association of intermediate filaments with vinculin-containing adhesion plaques of fibroblasts
TRANSCRIPT

Cell Motility and the Cystoskeleton 8:274-283 (1987)
Association of Intermediate Filaments With Vinculin-Containing Adhesion Plaques of
Fibroblasts
Alexander D. Bershadsky, lrena S. Tint, and Tatjana M. Svitkina
Cancer Research Center, Academy of Medical Sciences (A. D.B.) and Laboratory of Molecular Biology and Bioorganic Chemistry, Moscow State University
(I.S.T., T.M.S.), Moscow, USSR
Double immunofluorescence staining of quail embryo fibroblasts with rabbit antibody to vinculin and mouse monoclonal antibody to vimentin revealed a coincidence between fluorescence patterns for cell-substrate focal contacts and intermediate filaments. Most of the vinculin-containing adhesion plaques coin- cided with the ends of vimentin-positive fibrils.
This association was further corroborated by immunoelection microscopic observations of the cytoskeletons of quail and mouse fibroblasts using a platinum replica technique. The intermediate filaments were identified either by direct treatment with antivimentin IgM or by an indirect immunogold staining method.
Colcemid treatment of the cells caused a collapse of intermediate filaments and destroyed their association with focal contacts. During the early stages of the colcemid-induced collapse of the intermediate filaments, single vimentin fibrils appeared to retain their association with focal contacts.
The possible role of the intermediate filaments in the formation and mainte- nance of focal contacts is discussed.
Key words: focal contacts, vimentin filaments, microtubules, immunofluorescence, platinum replicas
INTRODUCTION
Spreading and locomotion of cultured cells on a solid substrate is accompanied by the formation of nu- merous discrete attachment sites, or focal contacts [Aber- crombie et al., 1971; Abercrombie and Dunn, 1975; Izzard and Lochner, 19761. The cytoplasmic face of a focal contact (adhesion plaque) contains several charac- teristic proteins, in particular, vinculin [Geiger, 1979; Burridge and Feramisco, 19801 and talin [Burridge and Connell, 19831. It is well established that adhesion plaques can be associated with the bundles of actin micro- filaments [Heath and Dunn, 1978; Wehland et al., 19791. Data on the association of adhesion plaques with cytoske- letal elements other than actin bundles are fragmentary. Evidence for a possible interrelationship between focal contacts and microtubules has recently been presented [Small and Rinnerthaler, 19851. A third element of the cytoskeleton, intermediate filaments, has been shown to
be associated with another type of specialized junctional membrane structures, ie, desmosomes [Drochmans et al., 1978; Skerrow and Skerrow, 1980; Kartenbeck et al., 1983, 19841. These structures are different from contacts: desmosomal plaques contain several specific proteins, called desmoplukins [Franke et al., 19821, but do not contain vinculin. In addition, desmosomes are not associated with actin filaments [Geiger et al., 19831.
The present work shows that vinculin-containing adhesion plaques of cultured fibroblasts may be associ- ated not only with bundles of actin filaments but also with vimentin intermediate filaments.
Received December 30, 1985; accepted April 14, 1987.
Address reprint requests to A.D. Bershadsky, Cancer Research Cen- ter, The USSR AMS, 24 Kashirskoye shosse, Moscow 115478, USSR.
0 1987 Alan R. Liss, Inc.

Intermediate Filaments and Focal Contacts 275
Fig. I . Association between intcrmcdiate filaments and focal contacts. Double-immunofluorescence labeling of thc same cell with antibody to vinculin (A) and to vimentin (B). Several corresponding sitcs are marked by the same digits. Bar = 10 pm.

276 Bershadsky, Tint, and Svitkina
MATERIALS AND METHODS Cell cultures
Secondary cultures of quail or mouse fibroblasts were used in all experiments. Cells were cultured in Ham’s F-12 medium or in the mixture (1:l) of 199 medium and 0.5% lactalbumin hydrolysate in Hank’s solution. Quail fibroblast medium was supplemented with 5 % fetal calf serum, 5 % tryptose phosphate broth, and 1% chicken serum; medium for mouse fibroblasts con- tained 10% bovine serum. Cells were grown on glass coverslips for 48 hr before fixation.
lmmunofluorescence
Cells were washed with PBS, extracted with a so- lution containing l % Triton X-100 and 4 % polyethylene glycol 40,000 in buffer I (50 mM imidazole, 50 mM KC1, 2 mM MgC12, 0.2 mM EGTA, and 0.05 mM EDTA, pH 6.8) for 3 min, washed with buffer 1, and fixed with 4 % formaldehyde in PBS.
Rabbit monospecific antibody to vinculin has been described previously [Neyfakh et al., 19831. Mouse monoclonal antibody to tubulin [Viklicky et al., 19821 was lundly donated by Dr. V. Viklicky (Inst. Mol. Ge- netics, Prague). Two types of mouse monoclonal anti- body to vimentin were used. Antibodies produced by “clone 30” has been described elsewhere [Troyanovsky et al., 19851. Another hybridoma clone (IIID3) was a generous gift of Drs. N. Romanova, L. Yakubov, and 0. Rokhlin (All-Union Cardiology Center, Moscow). We have shown that the culture medium of the IIID3 clone stained a radial filamentous network in the cytoplasm. After colcemid treatment the filaments collapsed in the perinuclear area, as vimentin filaments do. The antibody specifically reacted with a 55 KD polypeptide from ly- sates of quail embryo fibroblasts in immunoblotting.
Double indirect immunofluorescence was per- formed as follows. Fixed cells were incubated with a mixture of first antibodies (antivimentin-antivinculin or antitubulin-antivinculin) for 30 min at room temperature. After washing, a mixture of FITC-conjugated goat anti- rabbit Ig (Sigma, St. Louis, MD) and TRITC-conjugated sheep antimouse Ig (Cappel Laboratories, Cochranville, PA) was applied for 30 min.
When double stained for actin and vimentin, the cells were first incubated with the vimentin antibody and then with a mixture of a FITC-conjugated antibody to
580, and LP 590 respectively, for rhodamine and BP 450-490, FT 510, and BP 520-560, respectively, for fluorescein. These combinations of filters provide a good selectivity with almost no bleed-through from one flu- orescent image to the other in a double-stained prepara- tion. In some cases, however, we used another filter set for FITC (BP 450-490, FT 510, and LP 520) that permits a slight leakage of rhodamine fluorescence. The resulting pictures, with a small amount of bleed-through, help to reveal a correspondence between rhodamine and fluores- cein images.
lmmunoelectron Microscopy
Two methods of immunoelectron microscopic vi- sualization of intermediate filaments on platinum replicas of the cytoskeleton were used. According to the first method, cells were extracted with Triton X-100 in the same way as for immunofluorescence and then were stained with monoclonal antivimentin IgM (clone IIID3) for 15 min in buffer I before fixation. IgM molecules are sufficiently large to be visualized by electron microscopy without any additional markers [Svitkma et al., 19841.
Alternatively, we used an indirect immunogold staining method. In this case cells were extracted with 1 % Triton x-100 in buffer I with 1 % PEG 40,000, fixed with 2 % glutaraldehyde in 0.1 M cacodylate buffer, and treated with NaBH4. The cytoskeletons were stained with mouse monoclonal antivimentin and then with a solution of 5-nm colloidal gold particles coated with goat antibod- ies to mouse immunoglobulin (Janssen Pharmaceutica, Beerse, Belgium). The staining was performed according to the procedure recommended by the manufacturer. In both cases, treated cytoskeletons were fixed in glutaral- dehyde solution after staining and then processed for replication.
The technique of platinum-carbon replication of the cytoskeleton [Heuser and Kirschner, 19801 was used with modifications described previously [Svitkina et al., 19841. Briefly, this procedure included dehydration of speci- mens in graded acetones, critical point drying, and rotary shadowing with platinum and carbon in a Balzers appa- ratus. The replicas were released from the coverslips by hydrofluoric acid treatment and were cleaned with aqueous Cr03. The last step was omitted when immuno- gold staining was used. A Philips EM 400 M electron microscope was used for examination of the replicas.
rabbit Ig and rhodamine-conjugated phalloidin. The ;am- ple of rhodamine-phalloidin used in this work was a generous gift of Dr. T. Wieland (Heidelberg, FRG).
The immunofluorescence specimens were exam- ined in a Zeiss Photomicroscope I11 equipped with a 40/ With Vinculin and Vimentin antibodies
1 .OO Planapo oil immersion objective and fluorescence- Vimentin staining revealed a radial filamentous net- Condensor I11 RS. The excitation filters, dichromatic work in the cytoplasm of fibroblasts. The density of the beam splitters, and barrier filters were BP 510-560, FT network was very high in the perinuclear part of the cell
R ES U LTS Double-lmmunofluorescence Staining of Cells

Fig. 2. Leading edges of two cells double stained with antibodies to vinculin (A,C) and to vimentin (B,D). A,B: Cell having mainly dash- like vinculin-containing focal contacts. C,D: Cell with numerous dot- like contacts at the edge. Several sites of correspondence between the vinculin and vimentin pictures are numbered 1 to 4 in each pair. Note
the association of vimentin-positive fibrils with both types of vinculin- containing contacts. The weak staining of intermediate filaments in C is due to leakage of rhodamine fluorescence through the fluorescein filters (see Materials and Methods) facilitates the search of correspon- dence between dot contacts and vimentin fibrils. Bars = 10 pm.
so that separate fibrillar structures could not be resolved and small, dot-like contacts localized preferentially at the in this region. Thick vimentin-positive cables as well as active edge of the cell and not associated with actin thinner vimentin-positive fibrils (corresponding to small bundles. intermediate filament bundles or possibly even to individ- The combined staining of the cells with both types ual filaments) were distinguishable in the peripheral part of the antibodies revealed an apparent correspondence of the cytoplasm. between vinculin-positive regions (Figs. 1 A, 2A,C) and
The vinculin antibody stained a system of discrete vimentin-containing fibrils (Figs. lB, 2B, D). Unlike regions at the ventral surface of the cell, which have been actin microfilaments, intermediate filaments seemed to shown to correspond to focal contacts visualized by inter- associate not only with dash contacts but sometimes with ference reflection microscopy [Geiger, 19791 or by an dot contacts also (Fig. 2C,D). antibody exclusion method [Neyfakh et al., 19831. As we The correspondence between the focal contacts and have shown previously [Bershadsky et al., 19851, vincu- the ends of vimentin-positive fibrils can be established lin-positive regions can be divided into two major mor- reliably only at the peripheral part of the cell where phological types: elongated, dash-like focal contacts distinct cables and fibrils can be clearly resolved. The associated with the ends of actin microfilament bundles existence of an association between vimentin-positive

278 Bershadsky, Tint, and Svitkina
Fig. 3 . Association between the ends of actin microfilament bundles and intermediate filaments. Double- fluorescence labeling of the same cell with rhodarnine-phalloidin (A) and antibody to vimentin (B). Several sitcs of correspondence are numbered 1 to 4. Bar = 10 pm.
fibrils can be established reliably only at the peripheral part of the cell where distinct cables and fibrils can be clearly resolved. The existence of an association between vimentin filaments and focal contacts localized in the central part of the cell remains to be proved. It should be noted that even in the peripheral part of the cell the coincidence of focal contacts and intermediate filaments was not absolute. Some ends of vimentin-positive fibrils had no apparent association with any type of focal con- tacts and vice-versa. We would like to emphasize, how- ever, that the correspondence between intermediate filaments and both dot and dash focal contacts was very prominent in our experiments.
Double-lmmunofluorescence Staining of Cells With Vimentin Antibody and Rhodamine-Labeled Phalloidin
Since the ends of microfilament bundles appear to be coincident with dash focal contacts, which in turn are associated with vimentin filaments (see above), it seemed reasonable that the ends of the bundles would be also
associated with some vimentin-positive fibrils. We ob- served this association after a combined staining of fibro- blasts with vimentin antibody and rhodamine-labeled phalloidin (Fig. 3A,B). Usually, distal parts of actin microfilament bundles and the bundles of intermediate filaments ran parallel to each other for a considerable distance. Sometimes an actin bundle and a bundle of intermediate filaments radiated from the same focal con- tact, but their orientation was slightly different.
Double-lmmunofluorescence Staining of Cells With Vinculin and Tubulin Antibodies
The staining of cells with tubulin antibody revealed a radial microtubular system usual for fibroblasts. Some microtubule ends were perpendicular to the cell edges, and others curved along the edges for a considerable distance. Combined staining of the quail cells with anti- tubulin and antivinculin demonstrated that the free ends of some microtubules were localized in close proximity to the focal contacts of both dot and dash types (Fig. 4A,B). On the other hand, some focal contacts were
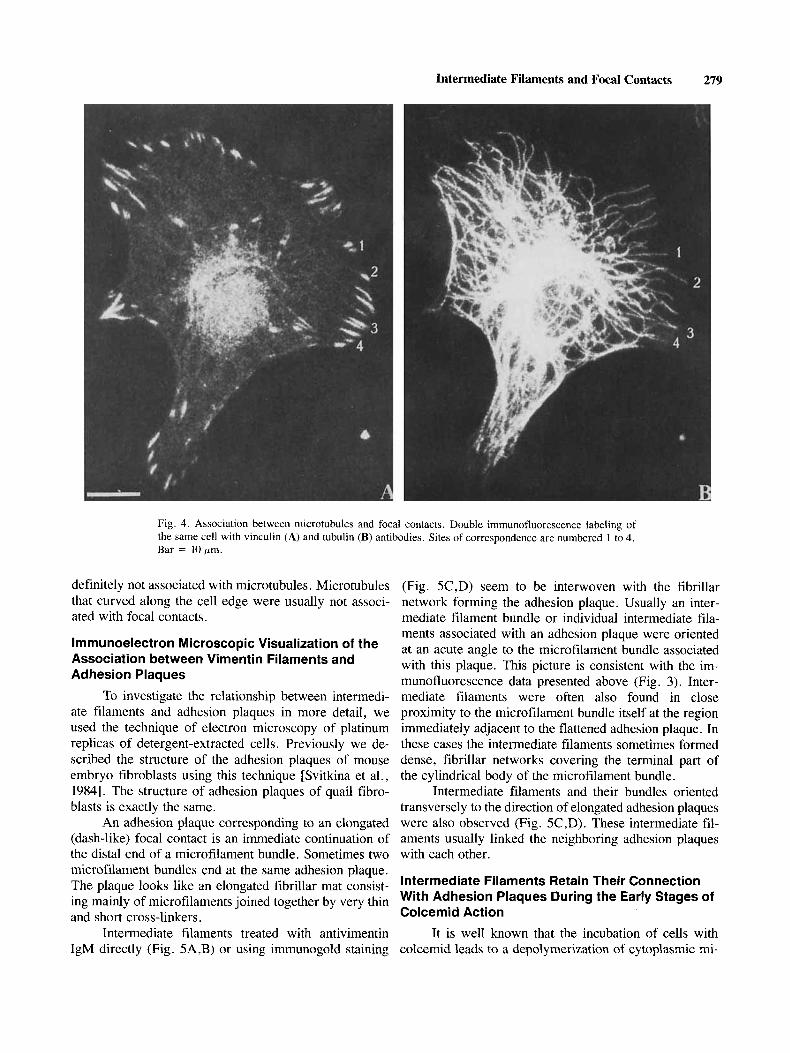
Intermediate Filaments and Focal Contacts 279
Fig. 4. Association between microtubules and focal contacts. Double immunofluorescence labeling of the same cell with vinculin (A) and tubulin IB) antibodies. Sites of correspondence are numbered 1 to 4. Bar = 10urn.
definitely not associated with microtubules. Microtubules that curved along the cell edge were usually not associ- ated with focal contacts.
lmmunoelectron Microscopic Visualization of the Association between Vimentin Filaments and Adhesion Plaques
To investigate the relationship between intermedi- ate filaments and adhesion plaques in more detail, we used the technique of electron microscopy of platinum replicas of detergent-extracted cells. Previously we de- scribed the structure of the adhesion plaques of mouse embryo fibroblasts using this technique [Svitkina et al., 19841. The structure of adhesion plaques of quail fibro- blasts is exactly the same.
An adhesion plaque corresponding to an elongated (dash-like) focal contact is an immediate continuation of the distal end of a microfilament bundle. Sometimes two microfilament bundles end at the same adhesion plaque. The plaque looks like an elongated fibrillar mat consist- ing mainly of microfilaments joined together by very thin and short cross-linkers.
Intermediate filaments treated with antivimentin IgM directly (Fig. 5A,B) or using immunogold staining
(Fig. 5C,D) seem to be interwoven with the fibrillar network forming the adhesion plaque. Usually an inter- mediate filament bundle or individual intermediate fila- ments associated with an adhesion plaque were oriented at an acute angle to the microfilament bundle associated with this plaque. This picture is consistent with the im- munofluorescence data presented above (Fig. 3). Inter- mediate filaments were often also found in close proximity to the microfilament bundle itself at the region immediately adjacent to the flattened adhesion plaque. In these cases the intermediate filaments sometimes formed dense, fibrillar networks covering the terminal part of the cylindrical body of the microfilament bundle.
Intermediate filaments and their bundles oriented transversely to the direction of elongated adhesion plaques were also observed (Fig. 5C,D). These intermediate fil- aments usually linked the neighboring adhesion plaques with each other.
Intermediate Filaments Retain Their Connection With Adhesion Plaques During the Early Stages of Colcemid Action
It is well known that the incubation of cells with colcemid leads to a depolymerization of cytoplasmic mi-

280 Bershadsky, Tint, and Svitkina
Figure 5

Intermediate Filaments and Focal Contacts 281
Intermediate filaments are associated not only with elongated vinculin-positive streaks (“dash contacts”), but also with certain vinculin-positive spots at the active edge of the cell (“dot contacts”). At the same time actin microfilament bundles are associated with dash contacts but not with dot contacts [Bershadsky et al., 19851. It has been suggested that dot contacts are initial forms of focal contacts and dash contacts are mature contacts [Bershad- sky et al., 1985; see also Izzard and Lochner, 1980; Small and Rinnerthaler, 19851. Thus, we speculate that at least in some cases the association of intermediate filaments with a focal contact precedes the formation of actin microfilament bundles and maturation of the contact.
In fibroblasts intermediate filaments are associated with microtubules [Goldman and Knipe, 1972; Singer et al., 1982; Green and Goldman, 19831. Moreover, some observations are consistent with the view that intermedi- ate filaments follow microtubules in their growth toward the periphery of the spreading cell [Hynes and Destree, 1978; Lyass, 19851. Here we have confirmed the data of Small and Rinnerthaler [1985] that the ends of certain microtubules are located in close proximity to vinculin- positive adhesion plaques.
Thus, an attractive hypothesis of the temporal de- velopment of focal contacts is that microtubules direct the intermediate filament attachment to the initial adhe- sion plaque, and this attachment, in turn, promotes the formation of the actin bundle associated with the plaque. Of course, this assumption needs experimental support.
It should be pointed out that the system of interme- diate filaments and microtubules is not absolutely neces- sary for the maturation of focal contacts, because both the cell spreading and dash contact formation can proceed in colcemid-containing media in which microtubules are depolymerized and most of the intermediate filaments are aggregated in the central part of the cell. However, in these conditions fibroblasts spread very slowly and with low efficiency, and the final cell morphology differs considerably from that of the intact cells [Vasiliev and Gelfand, 19761.
Thus, intermediate filaments together with micro- tubules may play a role in the formation of the system of correctly distributed and correctly oriented mature con- tacts connected with actin microfilament bundles, which is essential for a coordinate course of cell spreading and polarization.
crotubules and then to an aggregation of intermediate filaments in the perinuclear area. At the final stage of this process, the intermediate filaments collapsed at the central part of the cells and had apparently no connec- tions with the system of vinculin-positive focal contacts. However, after a 3-hr incubation with colcemid (2 pgl mi), vimentin-positive fibrils, which coincide with the focal contacts by their distal ends, were frequently seen (Fig. 6A,B). The proximal ends of these fibrils were usually associated with the bulk of intermediate filaments already concentrated near the nucleus. Sometimes short, wavy fragments of vimentin-positive fibrils were seen. These fragments lost their connection with the mass of collapsed intermediate filaments but appeared to remain associated with the focal contacts.
DISCUSSION
Our results provide evidence for an association of vimentin intermediate filaments and adhesion plaques of fibroblasts. The possibility of such an association was briefly mentioned in several earlier communications [Badley et al., 1978; Bershadsky and Tint, 1984; Green and Goldman, 19841. Here we have presented three lines of evidence to support this possibility.
First, the existence of the association between inter- mediate filaments and adhesion plaques can be inferred from a comparison of immunofluorescence pictures of the same quail cell stained with antivimentin and antivin- culin simultaneously. Second, immunoelectron micros- copy of intermediate filament distribution in the cytoskeleton of quail and mouse embryo fibroblasts also revealed an apparent association. Third, the distal ends of some vimentin-positive fibrils retain the association with adhesion plaques even after a colcemid-induced ag- gregation of the bulk of intermediate filaments in the central part of a cell. These data suggest that intermediate filaments and adhesion plaques may somehow be physi- cally connected.
Fig. 5. Platinum replicas of cytoskeletons treated with antivimentin. A: Fragment of the leading edge of a quail fibroblast directly stained with antivimentin IgM. Microfilament bundle (8) with adhesion plaque (P) at the distal end is seen. Bar = 1 pm. B: Detail of A. Numerous intermediate filaments decorated in a bead-like fashion (arrows) are attached to the adhesion plaque. Undecorated microfilaments forming the plaque are also seen. Bar = 0.2 pm. C: General view of the leading edge of a mouse fibroblast. Two adhesion plaques and asso- ciated bundles of microfilaments are seen. Vimentin filaments treated with indirect imniunogold staining are oriented transversely to the bundles and link the neighboring plaques with each other. Bar = 1 pm. C: Detail of C. Intermediate filaments treated with colloidal gold and microfilaments not showing colloidal gold in the adhesion plaque are seen. Bar = 0.2 pm.
ACKNOWLEDGMENTS
We thank Prof. J.M. Vasiliev and Drs. A.V. Lju- bimov and A.A. Neyfakh, Jr., for critical reading of the manuscript.

282 Bershadsky, Tint, and Svitkina
Fig. 6 . Retention of the association of intermediate filaments and focal contacts in the course of colcemid- induced collapse of intermediate filaments. The cell was treated with colcemid for 3 hr. Double- immunofluorescence labeling with antibody to vinculin (A) and to vimentin (B). Note vimentin-positive fibrils associated with the bulk of both intermediate filaments and focal contacts. Bar = 10 Fm.
NOTE ADDED IN PROOF
After the final version of this article had been pre- pared, the data on the association of vimentin filaments with the cell surface and extracellular matrix were pub- lished [Green and Goldman, 19861. Those results are in agreement with the suggestion that intermediate filaments of fibroblasts are involved in the formation of cell-sub- strate contacts.
REFERENCES
Abercrombie, M., and Dunn, G.A. (1975): Adhesion of fibroblasts to substratum during contact inhibition observed by interfer- ence reflection microscopy. Exp. Cell Res. 92:57-62.
Abercrombie, M., Heaysman, J.E.M., and Pegrum, S.M. (1971): The locomotion of fibroblasts in culture. IV. Electron micros- copy of the leading lamella. Exp. Cell Res. 67:359-367.
Badley, R.A., Lloyed, C.W., Woods, A,, Carruthers, L. , Allcock, C., and Rees, D.A. (1978): Mechanisms of cellular adhesion 111. Preparation and preliminary characterisation of adhesions. Exp. Cell Res. 117:231-244.
Bershadsky, A.D., and Tint, I .S. (1984): Association of cytoskeleton and vinculin-containing adhesion plaques: A difference be- tween normal and RSV-transformed quail cells. In 16th Meet- ing of the Federation of European biochemical Societies, Moscow. Abstracts. p. 224 (Abstr. IV-005).
Bershadsky, A.D., Tint, I.S., Neyfakh, A.A., Jr., and Vasiliev, J.M. (1985): Focal contacts in normal and RSV-transformed quail
cells. Hypothesis of the transformation induced deficient mat- uration of focal contacts. Exp. Cell Res. 158:433-444.
Burridge, K., and Connell, L. (1983): Talin: A cytoskeletal compo- nent conccntrated in adhesion plaques and other sites of actin- membrane interaction. Cell Motil. 3:405-417.
Burridge, K., and Feramisco, J.K. (1980): Microinjection and local- ization of a 130 kD protein in living fibroblasts: A relationship to actin and fibronectin. Cell 19:587-595.
Drochmans, P., Freudenstein, C., Wanson, J.-C., Laurent, L. , Keenan, T. W., Stddler, J . , Leloup, R., and Franke, W. W. ( 1978): Structure and biochemical composition of desmosomes and tonofilaments isolated from calf muzzle epidermis. J . Cell Biol. 79:427-442.
Franke, W.W., Moll, R., Schiller, D.L., Schmid, E., Kartenbeck, J . , and Mueller, H. (1982): Desmoplakins of epithelial and myocardial desmosomes are immunologically and biochemi- cally related. Differentiation 23: 189-205.
Geiger, B. (1979): A 130 kD protein from chicken gizzard: Its local- ization at the termini of microfilament bundles in cultured chicken cells. Cell 18: 193-205.
Geiger, B., Schmid, E., and Franke, W.W. (1983): Spatial distribu- tion of proteins specific for desmosomes and adhaerens junc- tions in epithelial cells demonstrated by double immuno- fluorescence microscopy. Differentiation 23: 189-205.
Goldman, R.D., and Knipe, D.M. (1972): Functions of cytoplasmic fibers in nonmuscle cell motility. Cold Spring Harbor Symp. Quant. Biol. 37:523-534.
Green, K.J., and Goldman, R.D. (1983): The effects of taxol on cytoskeleton components in cultured fibroblasts and epithelial cells. Cell Motil. 3:283-305.
Green, K.J., and Goldman, R.D. (1984): Evidence for an interaction

Intermediate Filaments and Focal Contacts 283
Singer, S.J., Ball, E.H., Geiger, B., and Chen, W.-T. (1982): Im- munolabeling studies of cytoskeletal associations in culturcd cells. Cold Spring Harbor Symp. Quant. Biol. 46:301-316.
Skerrow, C.J., and Skerrow, D. (1980): Desmosomes and filaments in mammalian epidermis. In Curtis, A.G., and Pitts, J.D. (eds): “Cell Adhesion and Motility.” Cambridge: Cambridgc University Press, pp. 445-464.
Small, J.V., and Rinnerthaler, G. (1985): Cytostructural dynamics of contact formation during fibroblast locomotion in vitro. Exp. Biol. Med. 1054-68.
Svitkina, T.M., Shevelev, A.A., Bershadsky, A.D., and Gelfand, V.I. (1984): Cytoskeleton of mouse embryo fibroblasts. Elec- tron microscopy of platinum replicas. Eur. J. Cell Bid . 34:64- 74.
Troyanovsky, S.M., Bannikov, G.S., Bershadsky, A.D., Gelfand, V.I., Ievleva, E.S., Karavanova, I.D., Lubimov, A.V., Mech- etner, E.B., Neyfakh, A.A., Jr., Rosinova, E.N., Svitkina, T.M., Tint, I.S., and Etkin, A.F. (1985): Production and characterization of monoclonal antibodies against intermediate filament proteins [in Russian]. Imniunologija 6:70-73.
Vasiliev, J.M., and Gelfand, I.M. (1976): Effects of colcemid on morphogenetic processes and locomotion of fibroblasts. In Goldman, R., Pollard, T., and Rosenbaum, J. (eds.) “Cell Motility.” Cold Spring Harbor, NY: Cold Spring Harbor Lab- oratory, pp. 365-395.
Viklicky, V., Draber, P., Hasek, J., and Bartek, J. (1982): Production and characterization of a monoclonal antitubulin antibody. Cell Biol. Int . Rep. 6:725-731.
Wehland, J., Osborn, M., and Weber, K. (1979): Cell-to-substratum contacts in living cells: A direct correlation between interfer- ence-reflexion and indirect immunofluorescence microscopy using antibodies against actin and a-actinin. J. Cell Sci. 37:257- 213.
between fibroblast intermediate filaments and fibronectin. J. Cell Biol. 99 (4):113a.
Green, K.I., and Goldman, R.D. (1986): Evidence for an interaction between the cell surface and intermediate filaments in cultured fibroblasts. Cell Motil. Cytoskeleton 6:389-405.
Heath, J.P., and Dunn, G.A. (1978): Cell to substratum contacts of chick fibroblasts and their relation to the microfilament sys- tem. J. Cell Sci. 29:97-212.
Heuser, J.E., and Kirschner, M.W. (1980): Filament organization revealed in platinum replicas of freeze-dried cytoskeletons. J. Cell Biol. 86:2 12-234.
Hynes, R.O., and Destree, A.T. (1978): 10 nm Filaments in normal and transformed cells. Cell 13: 151-163.
Izzard, C.S., and Lochner, L.R. (1976): Cell-to-substrate contacts in living fibroblasts: An interference reflexion study with an evaluation of the technique. J. Cell Sci. 21: 129-159.
Izzard, C.S., and Lochner, L.R. (1980): Formation of cell-to-sub- strate contacts during fibroblast motility: an interference-re- flexion study. J. Cell. Sci. 42:81-116.
Kartenbeck, J., Franke, W.W., Moser, J.G., and Stoffels, U. (1983): Specific attachment of desmin filaments to desmosomal plaques in cardiac myocytes. EMBO J. 2:735-742.
Kartenbeck, J . , Schwechheimer, K., Moll, R., and Franke, W.W. (1984): Attachment of vimentin filaments to desmosomal plaques in human meningiornal cell and arachnoidal tissue. J. Cell Biol. 98: 1072-1081.
Lyass, L.A. (1985): The correlated distribution of microtubules and intermediate filaments during spreading of cultured fibroblasts [in Russian]. Tsitologija 27:342-345.
Neyfakh, A.A., Jr., Tint, IS., Svitkina, T.M., Bershadsky, A.D., and Gelfand, V.I. (1983): Visualization of cellular focal con- tacts using a monoclonal antibody to 80 kD serum protein absorbed on the substratum. Exp. Cell Res. 149:387-396.