anti-idiotype monoclonal antibody - pnas. natl. acad. sci. usa vol. 89, pp. 2546-2550, april 1992...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 89, pp. 2546-2550, April 1992Immunology
Anti-idiotype monoclonal antibody elicits broadly neutralizinganti-gpl20 antibodies in monkeys
(anti-idiotype antibody/neutralizing antibody/human immunodeficiency virus vaccine)
CHANG-YUIL KANG*t, PETER NARAt, SOULAIMA CHAMAT*, VINCE CARALLI*, AGNES CHEN*,MAI-LAN NGUYEN*, HIRONORI YOSHIYAMA§, W. J. W. MORROW*, DAVID D. Ho§, AND HEINZ K6HLER**IDEC Pharmaceuticals Corporation, La Jolla, CA 92037; *The National Cancer Institute, Frederick, MD 21701; and §The Aaron Diamond AIDS ResearchCenter, New York, NY 10016
Communicated by Alfred Nisonoff, December 26, 1991 (received for review October 17, 1991)
ABSTRACT Murine monoclonal antibodies (mAbs) wereraised against human, polyclonal, anti-gpl20 antibodies (Abl)and were selected for binding to broadly neutralizing anti-gpl20 antibodies in sera positive for human immunodeficiencyvirus (HIV). One anti-idiotype mAb (Ab2), 3C9, was found tobe specific for human anti-gpl20 antibodies directed against anepitope around the conserved CD4 attachment site of gpl20.The 3C9 reactive human anti-gpl20 antibodies (3C9' Ab)neutralized MN, huB, RF, and four primary isolates of HIVtype 1 (HIV-1). Cynomolgus monkeys were immunized with3C9 in adjuvant to test whether this anti-idiotype mAb couldinduce neutralizing anti-gpl20 antibodies. The results showthat purified anti-anti-idiotype antibodies (Ab3) from 3C9immune sera bind to an epitope around the CD4 attachmentsite of gpl2OSF and gpl2OmII. Furthermore, purified gpl20-specific Ab3 neutralize MN, fIB, and RF isolates. Theseresults demonstrate that primates immunized with an anti-idiotype mAb produce broadly neutralizing anti-HIV-1 anti-bodies. Since this anti-idiotype mAb was selected by identifyinga clonotypic marker, its biological activity can be explained asthe result of clonotypic B-cell stimulation.
The immunodominant neutralizing site against human immu-nodeficiency virus type 1 (HIV-1) resides in the third hyper-variable region (V3) of gpl20. The hypervariability of theamino acid composition outside and within this region sug-gests that HIV may be able to escape the effects of V3-specific neutralizing antibodies by mutation (1). The secondmajor neutralizing epitope of gpl20 is the CD4 attachmentsite. A recent report has demonstrated the conformationaldependence of this region (2). Furthermore, our study hasshown that several epitopes reside around the CD4 attach-ment site (3). Antibodies specific for these epitopes generallyexhibit broad neutralizing activities against multiple strains ofHIV-1 (2-5). Thus, the CD4 attachment site of gpl20 is anattractive target for immunotherapy as the problems ofantigenic variation may be minimized. However, the naturalantibody response to this region is weaker, at least in earlyinfection (1, 2), and the neutralizing activities of the CD4site-specific antibodies are less potent than that of V3-specific antibodies (6).
In the present study, we explored the use of anti-idiotypeantibodies, to induce CD4 site-directed, broadly neutralizingantibodies in nonhuman primates. The reason for using thisapproach was to overcome the limiting immunogenic poten-tial of the CD4 epitopes. We believed that the inherentantigenically imposed restriction associated with virus-basedvaccines could be overcome by using antibodies directedagainst idiotopes on B-cell receptors with specificity for the
CD4 attachment site of gpl20. Targeting such B cells withspecific anti-idiotype (or anti-clonotype) antibodies maystimulate the synthesis of specific antibodies. In this regard,we utilized broadly neutralizing human polyclonal anti-gpl20antibodies as template idiotype antibody (Abl) to generatemurine anti-idiotype monoclonal antibodies (Ab2 mAbs),since it was believed that this approach will increase thechance of generating anti-anti-idiotype antibodies (Ab3) inhumans that are close to Abls in terms of clonality andspecificity. One anti-idiotype mAb, 3C9, that interacted withCD4 site-specific, broadly neutralizing antibodies was se-lected and tested in cynomolgus monkeys. The data indicatethat this anti-idiotype mAb induced broadly neutralizinganti-gp120 antibodies in nonhuman primates.
MATERIALS AND METHODSImmunizations. Four cynomolgus monkeys received intra-
muscular injections with 2.5 mg of3C9 in 1 ml ofSAF (Syntexadjuvant formulation) containing 0.6 mg of N-acetylmuram-yl-L-alanyl-D-isoglutamine. The injections were administeredbiweekly. Animals were bled before the injection series and7 days after each injection.Recombinant Proteins. gpl20IIIB is a recombinant glycopro-
tein of a HIV111B isolate that is secreted from a Drosophila cellline. The gpl2Om1B was received from SmithKline Beecham.gp120sF2 is a recombinant envelope glycoprotein ofan HIVs5isolate that is secreted from Chinese hamster ovary (CHO)cells. gp120sF2 was received from Chiron. Recombinant sol-uble CD4 (sCD4) was prepared by transfection of a truncatedCD4 gene into CHO cells. Secreted sCD4 in culture superna-tants of CHO cells (representing 95% of the amino-terminalextracellular portion of the molecule) was purified on ananti-CD4 antibody-conjugated Sepharose column.
Affinity Purification of Antibodies. All affinity columnswere prepared by conjugating 8 mg of protein with 5 ml ofCNBr-activated Sepharose 4B (Pharmacia). To purify the3C9-reactive antibody (3C9' Ab) from pooled sera of healthyHIV-infected individuals (6), heat (560C for 1 hr)- and deter-gent (1% Nonidet P40)-treated sera were applied to a 3C9-conjugated Sepharose affinity column. After being washedextensively, bound antibodies (3C9' Abs) were eluted withcitrate buffer (pH 2.8) and dialyzed against phosphate-buffered saline (PBS; pH 7.2). To purify various Ab3s frommonkey sera, all 3C9 immune sera were initially diluted 1:1with PBS. The diluted serum was then passed over an affinitycolumn of normal mouse immunoglobulin-conjugated Seph-
Abbreviations: Abl, idiotype antibody; Ab2, anti-idiotype antibody;Ab3, anti-anti-idiotype antibody; V3, the third variable region ofgpl20; sCD4, soluble CD4; mAb, monoclonal antibody; BSA, bovineserum albumin.tTo whom reprint requests should be addressed at: IDEC Pharma-ceuticals Corporation, 11099 North Torrey Pines Road, La Jolla,CA 92037.
2546
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 89 (1992) 2547
arose. The flow-through fraction was collected and set aside.The bound antibody (anti-isotype Ab) was washed, elutedwith citrate buffer, and dialyzed against PBS. The flow-through fraction from the above purification was then passedover the 3C9-Sepharose affinity column. The bound antibody(Ab3) was washed, eluted with citrate buffer, and dialyzedagainst PBS. Subsequently, the Ab3 was passed over agpl20sF-Sepharose affinity column, and the column waswashed extensively. The bound antibody (gpl20-specificAb3) was eluted with citrate buffer and dialyzed against PBS.RIA. gpl20SF2 (Chiron) and gp120I11B (SmithKline
Beecham) were labeled with 1251 by using a Bolton-Hunterreagent kit (NEN). RIA plates (Dynatech) were coated for 18hr with various amounts of antibody in PBS. The plates werewashed and then blocked for 1 hr with PBS containing 1%bovine serum albumin (BSA). 125I-labeled gp120jI1B orgp120sF2 in PBS containing 10%o (vol/vol) fetal calfserum and0.05% Tween 20 was added in the presence or absence ofdifferent concentrations of inhibitors. After 3 hr, the plateswere washed with PBS, and the bound radioactivity wasdetermined in a y counter.ELISA. To test the binding activity of 3C9' Abs to a V3
region-derived peptide (amino acids 301-338 of gp120s2),microtiter plates were coated overnight with 0.25 pkg ofprotein in PBS per well. After the adherent layer was blockedand washed, different concentrations of antibody in PBSwere added to the plates. Alkaline phosphatase-coupled goatanti-human immunoglobulin and phosphatase substrate (Sig-ma) were used to detect antibodies binding to the plates.
Neutralization Assays. Two quantitative neutralization as-says with HIV-1 laboratory strains were performed as de-scribed (7, 8). Briefly, monolayers of CEM-SS target cellswere cultured with different HIV strains in the absence orpresence of different neutralizing antibodies. After 3-5 daysin culture, syncytia-forming units were counted, and in thesame assay, supernatants were assayed in ELISA for p24concentration 7-12 days later. Neutralization assays withHIV primary isolates were performed as described (9).
RESULTSProperty of Anti-Idiotype mAb 3C9 as Candidate for Clo-
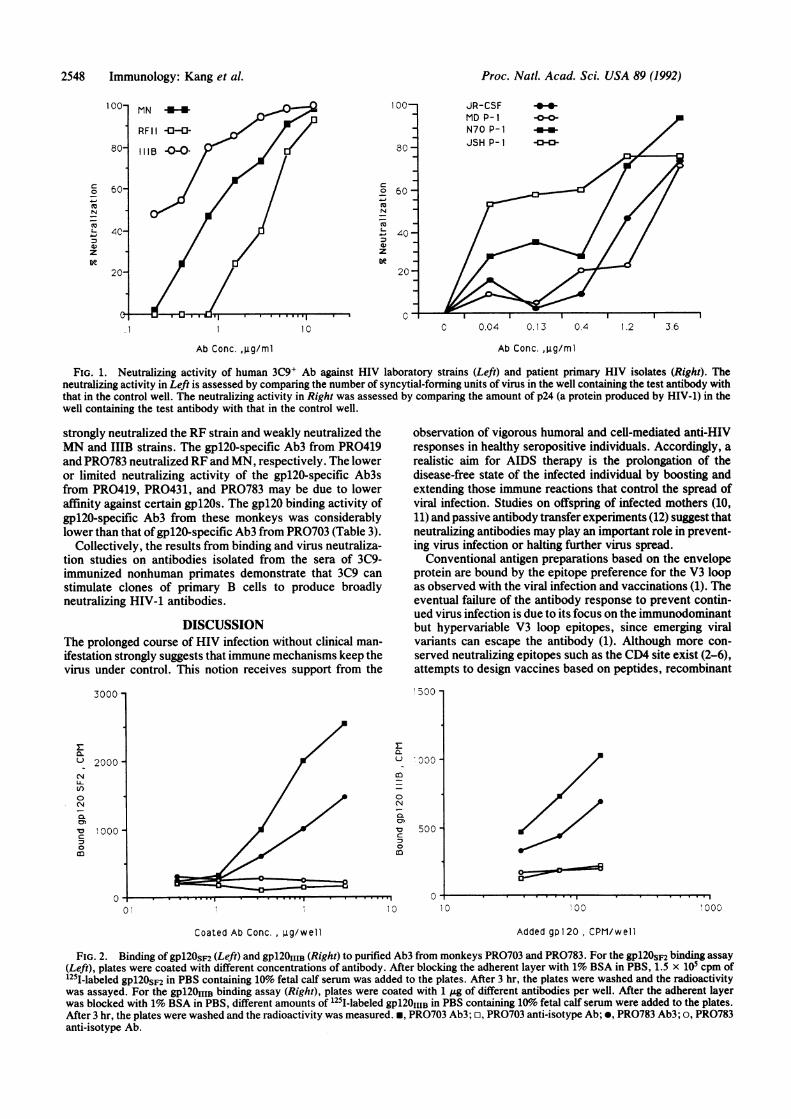
notypic Stimulation. Recently, we generated a series of anti-idiotypic antibodies against purified human anti-gpl20 anti-bodies from a pool of four healthy HIV-1-infected donors (3).Briefly, anti-gpl20 antibodies were purified on gp1205s2-Sepharose and were used to immunize mice for hybridomafusion. Anti-idiotype mAbs were selected for binding tohuman anti-gpl20 antibodies. One anti-idiotype mAb, 3C9,with specificity for CD4 site-directed anti-gpl20 antibodies,was further characterized as a candidate for induction ofanti-gpl20 antibodies. Pooled human HIV-1+ sera were pu-rified on a 3C9 immunoabsorbent, and eluted antibodies(3C9+ Ab) were tested for binding to gpl20 and V3 looppeptides. The results (Table 1) show that 3C9+ Ab bound totwo genetically distinct gpl20s derived from HIVs5 andHIVIIIB and did not bind to the V3 loop peptide. Furthermore,3C9+ Ab binding to gpl20 was inhibited by sCD4, indicatingthat 3C9+ Ab is specific for an epitope around the CD4attachment site of gpl20. Next, the biological function of3C9+ Ab was examined in virus neutralization assays. Hu-man 3C9+ Ab exhibited different spectra of broadly neutral-izing activities against genetically distinct HIV laboratorystrains (Fig. 1 Left) and four primary isolates established fromHIV-infected individuals (Fig. 1 Right). Collectively, theseresults indicate that anti-idiotype mAb 3C9 recognizes aclonotypic determinant shared by human polyclonal anti-gpl20 antibodies that are specific for a conserved epitopearound the CD4 attachment site and exhibit broadly neutral-izing activities. By this definition, 3C9 defines a marker on Bcells producing neutralizing anti-gpl20 antibodies.
Table 1. Immunological properties of human 3C9-reactiveantibodies (3C9' Ab)
Binding activity of 3C9' Ab in assayscpm in RIA with 1251_gp120tgpl20SF2 gp120IHlB A405 ingp1205~~ l2OI~lB ELISA with
Without With Without With V3 loopAb* sCD4 sCD4 sCD4 sCD4 peptidef
3C9+ Ab 5058 1539 8394 1963 0.07Normal hIg 145 151 282 278V3-specific hAb - 1.77
(polyclonal)*3C9+ Ab was purified from HIV+ human sera on a 3C9-conjugatedSepharose column, normal human immunoglobulin (hIg) was fromThe Binding Site (San Diego), and V3-specific human polyclonalantibodies (hAb) were purified from HIV+ human sera on a V3SF2-conjugated Affi-Gel column as described (6).
tPlates were coated with 0.3 1g of antibody per well. After additionof PBS containing 10%6 fetal calf serum to block the adsorbed layer,5 x 104 cpm of 125I-labeled gp12OsF2 or 125I-labeled gp120jII1 wasadded to each well in the presence or absence of sCD4 at 50 ,ug/ml.After 3 hr, the plates were washed and the radioactivity wasmeasured.
*Plates were coated with 0.25 ,ug of V3SF2 peptide in PBS per well.After addition of PBS containing 1% BSA to block the adsorbedlayer, 1 ,ug of antibodies per ml of PBS was added to the plates.Alkaline phosphatase-coupled goat anti-human Ig and phosphatasesubstrate were used to detect antibodies binding to the plates.Absorbance was measured at 405 nm.
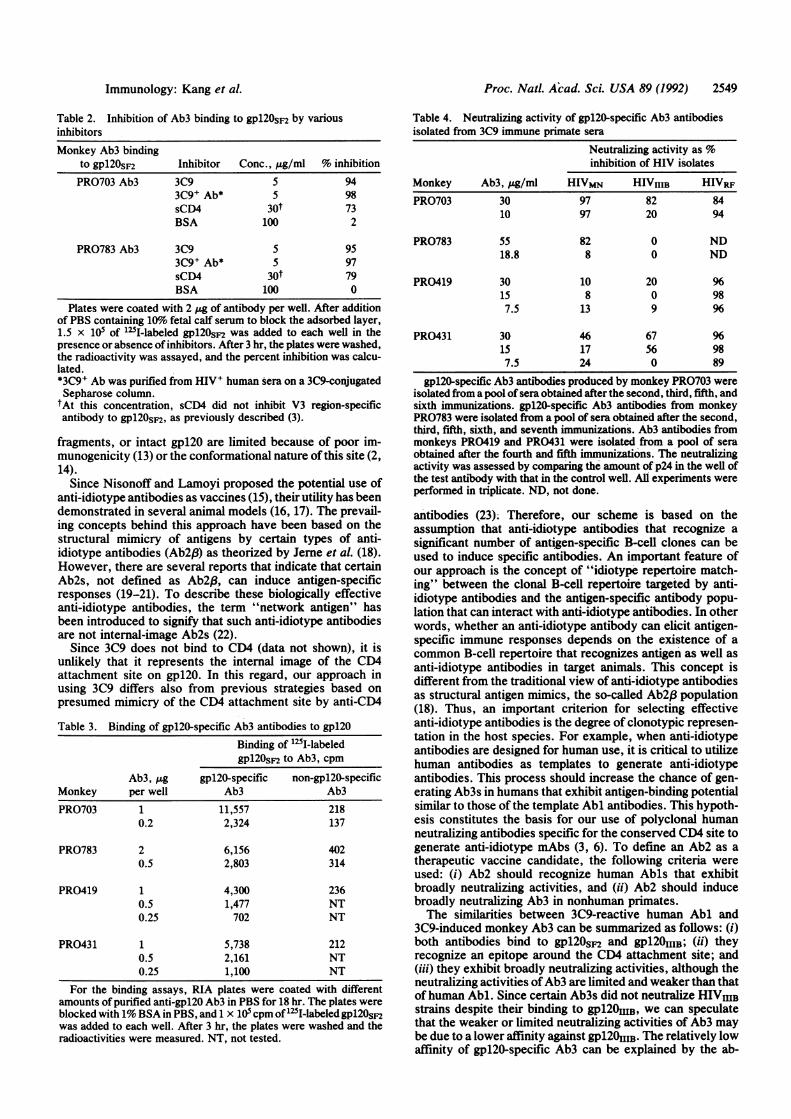
3C9 mAb Induces Anti-gpl20 Ab3 Antibodies. To testwhether 3C9 elicits anti-gpl20 antibodies, four cynomolgusmonkeys were immunized with 3C9 mAb in Syntex adjuvantformulation. Since the immune serum was expected to con-tain 3C9-specific antibodies (Ab3) and mouse immunoglob-ulin-specific antibodies (anti-isotype Ab), it was fractionatedon successive affinity columns and tested for binding togpl20. Ab3 from two monkeys bound to gpl20sF2 in anantibody concentration-dependent manner (Fig. 2 Left) andto gp120O1HB in an antigen concentration-dependent manner(Fig. 2 Right), whereas anti-isotype Abs did not bind to eithertype of gpl20. Similar results were obtained with Ab3s fromtwo other monkeys (data not shown). Inhibition assays wereperformed to analyze the specificity of Ab3. The results inTable 2 show that gp120 binding of Ab3 from monkeysPRO703 and PRO783 were inhibited by 3C9 anti-idiotypemAb (Ab2), human 3C9+ Ab (Abl), and sCD4. Similar resultswere obtained with Ab3s from other monkeys (data notshown). These results confirm the anti-clonotypic nature of3C9, as it is able to induce in primates gpl20-specific Ab3with similar specificity to the human anti-gpl20 antibodies(3C9+ Ab) purified on 3C9 immunoabsorbent (see Table 1).Monkey Ab3 antibodies were further fractionated on a
gp120sF2-Sepharose affinity column. The flow-through andbound fractions were collected as non-gpl20-specific Ab3and gpl20-specific Ab3, respectively. These antibodies werethen examined for gpl20 binding activity. The data in Table3 show that all gpl20-specific Ab3 from four monkeys boundto gpl20, whereas non-gpl20-specific Ab3 did not. Thisfinding indicates that only a fraction ofAb3 recognized gpl20and also suggests that the gpl20-specific Ab3 from all fourmonkeys had a range of affinities for gpl20. For example,gpl20-specific Ab3 of PRO703 exhibited a 3- to 4-fold higherbinding to gpl20 than that of gpl20-specific Ab3 from theother three monkeys.
Neutralizing Activity of gpl20-Specific Ab3. Finally, weexamined the HIV-1 virus-neutralizing activities of gpl20-specific Ab3 antibodies. Table 4 summarizes the results. Thegpl20-specific Ab3 from PRO703 neutralized MN, IIIB, andRF strains of HIV-1. The gpl20-specific Ab3 from PRO431
Immunology: Kang et al.
Proc. Natl. Acad. Sci. USA 89 (1992)
c
0
N
L.
-
=4)
z
.1 1 10
Ab Conc. ,jLg/ml
0 0.04 0.13 0.4 1.2 3.6
Ab Conc. ,gg/ml
FIG. 1. Neutralizing activity of human 3C9' Ab against HIV laboratory strains (Left) and patient primary HIV isolates (Right). Theneutralizing activity in Left is assessed by comparing the number of syncytial-forming units of virus in the well containing the test antibody withthat in the control well. The neutralizing activity in Right was assessed by comparing the amount of p24 (a protein produced by HIV-1) in thewell containing the test antibody with that in the control well.
strongly neutralized the RF strain and weakly neutralized theMN and IIIB strains. The gpl20-specific Ab3 from PRO419and PRO783 neutralized RF and MN, respectively. The loweror limited neutralizing activity of the gpl20-specific Ab3sfrom PRO419, PRO431, and PRO783 may be due to loweraffinity against certain gpl20s. The gpl20 binding activity ofgpl20-specific Ab3 from these monkeys was considerablylower than that ofgpl2O-specific Ab3 from PRO703 (Table 3).
Collectively, the results from binding and virus neutraliza-tion studies on antibodies isolated from the sera of 3C9-immunized nonhuman primates demonstrate that 3C9 canstimulate clones of primary B cells to produce broadlyneutralizing HIV-1 antibodies.
DISCUSSIONThe prolonged course of HIV infection without clinical man-ifestation strongly suggests that immune mechanisms keep thevirus under control. This notion receives support from the
3000
r
Q.
N
U')
0
(N
cl
0D
0
0
observation of vigorous humoral and cell-mediated anti-HIVresponses in healthy seropositive individuals. Accordingly, a
realistic aim for AIDS therapy is the prolongation of thedisease-free state of the infected individual by boosting andextending those immune reactions that control the spread ofviral infection. Studies on offspring of infected mothers (10,11) and passive antibody transfer experiments (12) suggest thatneutralizing antibodies may play an important role in prevent-ing virus infection or halting further virus spread.
Conventional antigen preparations based on the envelopeprotein are bound by the epitope preference for the V3 loopas observed with the viral infection and vaccinations (1). Theeventual failure of the antibody response to prevent contin-ued virus infection is due to its focus on the immunodominantbut hypervariable V3 loop epitopes, since emerging viralvariants can escape the antibody (1). Although more con-served neutralizing epitopes such as the CD4 site exist (2-6),attempts to design vaccines based on peptides, recombinant
500-
r-L
co0
N
CM
0co
000 '
500-
0
01
Coated Ab Conc., 4g/well
10 10 00 000
Added gpl2O0, CPM/well
FIG. 2. Binding of gpl2OSF2 (Left) and gp120OiIB (Right) to purified Ab3 from monkeys PRO703 and PRO783. For the gp120sF2 binding assay(Left), plates were coated with different concentrations of antibody. After blocking the adherent layer with 1% BSA in PBS, 1.5 x 105 cpm of125I-labeled gp12OSF2 in PBS containing 10% fetal calf serum was added to the plates. After 3 hr, the plates were washed and the radioactivitywas assayed. For the gpl20OjHB binding assay (Right), plates were coated with 1 pg of different antibodies per well. After the adherent layerwas blocked with 1% BSA in PBS, different amounts of 125I-labeled gpl2OIIIB in PBS containing 10%o fetal calf serum were added to the plates.After 3 hr, the plates were washed and the radioactivity was measured. *, PRO703 Ab3; o, PRO703 anti-isotype Ab; *, PRO783 Ab3; o, PRO783anti-isotype Ab.
c04-
N
L.
a)zBe
2548 Immunology: Kang et al.
Proc. Natl. Acad. Sci. USA 89 (1992) 2549
Table 2. Inhibition of Ab3 binding to gpl20sF2 by variousinhibitorsMonkey Ab3 binding
to gpl20sn Inhibitor Conc., jkg/ml % inhibition
PRO703 Ab3 3C9 5 943C9+ Ab* 5 98sCD4 30t 73BSA 100 2
PRO783 Ab3 3C9 5 953C9+ Ab* 5 97sCD4 30t 79BSA 100 0
Plates were coated with 2 pg of antibody per well. After additionof PBS containing 10%1 fetal calf serum to block the adsorbed layer,1.5 x 105 of 125I-labeled gp12OSF2 was added to each well in thepresence or absence of inhibitors. After 3 hr, the plates were washed,the radioactivity was assayed, and the percent inhibition was calcu-lated.*3C9+ Ab was purified from HIV+ human sera on a 3C9-conjugatedSepharose column.tAt this concentration, sCD4 did not inhibit V3 region-specificantibody to gp120sF2, as previously described (3).
fragments, or intact gpl20 are limited because of poor im-munogenicity (13) or the conformational nature of this site (2,14).
Since Nisonoff and Lamoyi proposed the potential use ofanti-idiotype antibodies as vaccines (15), their utility has beendemonstrated in several animal models (16, 17). The prevail-ing concepts behind this approach have been based on thestructural mimicry of antigens by certain types of anti-idiotype antibodies (Ab2,B) as theorized by Jerne et al. (18).However, there are several reports that indicate that certainAb2s, not defined as Ab2p, can induce antigen-specificresponses (19-21). To describe these biologically effectiveanti-idiotype antibodies, the term "network antigen" hasbeen introduced to signify that such anti-idiotype antibodiesare not internal-image Ab2s (22).
Since 3C9 does not bind to CD4 (data not shown), it isunlikely that it represents the internal image of the CD4attachment site on gpl20. In this regard, our approach inusing 3C9 differs also from previous strategies based onpresumed mimicry of the CD4 attachment site by anti-CD4
Table 3. Binding of gpl20-specific Ab3 antibodies to gpl20Binding of 125I-labeledgp120sF2 to Ab3, cpm
Ab3, jg gpl20-specific non-gpl20-specificMonkey per well Ab3 Ab3
PRO703 1 11,557 2180.2 2,324 137
PRO783 2 6,156 4020.5 2,803 314
PRO419 1 4,300 2360.5 1,477 NT0.25 702 NT
PRO431 1 5,738 2120.5 2,161 NT0.25 1,100 NT
For the binding assays, RIA plates were coated with different
Table 4. Neutralizing activity of gpl2O-specific Ab3 antibodiesisolated from 3C9 immune primate sera
Neutralizing activity as %inhibition of HIV isolates
Monkey Ab3, ug/ml HIVmN HIVnm HIVRFPRO703 30 97 82 84
10 97 20 94
PRO783 55 82 0 ND18.8 8 0 ND
PRO419 30 10 20 96
15 8 0 987.5 13 9 96
PRO431 30 46 67 96
15 17 56 987.5 24 0 89
gpl20-specific Ab3 antibodies produced by monkey PRO703 wereisolated from a pool of sera obtained after the second, third, fifth, andsixth immunizations. gpl2O-specific Ab3 antibodies from monkeyPRO783 were isolated from a pool of sera obtained after the second,third, fifth, sixth, and seventh immunizations. Ab3 antibodies frommonkeys PRO419 and PRO431 were isolated from a pool of seraobtained after the fourth and fifth immunizations. The neutralizingactivity was assessed by comparing the amount of p24 in the well ofthe test antibody with that in the control well. All experiments wereperformed in triplicate. ND, not done.
antibodies (23). Therefore, our scheme is based on theassumption that anti-idiotype antibodies that recognize asignificant number of antigen-specific B-cell clones can beused to induce specific antibodies. An important feature ofour approach is the concept of "idiotype repertoire match-ing" between the clonal B-cell repertoire targeted by anti-idiotype antibodies and the antigen-specific antibody popu-lation that can interact with anti-idiotype antibodies. In otherwords, whether an anti-idiotype antibody can elicit antigen-specific immune responses depends on the existence of acommon B-cell repertoire that recognizes antigen as well as
anti-idiotype antibodies in target animals. This concept isdifferent from the traditional view of anti-idiotype antibodiesas structural antigen mimics, the so-called Ab2J3 population(18). Thus, an important criterion for selecting effectiveanti-idiotype antibodies is the degree of clonotypic represen-tation in the host species. For example, when anti-idiotypeantibodies are designed for human use, it is critical to utilizehuman antibodies as templates to generate anti-idiotypeantibodies. This process should increase the chance of gen-erating Ab3s in humans that exhibit antigen-binding potentialsimilar to those of the template AbI antibodies. This hypoth-esis constitutes the basis for our use of polyclonal humanneutralizing antibodies specific for the conserved CD4 site togenerate anti-idiotype mAbs (3, 6). To define an Ab2 as atherapeutic vaccine candidate, the following criteria wereused: (i) Ab2 should recognize human Abls that exhibitbroadly neutralizing activities, and (ii) Ab2 should inducebroadly neutralizing Ab3 in nonhuman primates.The similarities between 3C9-reactive human Abl and
3C9-induced monkey Ab3 can be summarized as follows: (i)both antibodies bind to gp12OsF2 and gpl20mB; (ii) theyrecognize an epitope around the CD4 attachment site; and(iii) they exhibit broadly neutralizing activities, although theneutralizing activities ofAb3 are limited and weaker than thatof human Abl. Since certain Ab3s did not neutralize HIVmIIBstrains despite their binding to gpl2OIIIB, we can speculatethat the weaker or limited neutralizing activities of Ab3 maybe due to a lower affinity against gpl2OIIIB. The relatively lowaffinity of gpl2O-specific Ab3 can be explained by the ab-
amounts of purified anti-gpl20 Ab3 in PBS for 18 hr. The plates wereblocked with 1% BSA in PBS, and 1 x 10 cpm of125I-labeledgp12OsF2was added to each well. After 3 hr, the plates were washed and theradioactivities were measured. NT, not tested.
Immunology: Kang et al.

Proc. Natl. Acad. Sci. USA 89 (1992)
sence of HIV antigen-primed B cells in immunologicallynaive monkeys. In contrast, it is anticipated that anti-idiotypeantibodies will induce Ab3s ofhigher affinity in HIV-infectedindividuals who have preexisting, gp12O-primed, and affinity-matured B cells that can respond to stimulation by 3C9.
Since 3C9 was not raised against monkey anti-HIV anti-bodies, we cannot expect a high degree of clonotypic repre-sentation in the naive B-cell repertoire of uninfected mon-keys. Thus, it is not surprising to observe that the neutralizingresponse in the 3C9-immunized monkey is variable. Althoughthe immunonaive monkey is not an ideal model for a HIV-infected human, we consider the data gained from the studyof these animals to be very encouraging for developingtherapeutic AIDS vaccines.
We thank Drs. Nabil Hanna and Frank Norton for their helpfuldiscussions, Nancy Dunlop for her technical assistance, Dr. NancyHaigwood for providing gpl20, and Wendy Parker and Susette Finofor their assistance in the preparation of the manuscript. This studywas supported in part by National Institutes of Health Grant1R43AI31310 to C.-Y.K. and by the New York Life InsuranceCompany.
1. Nara, P. L., Garrity, R. D. & Goudsmit, J. (1991) FASEB J. 5,2437-2455.
2. Ho, D. D., McKeating, J. A., Li, X. L., Moudgil, T., Daar,E. C., Sun, N.-C. & Robinson, J. E. (1991) J. Virol. 65,489-493.
3. Chamat, S., Nara, P., Berquist, L., Whalley, A., Morrow,W. J. W., Kohler, H. & Kang, C.-Y. (1991) Modern Ap-proaches to New Vaccines Including Prevention of AIDS:Vaccines 91 (Cold Spring Harbor Lab., Cold Spring Harbor,NY), p. 26 (abstr.).
4. Tilley, S. A., Honmen, W. J., Racho, M. E., Hilgartner, M. &Pinter, A. (1991) Res. Virol. 142, 247-259.
5. Posner, M. R., Hideshima, T., Cannon, T., Mukherjee, M.,Mayer, K. H. & Byrn, R. (1991) J. Immunol. 146, 4325-4332.
6. Kang, C.-Y., Nara, P., Chamat, S., Caralli, V., Ryskamp, T.,Haigwood, N., Newman, R. & Kohler, H. (1991) Proc. Natl.Acad. Sci. USA 88, 6171-6175.
7. Nara, P. L. & Fischinger, P. J. (1988) Nature (London) 332,469-470.
8. Whalley, A. S., Nguyen, M.-L. & Morrow, W. J. W. (1991)Viral Immunol. 4, 201-213.
9. Daar, E. S., Li, X. L., Moudgil, T. & Ho, D. (1990) Proc. Natl.Acad. Sci. USA 87, 6574-6578.
10. Prince, M. A., Horowitz, B., Baker, L., Shulman, R. W.,Ralph, H., Valinsk, J., Cundell, A., Brotman, B., Boehle, W.,Rey, F., Piet, M., Reesink, A., Leslie, N., Tersmette, M.,Miedema, F., Barbosa, L., Nemo, G., Nastala, C. L., Allan,J. S., Lee, D. R. & Eichberg, J. W. (1988) Proc. Natl. Acad.Sci. USA 85, 6944-6948.
11. Karpas, A., Hill, F., Youle, M., Cullen, V., Gray, J., Byron,N., Hayhoe, F. G. J., Tenant-Flowers, M., Howard, L., Gil-gen, D., Oates, J. K., Hawkins, D. & Gazzard, B. (1988) Proc.Natl. Acad. Sci. USA 85, 9234-9237.
12. Emini, E. A., Schleif, W. A., Murthy, K., Eda, Y., Tokiyoshi,S., Putney, S. D., Matsushita, S., Nunberg, J. H. & Eichberg,J. W. (1991) Seventh International Conference on AIDS, Flo-rence, TH.A.64 (Abstr.).
13. Haigwood, N. L., Nara, P. L., VanNest, G., Scandella, C. J.,Eichberg, J. W. & Steimer, K. S. (1991) ModernApproaches toNew Vaccines Including Prevention of AIDS: Vaccines 91(Cold Spring Harbor Lab., Cold Spring Harbor, NY), pp.51-58.
14. Morrow, W. J. W., Williams, W. M., Whalley, A. S.,Ryskamp, T., Newman, R., Kang, C.-Y., Chamat, S., K6hler,H. & Kieber-Emmons, T. (1992) Immunology, in press.
15. Nisonoff, A. & Lamoyi, E. (1981) Clin. Immunol. Immunopa-thol. 44, 397-406.
16. Kennedy, R. C., Henkel, R. D., Pauletti, D., Allan, J. S., Lee,T. H., Essex, M. & Dreesman, G. R. (1986) Science 231,1556-1559.
17. Kohler, H., Kaveri, S., Kieber-Emmons, T., Morrow,W. J. W., Muller, S. & Raychaudhuri, S. (1989) MethodsEnzymol. 178, 3-35.
18. Jerne, N. K., Roland, J. & Cazenave, P.-A. (1982) EMBOJ. 1,243-245.
19. Schick, M. R., Dreesman, G. R. & Kennedy, R. C. (1987) J.Immunol. 138, 3419-3425.
20. Huang, J.-H., Ward, R. E. & Kohler, H. (1986) J. Immunol.132, 770-776.
21. Kennedy, R. C., Dreesman, G. R., Butel, J. S. & Lanford,R. E. (1985) J. Exp. Med. 161, 1432-1449.
22. Kohler, H., Kieber-Emmons, T., Srinivasan, S., Kaveri, S.,Morrow, W. J. W., Muller, S., Kang, C.-Y. & Raychaudhuri,S. (1989) Clin. Immunol. Immunopathol. 52, 104-116.
23. Healey, D. G., Dianda, L., Buch, D., Schroeder, K., Truneh,A., Sattentau, Q. J. & Beverly, P. C. (1991) Eur. J. Immunol.21, 1491-1498.
2550 Immunology: Kang et al.