aryl hydrocarbon (ah) receptor dna-binding activity hydrocarbon (ah) receptor dna-binding activity...
TRANSCRIPT

THE JOURNAL OF BIOLOCKXL CHEMISTRY Vol. 265. No. 16. Issue of June 5, pp. 9251-9258,199O 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
Aryl Hydrocarbon (Ah) Receptor DNA-binding Activity SEQUENCE SPECIFICITY AND Zn2’ REQUIREMENT*
(Received for publication, June 21, 1989)
Fahri SaatciogluS, David J. Perry, David S. Pasco, and John B. Fagant From the Molecular Biology Laboratory, Maharishi International University, Fairfield, Iowa 52556
The aryl hydrocarbon (Ah) receptor, also called the xenobiotic or TCDD receptor, mediates transcriptional activation of the cytochrome P-450~ (CYPlAl) gene by interacting with Ah or xenobiotic response ele- ments. This paper presents evidence that a metal ion, probably Zn’+, is an essential cofactor for the Ah recep- tor. This paper also maps in detail the interactions between the Ah response element XREl and the Ah receptor from the rat hepatocyte-derived cell line LCS7. Interactions were mapped by three methods, 1) methylation interference footprinting, 2) mobility shift competition experiments, using a series of synthetic oligonucleotides with systematic alterations in the Ah response element core sequence, and 3) orthophenan- throline/Cu+ footprinting. These findings suggest the following consensus sequence for DNA recognition by the Ah receptor: CNA/TNA/TCACGCA/TA/T. The che- lators l,lO-phenanthroline and oxalic acid inhibited the sequence-specific DNA-binding activity of the Ah receptor in a concentration dependent manner, sug- gesting that the DNA-binding activity of the receptor requires divalent metal ions. Inhibition was due to metal-chelation, since: 1) inhibition was almost com- pletely prevented by the presence of Zn’+, or other divalent metal ions having high affinity for the chela- tors used, while metal ions with low affinity did not protect; 2) the DNA-binding activity of the receptor could be restored by dialysis to remove l,lO-phenan- throline, but only in the presence of Zn’+, while dialysis in the absence of metal ions reversed inhibition by the nonchelating isomer 4,7-phenanthroline. The involve- ment of a divalent cation in receptor function, possibly bound via sulfhydryls, was also suggested by the find- ing that Cd2’ and Co2+ inhibited DNA-binding activity. Once bound to the XREl DNA sequence, the receptor could not be inhibited by l,lO-phenanthroline, sug- gesting that the essential metal ion must become inac- cessible to chelation when the receptor binds DNA. The Zn2+ requirement of the Ah receptor is similar to that of the estrogen and the glucocorticoid receptors and is consistent with the hypothesis that the Ah receptor is a member of the steroid and thyroid hormone receptor superfamily.
The polycyclic aromatic hydrocarbon-inducible cytochrome
* This project was supported by Research Grant ROl-CA 38655 from the National Cancerhrstituteand by a grant from the Maharishi International Universitv Research Fund. The costs of uublication of this article were defrayed in part by the payment of-page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of an MIU Predoctoral Fellowship. 5 To whom correspondence should be addressed.
P-450IA’ subfamily of the cytochromes P-450’ is capable of both detoxifying polycyclic aromatic compounds and convert- ing some compounds to highly active carcinogens and muta- gens. Treatment with polycyclic compounds, such as TCDD, 3-methylcholanthrene, and /3-naphthoflavone, causes P- 450IAl mRNA levels to increase loo-fold both in hepatocyte cultures and in liver (Fagan et al., 1986; Kawajiri et al., 1984; Pasco et al., 1988). Both transcriptional and post-transcrip- tional mechanisms have been shown to be important in the regulation of the CYPlAl’ gene of rat (P-450~; Pasco et al., 1988), mouse (Pi-450; Kimura et al., 1986), and human (Pi- 450; Thierry et al., 1987). DNA-mediated gene transfer studies have identified multiple regulatory elements, both positive and negative, associated with the CYPlAl gene. One of these is responsive to polycyclic compounds and is termed the Ah response element, TCDD response element, or xenobiotic response element (Fujisawa-Sehara et al., 1986, 1987; Sogawa et al., 1986; Jones et al., 1985, 1986a, 1986b; Gonzalez and Nebert, 1985; Neuhold et al., 1986).
The proposed model for the action of TCDD and other polycyclic compounds is remarkably similar to the current model of steroid hormone action (Ringold, 1985; Yamamoto, 1985), in which a hydrophobic ligand binds with high affinity to a soluble receptor protein and the resulting ligand-receptor complex acts as a modulator of transcription for several genes (Poland and Knutson, 1982; Poland and Kimbrough, 1984).
The Ah receptor, also known as the TCDD receptor and xenobiotic receptor, when complexed with its ligand can undergo temperature-dependent transformation to become a DNA-binding protein (Gasiewicz and Bauman, 1987; Hannah et al., 1986). The sequence-specific interactions of the murine Ah receptor with the Ah response element of the CYPlAl gene have been previously demonstrated by the mobility shift DNA-binding assay (Denison et al., 1988a, 198813; Fujisawa- Sehara et al., 1988; Hapgood et al., 1989). In this paper we map in detail, by three independent methods, the interactions of the rat Ah receptor with the Ah response element of the rat CYPlAl (P-450~) gene. This new, detailed information makes it possible to construct a much more precise model of the interactions between the Ah receptor and its cognate sequence.
We also report in this paper the effect of several divalent cations and of the chelating agents l,lO-phenanthroline and oxalic acid on the DNA-binding activity of the Ah receptor. Our data show that Cd2+, Co*+, l,lO-phenanthroline, and oxalic acid all inhibit binding of the Ah receptor to the Ah response element XREl and that Zn*+ can prevent inhibition by l,lO-phenanthroline and can reverse this inhibition when
i See Nebert et al. (1989) for current P-450 nomenclature. ‘The abbreviations used are: P-450, cytochrome P-450; TCDD,
2,3,6,7-tetrachlorodibenzo-p-dioxin; CDTA, 1,2-cyclohexanediamine- tetraacetic acid.
9251
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

9252 Ah Receptor DNA-binding Activity
present during removal of l,lO-phenanthroline by dialysis. Similarly, metal ions chelated by oxalic acid prevent the inhibitory effect of this agent. These results suggest that a metal ion, most likely Zn2+ is an essential cofactor for the DNA-binding activity of the Ah receptor. Since the “zinc finger” motif is a common feature of the DNA binding do- mains of all known members of the steroid and thyroid hormone receptor superfamily, and since Zn*+ is an essential cofactor for the glucocorticoid receptor (Freedman et al., 1988) and for the estrogen receptor (Sabbah et al., 19871, the find- ings reported here provide additional support for the earlier hypothesis that the Ah receptor is a member of this receptor superfamily.
The bulk of the work presented in this paper has been carried out in the rat hepatocyte-derived cell line LCS7 (Chou, 1983; Chou and Schlegel-Haueter, 1981), in which it has been demonstrated that TCDD and other polycyclic compounds induce the transcription of the P-450~ gene and probably influence post-transcriptional events as well (Teifeld et al., 1989). All previously reported research concerning cis-acting transcriptional regulatory sequences associated with the P- 450~ gene and concerning putative regulatory proteins that interact wit.h these sequences, has been carried out using a single cell line, the murine hepatoma-derived cell fine Hepal (Hankinson, 1979) and its derivatives, primarily because of the availability of Ah receptor regulatory variants of this line (Hankinson, 1979; Legraverend et al., 1982; Miller et aL, 1983). Even researchers who have been investigating the function of the rat P-450~ regulatory sequences have used this mouse derived cell line (Fujisawa-Sehara et al., 1986, 1987, 1988). We have chosen to use the LCS7 cells in order to examine the regulation of t.he rat P-45& gene in a cell line derived from that same species and to evaluate the regulation of this gene in a second cell culture system.
EXPERIMENTAL PROCEDURES
Ma&+z~-MgC&, h&C&, CoCl,, CaCl,, Cd&, and oxalic acid were obtained from Fisher, ZnClt from J. T. Baker Chemical Co., and l,lO-phenanthwline and 4,7-phenanthroline from GFS Chemicals (Columbus, OH).
Gel Electrophoresis DNA-binding Assay-Nuclear extracts were incubated with approximately 5000 cpm (0.02 ng) of an end-labeled, double-stranded DNA fragment in the presence of 0.5-1.0 tig of poly(dI-dC)-poly(dI-dC) (Pharmacia LKB Biotechnology, Inc. or Boehrineer Mannheim) in a final volume of 10 al. Binding reactions were carried out in 25.mM HEPES, pH 7.5, 1.m~ EDT& 0.7 mM dithiothreitol, 100 mM KCl, and 10% glycerol. Samples were layered onto 6% low ionic strength polyacrylamide gels (acrylamide/bisacryl- amide weight ratio 191) and electrophoresed at 100 V for 90 min in buffer consisting of 20 mM HEPES, 20 rnM Tris, 1 rn~ EDT& pH 8.0, at room temperature. They were then fixed in 10% (v/v) methanal and 10% (v/v) acetic acid, transferred to Whatman 3MM paper, dried, and autoradio~raphed. All figures present representative ex- periments that have been repeated at least three times. In sOme experiments, duplicate lanes are presented for some or ali conditions to demonstrate the reproducibility of the method.
~~igonucleotides and Probe Pre~rat~on-Single-stranded oliganu- cleotides, synthesized using a Pharmacia Gene Assembler, were end- labeled with (r-32PjATP using poiynucleotide kinase (Maniatis et al., 1982) and annealed with their complementary strands by heating to 90 “C in 100 mM NaCI, 50 mM Tris-Cl, pH 7.5, and 10 mM M&12, and cooling slowly to 4 “C.
Cell Culture-LCS7 cells were obtained from J. Chou (National Institutes of Health, Bethesda, MD). This line is a clonal derivative of line RALA255-cmG (Chou, 1983). which was generated by trans. formation of adult ;at primary hepatocytes with ihe tsA255 strain of SV40 virus, which is temperature-sensitive for maintenance of trans- formation. These cells grow prolifically at the permissive temperature but regain many hepatocyte characteristics when shifted to the non- permissive temperature. Ceils from frozen stocks were maintained at 34 “C (permissive temperature for transformation) in a 1:1 mixture
of Dulbecco’s modified Eagle’s medium (4.5 mg/ml glucose) and Ham’s F-12 medium (without hypoxanthine), with 4% fetal bovine serum (K. C. Biologicals, Kansas City, MO) and 1 pM cortisol (Sigma). When confluent, the cells were transferred to 40 “C (nonpermissive temperature for transformation) for 2 days before performing exper- iments. Medium was changed every three days at 34 “C and every day at 40 “C. Gentamicin sulfate (50 rg/ml) and amphotericin B (1.5 eg/ ml) from Gibco were present in the medium at all times. Cultures were maintained in a humidified atmosphere containing 5% CO2 and 95% air.
Preparation of Nuclear &tracts-Nuclear extracts, containing the Ah receptor, were prepared according to Whitlock and Galeazzi (1984), with minor changes. Briefly, confluent LCS7 cells were har- _ _._.. vested from tissue culture roller bottles by scraping into phosphate- buffered saline (without Ca2* or MI?‘) and centrifurrine at 2000 mm for 5 min in a Beckman JS7.5 rot&.‘The cellular &.&t was iez: pended in 5 volumes of 10 mM HEPES, pH 7.5. After swelling 10 min on ice, the cells were collected by centrifugation as before and resus- pended in 5 volumes of 3 mM MgClz, 1 mM dithiothreitol, 25 mM
HEPES, pH 7.5. Cells were again collected by centrifugation and resuspended in 1 volume of the same buffer. Cells were broken with 15 strokes in a Dounce homogenizer using a tight pestle. The cell homogenate was immediately centrifuged for 30 s in an Eppendorf __ _. -. . microcentrifuge and the crude nuclear p&let resuspended in 2 volumes of 0.1 hi KCl. 25 mM HEPES. DH 7.5. and 1 mM dithiothreitol. I\Ttirl*i . . - “A- -
were lysed bi adding 2 M KCi to a final concentration of 0.4 M and mixing gently at 4 “C for 30 min. The lysate was adjusted to 20% glycerol and centrifuged at 45,000 rpm for 30 min in a Beckman Ti- 70 rotor. The transparent supernatant was stored at -60 “C in small aliquots. All procedures were performed at 4 “C.
Methyl&on Interference Assay-Methyiation interference assays were performed as described previously (Gilman et al., 1986) with a few modifications. End-labeled oligonucleotides were treated with dimethyl sulfate as described (Hendrickson and SchLif, 1985). Par- tially methylated oligonucleotides were incubated with nuclear ex- tracts as described above for mobility shift assays, except that the reaction was scaledup to 100 ~1 and contained app~ximately 300,~O cpm of DNA/reaction. After electrophoresis, DNA was electrotrans- ferred from the gel to Whatman DEBI paper overnight at 12 volts. The DE81 paper was exposed to x-ray film for 1 h at -70 “C to locate the complexed and free DNA bands, which were excised, and the DNA was eluted by incubation in 10 mM Tris-Cl, pH 7.5, I mM EDTA, 1 M NaCl for several hours at 37 “C, Carrier tRNA (10 pg) was added to the eluates, which were extracted twice with phenol/ chforoform/isoamyl alcohol (25:24:1, v/v) and once with chloroform/ isoamyl alcohol (24:1, v/v). DNA was precipitated twice with EtOH, resuspended in 80% (v/v) formamide, 50 mM Tris borate, pH 8.3, and 1 mM EDTA, denatured at 100 “C for 2 min and applied to a standard 10% polyacrylamide sequencing gel.
ortho-YhenQnthroline/Copper Footprint&-A preparative scale mobility shift reaction was carried out and electrophoresed as de- scribed above. The gel was then treated with o&o-phenanthroline,’ Cu+ reagent, which cleaves the DNA while still in the gel (Kuwabara and Sigman, 1967). DNA was electrotransferred to DE&l paper, isolated, and run on standard sequencing gels, as described above.
RESULTS
DNA ~~~i~ Activity of the Ah Receptor from Rat Hepa- t~y~e-der~ue~ LCS7 Cells-The Ah receptor from the murine hepatoma cell line Hepal has been previously shown to bind sequence specifically to Ah response elements (Denison et al., 1988a, 1988b; Fujisawa-Seh~a et: aZ., 1988; Hapgood et ai., 1989). Fig. 1 shows that a similar DNA-binding activity is present in the nuclei of LCS7 cells, a rat hepatocyte-derived cell line. LCS? cells were treated for the specified times with the poIycycIic compound ~-naphtho~avone and nuclear ex- tracts prepared by the same procedure used to prepare extracts containing the Ah receptor from Hepal cells (Denison et al., 1988a, 198813). The mobility shift, or gel electrophoresis, DNA-binding assay (Fried and Crothers, 1981; Garner and Revzin, 1981; Strauss and Vaushavsky, 1984) was used to detect nucfear proteins that bound sequence specifically to the Ah response element XREl. A 32P-labeled double stranded, 40-base pair oligonucleotide, whose sequence was
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Ah Receptor DNA-binding Activity
5’-CTCCAGGCTCTTCTCACGCAACTCCGGGGCAC- 3’ (coding strand), was used as probe in these experiments. This oligonucleotide corresponded in sequence to the region from -997 to -1028 from the transcription start site of the P-450~ gene, with additional bases at each end to create BamHI- or BglII-ligatable ends. This sequence spans the core region of XREl, bases -1007 to -1021, one of the Ah response elements of the P-450~ gene (Fujisawa-Sehara et al., 1987). This domain is identical in sequence to one of the Ah response elements of the murine P-450IAl gene and has been used to characterize the DNA-binding properties of the murine Ah receptor from Hepal cells (Denison et al., 1988a, 1988b; Hapgood et al., 1989).
Fig. 1 shows that, following treatment with P-naphthofla- vone, XREl-binding activity rapidly appeared in the nuclei of LCS7 cells, with kinetics similar to those observed for the appearance of the Ah receptor in the nuclei of Hepal cells (Denison et al., 1988a). Fig. 1 also shows that the rapid appearance of XREl-binding activity in the nuclei of LCS7 cells was not blocked by inhibition of protein synthesis, as has been reported for the Ah receptor from Hepal cells (Denison et al., 1988a). These and other findings concerning the nuclear XREl-binding activity isolated from LCS7 cells suggests that this activity corresponds to the Ah receptor observed in nuclear extracts from Hepal cells. The findings supporting this conclusion include the DNA footprinting data presented below.
Mobility Shift Competition Experiments-A series of mo- bility shift competition studies were performed, first to deter- mine if the DNA-protein complexes detected in the experi- ments shown in Fig. 1 were sequence-specific, and second to precisely map these interactions.
The experiment presented in Fig. 2 tested the ability of a series of “mutant” double-stranded oligonucleotides to com- pete with ‘*P-labeled XREl-40 for binding to the Ah receptor present in nuclear extracts from LCS7 cells. As shown in Fig. 3, each mutant oligonucleotide differed by 3 bases from the sequence of XREl-15, a 15-base pair oligonucleotide contain- ing the core domain of XREl. Fig. 2 shows that the bands corresponding to the Ah receptor diminished when unlabeled XREl-40 (lanes 2-4) or XREl-15 (lanes 5-7) was included in DNA binding reactions at molar concentrations 5-, 25-, and loo-fold higher than that of “‘P-labeled XREl-40. In contrast, these bands were undiminished in DNA binding reactions carried out in the presence of the same concentra- tions of oligonucleotides M3 and M4 (lanes 14-19) and were
cy‘ 4-B ycleB
-‘4-F *. ‘4-F
FIG. 1. Ah receptor from rat LCS7 cells. A, LCS7 cells were treated with @-naphthoflavone (5 wg/ml) for O-120 min, as specified. Cells were harvested and nuclear extracts prepared as described under “Experimental Procedures.” Specific binding to XREl-40 was meas- ured by the mobility shift assay. B, before harvesting, cells were treated for 30 min with p-naphthoflavone (B) and/or 60 min with 10 gg/ml cycloheximide (X). B indicates specific DNA-protein com- plexes, and F indicates free XREl-40 cDNA.
C~ALlLaIL-blLJILdlL~ 1 2 3 4 5 6 7 tl 9 10 11 1213141516171819202122
rw +B
4-F
FIG. 2. Specific binding of the Ah receptor to the Ah re- sponse element XREl of the P-450~ gene. Nuclear extracts from P-naphthoflavone-treated LCS7 cells were incubated with XREl-40 (0.04 ng, 10,000 cpm), and the reactions were resolved by electropho- resis. Lane 2 contained no competitor DNA; each subsequent set of three lanes contained 0.1, 0.5, dr 2.0 ng (5-, 25, and lOO-fold molar excess, respectively) of competitor; XREl-40 (lanes 2-4), XREI 15- base pair core region (lanes 5-7), XREl-related oligonucleotides, Ml (lanes &IO), M2 (lanes II-13), M3 (lanes 14-16), M4 (lanes 17-19), M5 (lanes 20-22). The positions of the specific protein-DNA com- plexes (B) and free probe (F) are indicated.
WT TCTTCTCACGCAACT Ml AGA------em---- M2 ---A(-J,--------- M3 ------GTG------ M4 ---------CGT--- M5 ------------TG&
FIG. 3. Sequence of XREl-related oligonucleotides used in competition studies. The 15-base pair XREl core sequence is specified. Only those bases which differ from the wild-type (WT) XREl sequence are indicated for the mutant oligonucleotides.
decreased only slightly in reactions carried out in the presence of oligonucleotide Ml (lanes 8-10). Similar concentrations of oligonucleotide M2 competed weakly, while those of M5 com- peted almost as effectively as XREl-15 itself for binding of 32P-labeled XREl-40 by the Ah receptor.
These findings indicate that the band labeled B in Figs. 1 and 2 represents a sequence-specific DNA-protein complex, since the ability of oligonucleotides to compete for binding with 32P-labeled XREl-40 was sequence-dependent. These results also indicate which bases within the XREl core se- quence are most important for formation of this complex. For instance, in oligonucleotide Ml, the triplet TCT, spanning base pairs -1020 to -1018 in XREl, had been replaced with AGA. The greatly attenuated ability of this oligonucleotide to compete with XREl-40 for binding to form DNA-binding complexes indicated that at least one of these 3 base pairs was important in binding the Ah receptor. Similarly, the inability of oligonucleotides M3 and M4 to compete indicated that at least 1 base pair in the triplet spanning positions -1011 to -1009 and at least 1 base pair in the triplet spanning positions -1008 to -1006 must have been important in the interaction of the Ah receptor with XREl-40. Since oligonu- cleotides M2 and M5 competed to some extent, we conclude that the positions at which these oligonucleotides differ from XREl-15 must have been less important for binding of the Ah receptor. It should be pointed out that all of the mutant oligonucleotides which competed with XREl-40 for binding of the Ah receptor did so less strongly than the wild-type XREl core sequence. It is possible that alteration of three consecutive bases anywhere in the core sequence reduces binding somewhat, either by causing local alterations in DNA
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

9254 Ah Receptor DNA-binding Activity
conformation or by disrupting secondary DNA-protein con- tacts.
Methylation Interference Footprinting-To further charac- terize the interactions of the Ah receptor with XREl, the methylation interference footprinting technique was used (Hendrickson and Schleif, 1985). This procedure identifies guanine residues that, when methylated at the N-7 position in the major groove, prevent or weaken binding of a factor. End-labeled XREl-40 was partially methylated with dimethyl sulfate (Hendrickson and Schleif, 1985) and used as probe in a preparative scale mobility shift reaction. Free and bound DNAs were separated by electrophoresis, isolated from the acrylamide gel, cleaved with piperidine, and run on a standard 10% sequencing gel. When XREl-40, 5’-end-labeled on the coding strand, was used as a probe, one band corresponding to the guanine at position -1011 was dramatically reduced in intensity in complexes formed with the Ah receptor (Fig. 4), indicating that this guanine was important for the interaction of the receptor with XREl. When the contacts on the non- coding strand were mapped, it was found that methylation of the guanines located at -1010, -1012, and -1014 suppressed binding of the Ah receptor. Thus, the guanines at position -1011 on the coding strand and at positions -1010, -1012, and -1014 on the noncoding strand were involved in forma- tion of complexes with the Ah receptor.
ortho-PhenanthrolinelCopper Footprinting-Footprinting by the ortho-phenanthroline/Cu’ footprinting technique (Ku- wabara and Sigman, 1987) was used as a third technique to map the interactions of the Ah receptor with XREl. In this procedure, DNA-protein complexes were resolved from free DNA by electrophoresis and then subjected to cleavage by ortho-phenanthroline/Cu+ reagent while still in the gel. This takes advantage of the increased stability of DNA-protein
mmINGNoN4.3lmuK FBF FBF
c :: \i l -101.
-loll.:- I - - - \T C
2 a-1a1a - - --c*-1010
T
FIG. 4. Methylation interference analysis. Methylation inter- ference footprinting was performed as described under “Experimental Procedures” using XREl-40, “‘P-labeled on either strand, as indi- cated, and nuclear extracts from P-naphthoflavone-treated LC57 cells. F represents the free probe, showing cleavage at all guanines, and R represents DNA-protein complexes, as in Figs. 1 and 2. The nucleotide sequence corresponding to the region of interest is pre- sented at the side of the figure. The guanines at which methylation blocks binding are indicated by arrows on the sequence.
CODING FB FG
NON-CODING FBFG
FIG. 5. ortho-Phenanthroline/Cu+ chemical footprinting analysis. ortho-Phenanthroline/Cu’ footprinting was performed as described under “Exoerimental Procedures” using as probe XREl- 40, end-labeled on thk coding and noncoding strands ai indicated. B and F, DNA-protein complexes and free DNA, respectively; C, gua- nine sequencing ladder. Heavy and light bars, those bases within the DNA sequence that were protected completely or partially from ortho- phenanthroline/Cu+ cleavage, respectively. Light exposures indicated that radioactivity in excess of that in other lanes had been loaded in lanes R in both panels. This was taken into account when the boundaries of the footprints were defined.
complexes when they enter the gel matrix. After cleavage, the DNA was eluted from the gel and run on a standard sequenc- ing gel. As shown in Fig. 5, the Ah receptor protected and therefore interacted with the same general region of XREl- 40 shown by competition mobility shift and methylation interference mapping to be important for Ah receptor binding. The boundaries of the protected domain on the coding strand were at bases -1004 to -1005 and -1018 and on the noncod- ing strand at bases -1006 and -1022.
Inhibition of the DNA Binding Activity of the Ah Receptor by l,lO-Phenanthroline-We have investigated the possible role of divalent metal ions in the DNA-binding function of the Ah receptor from LCS7 cells. The experiment presented in Fig. 6 investigated the effect of the chelator l,lO-phenan- throline on the formation of complexes between “‘P-labeled XREl-40 and the Ah receptor present in nuclear extracts from /3-naphthoflavone-treated LCS7 cells. Incubation of nu- clear extract with l,lO-phenanthroline prior to the addition of “ZP-labeled XREl-40 inhibited formation of Ah receptor- XREl complexes in a concentration-dependent manner (Fig. 6a, compare lanes 1 and 2 with lanes 3-7). Half-maximal inhibition was observed at 1 mM l,lO-phenanthroline and maximum with 3.0 mM. Note that treatment with l,lO-phen- anthroline did not decrease the intensity of other bands, some of which have been shown to be due to sequence-specific interactions of other factors with XREl.’ This indicates that the effect of l,lO-phenanthroline on the Ah receptor was
” F. Saat.cioglu, D. J. Perry, D. S. Pasco, and J. B. Fagan, submitted for publication.
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Ah Receptor DNA-binding Actiuity 9255
1 1 3 4 5 6 7 a 9 10 II 12.34 56
FIG. 6. Inhibition of the Ah receptor DNA-binding activity by l,lO-phenanthroline and protection from inhibition by Zn *+. A nuclear extract was incubated at 23°C for 15 min with O-3 mM l,lO-phenanthroline (lanes 1-7) or 15 min with 1 inM divalent cation followed by 15 min with 2 mM l,lO-phenanthroline (lanes 8-12). DNA binding activity was measured by mobility shift method. B, the effect of the nonchelating isomer 4,7- phenanthroline on the binding of the Ah receptor to XREl-40 was assessed. Numbering system is as in Panel A, except that the effects of Ca’+, M$‘, and Mn”+ were not tested, and 4,7-phenanthroline was used in place of l,lO- phenanthroline.
specific. This inhibition was almost completely prevented if Zn’+ was added to the nuclear extract before addition of l,lO- phenanthroline (Fig. 6A, lane 8). Mg2+ protected much less effectively (less than 20X), whereas Ca2+ and Mn2+ were ineffective (Fig. 6A, lanes 9-11). When 4,7-phenanthroline, a phenanthroline isomer that largely lacks the ability to chelate divalent cations, was tested, inhibition was observed (Fig. 6B, lanes l-5), but an excess of Zn” or other divalent cations could not prevent this inhibition, suggesting inhibition by another mechanism (Fig. 6B, compare lanes 5 and 6).
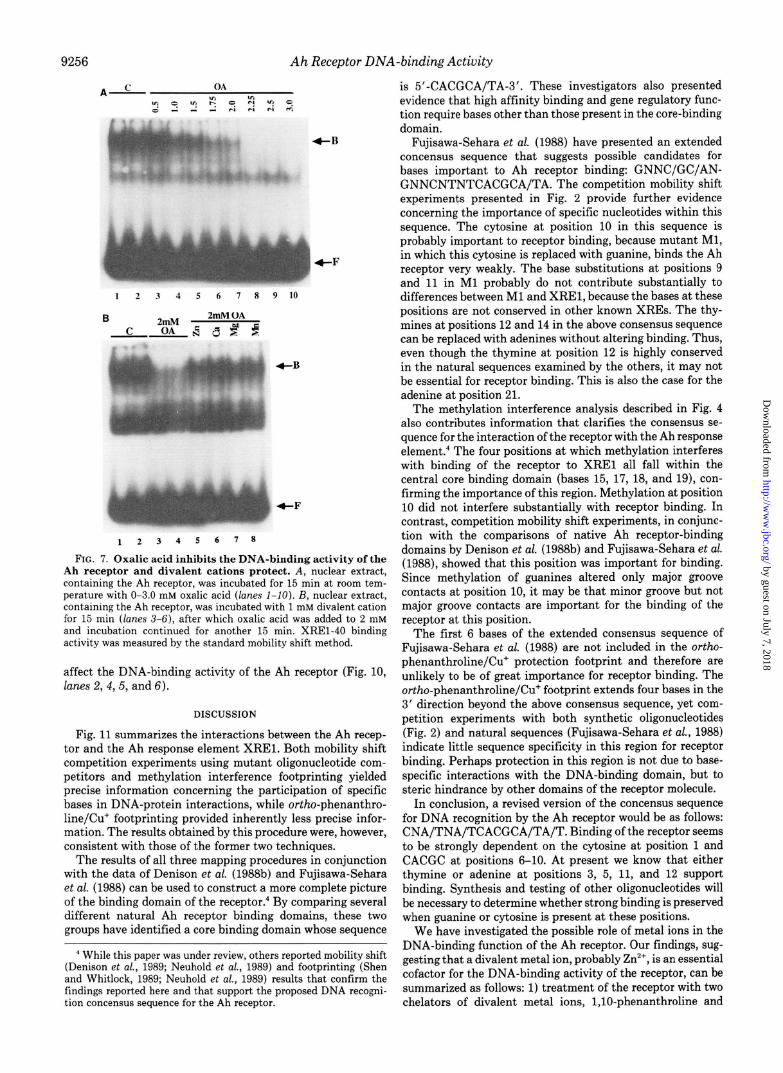
Inhibition of the DNA-binding Activity of the Ah Receptor by Oxalic Acid-EDTA and CDTA, both of which are effective chelators of several divalent cations, including Zn’+, had no significant effect on the DNA-binding activity of the Ah receptor, even at concentrations as high as 20 mM (data not shown). However, another chelator, oxalic acid, was found to inhibit the DNA-binding activity of the receptor. Fig. 7A shows that oxalic acid inhibited receptor activity in a concen- tration-dependent manner (compare lanes 1 and 2 with lanes 3-8). Half-maximal inhibition was achieved at 1.0 mM oxalic acid and complete inhibition at 2.25 mM. Zn’+, Ca”, Mg’+, and Mn*+, all of which are known to be chelated with similar affinities by oxalic acid, all prevented receptor inhibition by about 50% (Fig. 7B, compare lanes 3 and 4 with lanes 5-8). These results support the findings with l,lO-phenanthroline and suggest that the Ah receptor requires a metal ion for DNA-binding activity.
Inactivation of the Ah Receptor by l,lO-Phenanthroline Is Reversible-Although adding Zn*+ to nuclear extracts before the addition of l,lO-phenanthroline prevented inhibition of the DNA-binding activity of the TCDD receptor, once lost, this activity could not be restored by adding Zn*’ in excess of l,lO-phenanthroline to mobility shift reactions (data not shown). The same result was obtained when the experiment was carried out under an argon atmosphere in the presence of dithiothreitol (data not shown). This suggested that the oxidation of sulfhydryl residues, which may be essential to receptor-DNA interactions, was not responsible for the inhib- itory action of l,lO-phenanthroline. Therefore, we tried re- storing receptor activity by removing l,lO-phenanthroline.
When extracts treated with l,lO-phenanthroline were di- alyzed in the absence of Zn*+, inhibition persisted (compare lanes 3 and 4 with lanes 9 and 10, Fig. 8). However, dialysis of l,lO-phenanthroline-treated extracts in the presence of Zn*+ restored the DNA binding activity of the receptor more than 50% (Fig. 8, compare lanes 3 and 4 with lanes 11 and 12). These observations suggest that the mechanism of inhi- bition by l,lO-phenanthroline involves the reversible removal of a metal ion(s), probably Zn*+, from the Ah receptor through the formation of a metal-chelator complex. The findings presented in Fig. 8 also indicate that the mechanism by which 4,7-phenanthroline partially inhibits the DNA-binding activ- ity of the receptor must be different from that of l,lO- phenanthroline, since removal of 4,7-phenanthroline by di- alysis in the absence of Zn*+ restored receptor activity more than 90% (Fig. 8, compare lanes 5 and 6 with lanes 13 and 14). This was in contrast to the failure to restore receptor activity by removing l,lO-phenanthroline in the absence of Zn2+, as pointed out above.
DNA Binding Protects the Ah Receptor from Inhibition by l,lO-Phenanthroline-In contrast to the substantial inhibi- tion of the receptor when l,lO-phenanthroline was added to nuclear extracts prior to the DNA binding reaction, inhibition was decreased substantially when the receptor was bound to XREl prior to treatment with l,lO-phenanthroline (Fig. 9). This was not due to protection by poly(dI-dC) .poly(dI-dC) or to interference by any other component of the binding reac- tion. l,lO-Phenanthroline efficiently inhibited the DNA- binding activity of the receptor in the presence of all reaction components (Fig. 9, compare lanes 1 and 2 with lanes 5 and 6), as long as it was added prior to the addition of “‘P-labeled DNA (XREl-40) (Fig. 9, compare lanes 5 and 6 with lanes 7 and 8).
Influence of Divalent Cations on the DNA-binding Activity of the Ah Receptor-Several divalent cations were tested for their direct effect on the DNA-binding activity of the Ah receptor. CoCl, or CdC12, included at 1 mM in mobility shift reactions, completely abolished the binding of the Ah receptor to XREl (Fig. 10, compare lanes 1 and 2 with lanes 3 and 7). ZnC12, CaC12, MgC12, and MnCL at 1 mM did not measurably
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from
9256 Ah Receptor DNA-bindingActivity
fB
+F
12345678
FIG. 7. Oxalic acid inhibits the DNA-binding activity of the Ah receptor and divalent cations protect. A, nuclear extract, containing the Ah receptor, was incubated for 15 min at room tem- perature with O-3.0 mM oxalic acid (lanes I-IO). B, nuclear extract, containing the Ah receptor, was incubated with 1 mM divalent cation for 15 min (lanes 3-6), after which oxalic acid was added to 2 mM and incubation continued for another 15 min. XREl-40 binding activity was measured by the standard mobility shift method.
affect the DNA-binding activity of the Ah receptor (Fig. 10, lanes 2, 4,5, and 6).
DISCUSSION
Fig. 11 summarizes the interactions between the Ah recep- tor and the Ah response element XREl. Both mobility shift competition experiments using mutant oligonucleotide com- petitors and methylation interference footprinting yielded precise information concerning the participation of specific bases in DNA-protein interactions, while ortho-phenanthro- line/Cu+ footprinting provided inherently less precise infor- mation. The results obtained by this procedure were, however, consistent with those of the former two techniques.
The results of all three mapping procedures in conjunction with the data of Denison et al. (198813) and Fujisawa-Sehara et al. (1988) can be used to construct a more complete picture of the binding domain of the receptor.4 By comparing several different natural Ah receptor binding domains, these two groups have identified a core binding domain whose sequence
’ While this paper was under review, others reported mobility shift (Denison et al., 1989; Neuhold et al., 1989) and footprinting (Shen and Whitlock, 1989; Neuhold et al., 1989) results that confirm the findings reported here and that support the proposed DNA recogni- tion concensus sequence for the Ah receptor.
is 5’-CACGCA/TA-3’. These investigators also presented evidence that high affinity binding and gene regulatory func- tion require bases other than those present in the core-binding domain.
Fujisawa-Sehara et al. (1988) have presented an extended concensus sequence that suggests possible candidates for bases important to Ah receptor binding: GNNC/GC/AN- GNNCNTNTCACGCA/TA. The competition mobility shift experiments presented in Fig. 2 provide further evidence concerning the importance of specific nucleotides within this sequence. The cytosine at position 10 in this sequence is probably important to receptor binding, because mutant Ml, in which this cytosine is replaced with guanine, binds the Ah receptor very weakly. The base substitutions at positions 9 and 11 in Ml probably do not contribute substantially to differences between Ml and XREl, because the bases at these positions are not conserved in other known XREs. The thy- mines at positions 12 and 14 in the above consensus sequence can be replaced with adenines without altering binding. Thus, even though the thymine at position 12 is highly conserved in the natural sequences examined by the others, it may not be essential for receptor binding. This is also the case for the adenine at position 21.
The methylation interference analysis described in Fig. 4 also contributes information that clarifies the consensus se- quence for the interaction of the receptor with the Ah response element.4 The four positions at which methylation interferes with binding of the receptor to XREl all fall within the central core binding domain (bases 15, 17, 18, and 19), con- firming the importance of this region. Methylation at position 10 did not interfere substantially with receptor binding. In contrast, competition mobility shift experiments, in conjunc- tion with the comparisons of native Ah receptor-binding domains by Denison et al. (1988b) and Fujisawa-Sehara et al. (1988), showed that this position was important for binding. Since methylation of guanines altered only major groove contacts at position 10, it may be that minor groove but not major groove contacts are important for the binding of the receptor at this position.
The first 6 bases of the extended consensus sequence of Fujisawa-Sehara et al. (1988) are not included in the ortho- phenanthroline/Cu+ protection footprint and therefore are unlikely to be of great importance for receptor binding. The ortho-phenanthroline/Cu+ footprint extends four bases in the 3’ direction beyond the above consensus sequence, yet com- petition experiments with both synthetic oligonucleotides (Fig. 2) and natural sequences (Fujisawa-Sehara et al., 1988) indicate little sequence specificity in this region for receptor binding. Perhaps protection in this region is not due to base- specific interactions with the DNA-binding domain, but to steric hindrance by other domains of the receptor molecule.
In conclusion, a revised version of the concensus sequence for DNA recognition by the Ah receptor would be as follows: CNA/TNA/TCACGCA/TA/T. Binding of the receptor seems to be strongly dependent on the cytosine at position 1 and CACGC at positions 6-10. At present we know that either thymine or adenine at positions 3, 5, 11, and 12 support binding. Synthesis and testing of other oligonucleotides will be necessary to determine whether strong binding is preserved when guanine or cytosine is present at these positions.
We have investigated the possible role of metal ions in the DNA-binding function of the Ah receptor. Our findings, sug- gesting that a divalent metal ion, probably Zn*+, is an essential cofactor for the DNA-binding activity of the receptor, can be summarized as follows: 1) treatment of the receptor with two chelators of divalent metal ions, l,lO-phenanthroline and
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Ah Receptor DNA-binding Activity
FIG. 8. Reversal of l,lO-phenan- throline inhibition by dialysis in the presence of Zn2+. Nuclear extract, con- taining the Ah receptor, was left un- treated (C) or incubated at 23 “C with 2 mM l,lO-phenanthroline or 2 mM 4,7- phenanthroline for 15 min, as indicated. DNA-binding activity was measured by the mobility shift method either directly (lanes Z-6) or after dialysis for 3 h at 4 “C (lanes 7-14). The dialysis buffer contained 0.1 M KCl, 25 mM HEPES, pH 7.5, and 1 mM dithiothreitol, except lanes II and 12, which also contained 0.1 mM ZnC12.
No Dialysis Dialysed
C OP 47-P C OP OPlZn 47-P
1
123456 7 8 9 10 11 12 13 14
op c OP+R.M. DNA+OP ----
C Zn Cd Ca Mg Mn CO
1 2 3 4 5 6 1 8
FIG. 9. DNA binding by the Ah receptor prevents inhibition by 1 ,lO-phenanthroline. Nuclear extract, containing the Ah recep- tor, was left untreated (lanes 3 and 4) or treated with 2 mM l,lO- phenanthroline for 15 min in the absence (lanes I and 2, OP) or presence (lanes 5 and 6, OP + RM) of poly(dI-dC) .poly(dI-dC) and all other components of the standard binding reaction mixture except XREl-40 (R.M., reaction mixture components). In lanes 7 and 8 (DNA + OP), the extract was first incubated with “P-labeled XREl- 40 to form receptor-DNA complexes and then l,lO-phenanthroline was added to 2 mM.
oxalic acid, prevented the binding of the receptor to XREl; 2) inhibition by l,lO-phenanthroline and oxalic acid could be prevented almost completely by the presence of excess Zn*+, suggesting that inhibition was due to chelation; 3) the abilities of other divalent cations to prevent inhibition by l,lO-phen- anthroline or oxalic acid were strongly correlated with their affinities (Vallec and Wacker, 1970) for these chelators; 4) the inhibition by l,lO-phenanthroline was relieved after ex- tensive dialysis in the presence of Zn*+ but not by dialysis in its absence; 5) in contrast, inhibition by the nonchelating isomer 4,7-phenanthroline was not prevented by Zn2+ or other divalent cations and was reversed almost completely by di- alysis in the absence of Zn*+. The ability to restore receptor
4-F
I 2 3 4 S 6 I
FIG. 10. Influence of divalent cations on the XREl-40 bind- ing activity of the Ah receptor. Nuclear extract was incubated for 15 min under standard mobility shift reaction conditions in the absence (C) or the presence of 1 mM divalent cation. DNA-binding activity was measured by mobility shift method.
-L CTCCAGGCTCTTCTCACGCAACTCCGGGGCAC _
- : : ..' a _ ! GAGGTCCGAGAAGAGT$CFTTGAGGCCCCGTG
FIG. 11. Summary of the interactions between XREl and the Ah receptor. P-450~ sequences -1028 to -997 are represented. Heavy and Light lines indicate strong and weak protection from ortho- phenanthroline/Cu’, respectfully, by the Ah receptor. The positions of apparent protein contacts determined by methylation interference are indicated by dots. Those bases which were important for binding, based on competition experiments with mutant oligonucleotides, are indicated by bars above the coding strand.
activity by dialysis to remove l,lO-phenanthroline in the presence of Zn”‘, but not in its absence, indicated that the removal of a metal ion by l,lO-phenanthroline may cause a reversible change in the receptor that prevents its binding to XREl. Although these experiments suggest that this metal
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

9258 Ah Receptor ~NA-birding Actiuity
ion is probably Zn’+> another metal ion for which Zn2+ can substitute, and which we have not tested, may be the natural cofactor for receptor function.
Although the data presented show that Zn2’ is an essential cofactor for Ah receptor DNA-binding activity, they do not directly indicate whether Zn2+ is tightly bound to the receptor or is in rapid equilibrium with the solvent. We tested other chelators for their effect on the DNA-bin~ng activity of the Ah receptor. Interestingly, EDTA and CDTA, which are known to chelate Zn2+ effectively, did not inhibit binding of the receptor to XRE1, even at high concentrations. Both of these compounds are structurally quite different from l,lO- phenanthroline and oxalic acid. The fact that structurally diverse chelators vary in their ability to inactivate the receptor and that this variation is not correlated with their affinities for Zn*+ (e.g. EDTA, which is noninhibitory, has a much higher affinity for Zn2+ than does oxalic acid, which is inhib- itory} indirectly suggests that the essential Zn2+ is not in rapid equilibrium with the solvent. It is more likely to be sequestered such that access to it by chelators is sterically constrained, and, therefore, dependent upon the structure of the chelator used. The finding that association of the receptor with DNA protects from inhibition by l,lO-phenanthroline is also con- sistent with the possibility that Zn2+ is bound to the receptor. Direct evidence bearing upon this point must await purifica- tion of sufficient quantities of the Ah receptor to analyze its divalent cation content and the chemical nature of the asso- ciation of this ion with the receptor.
We have found that Cd” and Co2+ directly inhibited Ah receptor binding to XREl. The mechanism by which Cd*+ and Co*+ inhibit is unknown at this time. It is thought that the steroid receptor binds Zn2+ through cysteine sulfhydryls. If the Ah receptor is a member of the same receptor family, it is possible that Cd’+ and Co”+ inhibit the Ah receptor by displacing the physiological divalent cation from the receptor and forming a complex with the receptor that is incapable of binding DNA. Alternatively they may interact with other cysteines that are required for the DNA-binding activity of the receptor.
Many structural and functional similarities (Xannah et aZ., 1986; Okey et al., 1979, 1980; Wilhelmsson et al., 1986) based on hydrodynamic properties and signal transduction, as well as differences based on genetic findings (Yamamoto et al., 1976; Whitlock, 1983, have been observed between the Ah receptor and the steroid receptors, particularly the glucocor- ticoid receptor. Based on the extensive similarities reported, it has been suggested that the Ah receptor belongs to the steroid and thyroid hormone receptor superfamily (Evans, 1988). The members of this family have a multidomain struc- ture, the most highly conserved of which is the DNA-binding domain with zinc-finger motifs analogous to those of tran- scription factor TFIIIA (Evans, 1988). Zn2* has been shown to be an essential cofactor for both the glucoeortieoid receptor (Freedman et ai., 1988) and the estrogen receptor (Sabbah et ah, 1987). Here, we have presented parallel findings for the Ah receptor, showing that the sequence-specific DNA binding activity of this receptor requires a metal ion, most likely Zn2+. These resuits are consistent with the hypothesis that the Ah receptor is a member of the same receptor superfamily. Clon- ing, sequencing, and expression of the cDNA for the Ah receptor, in conjunction with mutagenesis and gene transfer experiments, will be required to conclusively establish
whether or not the Ah receptor is a member of this receptor superfamily.
REFERENCES
Chou, J. Y. (1983) Mol. Ceil Biol. 3,1013-1020 Chou, J. Y., and Schlegel-Haueter, S. E. (1981) J. Cell. B&l. 89, 216-222 Cresteil. T.. Jaiswal. A. K., and Eisen, H. J. (1987) Arch. Bosom. &o&s.
253,??331240 D&son. M. S.. Fisher. J. M.. and Whitlock. J. P.. Jr. (1988a) Proe. N&l. Acud.
&i U: S. A. ilk%, 25i8-2532 Denison, M. S., Fisher, J. M., and Whitl~k, J. P., Jr. (1988bf J. Eiol. Chem
263,17221-17224 Den&on, M. S.. Fisher, J. M., and Whitlock, J. P., Jr. (1989) J. B&E. C&en.
264.1647%i6482 Evans, R. M. (1988) Science 240.889-895 Fagan, J. B., Pastewka, J. V., Chalberg, S. C., Gozukara, E., Guengerieh, F. P.,
and Gelboin, H. V. (1986) Arch. Biochem Biophys. 244.261-272 Freedman, L. P., Luisi, B. F., Korszun, 2. R., Basavappa, R., Sigier, P. B., and
Yamamoto, K. R. (1988) Nature 334.543-546 Fried, M., and &others, D. M. (1981) Nucleic Acids At-s. 9.6505-6525 Fujisawa-Sehara, A., Sagawa, K., Nishi. C., and Fujii-Kuriyama, Y. (1986)
Nucleic Acids Res. 14, X465-1477 Fujisawa-Sehara, A., Sagawa, K., Yamane, M., and Fujii-Kuriyame, Y. (1987)
Nucleic Acids Res. 15,4179-4191 Fujisawa-Sehara, A., Yamane, M., and Fujii-Kuriyama, Y. (1988) Proc. NotL
Acad. Sci. U. S. A. 85,585~~5863 Garner, N. M., and Rezvin, A. (1981) Nucleic Acids Res. 9,3047-3060 Gasiewicz, T. A., and Bauman, P. A. (1987) J Bill. Chem. 262.2116-2120 Gilman, M. Z., Wilson, R. N., and Wienberg, R. A. (1986) Mol. Cell. Bill. 6.
4305-4316 Gonzalez, F. J., and Nebert, D. W. (1985) Nucleic Acids Res. 13,7269-7288 Hannah, R. R., Lund, J., Poellinger, L., Gillner, M., and Gustafsson, J. A.
(1986) Eur. J. B&hem. 156.237-242 Hapgood, J., Cuthill, S., Denis, M., Poellinger, L., and Gustafsson, J. (1989)
Proc. N&l. Acad. Sci. U. S. A. 86, 60-64 Hankinson, 0. (1979) Proc. Nan. Acad. Sci. U. S. A. 76, 373-376 Hendrickson, W., and Schleif, R. (1985) Proc. Natl. Acad. Sci. U. S. A. 82,
3129-3133 Jones. P. B. C.. Galeazzi. D. R.. Fisher, J. M.. and Whitlock, J. P., Jr. (1985)
S&me 227,‘1499-1502 Jones. P. B. C., Durrin, L. K., Galeazzi, D. R., and Whitlock, J. P., Jr. (1986a)
Pm. N&l. Acad. Sci. U. S. A. 83, 2802-2806 Jones, P. B. C., Durrin, L. K., Fisher, J. M., and Whitlock, J. P., Jr. (198613) J.
Eiol. Chem. 261 6647-aacn Kawaiiri, K.. Got& C
c, Y Y Y Y
)., Tagashira, Y., Sagawa, K., and Fujii-Kuriyama, Y. (1984) J. Biol. Chem. 259,10145-10149
Kimura. S.. Gonzalez, F. J., and Nebert, D. W. (1986) Mol. Cell. Biol. 6, 1471- 1477.
Kuwabara, M. D., and Sigman, D. S. (1987) Biochemistry 26.7234-7238 Legraverend, C., Hannah, R. R., Eisen, H. J., Owens, I. S., Nebert, D. W., and
Hank&on, 0. (1982) J. Biol. Chem. 257,6402-6407 Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning: A
Loooratory Manual, pp. X23-124, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Miller, A. G., Israel, D., and Whitlock, J. P., Jr. (1983) J. Biot. Chem. 258, 3523-3527
Nebert, D., Nelson, D., Adesnik, M., Coon, M., Estabrook, R., Gonzalez, F., Guengerich, F. P., Gunsalus, I., Johnson, E., Kemper, B., Levm, W., Phillips, I., Sam, R., and Waterman, M. (1989) DNA (N. Y.) 8, 1-13
Neuhold. L. A., Gonzalez, F. J., Kaiswsl, A. K., and Nebert, D. W. (1986) DNA (N. Y.) 5,403-411
Nenhold. L. A.. Shiravoshi. Y.. Or&o. K.. Jones, J. E.. and Nebert, D. W. (1989) Mol. Cell. Bik 9, $378-i38&
Okey, A. B., Bondy, G. P., Mason, M. EI, Kahl, G. S., Eisen, H. J., Guenthner, T. M., and Nebert, D. W. (1979) j. Brat. Chem. 254,11636-11648
Okey, A. B., Bandy, G. P., Mason, M. E., Nebert, D. W., Forster-Gibson, C. J., Munean, J., and Dufresne, M. J. (1989) J. BiDi. Chem. 265.11415-11422
Pasco. D. S., Bowm. K. W., Merchant, S. N., Chalberg, S. C., and Fagan, J. B. (1988) J. &oiChem. 263,8671-8676
Poland, A., and Kimbrough, R. D. (eds) (1984) Bio~g~c~ Mec~~~~ of Dioxin Action, Coid Spring Harbor Laboratory, Cold Spring Harbor, NY
Poland, A., and Knutson, J. C. (1982) Annrr. Rev. P~Q~~~~. 7’orieoL 22,517-
Ri%k G. M. (1985) Anna. Reu. Pharaoh. Toxicol. 25 529-566 ‘M., Redeuilh, G., Seeco, C., and Baulieu, E. E. (1987) J. Biol. Chem. 8631-8635
!&& S., and Whitlock, J. P., Jr. (1989) J. Biol. Chem. 264,17754-17758
K., Fuiisawa-Sehara, A., Yamane, M., and Fujii-Kuriyama, Y. (1986) Proc. N&l. A&d. Ser. U. S. A. 83,8044-8046
Strauss, F., and Varshavsky, A. (1984) Cell 37.889-901 Teifeld. R. M., Faenn~ .I. B.. and Pasco. D. S. 119891 DNA fN. Y.) 8.329-338 Vallec, B. L., and
Vol. 5, pp. 129-146, Academic Press, Or1 Whitlock, J. P., Jr. (1987) Phormacol. Reu. Whitlock, J. P., Jr., and Galeazzi, D. R. (II Wilhelmsson. A.. Wikstrom. A. C.. a
?&&er, W. E. C. (1970~~~oT~~~ (Neurath, H., edf
39, i47-161 x-384) J. Biol. Chem. 259.980985
,nd Poellinger. L. (1986) J. Biol. Chem 261.13458-13463
Yamamoto, K. R. (1985) Annu. Reu. Genet. 19,209252 Yamamoto, K. R., Gehring, U., Stampfer, M. R., and Sibley, C. H. (1976)
Recent frog. Horn. Res. 32,3-32
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

F Saatcioglu, D J Perry, D S Pasco and J B FaganZn2+ requirement.
Aryl hydrocarbon (Ah) receptor DNA-binding activity. Sequence specificity and
1990, 265:9251-9258.J. Biol. Chem.
http://www.jbc.org/content/265/16/9251Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/16/9251.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from