and jennifer r. shelton
TRANSCRIPT

Domain-Speci�c Knowledge Systems in theBrain: The Animate-Inanimate Distinction
Alfonso Caramazza and Jennifer R. SheltonHarvard University
Abstract
We claim that the animate and inanimate conceptual cate-gories represent evolutionarily adapted domain-speci�c knowl-edge systems that are subserved by distinct neural mechanisms,thereby allowing for their selective impairment in conditionsof brain damage. On this view, (some of) the category-speci�cde�cits that have recently been reported in the cognitiveneuropsychological literature—for example, the selective dam-age or sparing of knowledge about animals—are truly categori-cal effects. Here, we articulate and defend this thesis againstthe dominant, reductionist theory of category-speci�c de�cits,which holds that the categorical nature of the de�cits is theresult of selective damage to noncategorically organized visual
or functional semantic subsystems. On the latter view, thesensory/functional dimension provides the fundamental organ-izing principle of the semantic system. Since, according to thelatter theory, sensory and functional properties are differen-tially important in determining the meaning of the membersof different semantic categories, selective damage to the visualor the functional semantic subsystem will result in a category-like de�cit. A review of the literature and the results of a newcase of category-speci�c de�cit will show that the domain-speci�c knowledge framework provides a better account ofcategory-speci�c de�cits than the sensory/functional dichot-omy theory.
INTRODUCTION
How is conceptual knowledge organized in the brain?Evidence relevant to this issue is provided by the pat-terns of semantic processing de�cits in brain-damagedsubjects, especially those de�cits that seem to selectivelyaffect one or another semantic category. The �rst cleardemonstration of a semantic category-speci�c de�citwas made by Warrington and Shallice (1984). They re-ported four patients recovering from herpes simplexencephalitis who were disproportionately impaired inproducing and understanding the names of living things.This result, coupled with the observation of a brain-dam-aged subject who showed the reverse dissociation—thatis, relative sparing of the comprehension of living rela-tive to nonliving things1 (Warrington & McCarthy,1983)—invited the inference that brain damage couldresult in category-speci�c semantic de�cits.2
Although the four cases described by Warrington andShallice (1984) were not all tested with the same tasksor the same materials, they all showed poorer perfor-mance in producing and/or comprehending the namesof living as opposed to nonliving things. In some casesthe contrast in performance was quite striking: J.B.R. wasonly able to recognize or name 2 of 48 living things(animals or plants) but accurately described or named45 of 48 nonliving things; similarly, S.B.Y. failed to iden-tify any of the living items but correctly identi�ed 36 of48 nonliving things. For example, J.B.R. de�ned briefcase
© 1998 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 10:1, pp. 1–34
as “small case used by students to carry papers” butresponded “don’t know” to the word parrot; similarly,S.B.Y. de�ned towel as “material used to dry people” butresponded “bird that �ies” to the word wasp. The othertwo cases reported by Warrington and Shallice, K.B. andI.N.G., could not be tested with the de�nition produc-tion tasks because of their severe language productionde�cits, but their performance in word/picture matchingtasks was poorer for animals than for nonliving things.Thus, all four cases showed disproportionate dif�cultyfor living than for nonliving things.
Con�rmation of the dissociation of performance forliving and nonliving things was soon obtained in manyother studies of brain-damaged subjects of various etiolo-gies, including other cases of herpes simplex encephali-tis, trauma, cerebro-vascular accident, and neurosurgery(Basso, Capitani, & Laiacona, 1988; Damasio, Grabowski,Tranel, Hichwa, & Damasio, 1996; De Renzi & Lucchelli,1994; Farah, Hammond, Mehta, & Radcliff, 1989; Farah,McMullen, & Meyer, 1991; Hart & Gordon, 1992; Hillis &Caramazza, 1991; Laiacona, Barbarotto, & Capitani, 1993;Laiacona, Capitani, & Barbarotto, 1997; McCarthy & War-rington, 1988; Pietrini, Nertempi, Vaglia, Revello, Pinna, &Ferro-Milone, 1988; Powell & Davidoff, 1995; Sartori &Job, 1988; Sheridan & Humphreys, 1993; Silveri & Gai-notti, 1988; Sirigu, Duhamel, & Poncet, 1991). And therehave also been several reports documenting the reversepattern of dissociations—that is, cases of impaired per-formance in naming and/or recognizing nonliving ob-

jects in the face of spared ability with living things (Hillis& Caramazza, 1991; Sacchett & Humphreys, 1992; War-rington & McCarthy, 1983, 1987). It would seem, then,that the category of living things can be damaged inde-pendently of the category of nonliving things, thus invit-ing the inference that the semantic system might beorganized categorically. Furthermore because most ofthe patients with selective impairment of the living cate-gory had sustained damage to (at least) the left temporallobe, it might be further surmised that the latter neuralstructure is part of a neural network speci�cally dedi-cated to representing conceptual knowledge about liv-ing objects.3
Despite the seemingly categorical nature of the de�cit,the inference that the semantic system is organized cate-gorically has never been entertained seriously (but seeWarrington, 1981). The obstacle most often cited againsta categorical interpretation of the category-speci�c se-mantic de�cits was apparent from the earliest results:The patterns of semantic categories affected in any onepatient did not correspond to plausible, clear-cut cate-gorical distinctions such as the living/nonliving or natu-ral kind/artifact dichotomies. For example, the early,paradigm cases J.B.R. and S.B.Y were both impaired notonly in processing the animal and plant categories butalso foods. Clearly, there is no obvious way in which thefood category could be easily subsumed within the livingthing category (think of spaghetti, rice pudding, etc.).Furthermore, extensive testing of J.B.R. showed that hewas also impaired in de�ning musical instruments, pre-cious stones, cloths, and metals—all inanimate objects—but not body parts (which according to some authorsshould be associated with living things). These violationsof the simple dichotomy of living/nonliving things ledWarrington & Shallice (1984, see also Warrington &McCarthy, 1983, 1987) to propose the sensory/functionaltheory of category-speci�c de�cits. They argued thatfunctional and visual (sensory) attributes are not onlydifferentially important for the identi�cation of nonlivingand living things, respectively, but also for other catego-ries such as food. Damage to the visual semantic subsys-tem results in disproportionate impairment of thecategories of living things and foods because the iden-ti�cation of the members of these categories dependscrucially on their visual (sensory) features, and damageto the functional semantic subsystem results in dispro-portionate impairment of nonliving things because iden-ti�cation of the members of this category of objectsdepends crucially on their functional, nonsensory attrib-utes. In other words, the seemingly categorical nature ofsemantic de�cits does not re�ect the categorical organi-zation of conceptual knowledge in the brain but ismerely the accidental consequence of a more basic,noncategorical organizing principle of the semantic sys-tem.4
The rejection of the possibility that conceptual knowl-edge might be organized categorically in the brain may
have been premature. Here we show that the evidenceusually cited in favor of reductionist accounts of cate-gory-speci�c de�cits is far from compelling. However,before doing so we consider whether the reported cate-gory effects are real or just artifacts of uncontrolledstimulus variables.
Are Category-Speci�c De�cits Artifacts ofUncontrolled Stimulus Factors?
Warrington and Shallice (1984) raised and dismissed thepossibility that the category-speci�c de�cits they re-ported might have arisen from frequency or familiaritydifferences between the members of the living and non-living categories. In an identi�cation experiment withword and picture stimuli, their subject S.B.Y. performedfar worse with the living than the nonliving categoryeven when familiarity and frequency were taken intoconsideration through covariance analyses. This precau-tion has not always been observed in other studies (oreven the other cases in Warrington and Shallice’s paper).It is not inappropriate, therefore, for Funnell and Sheri-dan (1992), Gaffan and Heywood (1993), and Stewart,Parkin, and Hunkin (1992) to raise the question ofwhether putative cases of category-speci�c de�cits forliving things might not simply re�ect the fact that theseitems tend to be of lower frequency and familiarity andmore visually complex than nonliving things. This con-cern is not unfounded because frequency, familiarity, andvisual complexity have powerful effects on perfor-mance.
Stewart et al. (1992) provide the clearest demonstra-tion of the need for caution in interpreting putativecases of category-speci�c de�cit of living things. Theyreport a case, H.O., with herpes simplex encephalitis,whose performance in naming the Snodgrass and Van-derwart (1980) pictures showed a signi�cant categoryeffect, with poorer performance for the living than non-living category (65% versus 86%). The difference be-tween categories remained even when the lists werematched for word frequency (42% versus 80% for livingand nonliving, respectively). However, when the twocategories were matched jointly for frequency, familiarity,and visual complexity, the category effect disappeared(42% versus 36% for living and nonliving things, respec-tively). Furthermore, when the categories were rede�nedin terms of the extent to which sensory (animals, vege-tables, fruit, large buildings, and natural features) versusfunctional (tools and kitchen utensils) attributes are sup-posedly more important in determining the meaning ofcategory members, there again was no difference be-tween categories when frequency, familiarity, and visualcomplexity were jointly controlled (57% versus 58%,respectively). These results, together with those of Fun-nell and Sheridan (1992), who showed that item famili-arity can be a major determinant of naming accuracy, andthe results of Gaffan and Heywood (1993), who showed
2 Journal of Cognitive Neuroscience Volume 10, Number 1

that living and nonliving things differ in visual dis-criminability and consequent ease of recognition, indi-cate that putative cases of selective de�cit of the livingcategory may not, in fact, be true category-speci�cde�cits but may instead re�ect category nonspeci�cprocessing dif�culties that are more severe for low-frequency, low-familiarity, and hard-to-discriminate visualobjects.
The results reported by Funnell and Sheridan (1992),Gaffan and Heywood (1993), and Stewart et al. (1992)demand that we adopt a cautious stance in interpretingsupposed cases of category-speci�c de�cits. Nonethe-less, there is compelling evidence that not all putativecases of category-speci�c de�cits can be explained byappeal to between-category variation in item frequency,familiarity, and visual complexity. First, just as was thecase for S.B.Y. (Warrington & Shallice, 1984), other casesof disproportionate dif�culty in naming and recognizingliving things continue to show the effect even whenfrequency, familiarity, and visual complexity are taken inconsideration (e.g., Farah, Meyer, & McMullen, 1996; Gai-notti & Silveri, 1996; Hart & Gordon, 1992; Kurbat, 1997;Laiacona, Barbarotto, & Capitani, 1993, Laiacona et al.,1997; Sartori, Miozzo, & Job, 1993; Sheridan & Hum-phreys, 1993). Thus, for example, Sartori et al. retestedtheir subject Michelangelo (Sartori & Job, 1988) withanimals and artifacts matched for frequency, familiarity,and visual complexity and once again found that henamed animals signi�cantly less accurately than artifacts(30% versus 70%). Second, there are a number of reportsof disproportionate de�cit for the category of nonlivingthings (Hillis & Caramazza, 1991; Sacchett & Humphreys,1992; Warrington & McCarthy, 1983, 1987)—the suppos-edly easier-to-process category.
Especially informative in this context is Hillis andCaramazza’s (1991) report of two cases with contrastingcategory-speci�c de�cits who were tested with the samestimuli on the same tasks. One subject, P.S., who hadsustained bilateral temporal lobe and left frontal damagefrom a severe blow to the head, showed a disproportion-ate de�cit with living things; the other case, J.J., who hadsustained left temporal lobe and basal ganglia damagefollowing a thromboembolic stroke, showed a dispropor-tionate de�cit for inanimate objects. The results reportedfor these two cases are striking: over seven administra-tions of a naming test, at 4 months postinjury, J.J. namedanimals (n = 46 in each test) and inanimate objects(including fruits and vegetables; n = 98 in each test) withan average accuracy of 81% and 12%, respectively; P.S.named living things (animals, fruits, and vegetables) andnonliving things with an average accuracy of 47% and93%, respectively. The two subjects were tested again 13months postinjury, when their performance had im-proved substantially. Their respective category-speci�cde�cits were still clearly apparent: J.J. named correctlyall animals but only 68% of inanimate objects; P.S. namedcorrectly all nonliving objects but only 64% of living
things. These patients’ category-speci�c de�cits were notrestricted to naming tasks but extended to comprehen-sion tasks: J.J. demonstrated comprehension of all animalnames but de�ned incorrectly or gave ambiguous re-sponses to 23 of 98 nonanimal names; P.S. understoodcorrectly all names of artifacts but de�ned incorrectly orgave ambiguous de�nitions to 11 of 70 animal names.Clearly, it is not possible to explain these contrastingpatterns of category-speci�c de�cits by appealing simplyto between-category variation in item frequency, famili-arity, and/or visual complexity. Indeed, the selective spar-ing of the animals category in J.J. unambiguouslyindicates that conceptual knowledge can be damagedalong the boundaries of semantic categories.
The conclusion to be drawn from the fact that selec-tive impairment of the living category is not alwaysreducible to an effect of frequency, familiarity, and/orvisual similarity (e.g., Sartori et al., 1993) and the fact thatknowledge of the living category can be selectivelyspared (e.g., Hillis & Caramazza, 1991) is not, however,that all putative cases of category-speci�c de�cits are tobe taken as such. Funnell and Sheridan’s (1992) andStewart et al.’s (1992) admonition against the facile in-terpretation of performance differences between catego-ries of items as having a categorical basis remainsgenerally valid—it is valid for the brain-damaged subjectsthey studied, and it is likely to be valid for some of theother putative cases of category-speci�c de�cit.
There are at least three implications that follow fromthe foregoing discussion. These concern the frequencyof occurrence of different types of category-speci�cde�cits, the types of categories affected, and the puta-tively disproportionate impairment in processing visualversus functional attributes by subjects with category-speci�c de�cit for living things. The �rst two points areobvious, if not less important because of that. Althoughthere is a clear asymmetry in the number of reportedcases of living versus nonliving category-speci�c de�cits(e.g., Gainotti & Silveri, 1996; Saffran & Schwartz, 1994),we cannot be sure of the magnitude of the putativeasymmetry because most of the reported studies did notmatch category items on the variables of familiarity andvisual complexity. Similarly, although there are many re-ports of de�cit or sparing of one or another highlyspeci�c category (e.g., musical instruments, gemstones,geographical places, outdoor objects, etc.) their validitymust remain in question because the items in thesecategories were also not speci�cally controlled for nui-sance variables that might have led to their spuriouslylow or high levels of performance. And, as we shall arguemore extensively below, the cautionary moral to bedrawn from Stewart et al.’s (1992) paper, in particular,extends beyond the issue of whether a differential pat-tern of performance across categories is to be ascribeda categorical status. The implications also extend toclaims about various cases of putatively disproportionateimpairment in processing visual versus functional infor-
Caramazza and Shelton 3

mation for the category of living things, because in thesecases, too, there may have been inadequate control ofthe relative dif�culty of visual and functional test items.
Having argued that at least some category-speci�cde�cits are real category effects (i.e., not artifacts ofuncontrolled stimulus factors), we now consider indetail two reductionist accounts of category-speci�cde�cits.
The Sensory/Functional Theory
The received reductionist account of category-speci�cde�cits is based on the role of sensory and functionalproperties in determining the meaning of living andnonliving things (Allport, 1985; Gainotti & Silveri, 1996;Hart & Gordon, 1992; Shallice, 1988; Silveri & Gainotti,1988; Warrington & McCarthy, 1983, 1987; Warrington &Shallice, 1984). There are several variants of this theory,but they all share two assumptions: (1) conceptualknowledge is organized in the brain by modality (visual,olfactory, motor/functional, etc.) and (2) sensory andfunctional properties are differentially important in iden-tifying members of the living and nonliving categories,respectively. We refer to this class of accounts as thesensory/functional theory (SFT).
The most widely discussed variant of this modality-based theory of semantics is the multiple semanticstheory proposed by Warrington, Shallice, and McCarthy(Warrington & McCarthy, 1983, 1987; Warrington & Sal-lice, 1984), which draws a fundamental distinction be-tween so-called visual and verbal semantics. The othercrucial assumption of the SFT is that the visual proper-ties of concepts are much more important for distin-guishing among members of living than nonliving things.A consequence of these assumptions is that selectivedamage to the visual semantic subsystem should resultin disproportionate impairment for the living category—and, hence, a category-speci�c de�cit. A further expecta-tion derived from this model is that damage to the visualsemantic subsystem should result in disproportionatedif�culty with visual properties of the living things cate-gory.5 A computationally based variant of the SFT (Farah& McClelland, 1991) has con�rmed the basic predictionsderived from the original proposal by Warrington, Shal-lice, and McCarthy.
Two types of evidence have been cited in support ofthe SFT and against a categorical account of category-speci�c de�cits: (1) category-speci�c de�cits supposedlydo not fall out neatly along the boundaries on which theliving/nonliving categories divide, and (2) category-speci�c de�cits are supposedly also modality-speci�c—that is, subjects with selective de�cits for items in theliving things category are supposedly more impaired inprocessing their visual than their functional properties(although they may also be impaired in processing thelatter). We discuss these two types of evidence in turn.
What Are the Categories of Category-Speci�c De�cits?
The four cases of category-speci�c de�cit described inWarrington and Shallice’s (1984) seminal report were allimpaired in processing not only living things but alsofoods. Furthermore, one of the four patients who wastested with a more extensive set of categories (J.B.R.)was also impaired with the categories of musical instru-ments, precious stones, diseases, cloths, and metals. War-rington and Shallice speculated that the co-occurrenceof de�cits to speci�c categories observed in these pa-tients was not accidental but re�ects instead the fact thatthe meanings of the items in the impaired categoriesdepend more heavily on the same damaged semanticsubsystem. That is, the co-occurrence of de�cits for thecategories of animals, foods, plants, and musical instru-ments was assumed to re�ect the fact that the meaningsof the members of these categories are distinguishedprimarily by their visual properties, whereas the mem-bers of the spared categories (e.g., furniture, vehicles,tools, etc.) are distinguished primarily by their functionalattributes.
Although one of the more robust associations of cate-gory-speci�c de�cits is that between the categories ofanimals and foods (De Renzi & Lucchelli, 1994; Sheridan& Humphreys, 1993; Silveri & Gainotti, 1988; Warrington& Shallice, 1984), there are a number of violations to thisempirical generalization: There are patients with cate-gory-speci�c de�cits who are impaired for animals (andfruits and vegetables, in some cases) but not food (Hart& Gordon, 1992; case J.J.: Hillis & Caramazza, 1991; Laia-cona, Barbarotto, & Capitani, 1993) or who are impairedin naming food items but not animals (case P.S.: Hillis &Caramazza, 1991).6 Similarly, the association of de�cits ofliving things and musical instruments, which was ob-served in some of the early reports of category-speci�cde�cit (Silveri & Gainotti, 1988; Warrington & Shallice,1984), has not held up. Thus, for example, the patientreported by De Renzi & Lucchelli (1994), who wasotherwise very similar to those reported by Silveri andGainotti and by Warrington and Shallice, performed verywell with musical instruments. And although there havebeen a number of reports of selective damage or sparingof the category of living things (animals, fruits, and vege-tables; e.g., case P.S.: Hillis & Caramazza, 1991; Laiacona,Barbarotto, & Capitani, 1993), it too dissociates, but alongtheoretically interesting lines: The category of animalscan be damaged or spared independently of the categoryof fruits, vegetables, and plants (Hillis & Caramazza, 1991;Hart & Gordon, 1992), and the category of fruits andvegetables (or plants) can be damaged selectively (Farah& Wallace, 1992; Hart, Berndt, & Caramazza, 1985) fromother living and nonliving categories.
It is far from obvious that the SFT can make sense ofthe attested patterns of associations and dissociations ofcategory-speci�c de�cits. The �rst problem concerns
4 Journal of Cognitive Neuroscience Volume 10, Number 1

whether it is reasonable to assume that the observedassociations of category-speci�c de�cits re�ect the or-ganizing principle of modality-speci�c semantics. Thus,what underlying sensory/functional principles wouldgroup together the categories of animals, foods, andmusical instruments versus the categories of furniture,tools, and body parts? The mere assertion that the mean-ings of the items in the �rst set are more dependent onvisual or other sensory attributes and those of the sec-ond set are more dependent on functional attributes isnot particularly convincing. Consider, for example, thecontrast among carrot, celery, apple, orange, avocado,tomato, hamburger, scrambled eggs, pasta, beer, and wine.These items are clearly visually different from one an-other, but they are also functionally different. One mightgo so far as to contend that what is important fordiscriminating among them is not so much their visualshape but their functions: used to make juice, used fordessert, eaten at breakfast, Italian food, fast food, eatenraw, eaten cooked, used for minestrone, and so on. Thesame argument can be made for musical instrumentswith respect to the importance of their functional prop-erties. And certainly visual shape is as important fordistinguishing hands, heads, beds, stools, hammers, andscrewdrivers as is their function. Nonetheless, the claimthat the relative “weighting” of the sensory and func-tional properties for discriminating among members ofa semantic category provides the binding factor for someof the observed associations of category-speci�c de�citshas never been developed in enough detail to allowserious consideration of its merits.7
Far more damaging to the SFT is the pattern of disso-ciations of category-speci�c de�cits. Since part of themotivation for the SFT is the claim that certain semanticcategories tend to be damaged together because of theircommon dependence on a modality-speci�c semanticsubsystem, the observation that these same categoriescan be damaged independently of each other poses aserious threat to the theory. That is, in the measure towhich one is committed to explaining the observedco-occurrence of impairment of, for example, the cate-gories of animals and of fruits and vegetables as re�ect-ing their shared dependence on the hypothesized visualsemantic subsystem, one is thereby committed to theirnecessary co-occurrence, and their dissociation wouldconstitute evidence against the hypothesized semanticsubsystem.
Warrington and McCarthy (1987) were keenly awareof this danger to the SFT and tried to deal with it byhypothesizing further subdivisions within the visual se-mantic subsystem. They argued that the dissociation ofde�cits of the categories of animals and of fruits andvegetables may be explained by assuming that someperceptual features are more important for animals thanfor fruits and vegetables, and vice versa for some otherperceptual features. They speculated that shape may be
more important for the categories of �owers and animalsand that color may be more important for the categoryof fruits and vegetables. And, presumably, in order toexplain the other patterns of dissociations of category-speci�c de�cits (e.g., animals from �owers; animals frommusical instruments) further subdivisions would have tobe speci�ed within the visual and functional semanticsubsystems. However, no independent evidence hasbeen provided for the claim that distinction amongmembers of different semantic categories depend differ-entially on color, shape, or other sensory features. Norhas it been shown that impairment of particular seman-tic categories is associated with special dif�culties forspeci�c types of sensory information (e.g., greater dif�-culty processing color in patients with selective damageto the categories of fruits and vegetables). Furthermore,it should be noted that the more subdivisions one pro-poses, speci�cally in order to accommodate dissocia-tions of particular categories, the more this proposalbecomes indistinguishable from a categorical account ofcategory-speci�c de�cits.
The evidence and arguments presented in this sectionhave shown that one of the pillars of the SFT does nothold. Contrary to the early reports of category-speci�cde�cits that stressed the co-occurrence of conceptuallydisparate combinations of categories (e.g., animals, foods,and musical instruments), there are a number of well-documented cases of dissociations of these categories.And, as we have seen, there appears to be no clearlymotivated basis for the argument that attested patternsof associations and dissociations of semantic categoryde�cits are readily explicable in terms of selective dam-age to modality-speci�c semantic subsystems. These criti-cisms of the SFT undermine its supposed superiorityover categorical accounts in explaining the occurrenceof category-speci�c de�cits. We turn now to the othermajor argument for the SFT—the claim that patientswith selective damage to the living category are dispro-portionately impaired in processing their visual proper-ties.
Is Knowledge of Visual Properties More SeverelyImpaired Than Knowledge of Functional Propertiesin Individuals with Category-Speci�c De�cits forLiving Things?
The claim that semantic category-speci�c de�cits re�ectselective damage to the visual or to the functional8
semantic subsystems predicts that subjects with selec-tive damage to the living or the nonliving categoriesshould show disproportionate dif�culty with visual orfunctional properties, respectively. This expectation isderived not only from Warrington, Shallice, andMcCarthy’s theory (Warrington & McCarthy, 1983, 1987;Warrington & Shallice, 1984) of the semantic system butalso from the computational variant proposed by Farah
Caramazza and Shelton 5

and McClelland (1991). The results of several studieshave been interpreted as having con�rmed this predic-tion. That is, there are reports of subjects with dispropor-tionate impairment of the category of living things whoare also supposedly disproportionately impaired in proc-essing the visual attributes of living things.
The �rst studies that explicitly addressed the possibil-ity of a co-occurrence of category- and modality-speci�csemantic de�cits are those of Basso et al. (1988), Sartoriand Job (1988),9 and Silveri and Gainotti (1988). In allthree studies, it was reported that subjects with dispro-portionate dif�culty with living things also had dispro-portionate dif�culty responding to questions about thevisual attributes of living objects. For example, Basso etal. report that their subject (N.V.) correctly answered 25of 29 questions about the functional attributes of livingthings (e.g., Does it live in Italy or in the desert?) butonly 10 of 20 questions about their sensory attributes(e.g., Does it have a smooth back or is it humpbacked?).Similarly, subject L.A. (Silveri & Gainotti, 1988) was onlyable to name 1 of 11 animals in response to de�nitionsthat stressed visual attributes (e.g., a black and whitestriped wild horse) but could name 8 of 14 animals inresponse to de�nitions that stressed functional attributes(e.g., the king of the jungle).
Later studies also purportedly showed an associationof category- and modality-speci�c semantic de�cits (DeRenzi & Lucchelli, 1994; Farah et al., 1989; Gainotti &Silveri, 1996; Hart & Gordon, 1992). Thus, for example,Farah et al. report that their subject L.H. was dispropor-tionately impaired in answering questions about visualattributes of living things (e.g., Do ducks have long ears?vs. Is peacock served in French restaurants?). And DeRenzi and Lucchelli report that their subject Felicia wassigni�cantly worse at answering questions about physi-cal than “functional-encyclopedic” attributes of livingthings but that she showed no such difference for non-living things.
Despite the seeming convergence of results in favorof the possibility that category-speci�c de�cits are alsomodality-speci�c, there are reasons for skepticism ininterpreting these results. First, there are issues concern-ing the validity of the reported results in each of thestudies cited; second, there is growing evidence thatcategory-speci�c de�cits are not associated with dispro-portionate dif�culties in processing either the visual orthe functional attributes of objects.
Interpretation of some of the studies purporting toshow that patients with category-speci�c de�cits forliving things are also disproportionately impaired inprocessing these objects’ visual attributes is underminedby the fact that these studies failed to control the relativedif�culty of visual and functional attribute judgments.Stewart et al. (1992; Exp. 5) have shown that judgmentsabout the visual properties of living things are moredif�cult to evaluate than judgments about functionalproperties. They have also shown that the disproportion-
ate dif�culty for visual attributes for living things in apatient with a putative category-speci�c de�cit disap-pears when visual and functional judgments are matchedfor dif�culty (Exp. 6). In light of these results, it is notpossible to meaningfully interpret some of the earlystudies claiming to show a co-occurrence of category-and modality-speci�c de�cits, where no attempt wasmade to control the relative dif�culty of visual and func-tional attribute judgments (e.g., Basso et al., 1988; Sartori& Job, 1988; Silveri & Gainotti, 198810).
A related dif�culty is encountered by De Renzi andLucchelli’s (1994) study. Although the authors were ableto show that their subject, Felicia, was more severelyimpaired in processing visual attributes of living thannonliving category items, she was also impaired in proc-essing functional attributes of living things, as indicated,for example, by her performance in deciding whether ananimal was native of Italy (only 73% correct). Further-more, since no attempt was made to equate visual andfunctional judgments for dif�culty, it is not possible todirectly compare relative performance for the two typesof judgments.
Farah et al. (1989) argued that their subject, L.H.,presented with a category-speci�c de�cit for the livingcategory that was also modality-speci�c for visual prop-erties. However, inspection of their data (Table 1, Farahet al.) shows that normal subjects were also dispropor-tionately poor in answering questions about the visualproperties of the living category. Thus, no meaningfulconclusion concerning category-speci�c de�cits is pos-sible from this study.
Recently, Gainotti and Silveri (1996) retested theirsubject, L.A., with new materials designed to be free ofthe methodological problems encountered by their ear-lier report. They again showed that L.A. performed dis-proportionately poorly on tasks involving the processingof visual attributes of living things. For example, in a taskrequiring the identi�cation of the visual feature onwhich two objects differed, they found that L.A. per-formed worse with animals (15% correct) than artifacts(50% correct). However, the two categories were alsosigni�cantly different in familiarity (on a scale of 1 to 5,2.55 and 3.96 for animals and artifacts, respectively). Andin a color feature naming task in which L.A. again per-formed more poorly with living than nonliving things,there, too, the two categories differed in familiarity. Thus,the interpretation of L.A.’s performance regarding theputative association of category- and modality-speci�cde�cits remains problematic at best.
Finally, Hart and Gordon (1992) have also argued fora modality-speci�c organization of the semantic systemon the basis of the performance of their subject, K.R.,who presented with a category-speci�c de�cit restrictedto animals. K.R. performed very poorly in naming ani-mals (18 of 61) despite being perfectly normal in namingother objects (348 of 350). The striking feature of thissubject was that she could answer correctly all ques-
6 Journal of Cognitive Neuroscience Volume 10, Number 1

tions about functional attributes of animals, as well as allquestions about both visual and functional attributes ofinanimate objects but was quite poor in answering ques-tions about the visual attributes of animals. Thus, thiscase would seem to represent the perfect dissociation insupport of the SFT.11 However, there are aspects of thissubject’s performance that are problematic for the SFT,and there are other features of her performance that mayundermine its usefulness altogether.
The fact that K.R. was impaired in processing thevisual properties of only animals is inconsistent with theSFT. The theory predicts that damage to visual semanticswould differentially affect the living and nonliving cate-gories. It does not predict that it would lead to damageto only the category of animals. At the very least itpredicts impairment of other living things (e.g., fruits)besides animals, as well as less-marked impairment of thenonliving category. None of these conditions were metby the results of this study. Equally problematic for theSFT is the fact that K.R.’s de�cit in processing visualproperties of animals was restricted to language inputs.She only performed poorly with visual attributes whenshe was required to respond to verbal inputs or torespond verbally. Thus, for example, she performed per-fectly in distinguishing between appropriately and inap-propriately colored animals but could not perform thesame discrimination when the color alternatives weregiven verbally. This result is also highly problematic forthe SFT. The theory predicts equal dif�culties (barringadditional sensory or language de�cits) for visual andverbal tasks because damage to the visual semantic sub-system would presumably lead to impairments in alltasks involving the use of this system. Thus, K.R.’s selec-tive dif�culty for visual properties restricted to the ver-bal domain is uninterpretable within the SFT.
Perhaps, most problematic about the Hart and Gordon(1992) study is their subject’s performance in a forced-choice property judgment task. In this task K.R. wasrequired to choose between two visual or two nonphysi-cal properties of animals (e.g., Is an elephant orange orgray? or Is an elephant a food or an animal?). The prop-erties were chosen in such a way that they includedproperties that she had produced correctly and incor-rectly in a previous direct recall task (e.g., What’s thecolor of lemon? Yellow; What’s the color of an elephant?Orange). In this task she virtually always (186 of 187)picked the choice corresponding to the property shehad produced in the previously administered propertynaming task, whether correct or incorrect. Thus, forexample, consistent with her response in the propertyproduction task, she chose orange when given thechoice, Is an elephant orange or gray? She chose theincorrect, strange response in 54 out of 55 trials. Theunexpected systematicity of K.R.’s strange errors raisesquestions about the usefulness of this study for inform-ing theories of normal language because it appears thatshe may have come to have systematic “strange beliefs”
about some animals.12 Nevertheless, the most that canbe concluded for the SFT from the Hart and Gordonstudy is that those aspects of K.R.’s performance that arenot strange are not explicable within the theory.
This review of the experimental evidence cited insupport of the claim that subjects with a category-speci�c de�cit for living things are disproportionatelyimpaired in processing the visual features of conceptshas shown it to be inadequate for the stated purpose. Ineach case (with the exception of Hart and Gordon, 1992,but there are other problems with this study), the failureto adequately control for differential levels of dif�cultyin processing visual versus functional properties of con-cepts undermines the possibility of drawing clear infer-ences from the results.13 Thus, there is no compellingempirical evidence in favor of the claim that category-speci�c de�cits are also necessarily modality-speci�cde�cits. Furthermore, there is growing evidence thatsubjects with category-speci�c de�cits for living thingsare equally impaired in processing visual and functionalattributes of objects—evidence that is directly in con�ictwith the SFT.
Laiacona, Barbarotto, & Capitani (1993) and Laiaconaet al. (1997) have provided the clearest evidence to datefor equal levels of impairment for visual and functionalattributes in cases of category-speci�c de�cit for livingthings. In the earlier of the two studies, they reportedtwo trauma cases, F.M. and G.R., with disproportionateimpairment in processing living things (F.M.: 20% vs. 70%correct naming of living and nonliving things, respec-tively; G.R.: 13% vs. 73% correct naming of living andnonliving things, respectively). When these subjectswere tested in a forced-choice task with strictly matchedquestions about visual or functional properties of ob-jects (Laiacona, Barbarotto, Trivelli, & Capitani, 1993),they performed equally poorly with the two types ofattributes. Thus, F.M. responded correctly 73% and 69%of the time to questions about visual and functionalproperties of living things, respectively, and he re-sponded correctly 96% of the time to both visual andfunctional properties of nonliving things. Similarly, G.R.responded correctly 55% and 58% of the time to ques-tions about visual and functional properties of livingthings, respectively, and he responded correctly 91% ofthe time to visual properties and 84% of the time tofunctional properties of nonliving things. In a more re-cent study, Laiacona et al. (1997) again observed equallevels of impairment for knowledge of visual and func-tional attributes in two new cases of category-speci�cde�cit for living things.14
Two other studies (Funnell & De Mornay Davies, 1997;Sheridan & Humphreys, 1993) further con�rm this pat-tern of results. Sheridan and Humphreys reported a sub-ject with a category-speci�c de�cit for living things whowas equally impaired with visual and functional attributejudgments (14 of 20 and 13 of 20 for visual and func-tional attributes of animals, respectively). And in a new
Caramazza and Shelton 7

analysis of J.B.R., one of the cases of category-speci�cde�cit for living things originally reported by Warringtonand Shallice (1984), Funnell and De Mornay Davies(1997) found that he is equally impaired in verifyingvisual and functional attribute questions about livingthings.15 These results indicate that knowledge of visualattributes of living things is not necessarily dispropor-tionately impaired (relative to knowledge of functionalattributes) in subjects with a category-speci�c de�cit forliving things. Thus, the second of the two major predic-tions of the SFT is not con�rmed.
The Organized Unitary Content Hypothesis(OUCH)
Another class of reductionist hypotheses rejects the mo-dality-speci�c16 claim of semantic organization proposedby the SFT but also explains category-speci�c de�cits asresulting from noncategorical properties of semanticrepresentations. The OUCH is one such account(Caramazza, Hillis, Rapp, & Romani, 1990; Hillis, Rapp, &Caramazza, 1995; Rapp, Hillis, & Caramazza, 1993; seeRiddoch, Humphreys, Coltheart, & Funnell, 1988, for arelated proposal). This account is based on two funda-mental characteristics of natural kind objects and arti-facts: (1) the properties that de�ne an object (e.g., dog,chair, etc.) are highly intercorrelated, and (2) membersof a superordinate category share many features in com-mon (S. Gelman & Coley, 1990; Keil, 1989; Markman,1989; Rosch, 1973, 1975; Rosch, Mervis, Gray, Johnson, &Boyes-Braem, 1976). Thus, for example, objects that arecapable of a certain type of bio-mechanical motion (likea dog) tend to have particular types of shapes and to bemade of certain kinds of “stuff” (texture, color, odors,etc.), and objects that have a particular shape (like achair) and an inability to move tend to be made ofdifferent kinds of “stuff.” Furthermore, these bundles ofintercorrelated properties are differentially distributed inthe categories of living and nonliving things. A conse-quence of the latter property of concepts is that themultidimensional space of semantic properties is nothomogeneously occupied but, instead, is “lumpy”—thereare regions that are densely packed and others that areonly sparsely occupied.17 The denser regions representconcept domains characterized by highly correlatedproperties, with the densest regions most likely corre-sponding to natural kind concepts; however, some arti-facts (e.g., tools) can also occupy relatively denseregions, because they, too, may share many correlatedproperties (e.g., shape and function).
One implication of the inhomogeneous nature of theorganization of semantic attributes in the brain is thatfocal damage to the semantic system can result in cate-gory-speci�c de�cits. This expectation is based on thefollowing reasoning: Focal damage to a region of seman-tic space will result in an impairment of those categorieswhose members’ meaning depends on the affected se-
mantic properties, and because of the lumpiness of se-mantic space, the impairment is likely to be dispropor-tionate for one category or another, re�ecting whichdense region happened to have been damaged. Twoother implications follow from the OUCH model. One isthat semantic categories with highly correlated proper-ties (i.e., natural kinds) are more likely to be damaged asa category. This expectation follows from the assumptionthat categories with highly correlated properties occupydense regions in semantic space, thereby allowing forthe possibility of selective damage to the category. Fromthis it follows that natural kinds are more likely to �gurein category-speci�c de�cits. The other prediction thatfollows from OUCH is that various different patterns ofcategory-speci�c de�cits are expected to occur, re�ect-ing the vagaries of the distribution of damage to thesemantic space in different cases of brain damage. Thatis, because in this view the semantic system consists ofa seamless, but lumpy space, the particular categoriesthat are affected will depend on which speci�c regionsof the brain are damaged. Thus, for example, OUCH doesnot exclude the possibility of a subject with dispropor-tionate impairment to the category of living things,foods, musical instruments, and precious stones (as wasthe case for J.B.R., reported by Warrington & Shallice,1984).
The other major prediction made by OUCH is thatselective impairment of speci�c semantic categoriesshould not be associated with disproportionate de�citto either visual or functional attributes of objects. Thisexpectation is based on the fact that focal damage to thesemantic system should affect closely correlated proper-ties that are likely to involve both visual and functionalattributes. This prediction clearly distinguishes theOUCH model from the SFT because, as we have seen,the latter theory of conceptual organization predicts theopposite result—namely, that category-speci�c de�citsfor living things should be associated with a dispropor-tionate de�cit for the visual attributes of objects.
The patterns of category-speci�c de�cits reviewedhere are consistent with expectations derived fromOUCH. The model predicts the occurrence of category-speci�c de�cits without postulating a categorical organi-zation of semantic knowledge. It also allows for theexistence of various combinations of category-speci�cde�cits. And, �nally, it is consistent with the resultsshowing that category-speci�c de�cits are not necessar-ily also modality-speci�c in nature (contrary to the pre-diction derived from the SFT). Thus, OUCH offers abetter account than the SFT for the various empiricalphenomena related to category-speci�c de�cits.
Although OUCH can account for important aspects ofthe category-speci�c de�cit phenomenon, it is unsatis-factory in at least one respect: It is too vague on criticalissues about the nature and organization of semanticinformation, allowing it to account for almost any pat-tern of category-speci�c de�cits. All it needs to do for
8 Journal of Cognitive Neuroscience Volume 10, Number 1

the latter purpose is to assume, for example, that braindamage has accidentally affected one of the lumpy re-gions of semantic space, thereby resulting in a category-speci�c de�cit. Although this kind of explanatory powerwould be welcome in the context of an explicitly articu-lated theory of semantics, in the present case it merelyre�ects the unconstrained nature of the model’s centralassumptions. Furthermore, the model fails to provide aprincipled account for the patterns of attested forms ofcategory-speci�c de�cits. Speci�cally, the OUCH (justlike the SFT) fails to explain why it is that by far the mostprevalent form of category-speci�c de�cit involves thecategory of living things.
Categorical Organization of Knowledge in theBrain: A Speculative Hypothesis
The reservations we have voiced about the reductionistaccounts of category-speci�c de�cits (the SFT andOUCH models) encourage us to explore the alternativehypothesis that semantic knowledge may be organizedcategorically in the brain (Warrington, 1981). The hy-pothesis we wish to entertain is that evolutionary pres-sures have resulted in specialized mechanisms forperceptually and conceptually distinguishing animateand inanimate kinds (R. Gelman, 1990; Premack, 1990),leading to a categorical organization of this knowledgein the brain. We will call this hypothesis the domain-speci�c knowledge hypothesis. In the “Discussion” sec-tion, we will elaborate further on this view and offervarious arguments in its defense. Here we simply notethat the hypothesis receives initial plausibility from de-velopmental research, which has shown that infants asyoung as 3 months old are able to distinguish animatefrom inanimate objects, as indicated, for example, bytheir ability to distinguish biological from nonbiologicalmotion (Bertenthal, 1993; Bertenthal, Prof�tt, & Cutting,1984) and their ability to distinguish caused from non-caused motion (Leslie, 1982, 1988; and see Leslie &Keeble, 1987, for results on 6-month-old infants) and thatthe 8- to 10-month-old infant’s ability to distinguish ani-mals from nonanimals is unlikely to be merely perceptu-ally based (Quinn & Eimas, 1996) but is most likelyconceptually driven (Mandler, 1992). Thus, for exam-ple, using a dishabituation paradigm, Mandler andMcDonough (1993) have shown that 9-month-old infantsdishabituate to an animal after repeatedly seeing modelsof a vehicle (and vice versa) even though the dishabitu-ating stimulus may be highly visually similar to the ha-bituated objects (e.g., birds and airplanes). Theseobservations further encourage us to explore the possi-bility that a fundamental distinction in human knowl-edge—that between knowledge of animate andinanimate things—may be rooted in speci�c evolution-ary adaptations.18
The domain-speci�c knowledge hypothesis makes thefollowing two predictions about the nature of category-
speci�c de�cits. First, the only “true” category-speci�cde�cits are those involving categories of knowledge forwhich evolutionary pressures have led to the develop-ment of specialized neural mechanisms for their percep-tual and conceptual distinction. Plausible categoricaldistinctions are those among animals, plant life, and arti-facts.19 The other major expectation derived from thishypothesis is that (everything else being equal) category-speci�c de�cits should result in comparable impair-ments for the visual and functional attributes of aconcept. This prediction re�ects the assumption that thecategory-speci�c nature of the de�cit results from thefact that brain damage has affected a categorically organ-ized knowledge system that represents all types of infor-mation relevant to that semantic domain.20
The evidence on category-speci�c de�cits that wehave reviewed in the preceding sections is consistentwith the expectations derived from the hypothesis thatsemantic knowledge is organized categorically in thebrain, although it is also consistent with the OUCHmodel. Here we provide further experimental evidencefor the hypothesis that semantic knowledge is organizedcategorically. We present a case study of a brain-damagedsubject, E.W., who has a selective de�cit for the categoryof animals and who is equally impaired for the visual andfunctional attributes of the members of this category.This pattern of performance—which is predicted by thedomain-speci�c knowledge account—has not been re-ported previously. The results are organized as follows.First we show that subject E.W. is disproportionatelyimpaired in naming animals relative to other living thingsand artifacts. We then show that this de�cit is not merelydue to a lexical retrieval problem because E.W. is alsoseverely impaired in visually recognizing animals and inrecognizing them from their characteristic sounds. Third,we show that E.W.’s de�cit occurs at the conceptuallevel because she is not only impaired in processingpictures of animals but also in answering verbal ques-tions about them. In this section we also show that herdisproportionate semantic de�cit for animals is equallysevere for the visual and functional properties of mem-bers of this category.
RESULTS
Does E.W. Have a Category-Speci�c NamingDe�cit?
Naming the Snodgrass and Vanderwart Pictures
The �rst analysis of E.W.’s performance is directed atestablishing that E.W. has a category-speci�c namingde�cit. Naming performance is summarized in Table 1. Aresponse was considered correct if E.W. provided theSnodgrass and Vanderwart (1980) name or any accept-able response produced by the control subjects (n = 5).For both the control subjects and E.W., performance isbroken down by animacy and familiarity. Familiarity was
Caramazza and Shelton 9
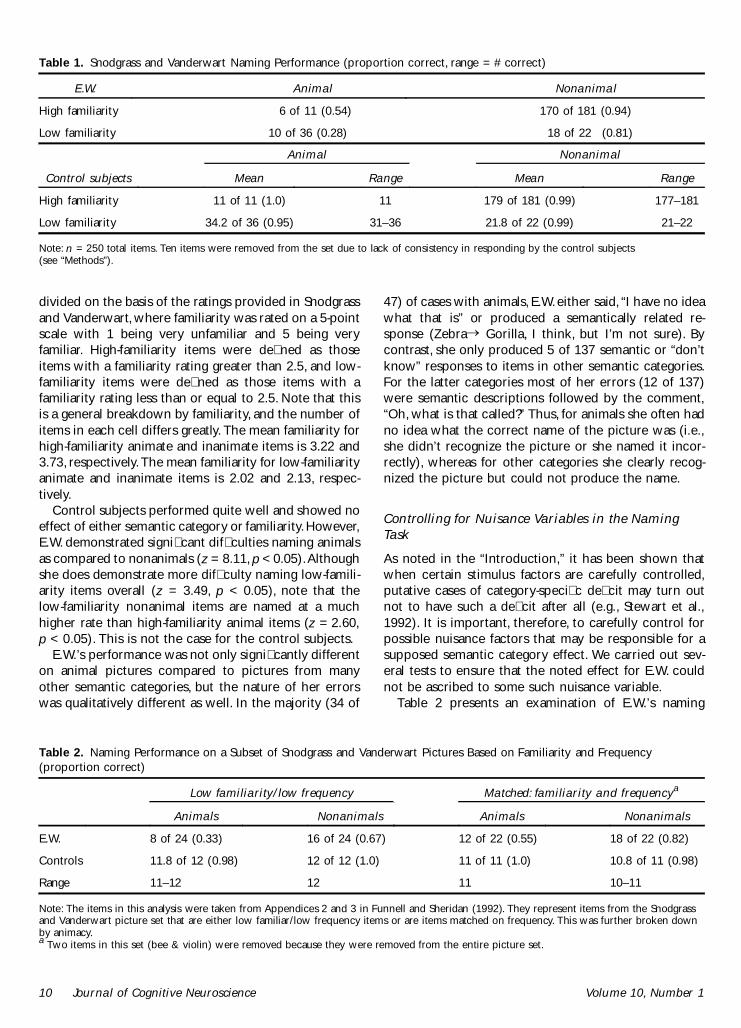
divided on the basis of the ratings provided in Snodgrassand Vanderwart, where familiarity was rated on a 5-pointscale with 1 being very unfamiliar and 5 being veryfamiliar. High-familiarity items were de�ned as thoseitems with a familiarity rating greater than 2.5, and low-familiarity items were de�ned as those items with afamiliarity rating less than or equal to 2.5. Note that thisis a general breakdown by familiarity, and the number ofitems in each cell differs greatly. The mean familiarity forhigh-familiarity animate and inanimate items is 3.22 and3.73, respectively. The mean familiarity for low-familiarityanimate and inanimate items is 2.02 and 2.13, respec-tively.
Control subjects performed quite well and showed noeffect of either semantic category or familiarity. However,E.W. demonstrated signi�cant dif�culties naming animalsas compared to nonanimals (z = 8.11, p < 0.05). Althoughshe does demonstrate more dif�culty naming low-famili-arity items overall (z = 3.49, p < 0.05), note that thelow-familiarity nonanimal items are named at a muchhigher rate than high-familiarity animal items (z = 2.60,p < 0.05). This is not the case for the control subjects.
E.W.’s performance was not only signi�cantly differenton animal pictures compared to pictures from manyother semantic categories, but the nature of her errorswas qualitatively different as well. In the majority (34 of
47) of cases with animals, E.W. either said, “I have no ideawhat that is” or produced a semantically related re-sponse (Zebra ® Gorilla, I think, but I’m not sure). Bycontrast, she only produced 5 of 137 semantic or “don’tknow” responses to items in other semantic categories.For the latter categories most of her errors (12 of 137)were semantic descriptions followed by the comment,“Oh, what is that called?” Thus, for animals she often hadno idea what the correct name of the picture was (i.e.,she didn’t recognize the picture or she named it incor-rectly), whereas for other categories she clearly recog-nized the picture but could not produce the name.
Controlling for Nuisance Variables in the NamingTask
As noted in the “Introduction,” it has been shown thatwhen certain stimulus factors are carefully controlled,putative cases of category-speci�c de�cit may turn outnot to have such a de�cit after all (e.g., Stewart et al.,1992). It is important, therefore, to carefully control forpossible nuisance factors that may be responsible for asupposed semantic category effect. We carried out sev-eral tests to ensure that the noted effect for E.W. couldnot be ascribed to some such nuisance variable.
Table 2 presents an examination of E.W.’s naming
Table 1. Snodgrass and Vanderwart Naming Performance (proportion correct, range = # correct)
E.W. Animal Nonanimal
High familiarity 6 of 11 (0.54) 170 of 181 (0.94)
Low familiarity 10 of 36 (0.28) 18 of 22 (0.81)
Animal Nonanimal
Control subjects Mean Range Mean Range
High familiarity 11 of 11 (1.0) 11 179 of 181 (0.99) 177–181
Low familiarity 34.2 of 36 (0.95) 31–36 21.8 of 22 (0.99) 21–22
Note: n = 250 total items. Ten items were removed from the set due to lack of consistency in responding by the control subjects(see “Methods”).
Table 2. Naming Performance on a Subset of Snodgrass and Vanderwart Pictures Based on Familiarity and Frequency(proportion correct)
Low familiarity/low frequency Matched: familiarity and frequencya
Animals Nonanimals Animals Nonanimals
E.W. 8 of 24 (0.33) 16 of 24 (0.67) 12 of 22 (0.55) 18 of 22 (0.82)
Controls 11.8 of 12 (0.98) 12 of 12 (1.0) 11 of 11 (1.0) 10.8 of 11 (0.98)
Range 11–12 12 11 10–11
Note: The items in this analysis were taken from Appendices 2 and 3 in Funnell and Sheridan (1992). They represent items from the Snodgrassand Vanderwart picture set that are either low familiar/low frequency items or are items matched on frequency. This was further broken downby animacy.a Two items in this set (bee & violin) were removed because they were removed from the entire picture set.
10 Journal of Cognitive Neuroscience Volume 10, Number 1

performance on the items included in Funnell and Sheri-dan’s (1992) tests of naming performance as a functionof animacy and familiarity (from their Appendixes 2 and3). These items are all taken from the Snodgrass andVanderwart picture set. The items in the �rst set are alllow-familiarity/low-frequency items, and the items in thesecond set are matched on familiarity and frequency(i.e., represent a range of familiarity and frequency).Since the number of items is quite low, we have includedE.W.’s naming performance from two administrations ofthe naming task. Control subjects only named theSnodgrass and Vanderwart picture set once.
It is immediately apparent that E.W.’s naming is af-fected by the animacy variable for both sets of items(z = 2.92, p < 0.05; z = 2.85, p < 0.05, respectively), evenwhen those items have been controlled for familiarity.Thus, taken together with the above analyses, E.W.’sde�cit in naming animals can be attributed to a de�citaffecting the category of animals and not an artifact dueto familiarity of the items.
However, Stewart et al. (1992; see also Gaffan & Hey-wood, 1993) argued that naming could also be in�u-enced by visual complexity. Table 3 contains E.W.’snaming performance on a subset of items from theSnodgrass and Vanderwart picture set that are matchedon visual complexity (n = 36) and a subset of items thatare matched (as closely as possible) on both visual com-plexity and familiarity. Once again, it is obvious that
neither the factor of visual complexity nor the combinedfactor of visual complexity and familiarity is responsiblefor E.W.’s decrement in performance for animate items.Instead, E.W.’s picture-naming performance is in�uencedstrongly by the factor of animacy and cannot be ex-plained by extraneous factors such as familiarity or visualcomplexity (z = 5.75, p < 0.05; z = 3.17, p < 0.05,respectively).
The Naming De�cit Is Restricted to Animals and IsNot General to Living Things
A crucial part of this investigation involves determiningthe boundaries of the category-speci�c de�cit. Spe-ci�cally, we are concerned with determining whetherE.W. is disproportionately impaired in processing livingthings or the more restricted category of animals. Forthis purpose we considered E.W.’s performance for spe-ci�c semantic categories.
Table 4 provides a breakdown of E.W.’s naming per-formance according to speci�c semantic categories. Sev-eral things should be noted. First, E.W.’s namingperformance is not affected by the category of livingthings but rather by the category of animate things.Thus, like several other reported cases (Farah & Wallace,1992; Hart et al., 1985; Hillis & Caramazza, 1991), E.W.’sde�cit respects a more �ne-grained distinction than liv-ing/nonliving. Although performance broken down by
Table 3. Naming Performance on a Subset of Snodgrass and Vanderwart Pictures Based on Visual Complexity and VisualComplexity/Familiarity (proportion correct)
Matched: Complexity Matched: Complexity and familiarity
Animals Nonanimals Animals Nonanimals
E.W. 10 of 36 (0.28) 32 of 36 (0.89) 7 of 17 (0.41) 16 of 17 (0.94)
Controls 35.4 of 36 (0.98) 35.4 of 36 (0.98) 16.6 of 17 (0.98) 16 of 17 (0.94)
Range 35–36 35–36 16–17 16–17
Note: The items in this analysis represent a subset of items from the Snodgrass and Vanderwart picture set that are matched on visual complex-ity (mean = 3.76, SD = 0.51) or on visual complexity (mean = 3.83, SD = 0.44) and familiarity (animate: mean = 2.78, SD = 0.84; inanimate:mean = 2.64, SD = 0.84). This was further broken down by animacy.
Table 4. E.W.’s Naming Performance as a Function of Speci�c Semantic Category (proportion correct)
Category Performance Category Performance
Animal 16 of 47 (0.34) Kitchenware 16 of 17 (0.94)
Body part 11 of 12 (0.92) Musical instrument 8 of 10 (0.80)
Clothing 27 of 27 (1.0) Tool 19 of 23 (0.83)
Fruit 12 of 12 (1.0) Vegetable 12 of 12 (1.0)
Furniture 15 of 15 (1.0) Vehicle 13 of 14 (0.93)
Other 53 of 61 (0.87)
Caramazza and Shelton 11

semantic category in this set of items must be inter-preted with caution because the items in each of thecategories are not controlled for potential nuisance vari-ables (i.e, familiarity, frequency, and visual complexity), itis clear from Table 4 that E.W. is not impaired in namingfruits and vegetables: Her performance for fruits andvegetables was perfect (100%) and strikingly differentfrom that for animals (34%).
E.W.’s Naming De�cit Is Not Restricted to VisualStimuli: Sound Identi�cation
In order to determine whether E.W.’s naming de�citmight not re�ect a category-speci�c visual agnosia, shewas tested with a sound identi�cation task in which theanimal and nonanimal sounds were matched as well aspossible on dif�culty level as determined by the perfor-mance of several control subjects (see “Methods”). E.W.was quite poor at identifying animal sounds (8 of 32 =0.25) as compared to nonanimal sounds (20 of 32 =0.63), a difference that was signi�cant (z = 3.06, p <0.05). These results suggest that E.W. does poorly innaming animals across modalities (i.e., the de�cit is notrestricted to identifying and naming animals from pic-tures).
Summary
The results we have reported in this section show thatE.W. has a semantic category-speci�c naming de�cit. Thede�cit is restricted to the category of animals and not tothe category of living things, as shown by the fact thatnaming of fruits and vegetables was virtually intact.E.W.’s poor naming performance for animals cannot bemerely ascribed to nuisance variables such as familiarityand visual complexity since her disproportionate dif�-culties with animals persisted when these variables werestrictly controlled. And, �nally, E.W.’s naming de�cit doesnot appear to be merely the result of a category-speci�cvisual agnosia because she also showed disproportion-ately poor performance for animals in a sound identi�ca-tion task.
Does E.W. Have a Category-Speci�c ObjectRecognition De�cit?
Sartori and Job (1988; see also De Renzi & Lucchelli,1994) reported that their subject Michelangelo, whoshowed a category-speci�c de�cit for living things, wasunable to distinguish between real and unreal animalsand to decide which of four disembodied parts of ananimal went with a body with a missing part. This typeof performance has been interpreted as indicating aselective de�cit for the visual properties of objects. How-ever, not all cases of category-speci�c de�cit are im-paired in object decision tasks. There are several reports
of subjects with category-speci�c de�cits for livingthings who perform within normal limits in object deci-sion tasks (e.g., Laiacona et al., 1997; Sheridan & Hum-phreys, 1993). Thus, impairment in visual object decisionperformance is not a necessary feature of category-speci�c de�cit for living things. It remains to be deter-mined what implications follow from the association/dis-sociation of impairments at the level of objectidenti�cation and naming. We will return to this issue inthe “Discussion.” Here we document that E.W. has asevere category-speci�c de�cit in recognizing animals.
Object Decision Task
In this task, E.W. and two control subjects were asked todetermine whether or not each picture represented areal object—either an animal or some type of artifact(see Figure 1). Performance for all subjects is summa-rized in Table 5. Normal control subjects performed fairlywell on this task. E.W. had signi�cant problems determin-ing which pictures represented real animals (and therewas a slight tendency to respond Yes in this task). Spe-ci�cally, she was barely above chance level (60% correct)in distinguishing between real and unreal animals. Shealso had a few dif�culties distinguishing between realand unreal artifacts (i.e., was willing to accept someunreal objects as real), but this is a problem shared bythe control subjects and probably re�ects the dif�cultyof the “unreal” inanimate objects. The important point isthat E.W. performs poorly, and well outside the normalrange, differentiating real and unreal animals but per-forms within normal range differentiating real and unrealobjects.21
Part Decision Task
In this task, E.W. and two control subjects were asked todecide which of two “heads” went with a headless body(see Figure 2). E.W. was severely impaired at selectingthe correct head for animals but not for artifacts (60%vs. 97%; z = 3.45, p < 0.05), and her performance with
Table 5. Object Decision Task (Hit = Real)
Subject Hit Correct rejection
E.W.:
Animals 21 of 30 (0.70) 15 of 30 (0.50)
Nonanimals 30 of 30 (1.0) 25 of 30 (0.83)
Controls:
Animals 29 of 30 (0.97) 25 of 30 (0.83)
Nonanimals 28.5 of 30 (0.95) 22 of 30 (0.73)
12 Journal of Cognitive Neuroscience Volume 10, Number 1

animals is not signi�cantly different than chance (z < 1).Normal controls had no dif�culty with either category(100% and 97% correct for animals and artifacts, respec-tively).
Summary
The results of the Object and Part Decision Tasks areclear: E.W. is severely impaired in discriminating betweenreal and unreal animals but not between real and unrealartifacts. She is at chance in recognizing animals, but herperformance with artifacts was indistinguishable fromthat of control subjects. This pattern of performancerules out the hypothesis that the locus of E.W.’s cate-gory-speci�c de�cit is at the stage of name retrieval.
E.W.’s Category-Speci�c Object RecognitionDe�cit Is Not the Result of Impaired VisualProcessing of Complex Objects.
The conclusion reached in the previous section regard-ing the locus of functional de�cit in E.W.’s category-speci�c impairment may be challenged. It could be ar-gued that the putative category-speci�c object recogni-tion impairment for animals is merely the result of ageneralized visual recognition de�cit. On this view, thereason that E.W. shows a category-speci�c de�cit foranimals is because this category is perceptually morecomplex, in the sense that its members are more similarto one another than the members of other categories(e.g., Gaffan & Heywood, 1993; Humphreys & Riddoch,1987). In order to rule out the possibility that E.W.’sdif�culties in visually recognizing animals was the resultof a general visual processing de�cit, we tested herability to perform dif�cult visual perceptual tasks.
Dif�cult Visual Matching and Categorization Tasks:The BORB
E.W. was administered those sections of the BirminghamObject Recognition Battery (BORB; Riddoch & Hum-phreys, 1993) that allow an assessment of her ability toperform complex visual matching and categorizationtasks (see “Methods”). The tests (Tests 7, 8, 11, and 12)were administered in the standardized manner. She per-formed quite well and within normal range on all tasks.
Tests 7 (Minimal Feature Test) and 8 (ForeshortenedView Test) of the BORB are complex visual processingtasks that involve matching a picture of an object seenfrom a “standard” viewpoint to the same object viewedfrom a different, nonstandard viewpoint. E.W. performedthese tasks quite well. She scored 88% and 84% cor-rect—well within normal limits. And she performed�awlessly in a similar task (Test 11: Object Matching Test)that required her to match an object (or a part of anobject) to a different view of that object, and very well
(93% correct) in an association matching task that re-quires the semantic interpretation of pictured objects.
Face Recognition
E.W. was asked to name pictures of famous people usingthe Boston Famous Faces Task (Albert, Butters, & Levin,1979). She was given unlimited time to respond. Acrossall the faces (across all decades and level of dif�culty),she performed very well (30 of 56 correct) and withinnormal range for her age group. This result suggests thatE.W. does not have a speci�c problem processing com-plex visual material, such as faces.
Summary
There are no indications in the results of our tests ofE.W.’s visual processing performance that she has anyimpairment in visual recognition or categorization otherthan speci�cally for animals. These results invite theinference that E.W.’s impairment in visually recognizinganimals is categorically based and most likely has thesame cause as her naming de�cit.
E.W.’s Category-Speci�c De�cit for Animals isNot Modality-Speci�c.
To this point we have documented that E.W. has a selec-tive de�cit in naming and recognizing animals, and wehave shown that the recognition impairment is unlikelyto merely re�ect a generalized visual processing defect.Here we provide direct evidence that her category-speci�c de�cit has a conceptual basis because she isalso impaired in verifying whether an animal has a par-ticular property or attribute (e.g., Does a giraffe havefour legs?). We also address directly the issue of whetherE.W. has more severe dif�culty in processing the visual/perceptual as opposed to functional/encyclopedic prop-erties of animals. As noted in the “Introduction,” thelatter issue is crucial for distinguishing between the SFTand amodal theories of semantic representation.
Attribute Processing Task 1: Central Attributes
In this task, E.W. and control subjects were asked todecide whether an attribute statement about an objectis true or false. In this and the following attribute tasks,only a subset of the total data was used so that itemscould be matched on familiarity (see “Methods” sectionfor a description of the matching procedure). The pat-terns of performance on the total data set did not differfrom the patterns reported here. Results for E.W. andcontrol subjects (n = 5) are presented in Table 6. E.W.has severe dif�culties making judgments about animals(z = 5.39, p < 0.05), independent of whether the state-ment involves perceptual or associative/functional infor-
Caramazza and Shelton 13

mation (z < 0.5). Thus, she only responded correctly to65% of the visual and the functional/associative attributestatements about animals. By contrast, her performanceon inanimate objects was very good and fell withinnormal limits.
Attribute Processing Task 2: Food/NonfoodAnimals
This task was designed to examine E.W.’s knowledge ofanimals that can be eaten (food animals, e.g., chicken,
pig, cow) and those that are not typically eaten (bypeople of her generation and cultural background) in theUnites States (nonfood animals, e.g., dog, horse, robin).The rationale was to examine her visual/perceptual andassociative/functional knowledge of animals that mightbe more familiar to her (food animals), even though theanimals in the “food” group were matched to the animalsin the “nonfood” group on familiarity.
As shown in Table 7, E.W. had severe problems answer-ing questions about animals, both for visual/perceptualand associative/functional information and for food andnonfood animals (all z’s < 1). She performed perfectly,however, when answering the question concerningwhether or not the animal is a food animal (the questionabout edibility was judged to be very easy by controlsubjects).
Attribute Processing Task 3: General/Speci�c PropertyJudgments
Some properties of a category are shared by all or mostmembers of a category; others are speci�c to one or onlya few members of the category. Those properties that aretrue of all members of the category may be more impor-tant or even essential for the inclusion of an object in acategory, but they do not contribute to distinguishingamong members of the category. Thus, for example,knowing that something has a mouth and eyes may besuf�cient to classify it as an animal but will not help indeciding whether it is a canary or an eagle. By contrastthe information that an object is about the size of achild’s �st and yellow may be insuf�cient to supporta decision about whether or not it is an animal butprovides suf�cient information for distinguishing be-tween an eagle or a canary. We will refer to the formerknowledge as general-attribute and the latter as speci�c-attribute knowledge.22 An important question concernsthe level of knowledge that is damaged in E.W. Is sheequally impaired with general- and speci�c-attributeknowledge?
The motivation for this question springs in part from
Table 6. Central Attributes Judgment Task (proportioncorrect; range for control subjects)
E.W. Animals Objects
Visual/perceptual:
True 38 of 59 (0.64) 54 of 59 (0.92)
False 26 of 39 (0.67) 37 of 39 (0.95)
Associative/functional:
True 20 of 30 (0.67) 30 of 30 (1.0)
False 17 of 27 (0.63) 26 of 27 (0.96)
Controls Animals Objects
Visual/perceptual:
True 0.92–0.98 0.92–1.0
False 0.85–1.0 0.86–1.0
Associative/functional:
True 0.93–1.0 0.93–0.97
False 0.96–1.0 0.92–1.0
Note: On the rating scale, 1 = just guessing and 5 = extremecon�dence. The mean (SD) visual/perceptual rating for true state-ments is 4.85 (0.22) and for false statements is 4.75 (0.31). Themean associative/functional rating for true statements is 4.92 (0.24)and for false statements is 4.88 (0.15).
Table 7. Food/Nonfood Animal Attributes Judgment Task (proportion correct; range for control subjects)
Subject Food Animal Nonfood Animal
E.W.:
Visual/perceptual 27 of 41 (0.66) 26 of 41 (0.63)
Associative/functional 32 of 41 (0.78) 32 of 41 (0.78)
Controls:
Visual/perceptual correct 0.83–0.93 0.85–1.0
Associative/functional correct 0.88–0.93 0.81–0.88
Note: On the rating scale, 1 = just guessing and 5 = extreme con�dence. The mean (SD) visual/perceptual rating is 4.69 (0.38). The mean asso-ciative/functional rating is 4.72 (0.38).
14 Journal of Cognitive Neuroscience Volume 10, Number 1

the observation that E.W. is quite capable of distinguish-ing animals from artifacts (though she cannot distinguishbetween real and unreal, but possible, animals). Thisraises the issue of whether her ability to categorize anobject as “animal” also allows her to access general-attribute knowledge about animals. In the eventualitythat she could systematically access such information, itwould establish that she is not confused about what ananimal is but only about detailed distinctions amongcategory members. To test this possibility, E.W. and con-trol subjects (n = 2) were asked to decide whethergeneral- and speci�c-attribute knowledge statementsabout an object are true or false (see “Methods” fordetails). The results are shown in Table 8 and clearlydemonstrate that E.W. has no dif�culty with questionsconcerning attributes that are shared by all (or almostall) members of a category. Because of this, controlsubjects did not complete these questions. E.W. per-formed almost perfectly with speci�c-attribute informa-tion about inanimate objects (and better than bothcontrol subjects) and much worse with speci�c-attributeinformation for animate objects (z = 4.69, p < 0.05).Similar to the pattern of performance on the previousattribute judgment tasks, her knowledge of speci�c at-tributes of animals is equally disrupted for visual and forassociative/functional information (z < 1).
Attribute Processing Tasks: Collapsing Across Tasks
The pattern of performance across the attribute judg-ment tasks reported thus far is clear: E.W. has severe
problems with the attributes of animate objects regard-less of whether or not they relate to visual properties orto associative properties and has no problem with attrib-utes of inanimate objects. For animals, the only questionson which she performs very well are general-attributeknowledge questions, indicating that she knows thatanimals have eyes and mouths and that they breathe anddigest food, but little else besides that.
To examine further her performance with visual/per-ceptual and associative/functional information for ani-mals and nonanimals, we collapsed her data across allthe attribute judgment tasks. This gives us a very largenumber of data points in each cell, as well as a range ofdif�culty of questions. Performance is summarized inTable 9 for matched items. Although there is a smalladvantage for associative/functional information over vis-ual/perceptual information for animals, this difference isfar from signi�cant (z = 0.7974, p > 0.05) and is onlyslightly larger than a similar difference for inanimateobjects.
Attribute Processing Task 4: Shared Attributes
Although the evidence presented thus far on attributejudgments would seem to indicate that E.W. has a cate-gory-speci�c de�cit for animals, there remains the possi-bility that the putative modality-speci�c nature of thede�cit merely re�ects a selective de�cit in processingcertain attributes that happen to be differentially distrib-uted across semantic categories. That is, there is thepossibility that the problem concerns the properties
Table 8. General/Speci�c Attributes Judgment Task (proportion correct; range for control subjects)
General Speci�c
Subject Visual Associative Visual Associative
E.W.:
Animals 130 of 130 (1.0) 129 of 130 (0.99) 74 of 100 (0.74) 115 of 150 (0.77)
Inanimate 60 of 60 (1.0) 75 of 75 (1.0) 100 of 100 (1.0) 149 of 150 (0.99)
Controls:
Animals – – 0.92–0.94 0.85–0.98
Inanimate – – 0.87–0.93 0.82–0.88
Note: On the rating scale, 1 = just guessing and 5 = extreme con�dence. The mean (SD) visual/perceptual rating is 4.91 (0.23). The mean asso-ciative/functional rating is 4.86 (0.23).
Table 9. Matched Data Collapsed Across All Attribute Judgment Tasks (proportion correct)
Animals Inanimate objects
Visual/perceptual 202 of 301 (0.67) 217 of 226 (0.96)
Associative/functional 223 of 300 (0.74) 217 of 219 (0.99)
Caramazza and Shelton 15

themselves and not the semantic category of animals. Ifthat were the case, a category-speci�c de�cit emergedonly because we queried different properties for animalsand nonanimals, and these properties just happened tobe differentially damaged. To circumvent this potentialproblem, E.W. was tested in two attribute judgmenttasks with the same attributes for animate and inanimateobjects.23
Attribute Processing Task 4a: Size Judgments. In thistask, E.W. was asked to decide which of two animate ortwo inanimate objects is the larger of the two. Perfor-mance is summarized in Table 10 and demonstrates thatE.W. had more dif�culty with animal size judgments thaninanimate size judgments (z = 1.83, p < 0.06) and isoutside the normal range for the animal category but notthe inanimate category.
Attribute Processing Task 4b: Other Shared Attributes.In this task, E.W. was asked to decide whether a propertystatement about animate and inanimate objects was trueor false (see “Methods” for examples of the propertiesincluded in the task). Performance is summarized in Table10 and indicates that E.W. had much more dif�culty withattributes of animate items than those same attributesassociated with inanimate items (z = 3.38, p < 0.05).
Attribute Processing Task 5: Visible and Not VisibleAttributes
E.W.’s performance in the preceding two tasks showsthat her de�cit involves the category of animate objectsand is not speci�c to attribute concepts found exclu-sively with animals. As a further test of her ability toprocess the attribute concepts that she is unable toverify in the context of judgments about animals, weasked her to answer questions about properties of pic-tured animals in one of two conditions: when the prop-
erties were visible and when they were not visible in thepictured animal. Thus, for example, E.W. was asked todecide whether a horse has a tail when shown a pictureof a horse with a tail and when shown a picture of ahorse from a perspective that did not show its tail. Thecrucial issue is whether she can answer attribute ques-tions correctly when the information is available in thepictured object but answers incorrectly when the infor-mation is not given in the picture. E.W. had no problems(100% correct) answering the attribute questions whenthe information was available in the picture but hadserious problems (55% correct) when the informationwas unavailable in the picture (z = 7.28, p < 0.05).
Summary
The results of the attribute processing tasks show thatE.W.’s category-speci�c de�cit extends beyond her nam-ing and picture recognition dif�culties to involve lan-guage comprehension. Her de�cit appears to beconceptually based and is not just a de�cit of visualperception or of lexical access in production. The resultsalso clearly show that E.W.’s de�cit involves equally thevisual/perceptual and the functional/associative proper-ties of animate objects. And, �nally, her dif�culties withobject attributes are the result of a category-speci�cimpairment for animate objects and not a generalizedde�cit for those attribute concepts. In short, the resultsshow that E.W. has a semantic category-speci�c de�cit.
DISCUSSION
The results we have reported can be summarized asfollows:
1. E.W. has a category-speci�c de�cit restricted to thecategory of animate objects: She is disproportionatelyimpaired in naming animals relative to other living things,such as fruits and vegetables, and relative to all otherinanimate categories tested, including food items. Thecategory-speci�c nature of the de�cit persisted understrict control of potential nuisance factors such as itemfamiliarity, visual complexity, frequency, or their combi-nation.
2. E.W. has a category-speci�c de�cit in visually (andauditorily) recognizing animate objects: She is dispropor-tionately impaired in recognizing animals relative toother living items and artifacts. This de�cit is not theresult of a generalized perceptual processing de�cit in-teracting with differential levels of visual complexityacross categories. E.W. performed within normal limits incomplex visual processing tasks.
3. E.W. has a category-speci�c de�cit in language com-prehension. In various types of attribute processing tasksshe performed very poorly with statements about ani-mate objects but performed within normal limits withstatements about other living things and artifacts. This
Table 10. Size Judgments Task and Matched AttributesJudgments Task (proportion correct; range for controlsubjects)
Size Judgments
Subject Animals Inanimate
E.W. 13 of 18 (0.72) 17 of 18 (0.94)
Controls 0.94–1.0 0.94–1.0
Matched Attributes Judgments
Subject Animals Inanimate
E.W. 32 of 42 (0.76) 42 of 42 (1.0)
Controls 0.94–0.97 1.0
Note: On the rating scale, 1 = just guessing and 5 = extremecon�dence. The mean (SD) rating is 4.97 (0.09).
16 Journal of Cognitive Neuroscience Volume 10, Number 1

selective de�cit persisted in the face of stringent controlsof various nuisance variables.
4. E.W.’s category-speci�c de�cit for animals is notalso modality-speci�c (for visual knowledge). In variousattribute processing tasks, she consistently performedequally poorly with visual and functional/associativestatements of animate objects and equally well, andwithin normal limits, for all attributes of inanimate ob-jects.
The facts 1 through 4 allow several conclusions aboutthe nature of E.W.’s de�cit. E.W.’s selective impairmentin recognizing and naming animate objects is not re-stricted to the recognition of visually presented stimulibut extends to auditory stimuli as well, and the fact thather impairment extends to language comprehensiontasks (facts 1 through 3) allows the inference that herde�cit involves the semantic system and not simply thevisual recognition or the word production systems. Fact4—E.W.’s selective impairment concerning animate ob-jects is equally severe for their functional/associative andvisual attributes—allows the inference that the damagedsystem is not organized by “modality;” that is, the dam-aged system does not represent visual and functional/associative knowledge in distinct subsystems.
These conclusions have important implications for theissues raised in the “Introduction” concerning the organi-zation of conceptual knowledge in the brain. First, theyare at variance with the sensory/functional theory (SFT)of semantic category-speci�c de�cits (Farah & McClel-land, 1991; Silveri & Gainotti, 1988; Warrington & Shal-lice, 1984) and thus provide a clear refutation of thistheory of semantic organization. Second, they are consis-tent with the hypothesis of amodal organization of thesemantic system, as proposed by the organized unitarycontent hypothesis (OUCH; Caramazza et al., 1990). And,third, they encourage the hypothesis of domain-speci�corganization of knowledge in the brain. We discuss eachof these issues in turn.
Refutation of the Sensory/Functional Theory ofCategory-Speci�c De�cits
The SFT makes two clear predictions concerning cate-gory-speci�c de�cits. One prediction is that category-speci�c de�cits should not honor “narrow” semanticcategory boundaries but should instead be determinedby the role played by visual (sensory) features in deter-mining the meaning of category members. This translatesinto the expectation that the categories of animals andfruits and vegetables (as well as other visually weightedcategories) should be damaged together. The other majorprediction is that category-speci�c de�cits for livingthings should necessarily also be modality-speci�c, in thesense that knowledge of visual properties must be dis-proportionately impaired relative to knowledge of func-tional/associative properties. Neither prediction held up
in our investigation of E.W. Her category-speci�c de�citis restricted to the category of animals, leaving unaf-fected her knowledge of fruits, vegetables, and artifacts,and her knowledge of functional/ associative attributesof animals is as impaired as her knowledge of their visualattributes.
Other results in the literature are highly problematicfor the SFT. The case of selective sparing of the categoryof animals (Hillis & Caramazza, 1991) and of subjectswith selective damage of the category of fruits andvegetables (Farah & Wallace, 1992; Hart et al., 1985)complement the results reported for E.W. (which showthat she has a selective de�cit for animals). These resultsjointly establish that category-speci�c de�cits can bevery narrow, involving separately the categories of ani-mate objects and the category of fruits and vegetables,thus undermining the expectation derived from the SFTthat the categories of living things should be damagedtogether. The other set of results that converges withE.W.’s performance in undermining the SFT concernsreports of subjects with category-speci�c de�cits forliving things who do not show a disproportionate de�citfor visual attributes of living things (Funnell & DeMornay Davies, 1997; Laiacona, Barbarotto, & Capitani,1993; Laiacona et al., in press; Sheridan & Humphreys,1993). Thus, on this issue too, there is no support for theSFT.
There remain two other sources of evidence that havebeen cited in support of the SFT—the results of someneuroimaging studies (e.g., Martin, Wiggs, Ungerleider, &Haxby, 1996) and the results of computational modeling(Farah & McClelland, 1991). We will discuss the neuro-imaging results in greater detail in a later section. Here,it suf�ces to say that some of the neuroimaging resultsare consistent with the SFT, but they are also consistentwith alternative interpretations. Thus, these results donot help distinguish between alternative theories of se-mantic organization. We begin with a discussion of theputative evidence from computational modeling.
Farah and McClelland (1991) purport to show thatdamage to the visual component of a modality-organizedsemantic system can explain the existence of category-speci�c de�cits. They did this by developing a paralleldistributed processing (PDP) model in which semanticknowledge is subdivided into visual and functional com-ponents. The two major assumptions of this model arethat visual and functional semantic properties are repre-sented in separate but interconnected networks and thatthe ratio of visual to functional properties for livingthings is much larger than that for nonliving things.When the visual components of various networks imple-menting these assumptions were lesioned, the resultingpatterns of performance mimicked the category-speci�cde�cit for living things, and the reverse dissociation wasobtained when the functional component of the net-work was lesioned. This pattern of results provides anexistence proof that a category-speci�c de�cit can be
Caramazza and Shelton 17

obtained without having to postulate categorical organi-zation of semantic knowledge.
As an existence proof that category-like effects canemerge from a noncategorically organized semantic sys-tem, the results reported by Farah and McClelland (1991)need not be controversial, nor even surprising for thatmatter. In some respects the results of the implementedmodel merely re�ect their assumption that the meaningof living things is much more dependent on visual prop-erties than is the case for nonliving things.24 In theirmodeling experiments, the ratio of visual to functionalproperties for living things was set to 16.1:2.1, and fornonliving things it was set to 9.4:6.7. Thus, damage tothe visual semantic network could hardly result in any-thing but more severe dif�culty for living than nonlivingthings. Put differently, the highly disproportionate ratiosof visual to functional properties for the categories ofliving and nonliving things can be seen as roughly ap-proximating a categorical distinction between the twosemantic domains. The model merely capitalizes on thiscorrelation. As noted by Farah and McClelland (1991),“The ability of a modality-speci�c semantic memory ar-chitecture to account for category-speci�c semanticmemory impairments depends, of course, on there beinga correlation between modality of knowledge and cate-gory of knowledge” (p. 355). If there were no suchcorrelation, it would not be possible to devise a modal-ity-based explanation for category- speci�c de�cits. Thus,the ultimate value of the modeling demonstration de-pends entirely on the validity of the empirical assump-tions made by the model, that is, the assumptions aboutthe ratios of visual to functional properties for the livingand nonliving categories, respectively.
Warrington and her collaborators (Warrington &McCarthy, 1983, 1987; Warrington & Shallice, 1984) hadsuggested that living things are known and distinguishedprimarily through their visual attributes, whereas nonliv-ing things are known primarily through their func-tional/associative properties. This suggestion was basedon mere intuition; however, Farah and McClelland (1991)attempted to provide an empirical foundation for thisintuition. They did this by counting the number of visualand functional descriptors that subjects identi�ed indictionary de�nitions of the living and nonliving thingsused by Warrington and Shallice (1984, Experiment 2).They found that subjects underlined more visual descrip-tors for living than nonliving things (an average of 2.68and 1.57, respectively) but many fewer functional de-scriptors for living than nonliving things (an average of0.35 and 1.11, respectively). Thus, the ratio of visual tofunctional properties was found to be far higher forliving (7.7:1) than nonliving things (1.4:1). These resultsseem to con�rm the speculation about the relative im-portance of visual and functional-associative propertiesin determining the meaning of living and nonlivingthings. However, there is a serious �aw in the experimen-tal procedure used by Farah and McClelland that makes
interpretation of their results problematic at best. Theproblem concerns the instructions given to subjects onwhat to count as visual and functional-associative de-scriptors.
Farah and McClelland (1991) asked their subjects tounderline either visual or functional descriptors inde�nitions of living and nonliving things. Subjects in thevisual descriptor condition were asked to underline allwords referring to the visual appearance of an item.Subjects in the functional descriptor condition wereasked to underline “all occurrences of words describingwhat the item does or what it is for” (p. 342; emphasisadded). Now, it is clear that all objects, living and nonliv-ing, can be described by reference to their visual appear-ance and, therefore, counting the visual descriptors inde�nitions of items in the two categories may provide afair estimate of the sensory properties known by sub-jects about various objects. However, asking subjects tounderline “what it is for” for living and nonliving thingswill not provide a fair estimate of the nonsensory prop-erties known for living things. That is, because the useof an item—what it is for—is a highly speci�c propertytypically found with artifacts but not natural kinds, hav-ing a “function” is almost a distinguishing feature be-tween living things and artifacts. It is meaningful to ask“What is a table or a knife for?” but barely felicitous toask “What is a lion or a squirrel for?” Thus, it is hardlysurprising that Farah and McClelland found large discrep-ancies in the ratios of visual to functional descriptors forliving and nonliving categories; these discrepancies areentirely due to the instructions given to subjects onwhat could count as a functional/associative feature. Wecan easily imagine reversing the ratios of visual to func-tional properties for living and nonliving things by in-structing subjects to underline visual properties that arefound almost exclusively in nonliving things, such as “haswheel-like features,” “has straight edges,” “has wood-liketexture,” and so on. In this case, we would most likely�nd that nonliving things would have a larger ratio ofvisual to functional/associative attributes than livingthings.
It is crucial that the criteria for identifying the sensoryand nonsensory properties of living and nonliving thingsnot contain a built-in category bias. However, the instruc-tions Farah and McClelland (1991) gave their subjectseffectively led them to exclude from consideration suchimportant nonsensory properties of living things as “livesin desert,” “ferocious,” “carnivore,” “herbivore,” “domesti-cated,” “solitary,” “edible,” “egg-laying,” and so on. If theseand other nonsensory properties had not been excludedby the instructions biasing subjects to consider only“what it is for”—principally a property of artifacts—thedifferences in the ratios of sensory to nonsensory prop-erties for living and nonliving things would not havebeen nearly as large as reported by Farah and McClel-land. Indeed, when we instructed two groups of subjectsto underline either all sensory properties or all non-
18 Journal of Cognitive Neuroscience Volume 10, Number 1

sensory properties in the de�nitions of the living andnonliving items used by Farah and McClelland,25 weobtained the following ratios of sensory to nonsensoryproperties for living and nonliving things: 2.9:2.5 and2.2:2.3 respectively.26 Similar results have recently beenobtained by McRae, de Sa, and Seidenberg (1997) usinga different procedure to estimate the ratio of visual tofunctional attributes for living and nonliving categories.These ratios do not provide empirical support for theassumption that there is a correlation between “modalityof knowledge and category of knowledge.”27
In this section we have argued that neither of themajor expectations derived from the SFT—the predictedcorrelation of category de�cits and the predicted co-occurrence of category- and modality-speci�c de�cits—receives support from E.W.’s performance or theperformance of other reported cases of category-speci�cde�cit. We have also argued that the computational mod-eling evidence (Farah & McClelland, 1991) offers nomore than an existence proof that category-like effectscan result from damage to noncategorically organizedsubsystems whose contents are correlated with seman-tic categories. However, the success of the computa-tional modeling demonstration does not require that thecontents of the subsystems be modality-speci�c. All thatit requires is that there be a correlation between thecontents of speci�c subsystems and semantic categories.The contents of subsystems can vary from completelynonoverlapping (categorical) representations to modal-ity-speci�c subsystems that are correlated with differentsemantic categories (SFT) to modality-neutral clumps ofintercorrelated semantic properties that are correlatedwith different semantic categories (OUCH). Thus, thecomputational modeling demonstration reported byFarah and McClelland would only be useful in distin-guishing between the SFT and alternative accounts ofcategory-speci�c de�cits if its assumptions about thecontents of the hypothesized subsystems could be mo-tivated empirically. However, we have argued that theempirical evidence provided by Farah and McClellandfor a putatively large discrepancy in the ratios of visualto functional/associative properties for living and nonliv-ing things is tainted by having inappropriately restrictedwhat could count as a functional/associative attribute(because only “what is it for” attributes were allowed,favoring the kinds of attributes that artifacts, but notanimals, are likely to have). And we have gone on toshow that the ratios of visual to functional/associativeproperties for living and nonliving things are, in fact,approximately equal when subjects are instructed tocount all perceptual and all nonperceptual (func-tional/associative) descriptors. Thus, the central assump-tions about the nature of the semantic subsystems in themodel proposed by Farah and McClelland are withoutempirical foundation, thereby undermining the value ofthe computational modeling demonstration as evidencein support of the SFT.
The Organized Unitary Content HypothesisRevisited
The organized unitary content hypothesis (OUCH)makes two clear predictions: Category-speci�c de�citscan be narrower than the living/nonliving distinctionand category-speci�c de�cits are not also modality-speci�c. E.W.’s performance is in agreement with bothpredictions from the OUCH: (1) Her de�cit is restrictedto the category of animate things, and (2) she is equallyimpaired in processing the visual and functional/associa-tive attributes of animals and of no other semantic cate-gory. The model’s major drawback is that OUCH does notprovide a principled explanation for the attested pat-terns of category-speci�c de�cits: It fails to explain whythe unambiguous cases of category-speci�c de�cit seemto respect the tripartite distinction animal/plant/artifact(and not some other pattern of category-speci�c de�-cits—i.e., why not a selective de�cit for aquatic animals)and why the most prevalent category-speci�c de�citinvolves the category of animals. Thus, although OUCHmay provide a useful noncategorical framework withinwhich to explore various aspects of semantic memoryde�cits, including their category-speci�c form, we mayhave to consider a radically different theoretical frame-work to explain the high prevalence of selective de�-cits for animals and the tripartite distinction animal/plant/artifact in the occurrence of category-speci�cde�cits.
Domain-Speci�c Organization of ConceptualKnowledge in the Brain: EvolutionaryAdaptations?
None of the evidence reviewed here, including E.W.’sperformance, compels us to adopt the view that concep-tual knowledge is organized categorically in the brain.However, doing so provides a natural explanation forprecisely that feature of the phenomenon of category-speci�c de�cits that has proven to be most problematicfor the reductionist accounts: the fact that the categoriesof animals, fruits and vegetables, and artifacts can bedamaged independently of each other. On this view, theempirically observed tripartite distinction of category-speci�c de�cits follows directly from the assumptionthat these, and only these, three categories form the basisfor the organization of conceptual knowledge. But theexplanatory power of this assumption would amount tonaught unless it could be given independent theoreticaland empirical justi�cation. That is, unless we can inde-pendently motivate the assumption of categorical organi-zation of conceptual knowledge, we would have merelyassumed what we are trying to explain—an infelicitouscircularity. It is important, therefore, to determinewhether there might not be theoretical arguments andempirical evidence that could be used to independently
Caramazza and Shelton 19

support the assumption that conceptual knowledge isorganized categorically.
In the “Introduction” we speculated that adoption ofan evolutionary perspective to the problem of how con-ceptual knowledge is organized in the brain might pro-vide the motivation for assuming that it is organizedcategorically. It is not implausible to assume that evolu-tionary pressures led to speci�c adaptations for recog-nizing and responding to animal and plant life (the latteroperationally represented by fruits and vegetables inresearch on category-speci�c de�cits). The �tness valuefor these adaptations are obvious: Animals are potentialpredators but also a potential source of food; plants area source of food and medicine. The ability to recognizeand respond quickly to types of animals has clear sur-vival and reproductive value, as does the ability to accu-rately distinguish among different plants for theiralimentary and medicinal value. In perceptual and cog-nitive terms, these adaptations might consist of special-ized processes for the rapid and accurate classi�cationof objects as animals, as plant life, or as neither of thesetwo categories of objects. In terms of neural mecha-nisms, the relevant adaptations might consist of dedi-cated neural circuits for processing animals and plantlife. And because of the clear affective/emotional compo-nent associated with �ight and feeding responses toanimals and plant life, it is not implausible to furtherassume that the neural adaptations would involve cir-cuits that include the limbic system. If these speculationswere to be even approximately correct, we would havethe independent motivation we have been seeking insupport of the assumption that conceptual knowledge isorganized categorically in the brain. The evolutionaryadaptations for recognizing animals and plant life wouldprovide the skeletal neural structures around which toorganize the rich perceptual, conceptual, and linguisticknowledge modern humans have of these categories.
An important implication that follows from the adop-tion of an evolutionary perspective on the organizationof conceptual knowledge in the brain is a restriction onwhat is likely to count as evolutionarily signi�cant se-mantic categories—that is, categories for which it isplausible to propose that their successful recognitionwould have �tness value. We have argued that animalsand plant life satisfy the latter criterion. Whether or notthe ability to recognize and use “tools” should also beaccorded �tness value is less obvious, although not im-plausible (see Hauser, in press). However, it is not neces-sary that tools be accorded such status for the argumentdeveloped here. It is suf�cient that the categories ofanimals and plant life be distinguished from “other” cate-gories. Thus, on this view, the only “true” category-speci�c de�cits28 will be those involving the categoriesof animals, plant life, and (by contrast) artifacts and notother �ner-grained distinctions such as land animals,fruits, kitchen utensils, and so on.
Another implication that follows from the assumption
that evolutionary adaptations provide the basis for thecategorical organization of conceptual knowledge in thebrain is that the incidence of category-speci�c de�citsfor living things should be greater than that for nonlivingthings. This implication is derived from the suppositionthat specialized neural mechanisms are more likely to behighly localized and consequently more prone to selec-tive damage. This expectation would seem to have somesupport. Most category-speci�c de�cits that have beenreported over the past 15 years have involved the cate-gories of living things. Of course, it could be that someof the putative category-speci�c de�cits for living cate-gories are merely artifacts of inadequate stimulus con-trols (Stewart et al., 1992; Funnell & Sheridan, 1992).However, as discussed extensively in the “Introduction,”there is now a substantial number of well-studied casesof category-speci�c de�cits for living categories that can-not be dismissed as methodological artifacts (see, forexample, cases in Laiacona, Barbarotto, & Capitani, 1993;Laiacona et al., 1997). Thus, even with a major correctionfor possible methodological artifacts in the early studiesof category-speci�c de�cits, the discrepancy betweenthe incidence of selective impairment for the living andnonliving categories remains striking.
A further implication of the view that there might bededicated neural circuits for the categories of animalsand plant life involves the category of foods. Because theassumed neural circuitry for animals and plant life in-volves the limbic system, it is not unreasonable to sup-pose that the semantic category of foods might sharesome of its neural circuitry. Consequently, damage toneural structures that affect the categories of animals orof fruits and vegetables are also likely (though not nec-essarily) to result in impairment of the category of foods,even though many members of the latter category aremore properly considered artifacts (consider, for exam-ple, spaghetti, fudge, hamburger, and so on). This assump-tion would explain why some cases of category-speci�cde�cits for living categories also show impairments forfoods.
The hypothesis that knowledge is organized categori-cally in the brain provides a good account of extantresults on semantic category-speci�c de�cits. However,it is important to stress here that the appeal to evolu-tionary pressures as a possible causal basis for the cate-gorical organization of knowledge in the brain is notnecessarily incompatible with the cognitive principlesthat characterize the OUCH and SFT proposals. Indeed,it is quite likely that both domain-speci�c and domain-general learning principles are involved in the organiza-tion of conceptual and linguistic knowledge in the brain.The domain-speci�c principles have a very narrow rangeof applicability and serve only to explain why it mightbe the case that knowledge of animals and of plant lifeis represented in distinct neural circuits. This account issilent on the larger issue of how conceptual knowledgewithin the broad categories of living and nonliving
20 Journal of Cognitive Neuroscience Volume 10, Number 1

things is organized. And it could very well turn out thatdomain-general principles such as those invoked underOUCH (or SFT, although no empirical support has beenfound for this account) provide the basis for the organi-zation of conceptual knowledge within the distinct cate-gories of animals, plant life, and artifacts.
Domain-Speci�c Organization of ConceptualKnowledge in the Brain: OntogeneticConsiderations
The idea that neural adaptations are responsible for theorganization of conceptual knowledge in the brain hasdirect implications for theories of acquisition of thisknowledge. Speci�cally, this thesis implies that thereshould be specialized mechanisms for the recognitionand categorization of the members of the categories forwhich speci�c adaptations have evolved. And, in fact, thecontrasting viewpoints about the possible bases for theexistence of semantic category-speci�c de�cits havetheir counterparts in the developmental literature. Therethe issue has been framed in terms of whether cognitivedevelopment is to be understood as the product ofdomain-general or domain-speci�c processes. The do-main-general view holds that a common set of verypowerful learning mechanisms underlies the acquisitionof all forms of knowledge: from mathematical to musicalskills and from language to the ability to play backgam-mon. This position is most clearly represented by theclassical empiricist theories of learning and memory(e.g., Skinner, 1957) and more recently by connectionisttheories (e.g., McClelland & Rumelhart, 1986). The do-main-speci�c view holds that highly specialized, dedi-cated mechanisms are involved in the acquisition ofspeci�c types of knowledge (Carey, 1985; Pinker &Bloom, 1990; Sperber, 1994). The best-known domain-speci�c theory is the theory of universal grammar oflanguage acquisition proposed by Chomsky (1980,1986). However, domain-speci�c theories have also beenproposed for speech perception (Liberman & Mattingly,1989), naive physics and psychology (Carey & Spelke,1994), number (Carey & Spelke, 1994; R. Gelman, 1990),the animate-inanimate distinction (R. Gelman, 1990), folkbiology (Atran, 1995; Sperber, 1994), and, more generally,various aspects of animal learning (Rozin & Schull, 1988).In each case, it could be proposed that specialized cog-nitive mechanisms, re�ecting adaptations in response toevolutionary pressures, are responsible for the acquisi-tion of a speci�c type of knowledge (for recent reviewssee papers in Hirschfeld & S. Gelman, 1994; Sperber,Premack, & Premack, 1995).
Of particular relevance here is the proposal that theacquisition of the animate-inanimate distinction involvesdomain-speci�c mechanisms. Although alternative pro-posals have been offered concerning the precise natureof the content of the domain-speci�c mechanism foracquiring the animate-inanimate distinction (see Atran,
1995; Carey, 1985; 1995; R. Gelman, 1990; R. Gelman &Brenneman, 1994; R. Gelman, Durgin, & Kaufman, 1995;Keil, 1994; Sperber, 1994), they all maintain that a set ofinnate principles guides its acquisition. These principlesdo their work by specifying the core content of the twocategories. Their major function is to direct attention tothe relevant exemplars of the categories by focusing onthe salient distinctions among them. Candidate featuresof the environment that could serve the function of“triggering” a �rst-pass distinction between categoriesinclude the motion patterns of objects and their physicalappearance (Mandler, 1992). However, the perceptualdistinctions are not themselves criteria for distinguishingbetween categories. They merely function as cues tocategory membership. The categorical distinction is fun-damental and precedes the acquisition of speci�c per-ceptual facts about their members. In other words, theperceptual features of objects are powerful cues tosameness of category, but they do not constitute thecore properties of the animate-inanimate distinction. Thelatter are more likely to be complex properties that donot correspond to any simple perceptual feature. Thus,candidate core properties for animate objects might betheir internal structure (e.g., having organs), whetherthey are capable of self-initiated motion, and whethertheir motion patterns can be construed as having inten-tional cause (Premack, 1990); for artifacts, a core prop-erty might be their intended function (Bloom, 1996).
There is considerable evidence in favor of the domain-speci�c thesis of the acquisition of the animate-inanimate distinction. Perhaps the most compelling re-sults are those concerning the infant’s ability to distin-guish physical from psychological (biological) causality.Recent experiments have shown that infants as young as3 months understand many aspects of the structure ofthe physical world. For example, they understand someof the distinguishing properties of the concepts of solid-ity and support, and they understand that two objectscannot occupy the same space at the same time (Baillar-geon, 1993; Spelke, 1988). Furthermore, this knowledgeis embedded in a larger conceptual scheme that involvesobject motion and the concept of causality. Spelke, Phil-lips, and Woodward (1995) have identi�ed a set of coreprinciples about the infant’s understanding of objects—the principles of cohesion, continuity, and contact—which function to constrain the interpretation of anobject’s motion. For example, the principle of contactunderlies the infant’s understanding that objects canonly act on each other by touching. There is a large bodyof evidence that very young infants understand the dis-tinction between caused and uncaused motion throughthe principle of contact. Thus, 4-month-old infants candistinguish between the implicit causality in a ball’smotion when it is launched by another moving objectand the very similar, but fundamentally different, casewhere the two objects are separated either by a brieftemporal lag or by a small spatial gap (Leslie, 1982, 1984;
Caramazza and Shelton 21

Leslie & Keeble, 1987). These results, together with otherrecent evidence that shows that infants suspend appli-cation of the contact principle when reasoning abouthuman (and perhaps animal) actions (Spelke et al., 1995),suggest that they are able to distinguish correctly be-tween animate and inanimate objects (for otherwisethey would not know when to apply and when tosuspend the contact principle). More generally, it hasbeen supposed that a core property of the infant’s con-cept of human (and perhaps animal) is the principle of“self-propelled motion”—a concept intimately related tothe distinction between mechanical and intentionalagency (see R. Gelman, 1990; Premack, 1990; Mandler,1992).
There are two other important sources of evidence insupport of the infant’s precocity in distinguishing ani-mate from inanimate objects. One is the infant’s well-documented ability to distinguish biological fromnonbiological motion. Bertenthal and his collaborators(Bertenthal, 1993; Bertenthal, Prof�tt, & Cutting, 1984)have shown that infants as young as 3 months canreliably distinguish point-light displays of motions pro-duced by animate and inanimate objects. The other evi-dence concerns the infant’s ability to categorize staticrepresentations of objects (toy �gures and pictures).Mandler and her collaborators (Mandler, 1992, 1994;Mandler, Bauer, & McDonough, 1991; Mandler &McDonough, 1993) have shown that 9-month-old infantscan distinguish between animals and vehicles evenwhen between-category similarity is controlled. Using adishabituation paradigm, they have found that infantsdishabituate to airplanes but not dogs, cats, or rabbitsafter seeing birds even though birds were shown withoutstretched wings and thus were more “perceptuallysimilar” to airplanes than they were to dogs, cats, orrabbits. This pattern of results suggests that infants makea global differentiation between animals and artifactsbefore they fully appreciate the basic-level categorieswithin these conceptual domains (but see Quinn &Eimas, 1996 for a different interpretation). Thus, the evi-dence from infants’ precocious categorization of animateand inanimate toy objects as well as the evidence fromtheir precocious distinction of point-light displays ofanimate and inanimate motion suggest that the distinc-tion may be subserved by specialized domain-speci�cmechanisms.
In this section we have argued that recent develop-ments in the area of concept acquisition provide impor-tant converging evidence for the proposal thatconceptual knowledge is organized categorically in thebrain. The evidence con�rms the expectation derivedfrom the claim that evolutionary adaptations form thebasis for the hypothesized categorical organization ofconceptual knowledge—namely, that the concepts ofanimal and artifact are acquired through the operationof innate domain-speci�c mechanisms.
Neuroanatomical Considerations
The neuroanatomical basis for semantic category-speci�c de�cits has been addressed both throughanatomo-clinical correlations and through functional im-aging of the intact brain. The investigation of anatomo-clinical correlations has produced relatively clear thoughnot unequivocal results. The picture is clearest for casesof selective de�cit for living things. Most of these caseshave sustained damage to the left temporal lobe (as wellas the right temporal lobe in patients recovering fromherpes simplex encephalitis; see Gainotti, Silveri, Daniele,& Giustolisi, 1995; Saffran & Schwartz, 1994, for reviews).However, some of the patients with disproportionatede�cit for living things have also sustained damage tofrontal and to inferior parietal areas (the case reportedhere; Hillis & Caramazza, 1991; Laiacona, Barbarotto, &Capitani, 1993) or extensive damage due to trauma(Farah et al., 1989; Laiacona, Barbarotto, & Capitani,1993). The pattern of anatomo-clinical correlations is lessclear for cases of selective impairment to the categoryof artifacts. Lesions associated with the latter pattern ofde�cit have been more variable and have typically in-cluded fronto-parietal areas but also the temporal lobein some cases (for reviews see Gainotti et al., 1995;Saffran & Schwartz, 1994). A rough generalizationfrom this literature is that the categories of livingthings are associated with temporal and sometimesfrontal and parietal lobe lesions, whereas impairmentof the category of artifacts is associated with moredorsal lesions involving temporal, parietal, and frontalareas.
Most cases of category-speci�c de�cits have been in-terpreted as involving damage to the semantic system,allowing inferences about the possible organization ofsemantic knowledge in the brain. An alternative proposalhas been offered by Damasio et al. (1996), who reportedthe most extensive investigation of the neuroanatomicalbasis for category-speci�c de�cits to date. These investi-gators argued that category-speci�c naming de�cits neednot result from damage to the semantic system but canresult from damage to the lexical representations medi-ating between the semantic and the phonological con-tent of words. On this view, the analysis of patterns ofcategory-speci�c de�cits can inform hypotheses aboutthe organization of lexical (as opposed to semantic)knowledge in the brain. In their investigation of a largenumber of patients with single, focal lesions whoshowed category-speci�c de�cits (N = 30), Damasio etal. found that impairment for the category of animals isassociated with damage to the left inferotemporal(mostly anterior) area, and impairment for tools is asso-ciated with damage to the posterolateral inferotemporalarea and the junction of temporo-occipital-parietal cor-tices. Thus, despite differences in the functional interpre-tation of category-speci�c de�cits,29 the results of this
22 Journal of Cognitive Neuroscience Volume 10, Number 1

study further con�rm the important role of the lefttemporal lobe in the representation of knowledge aboutthe category of animals and the role of more dorsal areasfor artifacts.
The results from functional neuroimaging experi-ments are somewhat unclear. Three PET studies havedirectly compared performance for living things andartifacts (Damasio et al., 1996; Martin et al., 1996; Peraniet al., 1995). All three studies show segregation betweencategories, but the speci�c areas involved differ acrossstudies. For the living things category, Perani et al. andMartin et al. found activation of the occipital lobes bilat-erally and the inferior temporal lobe (bilaterally by Per-ani et al.; left only by Martin et al.). For the artifactcategory, Perani et al. found mostly left-sided activationof the lingual, parahippocampal gyri, middle occipitalgyrus, and dorsolateral frontal regions; Martin et al. foundactivation of the fusiform gyri of the temporal lobes,bilaterally, and left inferior frontal region and left precen-tral gyrus. By contrast, Damasio et al. (whose study fo-cused on activations of regions of the temporal lobe)found that naming animals resulted in activation of theleft inferotemporal area, and naming of tools resulted inactivation of posterior middle and inferior temporal gyri.Thus, there is some agreement among the three studies:The inferior temporal lobe is activated in processinganimals; the posterior middle temporal area may be moreimportant for tools. In addition, there are indications thatthe frontal lobes may be important not only for tools(Martin et al. and Perani et al.) but also for animals(Martin et al.). Finally, animal naming more strongly acti-vates the occipital lobes, bilaterally.
It is not implausible, then, to conclude that the lefttemporal lobe is crucially involved in some aspect oflexical-semantic processing, as may be parts of the leftfrontal lobe. Although Damasio et al. (1996) interpret thefunction of the different areas of the left temporal lobeactivated in naming animals and artifacts as involvingmechanisms of lexical retrieval as opposed to semanticknowledge, this conclusion is premature at best (seeNote 29). A more problematic issue concerns the inter-pretation of the differential roles of the occipital lobesin naming animals and artifacts. The interpretation pro-posed by Martin et al. (1996) is that because animals arevisually more complex, there is a “top-down” modulationof visual processing to help visual identi�cation.Whether or not this interpretation is correct, there areno reports of either category-speci�c recognition orcategory-speci�c semantic de�cits following lesions ofthe left extrastriate cortex. Thus, the relevance of theobservation that the occipital lobes are more intenselyactivated in naming animals as opposed to artifacts forissues about semantic processing remains obscure. Whatis clear, however, is that different neural mechanisms areinvolved in processing the categories of animate andinanimate objects.
Category-Speci�c Organization: Conceptual,Perceptual, or Both?
A �nal issue that needs to be addressed here is therelation between object recognition and semantic cate-gory-speci�c de�cits. Sartori and Job (1988) were the�rst to report that a patient with a semantic category-speci�c de�cit was unable to distinguish between realand chimeric animals. E.W. also showed marked dif�cul-ties distinguishing between real and unreal animals.However, the association of a category-speci�c de�cit inobject decision tasks with category-speci�c dif�cultiesin lexical processing tasks is not a necessary one. Thereare reports of patients who show category-speci�cde�cits in naming and comprehension tasks but who arefully able to distinguish real from unreal animals (e.g.,Laiacona et al., 1997; Sheridan & Humphreys, 1993).Thus, category-speci�c object decision de�cits dissociatefrom category-speci�c comprehension performance. Apossible implication of these facts is that the category-speci�c organization of conceptual categories occurs atmultiple levels of processing. Speci�cally, it could beargued not only that the conceptual system is organizedcategorically but also that the object recognition systemis sensitive to category distinctions. In other words, cate-gory-speci�c specialization may be found at both cogni-tive and perceptual processing levels. This conclusionaccords well with the view that the categorical organi-zation of conceptual knowledge in the brain re�ects aneural adaptation in response to evolutionary pressures.With this view, the �tness value of the proposed cate-gorical distinction would depend in large measure onour ability to successfully make the correct perceptualdistinctions. It would be advantageous, therefore, forspecialization to occur as early as possible in the objectrecognition system.30
Conclusion
We have reported the detailed analysis of the perfor-mance of a brain-damaged subject with a clear-cut cate-gory-speci�c de�cit. Two features of this subject’sperformance are of crucial interest: The de�cit wasrestricted to the category of animate objects, and itinvolved equally visual and functional/associative knowl-edge of animals. This pattern of performance is highlyproblematic for the sensory/functional theory of cate-gory-speci�c de�cits. We have also shown through acritical review of the literature on category-speci�cde�cits that the SFT is without empirical support. In-stead, the evidence is consistent with the OUCH model,which maintains that categorical organization in the se-mantic system emerges from the correlational structureof the properties of members of a category. However, wehave also argued that the results invite the more intrigu-ing hypothesis that there are different levels of organiza-
Caramazza and Shelton 23

tion in the neural system. One very basic level of organi-zation re�ects neural adaptations in response to evolu-tionary pressures for the rapid, successful recognition ofanimate (and plant life) objects. This is the only truecategorical organization of conceptual (and perhaps per-ceptual) knowledge in the brain. In addition, however, itis possible that there is further structure within thesebroad domains of knowledge. We have proposed that thelatter structure is not categorical in form but re�ects thecorrelational properties of members of semantic catego-ries.
METHODS
Subjects
E.W. is a 72-year-old woman who suffered a left cerebralvascular accident in February 1988. CT scans in Januaryof 1995 revealed a large area of encephlomalacia withinthe left posterior frontal and parietal lobes consistentwith the old infarct (from 1988). At the time of thestroke, EW was working in the personnel department ofa government agency. She has a high school educationand a degree from a secretarial school (i.e., approxi-mately 14 years of schooling). E.W. was �rst seen by usin February 1996.
E.W. is �uent and produces well-formed, coherent sen-tences, both in constrained production tasks and spon-taneous speech. In response to the “Cookie Theft”picture (Goodglass & Kaplan, 1983), she produced thefollowing: “The boy is on the stool getting the cookie jar.The girl is washing dishes and the water is going all overthe �oor.”
E.W. does not present with any sentence comprehen-sion impairments, and reading of single words is almostperfect. E.W. has signi�cant problems with both writtenand oral spelling.
It �rst came to our attention that E.W. had dif�cultywith animals when she was asked to name picturesincluded in a screening battery. The only pictures shecould not name were pictures of animals. E.W. reportedthat, “I’m not good with animals. They always give metrouble.” Later, we asked E.W. whether she had problemswith animals because she never had much experiencewith them. She responded, “Well, I did know some thingsabout animals. I used to take my kids to the zoo and such.It’s worse since I had my stroke. Now, I have no ideaabout animals at all. They [her speech therapists] usedto give me pictures and I told them I couldn’t do animals.I can’t even think too much about what animals looklike.”
Five elderly subjects matched in age and educationlevel to E.W. participated in the experiments (mean age:72.8 years, range: 72 to 73 years; mean education: 12.4years, range: 12 to14 years). Not all subjects completedall experiments.
Procedures
Visual Naming Task
The picture set from Snodgrass and Vanderwart (1980)was randomized and presented to E.W. for naming. Eachpicture was presented on a separate sheet of paper. E.W.was asked to provide the name of each picture, and, ifunable to provide the name, to provide any informationpossible. She was given an unlimited amount of time torespond to each picture.
Five normal control subjects were presented with thecomplete picture set and asked to provide a name foreach picture. This was done to determine if certainpictures elicited alternative, but acceptable, responsesthat may be particular to the speci�c age and educationlevel of the patient and to determine if the controlsubjects would show a familiarity effect in naming. Somepictures were consistently named incorrectly by at leasttwo or more of the control subjects. In these cases, theyeither provided a superordinate term (e.g., beetle ®bug; artichoke ® vegetable), could not produce anyname (e.g., seahorse ® 2 subjects said, “can’t think ofthe name”), provided a semantic description (e.g., cater-pillar ® thing before it changes into a butter�y; chisel ®shaves things), or in one case, could not identify thepicture (peach ® canteen, fruit, ball, puffy thing). Therewere a total of 10 items that resulted in misnaming bytwo or more of the control subjects (ant, bee, �y, beetle,caterpillar, ostrich, seahorse, peach, artichoke, chisel),and these items were removed from all the analysesinvolving the Snodgrass and Vanderwart picture set.
Sound Identi�cation Task
Sixteen animal sounds and 16 nonanimal sounds weredownloaded from a digitized sound library (SoundEditPro©) onto an audio tape. Presentation order of thesounds was randomized. Three young control subjectslistened to each sound, played twice in a row, and wereasked to identify the sound. For most items, all threesubjects correctly identi�ed a sound. There were severalitems that elicited alternative but acceptable names, andthese were considered correct responses. The soundswere presented to E.W. for identi�cation. She was in-structed to listen to the sound and to try and identify it.She was told that some of the sounds were from animalsand some were not. This task was administered twice toincrease the number of observations.
Object Decision Task
This task was developed to examine E.W.’s ability todistinguish between real and unreal animals and objectsusing the same number of stimuli in the animal andnonanimal conditions. Also, unreal nonanimal items weredeveloped that were very dif�cult to distinguish fromreal nonanimal items to equate more closely the animal
24 Journal of Cognitive Neuroscience Volume 10, Number 1

and nonanimal stimuli on level of dif�culty (see exam-ples presented in Figure 1). Thirty animal pictures and30 nonanimal pictures from the Snodgrass and Vander-wart set were spliced with one another to create unrealanimals and objects. The object parts were chosen fromitems that were similar visually (and had similar parts)but did not belong together.
Part Decision Task
The real and unreal animals and objects used in theObject Decision Task were edited and used in this task.Each “body” was separated from its real and unreal“head” and arranged on a sheet of paper, with the “body”on the top half of the sheet and the two “heads” on thebottom half of the sheet of paper (head and body are inquotes because these terms do not apply directly to theobjects—however, we will refer to the parts as head andbody for ease of reference). Each head was correctlypaired with its body on one trial and incorrectly pairedwith another body on the second trial (e.g., on one triala dog body was paired with a donkey head and a doghead; on another trial a donkey body was paired with adonkey head and a dog head). Examples of animate andinanimate items are presented in Figure 2. Items wereblocked such that a head pair only appeared once ineach block.
Visual Processing Tests: BORB Tests 7, 8, 11, and 12
Several tests from the BORB (Riddoch & Humphreys,1993) were administered to E.W. to assess her ability to
perform complex visual analysis. The tests were admin-istered in the standardized manner. In all tasks, E.W. waspresented with a target picture at the top of the pageand two probe pictures at the bottom of the page.For each task, E.W. was required to pick the appro- pri-ate probe picture, depending on the instructionsgiven. Control data are taken from that reported in theBORB.
In the Minimal Feature View Task (Test 7), the targetpresents an object pictured in a standard viewpoint. Theprobe pictures represent either the same object from adifferent viewpoint or a different object that is visuallysimilar to the target picture and presented from thesame viewpoint as the matching probe picture. For ex-ample, the target picture of a bus displays a bus whenviewed from the side, and the two probe pictures displaya bus when viewed from the top and a tractor whenviewed from the top. Thus, the main identifying featureof the target object is typically hidden from view, but theshape of the object is relatively maintained. The meanfor control subjects was 23.3 (SD = 2.0, range: 18.5–25).
In the Foreshortened View Task (Test 8), the targetpicture presents an object pictured in a standard view-point and the probe pictures represent either the sameobject from a different viewpoint or a different objectthat is visually similar to the target picture and presentedfrom the same viewpoint as the matching probe picture.For example, the target picture of a car is pictured whenviewed from the side and the two probe pictures displaya car when viewed from the front and a bus whenviewed from the front. Thus, the main identifying featureof the target object is typically in view, but the shape of
Figure 1. Examples of itemsused in the Object Decisiontask. Each item was presentedseparately.
Caramazza and Shelton 25

the object is relatively obscured. The mean for controlsubjects was 21.6 (SD = 2.8, range: 16.7–25).
In the Item Match Task (Test 11), the target presentsan object from some semantic category, and the probepictures represent the same object or a different objectfrom the same category. For example, the target pictureof a tiger’s head is presented with the probe pictures ofa tiger viewed from the side and a polar bear viewedfrom the side. Although Riddoch and Humphreys (1993)claim that this task cannot be done on the basis of visualfeatures, this claim is easily questionable (e.g., one couldrely on the stripes of the tiger to match the tiger pic-tures). We view this task as another measure of visualprocessing and visual feature matching rather than ameasure of access to stored knowledge.
In the Associative Match Task (Test 12), the targetpresents an object that is associated with one of the twoprobe pictures. For example, the target of a car is pre-sented with the probe pictures of a road and railroadtracks. The mean for control subjects was 27.5 (SD = 2.4,range: 21–30). There are �ve animal pictures on this testand E.W.’s two errors were on two animal trials. For thepicture of the milk bottle, E.W. picked the sheep ratherthan the cow and for the picture of the egg, E.W. pickedthe owl rather than the chicken. These are clearly seman-tic association errors that appear to result from herde�cit comprehending information about animals.
Development and Analysis of New Attribute QuestionTasks
A number of attribute question tasks were developed toaddress speci�c questions regarding E.W.’s category-spe-ci�c knowledge de�cit. For each of these, �ve elderly
control subjects completed each task in two ways. Sub-jects were �rst told to decide whether or not the state-ment about an object was true (e.g., bears like honey;bears are bred for wool; gloves have a space for each�nger; gloves are worn on the head). After making theirdecision, subjects were asked to rate how con�dent theywere that they knew the answer to the statement on ascale of 1 to 5, with 1 representing “just guessing” and5 representing “extreme con�dence.” This was done toexamine familiarity of the question being asked aboutan item. Items were matched as closely as possible onfamiliarity of the concept itself, as determined from theSnodgrass and Vanderwart (1980) norms and/or the typi-cality ranking from the Battig and Montague (1969) cate-gory norms. Items that were unclear to the controlsubjects and/or were judged to be correct even thoughthey were created to be incorrect (e.g., elephants arebrown) were removed from the stimulus set.
Both the control subjects and E.W. completed all theattribute questions associated with each item in eachattribute judgment task. For each experiment task devel-opment is described accordingly. Data presented in Ta-bles 6 through 10 represent a subset of the attributequestions. For each task, items in the speci�c compari-son groups were matched one-to-one on dif�culty levelof the attribute, as determined by the familiarity ratingsgiven by the control subjects. For example, a comparisonof the perceptual attributes associated with their respec-tive animal and object concept is relevant to the inves-tigation. Thus, attribute questions in each of these cellsare matched perfectly for dif�culty level. Data are pre-sented this way to control for the in�uence of familiarityor dif�culty on the level of performance; however, itshould be noted that the pattern of performance is
Figure 2. Examples of itemsused in the Part Decision task.Each item was presented sepa-rately.
26 Journal of Cognitive Neuroscience Volume 10, Number 1

identical for both the matched data set and the wholedata set. To keep the number of items in each cell at areasonable level, we could not match attributes perfectlyfor every speci�c comparison. Note, however, that thereis little difference in dif�culty level between all the cells(e.g., perceptual animate and associative inanimate).
For the �rst attribute judgment task, we present databroken down by true and false statements. This was doneto demonstrate the lack of response bias for E.W.; thatis, she performs equally well (or equally poor) on trueand false statements. To simplify presentation of the datafor the rest of the tasks, performance is collapsed acrosstrue and false statements. E.W.’s performance was alwaysexamined for any indication of a response bias, but therewas little suggestion of this in any of the judgment tasks.
Attribute Property Task 1: Central Attributes. Forty ani-mals and 46 inanimate items (representing fruits, vegeta-bles, clothing, and tools) were selected and between 1and 15 attributes about each item was determined. Eachitem and attribute speci�c to the item were given to 10undergraduate students, who were asked to judgewhether or not an attribute was characteristic or impor-tant in de�ning that concept. From these judgments,three to six attributes that the largest number of subjectsagreed to be important in de�ning the concept wereused. Matching trials consisted of each attribute pairedcorrectly with the corresponding object, and nonmatch-ing trials consisted of attributes pseudorandomly as-signed to other items. Matching of items resulted in 98visual questions each for animals and objects and 57associative questions each for animals and objects.
Attribute Property Task 2: Food/Nonfood Animals.Nine food animals (chicken, cow, crab, lobster, pig, rabbit,salmon, tuna, turkey) and nine nonfood animals wereselected (dog, horse, octopus, pigeon, raccoon, robin,shark, turtle, whale) to serve as items in this task (foodand nonfood animals were considered as such for hergeneration). Six visual attributes and six associative/func-tional attributes were determined for each item based oninformation collected from normal undergraduates re-garding the attributes most associated with speci�c items(from the task described above). Nonmatching itemswere created by pseudorandomly assigning attributes toother items. Five control subjects completed this task.Matched items resulted in 41 questions each for food andnonfood animals in both the visual and associative cells,respectively.
Attribute Property Task 3: General/Speci�c PropertyJudgments. Sixty-�ve animals (15 each for birds, insects,sea animals; 20 mammals) and 45 objects (15 each forfruits, vegetables, buildings) were selected and werematched as best as possible on familiarity (not all itemshad familiarity ratings because they are not all repre-sented in the Snodgrass and Vanderwart set). Items
within each category (e.g., birds, fruit) were the �rst 15items listed in the Battig and Montague (1969) categorymembership norms. The attributes that were common toall members of a category (e.g., all animals or all build-ings) and attributes that were common to only somemembers of a category are summarized in Table 11. Be-cause of the number of trials (n = 1165), an equal num-ber of true and false statements were not created.Instead, each attribute was asked of each item in a cate-gory. The table shows the attributes that were queriedfor each item in each of the four categories tested.Matched items for speci�c attributes resulted in 100visual/perceptual questions for both animate and inani-mate items and 150 associative/functional questions forboth animate and inanimate items.
Attribute Property Task 4a: Size Judgments. Thirty ani-mate pairs and 30 inanimate pairs were developed. Onemember of the pair was judged to be larger than thesecond member of the pair, although the distinctionrequired a �ne discrimination (e.g., goat-raccoon; tomato-pea). Four control subjects completed this task. Matcheditems resulted in 18 items in the animate and inanimatecategories, respectively.
Attributes Property Task 4b: Other Shared Attributes.Seven attributes that are “shared” between animate andinanimate objects were selected (big, small, number oflegs, surface texture, short, tall, color) and 12 items thathad one of these features were paired with an item nothaving this feature, resulting in 42 trials in the animatecondition and 42 trials in the inanimate object. Matcheditems resulted in 42 questions in the animal and objectcategories, respectively.
Attributes Property Task 5: Visible and Not VisibleAttributes. Twenty animals were selected and two tofour pictures of these animals were developed. Eachpicture represented a different view of the animal suchthat only in one of the pictures showed the experimen-tally relevant visual attributes of the animal represented.For example, one picture of a horse might have all fourlegs, a tail, and a mane, another picture of a horse mightonly be a side view of the head. Attribute questions weredeveloped for each animal picture that were either rep-resented in the picture or were not represented in thepicture. This resulted in 95 attribute questions that wereanswerable based on the information in the picture and65 attribute questions that were not answerable basedon the information in the picture. Each animal was testedonly once in a session. E.W. was shown a picture of theanimal (e.g., horse) and told, “This is a picture of a horse.I’m going to ask you some questions about horses andyou can use the information in the picture to answerthem.” She was then questioned about attributes associ-ated with a horse (e.g., number of legs, tail, mane).
Caramazza and Shelton 27
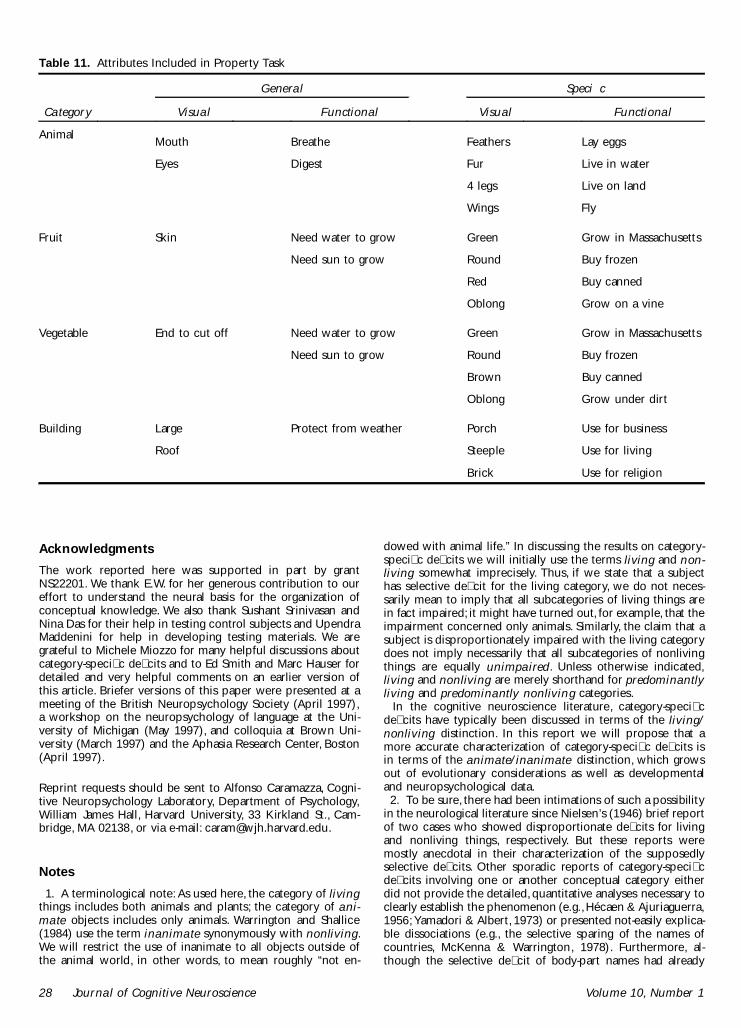
Acknowledgments
The work reported here was supported in part by grantNS22201. We thank E.W. for her generous contribution to oureffort to understand the neural basis for the organization ofconceptual knowledge. We also thank Sushant Srinivasan andNina Das for their help in testing control subjects and UpendraMaddenini for help in developing testing materials. We aregrateful to Michele Miozzo for many helpful discussions aboutcategory-speci�c de�cits and to Ed Smith and Marc Hauser fordetailed and very helpful comments on an earlier version ofthis article. Briefer versions of this paper were presented at ameeting of the British Neuropsychology Society (April 1997),a workshop on the neuropsychology of language at the Uni-versity of Michigan (May 1997), and colloquia at Brown Uni-versity (March 1997) and the Aphasia Research Center, Boston(April 1997).
Reprint requests should be sent to Alfonso Caramazza, Cogni-tive Neuropsychology Laboratory, Department of Psychology,William James Hall, Harvard University, 33 Kirkland St., Cam-bridge, MA 02138, or via e-mail: [email protected].
Notes
1. A terminological note: As used here, the category of livingthings includes both animals and plants; the category of ani-mate objects includes only animals. Warrington and Shallice(1984) use the term inanimate synonymously with nonliving.We will restrict the use of inanimate to all objects outside ofthe animal world, in other words, to mean roughly “not en-
dowed with animal life.” In discussing the results on category-speci�c de�cits we will initially use the terms living and non-living somewhat imprecisely. Thus, if we state that a subjecthas selective de�cit for the living category, we do not neces-sarily mean to imply that all subcategories of living things arein fact impaired; it might have turned out, for example, that theimpairment concerned only animals. Similarly, the claim that asubject is disproportionately impaired with the living categorydoes not imply necessarily that all subcategories of nonlivingthings are equally unimpaired. Unless otherwise indicated,living and nonliving are merely shorthand for predominantlyliving and predominantly nonliving categories. In the cognitive neuroscience literature, category-speci�cde�cits have typically been discussed in terms of the living/nonliving distinction. In this report we will propose that amore accurate characterization of category-speci�c de�cits isin terms of the animate/inanimate distinction, which growsout of evolutionary considerations as well as developmentaland neuropsychological data. 2. To be sure, there had been intimations of such a possibilityin the neurological literature since Nielsen’s (1946) brief reportof two cases who showed disproportionate de�cits for livingand nonliving things, respectively. But these reports weremostly anecdotal in their characterization of the supposedlyselective de�cits. Other sporadic reports of category-speci�cde�cits involving one or another conceptual category eitherdid not provide the detailed, quantitative analyses necessary toclearly establish the phenomenon (e.g., Hécaen & Ajuriaguerra,1956; Yamadori & Albert, 1973) or presented not-easily explica-ble dissociations (e.g., the selective sparing of the names ofcountries, McKenna & Warrington, 1978). Furthermore, al-though the selective de�cit of body-part names had already
Table 11. Attributes Included in Property Task
General Speci�c
Category Visual Functional Visual Functional
AnimalMouth Breathe Feathers Lay eggs
Eyes Digest Fur Live in water
4 legs Live on land
Wings Fly
Fruit Skin Need water to grow Green Grow in Massachusetts
Need sun to grow Round Buy frozen
Red Buy canned
Oblong Grow on a vine
Vegetable End to cut off Need water to grow Green Grow in Massachusetts
Need sun to grow Round Buy frozen
Brown Buy canned
Oblong Grow under dirt
Building Large Protect from weather Porch Use for business
Roof Steeple Use for living
Brick Use for religion
28 Journal of Cognitive Neuroscience Volume 10, Number 1

been clearly documented (e.g., Dennis, 1976; Selecki & Herron,1965; Weinstein, 1964), discussions of this de�cit did not spe-ci�cally focus on the possibility that the semantic system mightbe organized categorically. Instead, body-part naming de�citswere typically discussed in relation to disturbances of the“body image”—autotopagnosia (De Renzi & Scotti, 1970)—and,consequently, may have been seen as a nontypical semanticdomain. Thus, it wasn’t until Warrington and Shallice’s (1984)paper that a compelling empirical basis was provided for thepossibility that the semantic system might be organized cate-gorically. 3. Over the past several years there have been a number ofreports of category-speci�c de�cits in patients with degenera-tive brain disease (see, for example, Gonnerman, Andersen,Devlin, Kempler, & Seidenberg, 1997; Silveri, Daniele, Giustolisi,& Gainotti, 1991). However, most of these reports did notcarefully control for various nuisance variables such as differ-ences in familiarity and visual complexity across semantic cate-gories (see below). Furthermore, the results are not obtainedreliably, and whenever a category-like effect is obtained, it israther small by comparison to the effects obtained in the casesreviewed here. We will not discuss these reports. 4. There has also been a different sort of reluctance againstadopting the proposal that the semantic system might be or-ganized categorically. It has been voiced most clearly by Farahand her collaborators (Farah & McClelland, 1991; Farah et al.,1996) and represents a basic tenet of associationistic accountsof cognition. They have argued that the adoption of the viewthat semantic knowledge is organized categorically in the brainwould represent “a signi�cant departure from everything elsewe know about brain organization, according to which subsys-tems are delineated by function or modality but not by seman-tic content” (Farah et al., 1996; p. 143). Despite the baldnessof this assertion, there is hardly any evidence that speaksdirectly to the issue of how “semantic content” is representedin the brain. And, in fact, the authors do not provide anyrelevant citations to empirical evidence that would supporttheir contention. We will discuss recent neuroimaging resultsthat might be relevant to this issue later in the paper. 5. This does not imply that retrieval of nonvisual propertiesfor living things would remain unaffected by damage to visualproperties. On the contrary, damage to any de�nitionally im-portant subset of the semantic properties of objects will affectretrieval of other semantic properties. Thus, damage to thevisual semantic subsystem is also expected to affect retrievalof the functional properties of living things but to a lesserextent. See text below for more detailed discussion. 6. There are many other studies of category-speci�c de�citthan are considered in this section of our paper. However,because these studies did not explicitly mention the categoriesunder consideration here, we are unable to unambiguouslydetermine whether the relevant association/dissociation wasobtained. For example, Basso et al. (1988), Farah et al. (1989,1991), Pietrini et al. (1988), Sartori and Job (1988) do not reportperformance for foods. 7. We consider later Farah and McClelland’s (1991) effort toempirically support the (somewhat different) claim that themembers of the categories of living and nonliving things de-pend differentially on visual and functional attributes for theirmeanings. 8. In the literature on category-speci�c de�cits, the notion of“functional” property is used synonymously with “nonsensory”property. For example, the typical habitat of an animal wouldbe part of the functional properties of that animal. 9. Although the Sartori and Job (1988) are often cited asshowing an association of category- and modality-speci�c se-mantic de�cits, this attribution may be inaccurate. The investi-gation of subject Michelangelo by Sartori and Job focused
principally on his ability to process pictured objects. And, infact, the authors argue that Michelangelo’s de�cit may havebeen restricted to the processing of “structural descriptions”for recognizing objects and not the semantic system itself.Nevertheless, Michelangelo was not consistently better able toprocess visual as opposed to functional aspects of the mean-ing of living things. For example, he was 100% correct injudging which of two animals was the larger one.10. There are additional dif�culties with this study that makeits interpretation highly problematic. To test whether the sub-ject, L.A., had greater dif�culty processing the visual or thefunctional attributes of living category items, the authors useda naming-to-de�nition task. The de�nitions stressed either thevisual or the functional properties of an animal. However, theanimals used in the visual-emphasis de�nitions were harder toname than those used in the function-emphasis de�nitions, asindicated by L.A.’s performance in an independent naming task(Exp. C). Thus, the alleged disproportionate dif�culty with vis-ual as opposed to functional attributes of living things (Exp. D)cannot be meaningfully interpreted. Gainotti and Silveri (1996)acknowledge this problem and report the results of new ex-periments that are supposed to be free of interpretationdif�culties. These new results are discussed in the main text.11. It should be noted that this pattern of performance—dif�culty in processing only visual attributes of animals—is notcompatible with the Farah and McClelland (1991) version ofthe SFT. This version of the theory predicts disproportionatedif�culty for visual versus functional attributes but does notpredict complete sparing of functional judgments in the faceof severe naming dif�culty for living things.12. The point here is that K.R.’s performance is not explicableby assuming damage to the semantic system, because a conse-quence of such damage would presumably be the productionof nonsystematic errors. The systematicity of the errors mightsuggest some type of “psychiatric” disturbance. None of thismeans that the case is less interesting. It does, however, raisethe question of whether it is relevant to the issues underconsideration here.13. And even if one were to ignore these problems, there isthe additional dif�culty that the SFT does not predict justpoorer performance in processing visual attributes for theliving things category but also for the nonliving things category.This prediction clearly follows from the hypothesis that theunderlying cause of the category-speci�c de�cit is damage toa visual semantic subsystem that is presumably shared by allconcepts. Consequently damage to this subsystem should alsodisproportionately impair performance for visual attributes ofnonliving concepts (though not to the same degree). However,this interaction has not been systematically observed (e.g.,Gainotti & Silveri, 1996). Thus, it does not appear that theinvestigation of differential performance for visual and func-tional attributes of concepts has produced a coherent patternof results on which to base the SFT.14. The two subjects studied in the Laiacona, Barbarotto, &Capitani (1993) paper were trauma cases, raising the possibilitythat aetiology may play a role in determining whether or notknowledge of visual and functional attributes are differentiallyimpaired in subjects with category-speci�c de�cits. This possi-bility is excluded by their more recent study (Laiacona et al.,1997) in which they report two new herpes simplex encepha-litis cases with clear category-speci�c de�cits for living thingsbut who, like the trauma cases, show equal levels of perfor-mance for knowledge of visual and functional attributes.15. Shallice (personal communication, April 1997) has alsoobserved that J.B.R. was equally impaired in processing visualand functional attributes of living and nonliving things.16. The notion of modality-speci�c de�cit is multiply ambigu-ous. One sense of modality-speci�c refers to a selective de�cit
Caramazza and Shelton 29

of the modality of input or output; that is, the modality ofpresentation of the stimulus or the modality used for produc-tion of a response. In this sense, a modality-speci�c de�citmight refer to a subject who, for example, is disproportionatelyimpaired in processing auditorially presented stimuli. Anothersense of modality-speci�c refers to the type of stimulus orresponse. In this sense, pictures and written words are differentmodalities—visual and verbal, respectively—even though bothare presented visually. The third sense of modality-speci�crefers to a type of knowledge. In this latter sense, modalityrefers to the type of properties of objects. Thus, for example, adistinction is drawn between the visual and functional (verbal)attributes of objects. Researchers have not always been carefulto distinguish among these senses of modality-speci�c. Unlessotherwise indicated here, we will use this term to refer to atype of knowledge.17. The notion of “denseness” discussed here is to be distin-guished from the unrelated notion referring to the amount ofinformation one has about a concept—we have lots of infor-mation about some things and only sparse knowledge aboutothers. The latter notion of denseness is also likely to �gureinto the probability that a concept will be adversely affectedby brain damage. And it may even play a role in the possibilitythat semantic de�cits may be categorical in nature, in thesense that categories for which we only have little knowledgeare more likely to be disproportionately affected by braindamage.18. It is important to note that the emphasis being placed hereon domain-speci�c knowledge systems as the result of evolu-tionary adaptations is not necessarily at odds with the OUCHand SFT hypotheses of category-speci�c de�cits. It may turnout that the two types of theories provide explanations of thesame phenomenon at different levels of analysis. We explorethis possibility in the “Discussion” section.19. Note that the claim is not that the only types of category-speci�c de�cits that will be observed will involve the catego-ries of animals, plants, and artifacts. Other category-speci�ctypes of de�cits may be observed, but these will have, byhypothesis, different causes from those for the major categorieslisted above. We return to this issue in the “Discussion” section.20. Once again, however, as we shall see in the text below, thisaccount of category-speci�c de�cits does not preclude thepossibility of selective de�cits restricted to particular modali-ties of access, such as visual object recognition versus wordcomprehension.21. E.W. was administered two other object decision tasks. Inboth tasks she performed very poorly with animals. On Test 10of the BORB (Riddoch & Humphreys, 1993), she scored 70%correct for animals versus 94% correct for nonanimals. Onanother test with 36 items, of which half were real animals andthe other half were unreal animals created by combining twohalves of different animals, she scored 72% correct.22. Although the terms general and speci�c attributes arerelated to the concepts of “de�ning” and “characteristic” fea-ture (Smith, Shoben, & Rips, 1974), we use these terms some-what more loosely to capture the fact that some properties aremuch more likely to be shared by members of a category,whereas others may be speci�c to one or only a few membersof a category. Furthermore, the general attributes may notcorrespond to the technically important features (e.g., egg-laying, biped) of a category but to the perceptually and cultur-ally salient ones.23. The notions of “shared” or “same” attributes across catego-ries must not be given undue theoretical importance at thispoint. That is, by using these notions we do not wish to commitourselves to the view that meanings are built up from a set ofsemantic primitives that are shared across semantic domains.It may turn out that categories share very few if any attribute
concepts. Thus, for example, the attribute “has wings” (or “hasfour legs,” “has texture X,” etc.), which may be claimed to beshared across animate and inanimate categories may, in fact,correspond to different attribute concepts in the two semanticdomains. That is, perhaps there is no abstract attribute “haswings” that is shared by, say, birds and airplanes; the attribute“has wings” in the domain of animals might be a differentconcept altogether from the homonymous “has wings” in thedomain of vehicles.24. Farah and McClelland (1991) note that damage to thevisual semantic component of their network model also leadsto some de�cit on functional questions. This result, too, isunsurprising given that the meaning of a word depends on thewhole set of properties that de�ne it.25. We thank Martha Farah for making the de�nitions availableto us.26. We tested 16 subjects. Eight subjects were asked to under-line all sensory/perceptual descriptors and the other eightsubjects were asked to underline all nonsensory (functional/associative) descriptors for each de�nition of the set of wordsused by Farah and McClelland (1991).27. It is important to emphasize that if Farah and McClelland(1991) had intended all along to restrict the term functionalto precisely the sense of “having a function,” an equally seriouscriticism would be raised against the computer simulation. Forin this case the dichotomy sensory-functional would fail tocapture the bulk of the nonsensory information we have aboutanimals (as argued in the text above). And since functionalproperties (in the strict sense) do not play a signi�cant role indistinguishing among animals, the computer demonstrationwould have left out just the important nonsensory informationwe have about animals. That the narrow sense of functional is not the sense in-tended by proponents of the SFT is clearly indicated by thefact that in experiments in which they investigate the relativeloss of visual and “functional” properties, they have includedproperties such as carnivore, lives in the desert, etc. (see, forexample, Farah et al., 1989). These properties are not functionalin the strict sense of “having a function.”28. That is, those category-speci�c de�cits that are not theresult of uncontrolled factors such as familiarity, frequency,complexity, or other idiosyncratic factors concerning the par-ticular histories of individuals with various categories of knowl-edge.29. There are reasons for being skeptical of Damasio et al.’sinterpretation of the functional basis for their patients’ cate-gory-speci�c naming de�cits. They argue that the functionallocus of de�cit in their patients is at the level of lexical selec-tion and not at the semantic level. They support this claim bynoting that they included for analysis only those naming fail-ures for which it could be ascertained that the patient had“correctly recognized” the to-be-named item. The criterion forrecognition was that the patient either named the item cor-rectly or provided an adequate description. Thus, they rea-soned, if a patient shows disproportionate naming dif�culty fora particular semantic category, it could be concluded that thisdif�culty must arise at a level of processing beyond the seman-tic level since the to-be-named items were presumably cor-rectly recognized. The authors identify this postsemantic stageof processing as the level at which lexical selection occurs.There are several problems with this conclusion. The authors do not report evidence that would allow in-ferences about the integrity of semantic processing in theirpatients. Thus, for example, no data are reported on compre-hension performance; the only data reported are those con-cerning naming performance. This is unfortunate because toour knowledge all the cases of semantic category-speci�cde�cit in the literature have been reported to have had a de�cit
30 Journal of Cognitive Neuroscience Volume 10, Number 1

in semantic processing. This is certainly the case for patientswith herpes simplex encephalitis, who have all been shown tohave had both naming and comprehension category-speci�cde�cits. And, presumably, this would also have been true forthe �ve cases of herpes simplex encephalitis included in thesample of patients tested by Damasio et al. (1996) (who prob-ably were disproportionately impaired in naming animals). Inother words, it seems that the criteria used by the authors forestablishing that the relevant feature of their patients’ namingperformance is a de�cit restricted to mechanisms of lexicalaccess (as opposed to semantic representations) may havebeen inadequate. The latter problem is recognized by the authors, but they goon to argue: “Our scoring procedure ensures that the defectsreported here pertain to word retrieval rather than to recogni-tion impairments, even in subjects who showed defects in bothnaming and recognition” (Damasio et al., 1996; p. 500). Thisargument is based on the reasoning that the lexical retrievalfailure observed in the patients is in addition to whateverother dif�culty they may have had in recognizing speci�cto-be-named items. But this reasoning is suspect. Without otherinformation, we do not know whether it is only those patientswho showed disproportionate recognition/comprehensionfailure for the items in a speci�c category who also showeddisproportionate naming failures in that category. Were this thecase, we would have grounds for supposing that the locus offunctional de�cit in these patients probably concerned thesemantic level. The mere fact that a patient provides a descrip-tion of an item that cannot be named does not imply that thebasis for the naming failure does not reside at the semanticlevel of processing. That is, there is little indication that thedescriptions provided by the patients were unambiguous.Taken out of context, the “appropriate” descriptions actuallymay not clearly describe the to-be-named item. There are other problems of interpretation with the resultsof this study. Damasio et al. (1996) report that they includedfor analysis only those patients who “were able to recognizeat least 50% of items in each category” (p. 500). Again, withoutfurther information it is dif�cult to interpret the authors’ re-sults because the naming data for different subjects could bebased on radically different subsets of test items, depending onwhich items they failed to recognize. Another problem concerns the method used to determinethat a patient had a category-speci�c de�cit. A patient wasclassi�ed as having a category-speci�c de�cit if naming perfor-mance was more than two standard deviations below the meanfor the normal controls. This means that it is possible that apatient who scored 1.95 SD below the mean for one category,but 2.05 SD below the mean for another category would havebeen classi�ed as having a category-speci�c de�cit for thelatter category. Unfortunately, unlike other reports of category-speci�c de�cits, the authors do not provide a direct compari-son of performance between categories for each patient withputative category-speci�c de�cits. Furthermore, when theproblems introduced by this criterion are considered inthe context of the scoring procedure used by the authors (theexclusion of an unequal number of trials from different cate-gories for different subjects due to recognition failure), it be-comes extremely dif�cult to compare the putative cases ofcategory-speci�c de�cit reported by Damasio et al. (1996) withother cases in the literature. And, certainly, we cannot becon�dent that their patients represent cases of category-speci�c lexical as opposed to semantic processing de�cit.30. This speculation may provide a basis for the functionalimaging results that show extrastriate activation for the cate-gory of animals. However, on the negative side, it must be notedthat, to our knowledge, there are no cases of category-speci�cagnosias for the categories of animate and inanimate catego-
ries. Such de�cits ought to be observed if there were categori-cal specialization early in the object recognition system. Per-haps a possible case demonstrating this is the patient reportedby Sartori and Job (1988).
REFERENCES
Albert, M. S., Butters, N., & Levin, J. (1979). Temporal gradi-ents in the retrograde amnesia of patients with alcoholicKorsakoff’s disease. Archives of Neurology, 36, 211–216.
Allport, D. A. (1985). Distributed memory, modular subsys-tems and dysphasia. In S. K. Newman & R. Epstein (Eds.),Current perspectives in dysphasia (pp. 32–60). Edin-burgh: Churchill Livingstone.
Atran, S. (1995). Causal constraints on categories and cate-gorical constraints on biological reasoning across cultures.In D. Sperber, D. Premack, & A. J. Premack (Eds.), Causalcognition: A multidisciplinary debate (pp.205–233). NewYork: Oxford University Press.
Baillargeon, R. (1993). The object concept revisited: New di-rection in the investigation of infants’ physical knowledge.In C. Granrud (Ed.), Visual perception and cognition ininfancy. Carnegie Mellon Symposia on Cognition(pp. 265–315). Hillsdale, NJ: Erlbaum.
Basso, A., Capitani, E., & Laiacona, M. (1988). Progressive lan-guage impairment without dementia: A case with isolatedcategory speci�c semantic defect. Journal of Neurology,Neurosurgery, and Psychiatry, 51, 1201–1207.
Battig, W. F., & Montague, W. E. (1969). Category norms forverbal items in 56 categories: A replication and extensionof the Connecticut category norms. Journal of Experimen-tal Psychology Monographs, 80, (3, part 2).
Bertenthal, B. I. (1993). Infants’ perception of biomechanicalmotions: Intrinsic image and knowledge-based constraints.In C. Granrud (Ed.), Visual perception and cognition ininfancy. Carnegie Mellon Symposia on Cognition(pp. 175–214). Hillsdale, NJ: Erlbaum.
Bertenthal, B. I., Prof�tt, D. R., & Cutting, J. E. (1984). Infantsensitivity to �gural coherence in biomechanical motions.Journal of Experimental Child Psychology, 37, 213–230.
Bloom. P. (1996). Intention, history, and artifact concepts. Cog-nition, 60, 1–29.
Caramazza, A., Hillis, A. E., Rapp, B. C., & Romani, C. (1990).The multiple semantics hypothesis: Multiple confusions?Cognitive Neuropsychology, 7, 161–189.
Carey, S. (1985). Conceptual change in childhood. Cam-bridge, MA: Bradford/MIT Press.
Carey, S. (1995). On the origin of causal understanding. InD. Sperber, D. Premack, & A. J. Premack (Eds.), Causal cog-nition: A multidisciplinary debate (pp. 268–302). NewYork: Oxford University Press.
Carey, S., & Spelke, E. (1994). Domain-speci�c knowledgeand conceptual change. In L. A. Hirschfeld & S. A. Gelman(Eds.), Mapping the mind: Domain speci�city in cogni-tion and culture (pp. 169–200). New York: Cambridge Uni-versity Press.
Chomsky, N. (1980). Rules and representations. New York:Columbia University Press.
Chomsky, N. (1986). Knowledge of language: Its nature,origin, and use. New York: Praeger.
Damasio, H., Grabowski, T. J., Tranel, D., Hichwa, R. D., &Damasio, A. R. (1996). A neural basis for lexical retrieval.Nature, 380, 499–505.
Dennis, M. (1976). Dissociated naming and locating of bodyparts after left anterior temporal lobe resection: An experi-mental case study. Brain and Language, 3, 147–163.
Caramazza and Shelton 31

De Renzi, E., & Lucchelli, F. (1994). Are semantic systemsseparately represented in the brain? The case of living cate-gory impairment. Cortex, 30, 3–25.
De Renzi, E., & Scotti, G. (1970). Atotopagnosia: Fiction or re-ality? Archives of Neurology, 23, 221–227.
Farah, M. J., Hammond, K. M., Mehta, Z., & Ratcliff, G. (1989).Category-speci�city and modality-speci�city in semanticmemory. Neuropsychologia, 27, 193–200.
Farah, M. J., & McClelland, J. L. (1991). A computationalmodel of semantic memory impairment: Modality spe-ci�city and emergent category speci�city. Journal of Ex-perimental Psychology: General, 120, 339–357.
Farah, M. J., McMullen, P. A., & Meyer, M. M. (1991). Can rec-ognition of living things be selectively impaired? Neuropsy-chologia, 29, 185–193.
Farah, M. J., Meyer, M. M. & McMullen, P. A. (1996). The liv-ing/nonliving dissociation is not an artifact: Giving ana priori implausible hypothesis a strong test. CognitiveNeuropsychology, 13, 137–154.
Farah, M. J., & Wallace, M. A. (1992). Semantically-boundedanomia: Implications for the neural implementation ofnaming. Neuropsychologia, 30, 609–621.
Funnell, E., & De Mornay Davies, P. (1997). JBR: A reassess-ment of concept familiarity and a category-speci�c disor-der for living things. Neurocase, 2, 461–474.
Funnell, E., & Sheridan, J. S. (1992). Categories of knowledge?Unfamiliar aspects of living and nonliving things. Cogni-tive Neuropsychology, 9, 135–153.
Gaffan, D., & Heywood, C. A. (1993). A spurious category-speci�c visual agnosia for living things in normal humansand nonhuman primates. Journal of Cognitive Neurosci-ence, 5, 118–128.
Gainotti, G., & Silveri, M. C. (1996). Cognitive and anatomicallocus of lesion in a patient with a category-speci�c seman-tic impairment for living beings. Cognitive Neuropsychol-ogy, 13, 357–389.
Gainotti, G., Silveri, M. C., Daniele, A., & Giustolisi, L. (1995).Neuroanatomical correlates of category-speci�c semanticdisorders: A critical survey. In R. A. McCarthy (Ed.), Seman-tic knowledge and semantic representations: Memory,Vol. 3, Issues 3 & 4 (pp. 247–264). Hove, England:Erlbaum, Taylor & Francis.
Gelman, R. (1990). First principles organize attention to andlearning about relevant data: Number and the animate-inanimate distinction as examples. Cognitive Science, 14,79–106.
Gelman, R., & Brenneman, K. (1994). First principles can sup-port both universal and culture-speci�c learning aboutnumber and music. In L. A. Hirschfeld & S. A. Gelman(Eds.), Mapping the mind: Domain speci�city in cogni-tion and culture (pp. 369–390). New York: CambridgeUniversity Press.
Gelman, R., Durgin, F., & Kaufman, L. (1995). Distinguishingbetween animates and inanimates: Not by motion alone.In D. Sperber, D. Premack, & A. J. Premack (Eds.), Causalcognition: A multidisciplinary debate (pp. 150–184). NewYork: Clarendon Press/Oxford University Press.
Gelman, S., & Coley, J. D. (1990). The importance of knowinga dodo is a bird: Categories and inferences in 2-year-oldchildren. Developmental Psychology, 26, 796–804.
Gonnerman, L. M., Andersen, E. S., Devlin, J. T., Kempler, D.,& Seidenberg, M. S. (1997). Double dissociation of seman-tic categories in Alzheimer’s disease. Brain and Lan-guage, 57, 254–279.
Goodglass, H., & Kaplan, E. (1983). The assessment of apha-sia and related disorders, 2nd ed. Philadelphia, PA: Lea &Febiger.
Hart, J., Berndt, R. S., & Caramazza, A. C. ( 1985). Category-speci�c naming de�cit following cerebral infarction. Na-ture, 316, 439–440.
Hart, J., & Gordon, B. (1992). Neural subsystems for objectknowledge. Nature, 359, 60–64.
Hauser, M. D. (in press). The functional properties of artifacts:A nonhuman primate’s view. Cognition.
Hécaen, H., & Ajuriaguerra de, J. (1956). Agnosie visuellepour les objets inanimes par lesion unilaterale gauche. Re-vue Neurologique, 94, 222–223.
Hillis, A. E., & Caramazza, A. (1991). Category-speci�c namingand comprehension impairment: A double dissociation.Brain, 114, 2081–2094.
Hillis, A. E., Rapp. B., & Caramazza, A. (1995). Constrainingclaims about theories of semantic memory: More on uni-tary versus multiple semantics. Cognitive Neuropsychol-ogy, 12, 175–186.
Hirschfeld, L. A., & Gelman, S. A. (Eds.) (1994). Mapping themind: Domain speci�city in cognition and culture. NewYork: Cambridge University Press.
Humphreys, G. W., & Riddoch, M. J. (1987). On telling yourfruit from your vegetables: A consideration of category-speci�c de�cits after brain damage. Trends in Neurosci-ence, 4, 145–148.
Keil, F. C. (1989). Concepts, kinds, and cognitive develop-ment. Cambridge, MA: MIT Press.
Keil, F. C. (1994). The birth and nurturance of concepts bydomains: The origins of concepts of living things. In L. A.Hirschfeld & S. A. Gelman (Eds.), Mapping the mind: Do-main speci�city in cognition and culture (pp. 234–254).New York: Cambridge University Press.
Kurbat, M. A. (1997). Can the recognition of living thingsreally be selectively impaired? Neuropsychologia, 35, 813–827.
Laiacona, M., Barbarotto, R., & Capitani, E. (1993). Perceptualand associative knowledge in category speci�c impair-ment of semantic memory: A study of two cases. Cortex,29, 727–740.
Laiacona, M., Barbarotto, R., Trivelli, C., & Capitani, E. (1993).Category speci�c semantic defects: A standardised testwith normative data. Archivio di Psicologia, Neurologiae Psichiatria, 54, 209–248.
Laiacona, M., Capitani, E., & Barbarotto, R. (1997). Semanticcategory dissociations: A longitudinal study of two cases.Cortex.
Leslie, A. M. (1982). The perception of causality in infants.Perception, 11, 173–186.
Leslie, A. M. (1984). Spatiotemporal continuity and the per-ception of causality in infants. Perception, 13, 287–305.
Leslie, A. M. (1988). The necessity of illusion: Perception andthought in infancy. In L. Weiskrantz (Ed.), Thought with-out language: A Fyssen Foundation symposium (pp. 185–210). Oxford, England: Clarendon Press/Oxford UniversityPress.
Leslie, A. M., & Keeble, S. (1987). Do six-month old infantsperceive causality? Cognition, 25, 265–288.
Liberman, A. M., & Mattingly, I. G. (1989). A specialization forspeech perception. Science, 243, 489–494.
Mandler, J. M. (1992). How to build a baby: II. Conceptualprimitives. Psychological Review, 99, 587–604.
Mandler, J. M. (1994). Precursors of linguistic knowledge.Philosophical Transactions of the Royal Society of Lon-don, 346, 63–69.
Mandler, J. M., Bauer, P. J., & McDonough, L. (1991). Separat-ing the sheep from the goats: Differentiating global catego-ries. Cognitive Psychology, 23, 263–298.
32 Journal of Cognitive Neuroscience Volume 10, Number 1

Mandler, J. M., & McDonough, L. (1993). Concept formationin infancy. Cognitive Development, 8, 291–318.
Markman, E. M. (1989). Categorization and naming in chil-dren: Problems of induction. Cambridge, MA: MIT Press.
Martin, A., Wiggs, C. L., Ungerleider, L. G., & Haxby, J. V.(1996). Neural correlates of category-speci�c knowledge.Nature, 379, 649–652.
McCarthy, R. A., & Warrington, E. K. (1988). Evidence for mo-dality-speci�c meaning systems in the brain. Nature, 334,428–430.
McClelland, J. L., & Rumelhart, D. E. (1986). Parallel distrib-uted processing: Explorations in the microstructures ofcognition. Volume 2: Psychological and biological mod-els. Cambridge, MA: MIT Press.
McKenna, P., & Warrington, E. K. (1978). Category speci�cnaming preservation: A single case study. Journal of Neu-rology, Neurosurgery, and Psychiatry, 43, 781–788.
McRae, K., de Sa, V. R., & Seidenberg, M. S. (1997). On the na-ture and scope of featural representations for word mean-ing. Journal of Experimental Psychology: General, 126,99–130.
Nielsen, J. M. (1946). Agnosia, apraxia, aphasia: Their valuein cerebral localisation. 2nd ed. New York: Harper.
Perani, D., Cappa, S. F., Bettinardi, V., Bressi, S., Gorno-Tempini,M., Matarrese, M., Fazio, F. (1995). Different neural systemsfor the recognition of animals and man-made tools.Neuroreport, 6, 1637–1641.
Pietrini, V., Nertempi. P., Vaglia, A., Revello, M. G., Pinna, V., &Ferro-Milone, F. (1988). Recovery from herpes simplex en-cephalitis: Selective impairment of speci�c semantic cate-gories with neuroradiological correlation. Journal ofNeurology, Neurosurgery, and Psychiatry, 51, 1284–1293.
Pinker, S., & Bloom, P. (1990). Natural language and natural se-lection. Behavioral Brain Sciences, 13, 707–784.
Powell, J., & Davidoff, J. (1995). Selective impairments of ob-jects knowledge in a case of acquired cortical blindness.Memory, 3, 435–461.
Premack, D. (1990). The infant’s theory of self-propelled mo-tion. Cognition, 36, 1–16.
Quinn, P. C., & Eimas, P. D. (1996). Perceptual organizationand categorization in young infants. In C. Rovee-Collier &L. P. Lipsitt (Eds.), Advances in infancy research, Vol. 10(pp. 1–36). Norwood, NJ: Ablex.
Rapp, B. C., Hillis, A. E., & Caramazza, A. C. (1993). The roleof representations in cognitive theory: More on multiplesemantics and the agnosias. Cognitive Neuropsychology,10, 235–249.
Riddoch, M. J., & Humphreys, G. W. (1993). Visual aspects ofneglect dyslexia. In D. W. Willows, R. S. Kruk, & E. Corcos(Eds.), Visual processes in reading and reading disabili-ties (pp. 111–136). Hillsdale, NJ: Erlbaum.
Riddoch, M. J., Humphreys, G. W., Coltheart, M., & Funnell, E.(1988). Semantic systems or system? Neuropsychologicalevidence re-examined. Cognitive Neuropsychology, 5, 3–25.
Rosch, E. (1973). On the internal structure of perceptual andsemantic categories. In T. E. Moore (Ed.), Cognitive devel-opment and the acquisition of language (pp. 111–144).New York: Academic Press.
Rosch, E. (1975). Cognitive representations of semantic cate-gories. Journal of Experimental Psychology: General,104, 192–233.
Rosch, E., Mervis, C. B., Gray, W. D., Johnson, D. M., & Boyes-Braem, P. (1976). Basic objects in natural categories. Cogni-tive Psychology, 8, 382–439.
Rozin, P., & Schull, J. (1988). The adaptive-evolutionary point
of view in experimental psychology. In R. C. Atkinson, R. J.Herrnstein, G. Lindzey, & R. D. Luce (Eds.), Stevens’ hand-book of experimental psychology, Vol. 1: Perception andmotivation; Vol. 2: Learning and cognition (pp. 503–546).New York: Wiley.
Sacchett, C., & Humphreys, G. W. (1992). Calling a squirrel asquirrel but a canoe a wigwam: A category-speci�c de�citfor artifactual objects and body parts. Cognitive Neuropsy-chology, 9, 73–86.
Saffran, E. M., & Schwartz, M. F. (1994). Of cabbages andthings: Semantic memory from a neuropsychological per-spectve—A tutorial review. In C. Umilta & M. Moscovitch(Eds.), Attention and performance 15: Conscious andnonconscious information processing (pp. 507–536).Cambridge, MA: MIT Press.
Sartori, S., & Job, R. (1988). The oyster with four legs: Aneuropsychological study on the interaction of visual andsemantic information. Cognitive Neuropsychology, 5, 105–132.
Sartori, G., Miozzo, M., & Job, R. (1993). Category-speci�cnaming impairments? Yes. Quarterly Journal of Experi-mental Psychology, 46A, 489–504.
Selecki, B. R., & Herron, J. T. (1965). Disturbances of the ver-bal body image: A particular syndrome of sensory aphasia.Journal of Nervous and Mental Disease, 147, 42–52.
Shallice, T. (1988). Specialisation within the semantic system.Cognitive Neuropsychology, 5, 133–142.
Sheridan, J., & Humphreys, G. W. (1993). A verbal-semanticcategory-speci�c recognition impairment. CognitiveNeuropsychology, 10, 143–184.
Silveri, M. C., Daniele, A., Giustolisi, L., & Gainotti, G. (1991).Dissociation between knowledge of living and nonlivingthings in dementia of the Alzheimer’s type. Neurology, 41,545–546.
Silveri, M. C., & Gainotti, G. (1988). Interaction between vi-sion and language in category-speci�c impairment. Cogni-tive Neuropsychology, 5, 677–709.
Sirigu, A., Duhamel, J. R., & Poncet, M. (1991). The role of sen-sorimotor experience in object recognition: A case of mul-timodal agnosia. Brain, 114, 2555–2573.
Skinner, B. F. (1957). Verbal behavior. New York: Appleton-Century-Crofts.
Smith, E. E., Shoben, E. J., & Rips, L. J. (1974). Structure andprocess in semantic memory: A featural model for seman-tic decisions. Psychological Review, 81, 214–241.
Snodgrass, J., & Vanderwart, M. (1980). A standardised set of260 pictures: Norms for name agreement, familiarity, andvisual complexity. Journal of Experimental Psychology:Human Learning and Memory, 6, 174–215.
Spelke, E. S. (1988). The origins of physical knowledge. InL. Weiskrantz (Ed.), Thought without language: A FyssenFoundation symposium (pp. 168–184). Oxford, England:Clarendon Press/Oxford University Press.
Spelke, E. S., Phillips, A., & Woodward, A. L. (1995). Infants’knowledge of object motion and human action. In D. Sper-ber, D. Premack, & A. J. Premack (Eds.), Causal cognition:A multidisciplinary debate (pp. 44–78). New York: Claren-don Press/Oxford University Press.
Sperber, D. (1994). The modularity of thought and theepidemiology of representations. In L. A. Hirschfeld & S. A.Gelman (Eds.), Mapping the mind: Domain speci�city incognition and culture (pp.39–67). New York: CambridgeUniversity Press.
Sperber, D., Premack, D., & Premack, A. J. (1995). Causal cog-nition: A multidisciplinary debate. New York: ClarendonPress/Oxford University Press.
Stewart, F., Parkin, A. J., & Hunkin, N. M. (1992). Naming im-
Caramazza and Shelton 33

pairments following recovery from herpes simplex en-cephalitis: Category-speci�c? Quarterly Journal of Experi-mental Psychology, 44A, 261–284.
Warrington, E. K. (1981). Concrete word dyslexia. BritishJournal of Psychology, 72, 175–196.
Warrington, E. K., & McCarthy, R. (1983). Category speci�caccess dysphasia. Brain, 106, 859–878.
Warrington, E. K., & McCarthy, R. (1987). Categories of
knowledge: Further fractionations and an attempted inte-gration. Brain, 110, 1273–1296.
Warrington, E. K., & Shallice, T. (1984). Category speci�c se-mantic impairments. Brain, 107, 829–854.
Weinstein, S. (1964). De�cits concomittant with aphasia orlesions of either hemisphere. Cortex, 1, 154–169.
Yamadori, A., & Albert, M. L. (1973). Word category aphasia.Cortex, 9, 112–125.
34 Journal of Cognitive Neuroscience Volume 10, Number 1
