annual variation of temperature sensitivity of soil organic carbon decomposition in north peatlands:...
TRANSCRIPT

ORIGINAL ARTICLE
Annual variation of temperature sensitivity of soil organic carbondecomposition in North peatlands: implications for thermalresponses of carbon cycling to global warming
Wu Xiang Æ Chris Freeman
Received: 14 April 2008 / Accepted: 18 August 2008 / Published online: 3 September 2008
� Springer-Verlag 2008
Abstract Temperature sensitivities of microbial respira-
tion and dissolved organic carbon (DOC) production were
investigated by using a novel method, thermal gradient
(2–20�C) temperature bar, in two typical peatlands (bog
and fen) in North Wales, UK over 12 months. The study
indicated that temperature sensitivity of soil organic carbon
decomposition in North peatlands was regulated not only
by temperature but soil water content, dry–rewet event and
phenologies. Potential decreases of Q10 (CO2) with
increasing soil temperature were confirmed in both peat-
lands, but Q10 (DOC) increase with increasing soil
temperature in both bog and fen sites. These results imply,
if other factors such as the so-called CO2 fertilization effect
are simultaneously taken into account, that the feedback of
global warming induced CO2 release from peatlands to
climate change may be overestimated in current biogeo-
chemical models. However, global warming might have
been nonlinearly accelerating DOC thermal production,
and therefore it helps explaining the causes of remarkable
increase of DOC in surface water in the Northern Hemi-
sphere during last several decades.
Keywords Temperature sensitivity � Soil organic
carbon � Decomposition � Peatlands
Introduction
There is a general expectation that increases in temperature
can accelerate the decomposition of soil organic carbon,
and, consequently, global warming should increase the
release of soil organic carbon to the atmosphere (Davidson
et al. 2000) and prompt the release of dissolved organic
carbon (DOC) from land to aquatic ecosystems (Freeman
et al. 2001). Over the past several decades, extensive
research has been carried out on the thermal response of
soil systems to global warming (Kirschbaum 1995; Mars-
chner and Bredow 2002; Knorr et al. 2005). However, the
understanding of the mechanism and factors regulating
temperature sensitivity of carbon processes in soil is still
very limited and constrains our confidence in projected
changes in carbon cycling. The interpretations of obser-
vation of laboratory and experiments are highly dependent
on mathematic techniques and inadequate model assump-
tions. Despite much research, a consensus has not yet
emerged on the temperature sensitivity of soil carbon
decomposition (Davidson and Janssens 2006).
Peatlands occupy approximately 15% of boreal and
sub-arctic region, and contain approximately one-third of
the world’s soil carbon pool. The high latitudes occupied
by peatlands are expected to see the greatest amount of
climatic warming in the next several decades (Pastor et al.
2003). Whether the huge amounts of C in peatlands will
response positively or negatively to global warming
depends on not only the temperature sensitivity of net
primary productivity but also the temperature sensitivity
of soil organic carbon decomposition rate. There is
increasing evidence to suggest that the temperature sen-
sitivity of decomposition of soil organic matter can be
significantly influenced by temperature and multiple other
confounding factors such as soil types, substrate quality,
W. Xiang (&) � C. Freeman
School of Biological Sciences, University of Wales,
Bangor LL57 2UW, UK
e-mail: [email protected]
Present Address:W. Xiang
Key laboratory of Biogeology and Environmental Geology
of Education Ministry, China University of Geosciences,
Wuhan 430074, People’s Republic of China
123
Environ Geol (2009) 58:499–508
DOI 10.1007/s00254-008-1523-6

hydrological condition and plant activities (Davidson et al.
1998; Yuste et al. 2004). While there is a paucity of
information on seasonal and spatial variation of tempera-
ture sensitivity of soil respiration in some terrestrial
ecosystems (Janssens and Pilegaard 2003; Saiz et al. 2006;
Vincent et al. 2006), few investigations on annual varia-
tion of temperature sensitivity of soil organic carbon
decomposition in northern peatlands were performed.
Particularly, few data are available on the temperature
sensitivity of DOC production in northern peatlands,
which is very important for valuable insight into the cause
of obvious DOC increase in surface water across much of
Europe and north USA (Evans et al. 2006). Microbial
respiration and DOC production are the major factors in
relation to soil organic carbon decomposition. Therefore,
further understanding of thermal response of microbial
respiration and DOC production in northern peatlands is of
potential significance for global carbon cycling and cli-
mate warming.
Field investigations have generally involved relating soil
CO2 fluxes or DOC concentration to diurnal or seasonal
variations in temperature. However, while this approach
has the advantage of investigating thermal response in
undisturbed soils, interpretations of microbial responses to
temperature are usually confounded by some co-varying
factors, such as root respiration. Though laboratory incu-
bations are commonly criticized for being unnatural, it has
been regarded as a successful complementary tool to field
survey and widely used in studies of temperature sensi-
tivity of soil carbon decomposition. However, few
laboratory incubations were performed under varying
temperature conditions. In particular, thermal response
estimation by means of Q10 function based on only two
temperatures, e.g., field temperature and simulated refer-
ence soil temperature in some research, should be
interpreted with caution, because incubation results based
on limited temperature points might be masked by sea-
sonally shifting thermal optimum in soil carbon-cycling
processes. A novel laboratory incubation method based on
a thermal gradient (2–20�C) temperature bar has been
successfully used in recent works related to thermal
response of North peatlands (Freeman et al. 2001). This
novel technique provides us a more reliable and convenient
tool to investigate annual variation of temperature sensi-
tivity of microbial respiration and DOC production in
North peatlands.
The major objectives of this study were: (1) to inves-
tigate the annual and spatial variations of temperature
sensitivity of soil organic carbon decomposition in North
peatlands, and (2) to identify the possible factors regu-
lating the thermal response of North peatlands to climate
change.
Materials and methods
Sample collection, preparation and incubation
Samples were collected monthly from an ombrotrophic bog
and a minerotrophic fen from February 2006 to January
2007. The bog peat was collected from Marchlyn Mawr
located in North Wales dominated by Sphagnum species
and other familiar bog plants such as hare’s-tail cotton-
grass. The fen peat was collected from Cors Erddreiniog
located in Anglesey dominated by Sphagnum, Juncus and
great fen-sedge (Fig. 1). Both of the peatlands are typical
in North Wales. The surface layer of vegetation was
removed and peat was collected to a depth of approxi-
mately 10–15 cm. For each field site, water table and
temperature of air and soil were measured, while three
subsamples were collected and mixed. The mixed peat
samples were homogenized by hand after removing most
of larger visible roots. A 2-cm thick layer of each peat was
placed in temperature gradient bars within two layers of
cling-film to prevent water loss without inhibiting gas
exchange. Samples were incubated in a temperature gra-
dient bar for 2 weeks, along which the soil temperature was
respectively controlled as 2, 4, 6, 8, 10,…,18 and 20�C.
The details of the method by using gradient bar can be
found in Fenner et al. (2005).
Gas and hydrochemical analysis
Fluxes of carbon dioxide were determined following 2
weeks of incubation by removing the cling-film from the
Fig. 1 Location of studied area
500 Environ Geol (2009) 58:499–508
123
sample surface, placing nine or ten 90 mm 9 13 mm Petri
dish bases upside down on the peat at equal distances along
the temperature gradient bar. The dishes were pressed
approximately 2 mm into the soil. The increase in trace gas
concentration above the initial background concentration
after 45 min was used to estimate gaseous fluxes from the
soil. Four replicate gas samples from each position were
analyzed by gas chromatography.
Dissolved organic carbon was tested in peat leachable
water obtained from each sample. Briefly, a peat cube
(1 cm3) was gently homogenized for 60 s with 9 ml
deionized water in a stomacher (Seward Colworth model
400). Following centrifugation (14,2809g, 25 min),
leachable water was obtained after through 0.45 lm filter.
The water samples were stored at 4�C before analysis.
DOC was measured by using Total Organic Carbon ana-
lyzer (Shimadzu 5000, Japan).
The apparent molecular weight distribution of DOC in
leachable water was determined by using high-pressure
size exclusion chromatography method of Zhou et al.
(2000).
Calculation of temperature sensitivity of microbial
respiration and DOC production
Temperature sensitivity of microbial respiration or DOC
production was estimated by means of Q10 function:
Q10 ¼ ðR2=R1Þ½10=ðT2�T1Þ� ð1Þ
where R2 and R1 are the rates of CO2 emission or DOC
concentration at two temperatures, T2 and T1. In this study,
T2 = 20�C and T1 = 2�C. R2 and R1 were estimated by
calculating linear regression slopes of CO2 flux or DOC
concentration against temperature.
Results and discussion
Annual variation of temperature sensitivity of microbial
respiration
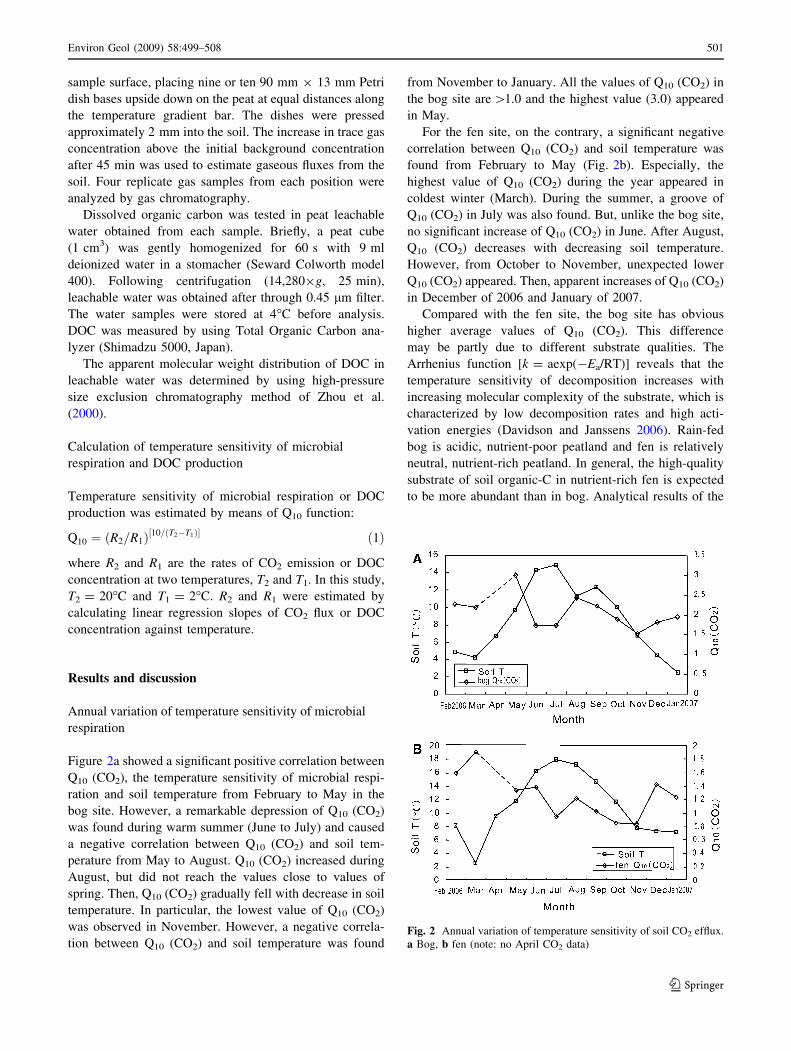
Figure 2a showed a significant positive correlation between
Q10 (CO2), the temperature sensitivity of microbial respi-
ration and soil temperature from February to May in the
bog site. However, a remarkable depression of Q10 (CO2)
was found during warm summer (June to July) and caused
a negative correlation between Q10 (CO2) and soil tem-
perature from May to August. Q10 (CO2) increased during
August, but did not reach the values close to values of
spring. Then, Q10 (CO2) gradually fell with decrease in soil
temperature. In particular, the lowest value of Q10 (CO2)
was observed in November. However, a negative correla-
tion between Q10 (CO2) and soil temperature was found
from November to January. All the values of Q10 (CO2) in
the bog site are [1.0 and the highest value (3.0) appeared
in May.
For the fen site, on the contrary, a significant negative
correlation between Q10 (CO2) and soil temperature was
found from February to May (Fig. 2b). Especially, the
highest value of Q10 (CO2) during the year appeared in
coldest winter (March). During the summer, a groove of
Q10 (CO2) in July was also found. But, unlike the bog site,
no significant increase of Q10 (CO2) in June. After August,
Q10 (CO2) decreases with decreasing soil temperature.
However, from October to November, unexpected lower
Q10 (CO2) appeared. Then, apparent increases of Q10 (CO2)
in December of 2006 and January of 2007.
Compared with the fen site, the bog site has obvious
higher average values of Q10 (CO2). This difference
may be partly due to different substrate qualities. The
Arrhenius function [k = aexp(-Ea/RT)] reveals that the
temperature sensitivity of decomposition increases with
increasing molecular complexity of the substrate, which is
characterized by low decomposition rates and high acti-
vation energies (Davidson and Janssens 2006). Rain-fed
bog is acidic, nutrient-poor peatland and fen is relatively
neutral, nutrient-rich peatland. In general, the high-quality
substrate of soil organic-C in nutrient-rich fen is expected
to be more abundant than in bog. Analytical results of the
Fig. 2 Annual variation of temperature sensitivity of soil CO2 efflux.
a Bog, b fen (note: no April CO2 data)
Environ Geol (2009) 58:499–508 501
123
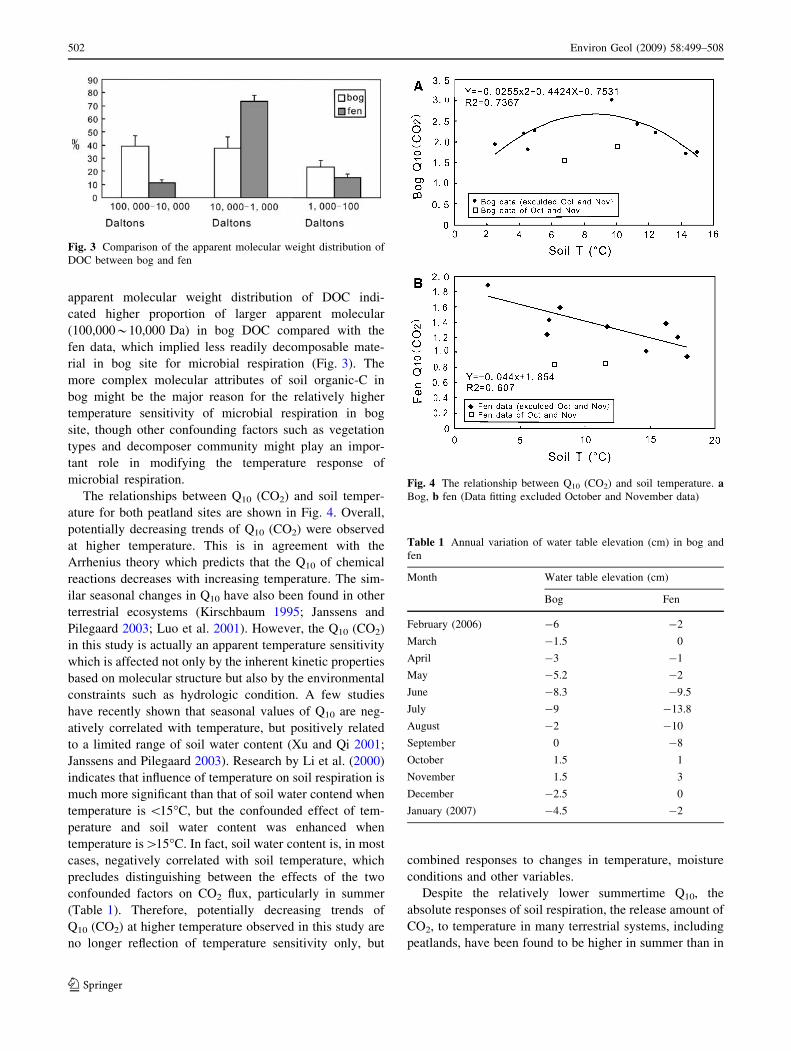
apparent molecular weight distribution of DOC indi-
cated higher proportion of larger apparent molecular
(100,000*10,000 Da) in bog DOC compared with the
fen data, which implied less readily decomposable mate-
rial in bog site for microbial respiration (Fig. 3). The
more complex molecular attributes of soil organic-C in
bog might be the major reason for the relatively higher
temperature sensitivity of microbial respiration in bog
site, though other confounding factors such as vegetation
types and decomposer community might play an impor-
tant role in modifying the temperature response of
microbial respiration.
The relationships between Q10 (CO2) and soil temper-
ature for both peatland sites are shown in Fig. 4. Overall,
potentially decreasing trends of Q10 (CO2) were observed
at higher temperature. This is in agreement with the
Arrhenius theory which predicts that the Q10 of chemical
reactions decreases with increasing temperature. The sim-
ilar seasonal changes in Q10 have also been found in other
terrestrial ecosystems (Kirschbaum 1995; Janssens and
Pilegaard 2003; Luo et al. 2001). However, the Q10 (CO2)
in this study is actually an apparent temperature sensitivity
which is affected not only by the inherent kinetic properties
based on molecular structure but also by the environmental
constraints such as hydrologic condition. A few studies
have recently shown that seasonal values of Q10 are neg-
atively correlated with temperature, but positively related
to a limited range of soil water content (Xu and Qi 2001;
Janssens and Pilegaard 2003). Research by Li et al. (2000)
indicates that influence of temperature on soil respiration is
much more significant than that of soil water contend when
temperature is \15�C, but the confounded effect of tem-
perature and soil water content was enhanced when
temperature is[15�C. In fact, soil water content is, in most
cases, negatively correlated with soil temperature, which
precludes distinguishing between the effects of the two
confounded factors on CO2 flux, particularly in summer
(Table 1). Therefore, potentially decreasing trends of
Q10 (CO2) at higher temperature observed in this study are
no longer reflection of temperature sensitivity only, but
combined responses to changes in temperature, moisture
conditions and other variables.
Despite the relatively lower summertime Q10, the
absolute responses of soil respiration, the release amount of
CO2, to temperature in many terrestrial systems, including
peatlands, have been found to be higher in summer than in
Fig. 3 Comparison of the apparent molecular weight distribution of
DOC between bog and fen
Fig. 4 The relationship between Q10 (CO2) and soil temperature. aBog, b fen (Data fitting excluded October and November data)
Table 1 Annual variation of water table elevation (cm) in bog and
fen
Month Water table elevation (cm)
Bog Fen
February (2006) -6 -2
March -1.5 0
April -3 -1
May -5.2 -2
June -8.3 -9.5
July -9 -13.8
August -2 -10
September 0 -8
October 1.5 1
November 1.5 3
December -2.5 0
January (2007) -4.5 -2
502 Environ Geol (2009) 58:499–508
123

winter (Chapman and Thurlow 1996), which implies that
importance of temperature decreased and soil moisture
assumed increased significance in controlling microbial
respiration. Nevertheless, it has also been found that over
drought-stress may cause rapid declines in soil respiration
rates (Davidson et al. 1998; Bowden et al. 1998). Drought
may reduces the thickness of soil water films, thus inhib-
iting diffusion of extracellular enzymes and soluble
organic-C substrates and lowering substrate availability at
reaction microsites (Davidson and Janssens 2006). There-
fore, drought mechanism might be useful to explain the
lower values of Q10 (CO2) in summer.
However, drought mechanism is unlikely to account for
the significantly lower Q10 (CO2) in rainy October and
November occurred in the bog and the fen, both of which
are significantly deviated from the optimal data fitting and
much lower than expected (square points in Fig. 4). The
reason for so lower Q10 values (or insensitivity) is pos-
sibly related to sharp transformation from oxidizing to
reducing conditions caused by rising water table. During
early autumn, rain caused soil water content to increase
but it is still under moderate moisture condition. After
September, in addition to decreased soil temperature,
precipitation remarkably increased and led to water table
rapidly covered the soil surface (Table 1). Higher water
table slows oxygen diffusion and sharply changed soil
environment from relatively aerobic to anaerobic condi-
tion. Bowden et al. (1998) found reduced soil respiration
rates under very dry and very wet condition. Harper et al.
(2005) used Mielnick–Dugas model to reveal the com-
bined responses of soil CO2 flux to soil temperature and
moisture, and indicated that the highest soil CO2 fluxes
occurred at high soil temperatures and intermediate soil
water contents. Furthermore, their results also confirmed
that both over drying and very high soil water content
strongly suppressed the temperature response of soil CO2
flux, while cool soil temperatures suppressed the soil
moisture response. In addition, extreme wet condition
might have changed soil microbial communities, and
consequently affected soil carbon decomposition pro-
cesses. Research by Waldrop and Firestone (2006)
supported the hypothesis that microbial community com-
position and function is altered when microbes are
exposed to new extremes in environmental conditions,
which caused not only generally slower degradative
enzymatic activities but also shit of soil microbial com-
munities. Unfortunately, our knowledge about the effect
of environmental constraints on microbial community
change is still poorly understood. Even such, it’s reason-
able to attribute the insensitive response of Q10 for
microbial respiration to changes in environmental tem-
perature in October and November mainly to the over wet
condition at lower temperature.
In addition, bog site showed a significant positive cor-
relation between Q10 (CO2) and soil temperature within
approximately 2*10�C which unlike monotonic decreas-
ing changes in Q10 (CO2) occurred in the fen (Fig. 4). The
similar results, increasing tendency in Q10 with increasing
temperature, have been reported in other soil systems
(Sjogersten and Wookey 2002). However, comparison
between the reported data should be very careful because
some of the tendency results were analyzed based on lab-
oratory incubation temperature rather than field
temperature. For example, Sjogersten and Wookey (2002)
calculated Q10 based on tundra soil incubation experiments
at three different temperature pairs. They found Q10 (CO2)
obtained from the 7–12�C range were significantly
(P \ 0.001) higher than the 2–7 and 12–17�C temperature
ranges. As Q10 calculation based on only two temperature
points could be easily masked by seasonally shifting ther-
mal optimum in soil carbon-cycling processes (Fenner
et al. 2005), their Q10 changes might be the reflection of
soil process (e.g., CO2 efflux) thermal optimum rather than
the response to seasonal temperature change.
The significant positive correlation between Q10 (CO2)
and soil temperature in bog site before May might result
from phenologies. It has been found that higher seasonal
Q10 may be affected by seasonal variation in carbon allo-
cation. Davidson et al. (2006) reported springtime Q10 of
soil respiration was always higher than springtime Q10 of
total ecosystem respiration, but the reverse was observed in
the autumn at the Howland forest of Maine, USA. They
suggested that these seasonal differences in apparent tem-
perature sensitivities of soil respiration and total ecosystem
respiration could partly result from different phenologies of
aboveground and belowground process, and different
temperature sensitivities of these processes. Boone et al.
(1998) reported Q10’s of 2.5 and 3.5, respectively, in
trenched plots without roots and control plots with roots in
temperate forests. Curiel et al. (2004) compared the sea-
sonal changes of Q10 for soil respiration between a
deciduous hardwood forest compared with an adjacent
evergreen conifer forest. They argued that the seasonal Q10
reflects the greater seasonality of photosynthesis and sub-
sequent supply of substrate belowground in the deciduous
hardwood site compared with the evergreen conifer site,
and the higher seasonal Q10 observed in the hardwood
forest can be explained partly by phenological responses to
seasons. In this study, for bog stand, the highest Q10 (CO2)
appeared in spring which is plant growing period. Though
most of visible larger roots, including living and dead
roots, were removed before incubation in this study, root-
derived factors might still influence the springtime appar-
ent Q10 (CO2) through following several pathways. First,
it’s very difficult or even impossible to remove all the roots
from peat materials during preparation for incubation.
Environ Geol (2009) 58:499–508 503
123
Many fine roots, including fresh root hairs growing during
spring periods, still make more or less contribution to the
apparent Q10 (CO2) though CO2 originating from root
respiration might be very small after removing most of the
roots. Second, even if all roots had been removed before
incubation, significant increase in root exudates exist in
peat samples might exert a strong influences on Q10 (CO2).
Strictly speaking, CO2 efflux measured in this study mainly
including not only the CO2 derived by microbial decom-
position of soil organic matter, but also CO2 evolved by
microbial utilization of exudates or other rhizodeposits.
Kuzyakov (2002) argued that root respiration contributes
only about 40–50% to the root-derived CO2 efflux, the
remaining 50–60% comprise the microbial decomposition
of root exudates and other rhizodeposits. Cheng et al.
(1993) found, in an experiment with 3-week-old wheat
plants, that root respiration and rhizo-microbial respiration
contributed, on average, 40.6 and 59.4% of total rhizo-
sphere respiration, respectively. In the spring and early
summer, the exudation intensity of growing plants has been
found to be very high, and it leads to increased microbial
growth and activity in the rhizosphere (Kuzyakov and
Cheng 2001). From winter to spring, root exudates increase
with growing root biomass and enhanced photosynthesis in
peatlands. Therefore microbial decomposition of root ex-
udates might exert a strong influence on spring apparent
Q10 (CO2) for nutrient-poor bog stand. In addition, stimu-
lation of microbial growth and activity around roots could
increase the mineralization of native soil organic matter or
corresponding rhizosphere priming effect (Kuzyakov
2002), which might consequently affect springtime Q10
(CO2) for the bog site in this study. Interestingly, no sig-
nificant ‘‘spring-stimulation’’ effect on Q10 (CO2) was
found for fen site. These might be partly explained by
different substrate availability. For nutrient-poor bog,
increasing supply of energy-rich root exudates could boost
up not only the labile substrate availability but also
microbial growth, which in turn influenced the temperature
sensitivity of microbial respiration. In contrast, nutrient-
rich fen may supply relatively abundant higher quality
substrates for microbial utilization, which leads to less
importance of root exudates effect, compared with other
environmental constraints, on apparent temperature sensi-
tivity of microbial respiration.
Annual variation of temperature sensitivity
of soil DOC production
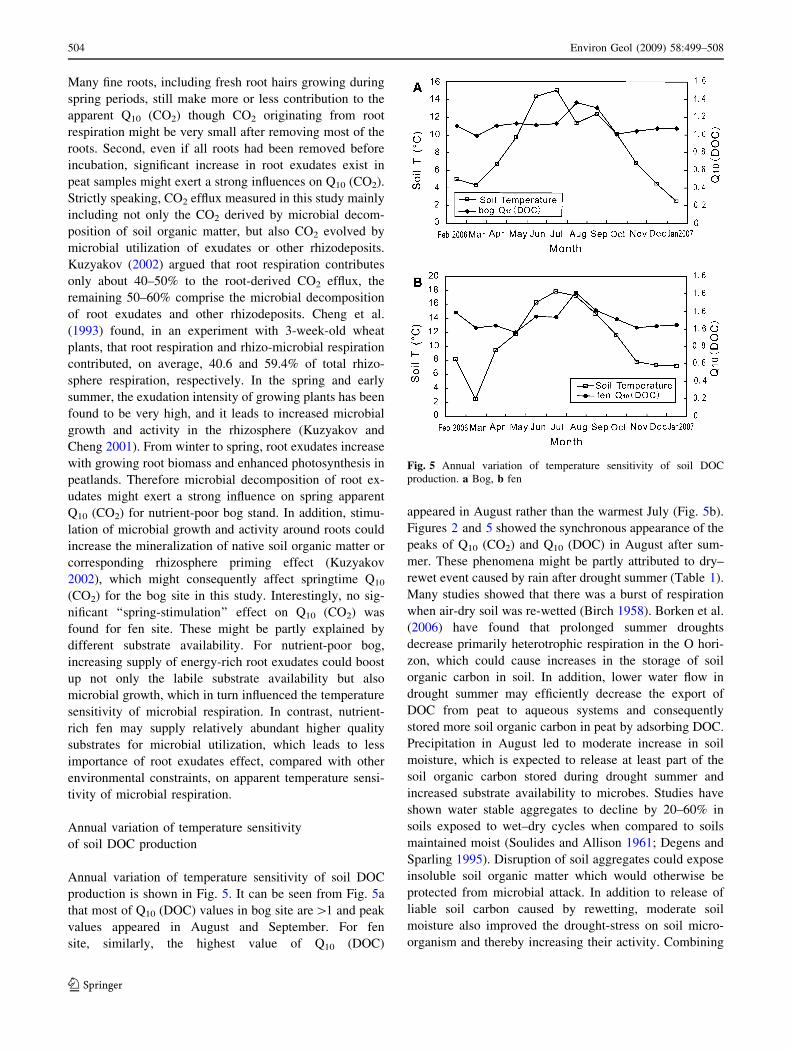
Annual variation of temperature sensitivity of soil DOC
production is shown in Fig. 5. It can be seen from Fig. 5a
that most of Q10 (DOC) values in bog site are[1 and peak
values appeared in August and September. For fen
site, similarly, the highest value of Q10 (DOC)
appeared in August rather than the warmest July (Fig. 5b).
Figures 2 and 5 showed the synchronous appearance of the
peaks of Q10 (CO2) and Q10 (DOC) in August after sum-
mer. These phenomena might be partly attributed to dry–
rewet event caused by rain after drought summer (Table 1).
Many studies showed that there was a burst of respiration
when air-dry soil was re-wetted (Birch 1958). Borken et al.
(2006) have found that prolonged summer droughts
decrease primarily heterotrophic respiration in the O hori-
zon, which could cause increases in the storage of soil
organic carbon in soil. In addition, lower water flow in
drought summer may efficiently decrease the export of
DOC from peat to aqueous systems and consequently
stored more soil organic carbon in peat by adsorbing DOC.
Precipitation in August led to moderate increase in soil
moisture, which is expected to release at least part of the
soil organic carbon stored during drought summer and
increased substrate availability to microbes. Studies have
shown water stable aggregates to decline by 20–60% in
soils exposed to wet–dry cycles when compared to soils
maintained moist (Soulides and Allison 1961; Degens and
Sparling 1995). Disruption of soil aggregates could expose
insoluble soil organic matter which would otherwise be
protected from microbial attack. In addition to release of
liable soil carbon caused by rewetting, moderate soil
moisture also improved the drought-stress on soil micro-
organism and thereby increasing their activity. Combining
Fig. 5 Annual variation of temperature sensitivity of soil DOC
production. a Bog, b fen
504 Environ Geol (2009) 58:499–508
123
both of the processes increased the temperature sensitivi-
ties of both of Q10 (CO2) and Q10 (DOC) in whatever
rewetting bog or fen site.
Though the highest values of Q10 (DOC) appeared in
August for both peat sites, the corresponding values of Q10
(CO2) are not the highest and even lower than expected
values on the basis of soil temperature alone. One of the
explanations is that some of the DOC, such as aromatic
carbon, released from peat during rewetting periods is
relatively stable for microbe utilization. Lundquist et al.
(1999) found that DOC content is not a reliable indicator of
C availability to microorganisms. Their finding was sup-
ported by Zsolnay and Steindl (1991) who found different
carbon mineralization rates in samples with comparable
DOC concentrations, which suggests that DOC is not the
only C substrate for mineralization. Another possible rea-
son is related to the converting reaction from SOC to DOC
by microbes utilization. The release of sequestered carbon
from soil aggregates in dry–rewet events including both
DOC adsorbed by peat and some SOC protected from
microbes attack during dry periods. The previously pro-
tected SOC is generally microbially labile organic carbon
sources, and some of them are converted to DOC while
some to CO2. However, the reaction converting SOC to
DOC is independent of CO2 production and has an inde-
pendent reaction rate constant (Chow et al. 2006).
Temperature effect on DOC production and CO2 release in
rewetting peat would be quite different from that before
drying. Unfortunately, the complex relationship between
carbon availability and DOC production, and the capacity
of the microbial community to utilize the previously pro-
tected SOC, remain poorly understood.
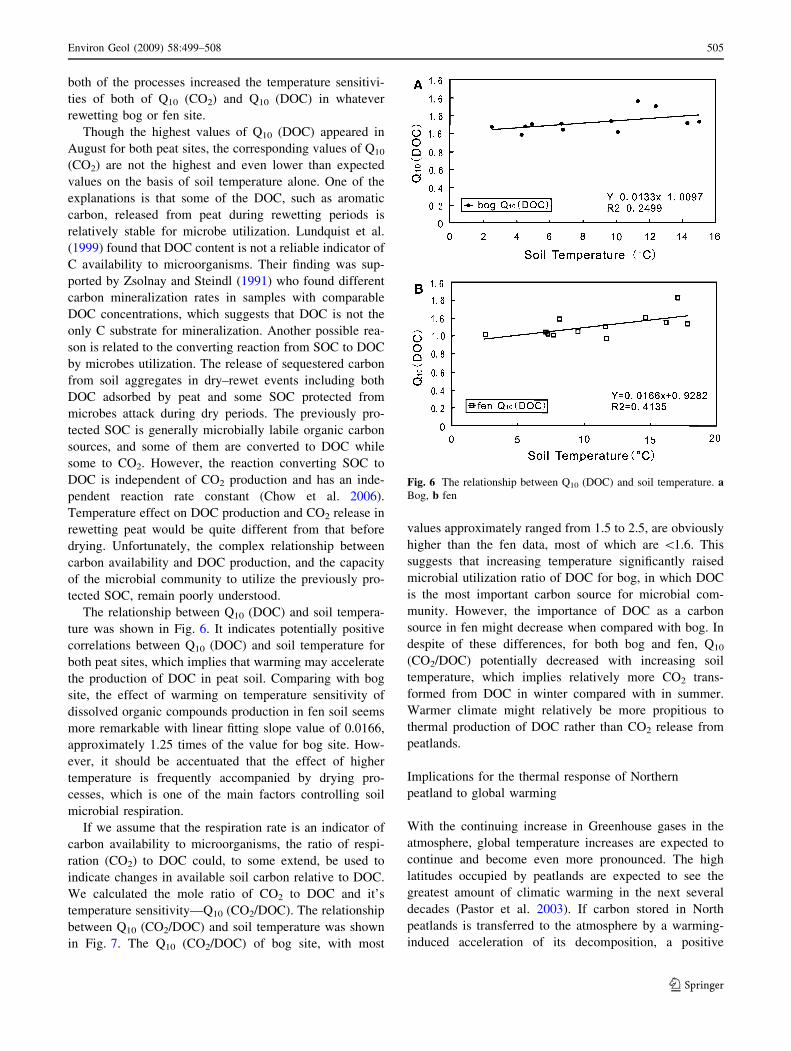
The relationship between Q10 (DOC) and soil tempera-
ture was shown in Fig. 6. It indicates potentially positive
correlations between Q10 (DOC) and soil temperature for
both peat sites, which implies that warming may accelerate
the production of DOC in peat soil. Comparing with bog
site, the effect of warming on temperature sensitivity of
dissolved organic compounds production in fen soil seems
more remarkable with linear fitting slope value of 0.0166,
approximately 1.25 times of the value for bog site. How-
ever, it should be accentuated that the effect of higher
temperature is frequently accompanied by drying pro-
cesses, which is one of the main factors controlling soil
microbial respiration.
If we assume that the respiration rate is an indicator of
carbon availability to microorganisms, the ratio of respi-
ration (CO2) to DOC could, to some extend, be used to
indicate changes in available soil carbon relative to DOC.
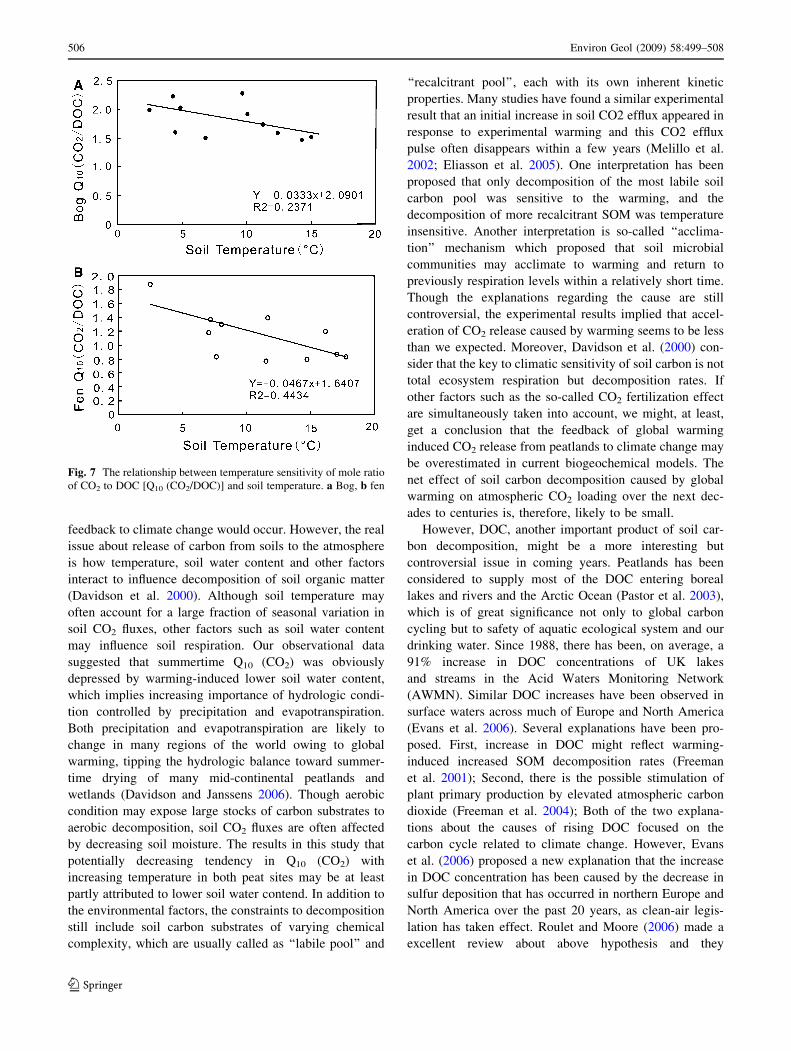
We calculated the mole ratio of CO2 to DOC and it’s
temperature sensitivity—Q10 (CO2/DOC). The relationship
between Q10 (CO2/DOC) and soil temperature was shown
in Fig. 7. The Q10 (CO2/DOC) of bog site, with most
values approximately ranged from 1.5 to 2.5, are obviously
higher than the fen data, most of which are \1.6. This
suggests that increasing temperature significantly raised
microbial utilization ratio of DOC for bog, in which DOC
is the most important carbon source for microbial com-
munity. However, the importance of DOC as a carbon
source in fen might decrease when compared with bog. In
despite of these differences, for both bog and fen, Q10
(CO2/DOC) potentially decreased with increasing soil
temperature, which implies relatively more CO2 trans-
formed from DOC in winter compared with in summer.
Warmer climate might relatively be more propitious to
thermal production of DOC rather than CO2 release from
peatlands.
Implications for the thermal response of Northern
peatland to global warming
With the continuing increase in Greenhouse gases in the
atmosphere, global temperature increases are expected to
continue and become even more pronounced. The high
latitudes occupied by peatlands are expected to see the
greatest amount of climatic warming in the next several
decades (Pastor et al. 2003). If carbon stored in North
peatlands is transferred to the atmosphere by a warming-
induced acceleration of its decomposition, a positive
Fig. 6 The relationship between Q10 (DOC) and soil temperature. aBog, b fen
Environ Geol (2009) 58:499–508 505
123
feedback to climate change would occur. However, the real
issue about release of carbon from soils to the atmosphere
is how temperature, soil water content and other factors
interact to influence decomposition of soil organic matter
(Davidson et al. 2000). Although soil temperature may
often account for a large fraction of seasonal variation in
soil CO2 fluxes, other factors such as soil water content
may influence soil respiration. Our observational data
suggested that summertime Q10 (CO2) was obviously
depressed by warming-induced lower soil water content,
which implies increasing importance of hydrologic condi-
tion controlled by precipitation and evapotranspiration.
Both precipitation and evapotranspiration are likely to
change in many regions of the world owing to global
warming, tipping the hydrologic balance toward summer-
time drying of many mid-continental peatlands and
wetlands (Davidson and Janssens 2006). Though aerobic
condition may expose large stocks of carbon substrates to
aerobic decomposition, soil CO2 fluxes are often affected
by decreasing soil moisture. The results in this study that
potentially decreasing tendency in Q10 (CO2) with
increasing temperature in both peat sites may be at least
partly attributed to lower soil water contend. In addition to
the environmental factors, the constraints to decomposition
still include soil carbon substrates of varying chemical
complexity, which are usually called as ‘‘labile pool’’ and
‘‘recalcitrant pool’’, each with its own inherent kinetic
properties. Many studies have found a similar experimental
result that an initial increase in soil CO2 efflux appeared in
response to experimental warming and this CO2 efflux
pulse often disappears within a few years (Melillo et al.
2002; Eliasson et al. 2005). One interpretation has been
proposed that only decomposition of the most labile soil
carbon pool was sensitive to the warming, and the
decomposition of more recalcitrant SOM was temperature
insensitive. Another interpretation is so-called ‘‘acclima-
tion’’ mechanism which proposed that soil microbial
communities may acclimate to warming and return to
previously respiration levels within a relatively short time.
Though the explanations regarding the cause are still
controversial, the experimental results implied that accel-
eration of CO2 release caused by warming seems to be less
than we expected. Moreover, Davidson et al. (2000) con-
sider that the key to climatic sensitivity of soil carbon is not
total ecosystem respiration but decomposition rates. If
other factors such as the so-called CO2 fertilization effect
are simultaneously taken into account, we might, at least,
get a conclusion that the feedback of global warming
induced CO2 release from peatlands to climate change may
be overestimated in current biogeochemical models. The
net effect of soil carbon decomposition caused by global
warming on atmospheric CO2 loading over the next dec-
ades to centuries is, therefore, likely to be small.
However, DOC, another important product of soil car-
bon decomposition, might be a more interesting but
controversial issue in coming years. Peatlands has been
considered to supply most of the DOC entering boreal
lakes and rivers and the Arctic Ocean (Pastor et al. 2003),
which is of great significance not only to global carbon
cycling but to safety of aquatic ecological system and our
drinking water. Since 1988, there has been, on average, a
91% increase in DOC concentrations of UK lakes
and streams in the Acid Waters Monitoring Network
(AWMN). Similar DOC increases have been observed in
surface waters across much of Europe and North America
(Evans et al. 2006). Several explanations have been pro-
posed. First, increase in DOC might reflect warming-
induced increased SOM decomposition rates (Freeman
et al. 2001); Second, there is the possible stimulation of
plant primary production by elevated atmospheric carbon
dioxide (Freeman et al. 2004); Both of the two explana-
tions about the causes of rising DOC focused on the
carbon cycle related to climate change. However, Evans
et al. (2006) proposed a new explanation that the increase
in DOC concentration has been caused by the decrease in
sulfur deposition that has occurred in northern Europe and
North America over the past 20 years, as clean-air legis-
lation has taken effect. Roulet and Moore (2006) made a
excellent review about above hypothesis and they
Fig. 7 The relationship between temperature sensitivity of mole ratio
of CO2 to DOC [Q10 (CO2/DOC)] and soil temperature. a Bog, b fen
506 Environ Geol (2009) 58:499–508
123

concluded that no proposed explanation is particularly
convincing. Our annual observational data in this study
indicated that temperature sensitivity of DOC production
[Q10 (DOC)] in both types of peatlands increased with
increasing temperature. This finding may help, at least in
part, in explaining the cause of obviously rising DOC,
because warming-induced soil DOC production rate in
peatlands is not evenly increasing but with nonlinear
acceleration. What is certain, unfortunately, is that our
finding still can not perfectly explain the cause of rising
DOC, further researches are still wanting. However, our
observational data underscore the importance of driving
mechanism related to climate change. In fact, more and
more evidences are converging to demonstrate that factors
related to climate change appear likely to be the most
important driving force to rising DOC.
Summary
This study confirmed that the annual variation of tem-
perature sensitivity of soil carbon decomposition in north
peatlands was influenced not only by the type of peat but
also by temperature and hydraulic condition, which are the
main factors controlling the thermal response of soil car-
bon decomposition. For bog, none linear thermal response
of Q10 (CO2) was found, while apparent monotone
decrease of Q10 (CO2) with increasing soil temperature
was observed in fen. Drought or extreme wet may
remarkably decrease the temperature sensitivity of
microbial respiration, but dry–rewet event may increase
Q10 (CO2), which suggests change of frequency and tim-
ing of drought and precipitation in the future will
remarkably influence the extent of thermal response of
peatlands to global warming. However, potentially
decreasing tendency in Q10 (CO2) with increasing tem-
perature in both peat sites in this study, combining with
other authors’ research results, implies that the feedback
of CO2 release from peatlands to global warming may be
overestimated based on current biogeochemical models.
Furthermore, the apparent increases of Q10 (DOC) with
increasing temperature in both bog and fen sites suggests
that climate change may accelerate DOC thermal pro-
duction, offering a potential explanation of the increase of
DOC in surface water in the Northern Hemisphere during
last several decades.
Acknowledgments This work was financial supported by grants
from Leverhulme Trust (F00174h) and Dr. W. Xiang want to thank
CSC and NSFC (40772205) for their support. We particularly want to
thank J. Williamson and C. Yvonne for their help with sample
collecting.
References
Birch HF (1958) The effect of soil drying on humus decomposition
and nitrogen availability. Plant Soil 10:9–31
Boone RD, Nadelhoffer KJ, Canary JD et al (1998) Roots exert a
strong influence on the temperature sensitivity of soil respiration.
Nature 396:570–572
Borken W, Savage K, Davidson EA, Trumbore SE (2006) Effects of
experimental drought on soil respiration and radiocarbon efflux
from a temperate forest soil. Glob Change Biol 12:177–193
Bowden RD, Newkirk KM, Rullo GM (1998) Carbon dioxide and
methane fluxes by a forest soil under laboratory-controlled
moisture and temperature conditions. Soil Biol Biochem
30:1591–1597
Chapman SJ, Thurlow M (1996) The influence of climate on CO2 and
CH4 emissions from organic soils. Agric For Meteorol 79:205–
217
Cheng WX, Coleman DC, Carroll CR, Hoffman CA (1993) In situ
measurement of root respiration and soluble C concentrations in
the rhizosphere. Soil Biol Biochem 25:1189–1196
Chow AT, Tanji KK, Gao S, Dahlgren RA (2006) Temperature, water
content and wet–dry cycle effects on DOC production and
carbon mineralization in agricultural peat soils. Soil Biol
Biochem 38:477–488
Curiel YJ, Janssens IA, Carrara A et al (2004) Annual Q10 of soil
respiration reflects plant phenological patterns as well as
temperature sensitivity. Glob Change Biol 10:161–169
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil
carbon decomposition and feedbacks to climate change. Nature
440:165–173
Davidson EA, Belk E, Boone RD (1998) Soil water content and
temperature as independent or confounded factors controlling
soil respiration in a temperate mixed hardwood forest. Glob
Change Biol 4:217–227
Davidson EA, Trumbore SE, Amundson R (2000) Soil warming and
organic carbon content. Nature 408:789–790
Davidson EA, Janssens IA, Luo YQ (2006) On the variability of
respiration in terrestrial ecosystems: moving beyond Q10. Glob
Change Biol 12:154–164
Degens BP, Sparling GP (1995) Repeated wet–dry cycles do not
accelerate the mineralisation of organic C involved in the macro-
aggregation of a sandy soil. Plant Soil 175:197–203
Eliasson PE, McMurtrie RE, Pepper DA, Stromgren M, Linder S,
Agren GI (2005) The response of heterotrophic CO2 flux to soil
warming. Glob Change Biol 11:167–181
Evans CD, Chapman PJ, Clark JM, Monteith DT, Cresser MS (2006)
Alternative explanations for rising dissolved organic carbon
export from organic soils. Glob Change Biol 12:2044–2053
Fenner N, Freeman C, Reynolds B (2005) Observations of a
seasonally shifting thermal optimum in peatland carbon-cycling
processes: implications for the global carbon cycle and soil
enzyme methodologies. Soil Biol Biochem 37:1814–1821
Freeman C, Evans CD, Monteith DT, Reynolds B, Fenner N (2001)
Export of organic carbon from peat soils. Nature 412:785
Freeman C, Fenner N, Ostle NJ et al (2004) Export of dissolved
organic carbon from peatlands under elevated carbon dioxide
levels. Nature 430:195–198
Harper CW, Blair JM, Fay PA, Knapp AK, Carlisle JD (2005)
Increased rainfall variability and reduced rainfall amount
decreases soil CO2 flux in a grassland ecosystem. Glob Change
Biol 11:322–334
Janssens IA, Pilegaard K (2003) Large seasonal changes in Q10 of soil
respiration in a beech forest. Glob Change Biol 9:911–918
Environ Geol (2009) 58:499–508 507
123

Kirschbaum MUF (1995) The temperature dependence of soil organic
matter decomposition, and the effect of global warming on soil
organic c storage. Soil Biol Biochem 27:753–760
Knorr W, Prentice IC, House JI, Holland EA (2005) Long-term
sensitivity of soil carbon turnover to warming. Nature 443:298–
300
Kuzyakov Y (2002) Separating microbial respiration of exudates from
root respiration in non-sterile soils: a comparison of four
methods. Soil Biol Biochem 34:1621–1631
Kuzyakov Y, Cheng W (2001) Photosynthesis controls of rhizosphere
respiration and organic matter decomposition. Soil Biol Biochem
33:1915–1925
Li LH, Wang QB, Bai YF (2000) Soil respiration of a Leymuschinensis grassland in the Xilin River Basin as affected by over-
grazing and climate. Acta Phytoecol Sin 24:680–686 (in
Chinese)
Lundquist EJ, Jackson LE, Scow KM (1999) Wet–dry cycles affect
dissolved organic carbon in two California agricultural soils. Soil
Biol Biochem 31(7):1031–1038
Luo Y, Wan S, Hui D et al (2001) Acclimatization of soil respiration
to warming in a tall grass prairie. Nature 413:622–625
Marschner B, Bredow A (2002) Temperature effects on release and
ecologically relevant properties of dissolved organic carbon in
sterilized and biologically active soil samples. Soil Biol
Biochem 34:459–466
Melillo JM, Steudler PA, Aber JD, Newkirk K, Lux H, Bowles FP,
Catricala C, Magill A, Ahrens T, Morrisseau S (2002) Soil
warming and carbon-cycle feedbacks to the climate system.
Science 298:2173–2175
Pastor J, Solin J, Bridgham SD, Updegraff K, Harth C, Weishampel P,
Dewey B (2003) Global warming and the export of dissolved
organic carbon from boreal peatlands. Oikos 100:380–386
Roulet N, Moore TR (2006) Browning the waters. Nature 444:283–
284
Saiz G, Green C, Butterbach-Bahl K, Kiese R, Avitabile V, Farrell E
(2006) Seasonal and spatial variability of soil respiration in four
Sitka spruce stands. Plant Soil 287:161–176
Sjogersten S, Wookey PA (2002) Climatic and resource quality
controls on soil respiration across a forest–tundra ecotone in
Swedish Lapland. Soil Biol Biochem 34:1633–1646
Soulides D, Allison F (1961) Effect of drying and freezing soils on
carbon dioxide productions, available mineral nutrients, aggre-
gation and bacterial population. Soil Sci 91:291–298
Vincent G, Shahriari AR, Lucot E, Badot PM, Epron D (2006) Spatial
and seasonal variations in soil respiration in a temperate
deciduous forest with fluctuating water table. Soil Biol Biochem
38:2527–2535
Waldrop MP, Firestone MK (2006) Response of microbial commu-
nity composition and function to soil climate change. Microb
Ecol 52:716–724
Xu M, Qi Y (2001) Spatial and seasonal variations of Q10 determined
by soil respiration measurement at a Sierra Nevadan forest. Glob
Biogeochem Cycles 15:687–696
Yuste JC, Janssens IA, Carrara A, Ceulemans R (2004) Annual Q10 of
soil respiration reflects plant phonological patterns as well as
temperature sensitivity. Glob Change Biol 10:161–169
Zhou Q, Cabaniss SE, Maurice PA (2000) Consideration in the use of
high-pressure size exclusion chromatography (HPSEC) for
determining molecular weights of aquatic humic substances.
Water Res 34:3505–3514
Zsolnay A, Steindl H (1991) Geovariability and biodegradability of
the water-extractable organic material in an agricultural soil. Soil
Biol Biochem 23:1077–1082
508 Environ Geol (2009) 58:499–508
123