ascorbic acid glycation of lens proteins produces uva sensitizers similar to those in human lens
TRANSCRIPT

Pho/rrhunii.strs r i d f‘holuhrolog\, Vol. 62, No. 3. pp. 454462, 1995 Printed in the United States. All rights reserved
003 I -X655/95 $05 00+0.00 C 1995 American Society for Photohiology
ASCORBIC ACID GLYCATION OF LENS PROTEINS PRODUCES UVA SENSITIZERS SIMILAR TO THOSE IN HUMAN LENS
B. J. ORTWERTH*, MIKHAII. LINETSKY and PAUL R. OLESEN Mason Institute of Ophthalmology. University of Missouri, Columbia, MO 652 12, USA
(Recriijed 9 Fehritnry 1995; czc-repred 29 April 1995)
Abstract-Soluble calf lens proteins were extensively glycated during a 4 week incubation with ascorbic acid in the presence of oxygen. Amino acid analysis of the dialyzed proteins removed at weekly intervals showed an increasing loss of lysine, arginine and histidine, consistent with the extensive protein cross- linking observed. Irradiation of the dialyzed samples with UVA light (1.0 kJ/cm’ total illumination through a 338 nm cutoff filter) caused an increasing loss of tryptophan, an additional loss of histidine and the production of micromolar concentrations of hydrogen peroxide. No alteration in amino acid content and no photolytic effects were seen in proteins incubated without ascorbic acid or in proteins incubated with glucose for 4 weeks. The rate of hydrogen peroxide formation was linear with each glycated sample with a maximum production of 25 nmol/mg protein illuminated. The possibility that the sensitizer activity was due to an ascorbate-induced oxidation of tryptophan was eliminated by the presence of a heavy metal ion chelator during the incubation and by showing equivalent effects with ascorbate-incubated ribonuclease A, which is devoid of tryptophan. The ascorbate-incubated samples displayed increasing absorbance at wavelengths above 300 nm and increasing fluorescence (340/430) as glycation proceeded. The spectra of the 4 week glycated proteins were identical to those obtained with a solubilized water-insoluble fraction from human lens, which is known to have UVA sensitizer activity .
The incubation of lens proteins with dehydroascorbic acid or L-threose, but not fructose, produced equivalent glycation, protein crosslinking and sensitizer activity. The relative sensitizer activity of the 4 week glycated sample was quantitatively very similar to that of a water-insoluble fraction from aged human lenses. These data are consistent with the hypothesis that the protein-bound brunescence in the lens may be advanced glycation endproducts, which are formed in large part by the oxidation products of ascorbic acid, and that these compounds may contribute significantly to the UVA sensitizer activity present in aged human lenses.
INTRODUCTION
Ultraviolet light has been identified as a risk factor for cat- aract formation in several epidemiological studies.’i The most effective wavelengths for cataract formation are in the UVB portion of the spectrum,s-K and recent work has cor- related the distribution of cortical cataracts in the lens with the quadrant of the lens receiving the greatest UV light ex- posure.F,” The spectrum of UV light impinging on the lens in v i v o , however, is almost exclusively UVA light,’” all of which is absorbed by the lens.” It has been postulated that this UV absorbance acts to protect the retina from photo- damage./2 In younger lenses the soluble chromophore, hy- droxylkynurenine glycoside, is an effective UVA filter and has little or no sensitizer activity.’-’.’4 In older lenses. and especially in brunescent lenses, however, there is an accu- mulation of yellow and then brown pigments, which are largely protein bound’-5 and which have UVA sensitizer ac- tivity.’“ Illumination of a sonicated lens water-insoluble frac- tion (W1SS)t’ from aged human lens leads to the photolysis
“To whom correspondence should be addressed. -1-Abhre~icrlions. ASA. ascorbic acid; BCA, bicinchoninic acid;
DTPA, diethylenetriaminepentaacetic acid; PAGE, polyacryl- amide gel electrophoresis; SDS, sodium dodecyl sulfate; WISS, a
of histidine, cysteine, methionine and tryptophan residues. The spectra of light causing these photolytic events exhibited sensitizer activity throughout the UVA region.”
Many laboratories have investigated the role of glycation in the formation of browning pigments, fluorophores and protein crosslinks in lens While many sugars have been suggested as being responsible for glycation in vivo, ascorbic acid (ASA) is present in highest concentration in the lens and is the most reactive agent in v i t r ~ . ~ ~ - ’ ~ Gly- cation does not occur with ASA directly, but through its oxidation products. This oxidation in normal tissues is pre- vented by the high concentrations of glutathione present in the lens.25 This protection, however, diminishes in older hu- man lenses and may result in the formation of protein-bound glycation chromophores. This investigation was undertaken to determine if ascorbate-glycated lens proteins have sensi- tizer activity and to what extent they resemble the sensitizers present in the water-insoluble fraction from aged human lens.
MATERIALS AND METHODS
Lens prutrin preparurion.s. Calf eyes were obtained from Pcl- Freer. Biologicals, Rogers, AR, shipped in fresh ice. The lenses were
washed water-insoluble fraction from human lens that was solu- bilized by sonication.
454

Glycated proteins as UVA sensitizers 455
removed and stored frozen at -70°C. The water-soluble fraction was prepared from the outer cortex of thawed, decapsulated lenses by removing the outer 1.0 cm of tissue with a cork borer. This tissue was homogenized in deionized water with a hand Dounce homog- enizer and the homogenate centrifuged at 30000 g for 20 min. The supernatant was dialyzed extensively against deionized water and used directly for glycation experiments. Normal human lenses were obtained from the Lions Eye Tissue Bank of Missouri and stored frozen in Hank’s basic salt solution until use. Twenty human lenses (55-75 years) were decapsulated, pooled and homogenized in 10 mL of deionized water. The 30000 g pellet was obtained and washed three times with an equal volume of deionized water. The pellet was resuspended once again and sonicated in ice for 5 min at a power setting of 4 and a duty cycle of 40% as described previ- o ~ s l y . ’ ~ , ~ ’ The solubilized protein was recovered after centrifugation at 30000g, and the pellet was resuspended and sonicated again. The second supernatant, when combined with the first, represented 95% of the total water-insoluble fraction and was designated the water- insoluble sonicate supernatant (WISS) fraction. All protein prepa- rations were filter-sterilized into sterile tubes and stored in the re- frigerator wrapped in aluminum foil.
The a-crystallin was prepared from a calf lens cortical extract by chromatography over a Sephadex G-200 column. The first-eluting peak was pooled and chromatographed on an Agarose A-5m column to remove any high molecular weight aggregates from the major 800 kDa a-crystallin peak. This material was used directly or lyophilized and stored at -15°C.
Glycation reactions. Dialyzed calf lens cortical proteins ( 10 mg/ mL) were incubated with 20 mM L-ascorbic acid in 0.1 M phosphate buffer containing 1 .0 mM diethylenetriaminepentaacetic acid (DTPA). Several reaction mixtures were prepared and sterile-filtered into sterile plastic centrifuge tubes. Each tube was wrapped in alu- minum foil and incubated in the dark at 37°C. Tubes were removed at weekly intervals and frozen at -70°C. At 4 weeks all samples were thawed and dialyzed before analysis. Reactions were also car- ried out with other carbohydrates each present at 20 mM in the reaction mixture described above.
Illumination conditions. Protein determinations were made on each lens fraction using the bicinchoninic acid (BCA) methodz6 ac- cording to the procedure described by the manufacturer (Pierce, Rockford, IL). Each protein preparation was diluted to 2.0 mg/mL and 3.5 mL were placed in a glass-stoppered quartz cuvette. Initial illuminations were carried out with a 200 W super pressure mercury lamp (Oriel Corp, Stratford, CT) equipped with a 20 cm water filter and 338 nm cutoff filter (50% transmission at 338 nm) to remove all UVB light. Illumination was carried out for 2 h, which resulted in an accumulated dose of 1.0 kJ/cm2 as measured with a YSI- Kettering model 65 radiometer. The solutions were continuously stirred throughout the irradiation period. Samples were withdrawn and placed immediately into cryogenic Nunc tubes and frozen in liquid nitrogen. Dark controls were kept in an airtight container at room temperature during the time of illumination and also stored in liquid nitrogen until analyzed.” The observations made with the ascorbate-glycated samples were repeated with new preparations and illuminated with a 1000 W mercury/xenon lamp. The light was fil- tered by a 10 cm water-jacketed filter (Oriel Cop, 6227) containing a 5% copper sulfate solution and the 338 nm cutoff filter. Both the copper sulfate filter and the water-jacketed cuvette holder were cooled with a circulating water bath set at 17°C. Temperature mea- surements in the cuvette showed temperatures no higher than 25°C at any time during the illumination. The protein solutions were stirred continuously during the illumination and the light was mea- sured by the black block detector mounted directly behind the cu- vette and connected to the YSI-Kettering radiometer. Illumination was continued for 1 h for a total illuminance of 1.5 kJ/cm2 for light transmitted by the 338 nm filter minus the light transmitted by a 475 nm filter. The absorbed light was measured to be 100 mW/cm2, after correcting for scattered light by determining the absorbance of the proteins dissolved in 4.0 M guanidine hydrochloride. This so- lution was optically clear and exhibited only half the absorbance compared to the slightly opalescent WISS fraction.
Spectral analysis. Absorption spectra were determined in a Perkin Elmer Corp. Lambda 3 dual beam spectrophotometer. Data collec- tion and plotting were accomplished using the IFA3 program and
the computer data station supplied by the manufacturer. Fluores- cence spectra were determined with a Perkin Elmer Model 650-40 spectrofluorometer. Spectra were gathered using either the PECUV program supplied by the manufacturer or the Surfer program (Gold- en Software, Golden, CO) to obtain the differential excitation and emission scans.
Chemical analyses. Hydrogen peroxide was assayed by the meth- od of Jiang et af.” Briefly, a 0.1 mL aliquot of illuminated protein was mixed with 0.1 mL each of 0. I M sorbitol, 0.25 M H2S0, and 1 .0 mM xylenol orange and 0.6 mL water. The reaction was initiated by the addition of 0.1 mL of 2.5 mM ferrous ammonium sulfate (made fresh daily) with vortexing. The absorbance at 560 nm was then determined after 45 min and compared to a hydrogen peroxide standard curve. Tryptophan content was determined by the method of Spies and Chambers,zx and SH groups were measured with DTNB by the method of Sedlack and Lindsay.29 The SH-group determi- nations were made in 5.1 M urea to assure accessibility as described previo~sly.’~ All of these assays were carried out in triplicate for both the irradiated and dark control samples. Histidine, lysine and arginine values were taken from a standard amino acid analysis-<” following acid hydrolysis. The values were expressed as either nmoll 1000 nmol total amino acids or nmollmL using a value of 8.0 pmol amino acids/mg protein.
RESULTS
T h e water-insoluble fraction from aged human lenses con- tains the bulk of the protein-bound chromophores and fluo- rophores. Illumination of this fraction with UVA light causes photolysis of the amino acids His, Cys, Met and Trp” with n o change in the other constituent amino acids. These pho- tolytic reactions were also accompanied by the production of p M concentrations of hydrogen peroxide:” Because the protein-bound fluorophores are thought to represent the ac- cumulated posttranslational modifications of lens pro- teins,”’.jj it seemed possible that the sensitizer activity in this fraction could be due to the presence of advanced glycation endproducts, generated a t least in part, f rom the reaction of lens proteins with the oxidation products of ASA
Dialyzed outer cortical proteins f rom calf lenses were in- cubated with 20 mM ASA under air for 0 4 weeks. Samples were removed a t weekly intervals and the amino acid com- position determined. Amino acid analysis exhibited losses in only three amino acids due to the ascorbate glycation; how- ever, reliable values for C y s and Met were not obtained. Figure 1 shows the content of Lys, Arg and His as a function of t ime of incubation. Ascorbate-dependent losses of 9.4 nmol of Lys (18%), 5.4 nmol of Arg (8.0%) and 2.5 nmol of His (6.3%) were seen. These losses were likely all due to glycation as opposed to oxidation, because the ascorbate- dependent generation of oxygen free radicals was prevented by the inclusion of a heavy metal chelator. A total o f 17.4 nmol/ l000 nmol of amino acid were modified after 4 weeks, which i s equivalent t o 140 nmol/mg protein, or slightly greater than three glycation-modified amino acids per crys- tallin subunit (assuming an average molecular weight of 25 kDa). A n exact repeat of this glycation experiment produced essentially the same rate of amino acid glycation as judged by amino acid analysis (data not shown). A reaction without A S A or with 20 mM glucose under the same conditions pro- duced n o change in the amino acid analysis with t ime of incubation.
The absorption spectrum of each protein sample is shown in Fig. 2. The initial calf lens protein preparation exhibited n o absorbance above 300 nm; however, the samples glycated

456 B. J. ORTWERTH et u /
70 0 0 0 -
60 0 0 0
50 0 - 2 C 40
Arg - ASA
Arg + ASA
Lys - ASA
T O -
Lys + ASA
His t ASA Hb - ASA
I I I I I 0 1 2 3 4
Incubation time (wks) Figure 1. The extent of glycation as measured by the Arg, Lys and His content of lens proteins incubated in air in the presence and absence of 20 mM ASA.
with ascorbate displayed increasing absorbance in the UVA region of the spectrum (320400 nm) in agreement with our prior report.j4 The spectrum for the 4 week glycated sample was indistinguishable from that of a WISS preparation iso- lated from human lenses of approximately 70 years of age. Both of these preparations absorbed light at higher wave- lengths, extending to 430 nm. These chromophores are pro- tein bound, since each preparation was extensively dialyzed prior to obtaining the spectra. All spectra were gathered on samples containing I .O mg/mL protein as determined by the BCA method. The 4 week glycated sample was straw-col- ored at a protein concentration of 10 mg/mL.
Each glycated protein preparation was illuminated with a 200 W mercury lamp for 2 h and analyzed for H202 pro- duction and amino acid photodestruction (Fig. 3). As the ascorbate-glycation reaction proceeded, there was a corre- sponding linear increase in the ability to generate H,O, upon UVA illumination (Fig. 3A). When the proteins were incu- bated under the same conditions without ascorbate, no per- oxide was generated. Similarly, dark-incubated samples of both the glycated and control proteins generated no peroxide (<1.0 nmol/mL) over the illumination period. The 4 week sample generated 33 nmol/mL H,O, following a total illu- mination of 1.0 kJ/cm2 (338475 nm). A repeat of this ex- periment with a different preparation of ASA-glycated lens proteins produced a maximum of 50 nmol/mL H,02 follow- ing an illumination with the 1000 W lamp (1.5 kJ/cm2). The glycation experiment with 20 mM glucose in place of ascor- bate produced no hydrogen peroxide when illuminated.
Figure 3B shows the effect of ascorbate-glycation on the UVA photolysis of tryptophan. A linear decrease was ob- served with a loss of 40 nmol/mL o r 18% of the total tryp- tophan after 2 h (1 .O kJ/cm2 illumination). The repeat of this experiment produced a loss of 65 nmol/mL of tryptophan after an irradiation of 1.5 kJ/cm2, which represented a 26% loss. There was no loss in Trp in the samples incubated without ascorbate or with glucose. Figure 3B shows the dark control values for each DreDaration to demonstrate that the
1.2{
I A
250 310 370 430 Wavelength (nm)
Figure 2. Absorption spectra of lens proteins incubated with 20 mM ASA in air for 0, 1, 2, 3 and 4 weeks and the absorption spectrum of a WISS fraction isolated from aged human lenses. The curves of the 2 and 3 week samples, as well as the 4 week samples and the WISS sample were identical.
Trp loss was not due to the ascorbate glycation and to show more directly the effect of only the UVA illumination.
Amino acid analysis of the samples indicated a UVA- dependent loss of histidine. Figure 3C shows a decrease in histidine content as the extent of ascorbate-glycation pro- ceeded, with again no loss in the samples incubated in the absence of ascorbate. A comparison with the dark controls, however, showed that much of the histidine loss in Fig. 3C was due to the ASA glycation (see Fig. 1 ) . After 4 weeks of glycation, the UVA-dependent photolysis of histidine was only 3.4 nmol/l000 nmol or 54 nmol/mL (9%) after an il- lumination with 1.0 kJ/cm2 of UVA light for 2 h and in the repeat experiment a loss of 5.2 nmol/l000 nmol or 83 nmol/ mL (15%) following an illumination with 1.5 kJ/cm2. The repeat experiment, however, produced only a slight loss of histidine after 2 weeks of glycation. No effect was observed with 20 mM glucose in place of ascorbate either due to gly- cation or illumination. No photolytic loss of tyrosine or phe- nylalanine was seen with any sample. During illumination, the ascorbate-glycated samples gave a slight, but consistent, increase in lysine and arginine content not seen in the dark controls (data not shown). This suggests a possible photo- destruction of Lys-Arg crosslinks.
Experiments were also carried out to determine the loss of SH groups as a result of UVA illumination. Definite SH destruction was seen in prior studies with the human WISS,’7 but only a slight loss was seen with the ascorbate-glycated samples. This was due in large part to the prior loss of SH groups, which occurred during the glycation period. The dark controls of the 4 week glycated sample were only 30% of the samples incubated without ascorbate. A loss was seen on illumination, but the values were too low to be reliable.
Figure 4 shows the rate of hydrogen peroxide production during the illumination with the 1000 W lamp (1.5 kJ/cm2 after 1 h of illumination). With each preparation there was a linear increase in the production of H,02 during the illu- mination period, arguing that there was no loss of sensitizer activitv in any samule due to photodestruction. Experiments
Glycated proteins as UVA sensitizers
g “01 560 E h
J
\
- 0
E 480-
0) C .- 0 $ 400- U
45;
C
K +ASA
I I I I I
321 A 24-
16-
8-
Q) ‘0 300- E Y
r n 200-
0
0 4
+ASA (DC) q I B
80- h
J E \ 60- u) Q, -
40- C Y
cv % 20- I
0 -_
2 400
m \
+ASA (UV) r
P 100
0 1 2 3 4
Time of incubation (wks) Figure 3. The effects of UVA irradiation ( I .0 kJ total) on lens proteins incubated for 0, 1, 2, 3 and 4 weeks in the presence and absence of 20 mM ASA. (A) The production of hydrogen peroxide with time of incubation. (B) The photolysis of tryptophan. (C) The photolysis of histidine. Panel B shows the dark control values for tryptophan content.
were camed out where a large excess of catalase (250 units/ mL) was included with the glycated proteins during the il- lumination. In those cases, no H,Oz formation was detected. Similarly, catalase added after the illumination but prior to the H,O, assay abolished all the peroxide that had formed. Clearly this is hydrogen peroxide formed during the illu- mination and not an oxidant produced during the glycation reaction. Assays for peroxide made immediately after the glycation reactions were terminated showed that no signifi- cant H,02 was produced (< 1 .O nmol/mL) during the 4 week incubation. The A,,, measurements for ascorbate (Em = 1.5 X lo4) showed that only 3% of the original 20 mM ascorbate remained after 4 weeks of incubation (data not shown).
Sensitizer formation was determined with different sugars and under different reaction conditions. All reactions were incubated for 4 weeks under the conditions described, dia- lyzed and subjected to both amino acid analysis and assays
wks
wks
wk
wks i- wks
Cat
0 15 30 45 60
Time of irradiation (mid Figure 4. Kinetics of hydrogen peroxide formation during the UVA irradiation ( 1 .5 kJ) of lens proteins that had been glycated for 0, 1, 2, 3 and 4 weeks, and a 4 week sample irradiated in the presence of 250 units/mL of catalase.
for sensitizer activity following UVA illumination. Table 1 shows the extent of glycation as the loss of lysine, arginine and histidine content and an estimate of the formation of advanced glycosylation endproducts by the absorbance at 350 nm. The lens proteins incubated for 4 weeks with 20 mM ascorbate, as well as with 20 mM dehydroascorbate and 20 mM L-threose, gave an equivalent extent of glycation. The values for Lys and Arg after glycation are very similar to those reported in Fig. 1 , and to the values obtained for a sonicated water-insoluble fraction from 60-70 year old hu- man lenses (WISS). The presence of an additional chelating agent, phytic acid, had no effect on the extent of glycation. A marked increase in 350 nm absorbance correlated directly with glycation. The reaction with fructose, like glucose, ex- hibited no apparent glycation. The RNase lost 25% of its lysine content upon glycation, as well as lesser amounts of arginine and histidine. The increase in 350 nm absorbance
Table 1. The effect of glycation on the amino acid composition and absorbance at 350 nm of calf lens proteins
nmoU1000 nmol
Argi- Histi- Reaction mixture* Lysine nine dine A,,,
LP - ASA 50 68 40 0.020 LP + ASA, zero time 51 67 40 0.067 LP + ASA 42 64 38 0.672 LP + ASA + 1.0 mM phytic acid 39 63 36 0.871 LP + DHA under nitrogen 40 62 37 0.752 LP + threose 41 61 38 0.526 LP + fructose 51 68 40 0.012 Human lens WISS (60-70 years) 43 73 32 0.480 RNase - ASA 86 34 33 0.009 RNase + ASA 65 30 32 0.795
*All reaction mixtures contained 10 mg/mL lens soluble protein, 2C mM sugar, 100 mM phosphate buffer, pH 7.0 and 1 .0 mM DTPA Incubations were for 4 weeks at 37°C in the dark. Analyses were carried out on the dialyzed proteins after 4 weeks of incubation The A,,, values are for 10 mg/mL solutions of dialyzed proteins

3.5 8 B. J . ORTWERTH et a1
94 67 43
0 7
x 30
r 20
14
1 2 3 4 5 6 7 8 9 1011121314
Figure 5. SDS-PAGE of lens proteins glycated with a variety of sugars. Lanes I , 10, 1 1 and 14 contain the molecular weight stan- dards, and lanes 2-8 contain lens proteins incubated for 4 weeks under the following conditions: lane 2, without ascorbate; lane 3, zero time with 20 mM ascorbate; lane 4 with 20 mM ascorbate; lane 5 , with 20 mM ascorbate and 1.0 mM phytic acid; lane 6, with 20 mM dehydroascorbate under nitrogen; lane 7, with 20 mnA4 L-threose; lane 8, with 20 mM fructose; lane 9, aged human WISS proteins; lane 12, RNnse alone; lane 13, RNase with 20 mM ascorbate.
with RNase was equivalent to the incubations with lens pro- teins.
Each of the preparations was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) to estimate the extent of protein crosslinking by the various glycation conditions. Figure 5 shows extensive protein cross- linking with ascorbic acid and its oxidation products. Again, phytic acid had no effect and fructose caused no protein crosslinking. The RNase was crosslinked, but mainly dimer formation was seen. The data show a direct relationship be- tween acid-stable glycation products (loss of Lys, Arg and His) and the formation of protein crosslinks with the lens proteins. A sample of a typical WISS fraction from aged human lenses is shown for comparison. The extent of cross- linking is typical of WISS preparation^'^.^^.^^ but some of the apparent monomers were likely due to a contamination by the proteins in the adjacent well.
measured and the data are presented in Table 2. The effect of ascorbate-glycation, as measured by H202 production and Trp and His photolysis, was quantitatively similar to the val- ues shown in Fig. 3 and to the photolysis of a human lens WISS under the same illumination conditions (1.5 kJ/h/cm2). The fact that both dehydroascorbic acid and L-threose were as effective as ascorbate in producing sensitizers is consis- tent with our prior observations on the relative glycation ability of ascorbate oxidation Sensitizer for- mation i s due to the glycation reactions, as opposed to the oxidation of tryptophan residues, because all reactions were carried out in the presence of 1.0 mM DTPA, and because the presence of an additional chelator, phytic acid, did not diminish the sensitizer activity. Also, ascorbate glycation of
The sensitizer activity of the various preparations w
Table 2. The effect of UVA illumination of glycated proteins on HzOz production and amino acid photolysis -
Reaction mixture"
HzOz produc-
tion TI-p loss His loss (nmol/ (nmol/ (nmol/ mL) mL) mL)
~
LP - ASA LP + ASA, zero time LP i ASA LP + ASA + 1.0 mM phytic acid LP + DHA under nitrogen LP + threose LP + fructose Human lens WISS (60-70 years) RNase ~ ASA RNase + ASA
0 0 0 0 0 0
50 50 91 72 68 80 .56 58 96 52 52 93 0 0 22
47 71 98 0 0 0
52 0 136
*Samples were those described in Table I . Solutions of the dialyzed proteins were illuminated with 1.5 kJ of UVA light over a I h period. The 100% value for Trp was 310 and for His was 640 nrnol/mL.
RNase A produced greater sensitizer activity, even though this protein is devoid of tryptophan. Fructose, like glucose, produced no sensitizer activity. The absorption values at 350 nm of the samples in Table 1 were consistent with the sen- sitizer activity in Table 2. The illumination with the 1000 W lamp produced a 40% loss in 350 nm absorbance in all the samples incubated with either ascorbate or the ascorbate oxidation products (data not shown). This argues for signif- icant photodestruction of the advanced glycation endprod- ucts.
To determine whether the sensitizer activity resided in the crosslink structures, the ascorbate-glycated proteins were subjected to Sephadex G-200 chromatography in the pres- ence of 4.0 M guanidine hydrochloride. The monomer and crosslinked peaks, when pooled and assayed, had equal sen- sitizer activity (data not shown). Therefore, advanced gly- cation products, which are bound to protein, but which are not crosslinks, also have sensitizer activity.
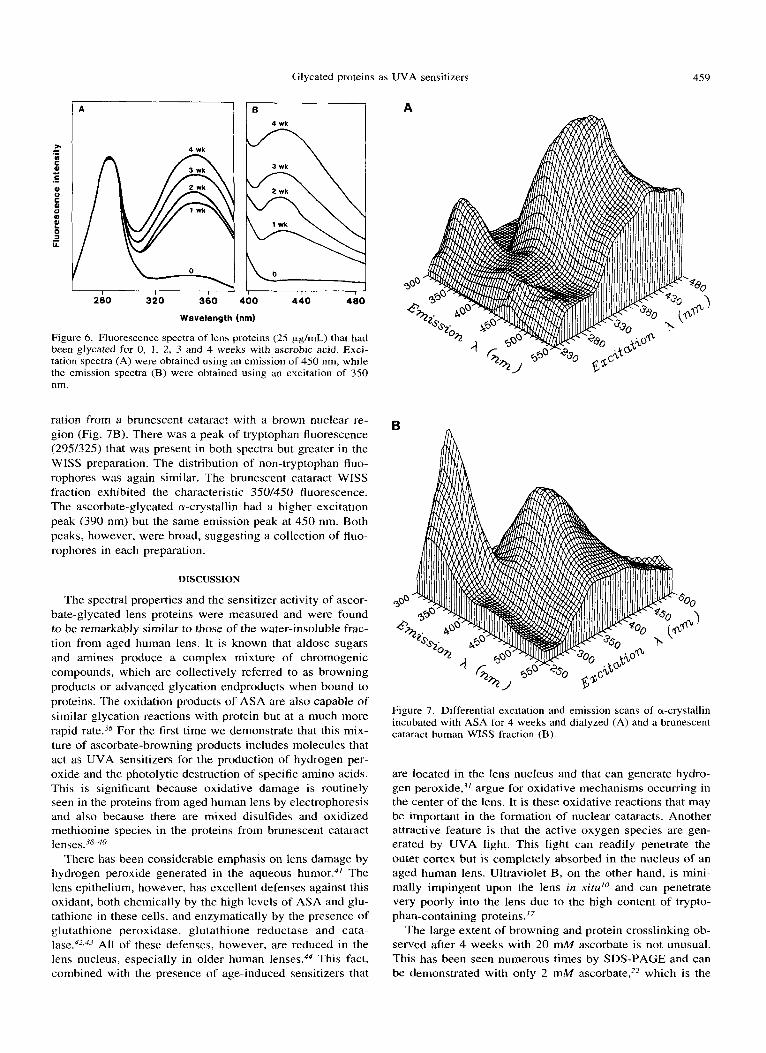
Previous work has shown that ascorbate glycation leads to the formation of fluorophores with E x E m of 350/430 nm.-j4 Fluorescence spectra of the glycated samples used for the UVA illumination experiments (Fig. 6) show increasing fluorescence with time of glycation. There i s little difference in the shape of the various spectra. The spectrum of the 4 week glycated sample exhibited excitation peaks at 290 nm and 340 nm, and a single X,,,, near 425 nm for emission. These values were very similar to those reported previously for the WISS fraction isolated from human lens (350/430).'7 It is known, however, that a large collection of protein- bound fluorophores is present in the aged human lens whose spectra vary with different excitation wavelengths. There- fore, a more detailed look at the fluorophores produced by ascorbate glycation was obtained. The major component of the lens WISS fraction is a-crystallin.'.' A sample of a-crys- tallin was isolated from bovine lens and glycated for 4 weeks with ASA in air. An extensively dialyzed sample of this material was subjected to a differential excitation and emis- sion scan and the data presented as a three-dimensional plot (Fig. 7A). The same was done for a washed WISS prepa-
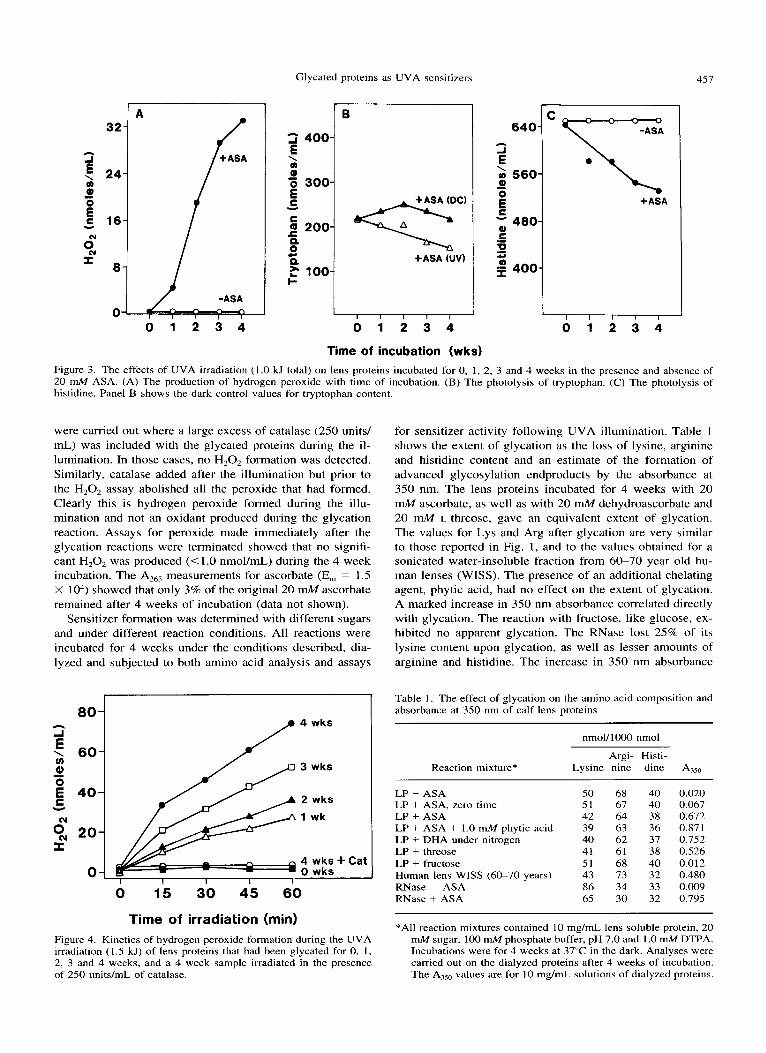
Glycated proteins as UVA sensitizers 459
A B 4 wk
i
280 320 360 460 440 460
Wavelength (nm)
Figure 6. Fluorescence spectra of lens proteins (25 pg/mL) that had been glycated for 0, I , 2, 3 and 4 weeks with ascrobic acid. Exci- tation spectra (A) were obtained using an emission of 450 nm, while the emission spectra (B) were obtained using an excitation of 350 nm.
ration from a brunescent cataract with a brown nuclear re- gion (Fig. 7B). There was a peak of tryptophan fluorescence (295/325) that was present in both spectra but greater in the WISS preparation. The distribution of non-tryptophan fluo- rophores was again similar. The brunescent cataract WISS fraction exhibited the characteristic 350/450 fluorescence. The ascorbate-glycated a-crystallin had a higher excitation peak (390 nm) but the same emission peak at 450 nm. Both peaks, however, were broad, suggesting a collection of fluo- rophores in each preparation.
DISCUSSION
The spectral properties and the sensitizer activity of ascor- bate-glycated lens proteins were measured and were found to be remarkably similar to those of the water-insoluble frac- tion from aged human lens. It is known that aldose sugars and amines produce a complex mixture of chromogenic compounds, which are collectively referred to as browning products or advanced glycation endproducts when bound to proteins. The oxidation products of ASA are also capable of similar glycation reactions with protein but at a much more rapid rate.j6 For the first time we demonstrate that this mix- ture of ascorbate-browning products includes molecules that act as UVA sensitizers for the production of hydrogen per- oxide and the photolytic destruction of specific amino acids. This is significant because oxidative damage is routinely seen in the proteins from aged human lens by electrophoresis and also because there are mixed disulfides and oxidized methionine species in the proteins from brunescent cataract lenses.3840
There has been considerable emphasis on lens damage by hydrogen peroxide generated in the aqueous humor.4/ The lens epithelium, however, has excellent defenses against this oxidant, both chemically by the high levels of ASA and glu- tathione in these cells, and enzymatically by the presence of glutathione peroxidase, glutathione reductase and cata- la~e .~’ .~’ All of these defenses, however, are reduced in the lens nucleus, especially in older human lenses.44 This fact, combined with the presence of age-induced sensitizers that
A
B
Figure 7. Differential excitation and emission scans of a-crystallin incubated with ASA for 4 weeks and dialyzed (A) and a brunescent cataract human WISS fraction (€3).
are located in the lens nucleus and that can generate hydro- gen peroxide,’/ argue for oxidative mechanisms occurring in the center of the lens. It is these oxidative reactions that may be important in the formation of nuclear cataracts. Another attractive feature is that the active oxygen species are gen- erated by UVA light. This light can readily penetrate the outer cortex but is completely absorbed in the nucleus of an aged human lens. Ultraviolet B, on the other hand, is mini- mally impingent upon the lens in s i tdo and can penetrate very poorly into the lens due to the high content of trypto- phan-containing proteins.”
The large extent of browning and protein crosslinking ob- served after 4 weeks with 20 mM ascorbate is not unusual. This has been seen numerous times by SDS-PAGE and can be demonstrated with only 2 mM ascorbate,z2 which is the

460 B. J. ORTWERTH er of.
level reported to be present in human l e n ~ . ~ ~ , ' ~ The loss of lysine was considerable, especially considering these sam- ples were not reduced with NaBH, prior to acid hydrolysis. This argues that only the advanced glycation products are being measured. These modified lysines, however, could not be detected as new peaks in the standard amino acid profile. In this regard it is significant that many investigators have reported a loss of protein amino groups during cataract for- m a t i 0 n , 4 ~ ~ ~ yet an amino acid analysis of these proteins also shows few, if any, modified amino acids. The lack of lysine modification with glucose and fructose does not necessarily indicate a lack of glycation but rather that the reaction pro- duced only Amadori products, which dissociated during the dialysis and acid hydrolysis steps. The low values for Lys content of the WISS suggest that this fraction may also be glycated. This is supported by the fact that even lower Lys values were obtained when a human WISS fraction was ex- tracted in the presence of NaBH, to stabilize Amadori prod- ucts.2.7
The loss of arginine and histidine during the reaction cor- responded with the crosslinking seen by SDS-PAGE. This has been reported previously." and argues for the formation of Lys-Arg and Lys-His crosslinks. There was a slight, but consistent, increase in Lys and Arg when the 4 week samples were illuminated. This could have represented a UVA pho- tolysis of some of the crosslinks. A similar increase in his- tidine may have also occurred but was masked by the his- tidine photolysis. Illumination caused a 40% decrease in absorbance at 350 nm, also suggesting photodestruction of the advanced glycation endproducts formed during the in- cubation with ASA. The SDS-PAGE showed no obvious re- lease of monomers upon illumination, but this again would have been masked by UVA-induced c r ~ s s l i n k i n g . ' ~ The gly- cation-initiated crosslinking of protein is often approximated by a measure of 350/450 fluorescence.so While there was a direct relationship between fluorescence and crosslinking, the sensitizer activity did not reside solely in the crosslinks. Separation of the glycated proteins into monomers and poly- mers showed equal sensitizer activity for both fractions. Also, the loss of 350 nm absorbance, which corresponded to the possible destruction of Lys-Arg crosslinks, did not result in a loss of sensitizer activity during the illumination. Gly- cated RNase, while displaying a higher sensitizer activity, exhibited less protein crosslinking, as only dimer formation was observed. Consistent with this was a smaller loss of His and Arg in the ascorbate-incubated RNase.
The extent of crosslinking, the absorption spectrum and the sensitizer-induced photolysis events were quantitatively almost identical to that seen for the WISS preparation from human lenses of 60-70 years of age. This fact argues for a similar content of sensitizers. Hydrogen peroxide formation proceeded linearly during the irradiation period after each week of glycation and was catalase sensitive as observed with the human WISS.-3' Peroxide formation by UVA light required glycation by ascorbate, as proteins incubated with- out ascorbate or even with glucose exhibited no peroxide formation when irradiated. This was not the result of an ascorbate-heavy metal ion oxidation of tryptophan, as gly- cated RNase was equally active. It has been shown that gly- cated proteins can cause the chemical generation of perox- ide,27.i' but this was insignificant in our ascorbate elvcated
samples, when assayed immediately after the 4 week incu- bation. Oxidation may have occurred during glycation, how- ever, as evidenced by the loss of S H groups. This oxidation may have depleted these low concentrations of glycation- generated H202 especially considering the length of the in- cubation period. The glycated proteins, however, were not capable of extensive peroxide generation, as no significant peroxide formation (<2 nmol/mL) was seen in our dark con- trols incubated for as long as 2 h.
The lack of sensitizer activity in the glucose-incubated samples argues that the formation of advanced glycation endproducts are necessary to produce the sensitizers. These endproducts can form, but not from glucose itself, but rather from the degradation of products of glucose that arise as a result of an attack by oxygen free radicals.5z These reactions were prevented in our incubations, because we routinely in- cluded DTPA in our reaction mixtures. This chelator has binding constants of 10I6, loz8 and lo2' for Fez+, Fe3+ and Cu2+, respectively,s3 and has been shown to prevent hydrox- yl radical formation in reactions with lens proteins.s4
Hydrogen peroxide was not directly responsible for the loss of either tryptophan or histidine but was likely due to singlet oxygen or hydroxyl radicals.4' The photolysis of his- tidine is consistent with the loss of histidine reported for the water-insoluble fraction isolated from brunescent cata- racts.-7s,S6 It must be pointed out, however, that a loss of tryptophan in cataractous lenses is still controversial. It is of interest in this regard that Dilley and Pirie56 stated that a decrease in tryptophan in bruncescent cataracts was observed using the tryptophan assay employed in this work.28
Ascorbate glycation of calf lens soluble proteins produced a modified protein fraction that had an identical absorption spectrum and a similar fluorescence spectrum to those rou- tinely obtained for isolated WISS preparations from human lens. The A,,, values vary somewhat, but our data agree with the 3451425 fluorescence reported by Bessems and Hoen- d e r ~ , ' ~ which is increased in nuclear cataract, and with the 3401420 fluorescence observed by Satoh et al.,58 which was highest in the water-insoluble fraction from human lens. The spectrum for the ascorbate-glycated a-crystallin was differ- ent than the glycated total lens proteins, but there was still a good deal of overlap with the spectrum from the brunes- cent cataract.
The results of these experiments indicate only a potential pathological reaction, but the UVA light absorbed during a 1 h irradiation in these in vitro experiments (360 J/cm2) was equivalent to about 6 months of lens exposure for 2 h of sunlight every day at noon./O.sy The large photodestruction of histidine was equivalent to that seen in brunescent cata- r a c t ~ . ~ - ~ Although it may seem that the radicals involved in this photolysis would readily be either quenched or enzy- matically destroyed in vivo, it must be remembered that the protein concentrations in the lens nucleus can be as high as 500 mg/mL. At these concentrations the likelihood of radi- cals reacting with a Trp (80 pmol/g) or His (130 pmol/g) in protein is very high compared to ascorbic acid (1-2 Fmol/ g) or GSH (2-5 pmol/g). Also, the protein-bound sensitizers in older lenses are located in the very large protein aggre- gates (?lo8 Da), which make up the water-insoluble frac- tion.2-7 The interior of these aggregates may represent a priv- ileged site. where antioxidant molecules mav not uenetrate

Glycated proteins as UVA sensitizers 46 1
and protective enzymes are surely excluded. It is likely that the site of damage may be very near the site of generation of the active oxygen species. The protein damage in cataract lenses, therefore, may not be initiated until later in life when these aggregates form and when a significant amount of UVA-absorbing glycation endproducts have accumulated. The generation of H,Op may not only be responsible for the oxidation of cysteine and methionine seen in human cata- racts but may also cause an accelerated oxidation of ASA, thereby leading to a greater formation of glycation endprod- uct sensitizers.
Acknowledgements-The authors express their gratidude to Dr. Robert Kuntz for his continuous advice and for configuring and mea- suring the light output of our UV instrumentation. We are also in- debted to Dr. Craig Eldred for determining the differential excitation and emission scans and to Sarah Boyce for her initial studies on this project. This work was supported in part by grants EY02035 and EY07070 from the National Eye Institute, in part by a Howard Hughes Training Fellowship award to Ms. Sarah Boyce and in part by Research to Prevent Blindness Inc.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
Zigman, S., M. Datiles and E. Torczynski (1979) Sunlight and human cataract. Invest. Ophthalmol. & Visual Sci. 18, 462467. Cruickshanks, K. J., B. E. K. Klein and R. Klein (1992) Ultra- violet light exposure and lens opacities: the Beaver Dam eye study. Am. J . Public Health 82, 1658-1662. Leske, M. C., L. T. Chylack, Jr. and S.-Y. Wu (1992) The lens opacities case-control study. Risk factors for cataract. Arch. Ophthalmol. 109, 244-25 1. The Italian-American Cataract Study Group (1991) Risk factors for age-related cortical, nuclear and posterior subcapsular cata- racts. Am. J . Epidemiol. 133, 541-553. Taylor, H. R., S. K. West, F. S. Rosenthal, B. Munoz, H. S. Newland, H. Abbey and E. A. Emmett (1988) Effect of ultra- violet radiation on cataract formation. N. Engl. J . Med. 319, 1429-1433. Jose, J. G . and D. G. Pitts (1985) Wavelength dependency of cataracts in albino mice following chronic exposure. Exp. Eye Res. 41, 545-563. Pitts, D. G., A. P. Cullen and P. D. Hacker (1977) Ocular effects of ultraviolet radiation from 295 to 365 nm. Invest. Ophthalmol. 16, 932-939. Li, D-Y. and R. F. Borkman ( 1 990) Photodamage to calf lenses in vitro by excimer laser radiation at 308, 337, and 350 nm. Invest. Ophthalmol. & Visual Sci. 31, 2180-2184. Schein, 0. D., S. West, B. Munoz, S. Vitale, M. Maguire, H. R. Taylor and N. M. Bressler (1994) Cortical lenticular opaci- fication: distribution and location in longitudinal study. Invest. Ophthalmol. & Visual Sci. 35, 363-366. Sliney, D. H. (1987) Estimating the solar ultraviolet radiation exposure to an intraocular lens implant. J. Cataracr Refractive Surg. 13, 296-301. Boettner, E. A. and J. R. Wolter (1962) Transmission of the ocular media. Invest. Ophthalmol. 1, 776-783. Collier, R. and S. Zigman (1987) The gray squirrel lens protects the retina from near-UV radiation damage. In Degepzerative Ret- inal Disorders: Clinical and Laboratory Investigations (Edited by J. G. Hollyfield, R. E. Anderson and M. M. Lavail), pp. 571- 585. Alan R. Liss, New York. Ellozy, A. R., R. H. Wang and J. Dillon (1994) Model studies on the photochemical production of lenticular fluorophores. Photochem. Photobiol. 59, 479484. Dillon, J. and S. J. Atherton (1990) Time resolved spectroscopic studies on the intact human lens. Photochem. Photobiol. 51, 465468. Pirie, A. (1968) Color and solubility of the proteins of human cataracts. Invest. Ophthalmol. 7, 634-650. Zigler, J. S., Jr, and J. D. Goosey (1984) Singlet oxygen as
possible factor in human senile nuclear cataract development. Curr. Eye Res. 3, 59-65.
17. Ortwerth, B. J. and P. R. Olesen (1994) UVA photolysis the protein-bound sensitizers present in human lens. Photochem. Photobiol. 60, 53-60.
18. Stevens, V. J. , C. A. Rouzer, V. M. Monnier and A. Cerami ( 1 978) Diabetic cataract formation: potential role of glycosyla- tion of lens crystallins. Proc. Natl. Acad. Sci. USA 75, 2918- 2922.
19. Lyons, T. J., G. Silvestri, J. A. Dunn, D. G. Dyer and J. W. Baynes (1991) Role of glycation in modification of lens crys- tallins in diabetic and nondiabetic senile cataracts. Diabetes 40, 101CL1015.
20. Swamy, M. S. and E. C. Abraham (1987) Lens protein com- position, glycation and high molecular weight aggregation in aging rats. Invest. Ophthalmol. & Visual Sci. 28, 1693-1701.
21. Araki, N., N. Ueno, B. Chakrabarti, Y. Morino and S. Horiuchi (1992) Immunochemical evidence for the presence of advanced glycation end products in human lens proteins and its positive correlation with aging. J. Biol. Chem. 267, 1021 1-10214.
22. Ortwerth, B. J., M. S. Feather and P. R. Olesen (1988) the pre- cipitation and cross-linking of lens crystallins by ascorbic acid. Exp. Eye Res. 47, 155-168.
23. Nagaraj, R. H., D. R. Sell, M. Prabhakaram, B. J. Ortwerth and V. M. Monnier (1991) High correlation between pentosidine protein crosslinks and pigmentation implicates ascorbate oxi- dation in human lens senescence and cataractogenesis. Proc. Natl. Acad. Sci. USA 88, 10257-10261.
24. Ortwerth, B. J. and P. R. Olesen (1988) Glutathione inhibits and glycation and crosslinking of lens proteins by ascorbic acid. Exp. Eye Res. 47, 737-750.
25. Ortwerth, B. J. and P. R. Olesen (1992) Studies on the solubi- lization of the water-insoluble fraction from human lens and cataract. Exp. Eye Res. 55, 777-783.
26. Smith, P. K., R. I. Krohn, G . T. Hermanson, A. K. Mallia, F. H. Gartner and M. D. Provenzano (1985) Measurement of pro- tein using bicinchoninic acid. Anal. Biochem. 150, 76-85.
27. Jiang, Z-Y., A. C. S. Woollard and S. P. Wolff (1990) Hydrogen peroxide production during experimental protein glycation.
28. Spies, I. R. and D. C. Chambers (1948) Chemical determination FEBS Lett. 268, 69-7 1.
of tryptophan. Anal. Chem. 20, 30-39. 29.
30.
31.
32.
33.
34.
35.
36.
37.
_. . Sedlak, J. and H. Lindsay (1968) Estimation of total, protein- bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 25, 192-205. Gehrke, C. W., P. R. Rexroad, R. M. Schisla, J. S. Absheer and R. W. Zumwalt (1987) Quantitative analysis of cystine, methi- onine, lysine, and nine other amino acids by a single oxida- t i o n 4 hour hydrolysis method. J. Assoc. Ofl Anal. Chem 70, 17 1-1 74. Linetsky, M., B. J. Ortwerth and P. R. Olesen (1995) Demon- stration of active oxygen species generated by UVA radiation of a human lens water-insoluble fraction. Photochem. Photobiol. (In press) Yu, N.-T., M. N. Bando and J. F. R. Kuck, Jr. (1983) Metabolic production of a blue-green fluorophor in lenses of dark-adapted mice and its increase with age. Invest. Ophthalmol. & Visual Sci. 24. 1157-1 161. Yu, N.-T., M.-Z. Cai, D. J.-Y. Ho and J. F. R. Kuck, Jr. (1988) Automated laser-scanning-microbeam fluorescence/Raman im- age analysis of human lens with multichannel detection: evi- dence for metabolic production of a green fluorophor. Proc. Narl. Acad. Sci. USA 85, 103-106. Ortwerth, B. J. and P. R. Olesen (1988) Ascorbic acid-induced crosslinking of lens proteins: evidence supporting a Maillard reaction. Biochim. Biophys. Acta 956, 10-22. Prabhakaram, M. and B. J. Ortwerth (1991) The glycation-as- sociated crosslinking of lens proteins by ascorbic acid is not mediated by oxygen free radicals. Exp. Eye Res. 53, 261-268. Slight, S. H., M. S. Feather and B. J. Ortwerth (1990) Glycation of lens proteins by the oxidation products of ascorbic acid. Biochim. Biophys. Acta 1938, 367-374. Ortwerth, B. J., J. A. Speaker, M. Prabhakaram, M. G. Lopez,

462 B. J . ORTWERTH et al
E. Y. Li and M. S. Feather (1994) Ascorbic acid glycation: the reactions of i--threose in lens tissue. Exp. Eye Res. 58, 665-674.
38. Truscott, R. J. W. and R. C. Augusteyn (1977) Oxidative changes in human lens proteins during senile nuclear cataract formation. Biochim. Biophys. Acta 492, 43-52.
39. Garner, M. H. and A. Spector (1980) Sulfur oxidation in se- lected human cortical cataracts and nuclear cataracts. Exp. Eye Res. 31, 361-369.
40. Lou, M. F. and J . E. Dickerson, Jr. (1992) Protein-thiol mixed disulfides in human lens. Exp. Eye Res. 55, 889-896.
41. Spector, A. and W. H. Garner (1981) Hydrogen peroxide and human cataract. Exp. Eye Res. 33, 673-68 I .
42. Babizhayev, M. A,, A. I. Deyev and A. V. Chernikov (1992) Pcroxide-metabolizing systems of the crystalline lens. Biochim. Biophvs. Actu 1138, 11-19.
43. Giblin, F. J . , J . R. Reddan, L. Schrimscher, D. C. Dziedzic and V. N. Reddy (1990) The relative roles of the glutathione redox- cyde and catalase in the detoxification of H202 by cultured rab- bit lens epithelial cells. Exp. Eye Res. 50, 795-804.
44. Pau, H., P. Graf and H. Sies ( I 990) Glutathione levels in human lens: regional distribution in different forms of cataract. Exp. Eye Res. 50, 17-20.
4.5. Heath, H. (1962) The distribution and possible functions of ascorbic acid in the eye. Exp. Eye Res. 1, 362-367.
46. Taylor, A,, P. F. Jacques, D. Nadler, F. Morrow, S . I. Sulsky and D. Shepard (1992) Relationship in humans between ascor- bic acid consumption and levels of total and reduced ascorbic acid in lens, aqueous humor, and plasma. Curr. Eve Re.s. 10, 7 s 1-79.
47. Garcia-Castineiras, S . and M. N. Miranda-Rivera (1983) Loss of frcc amino groups in the water-insoluble fraction of nuclear senile cataract. Invest. Ophthalmol. & Visual Sci. 24, 1181- 1187.
48. Rao, G. N. and E. Cotlier (1986) Free epsilon amino groups and 5-hydroxymethylfurfural contents in clear and cataractous human lenses. Invest. Ophthulmol. & Visual Sci. 27, 98-102.
49. Simonelli, F., L. Cotticelli, A. Iura, C . Manna, A. Nesti, E.
Rinaldi and G. Auricchio (1990) The decrease of free epsilon- amino groups in senile and diabetic cataracts. Ophthnlmic Res. 22, 160-165.
50. Richard S., C. Tamas, D. R. Sell and V. M. Monnier (1991) Tissue-specific effects of aldose reductase inhibition on fluores- cence and cross-linking of extracellular matrix in chronic galac- tosemia. Relationship to pentosidine cross-links. Diabetes 40, 1049-1 056.
51. Mullarkey, C. J., D. Edelstein and M. Brownlee (1990) Free radical generation by early glycation products: a mechanism for accelerated atherogenesis in diabetes. Biochenz. Biuphys. Rex Cornmun. 173, 932-939.
52. Fu, M.-X., K. J. Wells-Knecht, J. A. Blackledge, T. J. Lyons, S. R. Thorpe and J. W. Baynes (1994) Clycation, glycoxidation, and cross-linking of collagen by glucose. Kinetics, mechanisms. and inhibition of late stages of the Maillard reaction. Diabetes 43, 676-683.
53. Nishikimi, M. and T. Ozawa (1987) Stabilization of ascorbate solution by chelating agents that block redox cycling of metal ions. Biochem. Int. 14, I 1 1-1 17.
54. Hunt, J. V., R. T. Dean and S. P. Wolff (1988) Hydroxyl radical production and autoxidative glycosylation. Biochem. J . 256, 205-2 12.
55. Zigler, J . S . , Jr., J. B. Sidbury, Jr., B. S. Yamanashi and M. Wolbarsht (1976) Studies o n brunescent cataracts I. Analysis of free and protein-bound amino acids. Ophthalmic Res. 8, 379- 387.
56. Dilley, K . J . and A. Pirie (1974) Changes to the proteins of the human lens nucleus in cataract. Exp. Eye Res 19, 59-72.
57. Bessems, C . J. H. and J. H. Hoenders (1987) Distribution of aromatic and fluorescent compounds within single human lens. Exji. Eye Rex 44, 817-824.
58. Sato, K., M. Bando and A. Nakajima (1973) Fluorescence in human lens. Exp. Eye Rex 16, 167-172.
59. Sliney, D. H. (1986) Physical factors in cataractogenesis: am- bient ultraviolet radiation and temperature. Invest. Ophthalmol. & Visuul Sci. 27, 78 1-790.