are there general laws in parasite community ecology? the ... · national de baillarguet, ......
TRANSCRIPT

2.1 Introduction
There is an increasing interest in parasite and infectious disease population (Grenfell and Dobson1995; Hudson et al. 2001) and community (Eschet al. 1990; Poulin 1998a; Rohde 2001) ecology, andinterestingly this has developed at a time whenmainstream ecologists have shown increasing inter-est in metapopulation theory and habitat frag-mentation (Hanski and Gilpin 1997; Hanski 1999),population dynamics in fragmented landscape(Hassell and Wilson 1997; Ferguson et al. 1997;Grenfell and Harwood 1997; Rohani et al. 1999) andmacroecology (Brown 1995; Rosenzweig 1995;Maurer 1999; Lawton 2000; Gaston and Blackburn2000). The development of what is now called spatialecology is one of the great triumphs of modern popu-lation and community ecology (Tilman and Kareiva1997), which has showed the critical importance ofspace and spatial characteristics for understandinga wide range of ecological phenomena (Holt 1993,1999). There are clear analogies between modernspatial ecology and parasite–infectious diseasepopulation and community ecology, and this
chapter will be devoted to a review of the recentdevelopment in parasite–infectious disease popu-lation and community ecology within this fruitfulcross-fertilizing arena.
There are considerably more studies available onparasitic systems today than 10 years ago (seePoulin 1997, 1998a; Poulin et al. 2000 for a review),and many of these investigations have clearlyshowed the role of dynamical processes in a spatialcontext (Ferguson et al. 1997; Grenfell and Harwood1997; Morand and Guégan 2000a; Poulin andGuégan 2000; Morand et al. 2002). Most if not allparasites live as populations that are divided intometapopulations on several spatial scales, and eachhost operates as a patch. Many of the topics inmodern spatial ecology have also their parallels inwithin-host infection processes with the individualhost body forming an heterogeneous environment(see Holt 1999). From the perspective of a parasiticlarval form or microbe, an individual host is anextraordinary landscape to invade with het-erogeneity in resource availability and colonization–-extinction risks. An infrapopulation is thusdefined as all the members of a given parasitespecies within a single host individual, and aninfracommunity includes all of the infrapopulationswithin an individual host. The next hierarchicallevel includes all the infrapopulations sampled froma given host species within an ecosystem, and whichforms the metapopulation. Parallel to the metapopu-lation is the component parasite community which
22
CHAPTER 2
Are there general laws in parasitecommunity ecology? The emergenceof spatial parasitology andepidemiologyJ.-F. Guégan,1 S. Morand,2 and R. Poulin3
1 GEMI, UMR IRD-CNRS 2724, Centre IRD de Montpellier, 911Avenue Agropolis BP 64501, 34394 Montpellier Cedex 5, France.
2 CBGP, UMR INRA-IRD-CIRAD-Agro.M., Campus Inter-national de Baillarguet, CS 30016, 34988 Montferrier sur Lez cédex,France.
3 Department of Zoology, University of Otago, PO Box 56,Dunedin, New Zealand.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 22

represents all of the infracommunities within agiven host population. Then the highest level ofparasite organization is the suprapopulation whichrepresents all individuals of a given parasitespecies within an ecosystem. Next, the parasitecompound community consists of all the parasitecommunities within an ecosystem (see Esch et al.1990). This creates at least a third-order scaling ofhabitat fragmentation for the parasites which has asignificant impact on the development of theoryregarding the evolution of populations and commun-ities of parasites and pathogens. Infrapopulationsand infracommunities may form many replicatesfrom one host to another, thus providing a remark-able opportunity for comparative analyses of thevariability of organizational patterns at severalhierarchical levels, so much more difficult toexplore for free-living organisms. Furthermore, thishierarchical organization means that larger-scaleprocesses may have a strong influence on localcommunity structure (see Poulin 1998a; Poulin et al.2000) and dynamics of parasites and microbes (seeGrenfell and Harwood 1997; Rohani et al. 1999),indicating that these larger-scale phenomena can-not be ignored anymore. For instance, recentadvances in epidemiology of childhood diseaseshave clearly shown the influence of spatial fluxeson local disease dynamics, recognizing structuralsimilarities between the processes of metapopu-lation biology and infection dynamics (Grenfell andHarwood 1997; Rohani et al. 1999).
Price (1990) in his contribution to the seminalbook by Esch, Bush, and Aho (1990) argued thatparasite community ecologists should take a lead-ing role in advancing areas of ecology with manyparasite studies being attractive complements forinvestigating some of the major questions in popu-lation and community ecology. Nearly one decadeand a half after, the intent of the present chapter is tosynthesize the more recent developments in para-site and microbial community ecology, and to assesscurrent perspectives regarding our knowledge ofthese communities.
To see where we are heading, consider a few sim-ple questions one could ask about a parasite ormicrobial community. What determines the num-ber of parasite species one host individual can
E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 23
harbour? Why are some parasite species extremelyrare when others are very common? What is thelocal population abundance of a widely distributedmacroparasite when compared to that of a rarespecies? These very basic questions of (parasite andinfectious disease) community ecology haveextraordinary little to do with small-scaleprocesses, but on the contrary need that we explorelarger-scale phenomena. Having described the dif-ferent macroscopic patterns, we then explore theconsequences of this research framework in popu-lation and community ecology of parasites andmicrobes. We opted in the present chapter to useexamples from both the microbial and parasito-logical community literature to illustrate the variousconcepts. We conclude by highlighting what thesefindings mean for further study of populationdynamics and community ecology of parasitic andinfectious disease in wildlife and humans.
2.2 Parasite community organisationand species coexistence
2.2.1 The emergence of spatial ecology ininfectious and parasitic diseases population and communities dynamics
As for mainstream population and communityecology (see Putman 1994; Begon et al. 1996; Weiherand Keddy 1999), parasite population and commun-ity ecology has concentrated on local processeswith an emphasis on local interactions betweenparasite species, and between these species andtheir host environment (see Kennedy 1975; Cheng1986; Esch and Fernandez 1993; Combes 1995; Bushet al. 2001). Another aspect is the strong researcheffort made over the past decades on very untidysmall-scale studies, which do not take a stronglyquantitative approach to issues such as spatial pat-terns in species richness at very large scales or pat-terns in species distributions (but see Price 1980;Rohde 1982; Poulin et al. 2000; this chapter). Forinstance, what determines the structure of the localcommunity of human infectious diseases in a givenplace of Western Africa has certainly as much to dowith large-scale, biogeographical processes as it hasto do with local conditions. This view is shared by
Rena-Ch02.qxd 8/20/04 9:58 PM Page 23

an increasing number of researchers interested inthe influence of climate variability on regional dis-ease dispersion and diffusion, for instance (Dobsonand Carper 1993; Rogers and Williams 1993; Hayet al. 1996; Patz et al. 1996; McMichael and Haines1997; Rapport et al. 1998; Epstein 1999; Rogers andRandolph 2000; Aron and Patz 2001).
2.2.2 Some definitions and basic conceptualframework
Many processes studied by parasite and microbialcommunity ecologists have clear linkages withlarger-scale, regional phenomena, but since studieshave focused too largely on small spatial scales theyare obviously unable to put the communities intoperspectives (see Lawton 2000; Poulin et al. 2000; thischapter). The assembly of parasites or microbialcommunities as for free-living ones is a multistage,multilayered process, and this forms a conceptuallyimportant framework on which to base further
research investigations. First, it starts at the top ofFig. 2.1 with the largest-scale pools of species. Theexistence of a global-scale pool of parasite andmicrobe species is entirely relevant for many organ-isms like in the case of crop pests, viruses, bacteria,and fungi of many aquatic organisms, or humaninfectious and parasitic diseases (see Rapport et al.2002). This notion is at present made highly relevantby the development of transcontinental transportsand economical exchanges between two distant bio-geographical regions, making the Earth today aglobal village for many pathogens and diseases (seePoulin 2003a, and the later Section 2.3.2.4‘Latitudinal gradient in species richness’). Theregional pool of parasite and microbe species, ormetacommunity, is a more conventional andaccepted notion in community ecology, and it existswithin a biogeographic region like a continent or asubcontinent. Then, understanding the origin of theparasite or microbe pool requires a knowledge of theevolutionary history of host–parasite associations, of
24 PA R A S I T I S M A N D E C O S YS T E M S
Infra-community
Component community
Compound community
Regional species pool
Continental species pool
Global species pool
Environmentalfilters
Dispersalfilters
Top-downeffects
(screen filters)
Bottom-upeffects
(invasive species,pandemia,…)
Historicalfilters
Figure 2.1 Schematic illustration of the main determinants of parasite or pathogen species richness in hosts. On the left: processes influencingspecies diversity are arranged into hierarchies in which different temporal and spatial factors may act. Any given level includes all lower levels, and itis included within all higher levels. On the right: the hierarchical scheme indicates that parasite or pathogen community assemblages at lower-spatial scales, for example, an individual host, are strongly dependent on upper scales (top-down effect on community richness), but the oppositesituation where lower-spatial scales may influence higher levels (bottom-up effect) is also possible.
Notes: Invasive species like crops or pandemia like HIV are illustrative of global impact of parasites and pathogens. Further research should reveal therespective roles played by ‘top-down’ and ‘bottom-up’ effects on community assemblages of parasites and microbes.
AQ: Pleasecheck theyear ofDobson andCarper 1993is ok?
Rena-Ch02.qxd 8/20/04 9:58 PM Page 24

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 25
the geographic isolation of the continent, of the linkages between pathogens and the biologicaldiversity present within the area, and so on (seeBrooks and McLennan 1991; Combes 1995; Poulin1998a). Local communities like those of macropara-sites in fish populations or microbes in human pop-ulations assemble themselves from this regional poolthrough a series of filters (see Fig. 2.1). Differences inhost population size and density or the spatialarrangement of host population habitat patches(high to low connectivity), for instance, may beresponsible for the persistence or the extinction ofparasite or pathogen populations moulding localcommunities (see Grenfell and Dobson 1995;Grenfell and Harwood 1997; Keeling and Grenfell1997, 2002; Rohani et al. 1999). At the individual hostlevel, infracommunities of parasites and pathogensagain assemble themselves from the local pool ofavailable species (see Fig. 2.1). If species can reach ahost they may still find the environment unsuitable,species interactions may also operate or constitutiveand induced defences against invasion can intervene.Many processes described here have also clear paral-lels in within-host infection since each host indi-vidual is composed of many different sites more orless connected with one another, and available, ornot, for parasite or pathogen establishment (seeHolt 1999). This framework shows that differentenvironmental filters work on all these commun-ities representing important steps in communityassembly and constitution (see Murray et al. 2002;Rapport et al. 2002).
Mainstream community ecologists have longdebated on the important steps in communityassembly working down from the larger scaleslargely dominated by regional, not local processes(see Ricklefs and Schluter 1993; Brown 1995;Rosenzweig 1995; Maurer 1999; Lawton 2000;Gaston and Blackburn 2000). This can be contrastedwith a more traditional approach in both parasito-logy and epidemiology through the study of localphenomena for understanding the structure anddynamics of parasitic or microbe assemblages.However the ‘top-down’ and ‘bottom-up’ paths areclearly complementary, and the recognition of theimportance of a regional or even global perspectivein parasite and microbe population dynamics and
community assembly theory would clearly benefitfrom more detailed attention than would be possiblefrom either approach alone. Recent studies on theimpacts of global environmental changes on disease population and communities dynamics (seeHarvell et al. 1999, 2002; McMichael 2001; Martensand McMichael 2002) provide several goodexamples of how largest-scale studies are of parti-cular relevance to both parasitology in wildlife andhuman epidemiology. It is also obvious that the‘context’ of the beginning of an infectious diseaseoutbreak transmitted from wildlife is clearly local(see for instance the cases of HIV), some havingdramatically increased in incidence and expandedin geographic range panglobally (see Hahn et al.2000; Daszak and Cunningham 2002).
As we know today from the study of complex hier-archical systems inspired by physics, both processesfrom the top to the bottom and from the bottom to thetop of Fig. 2.1 (see also Allen and Hoekstra 1992; Allenet al. 1993) are certainly acting as forces controllingparasitic and infectious disease community assembly.Recognizing that often the determinants of both hostanimal (or plant) and individual human health mayoccur at levels higher within the ecosystem hierarchyis thus one of the major tasks of modern parasitologyand epidemiology.
2.3 Emergent properties of parasite and infectious disease communities
2.3.1 On the search for regularities in parasiteand infectious disease community structure and processes
One of the principal advantages of this two-wayviewpoint of community assembly organisation isthat it takes a sufficiently distant view of parasito-logical and epidemiological systems that the idio-syncratic details disappear, and only the importantgeneralities remain (see Brown 1995; Rosenzweig1995; Gaston and Blackburn 2000 for application inmainstream ecology; Poulin et al. 2000 and this vol-ume for parasitological–epidemiological investi-gations). This may reveal general patterns, orregularities, that would otherwise have beenentirely neglected (see Morand and Poulin 1998;
Rena-Ch02.qxd 8/20/04 9:58 PM Page 25

Morand 2000; Morand and Guégan 2000a). Any scientific discipline must pass through a phasewhere the phenomena of interest are clearly andquantitatively identified (this chapter), and thenthe mechanisms underlying the observed patternsare explored and challenged with rigourous theor-etical and empirical testing. Specifically, the recentdevelopments in population dynamics of infectiousdiseases have clearly shown how useful generaliza-tions, but not at the level of local human commun-ities, might be indicative of significant regulation inspatial dynamics of those diseases (Ferguson et al.2003). Nevertheless, the impossibility of usingmanipulative experiments in natural systems (caseof wildlife diseases) and anthropogenic systems(case of epidemiology) means that it is often diffi-cult to retain a single hypothesis among competingalternative solutions to understand the mech-anisms that underlie the patterns (see Morand andGuégan 2000a for an illustration). Consequently,this inability to exploit manipulative experimenta-tion over large-scales have forced parasitologists,plant-associated insect ecologists, and epidemio-logists to use comparative approaches (see Aho andBush 1993; Cornell 1993; Lawton et al. 1993; Poulin1995a; Cornell and Karlson 1997; Morand andPoulin 1998; Choudhury and Dick 2000; Morandand Guégan 2000a; Guégan et al. 2001; Brändle andBrandl 2003; Nunn et al. 2003; Guernier et al. 2004)as macroecologists did before (Brown 1995;Rosenzweig 1995; Maurer 1999). The followingsections attempt to illustrate this using differentexamples from the literature, and to discuss theconsistency of the results with the various hypo-theses used to explain the observed patterns in thelight of recent advances in macroecology (seeGaston and Blackburn 2000), and comparativeanalysis in parasitology-epidemiology (see Poulinet al. 2000).
2.3.2 Parasite and infectious disease speciesrichness
Undoubtedly, parasites and other kinds of microbesand associated organisms like phytophagousinsects may represent more than half of the livingorganisms (Price 1980; de Meeüs et al. 1998;
Morand 2000; Poulin and Morand 2000; Curtis et al.2002; Brändle and Brandl 2001, 2003; Nee 2003),even if very few attempts have been made to rigor-ously quantify and delineate the differences in rich-ness between free-living organisms on the onehand and their associated organisms on the otherhand (but see Strong and Levin 1975; Strong et al.1985; Hillebrand et al. 2001; Guernier et al. 2004 andhereafter). Most, if not all, organisms are hosts forparasites, comprising helminths, arthropods, fungi,or microbes, and if the pioneer work by Guernierand colleagues is representative of other (host)species, the overall biodiversity on Earth may becurrently underestimated by more than an order ofmagnitude due to the unsuspected species diversityof parasites and other kinds of microorganisms (seeGuernier et al. 2004 and hereafter). Investigationsattempting to identify determinants of species rich-ness of parasites and other kinds of associatedorganisms are now well represented in the recentparasitological literature (Poulin 1995a; Morand2000; Poulin and Morand 2000; Brändle and Brandl2001; Guernier et al. 2004) much more than a decadeago, and more research is needed on different sym-biotic systems to examine the extent to which theobserved patterns are supported or undermined.Here we draw attention to the four most popularand striking ‘macroecological’ patterns in thespecies richness of assemblages of parasitic andother kinds of associated organisms, and we pro-mulgate how the search for consistency in commonpatterns, or not, from other symbiotic systems willcontribute to the emergence of a more rigorousresearch agenda in parasitology and epidemiology.
2.3.2.1 Species–area relationshipThe variation in species richness of parasites andother associated organisms is generally not random,but shows one regular pattern which is thespecies–area relationship (Simberloff and Moore1997; Poulin 1998a,b; Morand 2000), a now-classicalfactor explaining the number of species likely to befound at any site in mainstream ecology (Brown1995; Rosenzweig 1995; Gaston and Blackburn2000). Usually, widespread hosts tend to have moreparasite or infectious disease species than hostswith a more restricted geographical range because
26 PA R A S I T I S M A N D E C O S YS T E M S
Rena-Ch02.qxd 8/20/04 9:58 PM Page 26

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 27
the increase of host range may allow the host toencounter more species (Gregory 1990). This patternseems to hold both within host species (see Freeland1979; Marcogliese and Cone 1991; Goüy de Bellocqet al. 2003; Calvete et al. 2004) and across host species(see Poulin 1998a; Morand 2000; Brändle and Brandl2003 for a review). Alternatively, host body size maybe taken to represent area size for parasites (seeGuégan et al. 1992; Guégan and Hugueny 1994).Indeed, in interspecific comparisons among hostspecies, many positive relationships have beenreported between host body size and parasitespecies richness; however, the relationship is notuniversal, and there are many exceptions (seereviews in Poulin 1997; Morand 2000).
Still, the species–area relationship is generally astrong pattern in parasite diversity studies. Forinstance, the species–area relationship explainsmore than 50% of the total variation in species rich-ness of phytophagous insects (Kennedy andSouthwood 1984; Brändle and Brandl 2001), andaround 14–30% of the variance in species richnessof parasitic fungi (Strong and Levin 1975; Brändleand Brandl 2003). More interesting than the exist-ence of a species–area relationship for symbioticsystems is what does this pattern mean? Or, put inother words, which processes may be responsiblefor the relationship?
One problem with this approach is that thesearch for correlates of species richness across popu-lations of host species characterized by differentgeographical ranges (or size area for isolatedsystems) is often not independent of sampling effort(see Gregory 1990), and most studies have thus con-trolled for the effects of differential sampling ofhost species (Gregory 1990; Poulin 1998a). Whetherit is correct still remains to be determined (Guéganand Kennedy 1996). Sampling effort may bestrongly correlated with host geographical rangeand other biological factors which may covary withhost range, for example, host size, host nichebreadth, extinction/colonization, and temporaldynamics (see Gaston and Blackburn 2000), whichthen disappear after such a statistical control (seeGuégan and Kennedy 1996).
The paper by Goüy de Bellocq et al. (2003) showsthat the parasite species richness in the woodmouse,
Apodemus sylvaticus, on western Mediterraneanislands depends on the surface areas of the differentsurveyed localities (but see Dobson et al. 1992a,b forcontradictory results). Other factors, that is, parasitespecies life-history traits and host species diversity,were also important as determinants of helminthrichness and composition across isolated rodentpopulations. Fig. 2.2 illustrates the relationshipbetween parasite species richness and surface areafor the eight Mediterranean islands and threecontinental regions used in the study.
More interestingly, the authors demonstrate thatthe positive relationship observed between parasitespecies richness and surface area across wood-mouse populations is not the result of a randomprocess, but, on the contrary, it shows the existenceof order, that is, nestedness (see correspondingSection, 2.3.2.4) in the presence/absence matrix ofparasite species across different localities. Helminthparasites are organized according to a hierarchy ofspecies, also called nested species pattern (seeGuégan and Hugueny 1994), across surveyed areas:some parasite species are widespread across local-ities, and some others are uncommon and found inonly fewer areas (see Goüy de Bellocq et al. 2003).The richness and distribution of the differenthelminth species across areas depends not only onarea size as previously shown but also on the type
0.4
0.6
0.8
1
1.2
1.4
1.6
0 1 2 3 4 5 6
Surface area (log-km2)
Par
asite
spe
cies
ric
hnes
s(lo
g-sc
ale)
Figure 2.2 The relationship between log-species number and log-surface area for parasites of the wood mouse, Apodemus sylvaticus, ondifferent islands and regions from Europe.
Notes: The line of best fit is a linear function of the form y � 0.11x � 0.72, r2 � 0.68, p � 0.0017). The three points in the upper rightside of the diagram are for Continental Europe, Spain, and Italy. Otherpoints are for western Mediterranean islands, that is, Mallorca, Menorca,Formentera, Ibiza, Corsica, Port-Cros, Porquerolles, and Sicila.Source: Redrawn from Goüy de Bellocq et al. (2003).
AQ: Pleasecheck theinsertion of sectionnumber is ok.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 27

of parasite life cycle and host mammal speciesdiversity. The explanations given by the authors arethat larger areas may sustain larger host popu-lations, an important parameter in epidemiologywhich determines the host resource needed for aparasitic or infectious disease agent to persist (seeGrenfell and Harwood 1997; Keeling 1997), andthus may favor the existence of a higher parasitediversity (see Morand and Guégan 2000b). Then thedifference observed in both the parasite speciesrichness and composition across areas may be dueto the fact that a parasite with a direct life cycle mayhave more chances to succeed in the colonization ofa new host population compared with a parasitewith an indirect life cycle which absolutely needs tofind a suitable intermediate or definitive hosts tocomplete its cycle. Finally a locality with high hostspecies diversity may be more favourable for para-sitic or disease persistence, for example parasiteswith a complex life cycle finding more definitivehosts species to achieve their development.
A reconsideration of the study by Goüy deBellocq et al. (2003) using Generalised LinearModels (see Wilson and Grenfell 1997; Venables andRipley 1999) instead of simple regressions indicatesthat the data are strongly flawed by sampling bias,a fact that makes previous findings questionable.The new results (see Table 2.1) show that helminthcomponent community richness variation acrossareas is strongly dependent on both sampling effortexpressed as the host sample size per area (F value � 345.97, p � 0.00034) and the interactionbetween sampling effort and surface area (F value � 30.35, p � 0.0118), the largest areas beingless sampled, on average, than smaller areas, withall other parameters kept constant in the model.Mammal host species diversity is then just mar-ginally significant in the multivariate analysis (seeTable 2.1). Surface area does not appear to be signi-ficant anymore. A stepwise elimination procedureusing the Akaike criterion yielded similar results.
This new result indicates that sampling effort mayexert strong bias in estimation of parasite species rich-ness variation across different areas, notably in thecase of the largest areas where rare parasite speciesmay be missed during parasitological investigations.This point has been made before (Gregory 1990;
Poulin 1998b), and sampling effort must be taken intoaccount in any investigations of species–arearelationships using parasite data. We also stronglyrecommend the use of multivariate analyses to takeinto account perverse effects exerted by samplingbias on statistics instead of a priori regressing speciesrichness data against sampling and the use of residu-als since sampling effort may also covary with otherindependent variables under study.
2.3.2.2 Species richness–isolation relationshipMore generally, species diversity is dependent onthe fragmentation and isolation of habitats(Whittaker 1998). Fragmentation and isolation havepromoted organism speciation and the build-up ofendemic faunas on Earth (Brown 1995). Isolationand fragmentation are, of course, one part of themany factors promoting species diversity and com-position (see Brown 1995; Rosenzweig 1995; Gastonand Blackburn 2000). Most organisms, includingparasitic and infectious diseases in hosts (this chap-ter), exhibit patterns of similarity in compositionand richness depending on geographic distanceand isolation (see Poulin and Morand 1999;Morand and Guégan 2000b).
Perhaps one of the most important lesson to belearned from recent studies in host–parasite systems
28 PA R A S I T I S M A N D E C O S YS T E M S
Table 2.1 Summary of Generalized Linear Model with a gaussianerror structure for explaining the parasite species richness variation inwoodmouse across 11 different areas
Deviance Resid. df Resid. dev F value p (F)
Null 678.73Sampling size 538.77 9 139.96 345.97 0.00034Host diversity 22.12 8 117.84 14.21 0.03269Surface area 3.33 7 114.51 2.14 0.23978Sampling � Host 6.33 6 108.18 4.06 0.13721Sampling � Area 47.26 5 60.92 30.35 0.01178Host � Area 40.53 4 20.39 26.02 0.01457Sampling � Host�Area 15.72 3 4.67 10.09 0.05021
Notes: Resid. df and Resid. dev are the residual degree of freedom and theresidual deviance at each step of the procedure, respectively; p(F) is theprobability statistics associated with the F test; Host diversity is the hostmammal species diversity per unit area; Sampling � Host is the two-wayinteraction term between Sample size and surface area, and so on (see textfor explanation). The rank of introduction of terms in the successive modelsdid not alter the main results as illustrated here.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 28

is the demonstration by Poulin and Morand (1999)that the geographical distance between componentcommunities of parasites in freshwater fish is oftenthe best, most general explanation of similarity inparasite species composition and to a lesser extent,of species richness across localities. Using multi-variate analysis based on permutation methods(see Legendre et al. 1994), these authors concludethat patterns of parasite species composition acrossdistinct isolated areas (i.e. distinct lakes in theirstudy) strongly depend on the distances that separ-ate the different localities, shorter geographicaldistances between isolated areas being associatedwith a greater similarity in parasite compositionbetween them, and nearby lakes harbouring num-bers of parasite species more similar than those ofdistant localities. Put in other words, there is inthese parasitological data a tendency for speciescomposition and richness to be autocorrelated overspace. As mentioned by Poulin and Morand (1999),it might be more accurate to say that it is the isola-tion of a given locality within the network of patchareas that here matters instead of geographical dis-tances among sites that roughly approximate thisisolation. It is possible to have exceptions to thispattern, and many other factors may promote sim-ilarity between close localities in the compositionand richness of parasite communities (see Kennedyet al. 1991; Hartvigsen and Kennedy 1993; Poulin1998a). Nevertheless, the authors strongly suggestto consider the effect of geographical distance as agood index of isolation in further comparativeanalyses of parasite communities, and it shouldtherefore become a basic requirement to control forthe confounding and often important effect of geo-graphical distance on the determinants of speciescomposition and richness in parasite componentcommunities.
Recently, Poulin (2003b) has shown that the influ-ence of geographical distance on the similaritybetween parasite communities may follow a regu-lar pattern. In the majority of parasite communitiesof fish and mammal hosts, the similarity in thespecies composition of communities decays expo-nentially with increasing geographical distancebetween localities (Poulin 2003b). Exponential ratesof decay in similarity have also been reported for
plant communities (Nekola and White 1999), andfurther emphasize the importance of geographicalisolation.
One illustrative example of the effect of geo-graphical distance and isolation on parasitic andinfectious disease communities is that of oceanicislands. Typically islands have fewer species perunit area than the mainland, and this distinction ismore marked the smaller the island and the farthestit is from a continental source (Rosenzweig 1995;Whittaker 1995). Very few studies have investi-gated parasite species community richness andcomposition on islands (see Kennedy 1978; Mas-Coma and Feliu 1984; Kennedy et al. 1986a; Dobson1988a; Dobson et al. 1992a; Miquel et al. 1996; Goüyde Bellocq et al. 2002, 2003). The main conclusionsreached by Dobson et al. (1992a) concerning para-site species richness and composition in Anolislizards from northern Lesser Antilles islands arethat they show a relatively depauperate parasitecommunity when compared with lizards sampledon the larger Carribean islands, for example, Cuba,or on continental areas, and that these differencesare associated with the life history attributes of thedifferent parasite species in the assemblages. Onthe whole, Goüy de Bellocq et al. (2003) reach thesame conclusions (see above), but they were unableto more formally characterize an effect of geo-graphical distance and isolation in-betweenMediterranean islands on parasite species richnessand composition in the wood mouse. Examples ofstudies of pathogen communities and species com-position in animals and humans on islands are evenmore scarce. Collares-Pereira et al. (1997) provide thefirst epidemiological data on pathogenic leptospiresserovars diversity in insectivore and rodent speciesin the Azores archipelago, and they conclude to alow serovars diversity of three within this group ofislands out of a total of nineteen serogroups morelargely represented over the world. Surprisingly,there are only few studies that have quantifiedpathogen species richness and composition onislands for human communities in a way similar towhat is traditionally done in community ecology.Based on unpublished data from one of us (Guéganand Guernier, unpubl. data), we show here that on atotal set of 197 different countries all over the world,
E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 29
AQ: Pleasecheck theyear ofWhittaker1995.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 29

the species richness of pathogens including viruses,bacteria, fungi, protozoa, and helminths is loweston the 73 islands compared with the mainlandcountries (see, Table 2.2). Even after controlling forconfounding effects that may be exerted by factorslike the socio-economical power, the population sizein number of inhabitants or the latitudinal positionof the country, the island factor still explains 24.1%of the total variation in pathogen species diversityacross areas. This result on pathogen species assem-blages in humans is indicative that isolation and/ordistance from a continent may be highly responsiblefor lower species richness in those localities. Assuggested by the work of Goüy de Bellocq et al.(2003), islands sample only from the dispersive por-tion of the mainland pool. This effect must, ofcourse, be distinguished from area size since thehuman pathogen study (see Table 2.2) has kept itseffect constant in a multivariate analysis. Present-day distances between isolated islands and aregional biogeographic pool, even if they do notstrictly reflect distances at the time of diseasecolonization, may however provide a rough indexof isolation.
From the present human diseases data set, inform-ation concerning the Caribbean islands and thesurrounding continental countries from Northern,Central, and Latin America representing themainland, it is informative to see that remoteislands from the Gulf of Mexico are poorest inpathogen species when compared to what isobserved on the continent and even on close islandslike Trinidad & Tobago, for instance (see Fig. 2.3).When considering the confounding effects exertedby covariate factors (notably the economic power ofa nation) on disease species richness across locali-ties, the influence of distance still remains (statisti-cal data not illustrated): distance from a continentalregional pool (49.1% of the total variationexplained; p � 0.00001), the total area size of islands(28.9% of the total variation explained, p � 0.00001),and to a lesser extent human community size (5.9%of the total variation explained, p � 0.028) are the bestpredictors of pathogen species richness in humanpopulations among the nineteen Caribbean islandsand the fifteen surrounding continental countries.These findings based on a study of communityassemblage of human diseases are in accordance
30 PA R A S I T I S M A N D E C O S YS T E M S
Table 2.2 Summary of Generalized Linear Model with a poissonianerror structure and a log-link for explaining the species richnessvariation in human infectious diseases across a set of 197 differentcontinental (124) or isolated (73) countries
Deviance Resid. df Resid. dev p (�2)
Null 265.56Island 64.07 196 201.49 0.00000GNP 2.63 195 198.86 0.10466Population size 40.32 194 158.54 0.00000Surface area 19.60 193 138.94 0.00001Latitude 8.80 192 130.14 0.00301Island � Surface area 2.98 191 127.16 0.08404
Notes: Dispersion parameter in the model is 0.67. Island is coded 1 andcontinental area is coded 0; GNP is the Gross National Product per country(in US $) to control for its effect on final statistics; Population size is thenumber of inhabitants; Latitude is the geographical position of each countryin degrees and minutes; see also Table 2.1; p (�2) is the probability statisticsassociated to �2 test. Models were built with independent variables andtheir two-way and three-way interaction terms. The rank of introduction ofterms in the successive models did not alter the main results as illustrated here.
4.9
5
5.1
5.2
5.3
5.4
5.5
0 2 4 6 8
1
2
3456
7
8
9
Distance from a continental source (in log)
Pat
hoge
n sp
ecie
sric
hnes
s (in
log)
Figure 2.3 The relationship between log-species number and log-distance (in km2) from the continent for human infectious diseasesin different Caribbean islands (n � 19) and surrounding continentalcountries (n � 15) from Americas.
Notes: Codes are 1 : USA; 2 : Brazil; 3 : Venezuela and Mexico;4 : Colombia and Panama; 5 : Surinam, Guyana, Honduras, Guatemala,and Costa Rica; 6 : French Guiana, San Salvador, Trinidad & Tobago,Puerto Rico, Nicaragua, Haiti, Dominican Republic, Belize; 7 : Guadeloupe,Cuba, Martinique, Jamaïque; 8 : Dutch Antilla, 9 : Aruba, Caïman Islands,Montserrat, Grenada, St Kitts & Neville, Antigua & Barbuda, Barbados,St Vincent and Dominica.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 30

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 31
with studies on population dynamics of infectiousdiseases on islands (see Black 1966). According toPrice (1990), as local host abundance increases, sothe effective population size for maintaining parasitepopulations increases, resulting in more parasitespecies being maintained in larger communities.Thus species dynamics of infectious diseases mightbe in many ways analogous to population dynam-ics (e.g. critical community size threshold), (seeGrenfell and Harwood 1997; Keeling 1997; Broutinet al. 2004). There are obviously similarities betweenspecies and population maintenance and dynamics(see Brown 1995), and all these points need to bedeveloped in further research on infectious diseasesin wildlife and humans as we learn more about thepatterns of variation in infectious disease speciesrichness with respect to island size, isolation, andhost community size. The above findings do notstipulate that modern events, and more particularlytranscontinental exchanges, do not influence infec-tious disease dispersal and maintenance in hetero-geneous environments, but they strongly suggestthat we need to explore the impact of spatial hetero-geneity on the course of infectious disease speciesdynamics, and the importance of these variables,for example, area size, isolation, community size,to better grasp the abundance, distribution, andidentity of pathogen species within local habitats.
A major outcome of fauna (or flora) isolation isendemism; thus old and/or remote islands tend togenerally have a large degree of endemism (seeWhittaker 1998). The term endemism refers to therestricted ranges of taxa in biogeography, and it isused differently in epidemiology–parasitology. Werefer here to the former definition. Little is knownabout parasite or infectious disease communityassemblages in endemic hosts, and Morandand Guégan (2000b) have concluded with somepredictions based on both empirical studies andmathematical modelling that hot spots of (host)endemism are also the foci for a large diversity ofendemic parasites and pathogens, and thatrestricted areas and/or low host community sizesare associated with a decrease in parasite orpathogen species numbers (see above). Regardingthe existence of endemic parasites or pathogens, we
can only speculate on their existence on Earth, andthey are probably legions. Many parasitologicalinvestigations have focused their efforts on theimpacts of exotic pathogens and pests on nativehost species, but an important advance in futureresearch should be the recognition of endemic parasites and pathogens, and the role they mayplay in maintaining and regulating biodiversity andecosystem dynamics. Recent research on emergentpathogens might shed light on their importance innature (see Aguirre et al. 2002).
2.3.2.3 Local–regional richness relationshipMuch parasitological literature on species diversitypatterns has been devoted to local mechanismswhereas in recent years large-scale processes havebeen regarded as important determinants of thespecies richness of local communities in free-livingorganisms (see Lawton 1999; Gaston and Blackburn2000). Implicit in many ecological studies is theimportant recognition that regional and historicalprocesses may profoundly affect local communitystructure (Brown 1995; Rosenzweig 1995; Lawton1999, 2000). Questions of spatial scale have beenaddressed only very recently in parasite commun-ity ecology (Price 1980; Aho 1990; Aho and Bush1993; Kennedy and Bush 1994; Kennedy andGuégan 1994; Barker et al. 1996) probably becausetraditional parasitology has been too medicallyorientated over a long time with a major focus onvery fine-scale studies.
Simply because every parasitic, parasitoid, oreven microbe species cannot be present every-where, we do not expect every species occurringwithin the regional pool to affect the composition ofevery local community. A central method used forthe recognition of the importance of regional andlocal processes is the regression of regional speciesrichness against local species richness plots (seeLawton 1999; Srivastava 1999; Hillebrand andBlenckner 2002), and several contributions havetried to disentangle the regional and local con-straints in free-living communities (see Srivastava1999; Shurin and Allen 2001; Hillebrand andBlenckner 2002, for recent reviews). The testrequires estimates of species richness for a given
Rena-Ch02.qxd 8/20/04 9:58 PM Page 31

group of organisms at both local and regionalspatial scales and a statistical evaluation of theirrelationship. When local richness is regressedagainst regional richness and the relationship islinear, the communities are unsaturated and are saidto exhibit ‘proportional sampling’ of the regionalspecies pools. If the relationship is a somewhatcurvilinear function, the possibility of saturationmay then arise (see Fig. 2.4). Although this typeof analysis appears to be straightforward, severalpitfalls have been discussed elsewhere (seeCreswell et al. 1995; Srivastava 1999; Shurin et al.2000). A major question is the definition of whatexactly regional and local richnesses mean forparasite communities, and the way their measure-ments are best comprehended (Kennedy andGuégan 1994; Barker et al. 1996). Usually, regional(Kennedy and Guégan 1994) or continental (Ahoand Bush 1993) parasite species richnesses, that isto say spatial scales reflecting a naturally occurringhierarchy from which local parasite communitiesmay be drawn, have been used to representregional pools. Morand et al. (1999) used another
estimate of regional species richness defined as thecomponent community richness, but the authorswere faced to the problem of defining exactly thespatial hierarchy at which processes may operate inopen marine systems. The measure of local parasitespecies richness adopted sometimes is the mean ormaximum number of parasite species in the para-site component communities known to the authors(see Aho and Bush 1993; Kennedy and Guégan1994), but a more correct definition of local speciesrichness seems to be the mean (Kennedy andGuégan 1994; Morand et al. 1999) or even maximum(Poulin 1996a, 1997; Calvete et al., in press) infra-community parasite species richness.
Generally, all the studies agree on an importantinfluence of both regional and local factors, buttheir relative importance may differ between cate-gories of organisms. Concerning parasite, para-sitoid, and microbe species communities, thediversity of species in local (at the host populationlevel) assemblages is intuitively regulated both bylocal (e.g. interspecific competition, habitat hetero-geneity) and by regional factors (e.g. evolution,
32 PA R A S I T I S M A N D E C O S YS T E M S
Boundary line
Type I
Type II
Regional species richness
Loca
l spe
cies
ric
hnes
s
Regional species richness
Loca
l spe
cies
ric
hnes
s
Regional species richness
Loca
l spe
cies
ric
hnes
s
(a)(b) (c)
Figure 2.4 The relationship between regional and local species richness for parasite, pest and pathogen organisms. (a) Because of the hierarchicalprocesses illustrated on Fig. 2.1, every parasite, pest or pathogen species present within the regional pool is unlikely to occur everywhere, which thusdetermines a boundary line never reached for which y equals x. Local species richness both at the infracommunity and component community scalesare usually less than regional richness, and two kinds of relationships may then exist. A type I curve indicates proportional sampling in which localspecies richness increases linearly with regional richness. A type II curve saturates with local species richness above a threshold for higher regionalrichness. In nature, parasite, pest or pathogen community assemblages lie anywhere between type I and type II systems (see Cornell and Karlson1997). (b) Example where the richest regional (component) communities are associated with the richest infracommunities and vice versa, suggestiveof no saturation in parasite species. Helminth parasites of introduced freshwater fish in the British isles (Guégan and Kennedy 1993) or of naturalpopulations of partridges in Spain (Calvete et al. 2003) are clear examples of unsaturated systems. (c) Examples where local species richness may befixed by internal constraints indicative of saturation in local species communities. Intestinal helminth infracommunities (see Aho 1990; Kennedy andGuégan 1994; Calvete et al. in press) generally are examples where saturation in parasite species may occur (see also text for contradictory results).
AQ: Pleaseupdate thereferenceCalvete.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 32

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 33
migration, history). The studies published so far onfree-living organisms have stressed the prevalenceof type I communities (see Fig. 2.4) interpreted asan indication of unsaturation of local assemblageswith generally weak or no effects of local interactionson species richness. Analyses of local to regionalspecies richness performed on host-associatedorganisms like parasites or parasitoids (Cornell1985; Aho 1990; Bush 1990; Hawkins and Compton1992; Aho and Bush 1993; Lawton et al. 1993;Dawah et al. 1995; Kennedy and Guégan 1994, 1996;Poulin 1996a, 1997; Morand et al. 1999; Frenzel andBrandl 2000; Calvete et al. in press) have, on thecontrary, shown the existence of both type I and IIcommunities (see Fig. 2.4). Particularly, the com-monness of type II communities in host–parasitesystems, and more specifically for helminths, maybe illustrative of limiting factors shaping these localcommunities. In addition to the occurrence ofsaturated assemblages shown in parasite systemscompared to other organisms, the three clearestresults that emerge from the publishedparasitological literature available today may besummarized as follows.
First, most studies on herbivorous insects (seeDawah et al. 1995; Frenzel and Brandl 2000) and fishectoparasites, that is, helminths and copepods (seeMorand et al. 1999), have identified that emptyniches are common and that local communities areunsaturated. As such, many natural enemy com-munities are subject to strong regional influencesthen providing opportunities for new invasivespecies to become established. The opposite can beobserved for many studies on internal parasites likeintestinal helminths of fish (Kennedy and Guégan1994), of amphibians and reptiles (Aho 1990), andof birds (Bush 1990; Calvete et al. in press) wherelocal forces may contribute to parasite communitystructure. Poulin (1996a, 1997) showed, on the con-trary, that for 31 intestinal helminth communities inbird hosts and 37 in mammal hosts the relationshipbetween the maximum infracommunity richnessand component community species richness waslinear, indicating the absence of species saturationand the availability of vacant niches in organismsaccepted to generally have species-rich helminth
communities (see Bush and Holmes 1986a,b; Stockand Holmes 1988). The contrasting results may inpart be due to the fact that Poulin’s (1996a, 1997)analysis included different host species, whereasmany of the studies that found a curvilinear rela-tionship between infracommunity richness andcomponent community richness included only dif-ferent host populations of the same host species(e.g. Kennedy and Guégan 1996; Calvete et al. inpress).
Then, second, the only intestinal helminth com-munities which exhibit unsaturated assemblagesare those occurring in introduced fish species in theBritish Isles, that consist of non native fishes nothaving had enough time to accumulate sufficienthelminth species from the native pool (Guégan andKennedy 1993) to develop ecologically interactivecommunities (Kennedy and Guégan 1994). A recentstudy by Torchin et al. (2003) which compared theparasite species richness between introduced andnative populations for 26 host species of molluscs,crustaceans, fishes, birds, mammals, amphibians,and reptiles also confirmed the reduced parasitiza-tion of introduced organisms suggestive of anabsence of saturation in those parasite communities(see Figs. 2(c) and (d)).
Third, major advances in our understanding ofsaturation versus non saturation of local parasitecommunities have been made in recent years.Cornell and Karlson (1997) and Srivastava (1999)drew attention to the necessity of the demonstra-tion of other lines of evidences of niche and habitatrelationships combined with information on localversus regional relationships. In particular, Rohde(1998) using randomization procedures highlightedthe many scenarios in which a curvilinear local toregional relationship might be generated withoutrequiring the necessity of species saturation. Usinga comprehensive survey of marine fish ectoparasitecommunities, Morand et al. (1999) examined theeffects of interspecific aggregation on the level ofintraspecific aggregation in infracommunities, andthey demonstrated that interspecific interactionswere reduced relative to intraspecific interactionsthus facilitating species coexistence in rich commun-ities (see Tokeshi 1999 for further details on species
Rena-Ch02.qxd 8/20/04 9:58 PM Page 33

coexistence). This pattern was highly coincidentalwith a positive linear relationship between infra-community species richness and total parasitespecies richness obtained after controlling for theconfounding effect exerted by phylogeny, indica-tive of no saturation in ectoparasite communities ofmarine fish (see Rohde 1991, 1998). In a recentstudy, Calvete et al. (in press) showed the existenceof a curvilinear relationship between local andregional species richnesses of intestinal helminthinfracommunities for eight populations of the red-legged partridge in Spain, even after checkingfor the confounding effect of geographical distanceamong localities on species richness calculations.Interestingly, this finding was confirmed by ademonstration of negative interspecific associ-ations for the helminth species community, espe-cially between cestodes and other helminthsparasitizing the bird intestines.
All these results illustrate a number of importantissues about the understanding of local–regionalrichness relationships in parasite or microbe com-munity assemblages. First, demonstrating theeffects of saturation, or not, in infracommunityassemblages requires that we simultaneously useadditional investigations of interspecific interact-ions, or that published examples of the types ofinteractions exist, to test for the possible existenceof interspecific competition. Neither of the two pat-terns for community assemblage organization, thatis, the local to regional richness relationship and thedemonstration of interspecific competition, is con-clusive on its own. Interestingly, conclusions aboutthe degree of concordance in the saturation of localcommunities, or not, between two or more methodsmay yield generality, but most of studies to datehave only considered one option to test for theshape of interspecific relationships (but see Calveteet al. in press). Second, the study of interspecificcompetition in local communities of parasites andpathogens has shaped the development of ourunderstanding of species interactions, that is,importance of local processes. Thus, considerationof local to regional richness relationships in parasiteor pathogen communities has lead to more atten-tion being paid to the importance of large-scale
patterns in parasitology and epidemiology. Third,one area in which the combination of these twomethods should be fruitful is in the connectionthat might exist with the density of parasitesor microbes. Indeed, if competition is importantwithin parasite or microbe communities, onewould expect to see density compensation in thosecommunities with few species. If completecompensation occurs, there should be no relationbetween parasite density or biomass and localspecies richness, while if there was no density com-pensation a linear trend would thus be expected(see Oberdorff et al. 1998; Griffiths 1999 for taxo-nomic groups others than parasites). Furthermore,the linkages between interspecific competition,(un)saturation and density compensation in para-site or microorganism community assemblages will require more research from community ecolo-gists, parasitologists and epidemiologists. Notably,these issues should be highly relevant in the field ofveterinary and medical sciences since any altera-tion of local habitats (from the point of view of oneparasite species, for example, one intestine) andother disturbances exerted by humans (e.g. the useof drugs like helminthicides or antibiotics) shouldreduce parasite or pathogen populations from timeto time, making ways for more resistant species oraliens to increase or to invade. The idea of satura-tion predicts that an invasive species (like a crop oran emerging virus) should not invade an infracom-munity in individual hosts, or should do so onlywith the consequence of excluding a resident mem-ber species, that is, density compensation by newindividual invaders. The study of interconnec-tedness between these patterns and our effortsat understanding the processes behind will require judicious choices of both host and parasiteor pathogen taxa, and at different levels of spatio-temporal organization. As discussed before, patterns of within-host microbial species richnesswill also likely profit from a greater considerationof dynamical processes in patchy and discontinu-ously distributed environments, microbial persist-ence, and abundance in a particular tissue-habitatbeing influenced in several ways by biogeographical-like processes within host individuals.
34 PA R A S I T I S M A N D E C O S YS T E M S
Rena-Ch02.qxd 8/20/04 9:58 PM Page 34

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 35
2.3.2.4 Latitudinal gradient in species richnessLatitudinal gradients in species richness of free-living organisms are one of the most consistentlarge-scale trends that we can observe in nature,and one of the best documented patterns in theecological literature (see Hawkins et al. 2003 for arecent review), but there are still exceptions (Brown1995; Rosenzweig 1995; Gaston and Blackburn2000). This pattern does hold not only for thehosts as a whole, but also for some parasitic andinfectious disease organisms (Rohde 1992; Poulinand Rohde 1997; Rohde and Heap 1998; Calveteet al. 2003; Guernier et al. 2004; but see Poulin 2001;Poulin and Mouritsen 2003), which means thatit could be a simple consequence of the observedlatitudinal cline in host species diversity.Parasitological or epidemiological investigationson large spatial scales are rare, and there isundoubtedly a need for more comparative studies.Expectations of rich low-latitude parasite commu-nities have been suggested by some authors(Kennedy 1995; Salgado-Maldonado and Kennedy1997) while it is intuitive from other studies (Poulinand Guégan 2000; Guégan et al. 2001) that a latitu-dinal richness gradient exists for fish ectopara-site and human infectious disease communities,respectively.
Across 80 localities from 16 Spanish provinces,there is a marked cline in helminth species richnessand composition in the red-legged partridge(Alectoris rufa L.) (Calvete et al. 2003). The highestlevels of helminth richness are encountered insouthern provinces of Spain (e.g. Badajoz, Huelva)and the lowest in northern ones (e.g. Alava BurgosSantander, Alava Navarra) (Calvete et al. 2003).Statistical analyses controlled for the effect of hostage, sex, body condition, and time at which thestudy was carried out, and thus it is unlikely thatthese variables contributed to the spatial variationin helminth distribution and species richness acrossthe study area. The cline in parasite species richnessand composition across Spain was mirrored by themeasures obtained for both infra- and componentcommunities levels, indicating that the poorestcomponent communities in the north correlatedwell with the poorest infracommunities. In the
north, parasite communities were characterized byhaving one, that is, Dicrocoelium sp., or a few dom-inant widespread species. In contrast, in southernprovinces, helminth communities were morespecies-rich with several codominant helminthspecies, Raillietina tetragona, Subulura suctoria,Cheilospirura gruweli (see Calvete et al. 2003 forfurther details). The answer to why there are morehelminth species in the red-legged partridge insouthern Spanish provinces than in the north is thatthe pattern may be related to variation in definitivehost densities and to the distribution and diversityof intermediate hosts. According to Calvete et al.(2003), red-legged partridge populations are usu-ally denser in the centre and south of Spain whichrepresent the core-area of their range. This findingwould tend to suggest that high densities of defini-tive hosts might be associated with a greaterabundance of helminths and greater helminthspecies richness as a whole. In addition, and thissecond hypothesis is not mutually exclusive of thedefinitive-host density hypothesis, a greater abund-ance or diversity of arthropod intermediate hosts inthe centre and south of Spain might result in theexposure of partridges to a wider variety of poten-tial parasite species (see Poulin 1995a). Calvete et al.(2003) argued, based on correlation statistics usingfactorial scores from multivariate analysis, that thenorth–south abundance and richness variation ofhelminths in partridges might be to a large extentdetermined by higher temperature in southernregions of Spain, environmental conditions causingan increase in the survival or activity of intermedi-ate forms of parasites with life-history stages out-side their definitive hosts. These results suggestthat variation in the distribution of helminthsin partridge hosts are probably associated withvariations in the distribution of their definitiveand intermediate hosts and the local ecologicalconditions that may act on these host–parasite relationships.
One recent study (see Guernier et al. 2004) goesone step further in the explanation of the existenceof a latitudinal gradient of species richness forpathogen species in human populations. Compilingdata on parasitic and infectious diseases for a total
Rena-Ch02.qxd 8/20/04 9:58 PM Page 35
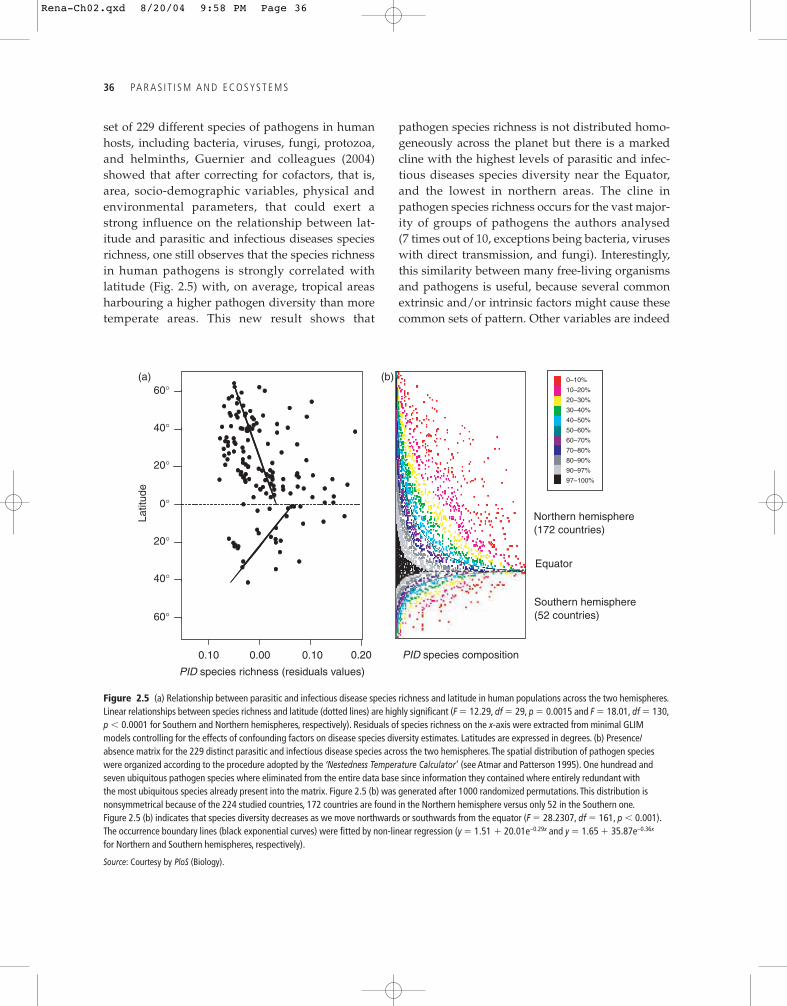
set of 229 different species of pathogens in humanhosts, including bacteria, viruses, fungi, protozoa,and helminths, Guernier and colleagues (2004)showed that after correcting for cofactors, that is,area, socio-demographic variables, physical andenvironmental parameters, that could exert astrong influence on the relationship between lat-itude and parasitic and infectious diseases speciesrichness, one still observes that the species richnessin human pathogens is strongly correlated withlatitude (Fig. 2.5) with, on average, tropical areasharbouring a higher pathogen diversity than moretemperate areas. This new result shows that
pathogen species richness is not distributed homo-geneously across the planet but there is a markedcline with the highest levels of parasitic and infec-tious diseases species diversity near the Equator,and the lowest in northern areas. The cline inpathogen species richness occurs for the vast major-ity of groups of pathogens the authors analysed (7 times out of 10, exceptions being bacteria, viruseswith direct transmission, and fungi). Interestingly,this similarity between many free-living organismsand pathogens is useful, because several commonextrinsic and/or intrinsic factors might cause thesecommon sets of pattern. Other variables are indeed
36 PA R A S I T I S M A N D E C O S YS T E M S
4060
4060
(a) (b)
0.10 0.00 0.10 0.20
60°
40°
20°
0°
20°
40°
60°
Latit
ude
PID species richness (residuals values)
Northern hemisphere(172 countries)
Southern hemisphere(52 countries)
Equator
PID species composition
0–10%
10–20%
20–30%
30–40%
40–50%
50–60%
60–70%
70–80%
80–90%
90–97%
97–100%
Figure 2.5 (a) Relationship between parasitic and infectious disease species richness and latitude in human populations across the two hemispheres.Linear relationships between species richness and latitude (dotted lines) are highly significant (F � 12.29, df � 29, p � 0.0015 and F � 18.01, df � 130,p � 0.0001 for Southern and Northern hemispheres, respectively). Residuals of species richness on the x-axis were extracted from minimal GLIMmodels controlling for the effects of confounding factors on disease species diversity estimates. Latitudes are expressed in degrees. (b) Presence/absence matrix for the 229 distinct parasitic and infectious disease species across the two hemispheres. The spatial distribution of pathogen specieswere organized according to the procedure adopted by the ‘Nestedness Temperature Calculator’ (see Atmar and Patterson 1995). One hundread andseven ubiquitous pathogen species where eliminated from the entire data base since information they contained where entirely redundant withthe most ubiquitous species already present into the matrix. Figure 2.5 (b) was generated after 1000 randomized permutations. This distribution isnonsymmetrical because of the 224 studied countries, 172 countries are found in the Northern hemisphere versus only 52 in the Southern one.Figure 2.5 (b) indicates that species diversity decreases as we move northwards or southwards from the equator (F � 28.2307, df � 161, p � 0.001).The occurrence boundary lines (black exponential curves) were fitted by non-linear regression (y � 1.51 � 20.01e–0.29x and y � 1.65 � 35.87e–0.36x
for Northern and Southern hemispheres, respectively).
Source: Courtesy by PloS (Biology).
Rena-Ch02.qxd 8/20/04 9:58 PM Page 36

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 37
important in explaining global-scale patterns ofhuman pathogens (e.g. modernization, urbaniza-tion, or impoverishment, especially in developingcountries), but the fact that the authors consideredsuch effects in their multivariate analyses tendsto indicate that biogeographical forces are alsoimportant indeed in shaping the distribution and abundance of pathogen species. These results thuschallenge the conventional wisdom that socio-economic conditions are of preponderant importancein controlling or eradicating diseases.
Over the last three decades, the number ofhypotheses advanced to explain the latitudinalgradient has increased from six (Pianka 1966) tonearly thirty, proffered by Rohde (1992). Some ofthese include spatial heterogeneity and patchiness, competition, predation, parasitism, mutualism,area, environmental stability, productivity, season-ality, solar energy (see Rohde 1992 for a generaldescription). In reality, several factors may beacting in concert or in series, and among thenumerous explanations given it is possible tosubstantially narrow the list of the most plausibleexplanations since many factors may be entirelyredundant or untestable (Hawkins et al. 2003). Forinstance, Gaston and Blackburn (2000) listed onlythree plausible explanations for latitudinal richnessgradients: area, energy, and time. They discardedthe possibility that random location of speciesmight be responsible for the latitudinal speciesgradient based on the absence of formal evidence.Area has often been cited as a simple hypothesis(see section on species–area relationships) toexplain that the tropics, which really harbour thehighest richness for many groups of organisms,also have the largest terrestrial surface area, that is,the geographical area hypothesis (see Rosenzweig1995). This explanation may not be plausible whenconsidering the latitudinal gradient of pathogenspecies in human populations as observed byGuernier and colleagues since both the surface areaand continental mass effects have been taken intoaccount in multivariate analysis.
Furthermore, and probably more important thanthe existence of a latitudinal gradient in speciesrichness is the demonstration of an overall patternof spatial distribution of parasitic and infectious
diseases species in human populations on Earththat conforms to a nested species subset hierarchy(see Guégan and Hugueny 1994; Guégan et al. 2001;Guernier et al. 2004). Nestedness structure indicatesthat species (here pathogens) that compose adepauperate community (here temperate condi-tions) statistically constitute a proper subset ofthose occurring in richer communities (herewarmer conditions in tropical areas), but theconverse situation, that is, pathogen species solelyoccurring in depauperate communities but not inthe richest ones, is either not found or not properlysubstantiated. This pattern, although not consid-ered in the ecological literature (but see Gaston andBlackburn 2000) who suspected the existence of aconnection between the two patterns; see later),was strongly associated with latitude, indicatingthat the progression of pathogen species richness isfrom species-poor countries in more temperateareas to species-rich ones when reaching tropicalzones (see Fig. 2.5).
Using Monte-Carlo simulations (see Manly 1991;Guégan and Hugueny 1994) to test the hypothesisof parasitic and infectious diseases spatial organi-sation on the largest scale, the authors assessedthe degree of nestedness of the system using two different but complementary programmes:(i) ‘Nestedness’ (Guégan and Hugueny 1994) and(ii) ‘Nestedness Temperature Calculator’ (Atmarand Patterson 1995). In the former programme,pathogen species were either selected with uniformprobability (R0) or with a probability proportionalto their incidence (R1) (Guégan and Hugueny 1994)whereas in the latter one only a R00 procedure wasretained (Atmar and Patterson 1995); (see alsoWright et al, 1998; Cook and Quinn 1998; Gastonand Blackburn 2000 for further details). Resultsfrom Monte Carlo simulations showed that theglobal distribution of human pathogens wasstrongly nested (Ns � 2481.4, R0 and R1 procedures,p � 0.0001), with some slight differences that werefound across the different groups of aetiologicalagents (all groups, p � 0.0001, but except for vector-borne viruses, with the R1 procedure (Ns � 1787, p � 0.0015). When considering the Northern andSouthern hemispheres separately, both were highlynested (R0 and R1 procedures, Ns � 6602, p � 0.0001
Rena-Ch02.qxd 8/20/04 9:58 PM Page 37

and Ns � 1230, p � 0.0001, respectively). This wasconfirmed by the R00 procedure used by the‘Nestedness Temperature Calculator’ program(Atmar and Patterson 1995), which provides a use-ful graphic representation of the results (Fig. 2.5),showing that parasitic and infectious diseasesspecies diversity decreases as we move northwardsor southwards from the equator (F � 28.2307, df � 161, p � 0.001), (see Guernier et al. 2004).Results from all three nestedness models (seeWright et al. 1998) explained reasonable amounts ofthe nested pattern in human pathogen speciesacross latitudes.
Wright et al. (1998) have suggested a cogentexplanation for nestedness as a series of probabili-stic filters, screening species with particular charac-teristics: local habitat suitabilities, differentialcolonization capacities of species, and sustainabil-ity of viable populations within their environment.Additionally, as pointed out by Gaston andBlackburn (2000), nestedness might be an inevitablesecond-order consequence of the same factors thatcause variation in species richness and range sizealong latitudinal gradients. It is exactly the viewwith which Guernier et al. (2004) totally agree withthe example of pathogen species diversity inhuman on a broad-scale.
Searching for the common causes explaining theexistence of both patterns, i.e. gradient in speciesrichness and nested structure, for parasitic andinfectious diseases diversity and composition inhuman communities, Guernier et al. (2004) retainedthe energy hypothesis as a likely candidate forexplanation. The energy hypothesis is a climate-based hypothesis that claims that energy avail-ability generates and maintains species richnessgradients (see Hawkins 2004 for a recent review).Many studies have successfully correlated gradi-ents in species diversity with variation in theclimatic environment, a relationship thought toshape large-scale biogeographic patterns (Hill et al.1999). The authors decomposed the potential effectof climate on pathogen diversity into Pearson’scorrelations to more deeply analyse the kindsof relationships between each of the four climatic variables and disease richness under study.The results show significant positive correlations
between pathogen species richness and the maxi-mum range of precipitation after Bonferonni multiplecorrection for all six of the parasite or infectiousdisease taxa considered: bacteria (r � 0.3545, df � 213,p � 0.0001), viruses directly transmitted from per-son-to-person (r � 0.2350, df � 215, p � 0.0001),viruses indirectly transmitted via a vector (r � 0.3575, df � 215, p � 0.0001), fungi (r � 0.3554, df � 216, p � 0.0001), protozoa (r � 0.3744, df � 216, p � 0.0001), and helminths (r � 0.4270, df � 215, p � 0.0001). On the other hand, the relationshipbetween pathogen species richness and monthlytemperature range was only significant for threegroups of pathogens: bacteria (r � 0.3016, df � 213, p � 0.0001), viruses directly transmitted (r � 0.2142,df � 214, p � 0.0015), and helminths (r � 0.2590, df � 213, p � 0.0001). No relationship between para-site species richness and mean annual temperatureappeared to be significant after Bonferroni correc-tions. Finally, only the relationship between bacter-ial species richness and mean annual precipitationwas significant (r � �0.1987, df � 213, p � 0.0034).No or very slight differences between total andcategories of pathogen species richnesses and someclimatic factors were observed between Northernand Southern hemispheres.
Many factors are involved in the determinationof the climate in an area, particularly latitude,altitude, and the position of the area relative tooceans and land masses. In turn, the climate largelydetermines the species of plants and animalsthat live in those areas. According to the resultsof Guernier et al. (2004), the maximum range ofprecipitation is highly correlated with latitudinalgradient of pathogen species, the parasitic speciesdiversity significantly increasing with this climate-based factor. Interestingly, the variation of precipi-tation around the mean was overall a betterpredictor of pathogen species distribution than itsaverage value, thus indicating that pathogenspecies, their vector and host populations mightbest function over only a wide range of precipita-tion, which is actually found in many tropicalregions of the world, those regions having more orless distinct wet and dry seasons during the year.Many parasites obviously require water as thebasic medium of their existence, and many others
38 PA R A S I T I S M A N D E C O S YS T E M S
Rena-Ch02.qxd 8/20/04 9:58 PM Page 38

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 39
strongly need wet conditions to complete their lifecycle, for example vector-borne diseases. Often,many microorganisms are also constrained by thehumidity of the atmosphere. Undoubtedly, thephysical factor of precipitation variation may affectparasitic and infectious microorganisms, vectorsand/or hosts over a range from low precipitationvariation at one extreme, for example, deserts, tohigh precipitation variation such like in the tropics.This relationship might be related to biologicalcycles and a variety of features in parasitic andinfectious stages that have evolved so that they arespecifically well adapted to the variability of pre-cipitation. So, prolonged drought should not befatal for some well-adapted microbes if wet condi-tions are encountered once to complete their lifecycle. Curiously, average precipitation was notretained as a good candidate for explaining the lat-itudinal gradient of pathogen species diversityexcept for bacteria (see Guernier et al. 2004). If weconsider the Earth as a simple body with an envi-ronmental gradient, such as the annual precipita-tion range, which runs from wet and hot equatorialregions northward and southward to Arctic andAntarctic areas with harsh conditions, then distanceand isolation from pathogen species-rich regions inthe tropics may screen pathogen species by theirextinction and colonization tendencies. Moreover,habitat suitability, for example presence of newhosts and reservoirs, and passive sampling mayscreen them by their habitat preference and avail-ability, and abundance, respectively. Guernier et al.(2004) reached the same conclusions as Calvete et al.(2003) indicating that parasite species richness,their spatial distribution and organization on verydifferent scales, climate-based forces, and the inter-play between habitat conditions and host–parasiteinteractions might be intimately connected to gen-erate the observed patterns of parasite speciesdiversity.
The overall conclusion that can be drawn fromthis section on the latitudinal gradient in speciesrichness for parasite and pathogen organisms is thata better understanding of parasitic and infectiousdiseases species diversity and community dynamicsover wide ranges of spatial scales is now clearlyneeded. The similarity in the patterns of some
parasitic or pathogen taxonomic groups and free-living organisms suggests that common mech-anisms are at work. Regardless of whether therichness of parasitic and infectious diseases simplytracks host diversity or, rather, is determined to agreater extent by exogenous factors, for example,climate-forced variables, is now a challenge thatneeds to be pursued in parasitology and epidemio-logy. In addition, the significant findings illustratedin this and other sections confirm that investigationson parasite and microorganism community assem-blages should also be performed at greater scalesthan the scale at which local variation in speciesrichness and composition is too often examined.Integration of systematics, biogeography, popu-lation and species dynamics, community ecology,and evolutionary biology is now essential for a complete understanding of the many scalingprocesses affecting parasitic and infectious diseases.
2.4 Linking parasite and microorganismcommunities and ecosystems. Directionsfor further work
The results summarized above underscore theimportant role of large-scale determinants on localparasite and pathogen community organization andassembly processes. They suggest that the success ofinvasive parasite species may vary depending onhow physical conditions, for example, size of thearea, geographical distance from a continentalsource, affect the rates of colonization. Additionally,the regional effects on local parasite or microorgan-ism species richness also tend to suggest thatresearch should be directed at regional processesand their effect on local diversity, regional processesbeing just as important as local ones in setting levelsof richness in parasite or microbe communities.Then, the existence of a latitudinal gradient of para-site and microbe species richness both at subregional(gut helminths in Spanish red partridges) and global(infectious diseases in human populations) scalesconfirms that investigations on parasites andmicroorganisms should expand the scale at whichlocal variation is traditionally examined. Moreinterestingly, the demonstration of the existence of a nested species subset pattern for human infectious
Rena-Ch02.qxd 8/20/04 9:58 PM Page 39

diseases (also true for gut helminths in Spanish redpartridges) suggests that the interplay between well-differentiated species of pathogen species withinassemblages and distinctive requirements forresources, including micro- and macro-habitats, con-tribute to the overall nestedness pattern and speciesdiversity we observed.
It is important to recognize that similar patterns,that is, species area, species isolation, local to globalrelationships, and latitudinal gradient in speciesdiversity, have been observed for many free-livingtaxonomic groups (see Gaston and Blackburn 2000;Hawkins et al. 2003), suggesting that commonmechanisms might be at work in generating theobserved patterns of species diversity for parasiticand infectious agents (see previous sections). Butwhat kinds of common properties and character-istics between, say, a virus species and a birdspecies, may produce similar patterns?
So far, the concern of parasitology and epidemi-ology has been widely aimed at defining the char-acteristics in which pathogens may differ fromother groups, focusing on details instead of search-ing for similarities. One pattern that appears to bepervasive across many communities, parasites,microbes, and free organisms alike, and at differentspatial hierarchical scales, is the nested species sub-set pattern. Nested patterns were observed in someparasitological–epidemiological studies (Guéganand Hugueny 1994; Hugueny and Guégan 1997;Guégan and Kennedy 1996; Guernier et al. 2004;Poulin and Valtonen 2001, 2002; Vidal-Martinezand Poulin 2003), but in many other cases commun-ities were observed to form random, unstructuredassemblages (Rohde et al. 1994, 1995, 1998; Poulin1996a,b; Worthen and Rohde 1996). The previoussections make clear something that should be obvi-ous, that is, that the four patterns documentedabove are not independent of each other. Morand et al. (2002) have provided a cogent explanationfor the generation of nested patterns in parasitecommunities, thus reinforcing the importance ofboth spatial and demographic stochasticity in para-site species distribution and composition. Fig. 2.6illustrates the way Morand and collaboratorssynthesize the hypothetical distributions of para-sites species among local communities and the
many factors that may be implied in shaping thesecommunities. Considering that nested versus non-nested species patterns are strongly linked to unimodal versus bimodal species frequencydistributions, respectively (Morand et al. 2002), theauthors concluded that observed patterns shouldbe the result of differential colonization/extinctionprocesses acting at the level of each parasite(or pathogen) species. Differential colonization/extinction processes, also called epidemiologicalprocesses in the field of epidemiology (seeAnderson and May 1991), are attributes of parasiteor microbe species, and are related to birthand death processes in population dynamics (seeMorand et al. 2002). Assemblages of microbesand parasites displaying nestedness on different hierarchical scales, ranging from within individualsof the same host population to the global scale, asshown in the present chapter, provide a strong indication that the same forces are at work acrossdifferent spatial and temporal scales.
So what emerges from this concept is that boththe recruitment of parasite (or pathogen) speciesfrom local (host) patches and the differential capa-cities of parasite (or pathogen) larval or adult formsto colonize those patches may have strong impactson the composition and richness of those commu-nities from the local to the global scale, and viceversa. This is an approach which is roughly similarto that adopted by metapopulation biologists(see Hanski 1999; Hanski and Gaggiotti 2004) andecologists working on the diffusion of infectiousdiseases (see Grenfell and Dobson 1995; Diekmannand Heesterbeek 2000), and in which invasibilityis a key factor for explaining local presence and distribution of a given species.
Invasibility is dependent on the availability oflocal resources and potential competition with localspecies, for example, presence of empty niches. Inmainstream ecology (see Elton 1958), it has beenhypothesized among other predictions that greaterdiversity should increase resistance to invasionsbecause the levels of limiting resources are gener-ally lower in more diverse habitats (Tilman et al.1996; Knops et al. 1999), thus giving rise to the so-called diversity–invasibility hypothesis. Adecrease in host species diversity allows remaining
40 PA R A S I T I S M A N D E C O S YS T E M S
Rena-Ch02.qxd 8/20/04 9:58 PM Page 40

E M E R G E N C E O F S PAT I A L PA R A S I TO L O G Y A N D E P I D E M I O L O G Y 41
species to increase in abundance as a resultof decreased competition for limiting ressources,a phenomenon called density compensation byecologists (see Begon et al. 1996). As a result, theincrease in remaining host abundances should facil-itate the invasibility of parasite or microbe species(Anderson and May 1978; Burdon and Chilvers1982; Antonovics et al. 1995). Indeed, host living inhigh densities may harbour a high diversity ofparasite species (Morand et al. 2000; Poulin andMorand 2000; Stanko et al. 2002), which in turnoften achieve high abundances (Arneberg et al.1998a). Not all available results agree with this,however (Stanko et al. 2002). Hence, both host diversity and disease invasibility are related to hostspecies abundances within communities (Burdonand Chilvers 1982; Mitchell et al. 2002). As alreadystated, this hypothesis has received some support ingeneral ecology (Tilman 1997), but further workneeds to be done in both parasitology and epidemi-ology to determine whether higher parasite (ormicrobe) species diversity at different hierarchicalscales, that is, within-host, within-population,and within-metapopulation community dynamicsprotects parasite or pathogen communities frominvaders (but see Torchin et al. 2003).
Another prediction available from the general lit-erature is that greater species diversity of microbesor parasites should decrease the severity of each of
the component diseases, a theorem which is calledthe species composition–disease hypothesis (seeElton 1958). This hypothesis predicts that anychanges in parasite or microbe community compos-ition within a host community should impact onthe severity and virulence of one, or a group of,diseases, which then proliferate to the detrimentof other parasite or microbe species. Both thesehypotheses, that is, the diversity–disease hypothesisand the species composition–disease hypothesis,have received some empirical support from theplant disease literature (Knops et al. 1999; Mitchelland Power 2003), but they have never been testedto our knowledge in the case of both animal orhuman pathogens. Research on community ecologyof parasite or microbe species, at the differenthierarchical scales discussed in the present paper,should clearly benefit from developing both experi-mental designs and comparative studies to explorein greater detail the potential linkages betweenspecies diversity and composition, invasibility, andother life-history traits such as virulence.
In addition, theoretical studies on parasite (ormicrobe) invasibility rest upon epidemiologicalprinciples, in which the local abundance of hosts isthe key determinant of whether a parasite can estab-lish into a naïve host population (Anderson and May1978; Burdon and Chilvers 1982; May and Anderson1978). Within the study of human infectious
Parasite species(by rank)
High probabilityof invasionLow probabilityof extinction
Large area sizeWide habitat typesLow isolation
Small area sizeSpecific habitat typesHigh isolation
Localities(by rank)
High probabilityof invasionLow probabilityof extinction
Figure 2.6 Nested (versus non-nested) speciessubset patterns are strongly associated with aninterplay of specific characteristics of parasite(or microbe) species themselves and localhabitat conditions to sustain or not a givenparasite (or microbe) species population and its life-cycle associates, for example, vectors,reservoirs. Parasite (or microbe) communityecology would benefit greatly by considering theimportance of epidemiological patterns ingenerating the processes observed.
Rena-Ch02.qxd 8/20/04 9:58 PM Page 41

diseases, recent investigations have clearly illus-trated the importance of local community sizefor the maintenance and diffusion of, in particular,childhood diseases (see Grenfell and Harwood 1997;Rohani et al. 1999; Dieckmann and Heesterbeek 2000;Grenfell et al. 2001). Unfortunately, very few studieshave gone a step further in demonstrating thathigher local abundance in hosts should yield higherparasite or microbe species diversity (but see Poulinand Morand 2000).
For macroparasites, and especially for contact-transmitted microparasites, both high parasiteabundance and high parasite species richnessshould be attained more easily at high host abund-ance or density. This is expected from epidemi-ological theory (see Grenfell and Dobson 1995)and supported by empirical studies on the deter-minants of parasite species richness (see Poulin andMorand 2000, and the present chapter). Indeed,host abundance/density is positively correlatedwith parasite (or microbe) species richness inseveral groups of vertebrates (Poulin and Morand2000). This trend has yet to be demonstrated forhuman populations (and plant pathogens as well)in which large communities as resources shouldharbour and sustain a greater diversity ofpathogens over space and time. Some, though veryfew, studies have even shown that parasite speciesrichness covaries positively with host species rich-ness across localities, an interesting finding that isyet to be more seriously quantified across differenthost taxonomic groups (Krasnov et al. 2004).
Any loss of host biological diversity should intu-itively have a direct influence on the diversity andabundances of parasites or pathogens, and in turnon their virulence and pathogenicity within hostpopulations. In addition, changes in host commun-ity composition should also influence the compon-ent community of parasites or microbes livingthere. Host species diversity should not randomlyaffect parasite species richness and composition assome of them will be more affected than others
depending on the exact causes of diversity loss.This loss may depend on habitat fragmentation,which should impact on rare host species livingat low abundance, or may be due to landscapechanges, which may influence the more adaptedhost species, and thus will favour either the spreadof rare species or invasion by exotic species(Daszak et al. 2001). Unfortunately, we cannot easilyrefer to parasitological or epidemiological studieson animals or humans that have specifically testedthese different hypotheses. Further empirical stud-ies are thus needed, which should contribute to abetter understanding of how native host speciesand their associated parasite or pathogen speciesmay prevent or limit the attacks of invaders (seeTorchin et al. 2003).
In summary, there are, of course, other interest-ing parasitological or epidemiological patterns thatare not discussed in the present chapter. However,our aim here was to argue that both parasitologyand epidemiology would first largely gain fromdeveloping both a much broader perspective and amore quantitative approach, as advocated recentlyby both macroecologists and community ecologists.Parasitology and epidemiology are two scientificfields that have evolved with an individual-centredresearch perspective, and which have then concent-rated their research efforts on only local phenom-ena over the past decades. One important messagefrom the present work is clearly that we need toadopt a much broader research perspective inparasite (or microbe) community ecology. Second,we are also intimately convinced that a better consi-deration of the linkages between host speciesdiversity and composition, and parasite or microbespecies diversity and composition within commun-ities on the one hand, and the risk of altering any(host) species and composition on host invasiveness,host defences and increase of virulence by parasitesor pathogens on the other hand. We are definitelyadvocating here a community ecology perspectivefor host–parasite systems.
42 PA R A S I T I S M A N D E C O S YS T E M S
Rena-Ch02.qxd 8/20/04 9:58 PM Page 42