assessment of probiotic properties in lactic acid bacteria isolated from wine
TRANSCRIPT

Accepted Manuscript
Assessment of probiotic properties in lactic acid bacteria isolated from wine
Almudena García-Ruiz, Dolores González de Llano, Adelaida Esteban-Fernández,Teresa Requena, Begoña Bartolomé, M.Victoria Moreno-Arribas
PII: S0740-0020(14)00152-X
DOI: 10.1016/j.fm.2014.06.015
Reference: YFMIC 2197
To appear in: Food Microbiology
Received Date: 20 February 2014
Revised Date: 6 May 2014
Accepted Date: 16 June 2014
Please cite this article as: García-Ruiz, A., González de Llano, D., Esteban-Fernández, A., Requena, T.,Bartolomé, B., Moreno-Arribas, M.V., Assessment of probiotic properties in lactic acid bacteria isolatedfrom wine, Food Microbiology (2014), doi: 10.1016/j.fm.2014.06.015.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
1
Assessment of probiotic properties in lactic acid bacteria isolated from wine 1
2
Almudena García-Ruiz, Dolores González de Llano*, Adelaida Esteban-Fernández, 3
Teresa Requena, Begoña Bartolomé, M.Victoria Moreno-Arribas 4
5
6
Instituto de Investigación en Ciencias de la Alimentación (CIAL), CSIC-UAM 7
C/Nicolás Cabrera 9, Campus de Cantoblanco, Universidad Autónoma de Madrid, 8
28049 Madrid, Spain 9
10
11
12
* Corresponding author: [email protected] (Dolores González de Llano) 13
14
15

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
2
Abstract 16
Probiotic properties are highly strain-dependent but rarely studied in enological lactic 17
acid bacteria (LAB). In this study, the probiotic features of 11 strains of Lactobacillus 18
spp., Pediococcus spp., and Oenococcus oeni, including saliva and acid resistance, bile 19
tolerance and exopolysaccharides’ production, were investigated. The assays included 20
two probiotic reference strains (L. plantarum CLC 17 and L. fermentum CECT 5716). 21
The Lactobacillus and Pediococcus strains showed high resistance to lysozyme (> 80% 22
resistance to 100 mg/L of lysozyme under conditions simulating the in vivo dilution by 23
saliva) and were capable of surviving at low pH values (pH 1.8) and bile salts, 24
suggesting good adaptation of the wine strains to gastrointestinal conditions. The ability 25
of the strains to adhere to the intestinal mucosa and the inhibition of the adhesion of 26
Escherichia coli to human intestinal cells were also evaluated. Adhesion levels of 27
enological LAB to Caco-2 cells varied from 0.37% to 12.2%, depending on the strain. 28
In particular, P. pentosaceus CIAL-86 showed a high percentage of adhesion to 29
intestinal cells (>12%), even higher than that shown by the probiotic reference strains, 30
and a high anti-adhesion activity against E. coli CIAL-153 (>30%), all of which support 31
this wine LAB strain as a potential probiotic. 32
33

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
3
1. Introduction 34
Probiotic food products are regarded as a significant part of the functional foods market, 35
a market that is truly expanding both in sales volume (60–70% of the total functional 36
food market) and in the variety of products offered (Mohammadi, et al., 2012). 37
According to the FAO/WHO (FAO/WHO, 2008) probiotics are live microorganisms 38
which when administered in adequate amounts confer a health benefit on the host. 39
These beneficial effects are mainly associated with the maintenance of a healthy gut 40
microbiota and an improvement of its resilience, as well as the modulation of lactose 41
intolerance, bowel function and gastrointestinal (GI) comfort, diarrhea prevention and 42
symptom alleviation, reduction of cholesterol levels and hypertension, and regulation of 43
the immune response, amongst others (Ouwehand et al., 2002; Leahy et al., 2005; de 44
Vrese and Schrezenmeir, 2008). 45
46
The most used probiotics belong to the genera of Lactobacillus and Bifidobacterium, 47
but other lactic acid bacteria (LAB) such as the Lactococcus, Streptococcus and 48
Enterococcus genera and certain yeast strains are also used as probiotics (Ouwehand et 49
al., 2002; de Vrese and Schrezenmeir, 2008; Ohland and MacNaughton, 2010). The 50
majority of the commercialized and most studied probiotics have been isolated from 51
dairy products and from the human GI tract. In fact, LAB are already used in many 52
probiotic dairy products. However, recent studies have evaluated the probiotic potential, 53
as a means of resistance to the extreme conditions of the GI tract (low pH in the 54
stomach, digestive enzymes, bile salts), adhesion to the intestinal mucosa, prolonged 55
and stable persistence in the intestinal tract, and antimicrobial and immunomodulatory 56

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
4
properties, amongst others, of bacteria of vegetable origin belonging to the genera of 57
Pediococcus and Leuconostoc (Jonganurakkun et al., 2008; Kang et al., 2009), as well 58
as bacterial strains isolated from alcoholic fermented beverages such as cider, in 59
particular Pediococcus parvulus (Fernández de Palencia et al., 2009). 60
61
LABs associated with the winemaking process mainly belong to the genera of 62
Oenococcus, Pediococcus and Lactobacillus, being Oenococcus oeni the main species 63
responsible of wine malolactic fermentation (MLF) (Fugelsang, 1997; Wibowo et al., 64
1985). These bacteria are adapted to grow in the hostile conditions imposed during the 65
elaboration of wine: low pH, high ethanol concentration, poor proportion of nutrients, 66
etc (Lonvaud-Funel et al., 2001; Mills et al., 2005; Spano and Massa, 2006). Resistance 67
to these factors, along with their structural and functional similarity with other bacterial 68
groups belonging to the most conventional probiotics, convert enological LAB into 69
potential probiotic candidates to exert beneficial effects on human health. To our 70
knowledge, only the study carried out by Foligné et al. (2010) has investigated the 71
probiotic properties of wine-related LAB. These authors explored the in vitro 72
immunomodulatory activities of strains belonging to O. oeni and Pediococcus parvulus, 73
found in wine, concluding that some O. oeni strains showed a measurable 74
immunomodulatory potential, although not at the level of certain conventional 75
probiotics. 76
77
The aim of this paper was to assess the probiotic potential of LAB from an enological 78
bacteria collection including strains from different genera and species. For that, selected 79

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
5
LAB strains were subjected to a series of in vitro analyses to evaluate: a) their resistance 80
to conditions in the GI tract; b) their adhesion to intestinal cells; and c) their effects on 81
the adhesion of pathogen bacteria to cultured human intestinal cells. These evaluations 82
were performed as an initial step toward establishing rational criteria for screening and 83
selecting wine-borne microorganisms with potential human probiotic properties. 84
85
2. Materials and Methods 86
2.1. Bacterial strains and culture media 87
Eleven strains of LAB belonging to Pediococcus pentosaceus (n=4), Lactobacillus casei 88
(n=3), Lactobacillus plantarum (n=1) and O. oeni (n=3) (Table 1) were selected from 89
the bacterial culture collection of CIAL (Instituto de Investigación en Ciencias de la 90
Alimentación, CSIC-UAM). These strains were previously isolated from red wines 91
during the early phase of MLF, and properly identified by 16S rRNA partial gene 92
sequencing as described by Moreno-Arribas and Polo (2008) and García-Ruiz et al. 93
(2013). In addition, two previously characterized probiotic strains (L. plantarum CLC 94
17 and L. fermentum CECT 5716) were used as reference controls. Besides in vitro 95
studies about its probiotic characteristics (Martín et al., 2005), L. fermentum CECT 96
5716 has been subjected to in vivo trials that have proven its tolerance, safety and 97
potential protection against gastrointestinal infections in infants (Gil-Campos et al., 98
2012). L. plantarum CLC 17 (also named L. plantarum LCH17) has been used in the 99
study of antimicrobial properties of phenolic acids and fungi extracts, among other 100
probiotic strains (Cueva et al., 2010; 2011). All strains were kept frozen at -70ºC in a 101
sterilized mixture of culture medium and glycerol (80:20, v⁄v). Lactobacillus and 102

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
6
Pediococcus strains were grown at 30ºC in MRS broth (Pronadisa, Madrid, Spain) and 103
MRS-Agar (Pronadisa) while Oenococcus strains were grown at 30ºC in MRS broth 104
supplemented with 5 g/L fructose (Panreac, Barcelona, Spain) and 1 g/L malic acid 105
(Panreac) (final pH 4.8) and MRS-Agar supplemented with 0.5 g/L cysteine (Sigma-106
Aldrich, St. Louis, MO, USA). 107
The strain Escherichia coli CIAL-153 (isolated from human feces) was used in the 108
assays of bacterial adhesion to intestinal cells. E. coli was grown at 37ºC in TSB 109
(Scharlau, Barcelona, Spain) and TSA (Scharlau) broths. 110
111
2.2. Assays of resistance to the gastrointestinal tract 112
2.2.1. Resistance to lysozyme 113
The lysozyme resistance assays were performed using the method described by Zago et 114
al. (2011). Enological strains grown overnight in 10 ml MRS broth at 30ºC were 115
pelleted by centrifugation, washed twice with phosphate buffer (0.1 M, pH 7.0), and 116
resuspended in 2 ml of Ringer solution (8.5 g/L NaCl, 0.4 g/L KCl, 0.34 g/L hydrated 117
CaCl2, Sigma-Aldrich). To simulate the in vivo dilution by saliva, the bacterial 118
suspensions (108-109 colonies forming units, CFU/mL (OD600=1)) were inoculated in a 119
sterile electrolyte solution (SES) (0.22 g/L CaCl2, 6.2 g/L NaCl, 2.2 g/L KCl, 1.2 g/L 120
NaHCO3) in the presence of 100 mg/L of lysozyme (Sigma-Aldrich). Bacterial 121
suspensions in SES without lysozyme were included as controls. Survival rate was 122
calculated as the percentage of the CFU/mL after 30 and 120 min compared to the 123

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
7
CFU/mL at time 0. CFU/mL was determined by cell counts in the appropriate agar 124
media were performed. Assays were carried out in triplicate. 125
126
2.2.2. Tolerance to simulated gastric juice 127
Overnight cultures of the strains were harvested by centrifugation (3000 g, 10 min, 5ºC) 128
and washed twice with phosphate buffer (pH 7). The cell suspension (108-109 CFU/mL) 129
was mixed (1:1) in 2x SES with 0.6% (w/v) pepsin. Samples were incubated with 130
agitation at 37ºC. Gastric environment was reproduced by progressive acidification 131
(addition of 1M HCl) from the initial pH value of 5.0 to 4.1, 3.0, 2.1 and 1.8. The 132
suspension was sequentially incubated for 20 min at each pH value; with the exception 133
of pH 1.8 which was incubated for 30 min. At 0, 20, 40, 60 and 90 min of incubation, 134
cell counts in the appropriate agar media were performed. Assays were carried out in 135
triplicate. 136
137
2.2.3. Bile resistance 138
The ability of the strains to grow in the presence of bile (w/v) was determined according 139
to the method of Vinderola and Reinheimer (2003). Each strain grown overnight was 140
inoculated (2% v/v) into appropriate broth with 0.06%, 0.125%, 0.25%, 0.5% and 1% of 141
bile (w/v) (Sigma-Aldrich). Cultures were incubated at 37ºC and, after 24 h, optical 142
density at 600 nm (OD600) was measured and compared to a control culture (without 143
bile salts). The results were expressed as the percentage of growth compared to the 144
control. Assays were carried out in triplicate. 145

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
8
146
2.3. Cell culture assays 147
Caco-2 cells from human colon adenocarcinoma were obtained from the American 148
Type Culture Collection and used in their terminally differentiated state to mimic small 149
intestine mature enterocytes. Caco-2 cells were grown and maintained in Dulbecco’s 150
modified Eagle’s medium (DMEM, Sigma-Aldrich) supplemented with 10% (v/v) fetal 151
calf serum at 37ºC in an atmosphere of 5% CO2/95% air at constant humidity. For the 152
experiments, Caco-2 cells were seeded in 24-well tissue plates at 25,000 cells/cm2 153
density and grown over 15 days to obtain a monolayer of differentiated and polarized 154
cells, and the culture medium was changed every 2 days. 155
156
2.3.1. LAB adhesion 157
The method described by Fernández de Palencia et al. (2008) was followed to study the 158
adhesion of the LAB strains to Caco-2 cells. Overnight cultures of the LAB strains 159
being studied were harvested by centrifugation (10,000 g, 10 min, 4ºC) and resuspended 160
in DPBS solution at a concentration of about 108 CFU/mL (OD600= 1). Then, 0.5 mL of 161
bacterial suspension was added to Caco-2 cell monolayers previously washed with 162
Dulbecco’s phosphate-buffered saline (DPBS Lonza Walkersville, Inc., USA).The ratio 163
of Caco-2 cells to bacteria was ≥ 1:100. 164
After 1 h of incubation at 37ºC under 5% CO2 atmosphere, wells were softly washed 165
three times with PBS solution to remove unbound bacteria. Caco-2 cells and adhered 166
bacteria were then detached using 0.05% trypsin-EDTA solution and the bacterial 167

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
9
counts were carried out in the appropriate agar media as described above. The adhesion 168
capacity was expressed as the number of adhered bacteria (CFU/mL) relative to the total 169
number of bacteria added initially (% Adhesion = [Adhered bacteria / Total of added 170
bacteria] x 100). CFU/mL was determined by cell counts in the appropriate agar media 171
were performed. Assays were performed in triplicate and three independent experiments 172
were carried out. 173
174
2.3.2. Competition between LAB and E. coli for cell adhesion 175
Competitiveness was tested by adding LAB strains and E. coli CIAL-153 176
simultaneously (in an initial ration of 1:1) to the Caco-2 cells followed by incubation for 177
1 h. Non-bound pathogens and bacteria were removed and the bacterial counts were 178
carried out as described above. Competitiveness was calculated as the percentage of 179
adhesion of E. coli added in combination with LAB strains relative to pathogen-bound 180
bacteria in the absence of LAB (control). 181
182
2.3.3. Inhibition of E. coli adhesion 183
To test the ability of the LAB strains to inhibit the adhesion of E. coli CIAL-153, LAB 184
strains were first added to the monolayer of Caco-2 cells and incubated for 1 h. Non-185
bound bacteria were removed by washing and E. coli was added to the wells and the 186
mixture was incubated for 1 h. Caco-2 cells and adhered bacteria (LAB/E. coli) were 187
then detached and the bacterial counts were carried out. The inhibition of the adhesion 188
of E. coli was expressed as a percentage using the following formula: Inhibition of 189

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
10
adhesion = 100×(1 − T1/T2), where T1 and T2 are the percentage of adhesion by E. coli 190
cells in the presence and absence of LAB strains, respectively. 191
192
2.3.4. Displacement of adhered E. coli 193
The ability of the LAB strains to displace previously adhered E. coli was assessed as 194
follows. E. coli CIAL-153 was first added to Caco-2 cells and incubated for 1 h. Non-195
bound E. coli bacteria were removed by washing and LAB strains were added to the 196
cells and the mixture incubated for 1 h. Caco-2 cells and adhered E. coli/LAB were then 197
detached and the bacterial counts were performed. Displacement of pathogens was 198
expressed as the percentage of adhesion by E. coli cells in the presence and absence of 199
LAB strains, as described above. 200
201
2.4. Production of exopolysaccharide 202
The screening for exopolysaccharide (EPS)-producing LAB strains followed the method 203
described by Garai-Ibabe et al. (2010), slightly modified. The LABs were grown in 204
MRS broth (pH 5.5) at 30ºC in an atmosphere containing 5% CO2 for 48h. The EPS-205
producing ability was evaluated by visual observation of the culture viscosity. 206
207
3. Results and Discussion 208
3.1. Resistance of wine LAB strains to gastrointestinal tract conditions 209
210

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
11
An important step towards the selection of potential probiotic candidates is to evaluate 211
their resistance to the extreme conditions of the GI tract. The first barrier that must be 212
overcome is the mouth, with a high concentration of lysozyme in the human saliva; then 213
the stomach, with low pH and digestive enzymes (i.e. pepsin); and the upper intestine, 214
which contains bile (Corzo and Gilliland, 1999). 215
216
Table 1 reports data of bacteria survival after treatment with lysozyme for 30 and 120 217
minutes. LAB strains of P. pentosaceus and L. casei -in particular CIAL-49, CIAL-86 218
and CIAL-92- and reference probiotic strain L. plantarum CLC 17 showed high 219
resistance to lysozyme, with survival percentages > 80 % even after 120 min of 220
incubation, which can be considered a severe treatment. L. plantarum CIAL-121 221
showed medium resistance to lysozyme, with a survival percentage ≥ 50 % after 120 222
min, and similar to that exhibited by the reference probiotic strain L. fermentum 223
CECT5716. In contrast, O. oeni strains were particularly sensitive to the action of 224
lysozyme, being mostly inactivated after 120 min (% of survival < 1 %). Resistance to 225
lysozyme has been attributed to the peptidoglycan structure in the cell wall, at the 226
physiological state of the cell and lysozyme structure in the medium (Cunningham et 227
al., 1991). This result confirms the high resistance of Lactobacillus strains to 100 mg/L 228
of lysozyme under conditions simulating the in vivo dilution by saliva observed by other 229
authors (Zago et al., 2011; Turchi et al., 2013). 230
231
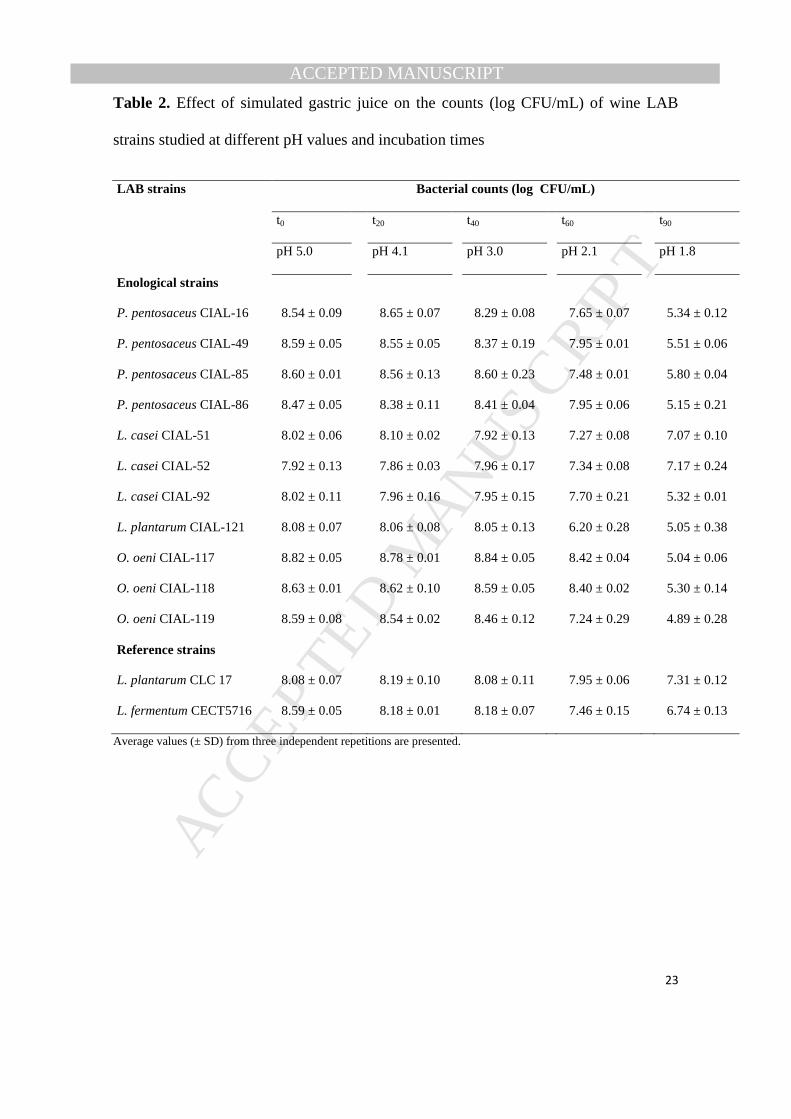
The wine LAB strains studied also showed great resistance to gastric juice conditions 232
(Table 2). There were no differences in any of their cell counts within the first 60 min of 233
incubation when pH decreased from 5.0 to 3.0. This was expected for the LAB isolated 234

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
12
from wine, since they are well adapted to wine conditions with a pH of about 3.5. 235
However, at the end of the treatment, when the simulated gastric juice reached pH 1.8, 236
the reduction of viability of the enological LAB strains was approximately 3 log-units, 237
except for L. casei CIAL-51 and L. casei CIAL-52, which exhibited a reduction of only 238
1 log-unit. It is noteworthy that the gastric juice resistance of these strains was similar to 239
the reference probiotic strain L. plantarum CLC 17 and superior to L. fermentum 240
CECT5716 (Table 2). These results suggest a good tolerance of the strains at pH 1.8, 241
which simulated the last gastric emptying of the intestine, as a strong discriminative pH 242
for the selection of high-acid tolerant strains. Other strains from Lactobacillus and 243
Pediococcus genera have also shown good tolerance to gastric juice conditions 244
(Fernández de Palencia et al., 2008; Bove et al., 2012; Jensen et al., 2012; Turchi et al., 245
2013). 246
247
The relevant physiological concentrations of human bile ranges from 0.3% to 0.5% 248
(Zavaglia et al., 1998; Dunne et al., 1999). It has also been reported that good bile 249
tolerance benefits the colonization in the host GI tract (Luo et al., 2012). In this regard, 250
it is important to evaluate the ability of potential probiotics to survive in the presence of 251
bile. For the enological LAB strains studied, the growth percentages at the maximum 252
concentration of bile assayed (1%) were higher than 70%, except for L. casei CIAL-51 253
and L. casei CIAL-52 (Table 1). Of special interest was the bile resistance of O. oeni 254
CIAL-117 (90.7%), which was even greater than that exhibited by the reference 255
probiotic strains, L. plantarum CLC 17 (73%) and L. fermentum CECT5716 (72%). 256
Mathara et al. (2008) established a limit of 0.3% bile to strain selection, considering 257
good resistance to be when the growth percentage in the presence of bile was above 258

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
13
50%. As mentioned, all strains assayed showed a percentage of growth above 50% in 259
the presence of bile, which reflected good bile resistance. This good bile tolerance is in 260
accordance with the results reported by Delgado et al. (2008), Turchi et al. (2013), 261
Jensen et al. (2012), and Chen et al. (2010) for Bifidobacterium, Lactobacillus strains, 262
Pediococcus pentosaceus and certain yeasts. 263
264
3.2. LAB adhesion to intestinal cells 265
266
Another important selection criterion for potential probiotic microorganisms is their 267
ability to adhere to the intestinal mucosa. This ability may provide beneficial effects, 268
such as the exclusion of pathogens (Collins et al., 1998; Lee et al., 2002; Ouwehand et 269
al., 2002) or host immunomodulation (Schiffrin et al., 1995). The difficulties of 270
studying bacterial adhesion in vivo have led to the development of in vitro model 271
systems for the preliminary studies of adhered strains (Verstelund et al., 2005). 272
Specifically, the human intestinal Caco-2 cell line is widely used in assays to evaluate 273
the adhesion properties of potential probiotic strains because this cellular model 274
expresses morphological and functional differentiation in vitro and shows 275
characteristics of mature enterocytes (Sambuy et al., 2005; Fernández de Palencia et al., 276
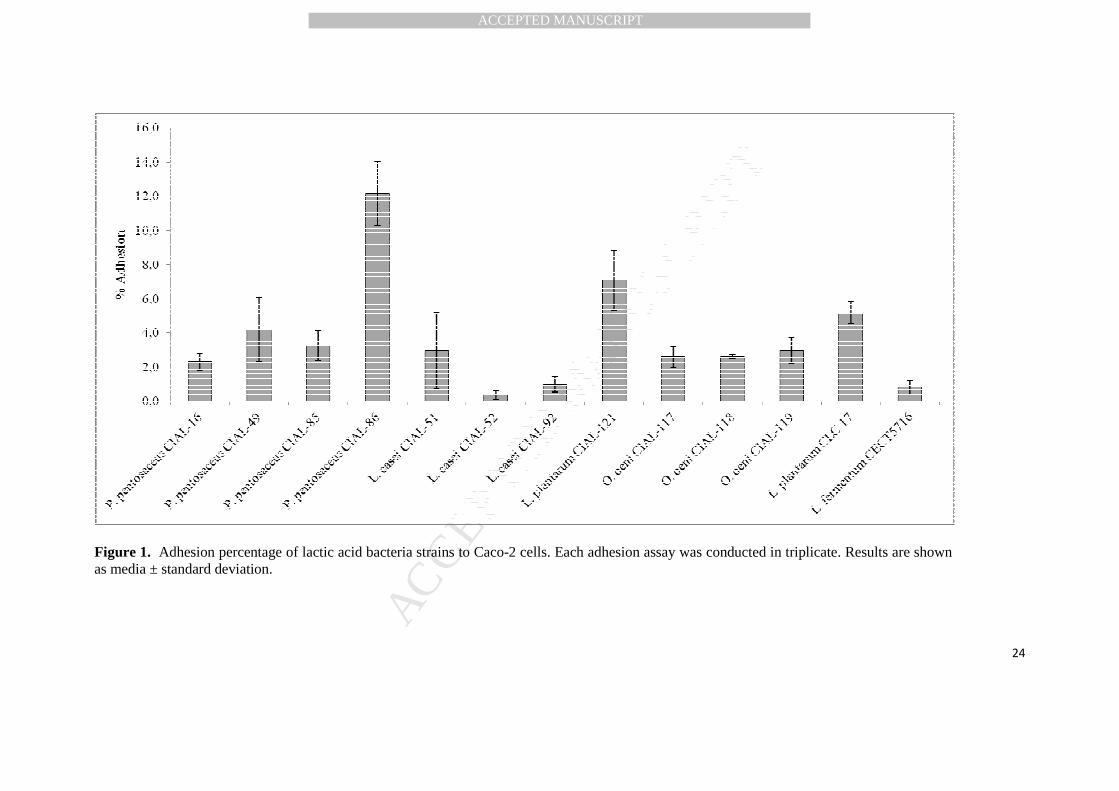
2008). Adhesion levels of enological LAB to Caco-2 cells varied from 0.37% to 12.2%, 277
depending on the strain, species and genera (Figure 1). This dependence is in line with 278
previously published data by Collado et al. (2006), whose adhesion values ranged from 279
0.9% (P. freudenreichii JS) to 20% (L. rhamnosus GG). P. pentosaceus CIAL-86 280
presented the highest adhesion percentage (12.2%), followed by L. plantarum CIAL-281
121 (7.10 %), both with adhesion values superior to the reference probiotic strain 282

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
14
(Figure 1). The adhesion level of P. pentosaceus CIAL-86 was also higher or similar to 283
that reported for the probiotic L. rhamnosus GG (9.7%) (Tuomola and Salminen, 1998), 284
P. pentosaceus BH105 (10.12%) (Uymaz et al., 2009) and VJ49 (12%) (Vidhyasagar 285
and Jeevaratnam, 2013), but lower than that measured for P. pentosaceus OZF (14.4%) 286
(Osmanagaoglu et al., 2010) and VJ13 (16%) (Vidhyasagar and Jeevaratnam, 2013), 287
The adhesion ability is affected by many factors, among which is the production of 288
EPS. In this work, the production of EPS was only assayed for the strains with a higher 289
percentage of adhesion (P. pentosaceus CIAL-86, L. plantarum CIAL-121) and for the 290
reference probiotic strain L. plantarum CLC 17. The so-called “ropy” character was 291
visually detected for all of them (results not shown). Therefore, the structure of EPS 292
may promote strain-specific interactions of bacteria with specific receptors and effectors 293
of Caco-2 cells. 294
295
3.3. Effects of LAB on E. coli adhesion to intestinal cells 296
297
Finally, and with the aim of evaluating the ability of enological LAB strains to prevent 298
the adhesion of pathogens to the intestinal mucosa, anti-adhesion assays were carried 299
out. In these assays, the ability of P. pentosaceus CIAL-86, L. plantarum CIAL-121 and 300
the probiotic control L. plantarum CLC 17, to compete, inhibit and displace the 301
attachment of E. coli CIAL-153 to Caco-2 cell lines was evaluated. Results of anti-302
adhesion assays are presented in the Figure 2. E. coli CIAL-153 showed an adhesion of 303
6.83%. When E. coli and LAB were added simultaneously (competition assay), the 304
degree of adhesion of E. coli was reduced by around 31–52% (Figure 2); this reduction 305
is in line with that reported by Lee et al. (2003) regarding the attachment inhibition of 306

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
15
E. coli strains to Caco-2 cells by L. rhamnosus. In a previous work, Lee and Puong 307
(2002) proposed that the competitiveness of two bacteria on the surface of host cells 308
depends on adhesive molecule-receptor interaction or their relative positions in the case 309
of steric hindrance. The ability to inhibit the adhesion of pathogens appears to depend 310
on both the probiotic and pathogen, indicating a very high specificity (Collado et al., 311
2006; Gueimonde et al., 2006). In our inhibition assays, P. pentosaceus CIAL-86 was 312
the most effective strain inhibiting the adhesion of E. coli CIAL-153, while L. 313
plantarum CIAL-121 showed the lowest values of inhibition with 8.89% (Figure 2). 314
The high values of exclusion observed by P. pentosaceus CIAL-86 and L. plantarum 315
CLC 17 (>30%) could indicate the competition of these strains with E. coli CIAL-153 316
for common adhesion receptors (Bernet et al., 1993; Lee and Puong, 2002; Leahy et al., 317
2005). The displacement of pre-adhered pathogens was also variable depending on the 318
tested strains and species (Figure 2). This displacement phenomenon could be explained 319
by the production of antimicrobial compounds or anti-adhesion factors, and also by the 320
competing for the same adhesion receptors (Lievin et al., 2000; Abedi et al., 2013). The 321
results of the inhibition and displacement assays were also different from each other. 322
This, together with previous observations (Lee et al., 2003; Collado et al., 2005; 2006; 323
Gueimonde et al., 2006), appears to confirm that different mechanisms are implied in 324
those processes. On the other hand, the absence of any correlation between the adhesion 325
and anti-adhesion results could suggest that the mechanisms implied in both phenomena 326
were different. This is in accordance with the results reported by other authors (Biblioni 327
et al., 1995; Collado et al., 2005; 2006). 328
In summary, the adhesion and anti-adhesion assays reflect very high strain specificity, 329
highlighting P. pentosaceus CIAL-86 has an excellent adhesion level and a good anti-330

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
16
adhesion activity against E. coli CIAL-153. In addition, the results obtained in the 331
present work show the convenient ability of 11 LAB strains isolated from wines to 332
resist the GI hostile environment; with values of resistance to lysozyme, gastric juice 333
and bile often similar or higher to those observed in the control probiotic strains L. 334
plantarum CLC 17 and L. fermentum CECT5716. As a whole, the results obtained 335
suggest that enological LAB strains, and particularly P. pentosaceus CIAL-86, display 336
promising probiotic properties, while further in vitro and in vivo investigations are still 337
necessary in order to confirm its beneficial role to human health. 338
339
Acknowledgments 340
This work has been funded by MINECO (Projects AGL2012-04172-C02-01, PRI-341
PIBAR-2011-1358 and Consolider Ingenio 2010 FUN-C-FOOD CSD2007-00063), the 342
Comunidad Autónoma de Madrid (ALIBIRD P2009/AGR-1469) and the INIA 343
RM2011-00003-00-00 Project. AGR is the recipient of a fellowship by the Danone 344
Institute. We also thank Dr. J.M. Rodríguez from the University Complutense of 345
Madrid (Spain) for providing us with the probiotic reference strains. 346
347
4. References 348
Abedi, D., Feizizadeh, S., Akbari, V., Jafarian-Dehkordi, A., 2013. In vitro anti-349
bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on 350
Escherichia coli. Research in Pharmaceutical Sciences, 8, 260-268. 351
352
Bernet, M. F., Brassart, D., Neeser, J.R., Servin, A.L., 1994. Lactobacillus acidophilus 353
la-1 binds to cultured human intestinal-cell lines and inhibits cell attachment and cell 354
invasion by enterovirulent bacteria. Gut, 35, 483-489. 355
356
Bibiloni, R., Fernando, P., De Antoni, G.L., 1999. Will a high adhering capacity in a 357
probiotic strain guarantee exclusion of pathogens from intestinal epithelia. Anaerobe, 5, 358
519-524. 359

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
17
360
Bove, P., Gallone, A., Russo, P., Capozzi, V., Albenzio, M., Spano, G., Fiocco, D., 361
2012. Probiotic features of Lactobacillus plantarum mutant strains. Applied 362
Microbiology and Biotechnology, 96, 431-441. 363
364
Chen, L. S., Ma, Y.; Maubois, J.L., Chen, L.J., Liu, Q.H., Guo, J.P., 2010. Identification 365
of yeasts from raw milk and selection for some specific antioxidant properties. 366
International Journal of Dairy Technology, 63, 47-54. 367
368
Collado, M.C., Gueimonde, M., Hernandez, M., Sanz, Y., Salminen, S., 2005. Adhesion 369
of selected Bifidobacterium strains to human intestinal mucus and the role of adhesion 370
in enteropathogen exclusion. Journal of Food Protection, 68, 2672-2678. 371
372
Collado, M.C., Jalonen, L., Meriluoto, J., Salminen, S., 2006. Protection mechanism of 373
probiotic combination against human pathogens: in vitro adhesion to human intestinal 374
mucus. Asia Pacific Journal of Clinical Nutrition, 15, 570-575. 375
376
Collins, J. K., Thornton, G., Sullivan, G.O., 1998. Selection of probiotic strains for 377
human applications. International Dairy Journal, 8, 487-490. 378
379
Corzo, G., Gilliland, S.E., 1999. Bile salt hydrolase activity of three strains of 380
Lactobacillus acidophilus. Journal of Dairy Science, 82, 472-480. 381
382
Cueva, C., Moreno-Arribas, M.V., Martín-Álvarez P.J., Bills, G., Vicente, M.F., 383
Basilio, A., López Rivas, C., Requena, T., Rodríguez J.M., Bartolome, B., 2010. 384
Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic 385
bacteria. Research in Microbiology, 161, 372-382. 386
387
Cueva, C., Moreno-Arribas, M.V., Bartolome, B., Salazar, O., Vicente, M.F., Bills, G., 388
2011. Antibiosis of vineyard ecosystem fungi against food-borne microorganisms. 389
Research in Microbiology, 162, 1043-1051. 390
391
Cunningham, F.E.; Proctor, V.A.; Goetsch, S.J., 1991. Egg-white lysozyme as a food 392
preservative - An overview. World's Poultry Science Journal, 47, 141-163. 393
394
de Vrese, M., Schrezenmeir, J., 2008. Probiotics, prebiotics, and synbiotics. In Stahl, U., 395
Donalies, U.E.B., Nevoigt, E., (Eds), Food Biotechnology. Advances in Biochemical 396
Engineering/Biotechnology Volume 111 ( pp 1-66). Springer-Verlag Berlin Heidelberg. 397
398
Delgado, S., O'Sullivan, E., Fitzgerald, G., Mayo, B., 2008. In vitro evaluation of the 399
probiotic properties of human intestinal Bifidobacterium species and selection of new 400
probiotic candidates. Journal of Applied Microbiology, 104, 1119-1127. 401
402
Dunne, C., Murphy, L., Flynn, S., O'Mahony, L., O'Halloran, S., Feeney, M., 403
Morrissey, D., Thornton, G., Fitzgerald, G., Daly, C., Kiely, B., Quigley, E.M., 404
O'Sullivan, G.C., Shanahan, F., Collins, J.K., 1999. Probiotics: from myth to reality. 405

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
18
Demonstration of functionality in animal models of disease and in human clinical trials. 406
Antonie van Leeuwenhoek, 76, 279-292. 407
408
FAO/WHO, 2006. Probiotics in food: health and nutritional properties and guidelines 409
for evaluation. FAO Food and Nutrition Paper 85. World Health Organization and Food 410
and Agriculture Organization of the United Nations Rome. 411
412
Fernandez de Palencia, P., Lopez, P., Corbi, A. L., Pelaez, C., Requena, T., 2008. 413
Probiotic strains: survival under simulated gastrointestinal conditions, in vitro adhesion 414
to Caco-2 cells and effect on cytokine secretion. European Food Research and 415
Technology, 227, 1475-1484. 416
417
Fernández de Palencia, P., Werning, M.L., Sierra-Filardi, E., Duenas, M.T., Irastorza, 418
A., Corbi, A.L., Lopez, P., 2009. Probiotic properties of the 2-substituted (1,3)-β-D-419
glucan-producing bacterium Pediococcus parvulus 2.6. Applied and Environmental 420
Microbiology, 75, 4887-4891. 421
422
Foligne, B., Dewulf, J.; Breton, J.; Claisse, O.; Lonvaud-Funel, A.; Pot, B., 2010. 423
Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by 424
Oenococcus oeni. International Journal of Food Microbiology, 140, 136-145. 425
426
Fugelsang, K.C., 1997. Wine Microbiology (pp1-247) Chapman & Hall, New York, 427
USA. 428
429
Garai-Ibabe, G., Areizaga, J., Aznar, R., Elizaquivel, P., Prieto, A., Irastorza, A., 430
Dueñas, M.T., 2010. Screening and selection of 2-branched (1,3)-β-d-glucan producing 431
lactic acid bacteria and exopolysaccharide characterization. Journal of Agricultural and 432
Food Chemistry,58, 6149-6156. 433
434
García-Ruiz, A., Tabasco, R., Requena, T., Claisse, O., Lonvaud-Funel, A., Bartolome, 435
B., Moreno-Arribas, M. V., 2013. Genetic diversity of Oenoccoccus oeni isolated from 436
wines treated with phenolic extracts as antimicrobial agents. Food Microbiology, 36, 437
267-274. 438
439
Gil-Campos, M., López, M.A., Rodriguez-Benítez, M.V., Romero, J., Roncero, I., 440
Linares, M.D., Maldonado, J., López-Huertas, E., Berwind, R., Ritzenthalerd, K.L., 441
Navas, V., Sierra C., Sempere, L., Geerlings, A., Maldonado-Lobón, J.A., Valero, A.D., 442
Lara-Villoslada, F., Olivares M., 2012. Lactobacillus fermentum CECT 5716 is safe and 443
well tolerated in infants of 1–6 months of age: A randomized controlled trial. 444
Pharmacological Research, 65, 231-238. 445
446
Gueimonde, M., Jalonen, L., He, F., Hiramatsu, M., Salminen, S., 2006. Adhesion and 447
competitive inhibition and displacement of human enteropathogens by selected 448
lactobacilli. Food Research International, 39, 467-471. 449
450

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
19
Jensen, H., Grimmer, S., Naterstad, K., Axelsson, L., 2012. In vitro testing of 451
commercial and potential probiotic lactic acid bacteria. International Journal of Food 452
Microbiology, 153, 216-222. 453
454
Jonganurakkun, B., Wang, Q., Xu, S.H., Tada, Y., Minamida, K., Yasokawa, D., Sugi, 455
M., Hara, H., Asano, K., 2008. Pediococcus pentosaceus NB-17 for probiotic use. 456
Journal of Bioscience and Bioengineering, 106, 69-73. 457
458
Kang, H., Oh, Y.J., Ahn, K.S., Eom, H.J., Han, N., Kim, Y.B., Sohn, N.W., 2009. 459
Leuconostoc citreum HJ-P4 (KACC 91035) regulates immunoglobulin E in an 460
ovalbumin-induced allergy model and induces interleukin-12 through nuclear factor-461
kappa B and p38/c-Jun N-terminal kinases signaling in macrophages. Microbiology and 462
Immunology, 53, 331-339. 463
464
Leahy, S.C., Higgins, D.G., Fitzgerald, G.F.; van Sinderen, D., 2005.Getting better with 465
bifidobacteria. Journal of Applied Microbiology, 98, 1303-1315. 466
467
Lee, Y.K.; Puong, K.Y., 2002. Competition for adhesion between probiotics and human 468
gastrointestinal pathogens in the presence of carbohydrate. British Journal of Nutrition, 469
88, S101-S108. 470
471
Lee, Y. K.; Puong, K.Y., Ouwehand, A.C., Salminen, S., 2003. Displacement of 472
bacterial pathogens from mucus and Caco-2 cell surface by lactobacilli. Journal of 473
Medical Microbiology, 52, 925-930. 474
475
Lievin, V., Peiffer, I., Hudault, S., Rochat, F., Brassart, D., Neeser, J. R., Servin, A. L., 476
2000. Bifidobacterium strains from resident infant human gastrointestinal microflora 477
exert antimicrobial activity. Gut, 47, 646-652. 478
479
Lonvaud-Funel, A., 2001. Biogenic amines in wines: Role of lactic acid bacteria. FEMS 480
Microbiology Letters, 199, 9-13. 481
482
Luo, Y., Ma, B.C., Zou, L.K., Cheng, J.G., Cai, Y.H., Kang, J.P., Li, B., Gao, X.H., 483
Wang, P., Xiao, J.J., 2012. Identification and characterization of lactic acid bacteria 484
from forest musk deer feces. African Journal of Microbiology Research, 6, 5871-5881. 485
486
Martin, R., Olivares, M., Marin, M.L.; Fernandez, L., Xaus, J., Rodriguez, J.M., 2005. 487
Probiotic potential of 3 lactobacilli strains isolated from breast milk. Journal of Human 488
Lactation, 21, 8-21. 489
490
Mathara, J.M., Schillinger, U., Kutima, P.M., Mbugua, S.K., Guigas, C., Franz, C., 491
Holzapfel, W.H., 2008. Functional properties of Lactobacillus plantarum strains 492
isolated from Maasai traditional fermented milk products in Kenya. Current 493
Microbiology, 56, 315-321. 494
495

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
20
Mills, D.A., Rawsthorne, H., Parker, C., Tamir, D., Makarova, K., 2005. Genomic 496
analysis of Oenococcus oeni PSU-1 and its relevance to winemaking. FEMS 497
Microbiology Reviews, 29, 465-475. 498
499
Mohammadi, R., Sohrabvandi, S., Mortazavian, A.M., 2012. The starter culture 500
characteristics of probiotic microorganisms in fermented milks. Engineering in Life 501
Sciences, 12, 399-409. 502
503
Moreno-Arribas, M. V., Polo, M.C., 2008. Occurrence of lactic acid bacteria and 504
biogenic amines in biologically aged wines. Food Microbiology, 25, 875-881. 505
506
Ohland, C.L., MacNaughton, W.K., 2010. Probiotic bacteria and intestinal epithelial 507
barrier function. American Journal of Physiology: Gastrointestinal and Liver 508
Physiology, 298, G807-G819. 509
510
Osmanagaoglu, O., Kiran, F., Ataoglu, H., 2010. Evaluation of in vitro probiotic 511
potential of Pediococcus pentosaceus OZF isolated from human breast milk. Probiotics 512
and Antimicrobial Proteins, 2, 162-174. 513
514
Ouwehand, A.C., Salminen, S., Isolauri, E., 2002. Probiotics: an overview of beneficial 515
effects. Antonie van Leeuwenhoek International Journal of General and Molecular 516
Microbiology, 82, 279-289. 517
518
Sambuy, Y., Angelis, I., Ranaldi, G., Scarino, M.L., Stammati, A., Zucco, F., 2005. The 519
Caco-2 cell line as a model of the intestinal barrier: influence of cell and culture-related 520
factors on Caco-2 cell functional characteristics. Cell Biology and Toxicology, 21, 1-26. 521
522
Schiffrin, E.J., Rochat, F., Linkamster, H., Aeschlimann, J.M., Donnethughes, A., 1995. 523
Immunomodulation of human blood-cells following the ingestion of lactic-acid bacteria. 524
Journal of Dairy Science, 78, 491-497. 525
526
Spano, G., Massa, S., 2006. Environmental stress response in wine lactic acid bacteria: 527
Beyond Bacillus subtilis. Critical Reviews in Microbiology, 32, 77-86. 528
529
Tuomola, E.M., Salminen, S.J. Adhesion of some probiotic and dairy Lactobacillus 530
strains to Caco-2 cell cultures. International Journal of Food Microbiology, 41, 45-51. 531
532
Turchi, B., Mancini, S., Fratini, F., Pedonese, F., Nuvoloni, R., Bertelloni, F., Ebani, 533
V.V., Cerri, D., 2013. Preliminary evaluation of probiotic potential of Lactobacillus 534
plantarum strains isolated from Italian food products. World Journal of Microbiology 535
and Biotechnology, 29, 1913-1922. 536
537
Uymaz, B., Simsek, O., Akkoc, N., Ataoglu, H., Akcelik, M., 2009. In vitro 538
characterization of probiotic properties of Pediococcus pentosaceus BH105 isolated 539
from human faeces. Annals Microbiology, 59, 485-491. 540
541

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
21
Vesterlund, S., Paltta, J., Karp, M., Ouwehand, A. C., 2005. Measurement of bacterial 542
adhesion - in vitro evaluation of different methods. Journal of Microbiological 543
Methods, 60, 225-233. 544
545
Vidhyasagar, V., Jeevaratnam, K., 2013. Evaluation of Pediococcus pentosaceus strains 546
isolated from Idly batter for probiotic properties in vitro. Journal of Functional Foods, 547
5, 235-243. 548
549
Vinderola, C. G., Reinheimer, J. A., 2003. Lactic acid starter and probiotic bacteria: a 550
comparative "in vitro" study of probiotic characteristics and biological barrier 551
resistance. Food Research International, 36, 895-904. 552
553
Wibowo, D., Eschenbruch, R., Davis, C. R., Fleet, G. H., Lee, T. H., 1985. Occurrence 554
and growth of lactic acid bacteria in wine: a review. American Journal of Enology and 555
Viticulture, 36, 302–313. 556
557
Zago, M., Fornasari, M.E., Carminati, D., Burns, P., Suarez, V., Vinderola, G., 558
Reinheimer, J., Giraffa, G., 2011 Characterization and probiotic potential of 559
Lactobacillus plantarum strains isolated from cheeses. Food Microbiology, 28, 1033-560
1040. 561
562
Zavaglia, A. G., Kociubinski, G., Perez, P., De Antoni, G., 1998 Isolation and 563
characterization of Bifidobacterium strains for probiotic formulation. Journal of Food 564
Protection, 61, 865-873. 565

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
22
Table 1. Resistance to lysozyme and bile of the LAB strains studied
Resistance to lysozyme
(% Survival)
Resistance to bile
(% Growth)
t30 t120 0.06% 0.125% 0.25% 0.5% 1%
Enological strains
P. pentosaceus CIAL-16 83.1 70.8 100 100 95.7 88.9 83.3
P. pentosaceus CIAL-49 100 83.3 100 100 91.8 83.4 78.1
P. pentosaceus CIAL-85 84.3 76.1 97.2 98.3 86.9 82.1 77.4
P. pentosaceus CIAL-86 93.9 88.6 99.1 100 88.3 89.5 84.1
L. casei CIAL-51 75.0 78.6 96.0 94.8 92.9 77.3 61.7
L. casei CIAL-52 71.1 70.7 98.2 96.6 97.5 78.0 64.3
L. casei CIAL-92 100 88.6 97.7 96.0 91.1 87.1 80.4
L. plantarum CIAL-121 65.1 50.8 93.6 91.2 89.0 89.5 88.7
O. oeni CIAL-117 62.1 < 1.00 100 100 100 96.8 90.7
O. oeni CIAL-118 60.0 1.00 90.7 89.8 82.7 87.5 80.5
O. oeni CIAL-119 32.0 < 1.00 100 100 90.6 87.1 72.9
Reference strains
L. plantarum CLC 17 100 86.0 93.8 89.1 77.9 76.8 73.0
L. fermentum CECT5716 65.4 61.5 100 100 88.0 78.7 72.0
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
23
Table 2. Effect of simulated gastric juice on the counts (log CFU/mL) of wine LAB
strains studied at different pH values and incubation times
LAB strains Bacterial counts (log CFU/mL)
t0 t20 t40 t60 t90
pH 5.0 pH 4.1 pH 3.0 pH 2.1 pH 1.8
Enological strains
P. pentosaceus CIAL-16 8.54 ± 0.09 8.65 ± 0.07 8.29 ± 0.08 7.65 ± 0.07 5.34 ± 0.12
P. pentosaceus CIAL-49 8.59 ± 0.05 8.55 ± 0.05 8.37 ± 0.19 7.95 ± 0.01 5.51 ± 0.06
P. pentosaceus CIAL-85 8.60 ± 0.01 8.56 ± 0.13 8.60 ± 0.23 7.48 ± 0.01 5.80 ± 0.04
P. pentosaceus CIAL-86 8.47 ± 0.05 8.38 ± 0.11 8.41 ± 0.04 7.95 ± 0.06 5.15 ± 0.21
L. casei CIAL-51 8.02 ± 0.06 8.10 ± 0.02 7.92 ± 0.13 7.27 ± 0.08 7.07 ± 0.10
L. casei CIAL-52 7.92 ± 0.13 7.86 ± 0.03 7.96 ± 0.17 7.34 ± 0.08 7.17 ± 0.24
L. casei CIAL-92 8.02 ± 0.11 7.96 ± 0.16 7.95 ± 0.15 7.70 ± 0.21 5.32 ± 0.01
L. plantarum CIAL-121 8.08 ± 0.07 8.06 ± 0.08 8.05 ± 0.13 6.20 ± 0.28 5.05 ± 0.38
O. oeni CIAL-117 8.82 ± 0.05 8.78 ± 0.01 8.84 ± 0.05 8.42 ± 0.04 5.04 ± 0.06
O. oeni CIAL-118 8.63 ± 0.01 8.62 ± 0.10 8.59 ± 0.05 8.40 ± 0.02 5.30 ± 0.14
O. oeni CIAL-119 8.59 ± 0.08 8.54 ± 0.02 8.46 ± 0.12 7.24 ± 0.29 4.89 ± 0.28
Reference strains
L. plantarum CLC 17 8.08 ± 0.07 8.19 ± 0.10 8.08 ± 0.11 7.95 ± 0.06 7.31 ± 0.12
L. fermentum CECT5716 8.59 ± 0.05 8.18 ± 0.01 8.18 ± 0.07 7.46 ± 0.15 6.74 ± 0.13
Average values (± SD) from three independent repetitions are presented.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
24
Figure 1. Adhesion percentage of lactic acid bacteria strains to Caco-2 cells. Each adhesion assay was conducted in triplicate. Results are shown as media ± standard deviation.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
25
Figure 2. Anti-adhesion assays (competition, inhibition and displacement) of E. coli CIAL 153 in presence of L. plantarum CLC 17, P. pentosaceus CIAL-86 and L. plantarum CIAL-121. Results are shown as media ± standard deviation.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTEnological LAB strains as potential probiotic candidates to exert beneficial effects on human health.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT• We assessed probiotic properties of eleven enological lactic acid bacteria strains
• Good adaptation of wine strains to gastrointestinal conditions was observed
• The adherence of the wine strains to the intestinal epithelium was strain dependent