitonealmente com 5x103 formas tripomastigotas sanguíneas da cepa Y T.
cruzi e divididos em grupos de 5 animais (n=5).
Cada grupo recebeu o composto MI80, escolhido na triagem in vitro, em suspensão
numa solução de carboximetilcelulose (CMC) a 0,5% e Tween80 (0,5%)(Sigma®) . Foram
cinco dosagens testadas (1:2) de 800 a 50mg/kg por via oral (gavagem) e via intraperitoneal.
Os grupos controles utilizados para a comparação de eficácia do tratamento foram: grupo
não tratado (NI) e grupo não infectado e tratado (NIT). O tratamento foi iniciado
imediatamente após a detecção de parasitemia patente (4ºdia após a infecção) e administrado
por 10 dias consecutivos, uma vez ao dia, para os grupos contendo os animais infectados.
3.7.2 Infecção e esquema de tratamento
Realizada a padronização da dose e da via de tratamento foi realizado um
experimento para avaliar a eficácia do composto MI80 na redução da parasitemia e sua
capacidade em alterar parâmetros biológicos, imunológicos e bioquímicos nos animais
tratados. Dessa forma, camundongos Swiss fêmeas foram infectadas, intraperitonealmente,
com cinco mil formas tripomastigotas sanguíneas do T. cruzi. Nesta etapa, foram utilizadas
duas cepas, Y e Cl, sendo uma parcialmente resistente ao Bz e a outra susceptível ao mesmo
fármaco. Os animais foram divididos em grupos de dez animais (n=10) e, após confirmada
a parasitemia, foram tratados por via oral com 50mg/kg da xantenodiona MI80, uma vez ao
dia, por 10 dias. Após 15 dias de infecção para a cepa Y e 18 dias para a cepa Cl os animais
foram eutanasiados. Foram coletados o plasma, coração, fígado e rim para análises
histológicas, imunológicas e bioquímicas.
3.7.3 Exame de sangue a fresco - parasitemia
Para a realização deste procedimento, 5µL de sangue da artéria coccígea ventral foi
coletado de cada animal e depositdos sobre uma lâmina de microscópio coberta com uma
lamínula 22x22mm para determinação do número de parasitos em 50 campos microscópicos.

37
O número encontrado foi multiplicado pelo fator do microscópio, calculado para cada
objetiva, levando em consideração o número de campos microscópios (Brener, 1962).
3.7.4 Mortalidade
A taxa de mortalidade foi calculada pela observação diária dos animais durante o
tratamento até a necropsia. Os resultados foram expressos em porcentagens.
3.7.5 Histopatologia do coração
Após o término do tratamento, os animais foram eutanasiados e, por meio de incisão
torácica, o coraçãofoi removido, lavado em PBS, pesado e seccionados longitudinalmente
para análise histopatológica. Os tecidos foram imersos em formalina tamponada a 10% por
no mínimo 48 horas. As etapas seguites foram realizadas em colaboração com o professor
Rômulo Novaes da Universidade Federal de Lavras. As amostras foram embebidas em resina
histológica de metacrilato de glicol e cortadas em micrótomo rotativo em seções de 3-mm
de espessura utilizando lâminas de vidro (Leica Biosystems, Wetzlar, Alemanha). Cinco
cortes histológicos foram coletados em semi-séries, utilizando uma de cada 50 seções, as
quais foram coradas com hematoxilina e eosina. A análise histopatológica foi realizada
avaliando-se a distribuição e organização do parênquima e do estroma conjuntivo, necrose
tecidual e infiltrado inflamatório, morfologia e distribuição de miócitos, vasos sanguíneos e
células intersticiais e presença de ninhos de amastigotas de T. cruzi (Novaes et al., 2016)
A intensidade da celularidade ou inflamação do coração foi avaliada comparando-se
a distribuição de células intersticiais no miocárdio de todos os grupos (Novaes et al., 2013).
Para isso, a celularidade tecidual, por área histológica, foi determinada como o número de
núcleos de células intersticiais em uma área de teste padronizada (At = 25×103μm2). Foram
avaliados a partir de 20 campos histológicos amostrados aleatoriamente de secções cardíacas
de cada animal usando um microscópio de campo claro com uma lente objetiva × 40
(ampliação de 400x; Axioscope A1, Carl Zeiss, Alemanha). Uma área total de tecido de
25,0×105 μm2 foi analisada para cada grupo. A celularidade intersticial total foi analisada
pelo software de análise de imagens Image-Pro Plus® (versão 4.5; Media Cybernetics Inc.,
Silver Spring, MA, EUA). Os resultados foram expressos como um diagrama de campo
octogonal organizado em domínios que representam a celularidade tecidual da seguinte
forma: (-) celularidade mínima / (+ -) leve, (+ +) moderada ou (+ + +) intensa/infiltrado
inflamatório (Felizardo et al., 2018).

38
3.7.6 Estereologia do coração
A amplitude da reorganização microestrutural cardíaca foi analisada a partir do método
estereológico (Novaes et al., 2013). A densidade volumétrica (Vv,%) do parênquima
cardíaco (cardiomiócitos [cmy]), estroma (tecido conjuntivo [cnt]) e vasos sanguíneos (bvs)
foi estimada como Vv = ΣPt / P; onde ΣPt é o número de pontos de teste que ativam a
estrutura de interesse e P é o número total de pontos no sistema de teste. Usamos um sistema
de teste com 72 pontos em uma área de teste padrão de (At = 25 × 103 μm2) ao nível do
tecido. A relação entre vasos sanguíneos e cardiomiócitos (Vv [bvs] / Vv [cmy]) foi utilizada
como índice morfológico de vascularização do miocárdio. A relação entre compartimentos
estruturais e funcionais do coração foi estimada como Vv [cnt] / Vv [cmy] (Novaes et al.,
2013). A densidade do volume foi estimada a partir de 20 campos histológicos amostrados
aleatoriamente de secções cardíacas de cada animal usando uma objetiva 40 × (ampliação
de 400x; Axioscope A1, Carl Zeiss, Alemanha). Uma área total de tecido de 25,0 × 105 μm2
foi analisada para cada grupo.
A densidade numérica de células intersticiais mononucleares (MN) e
polimorfonucleares (PMN) por área histológica foi estimada como QA = ΣQ / At; onde ΣQ
é o número de MN ou PMN no plano focal microscópico e At é a dimensão do teste (At =
8,56 × 103 μm2). A densidade do volume foi estimada a partir de 20 campos histológicos
amostrados aleatoriamente de secções cardíacas de cada animal usando uma objetiva × 100
(aumento de 1000x; Axioscope A1, Carl Zeiss, Alemanha) (Novaes et al., 2013). Uma área
total de tecido de 85,6 × 104 μm2 foi analisada para cada grupo. Todas as contagens foram
realizadas usando o software de análise de imagem AxionVision (Carl Zeiss, Alemanha).
3.7.7 Produção de citocinas
As citocinas IFN-γ, CCL2, IL-10 e RankL foram quantificadas no sobrenadante do
macerado do tecido cardíaco dos animais por meio de do teste imunoenzimatico ELISA
sanduíche (PeproTech, NJ, EUA) seguindo recomendaçoes do fabricante. Microplacas de
96 poços foram adsorvidas com anticorpo de captura monoclonal anti IFN-γ,CCL2, IL-10
ou RANKL na concentração adequada para cada citocina, em salina tamponadade fosfatos
(PBS), pH7,4 (100 µL/poço). As placas foram incubadas por 12h a temperatura ambiente .
Após a incubação as placas foram lavadas quatro vezes com PBS, contendo 0,05% de Tween-
20 (solução de lavagem) e posteriormente bloqueadas com 300 µL por poço com PBS

39
contendo 1% de SFB (solução de bloqueio) à temperatura ambiente por 1h. Em seguida as
placas foram lavadas quatro vezes com a solução de lavagem. Após lavagem, foram
adicionados a cada poço, 50µL do padrão de citocinas ou do sobrenadante do macerado do
tecido cardíaco. As placas foram incubadas a temperatura ambiente por 2h e lavadas quatro
vezes com a solução de lavagem. Em seguida foi adicionado 100µl por poço do anticorpo
de detecção (anticorpo anti-citocina de camundongo marcado com biotina) diluído na
concentração recomendada pelo fabricante em PBS. As placas foram incubadas à
temperatura ambiente por 12h e lavadas quatro vezes com a solução de lavagem, sendo então
adicionados 100µL/poço do conjugado avidina-HRP na diluição de 1:2000 em PBS
contendo 0.05% de Tween-20 e 0,1% de SFB (solução diluente). As placas foram incubadas
novamente em temperatura ambiente por 30 min. O cromógeno escolhido para revelação foi
o 2,2#-Azino-bis (ácido 3-etilbenzotiazolina-6-sulfonico) (ABTS – Sigma Aldrich,
Missouri, USA) e 100µL/poço deste cromógeno adicionados por vinte minutos de
incubação, em ausência de luz e temperatura ambiente. Interrompida a reação, a leitura da
intensidade de coloração foi realizada em leitor de ELISA utilizando-se o comprimento de
onda de 405nm, com correção de 650nm. A quantificação da quimiocina e citocinas
presentes nas amostras foi determinada baseada na densidade óptica obtida com a curva
padrão de concentração conhecida destes mediadores, analisadas pelo software SOFTmax
PRO 4.0.
3.7.8 Testes bioquímicos
Os testes bioquímicos descritos abaixo foram adquiridos e seguiram as
recomendações do fabricante - Bioclin®
3.7.8.1 Transaminase oxalacética (AST/TGO)
Para a dosagem da enzima transaminase oxalacética (AST/TGO) utilizou-se o
método colorimétrico de Reitman S (1957) que se baseia na reação catalisada pela AST/TGO
do grupamento amino de um alfa-aminoácido para um alfa-cetoácido segundo a reação a
seguir:
L-Aspartato + Alfa-Cetoglutarato L-Glutamato + Oxalacetato
Junto à reação foi adicionado um composto com propriedades corantes
(dinitrofenilhidrazina) que reage com o oxaloacetato fomando hidrazina. A intensidade de

40
cor foi diretamente proporcional à produção de oxaloacetato, que por sua vez, representa a
atividade enzimática. Foram utilizadas três soluções neste protocolo, a primeira contendo
PBS, ácido L-aspártico, ácido alfa cetoglutárico e azida sódica. A segunda solução continha
o reagente corante possuindo dinitrofenilhidrazina e ácido clorídrico e a terceira solução de
hidróxido de sódio. Para inicialização do procedimento, 250µl da primeira solução foi
incubada a 37°C por 3 minutos, em seguida foi adicionado 100µl da amostra de plasma e
novamente incubado por mais 30 minutos a 37°C. Adicionou-se então 250µl da segunda
solução e o sistema foi deixado em repouso por 20 minutos e finalmente adicionado 2,5ml
do hidróxido de sódio. Após 5 minutos a 37°C o conteúdo foi transferido para uma cubeta
de quartzo e lido em espectrofotômetro com comprimento de onda 505nm. O cálculo final
foi conseguido a partir dos valores de diluição de uma curva padrão pré-determinada pelo
kit.
3.7.8.2 Transaminsae pirúvica (ALT/TGP)
O método colorimétrico para a dosagem a enzima transaminase pirúvica (ALT/TGP)
é semelhante ao descrito para a AST/TGO (Reitman S, 1957). Entretanto com a alteração do
L-aspartato pela L-alanina e do oxaloacetado pelo piruvato.
L-Alanina + Alfa-Cetoglutarato L-Glutamato + Piruvato
Junto à reação foi adicionado um composto com propriedades corantes
(dinitrofenilhidrazina) que reage com o piruvato fomando hidrazina. A intensidade de cor
foi diretamente proporcional à produção de piruvato, que por sua vez, representa a atividade
enzimática. Foram utilizadas três soluções neste protocolo, a primeira contendo PBS, ácido
L-alanina, ácido alfa cetoglutárico e azida sódica. A segunda solução continha o reagente
corante possuindo dinitrofenilhidrazina e ácido clorídrico e a terceira solução de hidróxido
de sódio. Para inicialização do procedimento, 250µl da primeira solução foi incubada a 37°C
por 3 minutos, em seguida foi adicionado 100µl da amostra de plasma e novamente incubado
por mais 30 minutos a 37°C. Adicionou-se então 250µl da segunda solução e o sistema foi
deixado em repouso por 20 minutos e finalmente adicionado 2,5ml do hidróxido de sódio.
Após 5 minutos a 37°C o conteúdo foi transferido para uma cubeta de quartzo e lido em
espectrofotômetro com comprimento de onda 505nm. O cálculo final foi conseguido a partir
dos valores de diluição de uma curva padrão pré-determinada pelo kit.

41
3.7.8.3 Creatinina
Para a detecção da creatinina em plasma dos animais foi utilizado o método de Jaffe
(1986) que consiste em um método colorimétrico onde a creatinina reage com o ácido pícrico
formando uma solução com coloração amarelada. Com a adição de um reagente ácido,
ocorre uma redução do pH e consequentemente da intensidade de cor devido a reversão da
reação que formou creatinina-picrato. Pela diferença das leituras obtidas, alcança-se o valor
da creatinina. Para realização do método, utilizou-se um reagente alcalino contendo
hidróxido de sódio, carbonato de sódio e surfactante, uma solução é de ácido pícrico e um
reagente ácido contendo ácido acético. Em um tubo foi adicionado 2,0mL do reagente
alcalino com 250µL da amostra de plasma e 500µL da solução de ácido pícrico. Após 10
minutos em banho-maria a 37°C, as absorbâncias foram lidas utilizando-se uma cubeta de
quartzo e espectrofotômetro em comprimento de onda de 510nm. Em seguida foi adicionado
100µl do reagente ácido e aguardou-se 5 minutos à temperatura ambiente para realização da
nova leitura em comprimento de onda de 510nm. Para se chegar ao valor final de creatinina
foi utilizado o seguinte cálculo:
Creatinina (mg/dL ) = Leitura 1- Leitura 2 x 3/absorbância do padrão
3.7.8.4 Ureia enzimática
Para determinação dos níveis de uréia no plasma dos animais foi utilizado um teste
enzimático colorimétrico onde a uréia foi hidrolisada a iôns amônio e CO2 baseando-se na
seguinte reação:
Uréia + 3 H2O 2NH4+ + CO2 + OH-
A reação continha uma solução enzimática contendo urease, PBS, nitroprussiato de
sódio, salicilato de sódio, estabilizantes, conservantes, além de hidróxido de sódio e
hipoclorito de sódio. Então 1mL da solução enzimática foi acrescentado a 10µl da amostra
e levado ao banho-maria a 37°C por 5 minutos. Em seguida foi adicionado 1ml da solução
oxidante e novamente incubado por 5 minutos a 37°C em banho-maria. A leitura então foi
aferida utilizando cubeta de quartzo e espectrofotômetro com o comprimento de onda de
600nm. Para se chegar ao valor final de uréia foi utilizado o seguinte cálculo:
urease

42
Uréia (mg/dL) =Absorbância da amostra / absorbância do padrão
3.8 Análise estatística
Todos os dados foram submetidos ao teste de normalidade Kolmogorov–Smirnov.
Parâmetros com distribuição normal foram analisados pelo teste de análise de variância
univariada (ANOVA-one way) e o teste de Bonferroni utilizado posteriormente para
determinar as diferenças entre os grupos. Para as variáveis que não apresentaram distribuição
normal, realizou-se o teste de Kruskal-Wallis, seguido pelo teste de Dunn’s para
determinação das diferenças entre grupos. Os dados foram expressos como média ± erro
padrão e as diferenças consideradas significantes para p<0,05. Todas as análises foram
realizadas utilizando o software GraphPadPrism versão 6.0 para Windows (San Diego,
California, USA).

43
4. RESULTADOS
4.1 - Resultados in vitro
4.1.1 Avaliação da toxicidade em formas tripomastigotas do T.cruzi
e determinação do IC50
Com o objetivo de avaliar a toxicidade dos derivados de xantenodionas sob formas
tripomastigotas do T. cruzi, foi realizado um ensaio de MTT e o cálculo da curva de
concentração-resposta para determinação da IC50.
Todas as seis xantenodionas testadas em tripomastigotas apresentaram, em diferentes
níveis, atividade tripanocida de maneira dose-dependente. A viabilidade dos parasitos foi
reduzida significativamente (p<0,05) em relação ao controle não tratado, nas concentrações
maiores que 100µM/mL para os compostos MI80 e MI81. Para o composto MI17 essa
redução foi notada nas concentrações maiores que 50µM/mL enquanto para os compostos
MI79, MI24 e MI25 este padrão foi observado nas concentrações acima de 10µM/mL
(Fig.7). Os controles do ensaio consistiram em poços contendo parasitas não tratados com
as xantenodionas e poços contendo DMSO 0,01% (v/v) evidenciando a não toxicidade deste
diluente, além de poços contendo o Bz como controle de alta atividade tripanocida
Na determinação da IC50, destacaram-se os compostos MI17, MI80 e MI81 com
valores de IC50 de 69,21µM, 87,76µM e 137,8µM, respectivamente. Outros três derivados
de xantenodionas testados, MI24, MI25 e MI79 apresentaram concentração inibitória
parasitária, porém menos significativa que os compostos citados inicialmente (Tabela 3).

44
Figura 7 – Análise do efeito citotóxico em formas tripomastigotas T. cruzi (cepaY), pelo
método do MTT. 1x106 foram adicionados por poço, em placa de 96 poços e os compostos
MI17, MI24, MI25, MI79, MI80 ou MI81 foram nas concentrações de 5 a 300µM/mL e
incubados por 24h. Os resultados foram expressos como valores em porcentagem do número de
parasitos viáveis após 24 horas de incubação com os derivados de xantenodionas. Análise de
variância (ANOVA) e pos-teste de Tukey foram aplicadas para determinar a diferença estatística
entre os tratamentos em relação ao controle DMSO. (*) representa diferença estatística
determinada por p<0,05.

45
4.1.2 Efeito dos compostos na viabilidade celular demacrófagos
murinos J774A.1)avaliados por ensaios colorimétricos
Para avaliar a viabilidade celular em presença das seis xantenodionas (MI17, MI24,
MI25, MI79, MI80 e MI8), testamos sua toxicidade em macrófagos murinos. Após 48h de
exposição aos compostos, a molécula MI81 mesmo na concentração de 300µM/mL,
apresentou toxicidade negligenciável nesta linhagem celular, com 85,22% de células viáveis.
Estes resultados foram comparáveis ao composto MI17 que também manteve um padrão de
células viáveis superior a 70% na maior concentração. Entretanto seus análogos, MI79 e
MI80, na dosagem superior a 200µM/mL, reduziram a viabilidade celular em 30%. Também
os compostos MI24 e MI25 foram citotóxicos nas concentrações de 100µM/mL e 50µM/mL,
respectivamente. A viabilidade também foi avaliada na presença de DMSO 0,01% (v/v)
mostrando que o diluente, nesta concentração, não foi citotóxico às células (Fig.8). Os
valores de 50% da concentração citotóxica (CC50) estão expressos na tabela 3 e foram
utilizados para o cálculo do índice de seletividade.

46
Observou-se que o composto MI81 obteve o melhor índice de seletividade
Figura 8 – Análise de viabilidade celular de culturas de macrófagos murinos J774A.1
avaliadas pelo método do MTT frente aos derivados de xantenodionas. 1x105 celulas foram
adicionada aos poços em placa de 96 poços.Após a formação da monocamada os compostos
MI17, MI24, MI25, MI79, MI80 ou MI81 foram adicionados em concentrações decrescentes de
5 a 300µM/mL e as placas incubadas por 48h em estufa 37ºC 5% CO2.Após 48h foi realizado o
ensaio de MTT e os resultados foram expressos em porcentagem do número de células viáveis
após 48 horas de incubação com os derivados de xantenodionas .

47
(67,12µM), seguido pelo MI80 (5,37µM) e pelo MI17 (4,3µM). Apesar dos demais
compostos apresentarem ação tripanocida contra formas tripomastigotas, os índices de
seletividade, os quais expressam quantas vezes o composto é mais tóxico para o parasito em
relação as células de mamíferos, foram baixos, conforme evidenciados na Tabela 3.
Tabela 3 – Citotoxidade em macrófagos murinos J774, atividade tripanocida em formas
tripomastigotas do T.cruzi e índice de seletividade de compostos derivados de xantenodionas
Composto
Células J774A.1
CC50(µM)
Tripomastigotas(Y)
IC50(µM)
IS
MI17 303 69.25 4.3
MI24 79,82 199,6 ND
MI25 232.9 209,7 1.1
MI79 >300 >300 1
MI80 >300 87.70 5.37
MI81 >300 136.4 67.12
*Bz >300 22.69 15.51
CC50 = concentração citotóxica de 50% / IC50-concentração inibitória de 50% / IS- índice
de seletividade, razão entre CC50 e IC50 / Bz -Benznidazol (*Usado como fármaco controle)
4.1.3 Atividade tripanocida em formas amastigotas do T.cruzi (cepa
CLtdTomato)
Nesta etapa, utilizou-se um controle de células infectadas com o T. cruzi e não
tratadas e um controle de células infectadas e tratadas com Bz para comparação dos
resultados. Dos doze compostos testados, apenas três apresentaram ação sob as formas
amastigotas da cepa CLtdtomato do T. cruzi (Fig.9 e Fig.10), sendo eles o MI25 com IC50
de 14.11µM, seguido do MI24 (IC50=26.43) e do MI80 (IC50=30,65) (Tabela 4). Foi
calculado, mais uma vez, o IS dos compostos empregando o valor do CC50 das células vero

48
e o IC50 sob as formas amastigotas. O composto MI80, apesar de não ter o menor IC50
dentre as xantenodionas testadas, obteve o melhor IS, principalmente devido a sua
baixíssima ação sob a viabilidade celular (Fig.10). Por esta razão, o composto MI80 foi
selecionado para os ensaios in vivo que avaliarão sua potencial ação tripanocida em animais
infectados por cepas sensíveis e parcialmente resistentes ao Bz, além de sua ação anti-
inflamatória.
M I2 4
L o g C o n c .(M )
% o
f M
ax
ima
l G
row
th
1 0 0 .0 1 0 0 .5 1 0 1 .0 1 0 1 .5 1 0 2 .0
0
2 0
4 0
6 0
8 0
1 0 0
M I2 5
L o g C o n c .(M )
% o
f M
ax
ima
l G
row
th
1 0 0 .0 1 0 0 .5 1 0 1 .0 1 0 1 .5 1 0 2 .0
0
2 0
4 0
6 0
8 0
1 0 0
M I8 0
L o g C o n c .(M )
% o
f M
ax
ima
l G
row
th
1 0 0 .0 1 0 0 .5 1 0 1 .0 1 0 1 .5 1 0 2 .0
0
2 0
4 0
6 0
8 0
1 0 0
Figura 9 - Atividade das xantenodionas MI24, MI25 e MI80 sobre a infecção de células Vero
pela cepaCLtdtomato do T. cruzi. 2,5x104 células foram infectadas pela cepa CLtdtomato e,
posteriormente, incubadas na presença ou ausência de concentrações decrescentes de xantenodionas
MI24, MI25 e MI80, por 72 horas. Cada ponto das curvas de dose-resposta corresponde à média de
2 experimentos independentes, em triplicata. O cálculo de IC50 foi realizado utilizando-se o software
GraphPadPrism6.

49
Figura 10 - Fotomicrografias representativas dos efeitos anti-T.cruzi dos compostos MI24,
MI25 e MI80. Estão representadas fotomicrografias de células Vero infectadas com cepa
CLtdTomato sem tratamento e tratadas com as xantenodiona MI24, MI25 e MI80 ou com o
fármaco benznidazol (Bz) ao terceiro dia após a infecção in vitro.
Campo Claro Proteína Fluorescente
Vermelha Sobreposição
MI24
Bz
MI25
MI80
Não tratado

50
4.1.4 Efeito sobre a viabilidade celular em células Vero por ensaios
fluorimétricos e seu respectivo índice de seletividade
Foram avaliadas a toxicidade dos 12 compostos derivados de xantenodionas, onde
seis foram testados no experimento anterior (MI17, MI24, MI25, MI79, MI80 e MI81) e sete
ainda não haviam sido testados (MI18, MI39, MI45, MI49, MI68, MI73 e MI78). Realizou-
se o ensaio fluorimétrico AlamarBlue® e, as células Vero plaqueadas e incubadas com os
12 compostos, por 72h, em concentrações seriadas de 100 a 0,39µM/mL.
Dos doze compostos testados, cinco não foram tóxicos em nenhuma concentração,
sendo eles os análogos MI79, MI80 e MI81, além dos MI73 e MI39 (Fig.11). Já para os
compostos MI17, MI24, MI25, MI49 e MI45 a viabilidade celular foi reduzida em 30% ou
mais nas concentrações maiores que 50µM/mL, sendo considerados citotóxicos (Fig.12). Os
demais derivados de xantenodionas MI18, MI68 e MI78 foram tóxicos mesmo em baixas
concentrações, o que inviabilizou a utilização dessas moléculas nestes
experimentos(Fig.13).

51
Figura 11 – Análise da viabilidade celular em culturas de células Vero pelo método do
AlamarBlue. 2,5x104 foram plaqueadas e incubadas em estufa 37ºC, 5%CO2 por 72h com os
derivados de xantenodionas MI79, MI80, MI81, MI73 e MI39 em concentrações decrescentes de 100
a 0,39µM/mL. As barras representam os valores em porcentagem do número de células viáveis de
três experimentos independentes em triplicata e erro padrão (SE).

52
Figura 12 - Viabilidade celular em culturas de células Vero pelo método do AlamarBlue,
determinado após 72 horas de incubação com os derivados de xantenodionas MI17, MI24, MI25,
MI45 e MI49 em diferentes concentrações. As barras representam os valores em porcentagem do
número de células viáveis de três experimentos independentes em triplicata e erro padrão (SE).

53
M I1 8
C o n c e n t r a ç ã o (M )
% d
e c
élu
las
Ve
ro v
iáv
eis
Dm
so
0,3
9
0,7
8
1,5
6
3,1
2
6,2
5
12,5
025
50
100
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
M I6 8
C o n c e n t r a ç ã o (M )
% d
e c
élu
las
Ve
ro v
iáv
eis
Dm
so
0,3
9
0,7
8
1,5
6
3,1
2
6,2
5
12,5
025
50
100
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
M I7 8
C o n c e n t r a ç ã o (M )
% d
e c
élu
las
Ve
ro v
iáv
eis
Dm
so
0,3
9
0,7
8
1,5
6
3,1
2
6,2
5
12,5
025
50
100
0
5
2 0
4 0
6 0
8 0
1 0 0
1 2 0
Figura 13 - Viabilidade celular em culturas de células Vero pelo método do AlamarBlue,
determinado após 72 horas de incubação com os derivados de xantenodionas MI18, MI68 e MI78
em diferentes concentrações. As barras representam os valores em porcentagem do número de células
viáveis de três experimentos independentes em triplicata e erro padrão (SE).
O composto MI80, apesar de não ter o menor IC50 dentre as xantenodionas testadas,
obteve o melhor IS, principalmente devido a sua baixa ação sob a viabilidade celular
(Fig.10). Por esta razão, o composto MI80 foi selecionado para os ensaios in vivo que
avaliaram sua potencial ação tripanocida em animais infectados por cepas sensíveis e
parcialmente resistentes ao Bz, além de sua potencial ação anti-inflamatória.

54
Tabela 4 - Citotoxidade em células Vero, atividade tripanocida em formas amastigotas do T.
cruzi (cepa CL) e o índice de seletividade de compostos derivados de xantenodionas
Composto
Células Vero
CC50(µM)
Amastigotas-CL
IC50(µM)
SI
MI17 >100 Inativo ND
MI18 27,8 Inativo ND
MI24 81,47 26,43 3,08
MI25 >100 14,11 7.29
MI39 >100 Inativo ND
MI45 43,58 Inativo ND
MI49 42,38 Inativo ND
MI68 29,69 Inativo ND
MI73 >100 Inativo ND
MI78 2,66 Inativo ND
MI79 >100 Inativo ND
MI80 >100 30,65 170
MI81 >100 Inativo ND
*Bz >100 3.4 103
CC50-concentração citotóxica de 50%. IC50-concentração inibitória de 50%. IS - índice de
seletividade, razão entre CC50 e IC50. Bz-Benznidazol (*Usado como fármaco controle)

55
4.2 Resultados In vivo
4.2.1 Padronização da dose e via de tratamento
Para dar início aos experimentos em modelo animal, foi realizado um experimento
de padronização das doses do composto MI80 e da respectiva via para sua administração..
O composto MI80, administrado a partir do quarto dia de infecção, mostrou efeito
tripanocida em todas as doses (800 a 50mg/kg) e rotas testadas (oral e intraperitoneal),
quando comparada ao controle infectado e ao não tratado (Fig.14). Por via oral observou-se
redução na parasitemia (p<0,05) para os grupos tratados com 400mg/kg e 800mg/kg em
relação ao grupo infectado e não tratado. Enquanto para a rota intraperitoneal, houve
diminuição da parasitemia apenas para a dose de 800mg/kg. Ainda verificou-se que o
tratamento com Bz foi capaz de suprimir a parasitemia durante todo o período de
experimentação.

56
V ia o ra l
D ia s a p ó s a in fe c ç ã o
Pa
ras
ito
s e
m 0
.1m
L d
e s
an
gu
ex
10
3
1 2 3 4 5 6 7 8 910
11
12
13
14
15
16
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
5 0 m g /kg
1 0 0 m g /kg
2 0 0 m g /kg
4 0 0 m g /kg
8 0 0 m g /kg
C o n tro le N T
B z - 1 0 0 m g
*
V ia in tra p e rito n e a l
D ia s a p ó s a in fe c ç ã o
Pa
ras
ito
s e
m 0
.1m
L d
e s
an
gu
ex
10
3
1 2 3 4 5 6 7 8 910
11
12
13
14
15
16
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
*
*
Figura 14 - Curva de parasitemia. Camundongos Swiss foram infectados com 5.000 formas
tripomastigotas sanguíneas da cepa Y do T. cruzi (n=5) e tratados ou não com a xantenodiona
MI80 ou Benznidazol (Bz). Os dados em cada ponto da curva equivalem à média da
parasitemia/dia referente a cada grupo. O símbolo (*) indica p<0,05 em relação ao grupo controle
infectado e não tratado.O gráfico superior indica a parasitemia durante o tratamento por via oral
e o gráfico inferior por via intraperitoneal.

57
Apesar da redução da parasitemia nos animais que receberam a MI80 por via
peritonial, foi observada uma acentuada taxa de mortalidade, inviabilizando esta via em
experimentos futuros (Fig.15). Dessa forma, o tratamento por gavagem foi priorizado na
dose de 50mg/kg, uma vez que esta concentração foi capaz de reduzir em 60% o número de
parasitos no sangue periférico durante o pico de parasitemia e ainda manter 80% dos animais
vivos até o fim do experimento.
V ia O ra l
D ia s a p ó s a in fe c ç ã o
% d
e S
ob
rev
ivê
nc
ia
0 5 1 0 1 5
0
2 0
4 0
6 0
8 0
1 0 0
V ia In tr a p e r ito n e a l
D ia s a p ó s a in fe c ç ã o
% d
e S
ob
rev
ivê
nc
ia
0 5 1 0 1 5
0
2 0
4 0
6 0
8 0
1 0 0
5 0 m g /k g 1 0 0 m g /k g l 2 0 0 m g /k g 4 0 0 m g /k g 8 0 0 m g /k g N ã o tra ta d o
Figura 15 - Curvas de letalidade dos animais infectados e tratados com a MI80 por via oral ou
intraperitoneal. Animais Swiss fêmeas foram infectados com a cepa Y do T. cruzi, tratados ou não
com MI80 em doses decrescentes de 800 a 50mg/kg ou BZ (100mg/kg) (n=5). Curvas referentes ao
período compreendido entre o 1º e o 15 º dia de infecção. O gráfico a direita representa a taxa de
mortalidade dos animais tratados por via oral e o gráfico da esquerda a mortalidade dos camundongos
tratados intraperitonealmente.
4.3 Avaliação do tratamento com o composto MI80 na dose de
50mg/kg por via oral
4.3.1 Parasitemia e mortalidade
Nos animais infectados com a cepa Y, o tratamento foi rapaz de reduzir em 40% a
quantidade de parasitos circulantes no sangue periférico no pico da infecção. Para os animais
infectados com a cepa CL do T. cruzi, não foi observada redução da parasitemia quando
comparada ao grupo dos animais infectado e não tratados. Diferenças estatísticas (p<0,05)
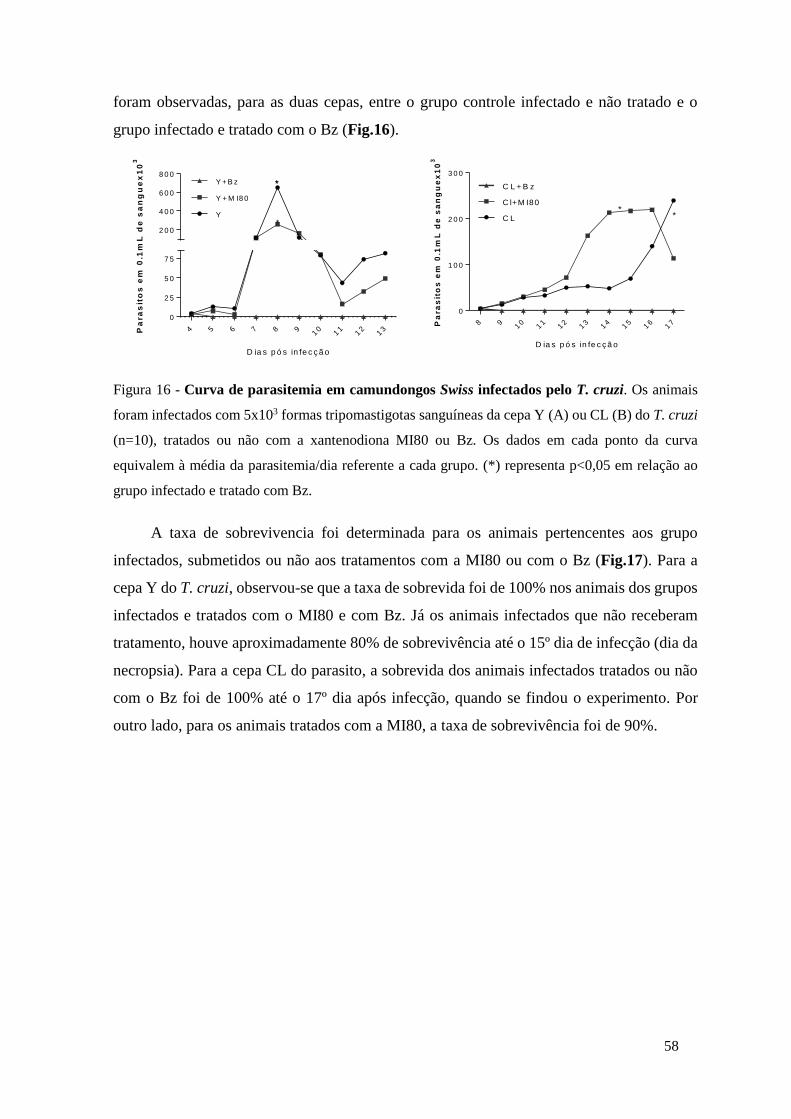
58
foram observadas, para as duas cepas, entre o grupo controle infectado e não tratado e o
grupo infectado e tratado com o Bz (Fig.16).
Figura 16 - Curva de parasitemia em camundongos Swiss infectados pelo T. cruzi. Os animais
foram infectados com 5x103 formas tripomastigotas sanguíneas da cepa Y (A) ou CL (B) do T. cruzi
(n=10), tratados ou não com a xantenodiona MI80 ou Bz. Os dados em cada ponto da curva
equivalem à média da parasitemia/dia referente a cada grupo. (*) representa p<0,05 em relação ao
grupo infectado e tratado com Bz.
A taxa de sobrevivencia foi determinada para os animais pertencentes aos grupo
infectados, submetidos ou não aos tratamentos com a MI80 ou com o Bz (Fig.17). Para a
cepa Y do T. cruzi, observou-se que a taxa de sobrevida foi de 100% nos animais dos grupos
infectados e tratados com o MI80 e com Bz. Já os animais infectados que não receberam
tratamento, houve aproximadamente 80% de sobrevivência até o 15º dia de infecção (dia da
necropsia). Para a cepa CL do parasito, a sobrevida dos animais infectados tratados ou não
com o Bz foi de 100% até o 17º dia após infecção, quando se findou o experimento. Por
outro lado, para os animais tratados com a MI80, a taxa de sobrevivência foi de 90%.
D ia s p ó s in fe c ç ã o
Pa
ra
sit
os
em
0.1
mL
de
sa
ng
ue
x1
03
8 910
11
12
13
14
15
16
17
0
1 0 0
2 0 0
3 0 0
C L
C l+M I80
C L + B z
**
D ia s p ó s in fe c ç ã o
Pa
ra
sit
os
em
0.1
mL
de
sa
ng
ue
x1
03
4 5 6 7 8 910
11
12
13
0
2 5
5 0
7 5
2 0 0
4 0 0
6 0 0
8 0 0
Y
Y + M I8 0
Y + B z **
*

59
Figura 17 - Curvas de letalidade de camundongos Swiss infectados com 5x103 formas
tripomastigotas sanguíneas da cepa Y ou CL do T. cruzi, tratados ou não com a MI80 ou Bz.
Animais Swiss fêmeas foram infectados com a cepa Y ou CL do T. cruzi, tratados ou não com
MI80(50mg/kg) ou BZ (100mg/kg) (n=10). Curvas referentes ao período compreendido entre o 1º e
o 15 º dia de infecção para cepa Y e 1º ao 18º para a cepa CL. O gráfico a direita representa a taxa
de mortalidade dos animais tratados para cepa CL o gráfico da esquerda a mortalidade dos
camundongos infectados coma cepa Y.Os resultados representam porcentagem de animais vivos.
4.3.2 Massa corporal e massa relativa do coração, baço e fígado
Observou-se a massa corporal nos animais infectados pela cepa Y, não tratados e
tratados com o MI80, foram maiores em relação aos animais não infectados (NI) e não
infectados tratados com MI80 (NIT) (Fig.18A). Nos animais infectados com a cepa CL, não
houve diferença significativa na massa corporal entre os grupos. Observamos que os animais
infectados com a cepa CL do T. cruzi tratados com o MI80 tiveram a massa relativa do
coração maior em relação aos controles NI e NIT (Fig.18B). Verificamos, também, que o
baço encontrava-se aumentado em todos os grupos infectados com a cepa Y do T. cruzi e
nem o tratamento com o MI80 nem com o Bz foram capazes de reverter este quadro. Para a
cepa CL do parasito, o tratamento com o MI80 não foi capaz de reverter o aumento da massa
esplênica, porém o Bz exerceu um papel protetor e preveniu o aumento deste órgão nos
animais infectados (Fig.18C). Também referente ao aumento relativo da massa hepática, o
MI80 não foi capaz de reverter o aumento observado com a presença do T. cruzi, sendo esta
redução observada apenas nos animais tratados com o Bz (Fig.19D).
D ia s a p ó s a in fe c ç ã o
% d
e S
ob
re
viv
ên
cia
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8
6 0
7 0
8 0
9 0
1 0 0
C l
C l+M I80
C l+ B z
NIT
NI
D ia s a p ó s a in fe c ç ã o
% d
e S
ob
re
viv
ên
cia
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5
6 0
8 0
1 0 0
Y
Y +M I80
Y + B z
NIT
NI

60
Ma
ss
a c
orp
ora
l (g
)
Y
Y+X
td
Y+B
z
CL
CL+X
td
CL+B
zN
IT NI
2 0
2 5
3 0
3 5
4 0
4 5
5 0
*_ _ _ _ _ _ _ _
#
Ma
ss
a r
ela
tiv
a
do
co
ra
çã
o (
mg
)
Y
Y+M
I80
Y+B
z
CL
CL+M
I80
CL+B
z
NIT N
I
0 .0 0 0
0 .0 0 5
0 .0 1 0
0 .0 1 5
*#
Ma
ss
a r
ela
tiv
a d
o b
aç
o (
mg
)
Y
Y+M
I80
Y+B
z
CL
CL+M
I80
CL+B
z
NIT N
I
0 .0 0
0 .0 1
0 .0 2
0 .0 3*_ _ _ _ _ _ _ _ _ _ _ _ _ _
_ _ _ _ _ _ _ _ _*
#
Ma
ss
a r
ela
tiv
a d
o f
íga
do
(m
g)
Y
Y+M
I80
Y+B
z
CL
CL+M
I80
CL+B
z
NIT N
I
0 .0 5
0 .1 0
0 .1 5
_ _ _ _ _ _ _ _*
*_ _ _ _ _ _ _ _
Figura 18 - Massa corporal e massa relativa do coração, baço e fígado. Camundongos Swiss
infectados e não infectados com a cepa Y ou CL do T. cruzi, tratados ou não com a MI80 ou Bz. A
massa corporal (A) dos animais foi avaliada após a eutanásia e, em seguida, o peso relativo do
coração (B), baço (C) e fígado (D), obtido a partir relação entre a massa do órgão dividido pela massa
corporal do animal. As caixas representam a mediana, máximo e mínimo. (*) representa p<0,05 em
relação aos controles NI e NIT. (#) mostra também diferença estatística em relação ao grupo
infectado e tratado com Bz. (Xtd) Xantenodiona MI80, (Bz) Benznidazol, (NI) Não infectado e (NIT)
Não infectado tratado com a MI80.
4.3.4 Efeito do tratamento com MI80 nos parâmetros inflamatórios
na infecção experimental pelo T.cruzi
4.3.4.1 Histopatologia e estereologia do miocárdio
O efeito do tratamento com a MI80 alterou aspectos histopatológicos cardíacos durante a
fase aguda da infecção experimental pelo T. cruzi e os resultados quantitativos e qualitativos estão
apresentados nas figuras 19 e 20, respectivamente. Foram observados ninhos de amastigotas nos
animais infetados e não tratados para as duas cepas estudadas (Fig. 20Y e CL), além de infiltrado
inflamatório caracterizado como leve a moderado com predominância de células mononucleares

61
(Fig.19C). Verificamos que o tratamento com a MI80 foi capaz de reduzir o infiltrado
inflamatório quando comparado aos animais que não receberam o composto (Fig.19A). O mesmo
perfil não foi percebido quando avaliamos as células polimorfonucleares, no qual o tratamento
não foi eficiente em reduzir o número de células para cepa Y (Fig.19B). Entretanto, o grupo
infectado pela cepa CL e tratado com a MI80 apresentou celularidade estatisticamente igual aos
controles não infectados. Observou-se ainda, redução do processo inflamatório no miocárdio nos
animais tratados com o Bz tanto para o grupo infectado com a cepa Y quanto com a CL do T.
cruzi (Fig. 19 A e B)
Figura 19 - Quantificação do processo inflamatório no músculo cardíaco. Camundongos Swiss
infectados com 5.000 formas tripomastigotas sanguíneas da cepa Y ou CL do T. cruzi foram
tratados ou não com MI80 ou Benznidazol (Bz) na fase aguda. Análise do infiltrado inflamatório
contendo células mononucleares (A) e polimorfonucleares (B) no miocárdio dos animais. (C)
Diagrama de campo octogonal organizado em domínios que representa a celularidade tecidual da
seguinte forma: (-) celularidade mínima / (+ ) leve, (+ +) moderada ou (+ + +) intensa / infiltrado
inflamatório. * p< 0,05 comparado com o grupo controle infectado e não tratado; # p<0,05 em
relação ao grupo tratado com Bz e & p<0,05 comparado ao grupo tratado com MI80. (NI) Não
infectado e (NIT) Não infectado tratado com a MI80.

62
Figura 20 - Fotomicrografia de cortes histológicos do tecido muscular cardíaco.
Camundongos Swiss foram infectados com 5.000 formas tripomastigotas sanguíneas da cepa Y
ou CL e tratados ou não com MI80 ou Bz, na fase aguda. As figuras superiores representam os
animais infectados pela cepa Y, além do controle não infectado. As imagens inferiores mostram
os animais infectados pela cepa CL e o controle não infectado e tratado com a MI80.Nos animais
infectados e não tratados (Y) ou (CL), o infiltrado inflamatório apresentou-se de maneira
moderada e foi possível identificar ninhos de amastigotas. Para o grupo de animais infectados e
tratados (Y+MI80) e (CL+MI80) verificou-se processo inflamatório discreto e focal. Nos animais
infectados e tratados com as doses 100 mg/Kg de Bz não houve detecção de infiltrado. Os
controles NI e NI+MI80 apresentaram aspecto histológico cardíaco normal. Barra 60μm2. (MI80)
Xantenodiona MI80, (Bz) Benznidazol, (NI) Não infectado e (NIT) Não infectado tratado com a
MI80.
Os animais infectados não tratados apresentaram redução no volume total dos
cardiomiócitos (Vv[cmy]) para cepa Y e CL, quando comparados aos grupos controle não
infectados e infectados e tratados com a MI80 (Fig.21A). Verificou-se ainda um aumento de
volume intersticial (Vv [cnt]) e vasos sanguíneos (Vv [bvs]) nos grupos Y e CL em relação aos
grupos Y+MI80, CL+MI80, NI e NIT (Fig.21B e C). A relação entre (Vv [bvs])/ (Vv [cnt])
indicou redução da vascularização do miocárdio no grupo Y e CL e o tratamento com a MI80 foi
capaz de controlar este processo em relação aos grupos infectados e sem tratamento (Fig.21D).
Contudo, há um aumento dos compartimentos estruturais e funcionais, obtidos da razão entre (Vv
[cnt])/ (Vv[cmy]), para os grupos Y, Y+MI80, CL e CL+MI80 quando comparados ao grupo
Y+MI80
60 µm
Y
(
60 µm
NI
60 µm
CL
60 µm
NI+MI80
60 µm
CL+MI80
60 µm
CL+Bz
60 µm
Y+Bz
60 µm

63
controle NI e NIT. O tratamento com Bz foi capaz de reverter todos os danos causados pela
infecção do ponto de vista estereológico.
Y
Y+M
I80
Y+B
zC
L
CL
+M
I80
CL
+B
zN
I
NI+
MI8
0
6 0
7 0
8 0
9 0
1 0 0
Vv
[c
my
] %
#_ _ _ _ _ _ _ _ _ _*
#_ _ _ _ _ _ _ _ _ _&
*
A
Y
Y+M
I80
Y+B
zC
L
CL
+M
I80
CL
+B
zN
I
NIT
0
1 0
2 0
3 0
4 0
Vv
[in
t] %
#_ _ _ _ _ _ _ _ _ _*
_ _ _ _ _ _ _ _ _ _ _ _ _ _&
*#
B
Y
Y+M
I80
Y+B
zC
L
CL
+M
I80
CL
+B
zN
I
NIT
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
Vv
[b
vs
] %
_ _ _ _ _ _ _ _ _ _ _ _ _ _&
*#
&*
& *#
C
Y
Y+M
I80
Y+B
zC
L
CL
+M
I80
CL
+B
zN
I
NIT
0 .0 0 0
0 .0 0 2
0 .0 0 4
0 .0 0 6
0 .0 0 8
0 .0 1 0
MV
I (
bv
s /
cm
y )
& *# &
D
Y
Y+M
I80
Y+B
zC
L
CL
+M
I80
CL
+B
zN
I
NIT
0 .0
0 .2
0 .4
0 .6
PS
R (
in
t /
cm
y )
_ _ _ _ _ _ _ _ _&
*#
_ _ _ _ _ _ _ _ _*#
E
Figura 21 – Aspectos estereológicos do miocárdio. Camundongos Swiss foram infectados com
5.000 formas tripomastigotas sanguíneas da cepa Y ou CL e tratados ou não com MI80 ou Bz, na
fase aguda. Confirmada a infecção, os animais foram tratados por 10 dias e o coração foi removido
e fixado em formol tamponado (10%). (A) Densidade volumétrica do caridiomiócito. (B)
Densidade volumétrica do tecido conjuntivo. (C) Densidade volumétrica dos vasos sanguíneos.
(D)Vascularização do miocárdio. (E) Compartimentos estruturais e funcionais. * p< 0,05
comparado com o grupo controle infectado e não tratado; # p<0,05 em relação ao grupo tratado
com Bz e & p<0,05 comparado ao grupo tratado com MI80. (NI) Não infectado e (NIT) Não
infectado tratado com a MI80. (Vv[cmy]) = volume total dos cardiomiócitos, (Vv [cnt])= volume
intersticial e (Vv [bvs])= volume de vasos sanguíneos.

64
4.3.4.2 Mediadores inflamatórios
A resposta imune após o tratamento com a xantenodiona MI80 foi também avaliada
neste estudo para citocinas inflamatórias e regulatórias. A proteína ligante de RankL não
sofreu alterações em nenhum dos grupos infectados pela cepa Y do T. cruzi. Já para a cepa
CL houve aumento na concentração desta proteína (p=0,009) no grupo infectado e tratado
com a MI80 em comparação aos grupos controle NI, NIT (Fig.22A). Observamos um
aumento de IFN e CCL2 para os animais infectados pela cepa CL com e sem tratamento com
a MI80, em relação aos grupos controle. Este perfil não foi observado nos animais infectados
pela cepa Y, que tiveram aumento na concentração de CCL2 no tecido cardíaco (Fig.22B).
Para as duas cepas do parasito, o tratamento com o Bz foi capaz de manter os níveis de IFN-
γ e CCL2 próximos aos dos grupos controles, sem diferença significativa (Fig.22B e C).
Embora o MI80 não tenha afetado os níveis de IL-10 no contexto da infecção com a cepa Y,
verificou-se aumento da produção desta citocina para os animais infectados pela cepa CL e
tratado com a MI80 em relação aos animais infectados e tratados com Bz (Fig.22D).
Ainda, avaliando em conjunto os 4 marcadores inflamatórios, o tratamento com a
dose 50mg/kg (NIT), durante 10 dias nos animais sem infecção, não apresentou redução ou
aumento da concentração destas proteínas quando comparado ao grupo não infectado sem
tratamento (Fig. 22).

65
Y
Y+M
I80
Y+B
zC
L
Cl+
MI8
0
Cl+
Bz
NIT N
I
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0*
Ra
nk
L (
pg
/10
0m
g)
Y
Y+M
I80
Y+B
zC
L
Cl+
MI8
0
Cl+
Bz
NIT N
I
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
CC
L2
(p
g/1
00
mg
)
_ _ _ _ _ _ _ _ _ _ _ _ _ _* *# #
Y
Y+M
I80
Y+B
zC
L
Cl+
MI8
0
Cl+
Bz
NIT N
I
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
6 0 0
IFN
(p
g/1
00
mg
)
*_ _ _ _ _ _
Y
Y+M
I80
Y+B
zC
L
Cl+
MI8
0
Cl+
Bz
NIT N
I
1 5 0 0
2 0 0 0
2 5 0 0
3 0 0 0
3 5 0 0
4 0 0 0
IL-1
0 (
pg
/10
0m
g)
#
A
A
C
B
Figura 22 - Perfil de marcadores inflamatórios no sobrenadante do macerado de miocárdio
de camundongos infectados ou não com a cepa Y ou CL do T.cruzi e tratados com a MI80
ou Bz. (A) Concentração de RankL. (B) níveis de CCL2, (C) IFN-gama no miocárdio e (D) IL-
10. O símbolo (#) representa p<0,05 em relação ao grupo infectado e tratado com Bz e (*) em
relação aos grupos NI e NIT. Os resultados estão expressos em caixas contendo a mediana,
máximo e mínimo. (Xtd) Xantenodiona MI80, (Bz) Benznidazol, (NI) Não infectado e (NIT) Não
infectado tratado com a MI80.
4.3.6 Tratamento com a xantenodiona MI80 e avaliação da
toxicidade renal e hepática
Lesões hepáticas e ou renais induzidas por fármacos constituem um dos principais
problemas a serem solucionados na busca por novos fármacos. Neste contexto foi dosado
no plasma dos animais, ureia e creatinina para verificação de possível lesão renal, além
de AST/TGO e ALT/TGP para avaliar se houve toxicidade hepática.
Verificou-se que o tratamento com a MI80 não gerou toxicidade renal e hepática,
uma vez que os valores desses marcadores no plasma nos animais não infectados e tratdos
com a MI80 foram similares ao grupo controle NI (Fig.23). Foi observada elevação das
enzimas hepáticas nos grupos infectados com a cepa Y com e sem tratamento com a MI80

66
em ralção aos controles não infectados. Para cepa CL, apenas o grupo infectado e sem
tratamento elevou a produção de ALT/TGP.
Figura 23 - Concentrações plasmáticas de TGO (A), TGP (B), ureia (C) e creatinina (D) de
camundongos infectados com as cepas Y e CL do T. cruzi, tratados com 50mg/kg durante
10 dias por via oral. O símbolo (*) representa diferença estatística (p<0,05) em relação aos
controles NI e NIT (#) representa p<0.05 em relação ao grupo infectado e tratado com Bz e (&)
indica diferença entre o grupo infectado e não tratado e o grupo tratado com a MI80. (, (Bz)
Benznidazol, (NI) não infectado e (NIT) não infectado tratado com a MI80
AL
T -
U/m
L
Y
Y+
MI8
0
Y+
BZ
CL
CL+
MI8
0
CL+
Bz
NIT N
I
5 0
1 0 0
1 5 0
2 0 0
2 5 0 _ _ _ _ _ _ _ _*#
#
Ure
ia-
mg
/dL
Y
Y+
MI8
0
Y+
BZ
CL
CL+
MI8
0
CL+
Bz
NIT N
I
0
2 5
5 0
7 5
AS
T -
U/m
L
Y
Y+
MI8
0
Y+
BZ
CL
CL+
MI8
0
CL+
Bz
NIT N
I
0
1 2 5
2 5 0
3 7 5
5 0 0 _ _ _ _ _ _ _ _*& #
Cre
ati
nin
a -
mg
/dL
Y
Y+
MI8
0
Y+
BZ
CL
CL+
MI8
0
CL+
Bz
NIT N
I
0 .0
0 .2
0 .4
0 .6
0 .8

67
5. DISCUSSÃO
Doenças negligenciadas como a tripanossomíase americana geram grandes impactos
em países de clima tropical (WHO, 2012). Há pouco incentivo, principalmente pela indústria
farmacêutica, para o desenvolvimento de novos compostos/fármacos que possam mudar esta
realidade (Nwaka e Hudson, 2006). Além disso, a complexidade genética dos
tripanosomastídeos, associado ao desequilíbrio da resposta imune provocado pela interação
parasita-hospedeiro, permanece parcialmente incompreendida e contribui negativamente
para a ausência de uma boa estratégia terapêutica (Tarleton, Rick L., 2003; Tarleton, R. L.,
2003; Tarleton et al., 2007)
Para sanar esta lacuna, muitos pesquisadores têm se esforçado na busca de compostos
alternativos, advindos de várias bibliotecas, com diferentes esqueletos moleculares e radicais
(Pinto et al., 2005; Bustamante e Tarleton, 2011). No presente estudo, foram testados doze
compostos, oriundos de uma biblioteca de xantenodionas (1-8-octahidroxantonas) com
potenciais atividades anti-T. cruzi, anti-inflamatória e cardioprotetora (Lin et al., 1996;
Garrido et al., 2001; Jiang, D.-J. et al., 2004; Dar et al., 2005; Jassar et al., 2005; Librowski
et al., 2005; Chomnawang et al., 2007; Marona et al., 2008; Bumrungpert et al., 2010;
Szkaradek et al., 2015; Shagufta e Ahmad, 2016; Daei Farshchi Adli et al., 2018). Por estas
razões, estas moléculas despertaram a atenção do nosso grupo pela possibilidade de alterar
positivamente a intrínseca relação existente entre a infecção pelo T. cruzi e a
imunopatogênese cardíaca desenvolvida em seu hospedeiro mamífero.
Encontrar e desenvolver um novo medicamento é um processo longo, complexo,
dispendioso e altamente arriscado. A pesquisa e o desenvolvimento, para a maioria dos
fármacos disponíveis hoje, exigiram de 12 a 24 anos, desde o início de seus projetos até
alcançarem as prateleiras das farmácias. Além disso, muitas pesquisas possuem custo
elevado e, ao final, não atingem os resultado esperado (Lombardino e Lowe Iii, 2004). Por
isso vários (e diferentes) protocolos para validação de compostos ativos contra o T. cruzi
encontram-se descritos na literatura (Canavaci et al., 2010; Romanha et al., 2010;
Bustamante e Tarleton, 2011; Chatelain e Konar, 2015; Osorio-Méndez e Cevallos, 2019).
Experimentos em modelos celulares são fundamentais para uma rápida triagem de

68
compostos, uma vez que o uso de animais para este fim está sendo descontinuado (Zingales
et al., 2009; Bustamante e Tarleton, 2011).
Neste trabalho, estratégias in vitro foram utilizadas inicialmente para testar a
atividade tripanocida das xantenodionas. A mudança da posição dos átomos dentro da
molécula alterou substancialmente sua atividade contra o T. cruzi, principalmente para os
compostos MI79, MI80 e MI81. Já foi demonstrado que xantonas extraídas de fungos e
plantas, como a Garcinea intermedia e Garcinea subelíptica possuem comprovada ação em
epimastigotas e tripomastigotas da cepa Tulahuen, com valores de IC50 menores do que os
encontrados por nosso grupo (Abe et al., 2003; Abe et al., 2004; Pontius et al., 2008).
Entretanto é interessante ressaltar que esta cepa Y é descrita como parcialmente resistente
ao Bz (Zingales et al., 2009), e, de uma maneira geral os trabalhos disponíveis utilizam
cepas sensíveis.
Este fato nos encorajou a avaliar a ação citotóxica dos mesmos compostos em
macrófagos de camundongos. O padrão internacional preconiza que a concentração a ser
usada para experimentação mantenha no mínimo 70% de viabilidade celular (ISSO:109935,
2009). Concentrações que reduzam este valor já padronizado são consideradas citotóxicas.
É sabido que mesmo apresentando uma boa ação contra parasitos, a toxicidade das xantonas
na maioria dos relatos é alta, o que inviabiliza seu uso para o fim parasitológico (Abe et al.,
2003; Pinto et al., 2005; Al-Massarani et al., 2013). Nossos achados corroboram
parcialmente com a literatura, uma vez que a maior parte das xantenodionas são tóxicas
apenas nas maiores concentrações, acima de 100µM/mL. Entre os compostos análogos,
MI79, MI80 e MI81, os valores de CC50 foram similares evidenciando que esta pequena
mudança na posição dos átomos dentro da molécula não foi prejudicial para células de
mamíferos.
Vale lembrar que os valores de IC50 e CC50 são fundamentais para a determinação
do índice de seletividade, que consiste na razão entre a concentração citotóxica de 50% para
as células de mamíferos e a concentração de inibição de 50% para o T. cruzi. O valor
encontrado para o IS indica quantas vezes o composto é mais toxico para o parasita do que
para as células hospedeiras (Nwaka e Hudson, 2006). Dessa maneira, foi calculado o IS para
as seis xantenodionas testadas e observou-se que, apesar de apresentarem atividade
tripanocida, o IS não foi satisfatório neste modelo. Há autores que estabelecem um índice de

69
seletividade >50 para que o composto chegue a próxima etapa em testes de atividade
tripanocida (Nwaka e Hudson, 2006).
A baixa seletividade também é um problema conhecido dos pesquisadores que
trabalham com esta molécula. Al-Massarani et al. (2013), mostrou um bom potencial
tripanocida de xantonas naturais, entretanto sua alta citotoxidade fez com que estes
compostos não se tornassem atrativos para estudos futuros. O mesmo perfil foi observado
por Abe et al. (2004) que encontrou ISs insatisfatórios, mesmo usando cepas do T. cruzi
susceptíveis ao Bz. Para reduzir os efeitos nocivos das xantonas em células de mamíferos e,
consequentemente, reduzir a sua toxicidade, vários autores têm sugerido mudanças
conformais na molécula. Esta prática é descrita principalmente para atividade anti-malárica
das xantonas e pode garantir a otimização dos compostos. Além disso, estas pequenas
mudanças conformacionais também podem aumentar ou diminuir a polaridade da molécula,
o que interfere diretamente em sua ação (Kelly et al., 2002; Fotie et al., 2003; Tjahjani,
2017).
Existe uma necessidade de novos compostos para o tratamento da infecção pelo T.
cruzi em todos os estágios da doença de Chagas (Rodriques Coura e De Castro, 2002;
Tarleton et al., 2007; Urbina, 2010). Apesar de vários estudos serem publicados anualmente,
há uma falha nos testes in vitro e in vivo que, em sua maioria, não são totalmente adequados
para comprovar a eficiência das drogas (Chatelain, 2015). Vários ensaios colorimétricos e
fluorimétricos foram desenvolvidos para rastrear compostos anti-T.cruzi (Muelas-Serrano et
al., 2000; Vega et al., 2005; Rolon et al., 2006). Esses testes são mais precisos e objetivos
do que o método tradicional que envolve contagem visual microscópica.
No protocolo de triagem de drogas anti-T.cruzi, proposto por Romanha e cols.
(2010), foi sugerido o uso de parasitos geneticamente modificados expressando o gene da
b-galactosidase descrito por Buckner et al. (1996). Mesmo que amplamente utilizado, este
método apresenta desvantagem, pois só é possível realizar uma leitura de ponto final após a
adição de detergente e com um substrato conversível. Linhagens de T. cruzi expressando
repórteres alternativos como proteínas fluorescentes (tdTomato) já foram descritas e
demonstram que os parasitas emitem fluorescência em todas as fases evolutivas sendo
possível avaliar o crescimento ou inibição dos parasitos de maneira contínua (Darocha et al.,
2004; Guevara et al., 2005; Hyland et al., 2008). Entre as vantagens do uso destes parasitas
fluorescentes para ensaios in vitro, está a eliminação da necessidade de fixação ou de

70
permeabilização e a detecção de fluorescência com o mínimo manuseio, usando um
fluorímetro e um “screener” baseado em microscópio ou mesmo outro sistema de imagem
(Canavaci et al., 2010).
Baseados em ensaios de alto rendimento para triagem de compostos com atividade
anti-T.cruzi, os experimento in vitro descritos a seguir foram realizados no laboratório do
Dr. Rick L Tarleton, na Universidade da Geórgia (USA), durante meu período de doutorado
sanduíche. Este laboratório desenvolve estratégias para ensaios de alta performance em
triagem de fármacos, tanto em modelos celulares, quando murinos (Canavaci et al., 2010;
Bustamante et al., 2014). Zingales et al. (2014) recomendam que a triagem de compostos
anti-T.cruzi deva ser realizada em amastigotas intracelulares. Apesar dos ensaios em
epimastigotas e tripomastigotas serem muito menos trabalhosos, os resultados não são
coerentes com os encontrados em amastigotas. Assim, em nosso estudo foram utilizadas
células Vero gamma irradiadas para os ensaios de atividade em formas intracelulares do T.
cruzi . A irradiação gamma nas células é capaz de inibir a replicação, fazendo com que não
haja crescimento celular exacerbado (Yan e Moreno, 1998). Optou-se por usar a cepa CL
tdtomato, uma vez que Sánchez-Valdéz et al. (2018) mostraram que o tratamento com o Bz
elimina de maneira rápida e eficiente a maior parte dos parasitas independentemente da
cepa, mantendo a taxa de “clearence” parasitário similiar.
Assim, os doze derivados de xantenodionas propostos em nosso estudo foram
testados utilizando a plataforma descrita previamente. Entre todos os compostos, apenas três
apresentaram ação tripanocida relevante. É importante chamar a atenção para os análogos
MI79, MI80 e MI81, dos quais apenas o MI80 mostrou ação anti-T.cruzi sem induzir
citotoxidade, ou seja, atuou diretamente sob o parasito. Outro aspecto relevante foi a maior
atividade da MI80, MI24 e MI25 em eliminar formas intracelulares, ou amastigotas. Este
resultado corrobora com o trabalho de De Oliveira Caleare et al. (2013) que testou xantonas
extraídas de uma árvore nativa do cerrado brasileiro, o pau santo, para ação anti- T.cruzi.
Isoladamente, outros autores avaliaram a ação de xantonas derivadas de fontes naturais e
verificaram maior atividade em formas flageladas do T. cruzi do que em amastigotas,
discordando dos nossos resultados (Pontius et al., 2008; Dua et al., 2009; Al-Massarani et
al., 2013)
Paralelamente foram realizados, ainda, ensaios para verificação da toxicidade celular
usando o reagente AlamarBlue. Nesta etapa, testou-se os mesmos compostos usados nos

71
experimentos com J774A.1 e foram adicionados outros seis. Para os compostos testados em
ambos os ensaios, colorimétricos e fluorimétricos (MI17, MI24, MI25, MI79, MI80 e MI81),
observou-se que o tipo celular e a estratégia metodológica para determinar a citotoxidade
não alteraram resultados, uma vez que os valores de CC50 foram similares e o perfil de
viabilidade celular também. Já para as xantenodionas não testadas previamente a maior parte
teve a viabilidade celular reduzida a 70% ou mais, em concentrações menores do que a
necessária para os experimentos em amastigotas (50µM/mL). Conforme discutido nos
parágrafos anteriores, a citotoxidade dessa classe de compostos não é a ideal para
experimentos de bioatividade. Além disso, a apolaridade de algumas xantenodionas pode ter
contribuído para a redução da viabilidade celular e atividade tripanocida. Mais uma vez,
reforçamos a necessidade de ajustes moleculares que permitam adequar a polaridade destas
moléculas e melhorar sua eficiência
Após avaliarmos o perfil de citotoxidade e a atividade tripanocida, elegemos o
composto MI80 para prosseguir com os ensaios in vivo. Nosso objetivo inicial era não
somente avaliar a ação anti-T.cruzi, mas também a ação destes compostos na resposta
inflamatória do hospedeiro frente a infecção. Assim, a abordagem tradicional, descritas nos
protocolos para descoberta de novos compostos com atividade anti-T.cruzi, pôde ser
flexibilizado para responder a estas perguntas. A escolha da linhagem animal (camundongo
Swiss fêmeas) foi feita baseada nos protocolos descritos por Romanha et al. (2010) e
Chatelain e Konar (2015). Este modelo é recomendado devido ao “background” genético
dos animais e por apresentarem um perfil de infecção aguda com alta parasitemia e níveis
de mortalidade, permitindo uma comparação entre o efeito do composto testado e o efeito
do Bz. Além disso, desde 2014, agências de pesquisa internacionais preconizam o uso de
fêmeas em experimentos nos quais a doença, objeto do estudo, não tenha correlação com
fatores genéticos como o gênero (Collins, 2014; Mccullough et al., 2014).
A escolha das cepas também foi baseada nos mesmos protocolos para descoberta de
novos fármacos para doença de Chagas descritos acima, por apresentarem resistência
moderada ou não ao Bz e ao nifurtimox (Filardi & Brener, 1987) e serem usadas
rotineiramente em estudos in vitro e in vivo para atividade de testes de fármacos. Além disso,
o uso destas cepas neste tipo de ensaio permite a identificação rápida (menos de 40 dias) da
dose ideal necessária para etapas subsequentes no processo de triagem (Romanha et al.,
2010; Bustamante e Tarleton, 2011; Zingales et al., 2014; Chatelain e Konar, 2015).

72
Não há trabalhos em modelo animal que envolvendo o T. cruzi e as xantendionas,
sendo necessária a verificação da dose máxima tolerada e sua influência na parasitemia.
Revisando a literatura para estabelecer o esquema de tratamento, foi observado que os
xantenos naturais possuem baixa biodisponibilidade oral em roedores (Gutierrez-Orozco e
Failla, 2013). Em camundongos C57BL/6 quando α-MG foi administrada por gavagem oral
em uma suspensão oleosa (100 mg / kg), uma concentração plasmática máxima de 1,38 μmol
/ L foi alcançada em 30 min (Petiwala et al., 2014). Além disso, o α-MG foi detectado no
plasma 24 h após a administração oral, sugerindo um padrão de eliminação lento (Atulkumar
et al., 2012). Por esta razão foram infectados animais com a cepa Y do T. cruzi e testadas
cinco doses da MI80 (800, 400, 200, 100 e 50mg/kg) em duas vias, oral e intraperitoneal,
por 10 dias.. Como os animais tratados por gavagem tiveram maior taxa de sobrevivência do
que os animais que receberam a MI80 por injeção intraperitoneal foi priorizada a via oral,
na menor dose capaz de reduzir a parasitemia (50mg/kg) sem causar a mortalidade dos
animais.
Observamos em nosso estudo uma redução da parasitemia em animais infectados
com a cepa Y do T.cruzi e tratados com a MI80. A partir destes resultados iniciais, foram
infectados novos camundongos com as cepas Y (parcialmente resistente ao Bz) e CL
(susceptível ao Bz) do T. cruzi com o objetivo de avaliar oefeito da dose de 50mg/kg na
redução da parasitemia e dos parâmetros inflamatórios. Nos animais infectados com a cepa
Y, dez dias de tratamento diário foi capaz de reduzir em 40% o número de parasitos
circulantes pelo exame de sangue a fresco, além de prevenir a mortalidade dos animais.
Parasitemia e mortalidade são critérios básicos no teste de composto com atividade anti-T.
cruzi (Romanha et al., 2010). Apesar desta redução não ser significativa (p<0,05) quando
comparada ao grupo controle infectado e não tratado, ela pode interferir diretamente na
inflamação do miocárdio (Caldas et al., 2014). Para a cepa CL, o período pré-patente ocorre
entre o 6º e o 7º dias após a infecção e há um aumento da parasitemia a partir do 16º dia,
atingindo o pico no 22º dia (Zingales et al., 1997). Nossos experimentos em animais
infectados com a cepa CL foram interrompidos ao final de 18 dias de infecção e 10 dias de
tratamento, não sendo possível avaliar alterações no auge da circulação de parasitos no
sangue.
Durante a fase aguda da infecção pelo T. cruzi, a resposta imunológica exacerbada é
frequentemente observada no miocárdio, gerando lesões associadas que, em casos extremos
pode levar a morte em consequência de uma resposta inflamatória sistêmica (Rossi e

73
Mengel, 1992; Gutierrez et al., 2009). Com o objetivo de avaliar o impacto do tratamento e
a consequente redução da parasitemia na evolução das lesões cardíacas, realizou-se a análise
quantitativa do número de células inflamatórias no miocárdio dos camundongos infectados.
Os resultados alcançados apontam que a existência e intensidade das lesões cardíacas estão
associadas com a cepa do parasito e à sua sensibilidade aos compostos utilizados no
tratamento. Apesar de não haver induzido a cura parasitológica, a MI80 mostrou-se efetiva
em reduzir as lesões cardíacas nos animais infectados com a cepa Y e CL. Em concordância
com os resultados alcançados em nosso trabalho, Caldas et al. (2019), mostrou em modelo
cão, que a redução do número de parasitas tem efeito benéfico e é capaz de prevenir lesões
cardíacas nos estágios iniciais da infecção.
Além da avaliação histológica do tecido cardíaco, avaliou-se também o efeito’ do
tratamento com a MI80 no padrão de produção de mediadores inflamatórios. O gatilho e a
manutenção da resposta inflamatória está diretamente relacionado a ativação de vias de
sinalização celular, como a NF-kb (Bistrian, 2007; Ang et al., 2011; Arthur e Ley, 2013;
Afonina et al., 2017). Neste contexto, foi avaliado a produção do ligante de Rank (RankL),
citocina caracterizada como receptor ativador da via NF-kb. Os resultados mostraram um
aumento significativo desta proteína nos animais infectados com a cepa CL para o grupo
controle não tratado e tratados com a MI80. Estes achados sugerem que a infecção pode ser
fator responsável pela ativação da via e o tratamento com a MI80 não foi capaz de limitar
este processo. Trabalhos recentes já revelaram que a mangiferina, uma xantona natural, tem
a capacidade de inibir a ativação de NF-kB induzida por RankL in vitro, entretanto, não estão
descritos na literatura o envolvimento desta citocina em modelos in vivo (Ang et al., 2011;
Ma et al., 2015; Ding et al., 2018). Para os animais infectados com a cepa Y não houve
diferença entre nenhum grupo avaliado, possivelmente relacionado a características
intrínsecas da cepa do T.cruzi (Brener e Chiari, 1967; Brener et al., 1976; Filardi e Brener,
1987; Callejas-Hernández et al., 2018). O aumento de CCL2 foi coerente com os resultados
da análise histológica, uma vez que houve um predomínio de células mononucleares.
Nos animais infectados com a cepa Y ou CL e tratados com a MI80, a redução da
intensidade das lesões não foi acompanhada de redução dos níveis de IFN e CCL2. Como
esperado e característico da infecção (Talvani e Teixeira, 2011), para os grupos controles
infectados e não tratados houve um aumento destes mediadores. A produção de CCL2 está
relacionado tanto ao crescimento parasitário e a morfogênese de tripomastigotas quanto ao

74
aumento na ativação e ao recrutamento de infiltrado inflamatório para o coração (Marino
Ana Paula et al., 2004; Guedes et al., 2010).
Ressaltamos que o grupo infectado com a cepa CL e tratado com a MI80 apresentou
aumento da produção da IL-10. Trabalhos anteriores do nosso grupo já mostraram que a
elevação desta citocina está associada a proteção do miocárdio e consequentemente redução
do dano cardíaco na infecção pelo T.cruzi em modelo murino(De Paula Costa et al., 2010;
Penitente et al., 2015; Leite et al., 2017). Possivelmente, alteraçoes relacionadas tanto pela
redução da parasitemia quanto ao quadro inflamatório após o tratamento com a MI80 possam
estar envolvidos neste evento. Para os demais grupos, não foi observada diferença nos níveis
de IL-10 e acreditamos que o tempo de experimentação não tenha sido longo o suficiente
para promover mudanças na produção desta citocinas anti-inflamatória (Reed et al., 1994;
Cunha-Neto et al., 1998)
Nossos resultados demonstraram que apesar de não induzir à cura parasitológica, o
tratamento com a MI80 foi eficaz em reduzir o infiltrado inflamatório nos animais infectados
com a cepa Y e CL do T. cruzi e, esta redução, pode estar associada com diminuição, mesmo
que não significativa, da parasitemia. Ademais, existem incontestáveis hipóteses que esta
classe de compostos possui ação cardioprotetora, principalmente por mecanismos
antioxidantes (Dai et al., 2004; Jiang, D. J. et al., 2004; Marona et al., 2008; Frangogiannis,
2014; Szkaradek et al., 2015; Imran et al., 2017). Sabendo-se que as cepas Y e CL do T.cruzi
são cardiotrópicas e não havendo relação entre a intensidade de inflamação no miocárdio e
redução de mediadores inflamatórios, sugere-se um potencial efeito cardioprotetor gerado
pela xantenodiona, não elucidado neste estudo.
O tratamento com a xantenodiona MI80 não induziu o aparecimento de reações
adversas detectáveis. Além disso, não foi verificada toxicidade hepática e renal, tendo como
parâmetros as concentrações de AST/TGO e ALT/TGP, ureia e creatinina dosados no plasma
dos animais não infectados e tratados com 50mg/kg da MI80. Foi observado um aumento
das enzimas hepáticas nos animais infectados, possivelmente pelos metabólitos gerados a
partir da infecção pelo T. cruzi (Penas et al., 2016). Há indícios que as xantonas exerçam
papel hepatoprotetor em modelos de toxicidade induzida por D-galactosamina (GAL),
revertendo os efeitos adversos causados por esta toxina (Das et al., 2012). Em discordância
dos resultados já descritos na literatura(Novaes et al., 2015), neste estudo o tratamento com

75
Bz não foi capaz de elevar as enzimas indicativas de dano hepático possivelmente pelo
rápido período de tratamento (10 dias).
Mesmo em condições experimentais, os modelos de infecção pelo T. cruzi envolvem
vários parâmetros muitas vezes não controlados, incluindo a relação entre a exposição e a
cura, via de administração, tempo e duração do tratamento, linhagem do camundongo,
interações parasita-hospedeiro e tipo de resposta imune (Chatelain e Konar, 2015). Deste
modo, levando em consideração estes parâmetros, o derivado MI80 do 1,8-
dioxooctahydroxanthenos apresenta-se como uma alternativa segura e promissora para
novos estudos envolvendo o tratamento da infecção pelo T. cruzi em modelo experimental.
São necessários ainda, mais estudos que demonstrem os mecanismos de cardioproteção neste
modelo de infecção experimental com o objetivo de estabelecer a interação destes compostos
com o T. cruzi e o hospedeiro vertebrado.

76
6. CONCLUSÃO
Nossos achados mostraram que o 1,8-dioxo-octa-hidroxanteno apresenta efeito anti-
parasitário limitado, entretanto na infeção aguda em modelo murino, o tratamento por um
período de 10 dias, com 50mg/kg, refletiu na redução dos parâmetros inflamatórios
associados ao T.cruzi.

77
7. REFERÊNCIAS
ABE, F. et al. Trypanocidal Constituents in Plants 3.<br>Leaves of <i>Garcinia intermedia</i> and Heartwood of <i>Calophyllum brasiliense</i>. Biological and Pharmaceutical Bulletin, v. 27, n. 1, p. 141-143, 2004.
ABE, F. et al. Trypanocidal constituents in plants 2.xanthones from the stem bark of Garcinia subelliptica. Biol Pharm Bull, v. 26, n. 12, p. 1730-3, Dec 2003. ISSN 0918-6158 (Print)
0918-6158 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/14646180 >.
ABERHAM, A. et al. Analysis of iridoids, secoiridoids and xanthones in Centaurium erythraea, Frasera caroliniensis and Gentiana lutea using LC–MS and RP-HPLC. Journal of Pharmaceutical and Biomedical Analysis, v. 54, n. 3, p. 517-525, 2011/02/20/ 2011. ISSN 0731-7085. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0731708510005649 >.
AFONINA, I. S. et al. Limiting inflammation—the negative regulation of NF-κB and the NLRP3 inflammasome. Nature Immunology, v. 18, p. 861, 07/19/online 2017. Disponível em: < https://doi.org/10.1038/ni.3772 >.
AL-MASSARANI, S. M. et al. Phytochemical, antimicrobial and antiprotozoal evaluation of Garcinia mangostana pericarp and alpha-mangostin, its major xanthone derivative. Molecules, v. 18, n. 9, p. 10599-608, Sep 2 2013. ISSN 1420-3049 (Electronic)
1420-3049 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24002136 >.
ALLAVENA, P. et al. Chemokines and chemokine receptors in the regulation of dendritic cell trafficking. Chem Immunol, v. 72, p. 69-85, 1999. ISSN 1015-0145 (Print)
0079-6034 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/10550931 >.
ANG, E. et al. Mangiferin attenuates osteoclastogenesis, bone resorption, and RANKL-induced activation of NF-kappaB and ERK. J Cell Biochem, v. 112, n. 1, p. 89-97, Jan 2011. ISSN 1097-4644 (Electronic)
0730-2312 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20683903 >.
ARTHUR, J. S.; LEY, S. C. Mitogen-activated protein kinases in innate immunity. Nat Rev Immunol, v. 13, n. 9, p. 679-92, Sep 2013. ISSN 1474-1741 (Electronic)
1474-1733 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23954936 >.
ATULKUMAR, R. et al. Single Dose Oral Pharmacokinetic Profile of α-Mangostin in Mice. Current Drug Targets, v. 13, n. 14, p. 1698-1704, 2012. ISSN 1389-4501/1873-5592. Disponível em: < http://www.eurekaselect.com/node/105552/article >.

78
BAFICA, A. et al. Cutting edge: TLR9 and TLR2 signaling together account for MyD88-dependent control of parasitemia in Trypanosoma cruzi infection. J Immunol, v. 177, n. 6, p. 3515-9, Sep 15 2006. ISSN 0022-1767 (Print)
0022-1767 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16951309 >.
BAGULEY, B. C.; SIEMANN, D. W. Temporal aspects of the action of ASA404 (vadimezan; DMXAA). Expert opinion on investigational drugs, v. 19, n. 11, p. 1413-1425, 2010. ISSN 1744-7658
1354-3784. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20964495
https://www.ncbi.nlm.nih.gov/pmc/PMC3583340/ >.
BHATTACHARYA, S. K.; SANYAL, A. K.; GHOSAL, S. Monoamine oxidase-inhibiting activity of mangiferin isolated from Canscora decussata. Naturwissenschaften, v. 59, n. 12, p. 651, Dec 1972. ISSN 0028-1042 (Print)
0028-1042 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/4540402 >.
BISTRIAN, B. Systemic response to inflammation. Nutr Rev, v. 65, n. 12 Pt 2, p. S170-2, Dec 2007. ISSN 0029-6643 (Print)
0029-6643 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18240543 >.
BONECCHI, R. et al. Up-regulation of CCR1 and CCR3 and induction of chemotaxis to CC chemokines by IFN-gamma in human neutrophils. J Immunol, v. 162, n. 1, p. 474-9, Jan 1 1999. ISSN 0022-1767 (Print)
0022-1767 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9886422 >.
BOOTS, A. W. et al. Oxidative damage shifts from lipid peroxidation to thiol arylation by catechol-containing antioxidants. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, v. 1583, n. 3, p. 279-284, 2002/08/08/ 2002. ISSN 1388-1981. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1388198102002470 >.
BRENER, Z. Therapeutic Activity and Criterion of Cure on Mice experimentally infected with Trypanosoma cruzi. Revista do Instituto de Medicina Tropical de Sao Paulo, v. 4, n. 6, p. 389-96, 1962. ISSN 0036-4665.
BRENER, Z.; CHIARI, E. [Susceptibility of different strains of Trypanosoma cruzi to various chemotherapeutic agents]. Rev Inst Med Trop Sao Paulo, v. 9, n. 4, p. 197-207, Jul-Aug 1967. ISSN 0036-4665 (Print)
0036-4665 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/5614998 >.
BRENER, Z.; COSTA, C. A.; CHIARI, C. Differences in the susceptibility of Trypanosoma cruzi strains to active chemotherapeutic agents. Rev Inst Med Trop Sao Paulo, v. 18, n. 6, p. 450-5, Nov-Dec 1976. ISSN 0036-4665 (Print)

79
0036-4665 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/828295 >.
BUCKNER, F. S. et al. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob Agents Chemother, v. 40, n. 11, p. 2592-7, Nov 1996. ISSN 0066-4804 (Print)
0066-4804 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/8913471 >.
BUMRUNGPERT, A. et al. Xanthones from mangosteen inhibit inflammation in human macrophages and in human adipocytes exposed to macrophage-conditioned media. J Nutr, v. 140, n. 4, p. 842-7, Apr 2010. ISSN 1541-6100 (Electronic)
0022-3166 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20181789 >.
BUSTAMANTE, J. M. et al. New, combined, and reduced dosing treatment protocols cure Trypanosoma cruzi infection in mice. J Infect Dis, v. 209, n. 1, p. 150-62, Jan 1 2014. ISSN 1537-6613 (Electronic)
0022-1899 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23945371 >.
BUSTAMANTE, J. M.; TARLETON, R. L. Methodological advances in drug discovery for Chagas disease. Expert Opin Drug Discov, v. 6, n. 6, p. 653-661, Jun 2011. ISSN 1746-045X (Electronic)
1746-0441 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21712965 >.
CALDAS, I. S. et al. Parasitaemia and parasitic load are limited targets of the aetiological treatment to control the progression of cardiac fibrosis and chronic cardiomyopathy in Trypanosoma cruzi-infected dogs. Acta Tropica, v. 189, p. 30-38, 2019/01/01/ 2019. ISSN 0001-706X. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0001706X18304637 >.
CALDAS, I. S. et al. Benznidazole therapy during acute phase of Chagas disease reduces parasite load but does not prevent chronic cardiac lesions. Parasitol Res, v. 103, n. 2, p. 413-21, Jul 2008. ISSN 0932-0113 (Print)
0932-0113 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18454349 >.
CALDAS, S. et al. Therapeutic responses to different anti-Trypanosoma cruzi drugs in experimental infection by benznidazole-resistant parasite stock. Parasitology, v. 141, n. 12, p. 1628-1637, 2014. ISSN 0031-1820. Disponível em: < https://www.cambridge.org/core/article/therapeutic-responses-to-different-antitrypanosoma-cruzi-drugs-in-experimental-infection-by-benznidazoleresistant-parasite-stock/4632E63DC0D989A89A3DB3C0E2F0BBE9 >.
CALLEJAS-HERNÁNDEZ, F. et al. Genomic assemblies of newly sequenced Trypanosoma cruzi strains reveal new genomic expansion and greater complexity. Scientific reports, v. 8, n. 1, p. 14631-14631, 2018. ISSN 2045-2322. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/30279473

80
https://www.ncbi.nlm.nih.gov/pmc/PMC6168536/ >.
CANAVACI, A. M. et al. In vitro and in vivo high-throughput assays for the testing of anti-Trypanosoma cruzi compounds. PLoS Negl Trop Dis, v. 4, n. 7, p. e740, Jul 13 2010. ISSN 1935-2735 (Electronic)
1935-2727 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20644616 >.
CARDOSO, M. S.; REIS-CUNHA, J. L.; BARTHOLOMEU, D. C. Evasion of the Immune Response by Trypanosoma cruzi during Acute Infection. Front Immunol, v. 6, p. 659, 2015. ISSN 1664-3224 (Print)
1664-3224 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26834737 >.
CHATELAIN, E. Chagas Disease Drug Discovery:Toward a New Era. v. 20, n. 1, p. 22-35, 2015. Disponível em: < https://journals.sagepub.com/doi/abs/10.1177/1087057114550585 >.
CHATELAIN, E.; KONAR, N. Translational challenges of animal models in Chagas disease drug development: a review. Drug design, development and therapy, v. 9, p. 4807-4823, 2015. ISSN 1177-8881. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26316715
https://www.ncbi.nlm.nih.gov/pmc/PMC4548737/ >.
CHEN, L.-G.; YANG, L.-L.; WANG, C.-C. Anti-inflammatory activity of mangostins from Garcinia mangostana. Food and Chemical Toxicology, v. 46, n. 2, p. 688-693, 2008/02/01/ 2008. ISSN 0278-6915. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0278691507004449 >.
CHOMNAWANG, M. T. et al. Effect of Garcinia mangostana on inflammation caused by Propionibacterium acnes. Fitoterapia, v. 78, n. 6, p. 401-8, Sep 2007. ISSN 0367-326X (Print)
0367-326X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17644272 >.
CIDADE, H. et al. In silico and in vitro antioxidant and cytotoxicity evaluation of oxygenated xanthone derivatives. Arabian Journal of Chemistry, 2017/01/25/ 2017. ISSN 1878-5352. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1878535217300096 >.
COLLINS, J. A. C. F. S. Policy: NIH to balance sex in cell and animal studies. Nature, v. 509, n. 7500, p. 282–283 May,15 2014.
CONLON J, B. D., SHARMA S, BHAT N, THOMPSON M, JIANG Z, RATHINAM VA,; MONKS B, J. T., XIAO TS, VOGEL SN, VANCE RE, FITZGERALD KA. STING, binds and signals in response to the vascular disrupting agent
5,6-dimethylxanthenone-4-acetic acid. Journal of Immunology, v. 190, n. 10, p. 5216-25, 2103.
CUNHA-NETO, E. et al. Cytokine production profile of heart-infiltrating T cells in Chagas' disease cardiomyopathy %J Brazilian Journal of Medical and Biological Research. v. 31, p. 133-137, 1998.

81
ISSN 0100-879X. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-879X1998000100018&nrm=iso >.
DA RE P, M. V., TOTH E, CIMA L. . Xanthone derivatives with centrally stimulating and analeptic activities. Arzneim Forsch (Drug Res) v. 18, p. 718-720, 1970.
DAEI FARSHCHI ADLI, A. et al. An overview on Vadimezan (DMXAA): The vascular disrupting agent. v. 91, n. 5, p. 996-1006, 2018. Disponível em: < https://onlinelibrary.wiley.com/doi/abs/10.1111/cbdd.13166 >.
DAI, Z. et al. 3,4,5,6-Tetrahydroxyxanthone protects against myocardial ischemia-reperfusion injury in rats. Cardiovasc Drugs Ther, v. 18, n. 4, p. 279-88, Jul 2004. ISSN 0920-3206 (Print)
0920-3206 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/15367825 >.
DAR, A. et al. Analgesic and antioxidant activity of mangiferin and its derivatives: the structure activity relationship. Biol Pharm Bull, v. 28, n. 4, p. 596-600, Apr 2005. ISSN 0918-6158 (Print)
0918-6158 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/15802793 >.
DARE, P., VALENTINI, P., BORRACCINI, A. & PRIMOFIORE, G.P. . S-Adrenergic blocking agents of the chromone and xanthone groups. Journal of Medicinal Chemistry, v. 15, p. 198-199, 1972.
DAROCHA, W. D. et al. Expression of exogenous genes in Trypanosoma cruzi: improving vectors and electroporation protocols. Parasitol Res, v. 92, n. 2, p. 113-20, Jan 2004. ISSN 0932-0113 (Print)
0932-0113 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/14634799 >.
DAS, J. et al. Mangiferin exerts hepatoprotective activity against D-galactosamine induced acute toxicity and oxidative/nitrosative stress via Nrf2–NFκB pathways. Toxicology and Applied Pharmacology, v. 260, n. 1, p. 35-47, 2012/04/01/ 2012. ISSN 0041-008X. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0041008X12000269 >.
DE OLIVEIRA CALEARE, A. et al. Trypanocidal activity of 1,3,7-trihydroxy-2-(3-methylbut-2-enyl)-xanthone isolated from Kielmeyera coriacea. Parasitology International, v. 62, n. 5, p. 405-411, 2013/10/01/ 2013. ISSN 1383-5769. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1383576913000585 >.
DE PAULA COSTA, G. et al. Enalapril prevents cardiac immune-mediated damage and exerts anti-Trypanosoma cruzi activity during acute phase of experimental Chagas disease. Parasite Immunol, v. 32, n. 3, p. 202-8, Mar 2010. ISSN 1365-3024 (Electronic)
0141-9838 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20398183 >.

82
DEEB, O.; ALFALAH, S.; CLARE, B. W. QSAR of aromatic substances: MAO inhibitory activity of xanthone derivatives. J Enzyme Inhib Med Chem, v. 22, n. 3, p. 277-86, Jun 2007. ISSN 1475-6366 (Print)
1475-6366 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17674808 >.
DESHPANDE, R. et al. Estradiol down-regulates LPS-induced cytokine production and NFkB activation in murine macrophages. Am J Reprod Immunol, v. 38, n. 1, p. 46-54, Jul 1997. ISSN 1046-7408 (Print)
1046-7408 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9266010 >.
DIAS, L. C. et al. Quimioterapia da doença de Chagas: estado da arte e perspectivas no desenvolvimento de novos fármacos %J Química Nova. v. 32, p. 2444-2457, 2009. ISSN 0100-4042. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-40422009000900038&nrm=iso >.
DINARELLO, C. A. Cytokines as mediators in the pathogenesis of septic shock. Curr Top Microbiol Immunol, v. 216, p. 133-65, 1996. ISSN 0070-217X (Print)
0070-217X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/8791739 >.
______. Historical insights into cytokines. Eur J Immunol, v. 37 Suppl 1, p. S34-45, Nov 2007. ISSN 0014-2980 (Print)
0014-2980 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17972343 >.
DINARELLO, C. A. et al. Suppression of innate inflammation and immunity by interleukin-37. Eur J Immunol, v. 46, n. 5, p. 1067-81, May 2016. ISSN 1521-4141 (Electronic)
0014-2980 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/27060871 >.
DING, L. Z. et al. Mangiferin inhibits apoptosis and oxidative stress via BMP2/Smad-1 signaling in dexamethasone-induced MC3T3-E1 cells. Int J Mol Med, v. 41, n. 5, p. 2517-2526, May 2018. ISSN 1791-244X (Electronic)
1107-3756 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/29484386 >.
DUA, V. K.; VERMA, G.; DASH, A. P. In vitro antiprotozoal activity of some xanthones isolated from the roots of Andrographis paniculata. Phytother Res, v. 23, n. 1, p. 126-8, Jan 2009. ISSN 1099-1573 (Electronic)
0951-418X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18683849 >.
DUTRA, W. O. et al. Cytokine mRNA profile of peripheral blood mononuclear cells isolated from individuals with Trypanosoma cruzi chronic infection. Scand J Immunol, v. 45, n. 1, p. 74-80, Jan 1997. ISSN 0300-9475 (Print)
0300-9475 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9010503 >.

83
FELIZARDO, A. A. et al. Impact of Trypanosoma cruzi infection on nitric oxide synthase and arginase expression and activity in young and elderly mice. Free Radic Biol Med, v. 129, p. 227-236, Dec 2018. ISSN 1873-4596 (Electronic)
0891-5849 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/30248443 >.
FILARDI, L. S.; BRENER, Z. Susceptibility and natural resistance of <em>Trypanosoma cruzi</em> strains to drugs used clinically in Chagas disease. Transactions of the Royal Society of Tropical Medicine and Hygiene, v. 81, n. 5, p. 755-759, 1987. ISSN 0035-9203. Disponível em: < https://doi.org/10.1016/0035-9203(87)90020-4 >. Acesso em: 2019/01/27.
FINNEGAN, R. A., STEPHANI, R. A., GANGULI, G., GANGULY, S. N., & BHATTACHARYA, A. K. Occurrence of Mangiferin in Hiptage madablota Geartn. Journal of Pharmaceutical Sciences, v. 57, p. 1039-1040, 1968.
FOTIE, J. et al. In-vivo antimalarial activity of some oxygenated xanthones. Annals of Tropical Medicine & Parasitology, v. 97, n. 7, p. 683-688, 2003/10/01 2003. ISSN 0003-4983. Disponível em: < https://doi.org/10.1179/000349803225002390 >.
FRANGOGIANNIS, N. G. The inflammatory response in myocardial injury, repair, and remodelling. Nature Reviews Cardiology, v. 11, p. 255, 03/25/online 2014. Disponível em: < https://doi.org/10.1038/nrcardio.2014.28 >.
FU, Q.; LI, C.; YU, L. Gambogic acid inhibits spinal cord injury and inflammation through suppressing the p38 and Akt signaling pathways. Mol Med Rep, v. 17, n. 1, p. 2026-2032, Jan 2018. ISSN 1791-3004 (Electronic)
1791-2997 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/29138827 >.
GARRIDO, G. et al. Analgesic and anti-inflammatory effects of Mangifera indica L. extract (Vimang). Phytother Res, v. 15, n. 1, p. 18-21, Feb 2001. ISSN 0951-418X (Print)
0951-418X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/11180516 >.
GAZZINELLI, R. T. Innate and Acquired Immunity in the Pathogenesis of Chagas Disease AU - Golgher, Denise. Autoimmunity, v. 37, n. 5, p. 399-409, 2004/08/01 2004. ISSN 0891-6934. Disponível em: < https://doi.org/10.1080/08916930410001713115 >.
GONG, Z. et al. Compound Libraries: Recent Advances and Their Applications in Drug Discovery. Curr Drug Discov Technol, v. 14, n. 4, p. 216-228, 2017. ISSN 1875-6220 (Electronic)
1570-1638 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28443514 >.
GRIFFITH, J. W.; SOKOL, C. L.; LUSTER, A. D. Chemokines and chemokine receptors: positioning cells for host defense and immunity. Annu Rev Immunol, v. 32, p. 659-702, 2014. ISSN 1545-3278 (Electronic)
0732-0582 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24655300 >.

84
GUEDES, P. M. et al. Inflammation Enhances the Risks of Stroke and Death in Chronic Chagas Disease Patients. PLoS Negl Trop Dis, v. 10, n. 4, p. e0004669, Apr 2016. ISSN 1935-2735 (Electronic)
1935-2727 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/27115869 >.
GUEDES, P. M. et al. Increased type 1 chemokine expression in experimental Chagas disease correlates with cardiac pathology in beagle dogs. Vet Immunol Immunopathol, v. 138, n. 1-2, p. 106-13, Nov 15 2010. ISSN 1873-2534 (Electronic)
0165-2427 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20619467 >.
GUEVARA, P. et al. Expression of fluorescent genes in Trypanosoma cruzi and Trypanosoma rangeli (Kinetoplastida: Trypanosomatidae): its application to parasite-vector biology. J Med Entomol, v. 42, n. 1, p. 48-56, Jan 2005. ISSN 0022-2585 (Print)
0022-2585 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/15691008 >.
GUTIERREZ-OROZCO, F.; FAILLA, M. L. Biological activities and bioavailability of mangosteen xanthones: a critical review of the current evidence. Nutrients, v. 5, n. 8, p. 3163-83, Aug 13 2013. ISSN 2072-6643 (Electronic)
2072-6643 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23945675 >.
GUTIERREZ, F. R. et al. The effects of nitric oxide on the immune system during Trypanosoma cruzi infection %J Memórias do Instituto Oswaldo Cruz. v. 104, p. 236-245, 2009. ISSN 0074-0276. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0074-02762009000900030&nrm=iso >.
HEAD, M. Pharmacokinetic evaluation of vadimezan (ASA404, 5,6-dimethylxanthenone-4-acetic acid, DMXAA) AU - Jameson, Michael B. Expert Opinion on Drug Metabolism & Toxicology, v. 7, n. 10, p. 1315-1326, 2011/10/01 2011. ISSN 1742-5255. Disponível em: < https://doi.org/10.1517/17425255.2011.614389 >.
HEADLAND, S. E.; NORLING, L. V. The resolution of inflammation: Principles and challenges. Semin Immunol, v. 27, n. 3, p. 149-60, May 2015. ISSN 1096-3618 (Electronic)
1044-5323 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25911383 >.
HILDERBRAND, S. A.; WEISSLEDER, R. One-Pot Synthesis of New Symmetric and Asymmetric Xanthene Dyes. Tetrahedron Lett, v. 48, n. 25, p. 4383-4385, Jun 18 2007. ISSN 0040-4039 (Print)
0040-4039 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/19834587 >.
HOSTETTMAN, K., & HOSTETTMANN, M. Xanthones. In: HARBORNE., P. M. D. A. J. B. (Ed.). Methods in plant biochemistry London, UK: Academic Press, v.1, 1989.

85
HSU, M. F. et al. Examination of the inhibitory effect of norathyriol in formylmethionyl-leucyl-phenylalanine-induced respiratory burst in rat neutrophils. Free Radic Biol Med, v. 23, n. 7, p. 1035-45, 1997. ISSN 0891-5849 (Print)
0891-5849 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9358247 >.
HYLAND, K. V. et al. Bioluminescent imaging of Trypanosoma cruzi infection. Int J Parasitol, v. 38, n. 12, p. 1391-400, Oct 2008. ISSN 1879-0135 (Electronic)
0020-7519 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18511053 >.
IGNATUSHCHENKO MV, W. R., RISCOE M. Xanthones as antimalarial agents: stage specificity. Am J Trop Med Hyg, v. 61, n. 1, p. 77-81, 2000.
IMRAN, M. et al. Mangiferin: a natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis, v. 16, n. 1, p. 84, May 2 2017. ISSN 1476-511X (Electronic)
1476-511X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28464819 >.
JAFFE, M. Ueber den Niederschlag welchen Pikrinsäure in normalen Harn erzeugt und über eine neue reaction des Kreatinins. Z Physiol Chem., v. 10, p. 391-400, 1986.
JASSAR, A. S. et al. Activation of tumor-associated macrophages by the vascular disrupting agent 5,6-dimethylxanthenone-4-acetic acid induces an effective CD8+ T-cell-mediated antitumor immune response in murine models of lung cancer and mesothelioma. Cancer Res, v. 65, n. 24, p. 11752-61, Dec 15 2005. ISSN 0008-5472 (Print)
0008-5472 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16357188 >.
JEONG, G.-S.; LEE, D.-S.; KIM, Y.-C. Cudratricusxanthone A from Cudrania tricuspidata suppresses pro-inflammatory mediators through expression of anti-inflammatory heme oxygenase-1 in RAW264.7 macrophages. International Immunopharmacology, v. 9, n. 2, p. 241-246, 2009/02/01/ 2009. ISSN 1567-5769. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1567576908003524 >.
JEONG, J.-J. et al. Mangiferin ameliorates colitis by inhibiting IRAK1 phosphorylation in NF-κB and MAPK pathways. European Journal of Pharmacology, v. 740, p. 652-661, 2014/10/05/ 2014. ISSN 0014-2999. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0014299914004555 >.
JIANG, D.-J.; DAI, Z.; LI, Y.-J. Pharmacological Effects of Xanthones as Cardiovascular Protective Agents. v. 22, n. 2, p. 91-102, 2004. Disponível em: < https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1527-3466.2004.tb00133.x >.
JIANG, D. J.; DAI, Z.; LI, Y. J. J. C. D. R. Pharmacological effects of xanthones as cardiovascular protective agents. v. 22, 2004. Disponível em: < https://doi.org/10.1111/j.1527-3466.2004.tb00133.x >.

86
KAYA, M.; DEMIR, E.; BEKCI, H. Synthesis, characterization and antimicrobial activity of novel xanthene sulfonamide and carboxamide derivatives. J Enzyme Inhib Med Chem, v. 28, n. 5, p. 885-93, Oct 2013. ISSN 1475-6374 (Electronic)
1475-6366 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/22803669 >.
KEISER, J.; VARGAS, M.; WINTER, R. Anthelminthic properties of mangostin and mangostin diacetate. Parasitol Int, v. 61, n. 2, p. 369-71, Jun 2012. ISSN 1873-0329 (Electronic)
1383-5769 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/22265670 >.
KELLY, J. X. et al. Optimization of xanthones for antimalarial activity: the 3,6-bis-omega-diethylaminoalkoxyxanthone series. Antimicrob Agents Chemother, v. 46, n. 1, p. 144-50, Jan 2002. ISSN 0066-4804 (Print)
0066-4804 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/11751125 >.
KNIGHT CG, S. T. Xanthene-dye-labelled phosphatidylethanolamines as
probes of interfacial pH. Studies in phospholipid vesicles. Biochem. Journal v. 15, n. 3, p. 683-687, 1989.
KUAN, L. C. Synthesis and characterisation of 1,3-dihydroxyxanthone derivatives and their antioxidant activities. 2012. (BACHELOR OF SCIENCE (HONS.) CHEMISTRY)
LACEY, D. L. et al. Osteoprotegerin Ligand Is a Cytokine that Regulates Osteoclast Differentiation and Activation. Cell, v. 93, n. 2, p. 165-176, 1998. ISSN 0092-8674. Disponível em: < https://doi.org/10.1016/S0092-8674(00)81569-X >. Acesso em: 2019/01/22.
LEE, H. Z. et al. Decreased protein kinase C activation mediates inhibitory effect of norathyriol on serotonin-mediated endothelial permeability. European journal of pharmacology, v. 353, n. 2-3, p. 303-313, 1998/07// 1998. ISSN 0014-2999. Disponível em: < http://europepmc.org/abstract/MED/9726661
https://doi.org/10.1016/S0014-2999(98)00385-9 >.
LEITE, A. L. J. et al. The immunomodulatory effects of the Enalapril in combination with Benznidazole during acute and chronic phases of the experimental infection with Trypanosoma cruzi. Acta Trop, v. 174, p. 136-145, Oct 2017. ISSN 1873-6254 (Electronic)
0001-706X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28720491 >.
LIBBY, P. Inflammatory mechanisms: the molecular basis of inflammation and disease. Nutr Rev, v. 65, n. 12 Pt 2, p. S140-6, Dec 2007. ISSN 0029-6643 (Print)
0029-6643 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18240538 >.

87
LIBROWSKI, T. et al. New xanthone derivatives as potent anti-inflammatory agents. Medicina (Kaunas), v. 41, n. 1, p. 54-8, 2005. ISSN 1648-9144 (Electronic)
1010-660X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/15687751 >.
LIN, C. N. et al. Synthesis and anti-inflammatory effects of xanthone derivatives. J Pharm Pharmacol, v. 48, n. 5, p. 532-8, May 1996. ISSN 0022-3573 (Print)
0022-3573 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/8799882 >.
LIU, S. H. et al. Effects of alpha-mangostin on the expression of anti-inflammatory genes in U937 cells. Chin Med, v. 7, n. 1, p. 19, Aug 24 2012. ISSN 1749-8546 (Electronic)
1749-8546 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/22920833 >.
LOMBARDINO, J. G.; LOWE III, J. A. The role of the medicinal chemist in drug discovery — then and now. Nature Reviews Drug Discovery, v. 3, p. 853, 10/01/online 2004. Disponível em: < https://doi.org/10.1038/nrd1523 >.
LOPACZYNSKI, W.; ZEISEL, S. H. Antioxidants, programmed cell death, and cancer. Nutrition Research, v. 21, n. 1, p. 295-307, 2001/01/01/ 2001. ISSN 0271-5317. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0271531700002888 >.
MA, J. et al. Gambogic acid inhibits osteoclast formation and ovariectomy-induced osteoporosis by suppressing the JNK, p38 and Akt signalling pathways. Biochem J, v. 469, n. 3, p. 399-408, Aug 1 2015. ISSN 1470-8728 (Electronic)
0264-6021 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26205493 >.
MADAN, B. et al. Xanthones as inhibitors of microsomal lipid peroxidation and TNF-α induced ICAM-1 expression on human umbilical vein endothelial cells (HUVECs). Bioorganic & Medicinal Chemistry, v. 10, n. 11, p. 3431-3436, 2002/11/01/ 2002. ISSN 0968-0896. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0968089602002626 >.
MALIK, L. H.; SINGH, G. D.; AMSTERDAM, E. A. Chagas Heart Disease: An Update. Am J Med, v. 128, n. 11, p. 1251 e7-9, Nov 2015. ISSN 1555-7162 (Electronic)
0002-9343 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26052027 >.
MARIN-NETO, J. A. et al. Pathogenesis of chronic Chagas heart disease. Circulation, v. 115, n. 9, p. 1109-23, Mar 6 2007. ISSN 1524-4539 (Electronic)
0009-7322 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17339569 >.
MARINO ANA PAULA, M. P. et al. Regulated on Activation, Normal T Cell Expressed and Secreted (RANTES) Antagonist (Met-RANTES) Controls the Early Phase of Trypanosoma cruzi–Elicited Myocarditis. Circulation, v. 110, n. 11, p. 1443-1449, 2004/09/14 2004. Disponível em: < https://doi.org/10.1161/01.CIR.0000141561.15939.EC >. Acesso em: 2019/02/13.

88
MARONA, H. et al. Antiarrhythmic and antihypertensive activity of some xanthone derivatives. Acta Pol Pharm, v. 65, n. 3, p. 383-90, May-Jun 2008. ISSN 0001-6837 (Print)
0001-6837 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18646559 >.
MAUSS, H. Über basisch substituierte Xanthon- und Thioxanthon-Abkömmlinge; Miracil, ein neues Chemotherapeuticum. v. 81, n. 1, p. 19-31, 1948. Disponível em: < https://onlinelibrary.wiley.com/doi/abs/10.1002/cber.19480810104 >.
MCCULLOUGH, L. D. et al. NIH initiative to balance sex of animals in preclinical studies: generative questions to guide policy, implementation, and metrics. Biology of sex differences, v. 5, p. 15-15, 2014. ISSN 2042-6410. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25780556
https://www.ncbi.nlm.nih.gov/pmc/PMC4360141/ >.
MENENDEZ, C. A. et al. Design, synthesis and biological evaluation of 1,3-dihydroxyxanthone derivatives: Effective agents against acetylcholinesterase. Bioorg Chem, v. 75, p. 201-209, Dec 2017. ISSN 1090-2120 (Electronic)
0045-2068 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28963904 >.
MOSMANN, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods, v. 65, n. 1-2, p. 55-63, Dec 16 1983. ISSN 0022-1759 (Print)
0022-1759 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/6606682 >.
MUELAS-SERRANO, S.; NOGAL-RUIZ, J. J.; GOMEZ-BARRIO, A. Setting of a colorimetric method to determine the viability of Trypanosoma cruzi epimastigotes. Parasitol Res, v. 86, n. 12, p. 999-1002, Dec 2000. ISSN 0932-0113 (Print)
0932-0113 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/11133116 >.
MUHAMMAD NISAR, I. A., MUHAMMAD RAZA SHAH, AMIR BADSHAH, MUGHAL QAYUM, HUMAYUN KHAN, INAMULLAH KHAND AND SAJID ALIA Amberlite IR-120H as a recyclable catalyst for the synthesis of 1,8-dioxo-octahydroxanthene analogs and their evaluation as potential leishmanicidal agents. RSC Advances, v. 3, n. 44, 2013.
MULAKAYALA, N. et al. Catalysis by molecular iodine: a rapid synthesis of 1,8-dioxo-octahydroxanthenes and their evaluation as potential anticancer agents. Bioorg Med Chem Lett, v. 22, n. 6, p. 2186-91, Mar 15 2012. ISSN 1464-3405 (Electronic)
0960-894X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/22365759 >.
MURDOCH, C.; FINN, A. Chemokine receptors and their role in inflammation and infectious diseases. Blood, v. 95, n. 10, p. 3032, 2000. Disponível em: < http://www.bloodjournal.org/content/95/10/3032.abstract >.

89
NABIH, I. Structure and activity in the schistosomicidal thiaxanthone and xanthone derivatives. v. 55, n. 2, p. 221-222, 1966. Disponível em: < https://onlinelibrary.wiley.com/doi/abs/10.1002/jps.2600550224 >.
NAKATANI, K. et al. Inhibitions of Histamine Release and Prostaglandin E<sub>2</sub> Synthesis by Mangosteen, a Thai Medicinal Plant. Biological and Pharmaceutical Bulletin, v. 25, n. 9, p. 1137-1141, 2002.
NARASIMHA MURTHY, Y. L. et al. Synthesis and Antibacterial Assay of 9-Substituted Aryl-1,8-dioxo-octahydroxanthenes. Asian Journal of Chemistry, v. 26, n. 15, p. 4594-4598, 2014. ISSN 09707077
0975427X.
NETEA, M. G. et al. A guiding map for inflammation. Nature Immunology, v. 18, p. 826, 07/19/online 2017. Disponível em: < https://doi.org/10.1038/ni.3790 >.
NISAR, M. et al. Amberlite IR-120H as a recyclable catalyst for the synthesis of 1,8-dioxo-octahydroxanthene analogs and their evaluation as potential leishmanicidal agents. RSC Advances, v. 3, n. 44, p. 21753-21758, 2013. Disponível em: < http://dx.doi.org/10.1039/C3RA43506G >.
NOVAES, R. D. et al. Trypanosoma cruzi infection induces morphological reorganization of the myocardium parenchyma and stroma, and modifies the mechanical properties of atrial and ventricular cardiomyocytes in rats. Cardiovasc Pathol, v. 22, n. 4, p. 270-9, Jul-Aug 2013. ISSN 1879-1336 (Electronic)
1054-8807 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23541389 >.
NOVAES, R. D. et al. Trypanosoma cruzi infection and benznidazole therapy independently stimulate oxidative status and structural pathological remodeling of the liver tissue in mice. Parasitology Research, v. 114, n. 8, p. 2873-2881, 2015/08/01 2015. ISSN 1432-1955. Disponível em: < https://doi.org/10.1007/s00436-015-4488-x >.
NOVAES, R. D. et al. Curcumin Enhances the Anti-Trypanosoma cruzi Activity of Benznidazole-Based Chemotherapy in Acute Experimental Chagas Disease. Antimicrob Agents Chemother, v. 60, n. 6, p. 3355-64, Jun 2016. ISSN 1098-6596 (Electronic)
0066-4804 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/27001816 >.
NUNEZ A, P. E., AMARO D, ACOSTA J, AGU¨ERO J, CAPOTE R, GARCIGA MR, MORALES I, GARCı´A O, GARRIDO G, MARTı´NEZ G, MORALES MA. Composiciones farmace´uticas y suplementos nutricionales a partir de
extractos de Mangifera indica L. OCPI. Cuba. 22846 2002.

90
NWAKA, S.; HUDSON, A. Innovative lead discovery strategies for tropical diseases. Nature Reviews Drug Discovery, v. 5, p. 941, 11/01/online 2006. Disponível em: < https://doi.org/10.1038/nrd2144 >.
OSORIO-MÉNDEZ, J. F.; CEVALLOS, A. M. Discovery and Genetic Validation of Chemotherapeutic Targets for Chagas' Disease. v. 8, n. 439, 2019-January-07 2019. ISSN 2235-2988. Disponível em: < https://www.frontiersin.org/article/10.3389/fcimb.2018.00439 >.
OUYANG, W. et al. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu Rev Immunol, v. 29, p. 71-109, 2011. ISSN 1545-3278 (Electronic)
0732-0582 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21166540 >.
PAL, P. B.; SINHA, K.; SIL, P. C. Mangiferin, a natural xanthone, protects murine liver in Pb(II) induced hepatic damage and cell death via MAP kinase, NF-kappaB and mitochondria dependent pathways. PLoS One, v. 8, n. 2, p. e56894, 2013. ISSN 1932-6203 (Electronic)
1932-6203 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23451106 >.
PARDO-ANDREU, G. L. et al. Mangifera indica L. (Vimang) protection against serum oxidative stress in elderly humans. Arch Med Res, v. 37, n. 1, p. 158-64, Jan 2006. ISSN 0188-4409 (Print)
0188-4409 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16314203 >.
PEDRO, M. et al. Xanthones as inhibitors of growth of human cancer cell lines and their effects on the proliferation of human lymphocytes in vitro. Bioorg Med Chem, v. 10, n. 12, p. 3725-30, Dec 2002. ISSN 0968-0896 (Print)
0968-0896 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/12413829 >.
PENAS, F. N. et al. Hepatic injury associated with Trypanosoma cruzi infection is attenuated by treatment with 15-deoxy-Δ12,14 prostaglandin J2. Experimental Parasitology, v. 170, p. 100-108, 2016/11/01/ 2016. ISSN 0014-4894. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0014489416302090 >.
PENITENTE, A. R. et al. Enalapril in Combination with Benznidazole Reduces Cardiac Inflammation and Creatine Kinases in Mice Chronically Infected with Trypanosoma cruzi. Am J Trop Med Hyg, v. 93, n. 5, p. 976-82, Nov 2015. ISSN 1476-1645 (Electronic)
0002-9637 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26350447 >.
PETIWALA, S. M. et al. Pharmacokinetic characterization of mangosteen (Garcinia mangostana) fruit extract standardized to α-mangostin in C57BL/6 mice. Nutrition research (New York, N.Y.), v. 34, n. 4, p. 336-345, 2014. ISSN 1879-0739
0271-5317. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24774070
https://www.ncbi.nlm.nih.gov/pmc/PMC6298434/ >.

91
PFISTER, J. R. et al. Xanthone-2-carboxylic acids, a new series of antiallergic substances. J Med Chem, v. 15, n. 10, p. 1032-5, Oct 1972. ISSN 0022-2623 (Print)
0022-2623 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/5069771 >.
PINAZO, M. J. et al. Biomarkers of therapeutic responses in chronic Chagas disease: state of the art and future perspectives. Mem Inst Oswaldo Cruz, v. 110, n. 3, p. 422-32, May 2015. ISSN 1678-8060 (Electronic)
0074-0276 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25946151 >.
PINTO, M. M.; SOUSA, M. E.; NASCIMENTO, M. S. Xanthone derivatives: new insights in biological activities. Curr Med Chem, v. 12, n. 21, p. 2517-38, 2005. ISSN 0929-8673 (Print)
0929-8673 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16250875 >.
POHLERS, G.; SCAIANO, J. C.; SINTA, R. A Novel Photometric Method for the Determination of Photoacid Generation Efficiencies Using Benzothiazole and Xanthene Dyes as Acid Sensors. Chemistry of Materials, v. 9, n. 12, p. 3222-3230, 1997/12/01 1997. ISSN 0897-4756. Disponível em: < https://doi.org/10.1021/cm970587p >.
PONTIUS, A. et al. Antiprotozoal Activities of Heterocyclic-Substituted Xanthones from the Marine-Derived Fungus Chaetomium sp. Journal of Natural Products, v. 71, n. 9, p. 1579-1584, 2008/09/01 2008. ISSN 0163-3864. Disponível em: < https://doi.org/10.1021/np800294q >.
POUPELIN, J. P. S.-R., G.; FOUSSARD-BLANPIN, O.; NARCISSE, G.; UCHIDA-ERNOUF, G.; LACROIX, R. Synthesis and anti inflammatory properties of bis 2 hydroxy 1 naphthyl methane derivatives. European Journal of Medicinal Chemistry, v. 13, n. 1, p. 67-71, 1978.
PRANTNER D, P. D., LAI W, WILLIAMS MS, SHARMA S, FITZGERALD KA, VOGEL SN. 5,6-Dimethylxanthenone-4-acetic acid (DMXAA) activates stimulator of interferon gene (STING)-dependent innate immune pathways and is regulated by mitochondrial membrane potential. J Biol Chem., v. 16, n. 287, p. 39776-88, 2012.
PRATA, A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect Dis, v. 1, n. 2, p. 92-100, Sep 2001. ISSN 1473-3099 (Print)
1473-3099 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/11871482 >.
QUIN, L. D., TYRELL, J. Fundamentals of Heterocyclic Chemistry: Importance in Nature and in the Synthesis of Pharmaceuticals. . New Jersey: 2010. 344.
RAHIMIFARD, M. et al. One-pot solvent-free synthesis of 1,8-dioxo-octahydroxanthene derivatives using sulfonic acid-functionalized LUS-1 and their antimicrobial activities. Research on Chemical Intermediates, v. 42, n. 4, p. 3847-3861, 2015. ISSN 0922-6168
1568-5675.

92
RAMPERSAD, S. N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors (Basel), v. 12, n. 9, p. 12347-60, 2012. ISSN 1424-8220 (Electronic)
1424-8220 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23112716 >.
RAPACZ, A. et al. Antiarrhythmic activity of some xanthone derivatives with β1-adrenoceptor affinities in rats. European Journal of Pharmacology, v. 738, p. 14-21, 2014/09/05/ 2014. ISSN 0014-2999. Disponível em: < http://www.sciencedirect.com/science/article/pii/S001429991400394X >.
RASSAMEEMASMAUNG, S. et al. Topical application of Garcinia mangostana L. pericarp gel as an adjunct to periodontal treatment. Complementary Therapies in Medicine, v. 16, n. 5, p. 262-267, 2008/10/01/ 2008. ISSN 0965-2299. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0965229908000320 >.
REDDI MOHAN NAIDU, K. et al. Design, synthesis and antiviral potential of 14-aryl/heteroaryl-14H-dibenzo[a,j]xanthenes using an efficient polymer-supported catalyst. Molecules, v. 17, n. 6, p. 7543-55, Jun 18 2012. ISSN 1420-3049 (Electronic)
1420-3049 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/22710828 >.
REED, S. G. et al. IL-10 mediates susceptibility to Trypanosoma cruzi infection. The Journal of Immunology, v. 153, n. 7, p. 3135, 1994. Disponível em: < http://www.jimmunol.org/content/153/7/3135.abstract >.
REITMAN S, F. S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am J Clin Pathol., v. 1 p. 56-63, 1957.
RIVERA, D. G. et al. Mangifera indica L. extract (Vimang) and mangiferin reduce the airway inflammation and Th2 cytokines in murine model of allergic asthma. J Pharm Pharmacol, v. 63, n. 10, p. 1336-45, Oct 2011. ISSN 2042-7158 (Electronic)
0022-3573 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21899550 >.
ROBERTS, Z. J. et al. The chemotherapeutic agent DMXAA potently and specifically activates the TBK1-IRF-3 signaling axis. J Exp Med, v. 204, n. 7, p. 1559-69, Jul 9 2007. ISSN 0022-1007 (Print)
0022-1007 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17562815 >.
ROCHA E SILVA, M. A brief survey of the history of inflammation. Agents and Actions, v. 8, n. 1, p. 45-49, 1978/01/01 1978. ISSN 1420-908X. Disponível em: < https://doi.org/10.1007/BF01972401 >.
RODRIQUES COURA, J.; DE CASTRO, S. L. A critical review on Chagas disease chemotherapy. Mem Inst Oswaldo Cruz, v. 97, n. 1, p. 3-24, Jan 2002. ISSN 0074-0276 (Print)
0074-0276 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/11992141 >.

93
ROLON, M. et al. Development of resazurin microtiter assay for drug sensibility testing of Trypanosoma cruzi epimastigotes. Parasitol Res, v. 99, n. 2, p. 103-7, Jul 2006. ISSN 0932-0113 (Print)
0932-0113 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16506080 >.
ROMANHA, A. J. et al. In vitro and in vivo experimental models for drug screening and development for Chagas disease %J Memórias do Instituto Oswaldo Cruz. v. 105, p. 233-238, 2010. ISSN 0074-0276. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0074-02762010000200022&nrm=iso >.
ROSSI, M. A.; MENGEL, J. O. Patogênese da miocardite chagásica crônica: o papel de fatores autoimunes e microvasculares %J Revista do Instituto de Medicina Tropical de São Paulo. v. 34, p. 593-599, 1992. ISSN 0036-4665. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0036-46651992000600015&nrm=iso >.
SAHA, S.; SADHUKHAN, P.; SIL, P. C. Mangiferin: A xanthonoid with multipotent anti-inflammatory potential. Biofactors, v. 42, n. 5, p. 459-474, Sep 10 2016. ISSN 1872-8081 (Electronic)
0951-6433 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/27219011 >.
SALOMON, C. J. First Century of Chagas' Disease: An Overview on Novel Approaches to Nifurtimox and Benznidazole Delivery Systems. Journal of Pharmaceutical Sciences, v. 101, n. 3, p. 888-894, 2012. ISSN 0022-3549. Disponível em: < https://doi.org/10.1002/jps.23010 >. Acesso em: 2019/01/27.
SANCHEZ-VALDEZ, F. J. et al. Spontaneous dormancy protects Trypanosoma cruzi during extended drug exposure. Elife, v. 7, Mar 26 2018. ISSN 2050-084X (Electronic)
2050-084X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/29578409 >.
SÁNCHEZ-VALDÉZ, F. J. et al. Spontaneous dormancy protects Trypanosoma cruzi during extended drug exposure. eLife, v. 7, p. e34039, 2018/03/26 2018. ISSN 2050-084X. Disponível em: < https://doi.org/10.7554/eLife.34039 >.
SAUER, N. N. Green Chemistry: Frontiers in Benign Chemical Syntheses and Processes Edited by Paul T. Anastas and Tracy C. Williamson (U. S. Environmental Protection Agency). Oxford University Press: New York, NY. 1999. 360 pp. $115.00. ISBN 0-19-850170-6. Journal of the American Chemical Society, v. 122, n. 22, p. 5419-5420, 2000/06/01 2000. ISSN 0002-7863. Disponível em: < https://doi.org/10.1021/ja995756g >.
SCARIM, C. B. et al. Current advances in drug discovery for Chagas disease. European Journal of Medicinal Chemistry, v. 155, p. 824-838, 2018/07/15/ 2018. ISSN 0223-5234. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0223523418305312 >.

94
SCHNEIDER, J. et al. Synthesis and biological activity of acronycine analogs. J Med Chem, v. 15, n. 3, p. 266-70, Mar 1972. ISSN 0022-2623 (Print)
0022-2623 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/5062070 >.
SHAGUFTA; AHMAD, I. Recent insight into the biological activities of synthetic xanthone derivatives. Eur J Med Chem, v. 116, p. 267-280, Jun 30 2016. ISSN 1768-3254 (Electronic)
0223-5234 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/27111599 >.
SHCHEKOTIKHIN, Y. M. N., T.G. Transformations of sym-octahydroxanthene-1,8-diones and 1,8-dioxo-sym-octahydroxanthylium salts in recyclization under the influence of amines. Chemistry of Heterocyclic Compounds, v. 42, n. 1, p. 28-33, 2002. ISSN 1573-8353.
SUGIMOTO, M. A. et al. Mediators of the Resolution of the Inflammatory Response. Trends in Immunology, v. 40, n. 3, p. 212-227, 2019. ISSN 1471-4906. Disponível em: < https://doi.org/10.1016/j.it.2019.01.007 >. Acesso em: 2019/05/06.
SUZUKI, O. et al. Inhibition of type A and type B monoamine oxidases by naturally occurring xanthones. Planta Med, v. 42, n. 1, p. 17-21, May 1981. ISSN 0032-0943 (Print)
0032-0943 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/7255567 >.
SZKARADEK, N. et al. Cardiovascular activity of the chiral xanthone derivatives. Bioorganic & Medicinal Chemistry, v. 23, n. 20, p. 6714-6724, 2015/10/15/ 2015. ISSN 0968-0896. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0968089615300316 >.
TALVANI, A.; TEIXEIRA, M. M. Chapter 8 - Inflammation and Chagas Disease: Some Mechanisms and Relevance. In: WEISS, L. M. e TANOWITZ, H. B. (Ed.). Advances in Parasitology: Academic Press, v.76, 2011. p.171-194. ISBN 0065-308X.
TANG, Y.-P. et al. Effect of a Mangosteen Dietary Supplement on Human Immune Function: A Randomized, Double-Blind, Placebo-Controlled Trial. v. 12, n. 4, p. 755-763, 2009. Disponível em: < https://www.liebertpub.com/doi/abs/10.1089/jmf.2008.0204 >.
TARLETON, R. L. Chagas disease: a role for autoimmunity? Trends in Parasitology, v. 19, n. 10, p. 447-451, 2003/10/01/ 2003. ISSN 1471-4922. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1471492203002228 >.
TARLETON, R. L. Trypanosoma cruzi and Chagas Disease: Cause and Effect. In: TYLER, K. M. e MILES, M. A. (Ed.). American Trypanosomiasis. Boston, MA: Springer US, 2003. p.107-115. ISBN 978-1-4419-9206-2.
TARLETON, R. L. et al. The challenges of Chagas Disease-- grim outlook or glimmer of hope. PLoS Med, v. 4, n. 12, p. e332, Dec 2007. ISSN 1549-1676 (Electronic)

95
1549-1277 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18162039 >.
TEIXEIRA, M. M.; GAZZINELLI, R. T.; SILVA, J. S. Chemokines, inflammation and <em>Trypanosoma cruzi</em> infection. Trends in Parasitology, v. 18, n. 6, p. 262-265, 2002. ISSN 1471-4922. Disponível em: < https://doi.org/10.1016/S1471-4922(02)02283-3 >. Acesso em: 2019/01/27.
TEWTRAKUL, S.; WATTANAPIROMSAKUL, C.; MAHABUSARAKAM, W. Effects of compounds from Garcinia mangostana on inflammatory mediators in RAW264.7 macrophage cells. Journal of Ethnopharmacology, v. 121, n. 3, p. 379-382, 2009/01/30/ 2009. ISSN 0378-8741. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0378874108006375 >.
THEEL, E. S.; PRITT, B. S. Parasites. v. 4, n. 4, 2016. Disponível em: < http://www.asmscience.org/content/journal/microbiolspec/10.1128/microbiolspec.DMIH2-0013-2015 >.
TJAHJANI, S. Antimalarial activity of Garcinia mangostana L rind and its synergistic effect with artemisinin in vitro. BMC Complement Altern Med, v. 17, n. 1, p. 131, Feb 28 2017. ISSN 1472-6882 (Electronic)
1472-6882 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28241761 >.
UDANI, J. K. et al. Evaluation of Mangosteen juice blend on biomarkers of inflammation in obese subjects: a pilot, dose finding study. v. 8, n. 1, p. 48, October 20 2009. ISSN 1475-2891. Disponível em: < https://doi.org/10.1186/1475-2891-8-48 >.
URBINA, J. A. Specific chemotherapy of Chagas disease: Relevance, current limitations and new approaches. Acta Tropica, v. 115, n. 1, p. 55-68, 2010/07/01/ 2010. ISSN 0001-706X. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0001706X0900357X >.
URBINA, J. A.; DOCAMPO, R. Specific chemotherapy of Chagas disease: controversies and advances. Trends in Parasitology, v. 19, n. 11, p. 495-501, 2003/11/01/ 2003. ISSN 1471-4922. Disponível em: < http://www.sciencedirect.com/science/article/pii/S1471492203002411 >.
VEGA, C. et al. A new pharmacological screening assay with Trypanosoma cruzi epimastigotes expressing beta-galactosidase. Parasitol Res, v. 95, n. 4, p. 296-8, Mar 2005. ISSN 0932-0113 (Print)
0932-0113 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/15682334 >.
VENNERSTROM, J. L. et al. Antimalarial dyes revisited: xanthenes, azines, oxazines, and thiazines. Antimicrob Agents Chemother, v. 39, n. 12, p. 2671-7, Dec 1995. ISSN 0066-4804 (Print)
0066-4804 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/8593000 >.

96
VIEIRA, L. M.; KIJJOA, A. Naturally-occurring xanthones: recent developments. Curr Med Chem, v. 12, n. 21, p. 2413-46, 2005. ISSN 0929-8673 (Print)
0929-8673 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/16250871 >.
VIOTTI, R. et al. Treatment of chronic Chagas' disease with benznidazole: Clinical and serologic evolution of patients with long-term follow-up. American Heart Journal, v. 127, n. 1, p. 151-162, 1994/01/01/ 1994. ISSN 0002-8703. Disponível em: < http://www.sciencedirect.com/science/article/pii/0002870394905215 >.
WENDER, P. A. H., S. L.; WRIGHT, D. L. . Towards the ideal synthesis. Chem. Ind., p. 765-769, 1997.
WILKINSON, S. R. et al. A mechanism for cross-resistance to nifurtimox and benznidazole in trypanosomes. Proceedings of the National Academy of Sciences, v. 105, n. 13, p. 5022, 2008. Disponível em: < http://www.pnas.org/content/105/13/5022.abstract >.
WINTER, R. W. et al. Potentiation of an antimalarial oxidant drug. Antimicrob Agents Chemother, v. 41, n. 7, p. 1449-54, Jul 1997. ISSN 0066-4804 (Print)
0066-4804 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9210664 >.
XU, D. et al. Evaluation of high efficiency gene knockout strategies for Trypanosoma cruzi. BMC Microbiol, v. 9, p. 90, May 11 2009. ISSN 1471-2180 (Electronic)
1471-2180 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/19432966 >.
XU KELLY, J. et al. A spectroscopic investigation of the binding interactions between 4,5-dihydroxyxanthone and heme. Journal of Inorganic Biochemistry, v. 86, n. 2, p. 617-625, 2001/09/01/ 2001. ISSN 0162-0134. Disponível em: < http://www.sciencedirect.com/science/article/pii/S0162013401002173 >.
YAN, W.; MORENO, S. N. A method to assess invasion and intracellular replication of Trypanosoma cruzi based on differential uracil incorporation. J Immunol Methods, v. 220, n. 1-2, p. 123-8, Nov 1 1998. ISSN 0022-1759 (Print)
0022-1759 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9839933 >.
YANG CHUN-HUI, M. L., WEI ZHEN-PING, HAN FENG, GAO JING. Advances in Isolation and Synthesis of Xanthone Derivatives. Chinese Herbal Medicines, v. 4, n. 2, p. 87-102, 2012. ISSN 1674-6384.2012.02.003.
ZARENA, A. S.; SANKAR, K. U. Screening of xanthone from mangosteen (Garcinia mangostana L.) peels and their effect on cytochrome c reductase and phosphomolybdenum activity. Journal of Natural Products (India), Uttar Pradesh, v. 2, p. 23-30, 2009. ISSN 0974-5211.

97
ZHANG, Q.; LENARDO, M. J.; BALTIMORE, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell, v. 168, n. 1-2, p. 37-57, Jan 12 2017. ISSN 1097-4172 (Electronic)
0092-8674 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/28086098 >.
ZINGALES, B. et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: second revision meeting recommends TcI to TcVI %J Memórias do Instituto Oswaldo Cruz. v. 104, p. 1051-1054, 2009. ISSN 0074-0276. Disponível em: < http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0074-02762009000700021&nrm=iso >.
ZINGALES, B. et al. Drug discovery for Chagas disease should consider Trypanosoma cruzi strain diversity. Memorias do Instituto Oswaldo Cruz, v. 109, n. 6, p. 828-833, 2014. ISSN 1678-8060
0074-0276. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25317712
https://www.ncbi.nlm.nih.gov/pmc/PMC4238778/ >.
ZINGALES, B. et al. Trypanosoma cruzi genome project: biological characteristics and molecular typing of clone CL Brener. Acta Trop, v. 68, n. 2, p. 159-73, Nov 1997. ISSN 0001-706X (Print)
0001-706X (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9386791 >.
ZUO, J. et al. Xanthone-rich dichloromethane fraction of Securidaca inappendiculata, the possible antirheumatic material base with anti-inflammatory, analgesic, and immunodepressive effects. Pharm Biol, v. 52, n. 11, p. 1367-73, Nov 2014. ISSN 1744-5116 (Electronic)
1388-0209 (Linking). Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25026334 >.