anticholinesterase pesticides (metabolism, neurotoxicity, and epidemiology) || gene expression
TRANSCRIPT

14GENE EXPRESSION
SHIRIN POURNOURMOHAMMADI
Faculty of Pharmacy, Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia
MOHAMMAD ABDOLLAHI
Faculty of Pharmacy and Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran, Iran
14.1 Introduction 175
14.2 Insecticide-Induced Gene Expression Alterationin Different Tissues 17614.2.1 Neural Cell Components 17614.2.2 Apoptosis 18014.2.3 Non-Neural Cell Components 18114.2.4 Cell Cycle Components 181
14.2.5 Cell Metabolism Components 18214.2.6 Cell Stress Components 18314.2.7 Other Components 184
14.3 Conclusion 185
References 185
14.1 INTRODUCTION
With the increasing demand for insecticide products, as wellas insecticide-contaminated food and different formulations,the question of their safety has become one of the seriousworld public health issues. Insecticides are categorized asanticholinesterase organophosphoruses (OPs), such asTEPP, phorate, mevinphos, fensulfothion, demeton, disulfo-ton, parathion, fonophos, famphur, coumaphos, ronnel,crufomate, trichlorfon, dichlorvos, chlorpyrifos, fenthion,diazinon, dimethoate, malathion, abate, and isoflurophate;and anticholinesterase carbamates (CM), such as aldicarb,oxamyl, carbofuran, benfuracarb, methomyl, formetanate,aminocarb, dimetilan, dimetan, dioxacarb, methiocarb, pro-poxur, bendiocarb, pirimicarb, bufencarb, MTMC, MPMC,isoprocarb, carbaryl; as well as organochlorines, and otherslike rodenticides, herbicides, fungicides, and fumigant com-pounds (see INCHEM, http://www.inchem.org).
OP insecticides and even new, more toxic, ones are amajor concern for human health because of their tendency
to damage different tissues at exposures below the thresholdfor signs of systemic intoxication. Generally, data indicatesthat most poisonings occur in young people by intentionaldrug overdose, followed by consumption of pesticides,especially OP and CM compounds. Studies also show thatOPs are one of the most common causes of poisoning-relateddeath. Several series of compounds are categorized as pesti-cides, but among them OP and CM compounds have anti-ChE activity causing demonstrating cholinergic effects inhuman acute exposure. OP and CM compounds are the mostcommonly used insecticides in the world and there are manycases of poisoning every year. They can be found in variousformulations, including dusting powder, flea collars, andshampoos for pets, etc. which are harmful over long periodsof time. The problem of pesticide residues in the environmentis still an important concern in terms of chronic toxicity eventhough people are commonly exposed to pesticides, which arenot well monitored (Soltaninejad and Abdollahi, 2009).
Several pesticides cause DNA strand breaks or fragmen-tation (Bolognesi, 2003). Variability in environmental
Anticholinesterase Pesticides: Metabolism, Neurotoxicity, and Epidemiology. Edited by Tetsuo Satoh and Ramesh C. GuptaCopyright # 2010 John Wiley & Sons, Inc.
175

exposure, coupled with genetic events like translocation,alters disease risk (Edwards and Myers, 2007). In vivoanimal studies of malathion exposure have shown positiveresults for chromosomal damage, though a study onmalathion to show genotoxicity and/or carcinogenicity hasbeen inconclusive in humans (Shadnia et al., 2005;Windham et al., 1998). Blasiak et al. (1999) analyzed theeffect of malathion exposure alone on human peripheralblood lymphocytes.
No studies have been published to date on chromosomalvariations related to malathion exposure. Some geneexpression changes after exposure to malathion are found inother cell culture models of chromosomal changes (Geiglet al., 2004). Discovery of genes altered after exposure topesticides may aid in future studies in environmental andoccupational epidemiology on pesticide exposures. Geneexpression profiling in response to toxic chemicals can beused to seek evidence for a chemical’s potential toxicityand carcinogenicity and, in future, may provide useful geneticbiomarkers that could be used in a clinical setting for earlydetermination of OP exposure as well as early diagnosis ofpesticide illness. Gene expression signatures for pesticideswith unknown side effects can be used to identify pesticideaction, which would make an association between pesticidesand known carcinogens or genotoxic agents.
In this chapter, data on pesticides and gene expression arereviewed (Table 14.1).
14.2 INSECTICIDE-INDUCED GENEEXPRESSION ALTERATION IN DIFFERENTTISSUES
14.2.1 Neural Cell Components
14.2.1.1 Cholinesterase Although it was originallythought that OPs act exclusively through inhibition of cholin-esterase (ChE) and consequent cholinergic hyperstimulation,it is now evident that there are multiple mechanisms that leadto neurodevelopmental abnormalities. OPs affect mammalianbrain development through a variety of mechanisms beyondtheir shared property of ChE inhibition.
14.2.1.2 Cell Growth Gene OPs differentially regulatemembers of the fibroblast growth factor (FGF) gene family,which has a role in brain development and proliferation,migration, differentiation, survival, synaptogenesis, andmyelination of cells. Slotkin and coworkers (Slotkin andSeidler, 2007; Slotkin et al., 2008) showed that subcutaneousadministration of chlorpyrifos (1 mg/kg/day) or diazinon(1 or 2 mg/kg/day) to neonatal rats 1 to 4 days postnatal atdoses that do not cause systemic toxicity or growth impair-ment and inhibition of ChE enzyme could induce major tran-scriptional changes in expression of many identified genes
involved in (1) neural cell growth and development of gliaand myelin; (2) transcription factors and cell signaling cas-cades that mediate neural cell differentiation; (3) cytotoxicevents, including oxidative stress and apoptosis; and (4)neurotransmitter pathways for acetylcholine, serotonin,norepinephrine, and dopamine, akin to different classes ofneurotrophic factors, including mRNAs encoding the neuro-trophins (NTFS), glial fibrillary acidic protein (GFAP), gluta-thione S-transferase (GST) etc., and the correspondingreceptors; although the expression of some genes like brain-derived neurotrophic factor (BDNF), the nerve growthfactor (NGF) has been much less changed (Table 14.1).Diazinon had similar effects on many of the same processesbut also showed major differences from chlorpyrifos whichbuttress the idea that different OPs target multiple pathwaysinvolved in neurodevelopmental outcomes.
Comparing the effects of the in vivo chlorpyrifos treat-ment with those in PC12 cells as a standard in vitro modelof mammalian neurodevelopment that reproduces many ofthe key mechanisms and features of the adverse effects ofOP on neural cell replication and differentiation, it is evidentthat effects in PC12 cells parallel many gene expressions formembers of the FGF and NTF families.
On the other hand, even oral administration of chlorpyrifosdaily to neonatal rats on days 10 to 20 postnatal at doses of 4.0or 6.0 mg/kg, which inhibits acetylcholinesterase (AChE)and growth, could alter the expression of mRNA for factorscritical to brain development such as BDNF and NGF inthe hippocampus and cortex. Conversely, oral administrationof chlorpyrifos daily on postnatal days 1 to 6 at doses of 1.5 or3.0 mg/kg could not change AChE activity in the forebrain,though body weight was significantly different from controlson postnatal day 7 in both males and females exposed to3.0 mg/kg chlorpyrifos. Increased NGF protein levels wereobserved in the hippocampus but not the cortex on postnatalday 20 with some reduction occurring on postnatal day 28 inboth regions. These changes did not correlate with thechanges in NGF mRNA. BDNF mRNA was increased inboth regions on postnatal days 20 and 28, whereas BDNFprotein levels were increased on postnatal day 20. On post-natal day 12, c-fos mRNA, a marker of neuronal activation,was increased in both regions. Total BDNF protein wasincreased in the hippocampus but decreased in the cortexwith no changes in NGF protein. The expression of NGF,reelin, and M1 mAChR mRNA was significantly reducedwith 1.5 or 3.0 mg/kg of chlorpyrifos in forebrain of bothsexes on postnatal days 1 to 6. b-III tubulin mRNAexpression remained unchanged after exposure, whereasmyelin-associated glycoprotein (MAG) mRNA expressionwas significantly decreased with both dosages of chlor-pyrifos in both sexes, suggesting effects on the developingoligodendrocytes. In contrast, glial fibrillary acidic protein(GFAP) mRNA levels were significantly increased with1.5 and 3.0 mg/kg of chlorpyrifos in both sexes, suggesting
176 GENE EXPRESSION
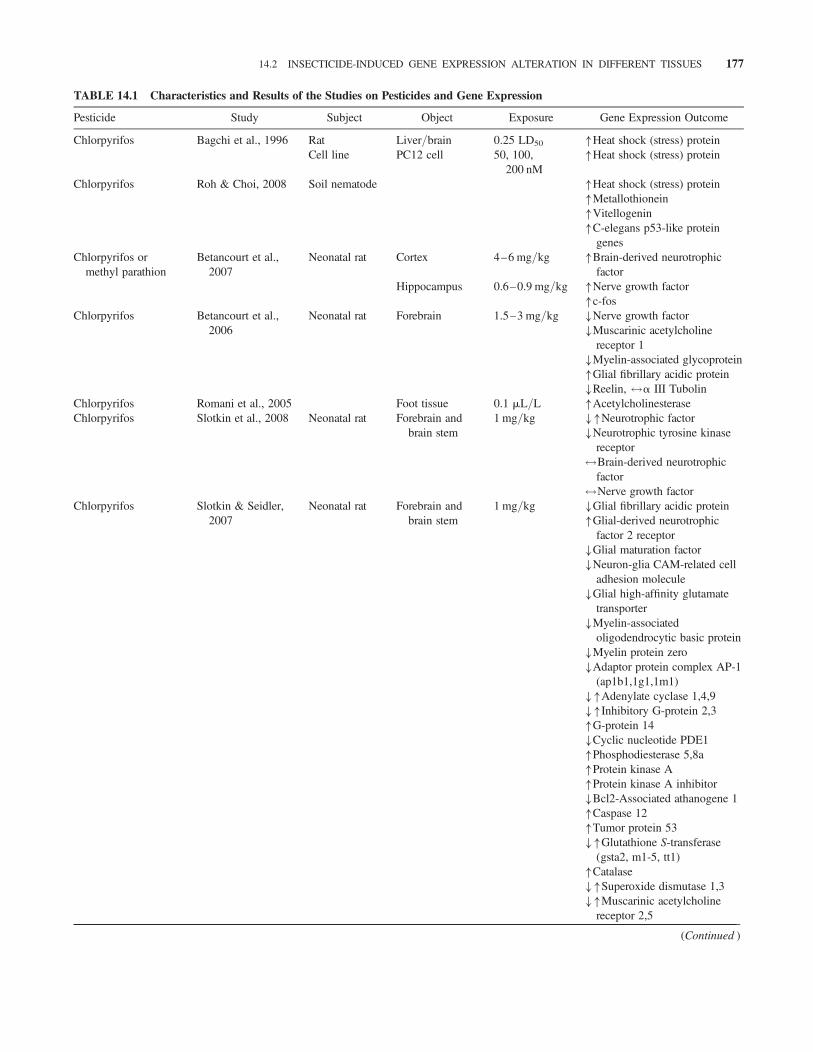
TABLE 14.1 Characteristics and Results of the Studies on Pesticides and Gene Expression
Pesticide Study Subject Object Exposure Gene Expression Outcome
Chlorpyrifos Bagchi et al., 1996 Rat Liver/brain 0.25 LD50 �Heat shock (stress) proteinCell line PC12 cell 50, 100,
200 nM�Heat shock (stress) protein
Chlorpyrifos Roh & Choi, 2008 Soil nematode �Heat shock (stress) protein�Metallothionein�Vitellogenin�C-elegans p53-like protein
genesChlorpyrifos or
methyl parathionBetancourt et al.,
2007Neonatal rat Cortex 4–6 mg/kg �Brain-derived neurotrophic
factorHippocampus 0.6–0.9 mg/kg �Nerve growth factor
�c-fosChlorpyrifos Betancourt et al.,
2006Neonatal rat Forebrain 1.5–3 mg/kg �Nerve growth factor
�Muscarinic acetylcholinereceptor 1
�Myelin-associated glycoprotein�Glial fibrillary acidic protein�Reelin, $a III Tubolin
Chlorpyrifos Romani et al., 2005 Foot tissue 0.1 mL/L �AcetylcholinesteraseChlorpyrifos Slotkin et al., 2008 Neonatal rat Forebrain and
brain stem1 mg/kg ��Neurotrophic factor
�Neurotrophic tyrosine kinasereceptor$Brain-derived neurotrophic
factor$Nerve growth factor
Chlorpyrifos Slotkin & Seidler,2007
Neonatal rat Forebrain andbrain stem
1 mg/kg �Glial fibrillary acidic protein�Glial-derived neurotrophic
factor 2 receptor�Glial maturation factor�Neuron-glia CAM-related cell
adhesion molecule�Glial high-affinity glutamate
transporter�Myelin-associated
oligodendrocytic basic protein�Myelin protein zero�Adaptor protein complex AP-1
(ap1b1,1g1,1m1)��Adenylate cyclase 1,4,9��Inhibitory G-protein 2,3�G-protein 14�Cyclic nucleotide PDE1�Phosphodiesterase 5,8a�Protein kinase A�Protein kinase A inhibitor�Bcl2-Associated athanogene 1�Caspase 12�Tumor protein 53��Glutathione S-transferase
(gsta2, m1-5, tt1)�Catalase��Superoxide dismutase 1,3��Muscarinic acetylcholine
receptor 2,5
(Continued )
14.2 INSECTICIDE-INDUCED GENE EXPRESSION ALTERATION IN DIFFERENT TISSUES 177
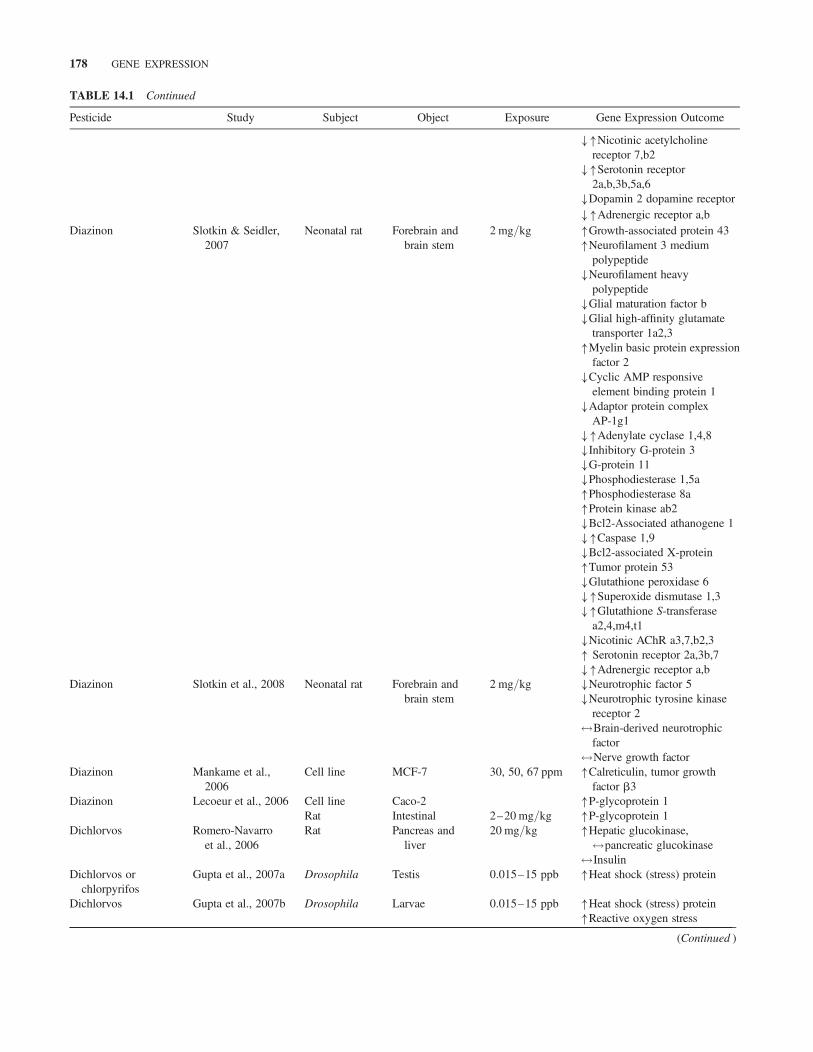
TABLE 14.1 Continued
Pesticide Study Subject Object Exposure Gene Expression Outcome
��Nicotinic acetylcholinereceptor 7,b2
��Serotonin receptor2a,b,3b,5a,6
�Dopamin 2 dopamine receptor��Adrenergic receptor a,b
Diazinon Slotkin & Seidler,2007
Neonatal rat Forebrain andbrain stem
2 mg/kg �Growth-associated protein 43�Neurofilament 3 medium
polypeptide�Neurofilament heavy
polypeptide�Glial maturation factor b�Glial high-affinity glutamate
transporter 1a2,3�Myelin basic protein expression
factor 2�Cyclic AMP responsive
element binding protein 1�Adaptor protein complex
AP-1g1��Adenylate cyclase 1,4,8�Inhibitory G-protein 3�G-protein 11�Phosphodiesterase 1,5a�Phosphodiesterase 8a�Protein kinase ab2�Bcl2-Associated athanogene 1��Caspase 1,9�Bcl2-associated X-protein�Tumor protein 53�Glutathione peroxidase 6��Superoxide dismutase 1,3��Glutathione S-transferase
a2,4,m4,t1�Nicotinic AChR a3,7,b2,3� Serotonin receptor 2a,3b,7��Adrenergic receptor a,b
Diazinon Slotkin et al., 2008 Neonatal rat Forebrain andbrain stem
2 mg/kg �Neurotrophic factor 5�Neurotrophic tyrosine kinase
receptor 2$Brain-derived neurotrophic
factor$Nerve growth factor
Diazinon Mankame et al.,2006
Cell line MCF-7 30, 50, 67 ppm �Calreticulin, tumor growthfactor b3
Diazinon Lecoeur et al., 2006 Cell line Caco-2 �P-glycoprotein 1Rat Intestinal 2–20 mg/kg �P-glycoprotein 1
Dichlorvos Romero-Navarroet al., 2006
Rat Pancreas andliver
20 mg/kg �Hepatic glucokinase,$pancreatic glucokinase$Insulin
Dichlorvos orchlorpyrifos
Gupta et al., 2007a Drosophila Testis 0.015–15 ppb �Heat shock (stress) protein
Dichlorvos Gupta et al., 2007b Drosophila Larvae 0.015–15 ppb �Heat shock (stress) protein�Reactive oxygen stress
(Continued )
178 GENE EXPRESSION

TABLE 14.1 Continued
Pesticide Study Subject Object Exposure Gene Expression Outcome
DFP Gupta et al., 1998 Hen Brain and spinalcord
1.7 mg/kg �CaM kinase II
DFP Gupta et al., 2000 Hen Brain and spinalcord
1.7 mg/kg �CaM kinase II��Tubolin�c-fos
DFP Damodaran &Abou-Donia2000
Hen Brain and spinalcord
1.7 mg/kg ��Glial fibrillary acidic protein��Vimentin
DFP Damodaran et al.,2001
Hen Brain and spinalcord
1.7 mg/kg ��a-tubolin
DFP Damodaran et al.,2002
Hen Brain and spinalcord
1.7 mg/kg ��Glyceraldehyde-3-phosphatedehydrogenase
�� Intermediate filamentproteins
Fenitrothion Yoshimi et al., 2002 MidgeChironomus
Larvae 0.4 mg/L �Heat shock (stress) protein
Malathion Gwinn et al., 2005 Cell line PrimaryNHMECs
25–100mg/mL
�Aldo–keto reductase 1C1�Aldo–keto reductase 1C2�Estrogen-responsive gene�Centromere protein F�Reactive factor C3�Thymidylate synthetase�Putative mitotic checkpoint
kinase (BUB1)�Plasminogen activator
Malathion Zhu et al., 2007 Lyguslineolaris
�Glutathione S-transferases
Methyl parathion Abel et al., 2004 Rat/mice/human
Cytosolicfraction
300 mM �Glutathione S-transferases
Monocrotophos Sharma et al., 2008 Blowfly Larvae 0.0005–0.005 ppm
�Heat shock (stress) protein
Adult 0.0001–0.001 ppm
�Heat shock (stress) protein
Parathion or malathionand 17 estradiol
Calaf & Roy, 2008 Cell line MCF-10F �Cyclin D, cyclin-dependentkinases 41
�C-Ha-ras oncogene�IGFBP, keratin 18, DVL1,
TP53�Minichromosome maintenance
deficient 2�Heat shock (stress) protein
Parathion Calaf & Roy, 2007 Cell line MCF-10F �Cyclin D, cyclin-dependentkinases 41
�c-kit, Trio, Rho-A, Rac-3, Dvl2,Notch4
�Epidermal growth factorreceptor
�Ezrin, b catenin, mutant p53protein
�Minichromosome maintenancedeficient
Prothiofos Lee et al., 2007 Plutellaxylostella
Head �Acetylcholinesterase
Tetrachlorvinfos De Sousa et al.,1997
Rat/human hepatocytes �Cytochrom 1A1
(Continued )
14.2 INSECTICIDE-INDUCED GENE EXPRESSION ALTERATION IN DIFFERENT TISSUES 179

increased astrocyte reactivity. These results show thatrepeated developmental OP exposure during the postnatalperiod alters NGF and BDNF in the cortex and the hippo-campus and the patterns of these alterations differ betweenregions.
14.2.2 Apoptosis
Methyl-parathion at doses of 0.6 or 0.9 mg/kg had similareffects on many of the gene families involved in brain deve-lopment (Betancourt et al., 2006, 2007). To clarify whetherchlorpyrifos or its product has a role in neurotoxicity, thereis a report showing chlorpyrifos and chlorpyrifos-oxon, butnot 3,5,6-trichloro-2-pyridinol (the breakdown product ofchlorpyrifos and chlorpyrifos-oxon), induce apoptosis inprimary cortical neurons cultured from embryonic day 17 ornewborn rats. It is generally agreed that chlorpyrifos-oxon isapproximately three orders of magnitude more potent thanchlorpyrifos in inhibition of brain AChE activity.
However, chlorpyrifos-oxon is only slightly more potentthan chlorpyrifos in inducing apoptosis. This indicates thatchlorpyrifos-induced apoptosis may occur independently ofAChE inhibition. Furthermore, chlorpyrifos activates theextracellular signal-regulated kinases 1/2 (ERK) and p38mitogen-activated protein (MAP) kinases. Surprisingly,blocking ERK 1/2 activation by the MEK inhibitor SL327caused a small but statistically significant inhibition of apop-tosis, while blocking of p38 with SB202190 significantlyaccelerated apoptosis induced by chlorpyrifos. This suggestsa pro- and anti-apoptotic role for ERK1/2 and p38, respecti-vely. Although chlorpyrifos did not stimulate total c-JunN-terminal kinases (JNK) activity, it caused a sustained acti-vation of a subpool of JNK in the nucleus and stimulated phos-phorylation of c-Jun, a downstream target of JNK. Transientexpression of a dominant negative c-Jun mutant inhibitedchlorpyrifos-induced apoptosis, suggesting a role for JNKand JNK-mediated transcription in this cell death (Caughlanet al., 2004). Together, these reports point to the fact thatOP neurotoxicity is not attributable to AChE inhibition.
Diisopropyl phosphorofluoridate (DFP) producesdelayed neurotoxicity, known as OP ester-induced delayedneurotoxicity (OPIDN), in hen, human, and other
sensitive species. A single dose of DFP (1.7 mg/kg, s.c.)produces first mild ataxia followed by paralysis in 7 to 14days in hens.
14.2.2.1 Glyceraldehyde-3-Phosphate DehydrogenaseThe effects of DFP administration on the mRNA expres-sion of some genes, including glyceraldehyde-3-phosphatedehydrogenase (GAPDH), an important glycolytic protein,alpha tubulin, one of the major subunit of tubulin polymersas an important constituent of cellular architecture, Ca2þ
calmodulin-dependent protein kinase II (CaM kinase II),and intermediate filament proteins such as glial fibrillaryacidic protein (GFAP) and vimentin as indicators of neuro-toxicity and astroglial pathology in different hen brain regions(cerebrum, cerebellum, brainstem, and spinal cord) at differ-ent time points up to 20 days determined the initial down-regulation of mRNA levels of tubulin, no elevation or slightdownregulation at day 1 for GAPDH, downregulation ofvimentin transcript levels at 20 days after an early reducedlevels. However, GFAP transcript levels reached near controllevels at 20 days post DFP treatment after an initial down-regulation in the highly susceptible tissues like brainstemand spinal cord indicating that DFP-induced stress responseresults in inhibition of cell metabolism and or cell injury/death. This was paralleled with a significant increase inc-fos expression as immediate-early gene in 30 min. Thiselevated level persisted at least up to 2 h in all brain regionssuggesting its role in the development of OPIDN. Increasein levels of tubulin mRNA at 5 days and thereafter coincidedwith increased tubulin transport, which may be due toincreased phosphorylation of tubulins in both axons,perikarya, and other intra-axonal changes resulting inimpaired axonal transport. DFP-induced decrease of tubulinpolymerization resulting in increased levels of free tubulinmonomers may be involved in the altered alpha tubulinmRNA expression at different time points by autoregulatorycircuits.
Nonsusceptible tissues like cerebrum have shown a dra-matic increase in GAPDH mRNA at day 1 post-treatment,and levels remained high at all time points suggestive of pro-tective mechanisms from the beginning. The cerebellumshowed only a moderate decline at day 2, while the alpha
TABLE 14.1 Continued
Pesticide Study Subject Object Exposure Gene Expression Outcome
Thiram Rath et al., 2007 Broilerchickens
growth platecartilage
100 ppm �Vascular endothelial growthfactor
�Bcl2Carbaryl Cheng et al., 2006 Cell line hGLCs 1–125 mmol/L �Steroidogenic acute regulatory
protein
Note: Increase (�), decrease (�), decrease or increase (��), no change in gene expression ($).
180 GENE EXPRESSION

tubulin remained near control levels at day 1; followed bymoderate downregulation of GFAP levels at 1, 2, and 5days then achieving near control values at 10 and 20 days.Vimentin was rapidly re-induced in cerebellum at 5 daysand remained at the same level at 10 days and then returnedto control values at 20 days after an initial downregulationat 1 and 2 days. Thus, these alterations were less drastic in cer-ebellum as indicated by initial susceptibility followed byrapid recovery.
On the other hand, both GFAP and vimentin levels wereupregulated from 2 days onwards in the nonsusceptibletissue cerebrum, implying protective mechanisms fromthe beginning. Delayed downregulation may be due to thecoordinated up- or downregulation of different subtypes ofalpha and beta tubulins as well as the differential responseof specialized cell types in cerebellum. Continuous over-expression of alpha tubulin in cerebrum from the beginningmay be its effective protective strategy to safeguard itselffrom neurotoxicity. DFP treatment also increases in vitroautophosphorylation of Ca2þ calmodulin-dependent proteinkinase II (CaM kinase II) and the phosphorylation of severalcytoskeletal proteins in the hen brain. The increase in CaMkinase II mRNA expression is consistent with increase inits activity. Data point to an important link in GAPDH andnitric oxide (NO), CaM kinase-II, protein kinase-A (PKA),c-fos, and phosphorylated-CREB (p-CREB) in DFP-inducedOPIDN. In addition, the DFP-induced astroglial pathology asindicated by the complex expression profile of GFAP andvimentin mRNA levels may be playing an important role inthe delayed degeneration of axons or is the result of progres-sive degeneration of axons in OPIDN. Overall, the differentialexpression pattern observed could be due to the differentialsusceptibility and variability in the rate of axonal transportof different regions besides the tubulin heterogenity ofCNS. Cerebellum, being the less susceptible tissue, showedmoderate increase initially followed by higher induction,suggestive of rapid recovery (Damodaran and Abou-Donia,2000; Damodaran et al., 2001, 2002; Gupta et al., 1998,2000).
14.2.3 Non-Neural Cell Components
14.2.3.1 Acethylcholinesterase The pesticide chlorpyri-fos could inhibit all enzymatic forms of true AChE poorlyafter a 4-day or a 15-day exposure to a sublethal concentration(0.1 mL/L) in foot or gill tissues of the benthonic bivalvemollusk Scapharca inaequivalvis. Also it could increase themaximum velocity (Vmax) and the Michaelis constant (Km)values. Consequently, catalytic efficiency of AChE, asdefined by the ratio Vmax/Km rises in the gills and drops inthe foot which would produce an overexpression of AChE-specific mRNAs suggesting that overexpression of enzymesin the presence of chlorpyrifos may be a consequence of its
resistance which seems to depend mainly on the resultingincrease in AChE content (Romani et al., 2005).
OPs have endocrine agonist or antagonist activities andalter hormone regulation of gene expression as has beenshown in MCF-7 cells, an estrogen-dependent human cellline, treated to 30, 50, or 67 ppm diazinon. Diazinon-treatedcells showed significant upregulation of calreticulin andTGF-b3 (Mankame et al., 2006).
14.2.4 Cell Cycle Components
In vitro analysis indicated that malathion’s two main meta-bolites, malaoxon and isomalathion, did in fact act as geno-toxic compounds after only one hour of exposure, whereaspure-grade malathion had no such effect. Gene expressionalterations in response to malathion exposure were found inall cell strains for nine genes:
1. Aldo-keto reductase 1 (AKR1C1)
2. Aldo-keto reductase 2 (AKR1C2)
3. An estrogen-responsive gene (EBBP)
4. Plasminogen activator (PLAT)
5. Centromere protein F (CPF)
6. Replication factor C (RFC3)
7. Thymidylate synthetase (TYMS)
8. Putative mitotic checkpoint kinase (BUB1)
9. A gene of unknown function (AI859865)
Of these, those that increased (1 to 3) are potentiallyinvolved in carcinogen and steroid metabolism, whereassome of those that decreased are associated with DNA repli-cation (5, 6, and 7) and cell cycle progression (8). The aldo-keto reductase gene family members are involved in thebreakdown and eradication of endogenous and exogenoussubstrates, including steroids and pesticides. Recent studieshave implicated these genes in the activation of polycyclicaromatic hydrocarbons, suggesting a potentiation of carcino-genicity in the presence of mixed exposures (Palackal et al.,2002; Penning et al., 1996).
Three of the six genes found to be decreased after mala-thion exposure are involved in DNA replication, with alteredexpression resulting in cell cycle arrest (Ellison and Stillman2003; Testa et al., 1994; Trinh et al., 2002), suggesting thatexposure to malathion may increase the possibility of carcino-genesis. Of the three genes found to be increased, AKR1C1and AKR1C2 were also increased after exposure to a non-OP pesticide, oxythioquinox (Gwinn et al., 2004).
Therefore, these two genes may be good general markersof pesticide exposure, with perhaps EBBP as a specificmarker for malathion exposure. Genes decreased in three ofthe four cell strains, including three genes involved in theregulation and progression of the cell cycle, CDC20,
14.2 INSECTICIDE-INDUCED GENE EXPRESSION ALTERATION IN DIFFERENT TISSUES 181

cyclinA2, and p16INK4a. CDC20 and cyclinA2 are bothinvolved in the cell’s progression from metaphase to ana-phase, with CDC20 being necessary to activate the anaphasepromoting complex leading to the degradation of cyclinA2(Dunican et al., 2002; Yih and Lee 2003). Malathion-inducedexpression showed some similar expression patterns to thosein the same cell strains exposed to benzo(a)pyrene (Gwinnet al., 2005).
The identification of genes involved in the process of neo-plastic transformation is essential for analyzing the pro-gression of breast cancer when induced by endogenous andexogenous agents, among which are the estrogens and theOP pesticides, respectively. Many studies have found anassociation between human cancer and exposure to OP pesti-cides. In a study of the effect of parathion as a ChE inhibitorand atropine as an antidote to AChE inhibitors it was shownthat parathion alone was able to induce increased cell pro-liferation, anchorage independence, and invasive capabilitiesof an immortalized human breast epithelial cell line, MCF-10F. In parallel, estrogen combined with either malathionor parathion could alter cell proliferation and induce celltransformation as well as significant invasive capabilities inthose cells. Several genes were upregulated by the effect ofall of the treatments, such as the cyclins, cyclin D1 andcyclin-dependent kinase 4, IGFBP3 and IGFBP5, and keratin18. The c-Ha-ras oncogene was upregulated by the effect ofmalathion alone and with the combination of estrogen andeither malathion or parathion. The DVL1 gene was upregu-lated with malathion alone and the combination of parathionwith estrogen. Expression of the Hsp 27, MCM2, and TP53inducible protein 3 genes was upregulated with malathionalone and with the combination of estrogen and eithermalathion or parathion, while TP53 (Li–Fraumeni syn-drome) was upregulated by estrogen alone and malathionalone. There was also an increase in c-kit, Trio, Rho-A,Rac-3, epidermal growth factor receptor (EGFR), Notch-4,Dvl-2, Ezrin, beta catenin, and mutant p53 protein expressionin the parathion-treated cells. However, atropine significantlyinhibited this increase. Among the genes affected were thecyclins, such as cyclin D3, the cyclin-dependent kinases(CDKs) such as CDK41, and the minichromosome main-tenance deficient (MCM) MCM2 and MCM3. It is suggestedthat pesticides alone or with estrogens affect humanbreast cells, inducing molecular changes indicative of trans-formation. Further, pesticides are an initiator factor in thetransformation process in breast cancer (Calaf and Roy,2007, 2008).
14.2.5 Cell Metabolism Components
Several studies have shown that OPs affect carbohydratemetabolism and induce metabolic pathways in brain, skeletalmuscles, and liver to produce hyperglycemia (Abdollahiet al., 2004a; Panahi et al., 2006; Pournourmohammadi
et al., 2005; Vosough-Ghanbari et al., 2007). As reviewedby Rahimi and Abdollahi (2007), OPs can influence bodyglucose homeostasis by several mechanisms, including phys-iological stress, oxidative stress, inhibition of paraoxonase,nitrosative stress, pancreatitis, inhibition of AChE, stimu-lation of adrenal gland, and disturbance in metabolism ofliver tryptophan (Rahimi and Abdollahi, 2007).
Pournourmohammadi et al. (2007) studied glucose-stimulated insulin secretion from freshly isolated pancreaticislets of rats pretreated with different concentrations ofmalathion. Their results show that hyperglycemia as an out-come of subchronic exposure to OPs is due to insulin resist-ance, disturbed insulin secretion, and damaged pancreaticLangerhans islets.
14.2.5.1 Enzymes Chronic exposure concerns are veryimportant especially considering the potential of OPs ininduction of diabetes. In the other side, the adverse effectsof the OP pesticide on glucose metabolism could be causedby enzyme activity or its expression variation. Glucokinaseis a tissue-specific enzyme expressed in liver and pancreaticbeta cells playing a crucial role in glycogen synthesis and glu-cose homeostasis. Pesticide affects pancreatic and hepaticglucokinase activity and expression differently, so that oneor three days of dichlorvos administration (20 mg/kg) inthe rat decreased glucokinase activity in the liver and, onthe contrary, glucokinase mRNA levels were increased. Incontrast, pancreatic glucokinase activity as well as mRNAlevels were not affected by the treatment. Insulin mRNAlevels were not modified by dichlorvos administration(Romero-Navarro et al., 2006).
Continuous and dominant use of chemical sprays likeinsecticides have facilitated development of resistance indifferent animals (Edwards and Myers 2007; Hemingwayet al., 2004). The cytochrome P450 (CYP) enzymefamily, together with the carboxylesterases and glutathioneS-transferases (GST), plays a central role in the detoxificationand in the metabolism of insecticides and other xenobiotics(Gilbert et al., 2005). Increase in tolerance to insecticides inmosquito Aedes albopictus is correlated with an inductionof CYP activity.
Further expression studies using real-time quantitativepolymerase chain reaction (PCR) confirmed that insecticidesand xenobiotics induced multiple CYP genes and one car-boxylesterase gene (Poupardin et al., 2008). GSTs make upa family of multifunctional enzymes that play an importantrole in detoxification of xenobiotic compounds contributingto insecticide resistance among insects, including mosquitoes(Brown 1986; Jirajaroenrat et al., 2001), and to multidrugresistance in tumor cell lines and cancer patients (Hayesand Pulford, 1995). Not all forms of resistance result fromgene amplification or compensatory mutation; rather, altera-tions of the structure and activity of the enzyme AChE leadto resistance to OP.
182 GENE EXPRESSION

Recently, microarray-based approaches have been used toinvestigate the effect of xenobiotic exposure on the expres-sion of detoxification genes. Hence, studying the inductionprofile of insect detoxification enzymes has been suggestedas a means to identify the major enzymes involved in insecti-cide detoxification.
Furthermore, the cross-induction of detoxificationenzymes involved in insecticide metabolism by environ-mental xenobiotics may enhance the tolerance to chemicalinsecticides. For example, it has been shown that mosquitoAedes aegypti larvae exposed to the herbicide Atrazinebecome more tolerant to the OP Temephos (Boyer et al.,2006). Similarly, exposure of mosquito Aedes albopictuslarvae to benzothiazole (a major leachate compound of auto-mobile tires) and pentachlorophenol (a wood-protectingagent) can increase their tolerance to CM, Rotenone, andTemephos (Suwanchaichinda and Brattsten, 2001). Ininsects, GSTs play an important role in OP resistance. Theplant bug Lygus lineolaris (Palisot de Beauvois) in cottonwith resistance to malathion had significantly higher(1.5-fold) GST activity than the susceptible strain; meanwhileGST gene transcripts were elevated in the resistant strain by1.3-fold. GST inhibitors like ethacrynic acid (EA) and diethylmaleate (DM) effectively could abolish resistance andincrease malathion toxicity against two resistant strains bymore than two- and threefold, whereas incorporation ofGST inhibitors did not significantly increase malathion tox-icity against a susceptible strain (Zhu et al., 2007). To identifythe role of GST in detoxification of OP, it has been reportedthat GST-mediated O-dealkylation occurs in cytosolicfractions isolated from rat, mouse, and human livers whichbiotransforms methylparathion (MeP). MeP O-dealkylationactivities of several purified recombinant GSTs belonging tothe alpha (human GSTs A1-1 and A2-2, mouse GSTA3-3, ratGSTA5-5), mu (human GSTs M1a-1a, M2-2, M3-3, M4-4),pi (human GSTP1-1, mouse GSTs P1-1, P2-2), and theta(human GSTT1-1) classes showed that in the presence of glu-tathione as a substrate, hGSTT1-1 and hGSTA1-1 exhibitedthe highest MeP O-dealkylation activities while expressionlevel is elevated, so hGSTA1-1 is responsible for most ofthe MeP O-dealkylation in human hepatic cytosol. In targetorgans such as brain and skeletal muscle, where hGSTT1-1is expressed, hGSTT1-1-mediated biotransformation ofMeP may be important (Abel et al., 2004). In Anopheles mos-quitoes, one of the GST genes, adgst1AS1, codes for (at least)four RNA splice variants that vary in their binding character-istics with regard to permethrin, a pyrethroid insecticide(Jirajaroenrat et al., 2001). Alternative RNA splicing couldexplain the rapid increase in permethrin resistance associatedwith GST upregulation observed among Culex mosquitoesselected for just one or three generations (Xu et al., 2005).In the other hand, alterations of the structure and activity ofthe enzyme AChE led to resistance to OPs in the orientalfruit fly, Bactrocera dorsalis (Hendel), a pest of great
economic importance in the Asia-Pacific region. The AChEenzyme from a strain selected for resistance to the insecticidefenitrothion shows substantially lower catalytic efficiency forvarious substrates and less sensitivity to inhibition by para-oxon, eserine, and fenitroxon, compared to that of the feni-trothion susceptible line. Mutations in the AChE gene ofBactrocera dorsalis were associated with resistance to OPinsecticides. The expression levels of AChE genes were notsignificantly different between the resistant and sensitivelines, indicating that quantitative changes in gene expressionwere not contributing significantly to the resistance pheno-type, supporting a direct causal relationship between themutations previously identified in AChE gene of this speciesand qualitative alterations of the structure and function of theAChE enzyme as the basis for the resistance phenotype (Hsuet al., 2008).
In vitro functional expression of AChE in Sf9 cellsrevealed that only resistant AChE1 is less inhibited with para-oxon, suggesting that resistant AChE1 is responsible forprothiofos resistance (Lee et al., 2007). CYP1A1 expressionin hepatocytes treated for three days with increasing nontoxicdoses of insecticides showed a strong and dose-dependentinduction with tetrachlorvinfos, fivefold in human andsevenfold in rat hepatocytes. However, CYP1A1 mRNAlevels were not correlated for tetrachlorvinfos, carbarylexerted less induction in both cell types, whereas malathionshowed no action on human hepatocytes (De Sousaet al., 1997).
14.2.5.2 P-Glycoprotein Other detoxification systems incells are efflux pumps, such as P-glycoprotein (P-gp), andserve both as natural defense mechanisms and to influencethe bioavailability and disposition of drugs. P-gp expressionand activity could be increased after the repeated exposure ofintestinal cell line, Caco-2 cells, to diazinon, suggesting theinvolvement of P-gp in the transfer of diazinon, leadingto potential consequences for xenobiotic interactions.Oral administration of diazinon (2 to 20 mg/kg, 5 days, or10 mg/kg, 2 to 12 days) increased intestinal mdr1a mRNAof rats, in both a dose- and time-dependent manner, increasedthe expression of intestinal P-gp, and showed that repeatedexposure to low doses of pesticide may lead to upregulatedP-gp functions in the intestine of mammals (Lecoeuret al., 2006).
14.2.6 Cell Stress Components
14.2.6.1 Oxidative Stress There are some reports thatshow oxidative stress and depressed AChE activity in bio-logic samples obtained from OP-formulating pesticideworkers, and correlation of genotoxicity and oxidativestress has led to the proposal of some mechanisms involvedin the ability of OP and CM compounds to induce freeradical damage (Abdollahi et al., 2004b; Ranjbar et al.,
14.2 INSECTICIDE-INDUCED GENE EXPRESSION ALTERATION IN DIFFERENT TISSUES 183

2002; Shadnia et al., 2005; Soltaninejad and Abdollahi,2009). Also, many studies on animals have revealed therole of oxidative stress in toxicity of anticholinesterase com-pounds. Most of these studies confirmed the role of OPs inaffecting body glucose metabolism and induction ofoxidative stress (Abdollahi et al., 2004a; Akhgari et al.,2003; Ghafour-Rashidi et al., 2007; Panahi et al., 2006;Pournourmohammadi et al., 2005, 2007; Rahimi et al., 2005).
14.2.6.2 Heat Shock Proteins Experimental evidence inpast years has confirmed that stress proteins play a vigorousrole in cellular defense. Heat shock proteins (Hsps) displaywide-ranging cellular functions such as protection fromstress-induced injury. Induction of Hsps in response to stres-sors is one of the first physiological events that defend cellsfrom the subsequent injury, and Hsps have been proposedas a biomarker of exposures to environmental toxicants,including certain chemical pesticides (Feige et al., 1996;Werner, 2001).
The stress proteins are believed to maintain the cell pro-teins in the correct conformation by aiding them to be prop-erly translocated to different organelles in the cell, and byguiding the newly formed as well as denatured proteins toachieve their proper conformation, thereby preventingmisfolding or aggregation of misfolded protein and proteo-lytic degradation of unstable proteins, or catalyzing properfolding of nascent polypeptides to their native functionalstate, thus protecting the cells from further damage (Federand Hofmann 1999; Nover 1991).
Among the Hsps, Hsp70 is one of the most commonlyexpressed stress proteins induced by OPs which provokeoxidative tissue damage and lipid peroxidation (Nadeauet al., 2001). Free radicals may modulate Hsp70 expressionand apoptosis, and consequently antioxidant defenseenzymes in the exposed organism. A concentration- andtime-dependent significant increase in reactive oxygenspecies (ROS) generation, along with significant upregula-tion of Hsp70, preceded changes in antioxidant defenseenzyme activities, contents of glutathione and malondialde-hyde level on third instar larvae of Drosophila melanogasterfed with dichlorvos at concentrations between 0.015 and15 ppb. An inhibitory effect on superoxide dismutase andcatalase activities significantly upregulated ROS generationand Hsp70 expression in the exposed organism, while inhi-bition of Hsp70 significantly affected oxidative stress markersinduced by dichlorvos. A comparison made among ROSgeneration, Hsp70 expression, and apoptotic markers showedthat ROS generation is positively correlated with Hsp70expression and apoptotic cell death end points, indicatinginvolvement of ROS in the overall adversity caused bydichlorvos to the organism.
Hsp70 expression was restricted only within the testislobes of male fly while it was not induced in the ovary ofthe female. Further, sex peptide (Acp70A) and accessory
gland protein (Acp36DE) expression, required to facilitatenormal reproduction, were significantly downregulated atthe higher concentrations, causing dose-dependent signi-ficant reduction of reproductive outcome. Dichlorvos ismore deleterious to fly reproduction than chlorpyrifos(Gupta et al., 2007b).
In blowflies Lucilia cuprina, whose heat shock Hsp70 andHsp60 responses have been well characterized, it has beenshown that in vitro exposure of larval and adult tissues todifferent concentrations of the OP Monocrotophos (0.0005to 0.005 ppm for larva and 0.0001 to 0.001 ppm for adult)induces the expression of Hsp60 and Hsp70 proteins. Theresponse was tissue specific in that gut tissues were relativelymore sensitive to pesticide toxicity than other tissues and,hence, they might serve as primary targets for early detectionof pesticide toxicity. Overall, the pattern of expression showsa positive correlation between the levels of pesticide-inducedHsp60 and Hsp70 and the tissue injury (Sharma et al., 2008).In the same way, expression of Hsp70 in the third-instar larvaltissues of transgenic Drosophila melanogaster (Hsp70-lacZ)following dietary exposure to the OP insecticide chlorpyrifosshowed an intense cytotoxic effect in salivary gland, proven-triculus, brain ganglia, and midgut at the lowest concentrationof the chemical following 24 and 48 h of exposure. Onthe other hand, a significant increase in Hsp70 expressionin the above larval tissues was observed in the next higherconcentration of CP after 6 h exposure when compared withthat of the lowest dose. A further increase in exposuretime caused 100% larval mortality. Further, reproductiveperformance was significantly affected even in the lowestdosed group. Nontarget larval tissues of Drosophila are vul-nerable to chlorpyrifos as evidenced by Hsp70 expression.Meanwhile, chlorpyrifos could alter expression of somestress genes in other animals akin to expression of Hsp 89aand Hsp 89b genes in hepatic and brain tissues of rats, aswell as enhancing Hsp 90 synthesis in cultured PC-12 cellsso heat shock protein, metallothionein, vitellogenin, andC. elegans p53-like protein genes on exposed soil nematodeCaenorhabditis elegans (Bagchi et al., 1996; Nazir et al.,2001; Roh and Choi, 2008).
On the other hand, Hsps have been involved in resistanceto OP in some animals as shown on the aquatic midgeChironomus yoshimatsui which is resistant to fenitrothion.The expression of Hsp70 gene in their larvae collectedabout one week after fenitrothion spraying in the fields wastwo- or threefold higher than that in the susceptible strainand was also much higher than those collected three monthsafter insecticide spraying ended (Yoshimi et al., 2002).
14.2.7 Other Components
14.2.7.1 Reproductive System Cholinesterases are ubi-quitous carboxylesterase type B enzymes capable of hydro-lyzing the neurotransmitter acetylcholine which are
184 GENE EXPRESSION

transiently expressed in multiple germline and embryoniccells, and have important roles in multiple cell types, sothat the amplification and overexpression of their corre-sponding genes might affect fertility (Soreq and Zakut,1990). Even though there is extreme variation in the anatom-ical and morphological organization of the male reproductivesystem in different species, it is characterized by similar pat-terns of male-dependent carboxylesterase overexpressioninduced by environmental reproductive xenobiotics. Therole of carboxylesterases in detoxification of environmentalchemicals such as OP pesticides suggests that various malegenital tract carboxylesterases may be characterized by asimilar physiological function, which protects the male repro-ductive system against xenobiotics and thus influences andprovokes its dysfunction resulting in sperm differentiationand maturation (Deakin and James, 2004). Carbaryl isknown to obstruct female reproductive function; however,the mechanisms through which the adverse effects aremediated are not clear. There is a report showing an inhibitionof basal and follicle-stimulating hormone (FSH)-inducedprogesterone production in primary human granulosa-luteincells (hGLCs) co-incubated with various concentrations ofcarbaryl at 0, 1, 5, 25, and 125 mmol/L in a dose-dependentmanner. Decreased progesterone biosynthesis was accom-panied by reduced cAMP loads in both basal and FSH-induced conditions.
Carbaryl could inhibit the gene expression of steroido-genic acute regulatory protein (StAR) as well as steroidogen-esis, at least in part, by obstructing the delivery of cholesterolover mitochondrial membranes and attenuating cAMP gener-ation (Cheng et al., 2006). Besides CM, the adverse effects ofOP compounds on reproduction system, dichlorvos is moredeleterious to fly reproduction compared to chlorpyrifos,with an adverse effect on Acp70A and Acp36DE expressionrequired to facilitate normal reproduction in the first instarlarvae of Drosophila melanogaster which caused a dose-dependent reduction in reproductive outcome, althoughmales seemed to be more sensitive than females (Guptaet al., 2007a).
14.2.7.2 Neuropathy Target Esterase Certain OPs reactwith the active-site serine residue of neuropathy target ester-ase (NTE) which is phosphorylated and aged and then causesaxonal degeneration and paralysis, and induces delayedneuropathy in human and some animals. NTE has been pro-posed to play a role in neurite outgrowth and processelongation during neural differentiation. The all-trans retinoicacid (ATRA)-induced differentiation of the neuroblastomacells with lowered NTE activity revealed that inhibition ofNTE expression does not affect neural differentiation inhuman neuroblastoma SK-N-SH cells, suggesting that OPsmay inhibit neural differentiation by initially acting on targetsother than NTE (Chang et al., 2005).
14.3 CONCLUSION
In reality, chlorpyrifos, the best-studied agent, affects braindevelopment through diverse targets such as oxidativestress, cell signaling cascades, expression and function ofnuclear transcription factors, and neuronal–glial cell inter-actions. Because many of the factors have overlapping func-tions, it is important to mention that alterations in expressionor function of any one factor may not necessarily lead to nastymorphological deficits but rather to more slight defects.
Overall, studies indicate that insecticides are potentialinducers of Hsps which could serve as potential biomarkersfor assessment and monitoring of toxicity induced by pesti-cides, even as sensitive indicators of low-level exposures tocertain insecticides, suggesting that these genes may bemechanistically involved in protecting tissues against oxi-dative stress induced by structurally diverse pesticides.Additionally, Hsp70 and antioxidant enzymes worktogether for cellular defense against pesticides.
Different mechanisms are involved in the resistance toOP and CM insecticides like insensitive AChE, GSTenzyme, P450 cytochrome oxidase family, and so on. Datasuggest that there are different regulatory mechanisms forCYP1A1 gene expression by different insecticides.
All studies point to the need to screen the various OPsfor similarities and differences in their targeting of the keypathways that contribute to their critical harmful outcomes.
REFERENCES
Abdollahi M, Donyavi M, Pournourmohammadi S, Saadat M(2004a) Hyperglycemia associated with increased hepaticglycogen phosphorylase and phosphoenolpyruvate carboxy-kinase in rats following subchronic exposure to malathion.Comp Biochem Physiol C Toxicol Pharmacol 137: 343–347.
Abdollahi M, Ranjbar A, Shadnia S, Nikfar S, Rezaiee A (2004b)Pesticides and oxidative stress: a review. Med Sci Monit 10:RA141–147.
Abel EL, Bammler TK, Eaton DL (2004) Biotransformation ofmethyl parathion by glutathione S-transferases. Toxicol Sci 79:224–232.
Akhgari M, Abdollahi M, Kebryaeezadeh A, Hosseini R,Sabzevari O (2003) Biochemical evidence for free radical-induced lipid peroxidation as a mechanism for subchronic tox-icity of malathion in blood and liver of rats. Hum Exp Toxicol22: 205–211.
Bagchi D, Bhattacharya G, Stohs SJ (1996) In vitro and in vivoinduction of heat shock (stress) protein (Hsp) gene expressionby selected pesticides. Toxicology 112: 57–68.
Betancourt AM, Burgess SC, Carr RL (2006) Effect of developmen-tal exposure to chlorpyrifos on the expression of neurotrophingrowth factors and cell-specific markers in neonatal rat brain.Toxicol Sci 92: 500–506.
REFERENCES 185

Betancourt AM, Filipov NM, Carr RL (2007) Alteration of neuro-trophins in the hippocampus and cerebral cortex of young ratsexposed to chlorpyrifos and methyl parathion. Toxicol Sci 100:445–455.
Blasiak J, Jaloszynski P, Trzeciak A, Szyfter K (1999) In vitrostudies on the genotoxicity of the organophosphorus insecticidemalathion and its two analogues. Mutat Res 445: 275–283.
Bolognesi C (2003) Genotoxicity of pesticides: a review of humanbiomonitoring studies. Mutat Res 543: 251–272.
Boyer S, Serandour J, Lemperiere G, Raveton M, Ravanel P (2006)Do herbicide treatments reduce the sensitivity of mosquito larvaeto insecticides? Chemosphere 65: 721–724.
Brown AW (1986) Insecticide resistance in mosquitoes: a pragmaticreview. J Am Mosq Control Assoc 2: 123–140.
Calaf GM, Roy D (2007) Gene expression signature of parathion-transformed human breast epithelial cells. Int J Mol Med 19:741–750.
Calaf GM, Roy D (2008) Cancer genes induced by malathion andparathion in the presence of estrogen in breast cells. Int J MolMed 21: 261–268.
Caughlan A, Newhouse K, Namgung U, Xia Z (2004) Chlorphyrifosinduces apoptosis in rat cortical neurons that is regulated bya balance between p38 and ERK/JNK MAP kinases. ToxicolSci 78: 125–134.
Chang PA, Wu YJ, Chen R, Li M, Li W, Qin QL (2005) Inhibition ofneuropathy target esterase expressing by antisense RNA does notaffect neural differentiation in human neuroblastoma (SK-N-SH)cell line. Mol Cell Biochem 272: 47–54.
Cheng S, Chen J, Qiu Y, Hong X, Xia Y, Feng T, Liu J, Song L,Zhang Z, Wang X (2006) Carbaryl inhibits basal and FSH-induced progesterone biosynthesis of primary human granu-losa-lutein cells. Toxicology 220: 37–45.
Damodaran TV, Abou-Donia MB (2000) Alterations in levels ofmRNAs coding for glial fibrillary acidic protein (GFAP) andvimentin genes in the central nervous system of hens treatedwith diisopropyl phosphorofluoridate (DFP). Neurochem Res25: 809–816.
Damodaran TV, Abdel-Rahman A, Abou-Donia MB (2001) Alteredtime course of mRNA expression of alpha tubulin in the centralnervous system of hens treated with diisopropyl phosphoro-fluoridate (DFP). Neurochem Res 26: 43–50.
Damodaran TV, Abdel-Rahman A, El-Sourady MH, Abou-DoniaMB (2002) Differential alteration of glyceraldehyde-3-phosphatedehydrogenase (GAPDH) mRNA in the central nervous systemof hens treated with diisopropylphosphorofluoridate (DFP).Neurochem Int 40: 371–379.
Deakin SP, James RW (2004) Genetic and environmental factorsmodulating serum concentrations and activities of the antioxi-dant enzyme paraoxonase-1. Clin Sci (London) 107: 435–447.
De Sousa G, Fontaine F, Pralavorio M, Botta-Fridlund D, Letreut Y,Rahmani R (1997) Insecticide cytotoxicity and CYP1A1/2induction in primary human and rat hepatocyte cultures.Toxicol In Vitro 11: 451–457.
Dunican DS, McWilliam P, Tighe O, Parle-McDermott A, CrokeDT (2002). Gene expression differences between the micro-satellite instability (MIN) and chromosomal instability (CIN)
phenotypes in colorectal cancer revealed by high-densitycDNA array hybridization. Oncogene 21(20):3253–3257.[Erratum in Oncogene 2002, 21(51):7912.]
Edwards TM, Myers JP (2007) Environmental exposures and generegulation in disease etiology. Environ Health Persp 115:1264–1270.
Ellison V, Stillman B (2003) Biochemical characterization ofDNA damage checkpoint complexes: clamp loader and clampcomplexes with specificity for 50 recessed DNA. PLoS Biol1(2): E33.
Feder ME, Hofmann GE (1999) Heat-shock proteins, molecularchaperones, and the stress response: evolutionary and ecologicalphysiology. Annu Rev Physiol 61: 243–282.
Feige U, Morimoto RI, Yahara I, Polla BS (1996) Stress InducibleCellular Responses. Boston: Birkhauser-Verlag.
Geigl JB, Langer S, Barwisch S, Pfleghaar K, Lederer G, SpeicherMR (2004) Analysis of gene expression patterns and chromoso-mal changes associated with aging. Cancer Res 64: 8550–8557.
Ghafour-Rashidi Z, Dermenaki-Farahani E, Aliahmadi A,Esmaily H, Mohammadirad A, Ostad SN, Abdollahi M(2007) Protection by cAMP and cGMP phosphodiesteraseinhibitors of diazinon-induced hyperglycemia and oxidative/nitrosative stress in rat Langerhans islets cells: molecular evi-dence for involvement of non-cholinergic mechanisms. PesticBiochem Physiol 87: 261–270.
Gilbert LI, Iatrou K, Gill S (2005) Comprehensive Molecular InsectScience. New York: Elsevier.
Gupta RP, Bing G, Hong JS, Abou-Donia MB (1998) cDNA clon-ing and sequencing of Ca2þ/calmodulin dependent proteinkinase II subunit and its mRNA expression in diisopropyl phos-phorofluoridate (DFP)-treated hen central nervous system. MolCell Biochem 181: 29–39.
Gupta RP, Damodaran TV, Abou-Donia MB (2000) C-fos mRNAinduction in the central and peripheral nervous systems of diiso-propyl phosphorofluoridate (DFP)-treated hens. Neurochem Res25: 327–334.
Gupta SC, Siddique HR, Mathur N, Mishra RK, Saxena DK,Chowdhuri DK (2007a) Adverse effect of organophosphatecompounds, dichlorvos and chlorpyrifos in the reproductivetissues of transgenic Drosophila melanogaster: 70 kDa heatshock protein as a marker of cellular damage. Toxicology 238:1–14.
Gupta SC, Siddique HR, Mathur N, Vishwakarma AL, Mishra RK,Saxena DK, Chowdhuri DK (2007b) Induction of hsp70,alterations in oxidative stress markers and apoptosis againstdichlorvos exposure in transgenic Drosophila melanogaster:modulation by reactive oxygen species. Biochim Biophys ActaGeneral Subjects 1770: 1382–1394.
Gwinn MR, Whipkey DL, Weston A (2004) The effect of oxythio-quinox exposure on normal human mammary epithelial cell geneexpression: a microarray analysis study. Environ Health 3: 9.
Gwinn MR, Whipkey DL, Tennant LB, Weston A (2005)Differential gene expression in normal human mammaryepithelial cells treated with malathion monitored by DNA micro-arrays. Environ Health Persp 113: 1046–1051.
186 GENE EXPRESSION

Hayes JD, Pulford DJ (1995) The glutathione S-transferasesupergene family: regulation of GST and the contribution ofthe isoenzymes to cancer chemoprotection and drug resistance.Crit Rev Biochem Mol 30: 445–600.
Hemingway J, Hawkes NJ, McCarroll L, Ranson H (2004) The mol-ecular basis of insecticide resistance in mosquitoes. InsectBiochem Mol Biol 34: 653–665.
Hsu JC, Wu WJ, Haymer DS, Liao HY, Feng HT (2008) Alterationsof the acetylcholinesterase enzyme in the oriental fruit flyBactrocera dorsalis are correlated with resistance to the organo-phosphate insecticide fenitrothion. Insect Biochem Mol Biol 38:146–154.
Jirajaroenrat K, Pongjaroenkit S, Krittanai C, Prapanthadara L,Ketterman AJ (2001) Heterologous expression and characteriz-ation of alternatively spliced glutathione S-transferases from asingle Anopheles gene. Insect Biochem Mol Biol 31: 867–875.
Lecoeur S, Videmann B, Mazallon M (2006) Effect of organopho-sphate pesticide diazinon on expression and activity of intestinalP-glycoprotein. Toxicol Lett 161: 200–209.
Lee DW, Choi JY, Kim WT, Je YH, Song JT, Chung BK, Boo KS,Koh YH (2007) Mutations of acetylcholinesterase1 contribute toprothiofos-resistance in Plutella xylostella (L.). Biochem BiophysRes Commun 353: 591–597.
Mankame T, Hokanson R, Fudge R, Chowdhary R, Busbee D(2006) Alteration of gene expression in human cells treatedwith the agricultural chemical diazinon: possible interaction infetal development. Hum Exp Toxicol 25: 225–233.
Nadeau D, Corneau S, Plante I, Morrow G, Tanguay RM (2001)Evaluation for Hsp70 as a biomarker of effect of pollutants onthe earthworm Lumbricus terrestris. Cell Stress Chaperon 6:153–163.
Nazir A, Mukhopadhyay I, Saxena DK, Chowdhuri DK (2001)Chlorpyrifos-induced hsp70 expression and effect on reproduc-tive performance in transgenic Drosophila melanogaster(hsp70-lacZ) Bg9. Arch Environ Contam Toxicol 41: 443–449.
Nover L (1991) Heat Shock Response. Boca Raton, FL: CRC Press.
Palackal NT, Lee SH, Harvey RG, Blair IA, Penning TM (2002)Activation of polycyclic aromatic hydrocarbon trans-dihydrodiolproximate carcinogens by human aldo-keto reductase (AKR1C)enzymes and their functional overexpression in human lungcarcinoma (A549) cells. J Biol Chem 277(27):24799–24808.
Panahi S, Vosough-Ghanbari S, Pournourmohammadi S, Ostad SN,Nikfar S, Minaie B, Abdollahi M (2006) Stimulatory effects ofmalathion on the key enzyme activities of insulin secretion inLangerhans islets, glutamate dehydrogenase and glucokinase.Toxicol Mech Methods 16: 161–167.
Penning TM, Ohnishi ST, Ohnishi T, Harvey RG (1996) Generationof reactive oxygen species during the enzymatic oxidation ofpolycyclic aromatic hydrocarbon trans-dihydrodiols catalyzedby dihydrodiol dehydrogenase. Chem Res Toxicol 9: 84–92.
Poupardin R, Reynaud S, Strode C, Ranson H, Vontas J, David JP(2008) Cross-induction of detoxification genes by environmentalxenobiotics and insecticides in the mosquito Aedes aegypti:impact on larval tolerance to chemical insecticides. InsectBiochem Mol Biol 38: 540–551.
Pournourmohammadi S, Farzami B, Ostad SN, Azizi E,Abdollahi M (2005) Effects of malathion subchronic exposureon rat skeletal muscle glucose metabolism. Environ ToxicolPharm 19: 191–196.
Pournourmohammadi S, Ostad SN, Azizi E, Ghahremani MH,Farzami B, Minaie B, Larijani B, Abdollahi M (2007)Induction of insulin resistance by malathion: evidence for dis-rupted islets cells metabolism and mitochondrial dysfunction.Pestic Biochem Physiol 88: 346–352.
Rahimi R, Abdollahi M (2007) A review on the mechanismsinvolved in hyperglycemia induced by organophosphorus pesti-cides. Pestic Biochem Physiol 88: 115–121.
Rahimi R, Nikfar S, Larijani B, Abdollahi M (2005) A review onthe role of antioxidants in the management of diabetes and itscomplications. Biomed Pharmacother 59: 365–373.
Ranjbar A, Pasalar P, Abdollahi M (2002) Induction of oxidativestress and acetylcholinesterase inhibition in organophospho-rous pesticide manufacturing workers. Hum Exp Toxicol 21:179–182.
Rath NC, Huff WE, Huff GR (2007) Thiram-induced changes inthe expression of genes relating to vascularization and tibialdyschondroplasia. Poult Sci 86(11):2390–2395.
Roh JY, Choi J (2008) Ecotoxicological evaluation of chlorpyrifosexposure on the nematode Caenorhabditis elegans. EcotoxEnviron Safe 71: 483–489.
Romani R, Isani G, De Santis A, Giovannini E, Rosi G (2005)Effects of chlorpyrifos on the catalytic efficiency and expressionlevel of acetylcholinesterases in the bivalve mollusk Scapharcainaequivalvis. Environ Toxicol Chem 24: 2879–2886.
Romero-Navarro G, Lopez-Aceves T, Rojas-Ochoa A, FernandezMejia C (2006) Effect of dichlorvos on hepatic and pancreaticglucokinase activity and gene expression, and on insulinmRNA levels. Life Sci 78: 1015–1020.
Shadnia S, Azizi E, Hosseini R, Khoei S, Fouladdel S, PajoumandA, Jalali N, Abdollahi M (2005) Evaluation of oxidative stressand genotoxicity in organophosphorus insecticide formulators.Hum Exp Toxicol 24: 439–445.
Sharma S, Rohilla MS, Reddy PVJ, Tiwari PK (2008) In vitroinduction of 60-kDa and 70-kDa heat shock proteins by endo-sulphan and monocrotophos in sheep blowfly Lucilia cuprina.Arch Environ Contam Toxicol 55: 57–69.
Slotkin TA, Seidler FJ (2007) Comparative developmentalneurotoxicity of organophosphates in vivo: transcriptionalresponses of pathways for brain cell development, cell signaling,cytotoxicity and neurotransmitter systems. Brain Res Bull 72:232–274.
Slotkin TA, Seidler FJ, Fumagalli F (2008) Targeting of neuro-trophic factors, their receptors, and signaling pathways in thedevelopmental neurotoxicity of organophosphates in vivo andin vitro. Brain Res Bull 76: 424–438.
Soltaninejad K, Abdollahi M (2009) Current opinion on the scienceof organophosphate pesticides and toxic stress: a systematicreview. Med Sci Monit 15: 75–90.
Soreq H, Zakut H (1990) Amplification of butyrylcholinesteraseand acetylcholinesterase genes in normal and tumor tissues:
REFERENCES 187

putative relationship to organophosphorous poisoning. PharmRes 7: 1–7.
Suwanchaichinda C, Brattsten LB (2001) Effect of exposure topesticides on carbaryl toxicity and cytochrome P450 activitiesin Aedes albopictus larvae (Diptera: Culicidae). PesticBiochem Physiol 70: 63–73.
Testa JR, Zhou JY, Bell DW, Yen TJ (1994) Chromosomal localiz-ation of the genes encoding the kinetochore proteins CENPE andCENPF to human chromosomes 4q24! q25 and 1q32! q41,respectively, by fluorescence in situ hybridization. Genomics23(3):691–693.
Trinh BN, Ong CN, Coetzee GA, Yu MC, Laird PW (2002)Thymidylate synthase: a novel genetic determinant of plasmahomocysteine and folate levels. Hum Genet 111(3):299–302.
Vosough-Ghanbari S, Sayyar P, Pournourmohammadi S, AliahmadiA, Ostad SN, Abdollahi M (2007) Stimulation of insulin andglucagon synthesis in rat Langerhans islets by malathion invitro: evidence for mitochondrial interaction and involvementof subcellular non-cholinergic mechanisms. Pestic BiochemPhysiol 89: 130–136.
Werner IK, Koger CS, Hamm JT, Hinton DE (2001) Ontogeny ofthe heat shock protein, hsp70 and hsp60, response and
developmental effects of heat-shock in the teleost, medaka(Oryzias latipes) Environ Sci 8: 13–30.
Windham GC, Titenko-Holland N, Osorio AM, Gettner S,Reinisch F, Haas R, Smith M (1998) Genetic monitoring ofmalathion-exposed agricultural workers. Am J Ind Med 33:164–174.
Xu Q, Liu H, Zhang L, Liu N (2005) Resistance in the mosquito,Culex quinquefasciatus, and possible mechanisms for resistance.Pest Manag Sci 61(11):1096–1102.
Yih LH, Lee TC (2003) Induction of C-anaphase and diplochro-mosome through dysregulation of spindle assembly check-point by sodium arsenite in human fibroblasts. Cancer Res63(20):6680–6688.
Yoshimi T, Minowa K, Karouna-Renier NK, Watanabe C, SugayaY, Miura T (2002) Activation of a stress-induced gene by insec-ticides in the midge, Chironomus yoshimatsui. J Biochem MolToxicol 16: 10–17.
Zhu YC, Snodgrass GL, Chen MS (2007) Comparative study onglutathione S-transferase activity, cDNA, and gene expressionbetween malathion susceptible and resistant strains of thetarnished plant bug, Lygus lineolaris. Pestic Biochem Physiol87: 62–72.
188 GENE EXPRESSION