applied plant anatomy: part 1: the origin of cells, tissues and systems in plants
TRANSCRIPT

Applied Plant Anatomy:Applied Plant Anatomy:
Part 1:
The origin of cells, tissues and systems in plants

outline
Where do cells come from?
What controls their formation?
What controls their organization?

tissue origins
meristemsmeristems
mother cellsmother cells
dermal fundamental vascular
protodermprotoderm
epidermisepidermis
parenchymaparenchyma
collenchymacollenchyma
sclerenchymasclerenchyma
stopstop
stopstop
stopstop
stopstop
procambiumprocambium
protophloemprotophloem
protoxylemprotoxylemprotoxylemprotoxylem
stostopp
metaphloemmetaphloemmetaphloemmetaphloem
metaxylemmetaxylem
stopstop
Go!Go!
Go!Go!
Go!Go!
Go!Go!
Go!Go!
Go!Go!

The shoot & root apex
The dermal tissue system arises in the shoot and root apical meristems
The protoderm produces epidermal cells
similar layers are formed in roots

differentiation
differentiation starts in meristematic cells, and new cells which are formed in specialized layers, such as the procambium shown at left. Here, you can see a strand of protophloem differentiating. These cells are not functional yet.
Question: Why does the protophloem differentiate before the protoxylem?
Question: Why do cells enlarge, what is the optimum size for cellsDo all cells have the same optima?

systems
cell and tissue systems
filling
protective
support
conductive
meristematic
functional
epidermis, periderm
parenchyma, collenchyma, sclerenchyma
collenchyma, sclerenchyma
phloem, xylem
procambium, cambium. periderm
storage, synthesis transport reproductive

the dermal system
the epidermis may become replaced by a new protective layer, called the periderm. This layer is also . This layer is also responsible for gas exchange, through structures responsible for gas exchange, through structures called called lenticels

Specialized cells
Plants are composed of many different cell types – all have specific functions and size, shape, wall structure and function, will be determined by position in the root, stem or leaf. Specialization does not equate to complexity.
For example, parenchyma cells in the cortex of roots and shoots, may well have a storage function.
in leaves, gas exchange is facilitated if intercellular spaces are present, as in these aerenchyma cells in the Canna leaf.
Remember;.. gas exchange requires the presence of stomata
OR even bigger:-

specialized cells
Gymnosperms contain modified cells which form ducts that contain resins and terpenoids
all leaves contain parenchyma cells which are specialized for photosynthesis, called mesophyll cells

supporting tissue 1
supporting tissue takes on many forms and can be simple or complex.

supporting tissue 1
pea stempea stem
waterlilly petiolewaterlilly petiolepea rootpea root
young young Pelargonium Pelargonium stemstemNew Zealand flaxNew Zealand flax
extensive fibre capsfibre caps support vascular tissue & leaf
11
2233
4455
XylemXylem in 1-4 provides support to roots, stems & leaves

supporting tissue 2 -- collenchyma
the shape in cross section varies - usually angular or lamellar
angular
lamellar
distribution varies (a) at corners in angular stems
in stems,
OR:-
it forms an ring, under the epidermis [outer cortex]; or it occurs mixed between other tissues, or as an inner cortical band

collenchyma - facts

supporting tissue 4
distribution varies (a) at corners in angular stems(see collenchyma).
it forms an ring, under the epidermis [outer cortex]; or it occurs mixed between other tissues, or as an inner cortical band
supports vascular bundles in leaves

sclerenchyma - facts

transport systems 1
phloemphloem
xylemxylem
bicollateralbicollateral
collateralcollateral
Vascular tissue is always arranged into vascular vascular bundlesbundles in stems and leaves.
In stems, xylemxylem is normally inside of (endarch toendarch to) the phloemphloem is described as being outside of (exarch toexarch to) the xylem. In a minority of families, phloem occurs on both sides of the xylem. These are bicollateral bicollateral vascular bundles.
xylemxylem
phloemphloem

transport systems 2
in roots
monocotyledonous roots contain many (more than 6-7 xylem strands.
and an equivalent number of phloem poles or strands

The xylem – cell organization
protoxylem
Protoxylem forms in regions where rapid cell elongation is still ongoing. As such, secondary thickening is limited, to accommodate stretching.

xylem development – juvenile to mature
from Esau: Anatomy of seed plants
xylem differentiation involves a number of critical steps – during each degradation of content occurs simultaneously with formation and synthesis of new, secondary cell walls. In the he final stages, the cytoplasmic content (nuclei, organelles etc.) are broken down, and flushed out to be recycled. The end product is a series of cells fit for rapid transport of water and water-soluble products.

the xylem – structural changes in development
Diagrams and micrographs from Esau: Plant Anatomy,

the xylem - an overview
although most of the cells of the xylem are dead at maturity (vessels, tracheids, fire-tracheids and fibers), xylem parenchyma cells are alive and contain cytoplasm a nuclei and organelles
Diagrams and micrographs from Esau: Plant Anatomy,

the xylem 3 cells and tissue
Xylem in dicots and monocots contain vessel members (V); tracheids (T); fibres (F) and xylem parenchyma elements
XT
F
In gymnosperms, vessels are absent
Diagrams and micrographs from Esau: Plant Anatomy,
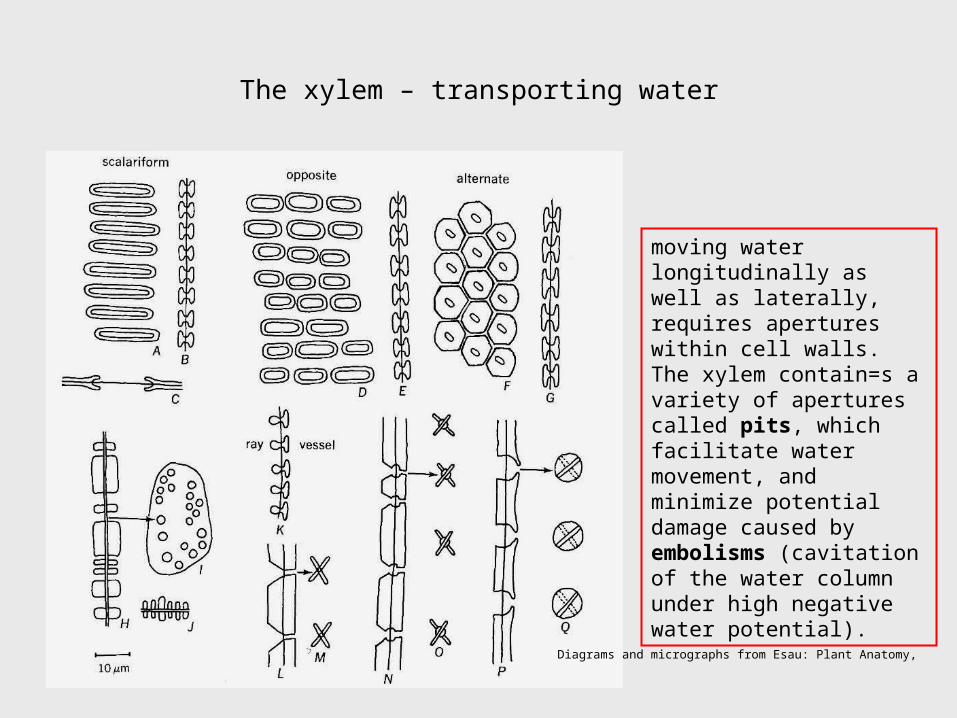
The xylem – transporting water
moving water longitudinally as well as laterally, requires apertures within cell walls. The xylem contain=s a variety of apertures called pits, which facilitate water movement, and minimize potential damage caused by embolisms (cavitation of the water column under high negative water potential).
Diagrams and micrographs from Esau: Plant Anatomy,

the xylem 4 – safer transport
Perforation plates in vessels are important structures, that will retard, or trap air bubbles which are formed during embolisms. Embolisms will, unless trapped, cause complete loss of functionality of the file of xylem vessels in which the bubbles occur.
unsafe

phloem tissue in angiosperms contains sieve tube members, joined end to end to form sieve tubes; as well as companion cells and phloem parenchyma cells.
in gymnosperms, the phloem comprises sieve cells, albuminous cells and parenchyma cells. It is also associated with transfusion tissue. This is a more primitive system than the angiosperm one.
vascular tissue in a pine needle
phloem
transfusion tissue
evolution of sieve tubes from Esau Anatomy of Seed Plants
CC
STsieve plate
phloem in melon petiolephloem in melon petiole
transport systems - the phloem 1

transport systems -the phloem in detail
light microscopy is only a starting point to understanding the structure of cells and tissues. The image to the right is a Transmission electron micrograph, which demonstrates the level of detail and power of TEM!
in the monocotyledon leaf, the phloem is made up of parenchyma cells, sieve tubes (ST) and companion cells (CC). Two types of sieve tubes occur - thick- (TWST) and thin-walled (ST) ones. TWST are not TWST are not associated with associated with CC.

transport systems - the phloem electron microscopy
in leaves, the sieve tubes are always narrower in diameter that associated parenchymatous elements, including the companion cells. This is because most phloem loading is an active process, either mediated by osmotic potential alone, or in combination with sucrose transporters, which are involved in loading the companion cell sieve tube complex.

moving carbohydrates -- the phloem
phloem is a complex tissue. Moving metabolites requires very specialized cells, called sieve tubes (in angiosperms) which, at maturity, do not have a vacuole, and do nto have a nucleus! Their end walls are perforate and the cells, joined end to end by these walls, form sieve tubes, through which assimilates move from a source (of the assimilate) to the sink (where they are used).
Diagrams and micrographs from Esau: Plant Anatomy,

sieve platessieve platessieve plates maintain cell integrity. Keep structures an proteins within cells, in place. The also have an important regulatory function

phloem is the principal carbohydrate transport channel – this channel is controlled
moving metabolitesmoving metabolites
The phloem is protected from damage (sudden pressure change) through the formation of callose on sieve plates and sieve areas
sieve areas
sieve plate

electron microscopy
sieve tubes contain plastids. These sieve type plastids have prominent protein bodies in them, with unknown function. Sieve tubes are relatively uncluttered with a clear lumen. Companion cells are associated nucleate cells

Origins: Control and regulation through genes
2. It is known that Knotted1-like homeobox1 (knox) genes are expressed in very specific patterns within shoot meristems and these genes play an important role in meristem maintenance. In plants, MADS box genes are most well known.
All differentiation is under gene control.
A great deal of work has been done using Arabidopsis
3. Misexpression of the knox genes, KNAT1 or KNAT2, in Arabidopsis produces a variety of phenotypes, including lobed leaves and ectopic stipules and meristems in the sinus, the region between lobes.
1. Genes affecting early stages of vascular patterning, prior to provascular network formation, may promote differentiation along wide pathways rather than narrow canals, because of failures to establish efficient canals of auxin flow.
For example:
----------------------1 A DNA sequence within genes involved in the regulation of development, about 180 base pairs long. It encodes protein (the homeodomain, which binds DNA http://en.wikipedia.org/wiki/Homeobox#Plants

More examples
In the Arabidopsis root meristem, initial cells undergo asymmetric divisions to generate the cell lineages of the root. The scarecrow mutation results in roots that are missing one cell layer owing to the disruption of an asymmetric division that normally generates cortex and endodermis. Cell 86: 432-433 Laurenzio et al. 1996
When differentiating cells leave the meristem field, they actively maintain a pool of uncommitted cells in the SAM. This suggests the maintenance of the meristem cells in an undifferentiated state - this is likely to be shared by other plant species
Shoot apical meristems (SAMs) do not share their set of regulatory factors with root apical meristems (RAMs), yet both adjust their cell populations according to the same basic mechanisms, such as intercellular signaling. In both SAMs and RAMs, these mechanisms involve interactions between two groups of cell populations, the pluripotent undifferentiated cells –in the organizing center of the SAM and in the quiescent center (QC) in the RAM – and the differentiating cells that will be incorporated into the plant body1.
1Nakajima and P. Benfey, Signalling in and out: control of cell division and differentiation in the
shoot and root. Plant Cell 14 Suppl. (2002), pp. S265–S276.
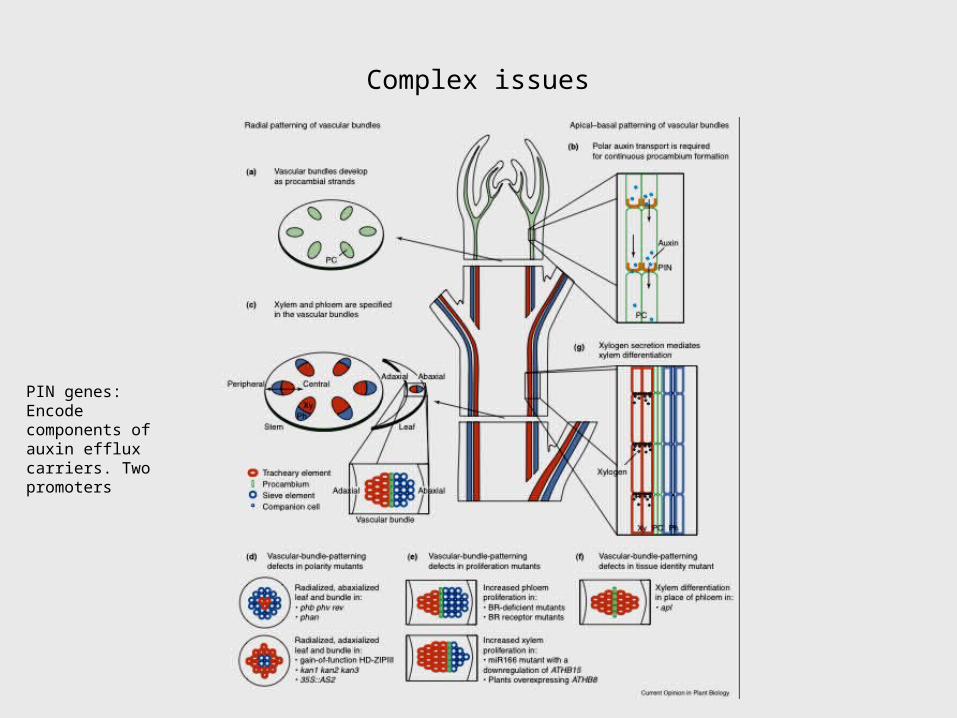
Complex issues
PIN genes: Encode components of auxin efflux carriers. Two promoters

More…zippers
it has been suggested that class III homeodomain leucine-zipper1 proteins (HD-Zip III) are involved in vascular development. However, little is known about the mechanisms of spatial and temporal organization in each vascular cell.
A protein structural motif involved in protein-protein interactions in many eukaryotic regulatory proteins (C/EBP prototype). Contain a repeat structure: Leu residues in every seventh position, causes a large amount of DNA to loop out. http://www.fhsu.edu/chemistry/twiese/glossary/biochemglossary.htm
Arabidopsis inflorescence stems develop extraxylary fibers at specific sites in interfascicular regions. The spatial specification of interfascicular fiber differentiation is regulated by the INTERFASCICULAR FIBERLESS1 (IFL1) gene because mutation of that gene abolishes the formation of normal interfascicular fibers in Arabidopsis stems.

Zipping vascular evolution
• See http://www.nature.com/nrm/journal/v5/n5/full/nrm1364.html
• Plant signals
Figure 1 Model of how CLASS III HD-ZIP1 and KANADI activities pattern lateral organs and vasculature. A centrally derived signal (red) activates CLASS III HD-ZIP genes, whose activity is antagonistic with that of KANADI activity. Both KANADI and MIR165/166 negatively regulate CLASS III HD-ZIP genes, (relationship between the two is not presently known). In lateral organs, CLASS III HD-ZIP activity promotes adaxial fates and KANADI activity promotes abaxial fates. In the vascular bundles, interactions between the two gene classes pattern the arrangement of xylem and phloem tissues. The vascular bundle shown is already differentiated, but the initial patterning events likely occur just below the apical meristem where provascular cells are being specified. –––––––––––––––-–1Class III homeodomain-leucine zipper proteins
Arabidopsis class III homeodomain-leucine zipper (HD-Zip III) proteins play overlapping, distinct, and antagonistic roles in key aspects of development that have evolved during land plant evolution.

Which comes first?
• The provision of nutrient and water becomes priority problems within the developing shoot or root axis. As the axis elongates and the diameter increases, so transport becomes more problematic
• Short-distance transport may be accommodated by:-diffusion, provided there are adequate cell to cell connections
•or by transmembrane transport, either through cells or along the cell wall free space interface

Size and shape; long is better for transport
Fig. 1. Changes in surface-to-volume ratios during cell expansion. When a cell (shown here as a cube), doubles its dimensions via (a) isotropic expansion its volume increases 8-fold whereas its surface increases only 4-fold and its surface-to-volume ratio is reduced from 6 to 3. (b) Anisotropic expansion of the same volume, producing long thin cells, increases the surface area to a greater extent and improves the surface-to-volume ratio. (c) The original ratio of surface-to-volume can be maintained when cell expansion is followed by cell division. From: Kondorosi et al, Current Opinion in Plant Biology Volume 3, Issue 6, 1 December 2000, Pages 488-492

Applied plant anatomy
Next: Part 2:
the root-stem-leaf continuum Intro Anatomy 1
