aspects of natural cold tolerance in ectothermic animals
TRANSCRIPT

Human Reproduction Vol. 15, (Suppl. 5) pp. 26-46, 2000
Aspects of natural cold tolerance in ectothermic animals
Hans Raml0y
Department of Life Sciences and Chemistry, Roskilde University, P.O.Box 260, DK 4000 Roskilde,Denmark
E-mail: [email protected]
Polar, alpine and temperate ectothermic (cold-blooded) animals encounter temperatures belowthe melting point of their body fluids eitherdiurnally or seasonally. These animals havedeveloped a number of biochemical and physio-logical adaptations to survive the low temper-atures. The problems posed to the animalsduring cold periods include changes in mem-brane and protein structure due to phasechanges in these molecules, changes in electro-lyte concentrations and other solutes in the bodyfluids as well as changes in metabolism. Cold-tolerant ectothermic animals can be divided intotwo groups depending which of two 'strategies'they employ to survive the low temperatures:freeze-tolerant animals which survive ice forma-tion in the tissues and freeze-avoiding animalswhich tolerate the low temperatures but notcrystallization of the body fluids. The adapta-tions are mainly directed towards the controlor avoidance of ice formation and include thesynthesis of low mol. wt cryoprotectants, ice-nucleating agents and antifreeze proteins. How-ever, some of the adaptations such as the syn-thesis of low mol. wt cryoprotectants are alsomore specific in their mechanism, e.g. directstabilizing interaction with membranes and pro-teins. The mechanisms employed by suchanimals may offer ideas and information onalternative approaches which might be usefullyemployed in the cryopreservation of cells andtissues frequently required in assisted repro-ductive technology.
26
Key words: antifreeze protein/cold tolerance/cryo-protectant/ice-nucleating agent/membrane
Introduction
Animals living in polar, temperate and alpineenvironments are either on a daily or yearly basissubjected to temperatures well below the freezingpoint of their body fluids. Liquid water is necessaryfor all life processes, therefore these animals musteither avoid freezing of body fluids but survivethe low temperatures or be able to endure iceformation in body fluids. A number of such animalsare endothermic and survive the low temperaturesby way of insulation such as fur or fat. Theectothermic (cold-blooded) animals, in which bodytemperature follows the surrounding environment,have a number of options when the temperaturefalls: they can hide in microhabitats where theyare not exposed to low temperatures; they canleave the area and return when conditions becomemore hospitable; or they can adapt to the lowtemperatures via a number of morphological, ana-tomical, biochemical and physiological features.The present review focuses on this last group ofanimals and their biochemical and physiologicaladaptations. In view of the temperature sensitivityof mammalian gametes and embryos, importantclues to the potential effects of cold and theiravoidance may be gained from comparative studies.
Damages due to cold per se
Animals living in areas where they are exposed tolow temperatures, or temperatures that are lower
© European Society of Human Reproduction and Embryology
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
than the temperature at which the animals arenormally active, may suffer damage due to thecold per se. The temperature does not need to fallbelow the melting point of the body fluids to causedamage. Damage caused by the cold per se isdue to either changes in metabolism or to phasetransitions in membranes and proteins as a resultof the low temperatures.
Phase changes in membranes
Biological membranes are bilayers of 20-80%lipids, mostly phosphoglycerides, with one primaryhydroxyl group esterified to phosphoric acid andthe other hydroxyl groups esterified to fatty acids.The phosphoglycerides also contain a polar headgroup often in the form of an amino alcohol, whichis esterified to the phosphoric acid via its hydroxylgroup. This arrangement gives rise to amphipathiccompounds or 'polar lipids', because of their polarheadgroups and their non-polar hydrocarbon tailsconsisting of 16, 18, 20 or 22 carbon atoms(Lehninger, 1975; Grout and Morris, 1987). Thebilayers are formed as a consequence of the amphi-pathic nature of the phospholipids. In the presenceof water the hydrophilic headgroups are exposedto the water and the hydrophobic hydrocarbonchains form the core of the membrane. The integrityof the biological membrane is determined byseveral factors such as Van der Waals forces(electrostatic interactions), salt bridges, hydrogenbonds and, perhaps more importantly, thermodyn-amic relations such as hydrophobic interaction andentropy. The hydrogen bonding of numerous watermolecules to each other is one of the strongestforces driving the membrane into its lamellarconfiguration (van Oss and Good, 1996). Mem-brane structure is dependent on temperature, pH,ionic strength of the surrounding medium and thestate of hydration (Williams, 1990).
During cooling, the initial effect is an increasein membrane viscosity (Grout and Morris, 1987).Upon further cooling, phase separations are likelyto occur. A characteristic of bilayers of a pure lipidis the phase transition temperature (Tc) abovewhich the lipid bilayers are found in a disorderedphase called the liquid crystalline state and belowwhich the bilayer is found in the more orderedgel state (Morris and Clarke, 1987). Efficient
membrane function requires the liquid crystallinephase, in which the membrane is strain-free, sothat hydrophobic regions of proteins and the lipidbilayer can be matched (Bloom, 1998). It shouldbe noted that if the hydration of the membranechanges, the lipids may go through the liquidcrystalline to gel transition and even reach a phasecalled the hexagonal II (Hn) phase, where thelipids organize into a non-lamellar three-dimen-sional matrix with the hydrophilic headgroupspointing inwards towards 'channels' of water whilethe hydrophobic hydrocarbon chains are pointingtowards each other (Quinn, 1985).
Biological membranes are not composed of onlyone pure lipid but rather of many—up to 200different lipids are found in the membranes ofsome biological systems (Morris and Clarke, 1987).Differential calorimetric studies have shown thata mixture of a saturated and a non-saturated lipidgives rise to two distinct endotherms if the mixtureis heated from below the liquid-crystalline phasetemperature (Tc) for both lipids to a temperatureabove the Tc for both lipids. These endothermsarise from the phase transitions of laterally phase-separated domains of the unsaturated and saturatedlipids respectively. The two lipids were separatedinto domains that consisted of the pure lipid ofone or the other. The saturated lipids stayed inthe gel phase at a higher temperature than theunsaturated lipids but on further heating these alsounderwent a phase transition and entered the liquidcrystalline phase (Quinn, 1985; Gennis, 1989).Chapman et al. (1977) have proposed a model forthe occurrences in the biological membrane duringcooling. They suggest that cooling a biologicalmembrane below the Tc of the lipids leads to lateralphase separations and that it also has profoundeffects on the distribution of the proteins boundto or integrated into the membrane. When themembrane is cooled, some lipids function as 'nuc-leation sites' and undergo a phase transition, crys-tallizing into 'islands' of the gel phase in whichproteins become trapped. Along the edges of these'islands' packing faults are likely and as theproximity of the proteins increases these may beginto aggregate (Quinn, 1985).
The consequences of the thermotropic behaviourof membrane lipids are diverse. A number of the
27
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
membrane components are free to diffuse withinthe membrane. Phase separations and the increasein membrane viscosity will have effects on thekinetics of diffusion-controlled processes (Groutand Morris, 1987). Enzymes associated with themembranes may become clustered into smalldomains of liquid lipid and this may have variouseffects on membrane function; there may be anincreased probability of the enzymes getting intocontact with their substrates which may lead toincreased enzyme activity (Grout and Morris,1987). In contrast, such an aggregation of theenzymes could also impede the transport of variousmolecules across the membrane, e.g. whereaggregations of enzymes may deplete an area ofsubstrate in the immediate vicinity of the enzymes.Here, transportation rates would decrease anddepend upon the diffusion of new substrate to theenzyme aggregation in question.
The boundaries between the gel phase and theliquid-crystalline phase are known to be especiallyleaky (Williams, 1990). Leaky areas will tend todispel the electrochemical gradients across themembranes as well as possibly causing the leakageof potentially damaging substances into the cells(e.g. increasing intracellular calcium). Aroundintegral proteins there is another possibility ofleakage because cooling may impair the abilityof non-bilayer forming lipids, also found in themembranes, to seal the boundaries between theseproteins and the membrane (Quinn and Williams,1985). Most of the evidence for the above-men-tioned effects of cooling on membranes was gainedvia studies of relatively simple model systemspossibly only consisting of a single or a mixture oftwo lipids and perhaps a single protein. Biologicalmembranes are of course infinitely more complexwith their large array of different lipids and bothbound and integral proteins. However, there areseveral examples of effects of cooling predictedby the above-mentioned models (for a review seeGrout and Morris, 1987).
Structural transitions in proteins
Proteins are of the utmost importance to all biolo-gical processes and to the structure of cells andliving systems in general. Therefore the mainten-ance of structure and function of proteins during
exposure to low temperature is imperative if thebiological system is to survive.
According to Franks and Hadley (1992), thecold denaturation temperatures of most proteinswhen examined in the pH range of maximalstability lie below the equilibrium freezing pointof water. For the majority of the examined proteinsthis means below -15°C. The stability of proteinsin solution is very limited (Franks, 1985) andpresumably based on a number of contributingfactors, which can be divided into stabilizing anddestabilizing factors. The former are hydrophobicinteractions, intrapeptide attractive effects (hydro-gen bonding, salt bridges, van der Waals inter-actions) whilst the latter are core repulsion,configurational entropy and solvation effects(Franks and Hadley, 1992). The increasing magni-tude of the latter, particularly solvation of non-polar(hydrophobic) moieties upon cooling, is thought tobe the molecular origin of cold denaturation (Hvidtand Westh, 1998). Denaturation of proteins at lowtemperatures is often reversible (Franks, 1985),e.g. tubulin depolymerizes at low temperaturesbut polymerizes when the temperature returns tonormal (Timasheff, 1978). However, some proteinsdenature non-reversibly during cooling, due mainlyto the formation of aggregates, e.g. urease (Hofstee,1949). Another example is phosphofructokinase, akey enzyme in glycolysis (Carpenter et ai, 1986)which is dissociated from a tetramer into twodimers, leading to the synthesis of sucrose fromfructose instead of the phosphorylation of fructose(Franks, 1985; Storey and Storey, 1992).
Activity and metabolism in animals at lowtemperatures above 0°C
When an ectothermic animal is cooled below itsusual temperature of activity, it passes through anumber of states before it eventually dies. The firststate, which is observed below the temperature ofoptimal activity, is called cold stupor (Klok andChown, 1997). In this state the animal becomesmore and more sluggish and eventually does notmove at all when reaching what is called chillcoma (Vannier, 1987; Block et al., 1992). It shouldbe noted that some animals do not survive thesestages for more than a few minutes (Lee, 1991)whereas other animals, namely the cold-adapted
28
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
can survive for several months or perhaps evenlonger (Block, 1990).
The reduction in activity is accompanied bya temperature-dependent reduction in respiration(Block, 1990; Block et al., 1998; Raml0v, unpub-lished observations). This can be assessed bymeasurements of Q10, defined as the ratio betweenmetabolic rates recorded at two temperatures andextrapolated to a 10°C difference:
Q10 = R,/R2*[10/(T1-T2)] .
Block (1977) reported that the Q10 for Antarcticland invertebrates is 3.04. Similar relationships areseen in other ectothermic animals although thevalue may vary depending on species and thetemperature range over which the Q10 is measured(Davenport, 1992).
The relationships between decreasing temper-ature, locomotion and respiration are not surprisingconsidering changes in membrane and proteinstructure mentioned above. Additionally, mostenzymes have a temperature range in which theyfunction optimally, and when, as the temperaturefalls outside this range, kinetics change and activityalso decreases (for discussions see Hochachka andSomero, 1984; Franks, 1985). Locomotion maybe impaired due to depolymerization of actinfilaments, and cytoplasmic streaming may slowdown or cease (Grout and Morris, 1987). Cyto-plasmic streaming is highly dependent on Ca2+
concentrations and ATP production and both maybe changed during cooling, e.g. intracellular Ca2+
concentration may change because of leakage ofCa2+ into the cells from the extracellular fluidbecause of lateral phase separations in the mem-brane (see 'Phase changes in membranes' above).Chilling injury can also be caused by the elasticstress that occurs when the membrane condensesto a greater extent than the contents of the cell(McGrath, 1987).
Protein synthesis may also be impaired or slowedby a decrease in temperature (Grout and Morris,1987). 'De novo' synthesis of proteins will nottake place and this may have a bearing on how longanimals are able to tolerate the low temperature. Ifleakage occurs, ion gradients and the membranepotential break down too. As the membrane poten-tial is important for all transport processes in
and out of cells, its breakdown interferes withmetabolite transfer across cell membranes. This inturn could lead to depletion of necessary substancesinside cells or to the accumulation of potentiallydangerous metabolic waste products. Thus, variousanimals are able to endure deceasing temperaturesto various extents and times depending upon theirability to withstand, counteract or repair the cellularchanges mentioned above.
The freezing process
When the temperature falls below the melting pointof the organism's body fluids there is a potentialfor ice formation in the organism. Nucleation ofice crystals may occur in water solutions at anytemperature below the melting point of the solution.If ice is not formed in the solution at its meltingpoint the solution is supercooled or undercooled.The supercooled solution is metastable as it isenergetically more favourable for the solution tobe in the frozen or partly frozen state than in theliquid state at this temperature. Crystallization maytherefore occur spontaneously at any time by thewater molecules aggregating into an ice nucleus(homogeneous nucleation), or by the water molec-ules aggregating around some substance or irregu-larity on a surface that lowers the activation energyof the crystallization (heterogeneous nucleation)(Franks, 1985; Williams and Carnahan, 1990).Nucleation is a time-dependent stochastic process(Vali, 1995) which is dependent on the probabilityof a sufficient number of water molecules to forma structure (cluster) that gives rise to an 'embryoice crystal' (Rasmussen and MacKenzie, 1973;Vali, 1995). These structures arise spontaneouslyas a function of random density fluctuations andtheir lifetime depends on their self-diffusion rate,which is temperature dependent (Franks, 1985).When the ice embryo reaches a critical size itbecomes a nucleus and this event is called nucle-ation (Vali, 1995).
There are therefore four factors to be consideredwhen dealing with nucleation events, (i) The tem-perature itself: the lower the temperature, thesmaller the number of water molecules requiredto make an ice embryo, thus the probability ofnucleation increases (Vali, 1995). (ii) The largerthe volume of the sample the higher the probability
29
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
Figure 1. The freezing process. Changes in cell size and solute concentration during freezing: (a) a single cell in solution, (b)extracellular nucleation occurs (*), (c) freeze concentration of the extracellular fluid with following cell shrinkage due toosmotic outflow of water from the cell, (d) totally solidified system; cell highly shrunken with intra- and extracellular eutecticprecipitation.
of a sufficient number of water molecules formingan ice embryo (Vali, 1995). Thus larger volumesusually freeze closer to the melting point thansmall volumes within any given time. In purewater, the homogeneous nucleation point is about-39°C, a temperature which can only be reachedin very small volumes (Angell, 1982). (iii) Acertain volume in the metastable supercooled statewill freeze at some time that depends on thetemperature (Vali, 1995). (iv) Presence of nucleat-ing agents in the system causing heterogeneousnucleation.
Once freezing is initiated, an amount of iceforms from water molecules which concentratesthe remaining solution to the extent where its watervapour pressure equals that of the vapour pressureof water over the ice at that specific temperature.This is called freezing concentration and occursbecause ice consists of pure water and only veryfew substances can be incorporated into the ice.
A biological entity (Figure 1) contains a large
number of various substances, many of which maygive rise to heterogeneous nucleation. Ice formationis usually initiated in the extracellular fluid (Figurelb). When ice begins to form, the extracellularfluid becomes increasingly concentrated (Figurelc). The vapour pressure of the remaining fluid isin equilibrium with the vapour pressure of the watervapour over the ice at the specific temperature. Theincrease in solute concentration of the extracellularfluid results in an osmotic outflow of water fromthe cells so that the osmolality of the intracellularfluid and thus the water vapour pressure is inequilibrium with the extracellular fluid which againis in vapour pressure equilibrium with the ice(Mazur, 1984). If the temperature continues todecrease further, more ice is formed and eventuallythe eutectic point of various substances is reached.At this point, ice and the solutes precipitate simul-taneously in a eutectic composition (Moore, 1981)(Figure Id) and the whole system is solidified.
30
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
Problems related to freezing of the body fluids
Freezing in an animal is usually initiated extra-cellularly either in the gut, due to the presence ofexogenous nucleating agents (Salt, 1953;Zachariassen, 1985; Duman et al., 1995; Worlandet al., 1997), in the tissues as a consequence ofaccidental ice-nucleating substances or specificice-nucleating proteins which may be membranebound (Baust and Zachariassen, 1983; Dumanet al, 1991a; Tsumuki and Konno, 1991) or insolution in the blood or haemolymph (Dumanet al., 1995). Freezing may also be initiated viainoculation, i.e. the nucleation of the body fluidsby contact with external ice (Salt, 1963; Shimadaand Riihimaa, 1988; Layne et al, 1990). Intracellu-lar freezing, except in a few cases (Salt, 1959a,1962; Wharton and Ferns, 1995), is lethal(Mazur, 1984).
When extracellular fluid freezes there are severalpossibilities of damage to the system. Freezing ofa part of the body fluid can be looked upon asdrying of the system (Lee, 1991; Ring and Danks,1998). As crystallization proceeds, more water isremoved from its role as a solvent for the dissolvedsolutes. The resulting increase in solute concentra-tion may lead to changes in enzyme activity andprecipitation or denaturation of proteins (saltingout) (Hochacka and Somero, 1984). These effectscan to a certain extent be predicted by theHofmeister series of neutral salts. Changes inpH are also likely to occur during freezing anddehydration (Franks et al., 1990), which mayaffect enzyme activity and possibly lead to proteindenaturation (Taylor, 1987), changes in membranepotential and changes in membrane transport(Franks, 1985).
Osmotic outflow of water from the cells willcause the cells to shrink. This may decrease thecell volume below the so-called minimum volume(Lee, 1991), which is typically reached at a bodyice content of -65% of the total body water (Storeyand Storey, 1993). When the cell volume decreasesto this value the membrane begins to rest on theintracellular structures, causing hydrostatic stresson the membrane as the cell cannot shrink further.The hydrostatic forces eventually rupture themembrane.
The dehydration of the system by crystallization
also causes phase changes in the membranes dueto the removal of the forces keeping the membranein its bilayer conformation (see 'Phase changes inmembranes'). Finally, the growth of ice crystals inthe tissues may lead to rupture of these andsharp ice crystals may penetrate cells (Grout andMorris, 1987).
Depending on the cooling rate, freezing of abiological system may affect the cells of the varioustissues differently, and one cooling rate may killsome cells whereas others survive (Grout andMorris, 1987). Indeed it seems as if there is anoptimum cooling rate for most cell types (Mazuret al., 1972). Survival of different cells at variouscooling rates is described by a bell-shaped curve(Mazur et al, 1972). The shape of these curves isdescribed as the two-factor hypothesis of freezinginjury (Mazur et al, 1972). The shape of the curvesis explained by the prolonged exposure to highsolute concentrations at low cooling rates (Mazuret al, 1912) or to the decrease in size of theunfrozen spaces in which the cells lie (Mazur,1984) and to intracellular freezing at high coolingrates. The optimal cooling rate for various cellsrelies on a number of factors: the water permeabil-ity of the cell membrane, the cell surface to volumeratio and the hydraulic conductivity (Grout andMorris, 1987). Hence cells with a high surface tovolume ratio and high membrane water permeabil-ity may tolerate high cooling rates, as such cellslose water very fast and therefore are not in dangerof intracellular freezing due to supercooling of theintracellular fluid. The consequence of this is thatthe survival of various tissues can vary consider-ably within an organism or between cells in solutionwhich are not either adapted to or artificiallyprotected from freezing at a specific cooling rate.
Adaptations to temperatures below themelting point of the body fluids in ectothermic
animals
Cold-adapted ectothermic animals employ one oftwo 'strategies' when exposed to cold; freezeavoidance and freeze tolerance (Table I).
Freeze-avoiding animals do not tolerate crystal-lization of their body fluids and thus their super-cooling point (SCP) is equal to their lower lethaltemperature (LLT). Often such animals have a
31
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
Table I. Adaptations to temperatures below the melting point of the body fluids in ectothermic animals
Freezing avoidance Freeze tolerance
Supercooling point = lower lethal temperatureIce formation is lethalHigh supercooling capacityPolyols function as antifreeze (may also stabilizemembranes and proteins)- Ice-nucleating agentsAntifreeze proteins often present
Supercooling point =£ lower lethal temperatureSurvival of extracellular icePoor supercooling capacityPolyols function as colligative and non-colligativecryoprotectants+ Ice-nucleating agentsAntifreeze proteins intracellular and inhibits RI
RI = recrystalisation inhibition.
large supercooling capacity (SCC), which meansthat there is a large temperature difference betweenthe melting point of the body fluids and theSCR Freeze-avoiding animals have to survive coldperiods in the metastable supercooled state. Theyare therefore in constant danger of ice formationin their tissues. However, there are a number ofexamples of freeze-avoiding animals survivingtemperatures as low as approximately -25 °C forprolonged periods or even as low as -50 to -60°C(Miller, 1982; Ring, 1982) or periods of up topossibly 30-40 years supercooled by about 1 °C inthe Antarctic ocean (e.g. Dissostichus mawsoniwhich reaches weights of up to around 80 kg inAntarctic waters).
To survive low temperatures in the supercooledstate, nucleating agents either have to be absentfrom the tissues or masked during the cold period(Lee, 1991). It has been observed that some freeze-avoiding species cease to eat and empty their gutduring autumn before the temperatures fall belowthe melting point of the body fluids, therebyenhancing these organisms SCC (Cannon andBlock, 1988).
Freeze-tolerant animals, however, survive theformation of ice within their tissues, consequentlythe SCP of these animals is different from theirLLT. Due to the presence of ice-nucleating agents(INA) (Zachariassen and Hammel, 1976) or inocu-lation of the body fluid, the SCC of these animalsis small and the body fluids typically crystallize ata relatively high temperature below the meltingpoint (Zachariassen, 1980), usually -5 and -10°C(Zachariassen and Hammel, 1976; Zachariassen,1980) [e.g. the New Zealand alpine weta Hemide-ina maori (Raml0v etal., 1992)] and the crysomelid
beetle Phyllodecta laticollis (Laak, 1982) (see alsoTable II). Some freeze-tolerant animals, however,have very low supercooling points due to theremoval of virtually all ice nucleators from thesystem (Miller, 1982; Ring, 1982). A number ofectothermic animals are freeze-tolerant even duringthe summer period (Laak, 1982; Raml0v et al.,1992), presumably because they are exposed to lowtemperatures even during summer (Raml0v, 1999).
The reason why the two strategies mentionedabove have evolved in parallel is not known.Freeze-avoiding animals are more likely to experi-ence freezing events the further the temperaturedecreases; it could therefore be speculated thatthese animals are found in areas where temper-atures do not fall to very low extremes, whereasfreeze-tolerant animals whose body fluids are inthermal equilibrium with their surroundings maybe found in areas with extremely low wintertemperatures (Zachariassen, 1980; Ring, 1982).However, recently a number of species inhabitingareas in which the temperature only rarely fallsbelow -10°C have been shown to be freeze tolerant(Sinclair, 1997; Raml0v, 1999). Lundheim andZachariassen (1993) proposed that freeze toleranceis an adaptation to the desiccating conditionsencountered during winter. At low temperaturesthe air is usually very dry and unless an animalhas an impermeable integument it risks losingwater to the frozen surroundings if its body fluidsare supercooled [due to the higher chemical poten-tial (vapour pressure) of supercooled water thanthat of ice]. Indeed Lundheim and Zachariassen(1993) have shown that some freeze-avoidingbeetles have low cuticular water permeability.Another explanation for this observation may be
32
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
that it reduces the risk of inoculation from externalice crystals (Lundheim and Zachariassen, 1993).An explanation for the observation that someanimals are freeze-tolerant even though ambienttemperatures do not fall to low extremes maytherefore be that these animals overwinter in hab-itats which are very moist and thus the chance ofinoculation is high (Klok and Chown, 1997;Raml0v, 1999). This seems to be the case both forH.maori (Raml0v, 1999) and the New Zealandalpine cockroach Celattoblatta quinquemaculata(Sinclair, 1997).
Freeze tolerance may also have evolved as anadaptation to areas where ambient temperaturesshow considerable variation. In such areas, animalsmay experience diurnal freeze/thaw cycles or per-haps prolonged periods of temperatures above themelting point of the body fluids. Such conditionsmay call for the possibility of these animals toforage and thus they are exposed to ingestion ofINA during periods when freezing temperaturesmay occur. Again, this is the case for H.maoriwhich may encounter sub-freezing temperatures atany time of the year and where the animals areregularly found with the cuticle covered in icecrystals during winter (Raml0v, 1999).
The control of ice formation
One of the most important features of adaptationsto cold in ectothermic animals is the control of iceformation. This is achieved either by completeavoidance of ice formation and inoculation or,alternatively, control of the site of ice formation(extracellular/intracellular), the crystallization tem-perature (and hence the ice growth rate and osmoticequilibration intra- and extracellularly), the amountof ice formed (and hence the extent of dehydrationand freezing concentration of the body fluids) andfinally the control of recrystallization. Recrystal-lization is the growth of large ice crystals at theexpense of smaller ones. Cold-adapted ectothermicanimals have evolved a number of physiologicaland biochemical adaptations to achieve this control(Figure 2).
Cryoprotective low mol. wt substances
Most cold-tolerant ectothermic animals synthesizehigh or low mol. wt substances (Zachariassen,1985) that protect the organisms either against ice
formation or damage due to ice occurring intheir tissues.
Protective low mol. wt substances in cold-adapted animals can be divided into two classesbased upon their actions: (i) colligative cryoprotec-tants* (which affect vapour pressure or freezingpoint, depending upon the number of moleculesinvolved) and (ii) cryoprotectants which stabilizemembranes and proteins (Storey and Storey, 1992).A number of requirements have to be fulfilled ifthese substances are to control ice formation: (i)they have to be highly soluble in aqueous solution;(ii) they are relatively non-toxic and non-reactivetowards cells and macromolecules, even in highconcentrations; (iii) they have to be compatiblesolutes (for definition see Hochachka and Somero,1984) so that they do not perturb protein structureand function; (iv) they have to counteract thedenaturing effects on proteins of cold, dehydrationand high ionic concentrations (Storey and Storey,1992).
A number of species employ dual or multiplecryoprotectant systems [multifactorial systems(S0mme, 1982)]. This may have the advantagethat the concentration of none of the substancesreaches poisonous levels in the organism's bodyfluids (Ring, 1980). It has also been proposedthat a combination of certain commonly foundcryoprotective substances increases the possibilityof vitrification of the body fluid (Wasylyk et al.,1988). However, this theory is based upon datawith artificial 'haemolymph' where the solutespecies and concentrations are chosen to emulatethose in the freeze-tolerant larva of the gall flyEurosta solidaginis. Vitrification has not beenobserved under natural conditions in animals.According to Storey and Storey (1992) the employ-ment of multiple cryoprotectant systems may alsohave metabolic advantages. For example, sorbitoland glycerol pools have different fates in spring;sorbitol being converted into glycogen, whereas
*Colligative comes from the word 'colligare' meaning'to glue together'. In the chemical context this meansthe properties of a solution that depend on the numberof particles (molecules) involved and not on thequality of the particles. The properties 'glued together'are, for example, vapour pressure, freezing (melting)point depression and boiling point elevation.
33
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018
H.Raml0v
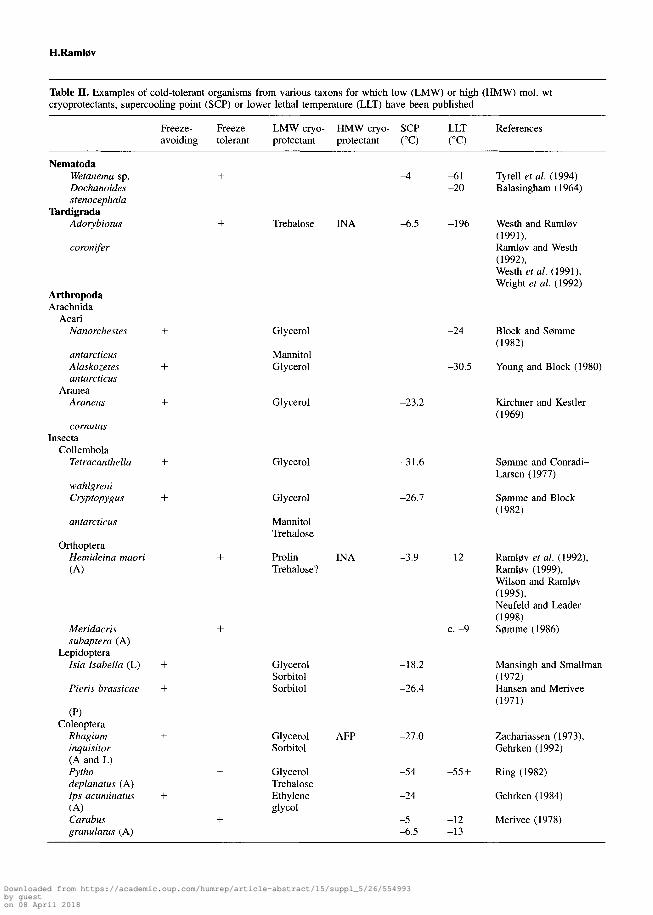
Table II. Examples of cold-tolerant organisms from various taxons for which low (LMW) or high (HMW) mol. wtcryoprotectants, supercooling point (SCP) or lower lethal temperature (LLT) have been published
Freeze- Freeze- LMW cryo- HMW cryo- SCP LLTavoiding tolerant protectant protectant (°C) (°C)
References
NematodaWetanema sp.Dochanoidesstenocephala
TardigradaAdorybiotus
coronifer
ArthropodaArachnida
AcariNanorchestes +
antarcticusAlaskozetes +antarcticus
AraneaAraneus +
cornutusInsecta
CollembolaTetracanthella +
wahlgreniCryptopygus +
antarcticus
OrthopteraHemideina maori(A)
Meridacrissubaptera (A)
LepidopteraIsia Isabella (L)
Pieris brassicae
(P)Coleoptera
Rhagiuminquisitor(A and L)Pythodeplanatus (A)Ips acuminatus(A)Carabusgranulatus (A)
Trehalose
Glycerol
MannitolGlycerol
Glycerol
Glycerol
Glycerol
MannitolTrehalose
ProlinTrehalose?
GlycerolSorbitolSorbitol
GlycerolSorbitol
GlycerolTrehaloseEthyleneglycol
INA
INA
AFP
-4
-6.5
-61-20
-196
-24
Tyrell et al. (1994)Balasingham (1964)
Westh and Raml0v(1991),Raml0v and Westh(1992),Westh etal. (1991),Wright et al. (1992)
Block and S0mme(1982)
-30.5 Young and Block (1980)
-23.2
-31.6
-26.7
-3.9
-18.2
-26.4
-27.0
-54
-24
-5-6.5
-12
c. -9
-55 +
-12-13
Kirchner and Kestler(1969)
S0mme and Conradi-Larsen (1977)
S0mme and Block(1982)
Raml0v et al. (1992),Raml0v (1999),Wilson and Raml0v(1995),Neufeld and Leader(1998)S0mme (1986)
Mansingh and Smallman(1972)Hansen and Merivee(1971)
Zachariassen (1973),Gehrken (1992)
Ring (1982)
Gehrken (1984)
Merivee (1978)
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018
Cold tolerance in ectotherms
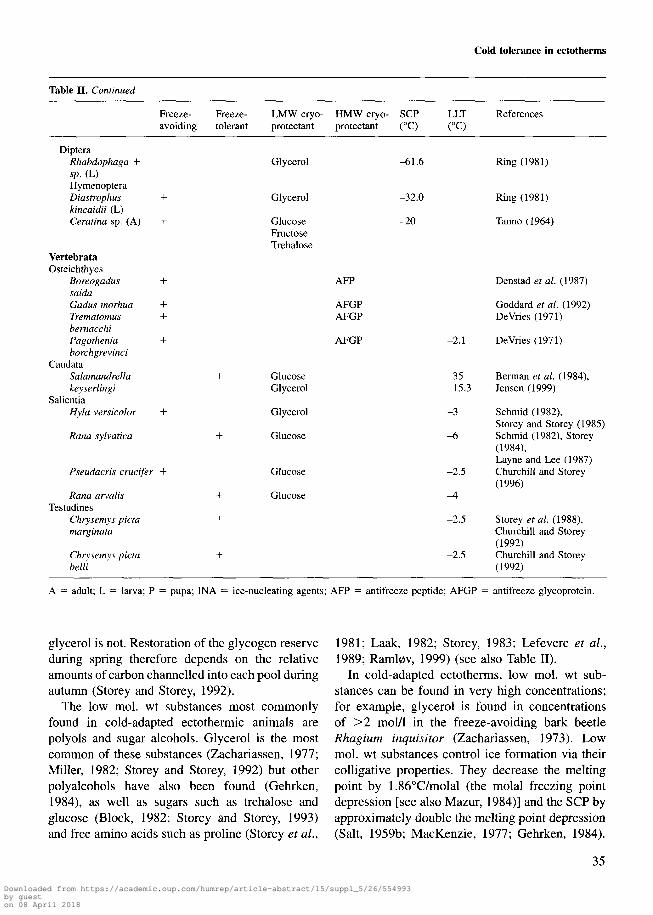
Table II. Continued
Freeze- Freeze- LMW cryo- HMW cryo- SCP LLTavoiding tolerant protectant protectant (°C) (°C)
References
DipteraRhabdophaga +sp. (L)HymenopteraDiastrophuskincaidii (L)Ceratina sp. (A)
VertebrataOsteichthyes
Boreogadus +saidaGadus morh.ua +Trematomus +bernacchiPagothenia +borchgrevinci
CaudataSalamandrellakeyserlingi
SalientiaHyla versicolor +
Rana sylvatica
Pseudacris crucifer +
Rana arvalisTestudines
Chrysemys pictamarginata
Chrysemys pictabelli
Glycerol
Glycerol
GlucoseFructoseTrehalose
-61.6
-32.0
-20
Ring (1981)
Ring (1981)
Tanno (1964)
GlucoseGlycerol
Glycerol
Glucose
Glucose
Glucose
AFP
AFGPAFGP
AFGP -2.1
-35-15.3
-3
-6
-2.5
-A
-2.5
-2.5
Denstad et al. (1987)
Goddard et al. (1992)DeVries (1971)
DeVries (1971)
Berman et al. (1984),Jensen (1999)
Schmid (1982),Storey and Storey (1985)Schmid (1982), Storey(1984),Layne and Lee (1987)Churchill and Storey(1996)
Storey et al. (1988),Churchill and Storey(1992)Churchill and Storey(1992)
A = adult; L = larva; P = pupa; INA = ice-nucleating agents; AFP = antifreeze peptide; AFGP = antifreeze glycoprotein.
glycerol is not. Restoration of the glycogen reserveduring spring therefore depends on the relativeamounts of carbon channelled into each pool duringautumn (Storey and Storey, 1992).
The low mol. wt substances most commonlyfound in cold-adapted ectothermic animals arepolyols and sugar alcohols. Glycerol is the mostcommon of these substances (Zachariassen, 1977;Miller, 1982; Storey and Storey, 1992) but otherpolyalcohols have also been found (Gehrken,1984), as well as sugars such as trehalose andglucose (Block, 1982; Storey and Storey, 1993)and free amino acids such as proline (Storey et al.,
1981; Laak, 1982; Storey, 1983; Lefevere et al.,1989; Raml0v, 1999) (see also Table II).
In cold-adapted ectotherms, low mol. wt sub-stances can be found in very high concentrations;for example, glycerol is found in concentrationsof >2 mol/1 in the freeze-avoiding bark beetleRhagium inquisitor (Zachariassen, 1973). Lowmol. wt substances control ice formation via theircolligative properties. They decrease the meltingpoint by 1.86°C/molal (the molal freezing pointdepression [see also Mazur, 1984)] and the SCP byapproximately double the melting point depression(Salt, 1959b; MacKenzie, 1977; Gehrken, 1984).
35
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
CONTROL OF ICE FORMATION
Low molecular weight cryoprotective substances
• Polyols(e.g. Glycerol, Ethylene glycol, Sorbitol)
• Sugars(e.g. Glucose, Trehalose)
• Free amino acids(e.g. Proline)
COLLIGATIVE CONTROL OF:Melting pointRate of ice formationIce fraction sizeSupercooling point
Proteinaceous cryoprotective substances
• Ice nucleating proteinsSynthesized or ingested
Proteins/lipoproteins77 kDa - 800 kDa
CONTROL OF:Supercooling point
• Antifreeze proteins
Proteins/glycoproteins3.2 kDa-32 kDa
CONTROL OF:Ice growth andrecrystallisation
Figure 2. Various adaptations to the control of ice in cold-tolerant ectothermic animals.
These two properties are important in freeze-avoiding animals which have to maintain liquidbody fluids during cold periods.
In freeze-tolerant animals, low mol. wt sub-stances decrease the amount of ice formed at anygiven temperature (Zachariassen, 1979) and slowthe rate of ice formation, e.g. the ice content ofE.solidaginis reaches -65% in -40 h (Lee andLewis, 1985). This animal accumulates sorbitoland glycerol during autumn amounting to 400 fimolglycerol/g dry weight and -120 |umol sorbitol/gdry weight as well as lower concentrations oftrehalose and glucose (Storey and Storey, 1992).In contrast, H.maori does not accumulate highconcentrations of low mol. wt substances. Theosmolality of its haemolymph increases from about350 mOsm in summer to -700 mOsm in winter,mainly due to the accumulation of proline (Raml0v,1999). In this animal, ice formation after nucleationis rapid and the ice content reaches -80% of thebody water in -6 h (Raml0v and Westh, 1993).The reduction of ice formed in the organism
limits the dehydration and is of significance inmaintaining a critical minimum cell volume. Inthe case of H.maori it must be assumed that itscells can survive extensive dehydration and thushave a very small critical minimum volume(Raml0v and Westh, 1993). Fat body cells andMalpighian tubules from H.maori undergo extens-ive dehydration during freezing, as observed bycryo-microscopy. Cells frozen to -8°C showedhigh survival upon thawing whereas cells cooledto -15°C and below showed a decline in survivalcorrelated with temperature (Sinclair andWharton, 1997).
Proteinaceous substances as special adaptationsto the control of ice formation
Cold-adapted ectothermic organisms synthesize anumber of proteinaceous substances that in variousways participate in the control of ice formation. Thesubstances in question are INA and the antifreeze/thermal hysteresis proteins (for reviews seeDeVries, 1982, 1986; Zachariassen, 1985; Cheng
36
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
and DeVries, 1991; Duman et al, 1991a, 1993,1995; Hew and Yang, 1992; Costanzo and Lee,1995).
Ice-nucleating agents
Extracellular INAs are found in a large number offreeze-tolerant animals (Aunaas, 1982; Duman,1982; Zachariassen, 1982, 1985; Duman andHorwath, 1983; Loomis, 1985; Duman et al.,1991a; Westh et al., 1991), but not in all species(Miller, 1982; Ring, 1982; Costanzo and Lee,1996). INAs inhibit extensive supercooling byinitiating ice formation via heterogeneous nucle-ation at relatively high temperatures below themelting point of the extracellular fluid(Zachariassen and Hammel, 1976; Zachariassen,1982; Duman and Horwath, 1983), preventinglethal osmotic shock (Lee, 1991; Zachariassen,1992) and intracellular ice (Zachariassen and Ham-mel, 1976; Duman, 1982; Zachariassen, 1982).INAs are thought to provide a template aroundwhich an embryonic ice crystal can form and growto become large enough to ensure freezing of asupercooled liquid; that is, a supercritical radius ata given supercooling (Burke and Lindow, 1990).INAs are found in the haemolymph (Duman et al.,1995), associated with different tissues (Baust andZachariassen, 1983; Duman et al., 1991a; Tsumukiand Konno, 1991) or in the gut contents (Dumanet al., 1995; Worland et al., 1997). There is muchevidence that INAs in the haemolymph of insectsand other animals are either proteins or lipoproteins(Duman and Horwath, 1983; Hayes and Loomis,1985; Neven et al., 1989; Duman et al, 1991b;Wilson and Raml0v, 1995) but can also be foundin some tissues as crystals (Mugnano et al., 1996).Ice-nucleating proteins are large, ranging in sizefrom 74 to 800 kDa, the latter having a diameterof 135A (Duman et al, 1984; Neven et al, 1989)presumably with rough surfaces and with manyhydrophilic residues extending into the solvent(Burke and Lindow, 1990; Wilson and Raml0v,1995). The size of the proteins determines at whichtemperature they initiate ice formation. Biologicalice nucleation theory predicts that, for example,an ice nucleator with a diameter of 300A willinitiate ice formation at a temperature of -3 to-8°C, depending on the geometry and contact
angle 0 between the ice nucleator and the iceembryo (Govindajaran and Lindow, 1988; Burkeand Lindow, 1990; Wilson, 1994; Vali, 1995).The lipoprotein ice nucleator (LPIN) is well-characterized, consisting of two apolipoproteins of265 and 81 kDa respectively. In LPIN the lipidcomponent is phosphatidylinositol (PI). It has beensuggested by Warner (1962) that inositol can orderwater molecules into an ice-like structure (Dumanet al, 1995). It was shown by Neven et al. (1989)that the PI component of LPIN is essential for ice-nucleation activity. Therefore it was speculatedthat the PI component of LPIN forms the templatearound which the water molecules orientate them-selves into the embryonic ice crystal (Dumanet al, 1991b).
It has also been speculated that some INAsaggregate and thereby form a more efficient icenucleator because more water molecules can bearranged into ice-like clusters (Mueller et al,1990). Duman et al (1992) showed that an increas-ing concentration of LPIN increased in ice-nucleat-ing activity, producing a maximal SCP of -6°C atconcentrations >1.7X10~7 mol/1. This indicatesthat aggregation and cooperation of several LPINmolecules is required to induce maximal nucle-ation activity.
In a number of cold-tolerant animals there is adifference between the SCP of the haemolymphand the whole body SCP, e.g. in H.maori the SCPof the haemolymph is about -7.5°C (Wilson andRaml0v, 1995) whereas the whole body SCP isabout -4°C (Raml0v et al, 1992). This differencecan have several reasons: (i) possibly there areINAs in the gut contents which induce the crystal-lization at about -4°C; (ii) the INAs which inducefreezing at -4°C are associated with various tissues;or (iii) the INAs in haemolymph are more activein vivo than in vitro. Why then does H.maori haveINAs in the haemolymph? One reason could bethat the ice nucleators in the haemolymph are justincidental ice nucleators, that is, some substance(in this case a protein) which has ice-nucleatingactivity but which serves a different function inthe animal. Another reason could be that H.maoriusually relies on ice nucleation from the gutcontent, but in some cases the animals may bestarved and the ice nucleators in the haemolymph
37
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
then serve as a 'back-up' system. There is probablya high selection pressure for ice nucleation as noice nucleation in a freeze-tolerant animal maybe lethal.
Antifreeze proteins
Another group of proteinaceous substances control-ling ice formation are the antifreeze or thermalhysteresis proteins. These proteins are found inpolar fishes that inhabit waters at or close to themelting point of seawater (-1.9°C) year round orseasonally (DeVries, 1982, 1986; Denstad et al,1987; DeVries and Cheng, 1992; Goddard et al,1992) and in invertebrates inhabiting areas wherethey at some time during the year are exposed tocold (Duman et al, 1991a, 1992, 1993). In contrastto agents which initiate ice formation the antifreezeproteins inhibit the growth of ice by interactingwith specific crystal planes on the ice crystal(DeVries and Cheng, 1992) or possibly stabilizethe metastable supercooled state by recognizingembryo ice crystals before they grow large enoughto initiate ice growth in the solution (Zachariassenand Husby, 1982). Further, antifreeze proteinshave been shown to inhibit recrystallization of ice(Ramsay, 1964; Knight and Duman, 1986; Knightet al., 1995). There may also be some proteins thatdo not show any 'antifreeze' activity but whichnevertheless inhibit recrystallization (Raml0vet al, 1996).
Substances with 'antifreeze' effects weredescribed for the first time by Ramsay (1964). Inhis comprehensive study of the cryptonephridialrectal complex of the mealworm Tenebrio molitor,Ramsay employed a melting point apparatus in thestudy of osmolalities in fluid from the rectalcomplex. During his study he observed that icecrystals decreased in size when the temperaturewas raised, but that they did not increase in sizewhen the temperature was lowered until a certaintemperature was reached at which the ice crystalssuddenly grew instantaneously and the wholesample solidified (Ramsay, 1964). On some occa-sions Ramsay observed that the ice crystals in thefluid did not increase in size before the temperaturewas lowered as much as 10°C. Antifreeze proteinsproduce, by a non-colligative mechanism (Westhet al., 1997), a separation of the melting point and
the temperature at which an ice crystal will grow.This is called the thermal hysteresis activity andthe temperature at which the ice crystal grows istermed the hysteresis freezing point (DeVries,1986). Although antifreeze activity was firstobserved in an insect larva, antifreeze proteinshave been studied the most in teleost fishes (Feeney,1974; DeVries and Cheng, 1992; Deng andLaursen, 1998). Scholander and colleagues (1957)were puzzled by the fact that teleost fishes in thearctic, whose blood had a melting point of -0.5to -0.8°C, live in close contact with ice in watersat temperatures around -1.7 to -1.8°C, thus beingsupercooled by almost 1°C but with no inoculationof the body fluids occurring. Scholander et al.(1957) did not identify the substances responsiblefor the inhibition of inoculation and/or formation ofice in the fishes. However, DeVries and Wohlschlag(1969), who worked in the Antarctic, discoveredthat a glycoprotein could explain 30% of thefreezing point depression in the serum obtainedfrom fishes in McMurdo Sound. Since then, anumber of different types of antifreeze proteinshave been discovered in polar fishes, all synthesizedin the liver and secreted into the blood (Cheng andDeVries, 1991). Today five types are known, (i)The glycoproteins found in Antarctic notothenioidfishes (DeVries and Wohlschlag, 1969; DeVrieset al, 1970, 1971; DeVries, 1982, 1986; DeVriesand Cheng, 1992). These glycoproteins are foundin eight distinct sizes named antifreeze glyco-protein (AFGP) 1-8 ranging from 2.6 to 34 kDa(DeVries et al, 1971; Duman and DeVries, 1972;Duman et al, 1993). They are composed of aglycotripeptide unit alanyl-alanyl-threonine with adisaccharide, Af-acetylgalactosamine and galac-tose [P-D-galactopyranosyl( 1 —>3)2-acetamido-2-deoxy-2-oc-D-galactopyranose] linked to the threon-ines (DeVries, 1971; DeVries et al, 1971; Dumanet al, 1993). In the small AFGP 7 and 8, thethreonines are sometimes substituted with proline(Lin et al, 1972; Kieran et al, 1980); these alsodiffer from the larger ones by having a somewhatlower antifreeze activity. AFGP identical to thosein the notothenioid fishes are also found in theunrelated species of gadoid fishes from the arctic,e.g. in Boreogadus saida and Gadus ogac (VanVoorhies et al, 1978), recognized as one of the
38
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
most important examples of convergent evolutionin biochemistry (Chen et al, 1997). (ii) The Type-I antifreeze proteins are found in winter flounder,Pseudopleuronectes americanus and other North-ern flatfishes and sculpins (Duman and DeVries,1976; Hew et al, 1985). These are alanine-rich,amphipathic cc-helix peptides with mol. wt of 3300-5000 Da and often contain 11 amino acid repeatingunits (Raymond et al, 1975; Davies and Hew,1990; Duman et al, 1993). (iii) The Type-IIantifreeze proteins are found in the sculpin searaven, Hemitripterus americanus. These do nothave an obvious repeat structure but are cysteinerich globular proteins that have a significant amountof (^-structures and mol. wt of -14 kDa (Slaugtheret al, 1981; Hayes et al, 1989). The Type-Illantifreeze proteins are found in eelpouts and zoar-cid fishes. These peptides have no amino acid biasbut lack histidine and tryptophan. They also haveno obvious repeating sequences and lack helicalstructures but contain compact (3-sheet structures(Davies and Hew, 1990). They have a mol. wt of-7000 Da (Li et al, 1985; Cheng and DeVries,1989). In 1998, Deng and Laursen described anew antifreeze protein (LS-12) from the long-hornsculpin, Myoxocephalus octodecimspinosis (Dengand Laursen, 1998). This antifreeze consists offour amphiphatic a-helices of similar length, foldedinto a four-helix bundle. This antifreeze may beconsidered the first example of a new type (TypeIV) fish antifreeze protein.
Despite the various classes of antifreeze proteinshaving few structures in common they all exhibitinhibition of the growth of ice crystals by adsorp-tion to or at least perturbation of different crystalfaces in the ice crystal (Raymond and DeVries,1977). It is interesting that, unlike most othermolecules, AFGP and antifreeze proteins areincorporated into the ice if a solution of thesemolecules freezes (Cheng and DeVries, 1991).Raymond and DeVries (1977) proposed that theAFGP hydrogen binds to the ice crystals via thehydrogen-bonding groups in the protein and thewater molecules in the ice lattice. This adsorptioninhibits the ice growth. Raymond and DeVries(1977) noted that the growth habit of ice crystalschanged in the presence of antifreeze proteins.When the antifreeze solutions reached the hyster-
esis freezing point the ice crystals grew as long,thin, parallel needles (spicules) whose axes werealigned with the ice c-axis. This is the thermodyn-amically non-preferred direction of growth. Usu-ally ice grows in the direction perpendicular to thec-axis, i.e. parallel to the a-axes (on the prismplane) (the thermodynamically preferred directionof growth) (Cheng and DeVries, 1991). Theobserved growth pattern indicated that the anti-freeze proteins preferentially adsorbed to the crystalfaces parallel to the c-axis (the prism planes) andinhibited growth in the direction perpendicular tothe c-axis (Raymond and DeVries, 1977; Raymondet al, 1989). From these observations it can bededuced that the spicular growth occurs when itbecomes thermodynamically preferable for the icecrystals to grow along the c-axis, i.e. on the basalplane. Later investigations have indicated that othertypes of antifreeze proteins adsorb to or perturbdifferent crystal planes in the ice crystals, e.g.primary prism planes and secondary prism planes(Knight et al, 1991; Ewart et al, 1999).
Antifreeze proteins are not restricted to marineteleost fishes, there are more than 30 knownspecies of terrestrial or tidal invertebrates in whichantifreeze activity has been shown in the bodyfluid (Theede et al, 1976; Duman, 1979; Husbyand Zachariassen, 1980; Block and Duman, 1989;Duman et al, 1991a). The functions of antifreezeproteins in terrestrial invertebrates may be thesame as in fish, namely inhibition of ice growth,which would be relevant in, for example, tidalinvertebrates which are moist and thus likely to beinoculated from the surroundings.
Terrestrial arthropods are often exposed toextremely low temperatures during winter e.g. -50°C(Zachariassen, 1985). The occurrence of antifreezeproteins in freeze-avoiding insects may thereforebe understood in one of two contexts. Usuallyinoculation of insects is not considered of greatimportance as these have a wax-coated hydro-phobic cuticle that is likely to prevent inoculation.However, S0mme (1982) showed that inoculationoccurs more often than usually thought. Thereforeantifreeze proteins may fortify the insects' defencesagainst inoculation (Gehrken, 1992). Another pos-sibility is that antifreeze agents stabilize the meta-stable supercooled state in freeze-avoiding insects.
39
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
Zachariassen and Husby (1982) have shown thatthere is an inverse log-linear relationship betweenthe hysteresis activity and the size of the ice crystalin the solution, that is, the smaller the size of theice crystal the larger the thermal hysteresis. Ifthese findings (Zachariassen and Husby, 1982) areextrapolated below the lower limit of observationof the ice crystal size used in the experiment, itwould appear that the antifreeze proteins may beable to inhibit the growth of clusters of watergiving rise to potential embryo ice crystals. Thesefindings are still under investigation (Kristiansenet al, 1999).
So far the antifreeze agents have primarily beenfound in the extracellular fluid in freeze-avoidinginsects, but antifreeze agents from the intestinalfluid (Olsen and Duman, 1997) and intracellularly(Kristiansen et al., 1999) have been described.
Antifreeze proteins are usually associated withfreeze-avoiding animals, but surprisingly antifreezeproteins have also been found in freeze-tolerantinsects (Duman et al., 1991a). In these animals theantifreeze effect is of no obvious function as theseanimals often promote freezing by the productionof INAs (see above); however, the effect of anti-freeze proteins on inhibiting recrystallization ofice already formed (Knight et al., 1995) may berelevant. Recrystallization is the growth of largeice crystals at the expense of smaller ones. Fora theoretical background on the grain boundarymigration involved in recrystallization see Knightet al. (1995). This change in size distribution ofice crystals over time may cause damage to thetissues either during thawing or during changesin ambient temperature. It has therefore beensuggested that the role of antifreeze proteins infreeze-tolerant animals is recrystallization inhibi-tion to inhibit lethal recrystallization (Knight andDuman, 1986). Recently recrystallization inhibitionwas described in relatively dilute homogenates ofthe freeze-tolerant Antarctic nematode Panagro-laimus davidi (Raml0v et al., 1996). As thisanimal is capable of surviving intracellular freezing(Wharton and Ferns, 1995) it may be of greatimportance that changes in crystal size, with theensuing disruption of cell membranes, are inhibited(Raml0v et al., 1996).
Specific protection by low molecular weightsubstances during cooling and freezing
Most cold-tolerant animals synthesize cryoprotec-tants either as a response to freezing or becauseof environmental cues during autumn. Apart fromcontrolling ice formation and supercooling points,these substances may also interact directly withvarious structures in the organisms.
Glycerol is the most commonly found low mol.wt protective substance in cold-tolerant animals(S0mme, 1982; Zachariassen, 1985; Storey andStorey, 1992); however, both sugars and free aminoacids are common in cold-tolerant animals, especi-ally trehalose and proline (Zachariassen, 1985;Storey, 1997; Raml0v, 1999). These substances areefficient stabilizers of membrane integrity andprotein structure during cooling and dehydrationdue to ice formation (Gekko and Timasheff,1981a,b; Rudolph and Crowe, 1985; Carpenteret al., 1986; Rudolph et al., 1986, Strauss et al,1986). The mechanism of stabilization of mem-branes by trehalose and other sugars is by pre-venting the formation of the gel phase inmembranes. This effect is caused by hydrogenbonding of trehalose to the head group of thephospholipids thereby spreading the monolayers(Rudolph et al., 1986; Strauss et al., 1986). Prolineseems to intercalate between the phospholipid headgroups (Rudolph et al., 1986) but its cryoprotectiveeffect is less well understood.
Summary
In the present article some of the important adapta-tions to low temperature have been reviewed,including changes in membrane and protein struc-ture during cooling and freezing, the two 'strat-egies' employed by cold-tolerant ectothermicanimals — freeze avoidance and freeze toler-ance — the control of ice formation via thecolligative effects of low mol. wt cryoprotectantsand non-colligative effects by INA and antifreezeproteins as well as the non-colligative actions oflow mol. wt cryoprotectants in the stabilization ofmembranes and proteins.
Although we know much about what we thinkare the adaptations to low temperatures in ecto-thermic animals, an animal not alreadycold-adapted cannot be made cold tolerant. Cryo-
40
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
protectants may be injected to induce ice formationvia INA, but not cold tolerance (apart from embryosand dispersed cells in a number of animals). Thisindicates that other adaptations are necessary inconjunction with those already investigated. It hasbeen shown that membranes change their lipidcomposition during cold exposure (Cossins andLee, 1985). It could be speculated that a specialmembrane lipid composition is necessary for theother adaptations e.g. low mol. wt cryoprotectants,to work optimally. Further, it has recently beenshown that antifreeze proteins also bind to biolo-gical membranes and change their properties(Ewart et al., 1999).
There are many subjects within the field ofadaptations to cold in ectothermic animals wherefurther investigations are necessary. It is unclearwhy a single animal species may contain severaldifferent sizes of antifreeze proteins. It could bespeculated that these may work in synergy or thatsome antifreeze proteins 'prepare' the surfaces ofthe ice crystals for the action of others. Themechanism of action of antifreeze proteins is stilla matter of debate, whether by direct bindingto, or perturbation of, the ice surface. Also, thestabilization of the supercooled state by antifreezeproteins is still not elucidated. Few of the structuresof the antifreeze proteins and ice nucleators foundin terrestrial invertebrates have been investigatedand elucidated (Liou et al, 1999).
Both cold-tolerant ectothermic invertebrates andvertebrates synthesize low mol. wt cryoprotectants,but there are large differences both in the types ofcryoprotectants synthesized and the environmentalcues which induce them. There are also largedifferences between the two groups concerning thesupercooling, the amount of ice, the ice formationrate and cooling rates tolerated.
Today the mechanisms controlling the synthesisof the various cryoprotective substances, photo-period, temperature, circadian rhythms and hor-monal control have only been investigated in fewcases leaving much to be learnt.
AcknowledgementsCharlotte Rasmussen is thanked for help in preparingthe manuscript.
ReferencesAngell, C.A. (1982) Supercooled water. In Franks, F.
(ed.), Water; A Comprehensive Treatise. Plenum Press,New York, Vol. 7, p. 1-81.
Aunaas, T. (1982) Nucleating agents in the haemolymphof intertidal invertebrates tolerant to freezing. Cryo-Letters, 3, 287.
Balasingham, E. (1964) Comparative studies on theeffects of temperature on free-living stages ofPlococonus laturis, Dochonoides stenocephala andAncylostoma caninum. Can. J. Zool., 42, 907-918.
Baust, J.G. and Zachariassen, K.E. (1983) Seasonallyactive cell matrix associated ice nucleators in aninsect. Cryo-Letters, 4, 65-71.
Berman, D.I., Leirikh, A.N. and Mikhailova, E.I. (1984)Winter hibernation of the Siberian salamanderHynobius keyserlingi. J. Evol. Biochem. Physiol., 3,323-327.
Block, W. (1977) Oxygen consumption of the terrestrialmite Alaskozetes antarcticus (Acari: Cryptostigmata).J. Exp. Biol, 68, 69-87.
Block, W. (1982) Cold hardiness in invertebratepoikilotherms. Comp. Biochem. Physiol., 73A, 581—593.
Block, W. (1990) Cold tolerance of insects and otherarthropods. Phil. Trans. R. Soc. Lond., B326, 613-633.
Block, W. and S0mme, L. (1982) Cold-hardiness ofterrestrial mites at Signy Island, Maritime Antarctic.Oikos, 38, 157-167.
Block, W. and Duman, J.G. (1989) Presence of thermalhysteresis producing antifreeze proteins in theAntarctic mite, Alaskozetes antarcticus. J. Exp. Biol.,250, 229-231.
Block, W., Davenport, J. and Bale, J. (1992) Animalsat low temperature. J. Zool., Lond., 226, 345-350.
Block, W., Worland, R.M. and Bale, J. (1998) Respiratoryresponses to chilling and freezing in two sub-antarcticinsects. Cryobiology, 37, 163-166.
Bloom, M. (1998) Evolution of membranes from aphysics perspective. Biol. Skr. Da. Vid. Selsk., 49,13-17.
Burke, M.J. and Lindow, S.E. (1990) Surface propertiesand size of the ice nucleation site in ice nucleationactive bacteria. Cryobiology, 27, 80-84.
Cannon, R.J.C. and Block, W (1988) Cold tolerance inmicroarthropods. Biol. Rev., 63, 23-77.
Carpenter, J.F., Hand, S.C., Crowe, L.M. and Crowe,J.H. (1986) Cryoprotection of phosphofructokinasewith organic solutes: Characterization of enhancedprotection in the presence of divalent cations. Arch.Biochem. Biophys., 250, 505-512.
Chapman, D., Cornell, B.A. and Quinn, P.J. (1977)Phase transitions, protein aggregation and a newmethod for modulating membrane fluidity. InSemenza, G. and Carafoli, E. (eds), Biochemistry andMembrane Transport. Springer-Verlag, New York, pp.72-85.
Chen, L., DeVries, A.L. and Cheng, C.-H.C. (1997)Convergent evolution of antifreeze glycoproteins inAntarctic notothenioid fish arctic cod. Proc. Natl.Acad. Sci. USA, 94, 3817-3822.
41
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
Cheng, C.C. and DeVries, A.L. (1989) Structures ofantifreeze peptides from the antarctic eel poutAustrolycichthys brachycephalus. Biochim. Biophys.Acta, 997, 55-64.
Cheng, C.C. and DeVries, A.L. (1991) The role ofantifreeze glycopeptides and peptides in the freezingavoidance of cold water fish. In di Prisco, G. (ed.), LifeUnder Extreme Conditions. Springer-Verlag Berlin.
Churchill, T.A. and Storey, K.B. (1992) Natural freezingsurvival by painted turtles Chrysemys picta marginataand C. picta bellii. Am. J. Physiol., 262, R530-R537.
Churchill, T.A. and Storey, K.B. (1996) Organmetabolism and cryprotectant synthesis duringfreezing in spring peepers Pseudacris crucifer. Copeia,3, 517-525.
Costanzo, J.P. and Lee, R.E. Jr (1995) Supercooling andice nucleation in vertebrate ectotherms. In Lee, R.E.Jr, Warren, G.J. and Gusta, L.V. (eds), Biological IceNucleation and its Applications. APS Press, St Paul,Minnesota, pp. 221-237.
Crowe, J.H. and Crowe, L.M. (1985) Effects ofdehydration on membranes and membranestabilization at low water activities. In Chapman, ?(ed.), Biological Membranes, Vol. 5. Academic Press,New York.
Crowe, J.H., Crowe, L.M. and Jackson, S. (1983)Preservation of structure and functional activity inlyophilised sarcoplasmatic reticulum. Arch. Biochem.Biophys., 220, 477^84.
Cossins, A.R. and Lee, J.A.C. (1985) The adaptation ofmembrane structure and lipid composition to cold.In Gilles, R. (ed.), Circulation, Respiration, andMetabolism. Springer-Verlag, Berlin, pp. 543-552.
Costanzo, J.P. and Lee, R.E. Jr (1996) Mini-review: Icenucleation in freeze-tolerant vertebrates. Cryo-Letters,17, 111-118.
Davenport, J. (1992) Animal Life at Low Temperature.Chapman and Hall, London.
Davies, P.L. and Hew, C.L. (1990) Biochemistry of fishantifreeze proteins. FASEB J., 4, 2460-2468.
Deng, G. and Laursen, R.A. (1998) Isolation andcharacterization of an antifreeze protein from thelong horn sculpin, Myoxocephalus octodecicspinosis.Biochim. Biophys. Ada, 1388, 305-314.
Denstad, J.P., Aunaas, T., Borseth, J.F. et al. (1987)Thermal hysteresis antifreeze agents in fishes fromSpitsbergen waters. Polar Res., 5, 171-174.
DeVries, A.L. (1971) Glycoproteins as biologicalantifreeze agents in Antarctic fishes. Science, 172,1152-1155.
DeVries, A.L. (1982) Biological antifreeze agents incold water fishes. Comp. Biochem. Physiol., 73A,627-640.
DeVries, A.L. (1986) Antifreeze glycopeptides andpeptides; interactions with ice and water. Meth.Enzymol., 127, 293-303.
DeVries, A.L. and Cheng, C.-H.C. (1992) The role of
antifreeze glycopeptides and peptides in the survivalof cold-water fishes. In Somero, G.N., Osmond, C.B.and Bolis, C.L. (eds), Water and Life: ComparativeAnalysis of Water Relationships at the Organismic,Cellular, and Molecular Levels. Springer-Verlag,Berlin, pp. 301-315.
DeVries, A.L. and Wohlschlag, D.E. (1969) Freezingresistance in some antarctic fishes. Science, 163,1073-1075.
DeVries, A.L., Vandenheede, J. and Feeney, R.E. (1971)Primary structure of freezing point depressingglycoproteins. J. Biol. Chem., 246, 305-308.
DeVries, A.L., Komatsu, S.K. and Feeney, R.E. (1970)Chemical and physical properties of freezing point-depressing glycoproteins from Antarctic fishes. J.Biol. Chem., 245, 2901-2908.
Duman, J.G. (1979) Subzero temperature tolerance inspiders: the role of thermal hysteresis factors. J. Comp.Physiol., 131B, 347-352.
Duman, J.G. (1982) Insect antifreezes and ice nucleatingagents. Cryobiology, 19, 613-627.
Duman, J.G. and DeVries, A.L. (1972) Freezing behaviorof aqueous solutions of glycoproteins from the bloodof an Antarctic fish. Cryobiology, 9, 469-472.
Duman, J.G. and DeVries, A.L. (1976) Isolation,characterization and physical properties of proteinantifreeze from the winter flounder, Pseudopluronectesamericanus. Comp. Biochem. Physiol., 53B, 375-380.
Duman, J.G. and Horwath, K. (1983) The role ofhaemolymph proteins in the cold tolerance of insects.Annu. Rev. Physiol, 43, 261-270.
Duman, J.G., Morris, J.P. and Castellino, F.J. (1984)Purification and composition of an ice nucleationprotein from queens of the hornet, Vespula maculata.J. Comp. Physiol., 154B, 79-83.
Duman, J.G., Xui, L., Neven, L.G. et al. (1991a)Hemolymph proteins involved in insect subzero-temperature tolerance: ice nucleators and antifreezeproteins. In Lee, R.E., Jr and Denlinger, D.L. (eds),Insects at Low Temperature. Chapman and Hall, NewYork, pp. 94-127.
Duman, J.G., Wu, D.W., Wolber, P.I. et al. (1991b)Further characterization of the lipoprotein icenucleator from freeze tolerant larvae of the cranefly Tipula trevitata. Comp. Biochem. Physiol., 99B,599-607.
Duman, J.G., Wu, D.W., Yeung, K.L. and Wolf, E.E.(1992) Hemolymph proteins involved in the coldtolerance of terrestrial arthropods: Antifreeze and icenuceator proteins. In Somero, G.N., Osmond, C.B.and Bolis, C.L. (eds), Water and Life: ComparativeAnalysis of Water Relationships at the Organismic,Cellular, and Molecular Levels. Springer-Verlag,Berlin, pp. 282-300.
Duman, J.G., Wu, D.W., Olsen, T.M. et al. (1993)Thermal-hysteresis proteins. In Steponkus, P.L. (ed.),
42
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
Advances in Low Temperature Biology. JAI Press,London, Vol. 2, pp. 131-182.
Duman, J.G., Olsen, T.M., Yeung, K.L. and Jerva, F.(1995) The roles of ice nucleators in cold tolerantinvertebrates. In Lee, R.E. Jr, Warren, G.J. and Gusta,L.V. (eds), Biological Ice Nucleation and itsApplications. APS Press, St Paul, Minnesota, pp.201-219.
Ewart, K.V., Lin, Q. and Hew, C.L. (1999) Structure,function and evolution of antifreeze proteins. CellMol. Life ScL, 55, 271-283.
Feeney, R.E. (1974) A biological antifreeze. Amer.Scient., 62, 712-719.
Franks, F. (1985) Biophysics and Biochemistry at LowTemperatures. Cambridge University Press,Cambridge.
Franks, F. and Hadley, R.H.M. (1992) Protein stabilityunder conditions of deep chill. In Steponkus, P.L.(ed.), Advances in Low Temperature Biology. JAIPress, London, Vol. 1, pp. 141-179.
Franks, F, Mathias, S.F. and Hadley, R.H.M. (1990)Water, temperature and life. Phil. Trans. R. Soc. Lond.,B326, 517-533.
Gehrken, U. (1984) Winter survival of an adult barkbeetle Ips acuminatus Gyll. J. Insect Physiol, 30,421^29.
Gehrken, U. (1992) Inoculative freezing and thermalhysteresis in the adult beetles Ips acuminatus andRhagium inquisitor. J. Insect. Physiol., 38, 519-524.
Gekko, K. and Timasheff, S.N. (1981a) Mechanism ofprotein stabilization by glycerol: preferential hydrationin glycerol-water mixtures. Biochemistry, 20, 4667-4676.
Gekko, K. and Timasheff, S.N. (1981b) Thermodynamicand kinetic examination of protein stabilization byglycerol. Biochemistry, 20, 4677^686.
Gennis, R.B. (1989) Biomembranes: Molecular Structureand Function. Springer-Verlag, New York.
Goddard, S.V., Kao, M.H. and Fletcher, G. L. (1992)Antifreeze production, freeze resistance, andoverwintering of juvenile northern atlantic cod (Gadusmorhua). Can. J. Fish. Aquat. Sci., 49, 516-522.
Govindarajan, A.G. and Lindow, S.E. (1988) Size ofbacterial ice-nucleation sites measured in situ byradiation and inactivation analysis. Proc. Natl. Acad.Sci. USA, 85, 1334-1338.
Grout, B.W.W. and Morris, G.J. (1987) The Effects ofLow Temperature on Biological Systems. EdwardArnold, London.
Hansen, T. and Merivee, E. (1971) Cold hardiness ofthe cabbage butterflies Pieris brassicae L. and Pierisrapae L. Esti NSV Tead. Akad. Toim. Biol., 20,298-303.
Hayes, D.R. and Loomis, S.H. (1985) Evidence for aproteinaceous ice nucleator in the haemolymph of thepulmonate gastropod, Melampus bidentatus. Cryo-Letters, 6, 418-421.
Hayes, PH., Scott, G.K., Ng, N.F.L. et al. (1989)Cysteine rich type II antifreeze protein precursor isinitiated from the third AUG codon of its mRNA.J. Biol. Chem., 264, 18761-18767.
Hew, C.L. and Yang, D.S.C. (1992) Protein interactionwith ice. Eur. J. Biochem., 203, 33—42.
Hew, C.L., Joshi, S., Wang, N.C. et al. (1985) Structuresof shorthorn sculpin antifreeze polypeptides. Eur. J.Biochem., 151, 167-172.
Hochachka, P.W. and Somero, G.N. (1984) BiochemicalAdaptation. Princeton University Press, NJ.
Hofstee, B.F. (1949) The activation of urease. / . Gen.Physiol., 32, 339-347.
Husby, J.A. and Zachariassen, K.E. (1980) Antifreezeagents in the body fluid of winter active insects andspiders. Experientia, 36, 963-964.
Hvidt, Aa. and Westh, P. (1998) Different views on thestability of protein conformations and hydrophobiceffects. / . Sol. Chem., 27, 395-402.
Jensen, M. (1999) Frysetolerance hos Salamandrellakeyserlingi. Masters thesis, Roskilde University, inDanish with a summary in English.
Kieran, G.F., Osuga, D.T., Ahmed, A.I. et al. (1980)Antifreeze glycoproteins from polar fish. J. Biol.Chem., 255, 663-667.
Kirchner, W. and Kestler, P. (1969) Untersuchungen zurkalteresistenz der schilfradspinne Araneus cornutus(Araneidae). J. Insect Physiol., 15, 41-53.
Klok, C.J. and Chown, S.L. (1997) Critical thermallimits, temperature tolernce and water balance ofa sub-Antarctic caterpillar, Pringleophaga marioni(Lepidoptera: Tineidae). J. Insect Physiol., 43, 685-694.
Knight, C.A. and Duman, J.G. (1986) Inhibition ofrecrystallization of ice by insect thermal hysteresisproteins: a possible cryoprotective role. Cryobiology,23, 256-262.
Knight, C.A., Cheng, C.C. and DeVries, A.L. (1991)Adsorbtion of oc-helical antifreeze peptides on specificice crystal surface planes. Biophys. J., 59, 409^418.
Knight, C.A., Wen, D. and Lauersen, R.A. (1995)Nonequilibrium antifreeze peptides and therecrystallization of ice. Cryobiology, 32, 23-34.
Kristiansen, E., Pedersen, S., Raml0v, H. andZachariassen, K.E. (1999) Antifreeze activity in thecerambycid beetle Rhagium inquisitor. J. Comp.Physiol, B169, 55-60.
Laak, S. van der (1982) Physiological adaptations to lowtemperature in freezing-tolerant Phyllodecta laticollisbeetle. Comp. Biochem. Physiol, 73A, 613-620.
Layne, J.R. and Lee, R.E. Jr (1987) Freeze toleranceand the dynamics of ice formation in wood frogs(Rana sylvaticd) from southern Ohio. Can. J. Zool.,65, 2062-2065.
Layne, J.R., Lee, R.E. and Huang, J.L. (1990) Inoculationtriggers freezing at high subzero temperatures in a
43
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
freeze tolerant frog (Rana sylvtica) and an insect{Eurosta solidaginis). Can. J. Zool., 68, 506-510.
Lee, R.E. Jr (1991) Principles of insect low temperaturetolerance. In Lee, R.E. Jr and Denlinger, D.L. (eds),Insects at Low Temperature. Chapman and Hall, NewYork, pp. 17-46.
Lee, R.E. Jr and Lewis, E.A. (1985) Effect of temperatureand duration of exposure on tissue ice formation inthe gall fly Eurosta solidaginis (Diptera, Tephritidae).Cryo-Letters, 6, 25-34.
Lefevere, K.S., Koopmanschap, A.B. and Kort, C.A.D.(1989) Changes in the concentrations of metabolitesin haemolymph during and after diapause in femaleColorado potato beetle Leptinotarsa decemlineata.J. Insect Physiol., 35, 121-128.
Li, X.M., Trinh, K.Y., Hew, C.L. et al. (1985) Structureof an antifreeze polypeptide and its precursor fromthe ocean pout, Macrozoarches americanus. J. Biol.Chem., 260, 12904-12909.
Lin, Y., Duman, J.G. and DeVries, A.L. (1972) Studieson the structure and activity of low molecular weightglycoproteins from an Antarctic fish. Biochim.Biophys. Res. Commun., 46, 87-92.
Liou, Y.-C, Thibault, P., Walker, V.K. et al. (1999) Acomplex family of highly heterogenous and internallyrepetitive hyperactive antifreeze proteins from thebeetle Tenebrio molitor. Biochemistry, 38, 11415-11424.
Loomis, S.H. (1985) Seasonal changes in freezingtolerance of the intertidal pulmonate gastropodMelampus bidentatus, Say. Can. J. Zool., 63, 2021-2025.
Lundheim, R. and Zachariassen, K.E. (1993) Waterbalance of overwintering beetles in relation tostrategies for cold tolerance. J. Comp. Physiol., B163,1-4.
MacKenzie, A.P. (1977) Non-equilibrium freezingbehaviour of aqueous systems. Phil. Trans. R. Soc.Lond., 287B, 167-189.
Mansingh, A. and Smallman, B.N. (1972) Variation inpolyhydric alcohol in relation to diapause and cold-hardiness in larvae of Isia Isabella. J. Insect Physiol.,18, 1565-1571.
Mazur, P. (1984) Freezing of living cells: mechanismsand implications. Am. J. Physiol., 247 {Cell Physiol.,16), C125-C142.
Mazur, P., Leibo, S.P. and Chu, E.H.Y (1972) A two-factor hypothesis of freezing injury. Evidence fromChinese hamster tissue culture cells. Exp. Cell Res.,71, 345-355.
Merivee, E. (1978) Cold-hardiness in Insects. Eesti NSVTead. Akad. Zool, 1082.
McGrath, J.J. (1987) Cold shock: thermoelastic stressin chilled biological membranes. In Diller, K.R. (ed.),Network Thermodynamics, Heat and Mass Transferin Biotechnology. ASME Bed, Vol. 5, HTD, Vol. 90,pp. 57-66.
Miller, L.K. (1982) Cold hardiness strategies of someadult and immature insects overwintering in interiorAlaska. Comp. Biochem. Physiol, 73A, 595-604.
Moore, W.J. (1981) Physical Chemistry. Longman,Essex.
Morris, G.J. and Clarke, A. (1987) Cells at lowtemperatures. In Grout, B.W.W. and Morris, G.J. (eds),The Effects of Low Temperature on Biological Systems.Edward Arnold, London, pp. 72-119.
Mueller, G.M., Wolber, P.K. and Warren, G.J. (1990)Clustering of ice nucleation protein correlates withice nucleation activity. Cryobiology, 27, 416^-22.
Mugnano, J.A., Lee, R.E. Jr and Taylor, R.T. (1996) Fatbody cells and calcium phosphate spherules induceice nucleation in the freeze-tolerant larvae of the gallfly Eurosta solidaginis (Diptera, Tephritidae). J. Exp.Biol, 199, 465-471.
Neufeld, D.S. and Leader, J.P. (1998) Freezing survivalby isolated malpighian tubules of the New Zealandalpine weta Hemideina maori. J. Exp. Biol, 201,227-236.
Neven, L.G., Duman, J.G., Low, M.G. et al (1989)Purification and characterization of an insecthaemolymph lipoprotein ice nucleator: Evidence forthe importance of phosphatidylinositol andapolipoprotein in the ice nucleator activity. J. Comp.Biochem. Physiol, 159, 71-82.
Olsen, T.M. and Duman, J.G. (1997) Maintenance ofthe supercooled state in overwintering pyrochroidbeetle larvae, Dendroides Canadensis: role of icenucleators and antifreeze proteins. J. Comp. Physiol,B167, 105-113.
Quinn, P.J. (1985) A lipid-phase separation model oflow temperature damage to biological membranes.Cryobiology, 22, 128-146.
Quinn, P.J. and Williams, W.P. (1985) Environmentallyinduced changes in chloroplast membranes and theireffect on photosynthesis function. In Barber, J. andBaker, N.P. (eds), Photosynthetic Mechanisms and theEnvironment. Elsevier Amsterdam, pp. 1^47.
Raml0v, H. (1999) Microclimate and variations inhaemolymph composition in the freezing-tolerantalpine weta Hemideina maori Hutton (Orthoptera:Stenopelmatidae). J. Comp. Physiol, B169, 224-235.
Raml0v, H. and Westh, P. (1992) Survival of theCryptobiotic Eutardigrade Adorybiotus coroniferduring cooling to -196 C: effect of cooling rate,trehalose level and short-term acclimation.Cryobiology, 29, 125-130.
Raml0v, H. and Westh, P. (1993) Ice formation in thefreeze tolerant alpine weta Hemideina maori Hutton(Orthoptera; Stenopelmatidae). Cryo-Letters, 14,169-176.
Raml0v, H., Bedford, J. and Leader, J.P. (1992) Freezingtolerance of the New Zealand alpine weta Hemideinamaori hutton (Orthoptera: Stenopelmatidae). J. Therm.Biol, 17, 51-54.
44
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

Cold tolerance in ectotherms
Raml0v, H., Wharton, D.A. and Wilson, P.W. (1996)Recrystallization in a freezing tolerant Antarcticnematode. Panagrolaimus davidi, and an alpine weta,Hemideina maori (Orthoptera; Stenopelmatidae).Cryobiology, 33, 607-613.
Ramsay, J.A. (1964) The rectal complex of the mealwormTenebrio molitor, L. (Coleoptera, Tenebrionidae). Phil.R. Soc. Lond., 248B, 748, 35-314.
Rasmussen, D.H. and MacKenzie, A.R (1973) Clusteringin supercooled water. J. Chem. Phys., 59, 5003-5013.
Raymond, J.A. and DeVries, A.L. (1977) Adsorbtioninhibition as a mechanism of freezing resistancein polar fishes. Proc. Natl. Acad. Sci. USA, 74,2589-2593.
Raymond, J.A., Lin, Y. and DeVries, A.L. (1975)Glycoproteins and protein antifreeze in two Alaskanfishes. J. Exp. Zooi, 193, 125-130.
Raymond, J. A., Wilson, P. and DeVries, A.L. (1989)Inhibition of nonbasal planes in ice by fish antifreezes.Proc. Natl. Acad. Sci. USA, 86, 881-885.
Ring, R.A. (1980) Insects and their cells. In Ashwood-Smith, M.J. and Farrant, J. (eds), Low TemperaturePreservation in Medicine and Biology. Pitman,London, pp. 187-217.
Ring, R.A. (1981) The physiology and biochemistry ofcold tolerance in arctic insects. J. Therm. Biol., 6,219-229.
Ring, R.A. (1982) Freezing tolerant insects with lowsupercooling points. Comp. Biochem. Physiol., 73A,605-612.
Ring, R.A. and Danks, H.V. (1998) Desiccation andcryoprotection: overlapping adaptations. Cryo-Letters,15, 181-190.
Rudolph, A.S. and Crowe, J.H. (1985) Membranestabilization during freezing: the role of two naturalcryoprotectants, trehalose and proline. Cryobiology,22, 367-377.
Rudolph, A.S., Crowe, J.H. and Crowe, L.M. (1986)Effects of three stabilizing agents I proline, betaine,and trehalose on membrane phospholipids. Arch.Biochem. Biophys., 245, 134-143.
Salt, R.W. (1953) The influence of food on cold hardinessof insects. Can. Entomol., 85, 261-269.
Salt, R.W. (1959a) Survival of frozen fat body cells inan insect. Nature, 184, 1426.
Salt, R.W. (1959b) Role of glycerol in the cold-hardeningof Bracon cephi. Can. J. Zool., 37, 59-69.
Salt, R.W. (1962) Intracellular freezing in insects. Nature,193, 1207-1208.
Salt, R.W. (1963) Delayed inoculative freezing in insects.Can. Entomol., 95, 1190-1202.
Schmid, W.D. (1982) Survival of frogs in lowtemperature. Science, 215, 697-698.
Scholander, P.F., van Dam, L., Kanwisher, J.W. et al.(1957) Supercooling and osmoregulation in arctic fish.J. Cell Comp. Physiol., 49, 5-24.
Shimada, K. and Riihimaa, A. (1988) Cold acclimation,inoculative freezing and slow cooling: essential factorscontributing to the freeze-tolerance in diapausinglarvae of Chymomyza costata (Diptera:Drosophilidae). Cryo-letters, 9, 5-10.
Sinclair, B. (1997) Seasonal variation in freezingtolerance of the New Zealand alpine cockroachCelattoblatta quinquemaculata. Ecol. Entomol., 22,462-467.
Sinclair, B. and Wharton, D.A. (1997) Avoidance ofintracellular freezing by the freezing-tolerant NewZealand alpine weta Hemideina maori (Orthoptera:Stenopelmatidae). J. Insect Physiol., 43, 621-625.
Slaughter, D., Fletcher, G.L., Ananthanrayanan, VS. andHew, L.L. (1981) Antifreeze proteins from the searaven, Hemitripterus americanus: further evidence fordiversity among fish polypeptide antifreezes. J. Biol.Chem., 256, 2022-2026.
Storey, K.B. (1983) Metabolism and bound water inoverwintering insects. Cryobiology, 20, 265-279.
Storey, K.B. (1984) Freeze tolerance in the frog Ranasylvatica. Experientia, 40, 1261-1262.
Storey, K.B. (1997) Organic solutes in freezing tolerance.Comp. Biochem. Physiol, 117A, 319-326.
Storey, K.B. and Storey, J.M. (1985) Freeze tolerancein the grey tree frog Hyla versicolor. Can. J. Zool.,63, 49-54.
Storey, K.B. and Storey, J.M. (1992) Biochemicaladaptations for winter survival in insects. InSteponkus, PL. (ed.), Advances in Low TemperatureBiology. JAI Press, London, Vol. 1, pp. 101-140.
Storey, K.B. and Storey, J.M. (1993) Cellular adaptationsfor freezing survival by amphibians and reptiles. InSteponkus, PL. (ed.), Advances in Low TemperatureBiology. JAI Press, London, Vol. 2, pp. 101-129.
Storey, K.B., Baust, J.G. and Buecher, P. (1981)Determination of water 'bound' by soluble subcellularcomponents during low-temperature acclimation inthe gall fly larva Eurosta solidaginis. Cryobiology,18, 315-321.
Storey, K.B., Storey, J.M., Brooks, S.P.J. et al. (1988)Hatchling turtles survive freezing during winterhibernation. Proc. Natl. Acad. Sci. USA, 85, 8350-8354.
Strauss, G., Schurtenberger, P. and Hauser, H. (1986) Theinteraction of saccharides with lipid bilayer vesicles:stabilization during freeze-thawing and freeze-drying.Biocim. Biophys. Acta, 858, 169-180.
S0mme, L. (1982) Supercooling and winter survival interrestrial arthropods. Comp. Biochem. Physiol., 73A,519-543.
S0mme, L. (1986) Tolerance to low temperatures anddesiccation in insects from Andean paramos. Arct.Alp. Res., 18, 253-257.
S0mme, L. and Conradi-Larsen, E.-M. (1977) Cold-hardiness of collembolans and oribatid mites fromwindswept mountain ridges. Oikos, 29, 118-126.
45
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018

H.Raml0v
S0mme, L. and Block, W. (1982) Cold-hardiness ofcollembola at Signy Island, maritime Antarctic. Oikos,38, 168-176.
Tanno, K. (1964) High sugar levels in the solitary beeCeratina. Low Temp. Sci., 22B, 51-57.
Taylor, M.J. (1987) Physico-chemical principles in lowtemperature biology. In Grout, B.W.W. and Morris,G.J. (eds), The Effects of Low Temperatures onBiological Systems. Edward Arnold, London, pp.3-71.
Theede, H., Schneppenheim, R. and Beress, L. (1976)Frostschutz-glykoproteine bei Mytilus edulisl Mar.Biol., 36, 183-189.
Timasheff, S.N. (1978) Thermodynamic examination ofthe self-association of brain tubulin to microtubulesand other structures, In Catsimpoolas, N. (ed.),Physical Aspects of Protein Interactions. Elsevier/North-Holland, New York, pp. 219-273.
Tsumuki, H. and Konno, H. (1991) Tissue distributionof the ice-nucleating agents in the larvae of the ricestem borer, Chilo supressalis Walker (Lepidoptera:Pyralidae). Cryobiology, 28, 376-381.
Tyrell, C , Wharton, D., Raml0v, H. and Moller, H(1994) Cold tolerance of an endoparasitic nematodewithin a freezing-tolerant host. Parasitology, 109,367-372.
Vali, G. (1995) Principles of ice nucleation. In Lee,R.E., Warren, G.J. and Gusta, L.V. (eds), BiologicalIce Nucleation and its Applications. APS Press, StPaul, Minnesota, pp. 1-28.
Van Oss, C.J. and Good, R.J. (1996) Hydrogen bonding,interfacial tension and the aqueous solubility oforganic compounds. J. Dispers. Sci. Technol., 17,433-449.
Van Voorhies, W.V., Raymond, J.A. and DeVries, A.L.(1978) Glycoproteins as biological antifreeze agentsin the cod Gadus ogac (Richardson). Physiol. Zool.,51, 347-353.
Vannier, G. (1987) Mesure de la thermotorpeur chez lesinsects. Bull. Soc. Ecophysiol., 12, 165-186.
Warner, D.T. (1962) Some Possible relationships ofcarbohydrates and other biological components withthe water structure at 37°C. Nature, 196, 1055-1058.
Wasylyk, J.M., Tice, A.R. and Baust, J.G. (1988) Partialglass formation: a novel mechanism of insectcryoprotection. Cryobiology, 25, 451^58.
Westh, P. and Raml0v, H. (1991) Trehalose accumulationin the tardigrade Adorybiotus coronifer duringanhydrobiosis. / . Exp. Zool., 258, 303-311.
Westh, P., Kristiansen, J. and Hvidt, A. (1991) Icenucleating activity in the freeze tolerant tardigradeAdorybiotus coronifer. Comp. Biochem. Physiol., 99A,401^04.
Westh, P., Raml0v, H., Wilson, P.W. and DeVries,A.L. (1997) Vapour pressure of aqueous antifreezeglycopeptide solutions. Cryo-Letters, 18, 277-282.
Wharton, D.A. and Ferns, D.J. (1995) Survival of
intracellular freezing by the Antarctic nematodePanagrolaimus davidi. J. Exp. Biol., 198, 1-7.
Williams, R. J. and Carnahan, D.L. (1990) Fracturefaces and other interfaces as ice nucleation sites.Cryobiology, 27, 479-482.
Williams, W.P. (1990) Cold induced lipid phasetransitions. Phil. Trans. R. Soc. Lond., 326B, 555-570.
Wilson, P. (1994) The physical basis of action ofbiological ice nucleation agents. Cryo-Letters, 15,119-126.
Wilson, P. and Raml0v, H. (1995) Hemolymph icenucleating proteins from the New Zealand alpineweta Hemideina maori (Orthoptera: Stenopelmatidae).Comp. Biochem. Physiol, B112, 535-542.
Worland, R.M., Sinclair, B.J. and Wharton, D.A. (1997)Ice nucleator activity in a New Zealand alpinecockroach Celatoblatta quinquemaculata(Dictyoptera: Blattidae). Cryo-Letters, 18, 327-334.
Wright, J., Westh, P. and Raml0v, H. (1992) Cryptobiosisin Tardigrada. Biol. Rev., 67, 1-29.
Yeung, K.L., Wolf, E.E. and Duman, J.G. (1991) Ascanning tunnelling microscopy study of an insectlipoprotein ice nucleator. J. Vac. Sci. Technol., B9,1197-1201.
Young, S.R. and Block, W. (1980) Experimental studieson the cold tolerance of Alaskozetes antarcticus.J. Insect Physiol, 26, 189-200.
Zachariassen, K.E. (1973) Seasonal variation inhaemolymph osmolality and osmotic contribution ofglycerol in adult Rhagium inquisitor L. (Col.Cerambycidae). Norweg. J. EntomoL, 20, 259-262.
Zachariassen, K.E. (1977) Effects of glycerol in freezetolerant Pytho depressus L. (Col. Pythidae). Norweg.J. EntomoL, 24, 25-29.
Zachariassen, K.E. (1979) The mechanism of thecryoprotective effect of glycerol in beetles tolerant tofreezing. J. Insect. Physiol, 25, 29-32.
Zachariassen, K.E. (1980) The role of polyols andnucleating agents in cold hardy beetles. J. Comp.Physiol, 140, 227-234.
Zachariassen, K.E. (1982) Nucleating agents in coldhardy insects. Comp. Biochem. Physiol, 73A, 557-562.
Zachariassen, K.E. (1985) Physiology of cold tolerancein insects. Physiol. Rev., 65, 799-837.
Zachariassen, K.E. (1992) Ice nucleating agents in coldhardy insects. In Somero, G.N., Osmond, C.B. andBolis, C.L. (eds), Water and Life: ComparativeAnalysis of Water Relationships at the Organismic,Cellular, and Molecular Level. Springer-Verlag,Berlin, pp. 261-281.
Zachariassen, K.E. and Hammel, H.T. (1976) Nucleatingagents in the haemolymph of insects tolerant tofreezing. Nature, 262, 285-287.
Zachariassen, K.E. and Husby, J.A. (1982) Antifreezeeffect of thermal hysteresis agents protects highlysupercooled insects. Nature, 298, 865-867.
46
Downloaded from https://academic.oup.com/humrep/article-abstract/15/suppl_5/26/554993by gueston 08 April 2018