anderson delano santos araÚjo degeneraÇÃo …livros01.livrosgratis.com.br/cp098504.pdf · for...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO - UFRJ
INSTITUTO DE CIÊNCIAS BIOMÉDICAS - ICB
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MORFOLÓGICAS
ANDERSON DELANO SANTOS ARAÚJO
DEGENERAÇÃO GORDUROSA EM MÚSCULO ESQUELÉTICO DO MANGUITO ROTATOR DE OVELHAS: UM ESTUDO
EXPERIMENTAL
SALVADOR

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2008
ANDERSON DELANO SANTOS ARAÚJO
DEGENERAÇÃO GORDUROSA EM MÚSCULO ESQUELÉTICO
DO MANGUITO ROTATOR DE OVELHAS: UM ESTUDO EXPERIMENTAL.
Dissertação apresentada ao Programa de Pós-graduação em Ciências Morfológicas do Instituto de Ciências Biomédicas da UFRJ realizado em parceria com o Instituto de Ciências da Saúde da UFBA, como requisito parcial para obtenção do grau de Mestre em Ciências Morfológicas.
Orientador: Prof. Dr. Radovan Borojevic
Co-orientador: Prof. Dr. Gildásio Cerqueira Daltro

SALVADOR 2008
Araújo, Anderson Delano Santos
DEGENERAÇÃO GORDUROSA EM MÚSCULO ESQUELÉTICO DO MANGUITO ROTATOR DE OVELHAS: UM ESTUDO EXPERIMENTAL/ Anderson Delano Santos Araújo
42 f.; 30 cm; il.
Dissertação (mestrado) - Universidade Federal do Rio de Janeiro. Instituto de Ciências Biomédicas e Universidade Federal da Bahia. Instituto de Ciências da Saúde, 2008. Orientador: Prof. Radovan Borojevic. Co-orientador: Prof. Gildásio Daltro
1. Degeneração gordurosa 2. Músculo esquelético 3. Modelo experimental 4. Manguito rotador.

TERMO DE APROVAÇÃO
ANDERSON DELANO SANTOS ARAÚJO
DEGENERAÇÃO GORDUROSA EM MÚSCULO ESQUELÉTICO DO MANGUITO ROTATOR DE OVELHAS: UM ESTUDO EXPERIMENTAL.
Dissertação apresentada ao Programa de Pós-graduação em Ciências Morfológicas do Instituto de Ciências Biomédicas da UFRJ realizado em parceria com o Instituto de Ciências da Saúde da UFBA, como requisito parcial para obtenção do grau de Mestre em Ciências Morfológicas.
Prof. Dr. Maria Isabel Doria Rossi Professor da Universidade Federal do Rio de Janeiro (UFRJ) _______________________________________________ Prof. Dr. Luciana Maria Pedreira Ramalho Professora da Universidade Federal da Bahia (UFBA) ________________________________________________ Prof. Dr. Vitor Antonio Fortuna Professor da Universidade Federal da Bahia(UFBA) __________________________________________________ Prof. Dr. Márcia Cury El-Cheikh Revisor e Suplente, Universidade Federal do Rio de Janeiro __________________________________________________ Radovan Borojevic Orientador, Professor Titular da Universidade Federal do Rio de Janeiro (UFRJ) ___________________________________________________ Gildásio de Cerqueira Daltro Co-Orientador, Professor Adjunto da Universidade Federal da Bahia ___________________________________________________ Anderson Delano Santos Araújo Mestrando ___________________________________________________ Salvador ____ de _____________ de 2008

RESUMO
O presente estudo teve como objetivo avaliar a degeneração muscular do manguito rotador em ovelhas, visando estabelecer um modelo experimental em animais de médio porte para o desenvolvimento de pesquisa da manipulação de células-tronco autólogas, como uma opção de tratamento. Foram utilizadas ovelhas adultas da raça Santa Inês, fêmeas, de 2 a 4 anos, com 25 a 35 kg, clinicamente saudáveis. Os Ovinos foram anestesiados e foi realizada a tenotomia do tendão do músculo infra-espinal. Para que o tendão lesionado não se recuperasse, um dreno “Penrose” que foi suturado à porção final do tendão. Foram coletadas biópsias musculares com 21, 35 e 65 dias pós-lesão. As amostras foram processadas pelo método de rotina histopatológica. Os procedimentos deste trabalho foram avaliados e aprovados pela Comissão de Bioética. Nas amostras de 21 dias pós-lesão apresentaram alguns pontos poucos adipócitos entre os fascículos (perimísio) e poucos adipócitos entres as fibras musculares (endomísio). Com 35 dias pós-lesão foi observada maior infiltração de adipócitos no perimísio e no endomísio, do que com 21dias. Nas amostras de 65 dias pós-lesão foi observada uma acentuada infiltração de tecido adiposo e havia diminuição do tamanho das fibras musculares e desarranjo estrutural; e formação de tecido de granulação próximo a região onde foi colocado o dreno, apresentando infiltrado de células mononucleares, proliferação de tecido conjuntivo e neovascularização. Este modelo experimental mostrou-se adequado para o estudo da infiltração gordurosa do tecido muscular do manguito rotator. Palavras chave: Degeneração gordurosa, músculo esquelético, modelo experimental, manguito rotador.

ABSTRACT
This study aimed to evaluate the muscle degeneration of the rotator cuff in sheep, to establish an experimental model of animal midsize search for the development of the manipulation of autologous stem-cells, as an option to treatment. Take used the adult sheep Santa Inez race, female, from 2 to 4 years, with 25 to 35 kg, clinically healthy. The sheep were anesthetized and was held to tenotomy infraespinal. For the injured tendon is not recover, a drain "Penrose" that was sutured to the final portion of the collected tendão. Take used muscle biopsies with 21, 35 and 65 days post-injury. The samples were processed by means of routine histopathology. The procedures of this work were reviewed and approved by the Commission on Bioethics. In samples of 21 days post-injury showed some few points between the issues adipocytes (perimysion) and few adipocytes between the muscle fibers (internal perimysium). With 35 days post-injury was observed in greater infiltration of adipocytes perimysion and internal perimysion, than with 21days. In samples of 65 days post-injury was observed a marked infiltration of fat and there was reduction in the size of muscle fibers and structural breakdown and formation of granulation tissue near the region where the drain was placed, giving infiltrate of mononuclear cells, proliferation connective tissue and neovascularization. This experimental model proved to be suitable for the study of fat infiltration of the muscle tissue of rotator cuff. Key words: fatty degeneration, skeletal muscle, experimental model, rotator cuff.

DEDICATÓRIA
À todos as pessoas que contribuíram para a minha formação, científica ou pessoal e que sempre acreditaram na palavra de um homem.

AGRADECIMENTO
Á Deus, que sempre guiou a minha vida e sempre mostrou-me isso ao longo da
minha caminhada.
Aos Meus Pais e Minhas Irmãs pelo amor, pela dedicação incansável, pelos
exemplos e por serem o meu combustível.
À Denise, simples assim!
Aos Meus Avós pelos exemplos e pelo amor e à Minha Mamãe Norma pelo
amor incondicional.
Aos Meus familiares (Tios e Primos) pelo apoio de sempre.
Aos Meus Pais, Irmãos e Avós paulistas que me acolheram e sempre me
ofertam um amor verdadeiro. Incondicional.
Ao Meu Professor Radovan Borojevic por ter me acolhido, ajudado, acreditado
em mim e por sempre me ensinar a ser Professor.
Ao Meu Professor Gildásio Daltro pela acolhida e pelos ensinamentos na
Ortopedia do Hospital Universitário Professor Edgard Santos.
À Minha Professora Maria Eugênia pelo carinho, atenção e incentivo.
Ao amigo Paulo Itamar Ferraz Lessa pela amizade e pelo incentivo.
À toda equipe do Laboratório de Histologia da UFRJ e à equipe de Medicina
Veterinária da UFF e em especial à Minha Professora Ana Maria Ferreira.
À equipe da Ortopedia e Traumatologia do HUPES pelos ensinamentos e pela
amizade.

À Minha Amiga Gorete (Laboratório da UFRJ) pelo carinho e atenção de
sempre.
Ao Meu Amigo Bruno Adelmo pela amizade, pelas risadas e pelo apoio na
informática.
À Daniela Heine pelo amor e amizade constantes.
Aos Meus alunos pela confiança e carinho devotados.
Aos Meus Amigos Marcelo Rios e Jaqueline pela amizade e confiança.
Ao Meus Amigos Fábio Luciano e Janice pela amizade e pelo incentivo.
Ao Meu C.R. do Flamengo pelo sorriso que sempre coloca no meu rosto e por
sempre me ensinar, seja nas vitórias, seja nas derrotas.

SUMÁRIO
1 INTRODUÇÃO E JUSTIFICATIVA 11
2 REVISÃO DE LITERATURA 14 2.1 DESENVOLVIMENTO DO MÚSCULO ESQUELÉTICO 14 2.2 ESTRUTURA DO MÚSCULO ESQUELÉTICO 16 2.3 CARACTERÍSTICAS DO MÚSCULO ESQUELÉTICO ADULTO 18 2.4 CARACTERÍSTICAS MORFOLÓGICAS DA REGENERAÇÃO DO TECIDO ESQUELÉTICO.
19
2.5 O MANGUITO ROTADOR E A INFILTRAÇÃO ADIPOSA 22
3 OBJETIVOS
25
3.1 OBJETIVO GERAL 25 3.2 OBJETIVOS ESPECÍFICOS 25
4 MATERIAIS E MÉTODOS 26 4.1 ANIMAIS 26 4.2 PROCEDIMENTO CIRÚRGICO 26 4.2.1 PROCEDIMENTO PRÉ-ANESTÉSICO 26 4.2.2 INDUÇÃO ANESTÉSICA 26 4.3 CIRURGIA 26 4.4 COLETA E PROCESSAMENTO DO MATERIAL 27 4.5 ANÁLISE HISTOLÓGICA DA BIÓPSIA MUSCULAR 27 4.6 PROTOCOLO PARA A DETECÇÃO DOS CARBOIDRATOS REATIVOS COMA A LECTINA DA GRIFFONIA SIMPLICIFORA
28
4.7 HISTOMORFOMETRIA 29 4.8. ANÁLISE ESTATÍSTICA 29
5 RESULTADOS 30 4.1 ASPECTOS CLÍNICOS E MACROCÓPICOS 30 4.2 AVALIAÇÃO HISTOPATOLÓGICA 30
5 DISCUSSÃO 32
6 CONCLUSÕES 36
ANEXOS 37
REFERÊNCIAS 39

LISTA DE ABREVIAÇÕES E SIGLAS ATP - Adenosia tri fostato BS-I - Lectina Griffonia simplicifora CK - Creatina quinase CCR2 - Receptor proteína DM - Dermomiotomo E - Ectoderma LP - Placa lateral MCP-1 - Monocyte chemotactic protein 1 MCK - Creatina quinase muscular MHC - Miosina de cadeia pesada MRF - Fator regulatorio miogenico MyoD - Proteína de regulação da diferenciação muscular Myf5 - Fator miogenico 5 MT - Miotomo NC - Notocorda NT - Tubo neural NCAM - Moléculas de adesão neuronal PAS - Ácido periódico reativo de Schiff SC - Esclerotomo

1. INTRODUÇÃO E JUSTIFICATIVA
A degeneração e ruptura do tendão são causas freqüentes de incapacidade
funcional dolorosa no homem. A forma usual de tratamento consiste na
reparação cirúrgica e re-inserção dos tendões rompidos. Tanto a reparação
espontânea quanto o tratamento conservador com a cicatrização do tendão em
posição retraída freqüentemente levam a uma função não satisfatória devido à
retração músculo-tendinosa, atrofia e infiltração adiposa nos músculos
esqueléticos. O tecido adiposo pode, eventualmente, substituir a maioria das
fibras musculares no fascículo muscular (Goutallier et al, 1994). Após algumas
semanas, as alterações retrácteis se tornam irreversíveis (Matsumoto et al,
2002), representando a principal causa da perda de função permanente,
apesar do sucesso do ponto de vista técnico, no reparo do tendão (Goutallier et
al, 1995). A lesão muscular associada é hoje, considerada como um processo
degenerativo não específico (Tomanek & Cooper, 1972; Baker & Hall-Craggs,
1980; Bjorkenheim, 1989; Josza et al, 1990).
Experimentalmente já foi verificado que a liberação do tendão leva a
profundas alterações estruturais e funcionais no músculo esquelético
(Bjorkenheim, 1989). Os músculos se tornam atróficos, sendo que as fibras de
contração lenta (fibras do tipo I) são mais afetadas que as fibras de contração
rápida (tipo II). O tecido conjuntivo intramuscular aumenta enquanto que o
número de capilares diminui (Josza et al, 1990). O músculo esquelético encurta
e perde parcialmente sua capacidade de criar tensão (Crawford, 1977). Após
um tempo relativamente curto as alterações se tornam irreversíveis (Jozsa et
al, 1990).
Na prática médica são observadas alterações musculares semelhantes
após a lesão por ruptura dos tendões dos músculos do manguito rotador. A
atrofia muscular é freqüentemente pronunciada e parece depender do tamanho
da ruptura e do tempo pós-lesão (Nakagaki et al, 1996; Gerber et al, 2000).
Imagens obtidas por tomografia computadorizada e ressonância magnética
demonstram a progressiva infiltração ou substituição dos músculos do
manguito rotador pelo tecido adiposo, processo denominado de degeneração
adiposa muscular (Goutallier et al, 1989; Goutallier et al, 1994; Nakagaki et al,

1996; Fuchs et al, 1999). Estas alterações musculares têm uma grande
relevância clínica uma vez que a degeneração adiposa pode predizer a
possibilidade de reparação das rupturas do tendão do manguito rotador
(Thomazeau et al, 1997; Goutallier et al, 1999) e influenciar de forma
substancial o prognóstico da reparação do manguito rotador (Goutallier et al,
1999; Pfirrmann et al, 1999; Gerber et al, 2000) ou da artroplastia total do
ombro (Edwards et al, 2002). A recuperação parcial da lesão pode ocorrer
quando há presença de atrofia muscular (Thomazeau et al, 1997; Gerber et al,
2000). Por outro lado, quando há presença de degeneração adiposa, a lesão
no ombro humano parece ser irreversível (Goutallier et al, 1999; Gerber et al.
2000). Além disso, a degeneração gordurosa muscular pode aumentar após a
reparação mesmo quando se obtém a reparação do tendão para o osso
(Gerber et al, 2000).
A degeneração gordurosa no músculo esquelético é observada em uma
variedade de doenças neuromusculares incluindo a distrofia muscular de
Duchene (Goutallier et al, 1994), miopatia mitocondrial (Goutallier et al, 1995),
miopatia nemalínica (Matsumoto et al, 2002), e esclerose lateral amiotrófica
(Baker e Hall-Craggs, 1980), em lesões tóxicas musculares (Bjorkenheim,
1989), lesão isquêmica (Tomanek e Cooper, 1972) e após denervação (Josza
et al, 1990; Crawford, 1977).
A degeneração gordurosa tem como conseqüência importante a perda da
função muscular e de amplitude de movimento, ocasionando uma mudança
funcional e estrutural nos músculos esqueléticos(Goutallier et al, 1995).
Baseado nestas informações e sabendo-se que a atrofia muscular e o infiltrado
de células de gordura são importantes fatores prognósticos é necessário o
entendimento dos mecanismos celulares e moleculares que disparam e
mantém a degeneração adiposa no músculo esquelético durante a reparação
da lesão tendinosa.
Na última década, as terapias celulares abriram novas perspectivas em
tratamento de doenças degenerativas. O aporte de células progenitoras
mostrou-se capaz de restaurar a estrutura e função dos tecidos atrofiados ou
degenerados, inclusive de músculo esquelético (Decary,1997). No caso da
atrofia e degeneração gordurosa a questão de terapia celular permanece

aberta. O aporte de novas células pode promover a regeneração do músculo,
mas pode também trazer novos progenitores de células adiposas. A
diferenciação adiposa é a via de diferenciação espontânea defaut de células-
tronco mesenquimais, quer fazem parte de maioria dos protocolos de terapias
celulares de tecidos de origem mesodérmica. O aporte desse tipo de
progenitores pode, portanto, tornar-se deletério na terapia de degeneração
associada com hipertrofia de tecido adiposo ectópico.
Com objetivo de esclarecer a possibilidade de uma nova proposta de
terapias para a degeneração gordurosa do músculo, estamos propondo um
estudo da lesão muscular provocada pela ruptura do tendão do músculo infra-
espinhal em ovelhas para futuramente propor protocolos de terapia celular
visando a reversão deste processo de degeneração.

2. REVISÃO DA LITERATURA
2.1 Desenvolvimento do músculo esquelético
Todos os músculos esqueléticos, exceto os músculos da cabeça são
derivados de células precursoras mesodérmicas que se originam nos somitos.
Durante o desenvolvimento embrionário a especificação das células
precursoras mesodérmicas para a linhagem miogênica é regulada por sinais
positivos e negativos originários dos tecidos circunjacentes (FIGURA 1A)
A especificação para a linhagem miogênica requer um aumento da
expressão de MyoD e Myf5, ativadores transcripcionais da família do fator
regulatório miogênico (MRF). Células miogênicas proliferativas positivas para
MyoD e/ou Myf5 são denominadas mioblastos. A partir do momento em que
elas saem do ciclo celular se tornam miócitos terminalmente diferenciadas que
expressam as MRFs tardias, a miogenina e a MRF4 e subseqüentemente os
genes músculo-específicos tais como a miosina de cadeia pesada (MHC) e a
creatina quinase muscular (MCK) (FIGURA 1B).
FIGURA 1A. Sinalização e eventos celulares na formação do músculo
esquelético embrionário. Células mesodérmicas dos somitos localizadas na

porção dorsal do somito (dermomiotomo – DM) recebem sinais dos tecidos
circunjacentes que induzem Wnts, sonic hedgehog (Shh) e Noggin ou inibem
(BMP 4) a expressão de MRFs primárias (Myf 5 e MyoD) e o comprometimento
para a linhagem miogênica. Mioblastos comprometidos migram lateralmente e
formam o miótomo (MT), o qual, eventualmente forma a musculatura
esquelética. Pax 3 promove a miogênese no miótomo lateral. . E, ectoderma;
LP, placa lateral; SC, esclerotomo; NC, notocorda; NT, tubo neural. (Retirado
de Chargé & Rudnicki, 2004).
FIGURA 1B. A expressão de Pax 3 nas células precursoras contribui para a
expansão das células miogênicas. Após a indução de Myf5 e/ou MyoD, as
células mesodérmicas do somito são comprometidas para a linhagem
miogênica (mioblastos). Posteriormente a upregulation de MRFs secundárias
(miogenica e MRF4) induz a diferenciação terminal de mioblastos para
miócitos. Finalmente a fusão de miócitos dá origem a miofibras multinucleadas
(retirado de Chargé & Rudnicki, 2004).
Por fim, os miócitos mononucleados se fundem formando um sincício
multinucleado o qual eventualmente se torna uma fibra muscular madura
contráctil (FIGURA 1B). Durante o desenvolvimento muscular uma

subpopulação de mioblastos não se diferencia permanecendo associada à
superfície das miofibras em desenvolvimento como células musculares
satélites, quiescentes (FIGURA 1B). Após a maturidade sexual o músculo
esquelético se torna um tecido estável caracterizado pela presença de fibras
musculares pós-mitóticas multinucleadas (Revisto por Chargé & Rudnicki,
2004)
2.2 Estrutura do Músculo esquelético
O tecido muscular é constituído por células alongadas, que contem
grande quantidade de filamentos citoplasmáticos de proteínas contráteis
geradoras de forças necessárias para a contração desse tecido utilizando a
energia contida nas moléculas de ATP. A célula muscular tem origem no
mesoderma e sua diferenciação ocorre pela síntese de proteínas filamentosas
concomitante com o alongamento das células. De acordo com as
características morfológicas e funcionais, distinguem-se três tipos de tecido
muscular; o músculo estriado esquelético, o músculo estriado cardíaco e o
músculo liso (Junqueira. 2004).
O músculo esquelético tem sido objeto de estudo em pesquisas nos
variados campos do conhecimento. O tecido muscular é capaz de exercer um
trabalho dinâmico, que permite a movimentação articular e um trabalho estático
que permite a estabilização articular (McComas, 1996). Afecções de diversas
origens podem acometer a musculatura e vem despertando um crescente
interesse por sua organização estrutural e funcional.
O músculo estriado esquelético é formado por feixes de células
cilíndricas muito longas e multinucleadas que apresentam estriações
transversais. Essas células ou fibras têm contração rápida e vigorosa e estão
sujeitas ao controle voluntário. Alguns componentes das células musculares
receberam nomes especiais como: sarcolema (membrana celular),
sarcoplasma (citossol) e o retículo sarcoplásmatico (retículo endoplasmático
liso) (Junqueira, 2004).
No músculo-esquelético os numerosos núcleos se localizam na periferia
das fibras, nas proximidades do sarcolema, fato este que ajuda a distinguir este

do músculo cardíaco, já que ambos possuem estriações transversais, porém no
músculo cardíaco os núcleos são centrais. As fibras musculares estão
organizadas em grupos de feixes envolvidos por uma camada de tecido
conjuntivo chamada epimísio que recobre o músculo inteiro. Do epimísio
partem finos septos de tecido conjuntivo que se dirigem para o interior do
músculo separando os feixes, com estes septos constituindo o perimísio
envolvendo os feixes de fibras. Cada fibra muscular, individualmente, é
envolvida pelo endomísio que é formado pela lâmina basal da fibra muscular,
associada às fibras reticulares. O endomísio apresenta escassa população
celular constituída por algumas células do conjuntivo, principalmente
fibroblastos (Junqueira, 2004). A unidade estrutural contrátil do músculo
esquelético é a fibra muscular esquelética ou miônio, célula cilíndrica - com 1 a
30 cm de comprimento e diâmetro de 10 a 100 µm - em que os vários núcleos
se dispõem em posição subsarcolêmica e com numerosas miofibrilas no
citoplasma (McComas, 1996).
Um segundo tipo celular da linhagem miogênica é a célula satélite,
mononucleada, sem miofibrilas, que se apóia na membrana basal do miônio.
Essas células persistem na fase G0 do ciclo celular, mas, em resposta a
determinados estímulos, podem entrar no ciclo mitótico, proliferar e se fundir ao
miônio. Desse modo, contribuem para o crescimento, reparo e hipertrofia pós-
natal do músculo-esquelético. Quando em mitose, as células satélites
expressam, temporariamente, moléculas específicas como os fatores de
transcrição da família MyoD, as isoformas da cadeia pesada da miosina e a
desmina (proteína de filamento intermediário), tanto in vitro quanto in vivo.
O MyoD é um membro da família de proteínas cuja expressão ectópica
pode induzir células não-miogênicas a se submeterem à diferenciação
miogênica. A família MyoD compreende MyoD, miogenina e Myf-5. Todas
possuem um domínio protéico conservado, o motivo bHLH e podem ativar a
expressão de genes musculares específicos. Cada membro da família pode
regular um estágio diferente do desenvolvimento muscular. Myf-5 e MyoD
parecem agir no estágio inicial da diferenciação dos mioblastos e a miogenina
no estágio mais tardio do mesmo (Koishi et al, 1994; Fuchtbauer et al, 1992).
As células satélites também expressam as moléculas de adesão NCAM
e M-caderina, no estado quiescente. A M-caderina está presente em células

satélites de músculos normais, desnervados ou em regeneração (Kuschel et
al., 1999). Os miônios apresentam no citoplasma miofibrilas cilíndricas de 1 a 2
µm de diâmetro que, freqüentemente, são tão longas quanto à própria célula
muscular. Estas miofibrilas consistem de pequenas unidades contráteis
repetidas, em série – sarcômeros - cada um com comprimento de 2,2 µm, que
conferem à miofibrila esquelética sua aparência estriada (Alberts et al., 2004).
Cada sarcômero é formado por filamentos finos e espessos. Os primeiros são
compostos de actina e proteínas associadas, sendo ligados por uma de suas
extremidades à linha Z, em cada extremidade do sarcômero. As extremidades
livres dos filamentos finos se estendem em direção ao centro do sarcômero,
onde se sobrepõem aos filamentos espessos. Estes, por sua vez, são
formados a partir de isoformas de miosina II específicas de músculo (Alberts et
al., 2004). As proteínas do disco Z estão em troca dinâmica com um pool
citoplasmático – incluindo até mesmo as proteínas previamente caracterizadas
como componentes estruturais estáticos do disco Z, como a -actinina e a
miotilina (LANGE et al., 2006).
2.3 Características do Músculo Esquelético Adulto
As fibras musculares são circundadas individualmente por tecido
conjuntivo e agrupadas em feixes que formam o músculo esquelético. São
altamente vascularizados para prover os nutrientes essenciais para a função
muscular. A fibra muscular madura está em contato com um neurônio motor
único e expressa moléculas características para a contração muscular,
principalmente as isoformas de miosina de cadeia pesada e enzimas
metabólicas. A especificação para as propriedades contráteis do músculo
esquelético é dada tanto pelo neurônio motor quanto a origem do mioblasto
muito embora os mecanismos precisos ainda não estejam clarificados
(Wigmore & Evans, 2002). Os músculos esqueléticos adultos são compostos
por uma mistura de miofibras com diferentes propriedades fisiológicas, desde
um tipo de contração lenta, resistente à fadiga a um tipo de contração rápida,
não resistente à fadiga. A proporção de cada tipo de fibra dentro do músculo
determina sua capacidade contrátil global. Apesar destas diferenças

fisiológicas, o mecanismo básico da contração muscular é semelhante em
todas as miofibras e é o resultado do mecanismo de escorregamento dos
filamentos espessos ricos em miosina sobre os filamentos finos ricos em
actina, após ativação neuronal (Huxley, 2000). A rede de tecido conjuntivo no
músculo esquelético transforma as contrações das miofibras em uma unidade
funcional onde a contração das miofibras é transformada em movimento via as
junções miotendíneas nas extremidades onde as miofibras se ligam ao
esqueleto através dos tendões. Portanto, as propriedades funcionais do
músculo esquelético dependem da manutenção de uma rede complexa de
miofibras, neurônios motores, vasos sanguíneos e matriz extracelular.
2.4 Características morfológicas da regeneração do tecido esquelético.
O músculo esquelético de adulto é um tecido estável com baixo turnover
(Decary et al, 1997; Schmalbruch & Lewis, 2000). Lesões pouco extensas
suscitam somente um baixo turnover muscular. Estima-se que no músculo
esquelético adulto de rato não mais que 1 – 2% das células sejam substituídas
cada semana (Schmalbruch & Lewis, 2000). Entretanto, o músculo esquelético
de mamíferos tem uma capacidade rápida e extensa de resposta frente a uma
lesão grave.
Quando ocorre uma lesão muscular causada por trauma direto (como
uma atividade física extensa e mais especificamente um treino de resistência)
ou por defeitos inatos genéticos, a regeneração muscular é caracterizada por
duas fases: uma fase degenerativa e uma fase regenerativa. O evento inicial da
degeneração muscular é a necrose das células musculares. Este evento é
geralmente iniciado pela ruptura do sarcolema da miofibra resultando em
permeabilidade da miofibra. A ruptura da integridade da miofibra é indicada
pelo aumento dos níveis séricos das proteínas musculares tais como a creatina
quinase - CK que usualmente está restrita ao citosol da miofibra. Em humanos
e em modelos animais, o aumento sérico de CK também é observado em
doenças musculares degenerativas como nas distrofias musculares. A ruptura
da integridade da membrana sarcolemal resulta em aumento do influxo de
cálcio resultando em perda da homeostasia de cálcio e aumento da proteólise

dependente do aumento do influxo deste íon (Belcastro et al, 1997; Alderton &
Steinhardt, 2000). Implica-se as calpaínas, que são proteases ativadas pelo
cálcio, capazes de clivar as proteínas miofibrilares e do citoesqueleto, no
processo inicial de degeneração miofibrilar (Belcastro et al, 1997).
A fase inicial da lesão muscular está geralmente acompanhada pelo
recrutamento de células inflamatórias e miogênicas. Fatores liberados pelo
músculo lesado ativam as células inflamatórias residentes no músculo que
liberam sinais quimiotáticos para a circulação. Granulócitos neutrófilos são as
células que inicialmente invadem o músculo lesado, sendo vistas nas primeiras
6 horas após a lesão. Após 48 horas, os macrófagos se tornam as células
predominantes no infiltrado inflamatório (Tidball, 1995). Além de seu papel na
degradação da área lesada e fagocitose dos debris celulares tem sido
verificado seu importante papel na regulação dos eventos essenciais na
regeneração do músculo esquelético (Lescaudron et al, 1999; Merly et al,
1999). Diferentes subpopulações macrofágicas estão presentes tanto no
músculo lesado quanto no músculo em regeneração de ratos (McLENNAN,
1996). Tanto a proteína MCP-1 e seu receptor CCR2 estão expressas no
músculo esquelético lesado (Summan et al, 2006; Warren et al, 2004). Além
disso, a imunoexpressão de CCR2 foi vista em macrófagos e em células
precursoras musculares miogenina-positivas (Warren et al, 2005). O papel do
eixo MCP-1 – CCR2 na regeneração muscular foi evidenciado utilizando
animais nocautes para CCR2. Os animais nocautes apresentaram defeito na
regeneração muscular com aumento da adipogênese e fibrose intramuscular
(Warren et al, 2004).
A degeneração muscular é seguida de ativação do processo
regenerativo. A reparação e a manutenção do sincício são feitas por um pool
de células miogênicas precursoras, indiferenciadas, denominadas células
satélites que se localizam abaixo da lamina basal que circunda a miofibra.
Durante o desenvolvimento pós-natal, as células satélites se dividem para
formar novos núcleos para as fibras musculares em crescimento (Moss and
Leblond, 1971), antes de se tornarem quiescentes enquanto as miofibras se
tornam maduras (Schultz et al, 1978). Entretanto as células satélites retém sua
capacidade de proliferar e se diferenciar em resposta a necessidades para o

turnover do núcleo e hipertrofia da miofibra (Snow, 1977,1978; Bischoff, 1986;
Schmalbruch & Lewis, 2000).
A ativação e proliferação das células satélites é provavelmente a
responsável pela enorme capacidade e eficiência de regeneração do músculo
esquelético lesado, com geração de grande número de novos miotubos nos
primeiros 3 dias após uma lesão aguda grave (Whalen et al, 1990). A taxa e
grau deste reparo é enorme considerando-se a escassez de células satélites
associadas com cada miofibra implicando em uma rápida ativação e expansão
dos precursores para a restauração da fibra muscular (Zammit et al, 2002).
A ativação das células satélites ou, seja, sua passagem do estado
quiescente para as etapas subseqüentes ao longo da linhagem miogênica
miogênese é controlada por vários fatores de transcrição, principalmente os
fatores regulatórios miogênicos (MRFs), o Myf5,MyoD, miogenina, and MRF4
(revisto por Zammit & Beauchamp, 2001). Os fatores Myf5 and MyoD
determinam a linhagem miogênica (Rudnicki et al, 1993; Tajbakhsh et al, 1996)
enquanto que a miogenina é essencial para a diferenciação muscular (Hasty et
al., 1993). No músculo adulto, o locus do Myf5 está ativo nas células satélites
quiescentes com Myo D aparecendo durante a ativação e a miogenina assim
que se inicia a diferenciação muscular (Fuchtbauer & Westphal, 1992;
Grounds et al, 1992; Yablonka-Reuveni & Rivera, 1994; Cooper et al, 1999;
Beauchamp et al, 2000).
Histologicamente, as fibras neoformadas são reconhecidas em cortes
longitudinais e em fibras isoladas como fibras contendo núcleos centrais
presentes por vezes ao longo de toda a extensão da fibra regenerada, o que
sugere que a fusão celular não ocorre de forma difusa durante a regeneração
mas, de forma focal no sítio da lesão (Blaveri et al, 1999). As fibras
neoformadas são geralmente basofílicas o que reflete a alta taxa de síntese
protéica e expressam formas embrionárias de miosina de cadeia pesada (Hall-
Craggs & Seyan, 1975; Whalen et al, 1990). Quando a fusão miocitária se
completa, as novas miofibras formadas aumentam em tamanho e o núcleo se
move para a periferia da fibra muscular.

2.5 O Manguito Rotador e a Infiltração Adiposa
O manguito rotador consiste de quatro músculos: o subescapular, o
supraespinal, o infraespinal e o redondo menor. Todos eles se originam na
escápula e inserem-se nas tuberosidades da cabeça umeral que possui três
funções essenciais: estabilidade, mobilidade e nutrição da articulação gleno-
umeral. Os músculos do manguito rotador terminam em tendões largos e
achatados, contínuos com a cápsula articular para formar o manguito
músculotendíneo. O músculo subescapular origina-se da porção anterior da
escápula e insere-se na pequena tuberosidade. O supraespinal origina-se na
fossa supra-espinhal, passa sob o acrômio e insere-se no topo da grande
tuberosidade. Já o infraespinal origina-se na fossa infra-espinhal e insere-se na
grande tuberosidade, atrás do músculo supraespinal e o redondo menor
origina-se da porção látero-inferior da escápula e insere-se na porção posterior
da grande tuberosidade (Sobotta, 2000)
A anatomia vascular do manguito rotador constitui um dos fatores
etiológicos do desenvolvimento das lesões do manguito, com o supraespinal
recebendo sua irrigação sanguínea da massa muscular. A chamada “área
crítica de Codman”, localizada próxima à inserção do supraespinal, é o local
onde a patologia degenerativa do manguito se inicia. Esta área é
hipovascularizada, pois se encontra continuamente comprimida entre o acrômio
e a grande tuberosidade. Sabe-se ainda que há mais abundância de vasos
sanguíneos na porção superior do que na porção inferior. A patogênese das
lesões do manguito rotador é muito controversa tendo no trauma, atrito
(degeneração), hipovascularização e impacto subacromial são os fatores mais
frequentemente citados (Sizinio, 2003).
Charles Neer (1972) descreveu as três fases evolutivas da síndrome
compressiva do manguito rotador. Na fase I ocorrem edema e hemorragias
reversíveis e tem relação direta com o uso excessivo do membro superior. A
fase II é marcada por fibrose e tendinite do manguito rotador e ocorrem de
maneira crônica e intermitente. Já na fase III ocorre a ruptura completa do
manguito com alterações ósseas, cistos subcondrais e formação de osteófitos.
A ruptura dos tendões dos músculos do ombro pode ser responsável pela

degeneração muscular e esta é caracterizada pela infiltração de células
gordurosas no tecido muscular, fato este já confirmado por estudos histológicos
(Goutallier et al, 1994). A infiltração gordurosa e a atrofia são consideradas
como sendo conseqüentes do desuso e da degeneração ou denervação do
músculo, sendo que essa última depende da espessura do tendão e do tempo
ocorrido após a lesão, ainda que a degeneração não tenha relação com o
envelhecimento, somente ocorrendo com o músculo lesionado (Ryan et al.
2002). Ocorre um aumento da densidade do tecido conjuntivo intramuscular e
uma diminuição do número de capilares e os músculos encurtados e
parcialmente lesados perdem a capacidade de desenvolver tensão. Após um
período relativamente curto, estas mudanças tornam-se irreversíveis,
ocasionando uma mudança funcional e estrutural nos músculos esqueléticos
(Gerber et al, 2004).
A degeneração do músculo supra-espinal é global, porém geralmente
leve já a do infra-espinal é global e se mais de meio tendão estiver
despedaçado, pode ser muito severa (Goutallier et al, 1994). O subescapular
também pode ser acometido de forma severa, mas envolve somente a parte do
músculo que está com o tendão rompido e o infra-espinal pode estar com
degeneração se existir uma ruptura larga e ântero-superior envolvendo o
supra-espinal e o subescapular (Goutallier et al, 1994; Gerber et al, 2004;
Meyer et al, 2005). A degeneração gordurosa é um importante fator de
prognóstico para as cirurgias do manguito rotador. Esta é progressiva com o
tempo e é acompanhada por uma debilidade da função muscular, embora
diminua após o reparo, sendo um fator que prediz a reparabilidade do tendão e
influencia substancialmente no resultado clínico da intervenção cirúrgica
(Goutallier et al, 1994; Gerber et al, 2004; Meyer et al, 2005).


3. OBJETIVOS
3.1 Geral
Desenvolver e caracterizar um modelo de lesão muscular em ovelhas pela
ruptura do tendão do músculo infra-espinal que se assemelhe à lesão do
manguito rotator.
3.2 Específicos
3.2.1. Descrever as alterações degenerativas e reparadoras no músculo infra-
espinhal após 21, 35 e 60 dias da ruptura do tendão provocada
experimentalmente.
3.2.2. Quantificar, através de estudo morfométrico, a densidade volumétrica da
infiltração / degeneração gordurosa no músculo infra-espinhal após 35 e 60
dias da ruptura do tendão provocada experimentalmente.

4. MATERIAL E MÉTODOS
4.1 Animais
Todos os procedimentos foram realizados em conformidade com as
recomendações éticas e legais especificadas para a experimentação animal do
Colégio Brasileiro de Experimentação Animal (1991).
Vários estudos experimentais tem demonstrado que os ombros das ovelhas
são modelos adequados para o estudo de desordens do manguito rotador em
humanos. Com base nessas informações foram selecionadas oito ovelhas
fêmeas, jovens, com idade entre 18 e 24 meses e com pesos entre 20 e 30 kg.
4.2 Procedimento cirúrgico
4.2.1 Procedimento pré-anestésico
Foram administrados, por via intravenosa, 0,1 mg/kg de Maleato de
Acepromazina 0,2% (Acepromazina, Syntec) e 0,2 mg/kg de Midazolam
(Induson, Cellofarm Farmacêutica). Além disso, foi administrado 0,4 mg/kg de
morfina, injetada via intramuscular.
4.2.2 Indução anestésica
Para a indução anestésica foram injetados por via intravenosa, 5 mg/kg
de Cloridrato de Cetamina 10% (Cetamin, Syntec) e 0,1 mg/kg de Midazolam
(Induson, Cellofarm Farmacêutica). Para manutenção da narcose anestésica
foi administrado por via inalatória Isoflurano (Isoforine, Cristália).
4.3 Cirurgia
Todos os procedimentos foram realizados em campo cirúrgico estéril, no
Hospital Veterinário da Universidade Federal Fluminense, segundo a técnica
descrita por Gerber e colaboradores (2004). Foi realizada uma incisão
cirúrgica curva, de 15 cm, na região posterolateral da articulação do ombro
direito. A incisão foi aprofundada e a origem acromial do músculo deltóide foi
identificada. O músculo foi elevado na sua porção cranial para a exposição da

inserção tendinosa do músculo infra-espinhal e sua inserção na porção
proximal do úmero. O tendão infra-espinhoso foi liberado de sua inserção da
unidade músculotendínea, na porção proximal do úmero, de forma a mimetizar
uma lesão crônica do manguito rotador. O ombro contralateral serviu como
controle. Relatos prévios mostraram que os tendões do infraespinal das
ovelhas se reparam espontaneamente após uma tenotomia. Para impedir esse
reparo espontâneo, a extremidade do tendão foi coberta com um tubo de
silicone de 5 cm (tubo de dreno de silicone com diâmetro de 12 milímetros,
instrumento médico da Fortuna, Taipei, Formosa) que foi fechada com sutura
na extremidade distal, técnica descrita por Gerber e colaboradores (2004).
4.4 Coleta e processamento do material
Foi realizada a dissecação e biopsias dos músculos infraespinhais, nos
tempos pós-cirúrgicos de 21, 35 e 60 dias, com cerca de 0,5 cm por 0,5 cm,
sendo fixadas imediatamente após a retirada cirúrgica em formaldeído 10%
tamponado. O material foi posteriormente submetido à técnica de rotina para
inclusão em parafina. Resumidamente, os fragmentos foram submetidos à
desidratação em soluções crescentes de álcool etílico, clarificados em xileno e
embebidos em parafina. Cortes de 3 µm de espessura foram obtidos em
micrótomo rotatório e corados com hematoxilina-eosina (técnica histológica
para estudo topográfico), tricrômico de Masson, PAS (ácido periódico-reativo
de Schiff) e Picro-Sirius para colágeno.
4.5 Análise histológica da biópsia muscular:
A organização geral de tecidos foi avaliada assim como as alterações
distróficas (degeneração / regeneração) existentes no tecido. Foi utilizada uma
escala histológica de 4 pontos, modificada de Hack e colaboradores (1999),
considerando-se o percentual da secção histológica apresentando os seguintes
parâmetros: fibrose, infiltração adiposa e degeneração e inflamação.
0 – nenhuma alteração;
1 – 5 -10% da superfície apresentando lesão;

2 - >10–25% da superfície apresentando lesão;
3 - > 25% - 50% da superfície apresentando lesão;
4 - > 50% da superfície apresentando lesão.
4.6 Protocolo para a detecção dos carboidratos reativos coma a lectina da
Griffonia simplicifora
A lectina Griffonia simplicifora (BS-I) se liga especificamente a resíduos
galactosil, cora o endotélio vascular de várias espécies animais inclusive o de
rato (Alroy et al, 1987). Portanto, utilizamos a lectina Griffonia simplicifora
biotinilada (B-1105, Vector Laboratories, USA) para a detecção do endotélio,
sendo utilizada para a quantificação dos capilares.
Em resumo, os cortes histológicos foram desparafinados e hidratados,
seguindo-se então a inibição da peroxidase endógena em solução de H2O2 a
3% em metanol por 15 minutos, lavagem em tampão fosfato salina (PBS) pH
7,4 após o bloqueio das ligações inespecíficas com solução de PBS-BSA 5%.
Os cortes histológicos foram então incubados com a lectina biotinilada na
diluição de 1: 100, em solução tampão contendo sais de metais (Kiernan,
1990), por 16 horas, na geladeira, em câmara úmida. Posteriormente, as
lâminas foram submetidas a lavagens com solução de PBS e incubadas com a
estreptavidina conjugada à peroxidase (Sigma, USA) na diluição de 1:50, em
PBS. Para a visualização da peroxidase foi utilizado o substrato cromógeno
DAB (Liquid DAB, DAKOCYTOMATON), seguindo-se lavagens com PBS e
água destilada, e contracoloração pela hematoxilina.
4.7 Histomorfometria
O estudo morfométrico foi realizado através de um sistema de captura
de imagens, constituído de câmara fotográfica digital EVOLUTION (Media
Cybernetics, USA) acoplada a microscópio de luz E 500 NIKON (Japan) e a um
computador contendo o programa de análise de imagens MAGEPRO–PLUS
5.0 (Media Cybernetics, USA). Imagens digitalizadas de alta qualidade (2048 x
1536 pixels buffer) obtidas das lâminas coradas pela HE e Griffonia simplicifora

utilizando a lente objetiva de 20x foram utilizadas para a quantificação da
densidade capilar (número de secções transversais de capilares / µm2).
4.8. Análise estatística
Todos os dados foram submetidos a análise estatística com auxílio do
programa Microsoft Excel® e SigmaStat®.
Os resultados da análise histomorfométrica dos grupos foram expressos
sob a forma de média ± desvio padrão. Na comparação de mais que duas
médias em uma única variável categórica independente (fator), aplicou-se o
teste de Análise de Variância (ANOVA) e o pós-teste Student-Newman-Keuls.
Um p<0,05 foi considerado como significativo.

5. RESULTADOS
5.1 Aspectos clínicos e macroscópicos
Os animais apresentaram–se após o procedimento cirúrgico
aparentemente bem, levantando-se imediatamente e andando. Nas
observações realizadas 21, 35 e 60 dias pós-lesão os animais apenas
apresentaram no galope uma claudicação leve, com redução da fase de apoio
na marcha, embora as patas dianteiras dos ovinos sejam de utilização para
propulsão.
No procedimento da biópsia aos 60 dias, foi feita uma incisão também
no local onde foi realizada a incisão do tendão. Foi observado que
macroscopicamente o tendão parecia ter se regenerado, apresentando aspecto
semelhante ao tendão original, apesar da presença de um tubo protetor que foi
envolvido pelo tecido conjuntivo.
5.2 Análise Histopatológica
O músculo infra-espinhal dos animais com 21 dias de lesão exibiu uma
organização relativamente pouco distorcida, com feixes musculares paralelos.
Algumas fibras estavam degeneradas (Escore=2, Anexo A), células em
regeneração caracterizadas pela presença de núcleos centrais enfileirados
(Escore=1, Anexo A) e processo inflamatório do tipo mononuclear com raros
granulócitos polimorfonucleares (Escore 3, Anexo A). Notava-se presença de
muito raras e pequenas células adiposas em meio às fibras musculares
fasciculares (Escore=1, Anexo A) e em maior numero no espaço interfascicular
(Escore=1, Anexo A). Todos os adipócitos, incluindo os menores no início da
diferenciação, eram tipicamente uniloculares. Nos espaços interfasciculares os
focos nascentes eram associados aos vasos sanguíneos de pequeno e médio
calibre, com rico tecido adventicial. Os pequenos adipócitos eram
freqüentemente próximos ao lado abluminal dos vasos. Com diferenciação
plena e aumento do volume do lóculo gorduroso, formavam-se cachos
preenchendo os espaços interfasciculares adjacentes. O infiltrado inflamatório

não era mais pronunciado nos focos interfasciculares de adipogênese, sendo
freqüentemente mais intenso nas interfaces interfasciculares ao longo dos
perimísios.
Após 35 dias de lesão notou-se a presença ainda de algumas células
degeneradas (Escore=2, Anexo A), raras células em regeneração (Escore=0,
menos de 5% da superfície examinada, Anexo A) e inflamação constituída por
células mononucleares presentes tanto em regiões inter- quanto
intrafasciculares (Escore=2, Anexo A). Infiltração gordurosa também foi vista
nas duas regiões, sendo mais abundantes em região interfascicular (Escore=2,
Anexo A). Ela se tornou dissecante, separando totalmente os feixes
musculares pela infiltração contínua ao longo dos perimísios.
Progressivamente, grupos de adipócitos invadiam também os feixes
musculares, separando os grupos cada vez menores. Deposição de fibras
colágenas foi vista em torno de células adiposas em áreas interfasciculares
(Escore=1, Anexo A) e entre fibras musculares (Escore=1, Anexo A; Figuras 1,
Anexo B).
Após 60 dias verificamos raras células degeneradas (Escore=1),
diminuição da inflamação em relação ao ponto 21 e 35 (Escore=1) e aumento
da infiltração gordurosa, tanto entre as fibras musculares, dentro do fascículo
muscular (Escore=2), quanto nas regiões interfasciculares (Escore=2). Fibras
colágenas também se mostraram mais abundantes dentro dos fascículos
musculares, em menor quantidade em regiões interfasciculares (Anexo B,
figuras 1-8).

6. DISCUSSÃO
Este estudo foi feito com objetivo de caracterizar descritivamente, do
ponto de vista histológico, as mudanças estruturais que ocorrem no músculo
esquelético após a desinserção da unidade músculo-tendínea do manguito
rotador de ovelhas. Nesta situação cria-se a inatividade do manguito rotador. O
protocolo cirúrgico utilizado para a desinserção do manguito rotador das
ovelhas foi o de Gerber e colaboradores (2004). A vantagem deste protocolo,
onde o tendão liberado é colocado e fixado dentro de um tubo de silicone, é a
criação de um modelo de lesão onde há o aparecimento de alterações no
músculo esquelético, devido a inatividade muscular, como a degeneração
adiposa e a atrofia muscular (Eccles, 1948; Björkenheim, 1989; Josza et al
1990), sem evidências de reparação muscular espontânea (Gerber et al, 2004).
Foi verificado que, a ruptura tendínea induzida, do músculo supraespinal de
coelhos seguida da inserção de uma membrana de polivinilidina (Durapore 7
SVLP, Millipore), para prevenir a reinserção espontânea, de forma semelhante
ao do tubo de silicone, ocasionou uma atrofia muscular, infiltração adiposa e
retardo da regeneração muscular (Matsumoto et al, 2002). Em humanos, o
retardo do tratamento cirúrgico também agrava as lesões no músculo, tanto a
infiltração adiposa quanto a atrofia muscular, sendo que após o reparo cirúrgico
com a re-inserção do tendão não levou a melhora do quadro nem reversão das
lesões (Nakagaki et al, 1996; Matsumoto et al, 2002; Gladstone et al, 2007),
aspecto também visto em coelhos (Rubino et al, 2007).
É bem conhecido que a desinserção do tendão leva a retração músculo-
tendínea provocando mudanças estruturais e arquiteturais profundas no
músculo esquelético (Hans et al, 2003; Meyer et al, 2004). O músculo sofre
atrofia sendo que as fibras lentas do tipo I são mais afetadas que as rápidas do
tipo II (Josza et al, 1990) e perda do volume muscular (Matsumoto et al, 2002;
Gerber et al, 2004; Meyer et al, 2004). Ocorre um encurtamento da fibra
muscular provavelmente devido a diminuição dos sarcômeros (Baker & Hall-
Craggs, 1978) e infiltração adiposa substancial nos espaços criados pela
modificação arquitetural muscular, porém sem evidências de um processo
degenerativo da unidade músculo-tendínea (Meyer et al, 2004). O tecido
adiposo e também o tecido fibroso invadem o músculo. Alguns autores

consideram que este processo visa preencher o espaço criado pela atrofia
muscular (Uhthoff et al, 2003). Há tanto uma infiltração adiposa intrafascicular
quanto extrafascicular (Matsumoto et al, 2002; Gerber et al, 2004), muito
embora o mecanismo que inicia o processo da infiltração adiposa ainda não
seja esclarecido. A implantação de estruturas tubulares, independente do tipo
de material, dentro do tecido muscular, provoca o aparecimento de lipoblastos
no tecido de granulação formado (Xaymardan et al, 2002).
No presente estudo, não pudemos realizar um estudo morfométrico
comparativo no sentido de inferirmos uma diminuição da área das fibras
musculares (atrofia) e/ou aumento da área de tecido adiposo. Por esta razão foi
realizado um estudo semi-quantitativo com a utilização de escores para
determinados parâmetros morfológicos. Diversos estudos em patologia
muscular tem utilizado este tipo de análise semi-quantitativa (Hack et al, 1999;
Fanin et al, 2007). Verificamos que tanto após 35 dias quanto 60 dias da
realização da desinserção tendínea há mínima evidencia de um processo
regenerativo dentro do músculo, o que corrobora o modelo proposto por Gerber
e colaboradores (2004). Além disso, pudemos evidenciar tanto o acúmulo de
tecido adiposo intrafascicular e interfascicular assim como aumento do tecido
conjuntivo extrafascicular e intrafascicular semelhante ao já descrito em
coelhos e em ovelhas (Björkenheim et al, 1989; Gerber et al, 2004; Meyer et al,
2004; Rubino et al, 2007; Coleman et al, 2008). Além disso, pudemos verificar
que o processo angiogênico já se encontra em fase de remodelamento, tendo
sido possível demonstrar a diminuição do número de capilares no tempo 60
dias. O uso da lectina GS permitiu evidenciar a célula endotelial, permitindo
desta forma, aferir o número de capilares existentes no músculo.
Gerber e colaboradores (2004) detectaram em exames de ressonância
magnética (RNM) que nem todas as partes do supra-espinal sofrem o mesmo
grau de atrofia. Nas atrofias conseqüentes a lesões nervosas, o infiltrado de
tecido gorduroso ocorre nas áreas desnervadas e nas atrofias por lesões
completas do tendão, o acúmulo de tecido adiposo é mais homogênea. A parte
mais superficial do músculo que se origina da fáscia reage diferentemente à
lesão e à retração do tendão do que a parte profunda que se origina da
escapula. A parte mais próxima à fáscia contribui mais para a atrofia muscular

do que a parte próxima à inserção muscular que por sua vez sofre mais com o
infiltrado de tecido gorduroso.
A origem dos adipócitos presentes no músculo em regeneração ainda
não é conhecida. Há evidências de que elas possam ser derivadas de células
precursoras mesenquimais presentes no tecido muscular. No tecido adiposo
subcutâneo, células estromais vasculares do tecido muscular podem ser a
fonte dos pré-adipócitos, que geram novos adipócitos. O processo semelhante
pode ocorrer na degeneração gordurosa intramuscular (Hausman &
Eichardson, 1998; Hausman & Poulos, 2005).
Sabe-se atualmente que os pericitos perivasculares têm características
pluripotenciais e estão presentes em todos os tecido adultos inclusive no tecido
muscular (Diaz-Flores et al, 1991; 1992; Brighton et al, 1992; Doherty et al,
1998; Shi & Gronthos, 2003; Brachvogel et al, 2005; Meirelles et al, 2007). Seu
potencial adipogênico, em cultura, já foi demonstrado (Farrington-Rock et al,
2004). Outra possibilidade para a origem dos adipócitos na lesão é a partir da
célula satélite muscular. Ela é uma célula multipotencial, capaz de exibir
diferenciação miogênica, osteogênica e adipogênica (Asakura et al, 2001) e
que, as condições de hiperóxia, também pode ser induzida em linhagem
adipogênica (Csete et al, 2001). Finalmente, o infiltrado inflamatório
mononuclear pode conter os progenitores pluripotentes de origem medular, que
são mobilizados da medula óssea para os tecidos em processo de inflamação
e regeneração, assim como monócitos que podem conter uma subpopulação
com características de progenitores pluripotentes (Zhao et al. 2003).
As vias de indução ectópica da diferenciação adipocítica no músculo em
degeneração não são conhecidas. A sinalização via MCP1 e seu receptor
CCR2 está expressa no músculo esquelético lesado (Summan et al, 2006;
Warren et al, 2004). A mesma via é responsável da mobilização de macrófagos
em tecido adiposo na síndrome metabólica e diabete (Kanda et al, 2006). A
mobilização de macrófagos e o intenso infiltrado mononuclear no interior de
feixes musculares representam aparentemente uma resposta à mesma
sinalização, que simultaneamente atrai e ativa a linhagem mono-macrofágica.
Em conseqüência da ativação, os macrófagos secretam grande quantidade de
TNFα cujo alvo, entre outros, é a via de sinalização do PPARγ Este gene é o

gene-master de diferenciação de pré-adipócitos em adipócitos, e a sua
ativação pode ser suficiente para converter os progenitores mesenquimais
perivasculares em adipócitos. Semelhante ao tecido adiposo em diabete, os
adipócitos ectópicos podem por sua vez ativar a mesma sinalização, gerando
um circulo auto-sustentado de hipertrofia adiposa. Este quadro se reflete na
dificuldade de reversão da atrofia muscular gordurosa, mesmo depois do
restabelecimento da ligação teno-muscular e solução da sua causa inicial.
As hipóteses citadas devem ser verificadas em estudos futuros, assim
como a viabilidade da proposta de uma terapia celular, que poderia, por vias
parácrinas, reverter o quadro de hipertrofia de adipócitos. Alternativamente, o
aporte de novos progenitores mesenquimais poderá aumentar a diferenciação
de novos adipócitos, e este perigo deve ser adequadamente avaliado.

7. CONCLUSÕES
O modelo de lesão muscular em ovelhas pela ruptura do tendão do
músculo infra-espinal que se assemelhe à lesão do manguito rotator se
mostrou representativo das lesões degenerativas e atrofia gordurosa de
músculo em humanos.
A infiltração progressiva de adipócitos envolveu seqüencialmente
perimísio e endomísio, com evidências morfológicas de atrofia das fibras
musculares em estágios avançado.
O início de diferenciação adipocítica envolveu as regiões perivasculares,
sugerindo a origem de novos adipócitos a partir de pericitos perivasculares.
Os achados sugerem cautela em propostas de terapia celular que
contemple a introdução de células mesenquimais nas regiões em degeneração
gordurosa, pois elas podem exacerbar a proliferação e diferenciação de
progenitores adipocíticos in situ.

FIGURA 1A. Animal após 21 dias de lesão. Microfotografia exibindo feixes musculares separados por células adiposas (seta) e infiltrado inflamatório ( ponta de seta ). Coloração: HE. Aumento original: 100X

FIGURA 1B. Animal após 21 dias de lesão. Microfotografia exibindo espaço interfascicular com infiltração adiposa (seta). Em meio ao fascículo muscular, fibras musculares em degeneração circundadas por infiltrado inflamatório (ponta de seta). Coloração: HE. Aumento original: 200X.

FIGURA 1C. Animal após 21 dias de lesão. Microfotografia exibindo detalhe do infiltrado inflamatório em meio as fibras musculares esqueléticas (seta). Coloração: HE. Aumento original: 400X

FIGURA 1D. Animal após 21 dias de lesão. Microfotografia exibindo fascículos musculares corados intensamente pelo PAS. Nota-se entre fibras musculares presença de células adiposas (seta). Aumento original: 100X

FIGURA 2A. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo ( ∗ ). Coloração: HE. Aumento original: 100X

FIGURA 2B. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo e raros adipócitos presentes em meio ao fascículo muscular. Coloração: HE. Aumento original: 200X.
FIGURA 2C. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares em corte transversal, tendo em área, em torno de célula em degeneração, infiltrado inflamatório mononuclear (seta). Coloração: HE. Aumento original: 400X.
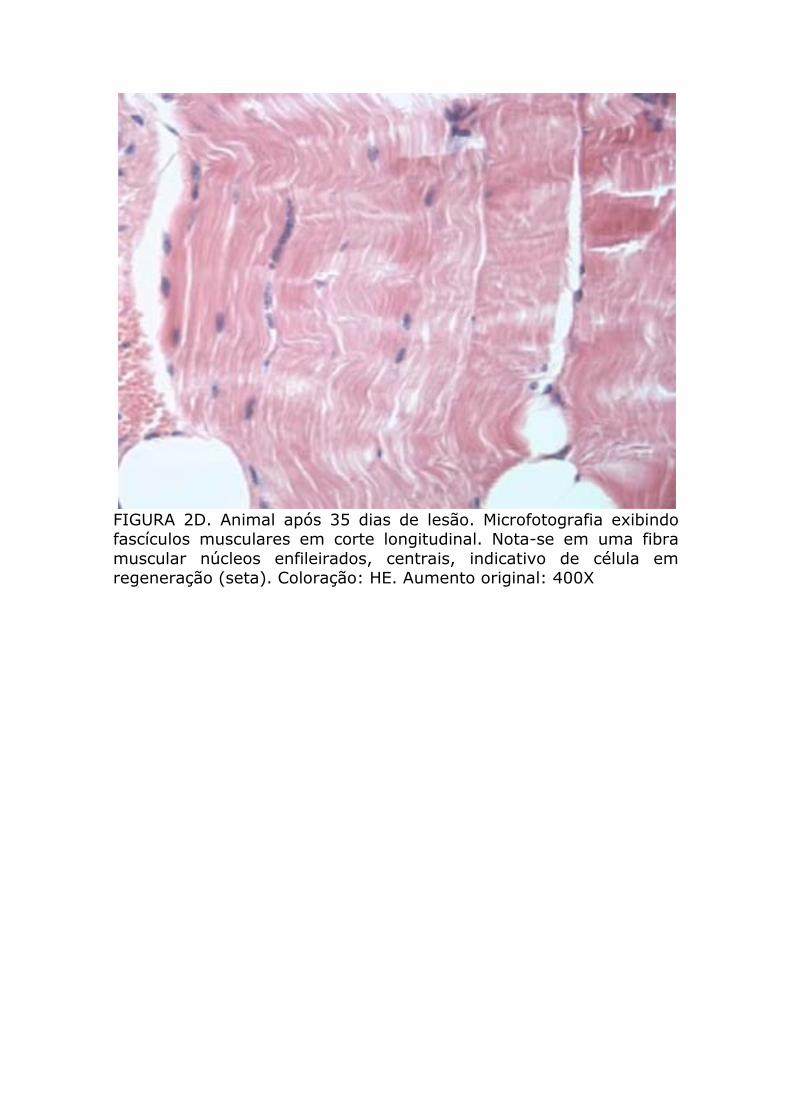
FIGURA 2D. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal. Nota-se em uma fibra muscular núcleos enfileirados, centrais, indicativo de célula em regeneração (seta). Coloração: HE. Aumento original: 400X

FIGURA 3A. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo ( ∗∗∗∗ ) e fibras musculares heterogêneas em relação ao conteúdo de glicogênio ( seta ). PAS. Aumento original: 100X

FIGURA 3B. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo e fibras musculares com ausência quase total de glicogênio (seta). PAS. Aumento original: 100X.

FIGURA 3C. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal exibindo traves fibrosas entre os fascículos musculares (seta). Picro-Sirius. Aumento original: 100X.

FIGURA 3D. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal. Traves fibrosas interfasciculares e mínima deposição de colágeno intrafascicular demonstradas pelo Picro-Sirius. Aumento original: 200X

FIGURA 4A. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo ( ∗ ). Coloração: HE. Aumento original: 100X

FIGURA 4B. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo e adipócitos presentes em meio ao fascículo muscular. Coloração: HE. Aumento original: 200X.
FIGURA 4C. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares em corte transversal, tendo em área, em torno de célula em degeneração, infiltrado inflamatório mononuclear (seta). Coloração: HE. Aumento original: 400X.

FIGURA 4D. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal com alguns adipócitos intrafasciculares ( ∗ ). Nota-se em uma fibra muscular núcleos enfileirados, centrais, indicativo de célula em regeneração (seta). Coloração: HE. Aumento original: 400X

FIGURA 5A. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo ( ∗∗∗∗ ) e fibras musculares com ausência quase completa do conteúdo de glicogênio (seta). Coloração: PAS. Aumento original: 100X

FIGURA 5B. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares separados por tecido adiposo e fibras musculares com ausência total de glicogênio (seta). Coloração: PAS. Aumento original: 200X.
FIGURA 5C. Animal após 60 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal exibindo intensa degeneração adiposa e traves fibrosas entre os fascículos musculares (seta). Coloração: Picro-Sirius. Aumento original: 100X.

FIGURA 5D. Animal após 35 dias de lesão. Microfotografia exibindo fascículos musculares em corte longitudinal. Traves fibrosas interfasciculares e mínima deposição de colágeno intrafascicular demonstradas pelo Picro-Sirius. Aumento original: 200X

FIGURA 6A. Animal após 35 dias de lesão. Microfotografia exibindo vasos capilares corados pela Griffonia simplicifora isolectina I (GS-I) biotinilada e revelada pela estreptavidina-peroxidase, utilizando a diaminobenzidina como substrato cromógeno. Contracoloração: hematoxilina. Aumento original: 100X.

FIGURA 6B. Animal após 60 dias de lesão. Microfotografia exibindo a reatividade da lectina (GS-I) em vasos. Reação histoquímica para lectina. Contracoloração: hematoxilina Aumento original: 100X.
10
20
30
*
***
*
***
mer
o d
e V
aso
s

FIGURA 6C. Representação gráfica da densidade capilar no músculo esquelético de ovelhas após a desinserção do tendão do músculo infraespinhal. Houve diferença significativa da densidade vascular no músculo infraespinhal animais. Dados representados pela média e desvio padrão da densidade capilar após 35 e 60 dias da lesão. ∗∗∗ ; ∗ : grupos diferentes entre si, com significância estatística (p<0,05).

FIGURA 7A. Animal após 21 dias de lesão. Microfotografia exibindo fascículo muscular tendo no espaço interfascicular algumas células adiposas, algumas delas em íntimo contacto com a parede da vênula (seta). Coloração: HE. Aumento original: 400X.

FIGURA 7B. Animal após 21 dias de lesão. Microfotografia exibindo o mesmo aspecto anterior. Coloração: HE. Aumento original: 400X.

FIGURA 7C. Animal após 21 dias de lesão. Microfotografia exibindo infiltração adiposa em espaço interfascicular. Notar a parede venular em contato com adipócitos (seta). Coloração: HE. Aumento original: 400X.

FIGURA 7D. Animal após 60 dias de lesão. Microfotografia exibindo espaço inerfascicular com infiltração adiposa e células inflamatórias (seta). Coloração: HE. Aumento original: 200X

FIGURA 8A. Animal após 35 dias de lesão. Microfotografia de corte histológica de músculo infra-espinhal lesado corado pela técnica imunohistoquímica para a detecção da isoforma alfa de músculo liso revelada pela técnica da avidina-biotina-peroxidase. A cor marrom da diaminobenzidina identifica a actina alfa de músculo liso existente em pericitos, em torno de capilares (seta). Notar a íntima relação da parede capilar com o adipócito. Aumento original: 400X.

FIGURA 8B e 8C. Animal após 35 dias de lesão. Microfotografia de músculo infra-espinhal lesado corado pela técnica imunohistoquímica para a detecção da isoforma alfa de músculo liso revelada pela técnica da avidina-biotina-peroxidase. A cor marrom da diaminobenzidina identifica a actina alfa de músculo liso existente em pericitos, em torno de capilares (seta). Células adiposas presentes em íntima relação com a parede de capilares e/ou vasos de pequeno calibre. Aumento original: 400X.

FIGURA 8D. Animal após 35 dias de lesão. Microfotografia de tecido muscular lesado corado para a detecção da isoforma alfa de músculo liso e revelada pela técnica da avidina-biotina-peroxidase. Reatividade para pericitos e camada muscular de vasos evidente no espaço interfascicular (seta). Células adiposas presentes em íntima relação com a parede de capilares e/ou vasos de pequeno calibre. Aumento original: 200X.

8 REFERÊNCIAS BIBLIOGRÁFICAS
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Biologia molecular da célula. São Paulo: Artmed, 2004. Alderton JM & Steinhardt RA. How calcium influx through calcium leak channels is responsible for the elevated levels of calcium-dependent proteolysis in dystrophic myotubes. Trends Cardiovasc Med 2000; 10: 268–272. Asakura A, Komaki M, Rudnicki M. Muscle satellite cells are multipotential stem cells that exhibit myogenic, osteogenic, and adipogenic differentiation. Differentiation 2001, 68: 245–253
Bischoff R. Proliferation of muscle satellite cells on intact myofibers in culture. Dev Biol. 1986, 115(1):129-39.
Beauchamp JR, Heslop L, Yu DSW, Tajbakhsh S, Kelly RG, Wernig A, Margaret E. Buckingham, Terence AP, Zammit PS. Expression of CD34 and Myf5 Defines the Majority of Quiescent Adult Skeletal Muscle Satellite Cells. Journal of Bone and Joint Surgery, 2000, 54:41-50.
Baker H, Hall-Craggs ECB. Recovery from the central core degeneration of the tenotomized rat soleus muscle. Muscle Nerve 1980; 3:151–9. Belcastro AN, Shewchuk LD, and Raj DA. Exercise-induced muscle injury: a calpain hypothesis. Mol Cell Biochem 1998; 179: 135–145. Bjorkenheim JM. Structure and function of the rabbit’s supraspinatus muscle after resection of its tendon. Acta Orthop Scand. 1989; 60:461-3. Blaveri K, Heslop L, Yu DS, Rosenblatt JD, Gross JG, Partridge TA, Morgan JE. Patterns of repair of dystrophic mouse muscle: studies on isolated fibers. Dev Dyn 1999; 216: 244–256. Brachvogel B, Moch H, Pausch F, Schlotzer-Schrehardt U, Hofmann C, Hallmann R, von der Mark K, Wrinkler T, Poschl E. Perivascular cells expressing annexin A5 define a novel mesenchymal stemcell-like population with the capacity to differentiate into multiple mesenchymal lineages. Development 2005; 132:2657-2668. Brighton CT, Lorich DG, Kupcha R, Reilly TM, Jones AR, Woodbury RA 2nd. The pericyte as a possible osteoblast progenitor cell. Clin Orthop 1992; 275:287-299. Chargé SBP & RudnickI MA. Cellular and Molecular Regulation of Muscle Regeneration. Physiol Rev 2004; 84: 209–238.

Coleman SH, Fealy S, Ehteshami JR, MacGillivray JD, Altchek DW, Warren RF, Turner AS. Chronic Rotator Cuff Injury and Repair Model in Sheep. J Bone Joint Surg Am 2003; 85: 2391-2402 Colégio Brasileiro de Experimentação Animal Princípios éticos na experimentação animal. São Paulo. Colégio Brasileiro de Expeimentação Animal, julho 1991. Cooper RN, Tajbakhsh S, Mouly V, Cossu G, Buckingham M, Butler GSB. In vivo satellite cell activation via Myf5 and MyoD in regenerating mouse skeletal muscle. J. Cell Sci. 1999, 112:2895–2901. Crawford GN. Some effects of tenotomy on adult striated muscles. J Anat.1977; 123:389-96. Csete M, Walikonis J, Slawny N, Wei Y, Korsnes S, Doyle JC, Wold B. Oxygen-mediated regulation of skeletal muscle satellite cell proliferation and adipogenesis in culture. J Cell Physiol 2001; 189:189-196. Decary S, Mouly V, Hamida CB, Sautet A, Barbet JP, Butler-Browne GS. Replicative potential and telomere length in human skeletal muscle: implications for satellite cell-mediated gene therapy. Hum Gene Ther 1997; 8: 1429–1438. Diaz-Flores L, Gutierrez R, Lopez-Alonso A, Gonzalez R, Varela H. Pericytes as a supplementary source of osteoblasts in periosteal osteogenesis. Clin Orthop Relat Res. 1992; 275:280-286. Diaz-Flores L, Gutierrez R, Gonzalez P, Varela H. Inducible perivascular cells contribute to the neochondrogenesis in grafted perichondrium. Anat Rec. 1991; 229:1-8. Doherty MJ, Ashton BA, Walsh S, Beresford JN, Grant ME, Canfield AE. Vascular pericytes express osteogenic potential in vitro and in vivo. J Bone Miner Res. 1998;13:828-838. Edwards TB, Boulahia A, Kempf JF, Boileau P, Nemoz C, Walch G. The influence of rotator cuff disease on the results of shoulder arthroplasty for primary osteoarthritis: results of a multicenter study. J Bone Joint Surg Am. 2002; 84:2240-8. Eccles JC. Investigations on muscle atrophies arising from disuse and tenotomy. J Physiol 1943;103: 253–266. Fanin M, Nardetto L, Nascimbeni AC, Tasca E, Spinazzi M, Padoan E, Angelini C. Correlations between cçlinical severity, genotype and muscle pathology in limb girdle muscular dystrophy type @a. J Mwed Genet 2007; 44:609-614.

Farrington-Rock C, Crofts NJ, Doherty MJ, Ashton BA, Griffin-Jones C, Canfield AE. Chondrogenic and adipogenic potential of microvascular pericytes. Circulation. 2004;110:2226-2232. Fuchs B, Weishaupt D, Zanetti M, Hodler J, Gerber C. Fatty degeneration of the muscles of the rotator cuff: assessment by computed tomography versus magnetic resonance imaging. J Shoulder Elbow Surg. 1999; 8:599-605. Fuchtabuer EM, Westphal H. MyoD and Myogenin are coexpressed in regenerating skeletal muscle of the mouse. Developmental Dynamics, 1992, v.193, p.34-39. Gerber C, Fuchs B, Hodler J. The results of repair of massive tears of the rotator cuff. J Bone Joint Surg Am. 2000; 82:505-15. Gerber C, Meyer DC, Schneeberger AG, Hoppeler H, von Rechenberg B. Effect of tendon release and delayed repair on the structure of the muscles of the rotator cuff: an experimental study in sheep. J Bone Joint Surg Am 2004; 86:1973-1982.
Gladstone JN, Bishop JY, Lo IKY, Flatow EL. Fatty Infiltration and Atrophy of the Rotator Cuff Do Not Improve After Rotator Cuff Repair and Correlate With Poor Functional Outcome. The American Journal of Sports Medicine, 2007,Vol. 35, No. 5.
Gordon LW, Tracy H, Dawn M, Xin G, Lyndell M, Laura O, William AK, Petia PS. Chemokine receptor CCR2 involvement in skeletal muscle regeneration. FASEB J. 2005 Mar;19(3):413-5. Goutallier D, Postel JM, Bernageau J, Lavau L, Voisin MC. Fatty muscle degeneration in cuff ruptures. Clin Orthop Relat Res 1994; 304:78–83. Goutallier D, Postel JM, Bernageau J, Lavau L, Voisin MC. Fatty infiltration of disrupted rotator cuff muscles. Rev Rheum Engl Ed 1995; 62:415–22. Goutallier D, Bernageau J, Patte D. L’évaluation par le scanner de la trophicité des muscles des coiffes des rotateurs avant une rupture tendineuse. Rev Chir Orthop. 1989;75:126-7. Goutallier D, Postel JM, Bernageau J, Lavau L, Voisin MC. Fatty muscle degeneration in cuff ruptures. Pre- and postoperative evaluation by CT scan. Clin Orthop. 1994; 304:78-83. Goutallier D, Postel JM, Lavau L, Bernageau J. Impact of fatty degeneration of the supraspinatus and infraspinatus muscles on the prognosis of surgical repair of the rotator cuff. Rev Chir Orthop Reparatrice Appar Mot. 1999; 85:668-76.

Grounds MD, Garret KL, Lai MC, Wright WE, Beilharz MW. Identification of muscle precursor cells in vivo by use of MyoD1 and myogenin probes. Cell Tiss. Res. 1992, 267:99–104. Hack AA, Cordier L, Shoturma DI, Lam MY, Sweeney HL, McNally EM. Muscle degeneration without mechanical injury in sarcoglycan deficiency. Proc Natl Acad Sci U S A 1999 Sep 14;96(19). Hack AA, Cordier L, Shoturma DI, Lam MY, Sweeney HL, McNally EM. Muscle degeneration without mechanical injury in sarcoglycan deficiency. Proc Natl Acad Sci U S A. 1999; 96: 10723–10728. Hall-Craggs EC and Seyan HS. Histochemical changes in innervated and denervated skeletal muscle fibers following treatment with bupivacaine (marcain). Exp Neurol 1975; 46:345–354.
Hasty P, Bradley A, Morris JH, Edmondson DG, Venuti JM, Olson EN, Klein WH. Muscle deficiency and neonatal death in mice with a targeted mutation in the myogenin gene. Nature, 1993, 364, 501 – 506.
Hausman GJ & Poulos SP. A method to establish cocultures of myotubes and preadipocytes from collagenase digested neonatal pig semitendinous muscles. J Anim Sci 2005; 83:1010-1016. Hausman GJ & Richardson RL. Newly recruited and pre-existing preadipocytes in cultures of porcine stromal-vascular cells: morphology, expression of extracellular matrix components, and lipid accretion. J Anim Sci 1998; 76:48-60. Huxley AF. Cross-bridge action: present views, prospects, and unknowns. J Biomech 2000; 33: 1189–1195. Josza L, Kannus P, Thoring J, Reffy A, Jarvinen M, Kvist M. The effect of tenotomy and immobilisation on intramuscular connective tissue. A morphometric and microscopic study in rat calf muscles. J Bone Joint Surg Br. 1990; 72 B:293-7. Junqueira LC, Carneiro J. Histologia Básica. 10ed. Rio de janeiro: Guanabara Koogan, 2004. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, Kitazawa S, Miyachi H, Maeda S, Egashira K, Kasuga M. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest 2006; 116: 1494–1505. Kiernan J. A. Histological & Histochemical methods. Theory and practice. Pergamon Press, Oxford, Second edition. 1990. pp 433.

Koishi K, Zhang M, Mclennan IS, Harris J. MyoD protein accumulates in satellite cells and is neurally regulated in regenerating myotubes and skeletal muscle fibers. Developmental Dynamics,1994 v.202, p. 244-254. Kuschel R, Reuveni ZY, Bornemann A. Satellite cells on isolated myofibers from normal and denervated adult rat muscle. The Journal of Histochemistry & Citochemistry, 1999, v.47, n.11, p. 1375-1383. Lange S, Ehler E, Gautel M. Multicompartment proteins in the sarcomere. Trends in Cell Biology, 2006, v.16, n.1, p. 11-18. Lescaudron L, Peltékian E, Fontaine-Pérus J, Paulin D, Zampieri M, Garcia L, Parrish E. Blood borne macrophages are essential for the triggering of muscle regeneration following muscle transplant. Neuromuscul Disord 1999; 9:72-80. Matsumoto F, Uhthoff HK, Trudel G, Loehr JF. Delayed tendon reattachment does not reverse atrophy and fat accumulation of the supraspinatus––an experimental study in rabbits. J Orthop Res 2002; 20:357–63. Matsumoto F, Uhthoff HK, Trudel G, Loehr JF. Delayed tendon reattachment does not reverse atrophy and fat accumulation of the supraspinatus – an experimental study in rabbits. J Orthop Res 2002; 20: 357-363. MCComas, AJ. Skeletal muscle: form and function. Champaig: Human Kinetics,1996. McLENNAN IS. Degenerating and regenerating skeletal muscles contain several subpopulations of macrophages with distinct spatial and temporal distributions. J. Anat. 1996; 188: 17-28. Meyer DC, Pirkl C, Pfirrmann CWA, Zanetti M, Christian Gerber Asymmetric atrophy of the supraspinatus muscle following tendon tear. Journal of Orthopaedic Research 23; 2005, 254–258. Meyer DC, Hoppeler H, von Rechenberg B, Gerber C. A pathomechanical concept explains muscle loss and fatty muscular changes following surgical tendon release. J Orthop Res 2004; 22: 1004-1007. Merly F, Lescaudron L, Rouaud T, Crossin F, Gardahaut MF. Macrophages enhance muscle satellite cell proliferation and delay their differentiation. Muscle Nerve 1999; 22:724-732. Moss, F.P. & Leblond CP. Satellite cells as the source of nuclei in muscles of growing rats. Anat. Rec. 1971; 170:421–435.
Mukesh S, Gordon LW, Robert RM, Rebecca C, Tracy H, Nico VR, Petia PS. Macrophages and skeletal muscle regeneration: a clodronate-containing

liposome depletion study. Am J Physiol Regul Integr Comp Physiol, 2006, 290: R1488-R1495.
Nakagaki K, Ozaki J, Tomita Y, Tamai S. Fatty degeneration in the supraspinatus muscle after rotator cuff tear. J Shoulder Elbow Surg 1996; 5: 194–200. Nakagaki K, Ozaki J, Tomita Y, Tamai S. Fatty degeneration in the supraspinatus muscle after rotator cuff tear. J Shoulder Elbow Surg. 1996; 5:194-200.
Neer CS. Anterior Acromioplasty for the Chronic Impingement Syndrome in the Shoulder. J Bone Joint Surg Am. 1972 Jan;54(1):41-50.
Padilla J, Fielding WC, Belcastro AN, Gardiner PF, Taylor AW. The effect of diazepan and exercise training on selected biochemical and histochemical properties of rat skeletal muscle. Acta Physiol Pharmacol Ther Latinoam; 1997, 47(4):203-10. Pfirrmann CW, Zanetti M, Weishaupt D, Gerber C, Hodler J. Subscapularis tendon tears: detection and grading at MR arthrography. Radiology. 1999; 213:709-14. Rudnicki MA, Schnegelsberg PNJ, Stead RH, Braun T, Arnold HH, Jaenisch R. MyoD or Myf-5 is required for the formation of skeletal muscle. Cell., 1993, 75, 1351-1359. Ryan AS, Nicklas BJ, Berman DM. Racial differences in insulin resistance and mid-thigh fat deposition in postmenopausal women. Obes Res, 2002, 10:336–44. Rubino LJ, Stills HF, Sprott DC, Crosby LA. Fatty infiltration of the torn rotator cuff worsens over time in a rabbit model. Arthroscopy. 2007; 23: 717-722. Schmalbruch H & Lewis DM. Dynamics of nuclei of muscle fibers and connective tissue cells in normal and denervated rat muscles. Muscle Nerve 2000; 23: 617–626. Snow MH. Myogenic cell formation in regenerating rat skeletal muscle injured by mincing. II. An autoradiographic study. Anat Rec 1977; 188: 201–217. Snow MH. An autoradiographic study of satellite cell differentiation into regenerating myotubes following transplantation of muscles in young rats. Cell Tissue Res 1978; 186: 535–540. Schultz, E., Gibson MC, Champion T. Satellite cells are mitotically quiescent in mature mouse muscle: an EM and radioautographic study. J.Exp. Zool. 1978; 206:451–456.

Shi S & Gronthos S. Perivascular niche of post-natal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res 2003; 18:696-704. Sizinio et al. Ortopedia e Traumatologia: Princípios e Pratica. 3º Ed. São Paulo: Artmed, 2003. p.185-237 Sobotta,J. Atlas de Anatomia Humana. Rio de Janeiro, Editora Guanabara Koogan, Vol.I e II, 2000 Summan M, Warren GL, Mercer RR, Chapman R, Hulderman T, Van Rooijen N, Simeonova PP. Macrophages and skeletal muscle regeneration: a clodronate-containing liposome depletion study. Am J Physiol Regul Integr Comp Physiol.; 2006, 290(6).
Tajbakhsh S, Rocancourt D, Buckingham M. Muscle progenitor cells failing to respond to positional cues adopt non-myogenic fates in myf-5 null mice. Nature, 1996, 384, 266 – 270.
Thomazeau H, Boukobza E, Morcet N, Chaperon J, Langlais F. Prediction of rotator cuff repair result by magnetic resonance imaging. Clin Orthop. 1997; 344:275-83. Tidball JG. Inflammatory cell response to acute muscle injury. Med Sci Sports Exercise 1995; 27: 1022–1032. Tomanek R, Cooper RR. Ultrastructural changes in tenotomized fast- and slow-twich muscle fibres. J Anat 1972; 113: 151–9. Uhthoff HK, Trudel G, Himori K. Relevance of pathology and basic research to the surgeon treating rotator cuff disease J Orthop Sci 2003; 8: 449–456 Xaymardan M, Gibbins JR, Zoellner H. Adipogenic healing in adult mice by implantation of hollow devices in muscle. Anat Rec, 2002, 267:28–36. Warren, G. L., O'Farrell, L., Summan, M., Hulderman, T., Mishra, D., Luster, M. I., Kuziel,W. A., Simeonova, P. P. Role of CC chemokines in skeletal muscle functional restoration after injury. Am. J. Physiol. Cell Physiol. 2004; 286, C1031–C1036. Wigmore PM, Evans DJR. Molecular and cellular mechanisms involved in the generation of fiber diversity during myogenesis. International Review of Cytology,2002, 216, 175-232. Wigmore PM & Evans DJ. Molecular and cellular mechanisms involved in the generation of fiber diversity during myogenesis. Int Rev Cytol, 2002, 216: 175–232.

Whalen RG, Harris JB, Browne GS, Sesodia S. Expression of myosin isoforms during notexin-induced regeneration of rat soleus muscles. Dev. Biological, 1990, v. 141, p. 24-40.
Yablonka RZ, Rivera AJ. Temporal expression of regulatory and structural muscle proteins during myogenesis of satellite cells on isolated adult rat fibers. The Journal of Cell Biology, 2000, Volume 151, Number 6, 1221-1234.
Zammit, P. & Beauchamp J. The skeletal muscle satellite cell: stem cell or son of stem cell? Differentiation. 2001; 68:193–204. Zammit, P.S., L. Heslop, V. Hudon, J.D. Rosenblatt, S. Tajbakhsh, M.E. Buckingham, J.R. Beauchamp, and T.A. Partridge. Kinetics of myoblast proliferation show that resident satellite cells are competent to fully regenerate skeletal muscle fibers. Exp. Cell Res. 2002; 281:39–49. Zhao Y, Glesne D, Huberman E. A human peripheral blood monocyte-derived subset acts as pluripotent stem cells. Proc Natl Acad Sci 2003; 100: 2426–2431.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo