anatomy of the tongue and taste buds · 2020-03-29 · 640 chapter 29 anatomy of the tongue and...
TRANSCRIPT

Chapter 29
Anatomy of the Tongue and Taste Buds
Martin Witt and Klaus Reutter
29.1 INTRODUCTION
“… Many papillae are evident, I might say, innumer-able, and the appearance is so elegant that they catch theview and thoughts of the observer, and control him for along time and not without enjoyment … All this delightsthe curious mind, when observing with an engyscope;and if anyone asks to what they are similar, I am unsurewhether I should compare this huge number of papillaefirst with grapes or fruits of the bay, or innumerable mush-rooms emerging between fine, densely standing blades ofgrass …”
These are the first detailed and enthusiastic wordson papillae and their “membranes” in human tongues byLorenzo Bellini (1665) (Figure 29.1), who knew that Mar-cello Malpighi (1628–94) had reported on lingual papillaethe year before. Malpighi discovered mucosal elevationsassociated with nerve fibers, naming these elevations“papillae,” as the morphological substrates for gustatorysensation (Figure 29.2).
The peripheral taste apparatus includes the gustatorysensory organs or taste buds and their innervation, and thespecific papillae within which taste buds are assembled.What one needs to know about taste buds in relation to howone perceives tastants depends on the approach to taste per-ception. One aim of this chapter is to provide a sufficientknowledge on peripheral gustatory anatomy as a basis forunderstanding other chapters of this book. Some structuraldetails about the human peripheral taste system are wellknown, but it is also worthwhile to provide comparativeanatomical information to fill in the gaps or understand andestablish basic principles. The fundamental question of howtastants are perceived has been addressed for more than twomillenia, and the majority of concepts, theories, and exper-imental “proofs” that have been proposed have since given
Handbook of Olfaction and Gustation, Third Edition. Edited by Richard L. Doty.© 2015 Richard L. Doty. Published 2015 by John Wiley & Sons, Inc.
way to present-day concepts. This chapter incorporates thisrich history which sculpted our contemporary views of gus-tatory anatomy and physiology.
Aristotle (384–322 BC), applying Platonic concepts,argued that taste sensation was carried from the tongue viathe blood, to the liver or heart, which was the common seatof the soul and all sense perception [De sensu 438b 26; Onyouth and old age, 469a 5–13; cited after Siegel (1970)].Galen’s (Claudius Galenus, 129–201 AD) anatomicalanalyses challenged this notion: his detailed studies on theinnervation of the tongue describe correctly the differentfunctions of the three principal nerves supplying the tongue(lingual, glossopharyngeal, and hypoglossal nerves), anddemonstrate their origin at the base of the brain. Galenposited that the lingual nerve communicated gustatorysensations, a concept yet resonant in contemporary neuro-biology. One strand of nerve fibers corresponding to cranialnerve IX (CN IX or glossopharyngeal nerve) of presentterminology (rediscovered in humans by Panizza, 1834was already known to Galen as the principal gustatorynerve of the tongue. CN IX also carries some motor fibersto the pharynx. Galen also noted excretory ducts of thelingual and sub-mandibular glands (in ox). The particularstructure of the tongue surface was initially describedby Casserius (1609) and, later by Malpighi (1686) andBellini (1665) (reviewed by Jurisch, 1922. Further evi-dence for the significance of the papillary epithelium andits cells came from observations in taste organs of thefrog (Fixsen, 1857, Waller, 1847, 1849). Taste buds wereidentified initially on the barbels and skin of fishes byLeydig (1851) and described as “becherförmige Organe”(goblet-shaped organs), whose function he associated withtactile sensitivity. Schulze (1863) subsequently suggestedthey were chemosensory structures. Similar organs inmammals were described as Schmeckbecher (taste goblets)
637

638 Chapter 29 Anatomy of the Tongue and Taste Buds
Figure 29.1 The cover sheet of Lorenzo Bellini’s book thatsummarizes examinations on the anatomy of the kidney and the tasteorgan. (Langerack, Leiden, 1711).
Figure 29.2 Depiction of a bovine tongue by Marcello Malpighi(1686) showing “patches” where papillae were observed. Note theconcentration of fungiform papillae (dots) on the tip of the tongue. Thedrawing of vallate and foliate papillae is still rather vague.
(Lovén, 1868) and Geschmacksknospen (taste buds) orGeschmackszwiebeln (“taste onions”) (Schwalbe, 1868).Herrick (1904) translated “Geschmackskospen” as “tastebuds.” Their location within lingual papillae, the latteralready associated with the loci of taste perception, lentcredence to their identity as taste sensor organs.
Nineteenth-century studies focused on cytologicalfeatures, nerve supply (Figures 29.3 and 29.5), and thedevelopment of taste buds. While the nerve-dependentnature of taste sensation was known since Galen’s time, itssignificance for sensory organ physiology blossomed in themid-1800s. With development of the neuron doctrine (seeKoelliker, 1844), previously described “ganglion globules”(Ganglienkugeln: Ehrenberg, 1833) could now be acknowl-edged as part of a specialized cellular system (cell theoryof Schleiden and Schwann) (Schwann, 1839). Two majorprerequisites favored the expansion of scientific knowl-edge in the nineteenth century: (1) replacement of the oftenspeculative natural philosophy by the experimentally basednatural sciences, mainly represented by Francois Magendie(1783–1855) in France and Johannes Müller (1801–58)in Germany, and (2) technical advances which includedimproved microscopes (K. Zeiss, 1816–88, together withErnst Abbé, 1840–1905, and E. Leitz, 1843–1920) as wellas use of histological techniques, for example, introductionof chromic acid for histological examination of neural tis-sue (Hannover, 1840), allowing for the distinction betweencells and fibers. Further, gold chloride staining facilitateddiscriminating finely ramifying nerve fibers (Gerlach,1858). Helmholtz (1842) observed a direct continuitybetween nerve cells (globuli gangliosi) and their fibers inevertebrates. The demand for the neuron doctrine in ver-tebrates was established by Koelliker (1844). Introductionof the methylene blue staining method facilitated observa-tions that fine nerve fibers in the frog taste disc widenedwith varicosities, penetrated the gustatory epithelium andapproached the sensory cells “with extremely sharp smallknobs, [which] … connect taste cells not continuously,but per contiguitatem” (Ehrlich, 1886). This pioneeringobservation of secondary sensory cells in a taste organ thatwas clearly different from primary sensory cells, as hadalready been described in the olfactory epithelium, albeitwithout the understanding of the “contact” nature of tastebud cells and their innervating afferents, as later describedultrastructurally, was opposed by Retzius (1892). Using theGolgi silver impregnation method (Golgi, 1873), Retziusattributed a sensory function only to free nerve endings(Figure 29.5b). Later, Krause (1911) clearly demonstratedthat the finest nerve fibers enter the taste bud and ascend tothe taste pore, but do not merge with taste bud cells whosebasal processes end near the basal cells.
29.2 LINGUAL PAPILLAE ANDTASTE BUD DISTRIBUTION
29.2.1 Nongustatory PapillaeApproximately 60 years before taste buds were identifiedas gustatory organs, an illustration of the human tongue

29.2 Lingual Papillae and Taste BUD Distribution 639
(a) (b)
Figure 29.3 These illustrations of Bourgery and Jacob (1839) show the human tongue including nerve and blood supply, as well as the lingualmuscle system. In spite of precise macroscopical observations on the innervation of glands and muscles, it still represents indirectly the (nowadaysrevised) morphological concept of gustation in the early nineteenth century, according to which the principal taste nerves are the lingual (p in (a) and(b); CN V3) and glossopharyngeal (t in (b); CN IX) nerves. The chorda tympani, although depicted near the submandibular ganglion (q in (a) and(b); CT), does not reach the lingual dorsum. Ebner’s glands are not known to the authors. (a): D, submandibular gland with q, chorda tympani andsubmandibular ganglion and p, lingual nerve; s, hypoglossal nerve (CN XII); E, sublingual gland; F, Nuhn’s gland. (b): C styloid process; b,stylopharyngeus muscle, the leading muscle for t, glossopharyngeal nerve (CN IX), which overlaps partly with the innervation area of the lingualnerve; k, lingual artery; p, lingual nerve, interrupted to show the glossopharyngeal nerve; q, part of the chorda tympani nerve; s, hypoglossal nerve(CN XII); X, trunk of the facial nerve. (See plate section for color version.)
by Sömmering (1806) (Figure 29.4) accurately showedthe regional distribution of lingual papillae. A line calledthe sulcus terminalis (an ontogenetic remnant, see below),which is located posterior to the vallate papillae, separatesthe body of the tongue from the lingual root. It may be seenthat the sulcus terminalis extends laterally to the pharyn-geal wall from the foramen caecum (also an ontogeneticremnant) near the midline (see Figures 29.4 and 29.11).The root of the tongue is covered by a papilla- free smoothepithelium, and beneath this epithelium lie mucous glandsand a reticular connective tissue filled with lymphaticfollicles which lead to the designation of “lingual tonsil.”Ducts of intralingual salivary glands (Ebner, 1873) emptyinto the troughs of vallate and foliate papillae (see below).
The dorsal surface of the tongue is covered with fili-form and conical papillae from the sulcus terminalis to thetongue tip. Filiform papillae are the most prevalent type,while the number of conical papillae may vary. Both typesof papillae are sparse along the lingual margin and abun-dant in the middle regions. Conical papillae have a cylindri-cal base, and taper to a sharp point at their apex. Filiformpapillae (L. filum = thread) have a pyramidal shape, anda narrow tail of cornified cells extending from their apical
tips as a pennant. The fila are part of the fibrous mat on thetongue’s surface in the hypertrophic condition called “hairytongue.”
29.2.2 Gustatory PapillaeTaste buds occur in distinct papillae of the tongue, theepithelium of the palate, oropharynx, larynx (epiglottis),and the upper esophagus. Taste buds of most vertebratesare bulb-shaped structures which are composed of about50–120 bipolar cells (see Figures 29.9 and 29.10). Withthe exception of basal cells, the slender taste bud cells arisefrom an interrupted basal membrane and converge withtheir apical protrusions, the microvilli, into the mucus-filledtaste pit. Together, these cells form the organ’s sensoryepithelium. The nuclei of the cells are located in the lowerthird of the taste bud, which is approximately the regionwhere most afferent nerve fiber terminals are distributed.Sensory cells possess transmembrane receptors and/or ionchannels for specific taste stimuli at apical and lateral por-tions of the cell membrane. Taste buds are demarcated from

640 Chapter 29 Anatomy of the Tongue and Taste Buds
Figure 29.4 The first precise depiction of a human tongue bySamuel Sömmering (1806). (Above) The anterior part (left side) of thetongue shows numerous fungiform papillae. Behind the V-likearrangement of vallate papillae (right side), the lingual tonsils and theintrance to larynx with the epiglottis are visible. (Below) Lateral viewof the tongue shows the left lingual artery and its ramifications intogustatory papillae, which appear as red dots after injection of the arterywith a red dye (observation of Sömmering).
surrounding nongustatory epithelial cells by specializedepithelial cells (marginal cells).
29.2.2.1 Distribution of Lingual Taste Buds.The pattern of taste bud distribution over the tongue sur-face is similar among humans and other mammals. Lingualtaste buds are found exclusively within gustatory papillae,that is, those bearing taste buds. Similar types of gustatorypapillae are located on homologous regions of adult mam-malian tongues. The gustatory papillae include the vallate,foliate, and fungiform papillae. As the term suggests,typical fungiform papillae are mushroom-shaped, with aslender neck and an enlarged head (Figures 29.6–29.8).But the majority of fungiform papillae vary in form andthe filiform papillae are intermingled among them. Shortlyafter the published discovery of taste buds in humans(Lovén, 1868; Schwalbe, 1868), the first systematic inves-tigations on the distribution of human taste buds withinthe oral cavity were carried out by the medical studentHoffmann (1875). He emphasized that taste buds are moresparse within foliate papillae and the soft palate includingthe uvula. Hoffmann concluded that the development oftaste perception is dependent on the number of taste budson a particular location.
Of the approximately 4,600 total taste buds in allthree lingual fields in humans, vallate buds compriseabout 48% (2200), foliates about 28% (1280) and fungi-forms 24% (1120). However, taste bud numbers varygreatly among individuals (Miller and Reedy, 1990a),with some adults possessing a total of only 500 taste buds
bcde
g
f
a
(a) (b)
sp
sp
bz
Figure 29.5 (a) First description of a mammalian taste bud (vallate papilla of the swine) by Schwalbe (1868). Minor bundles and fibrils of nervesare lost in the “interior of taste goblets”. Note the incorrect arrangement of basal cells. According to Schwalbe, two cell types, pin cells(Stiftchenzellen), and rod cells (Stabzellen), might mediate different taste sensations. (b) Distinguished observation of intra- and intergemmal nerveterminals in the rabbit foliate papillae by Retzius (1892) using the Golgi silver impregnation method. Taste buds are directed to the trench of thepapilla. Retzius also depicts a slender bipolar “sensory” intragemmal cell and a multipolar cell (bz) (right side). Direct contacts between nerve fibersand taste cells were not noticed. Retzius believed that exclusively nerve fibers rather than taste cells were responsible for gustatory sensations.
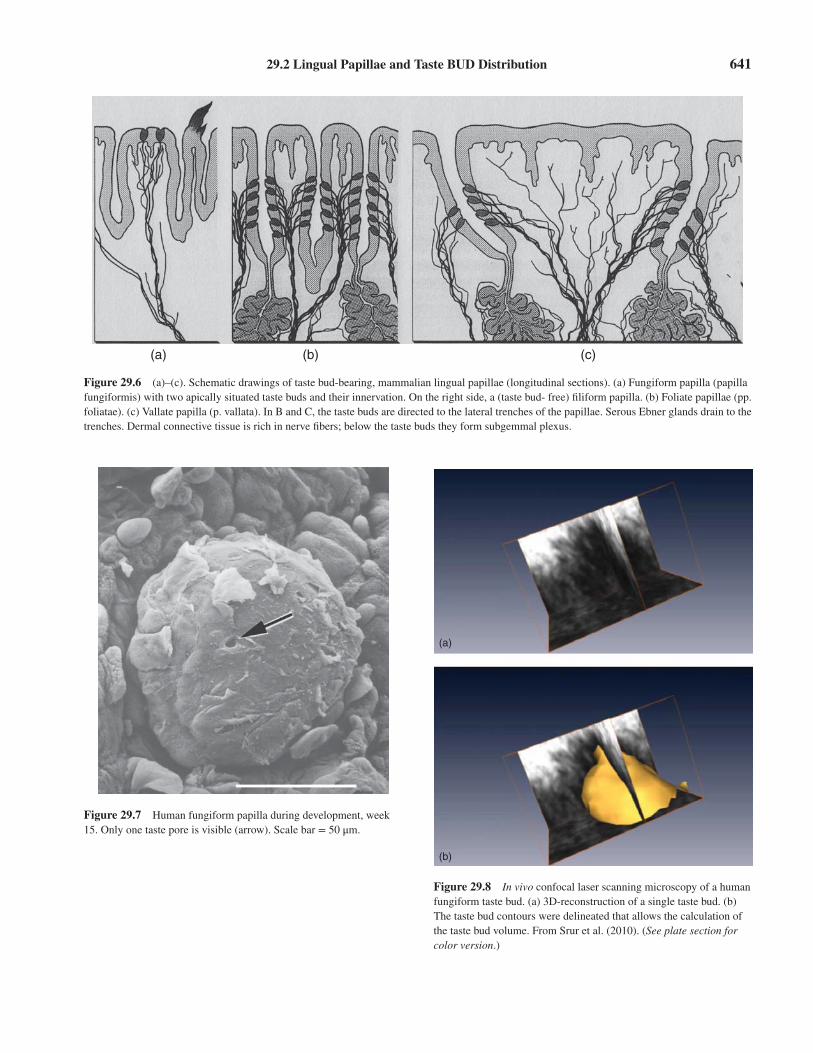
29.2 Lingual Papillae and Taste BUD Distribution 641
(a) (b) (c)
Figure 29.6 (a)–(c). Schematic drawings of taste bud-bearing, mammalian lingual papillae (longitudinal sections). (a) Fungiform papilla (papillafungiformis) with two apically situated taste buds and their innervation. On the right side, a (taste bud- free) filiform papilla. (b) Foliate papillae (pp.foliatae). (c) Vallate papilla (p. vallata). In B and C, the taste buds are directed to the lateral trenches of the papillae. Serous Ebner glands drain to thetrenches. Dermal connective tissue is rich in nerve fibers; below the taste buds they form subgemmal plexus.
Figure 29.7 Human fungiform papilla during development, week15. Only one taste pore is visible (arrow). Scale bar = 50 μm.
(a)
(b)
Figure 29.8 In vivo confocal laser scanning microscopy of a humanfungiform taste bud. (a) 3D-reconstruction of a single taste bud. (b)The taste bud contours were delineated that allows the calculation ofthe taste bud volume. From Srur et al. (2010). (See plate section forcolor version.)

642 Chapter 29 Anatomy of the Tongue and Taste Buds
(Linden, 1993). The taste bud density of foliate papil-lae seems to be constant in life (Hou-Jensen, 1933), butage-related differences have been reported for vallatepapillae, which are more numerous and containing moretaste buds in younger individuals (Jurisch, 1922). There arealso more marginal fungiform papillae during the late fetaland newborn period, but they usually lack taste buds andare referred to as “sucking papillae” (Habermehl, 1952;Yamasaki and Takahashi, 1982).
29.2.2.2 Vallate Papillae. Vallate papillae, firstcomprehensively described by Haller (1766) and Sömmer-ing (1806), lie directly anterior to the sulcus terminalis andextend in a V-shaped line across the root of the tongue(Figures 29.4 and 29.12). They are round and measurebetween 2 and 8 mm in diameter. The pores of taste budsopen into the trenches around the bases of each vallatepapilla (Figure 29.6). The papillae are innervated by anenormously large nerve fiber plexus originating from CNIX (see below) compared to foliate or fungiform papillae.The number of vallate papillae per human tongue variesbetween 4 and 18 (n = 2,264 tongues), with an average of9.2 ±1.8 papillae (Münch, 1896). Ninety-eight percent ofall tongues have a central median papilla (Figure 29.4).The presence of three or four lateral papillae on eachtongue half was observed most often (20%). Atrophicchanges were observed in papillae of some men > 40 yearsold and some women > 55–60 years old, though Jurisch(1922) reported that the number of vallate papillae did notappear to change systematically as a function of age. Theaverage numbers of taste buds per papilla are summarizedin Table 29.1.
29.2.2.3 Foliate Papillae. Foliate papillae inhumans were first reported by Albinus (1754) and histo-logically described by Hönigschmied (1873), but did notbecome the focus of scientific attention in humans until the
twentieth century. These papillae, located bilaterally alongthe posterolateral margins of the tongue surface, consistof parallel rows of ridges (folia) and valleys which lieadjacent to the lower molar teeth. Ducts located betweenthe folia transmit secretions from mostly serous lingualglands within the root of the tongue.
Scanning electron microscopy of human foliate papil-lae and transmission electron microscopy of their residenttaste buds was reported by Svejda and Janota (1974) andAzzali et al. (1996). The number of taste buds in humanfoliate papillae was reported by Hou-Jensen (1933) andMochizuki (1939) (see Table 29.1). Confusion in find-ing taste buds within the folds of human foliate papillaereflects papilla structure. As many as 20 parallel ridgesand furrows are found on the posterolateral margin of thehuman tongue. The rostralmost furrows (lateral rugae)contain no glandular ducts or taste buds, and their epithe-lia are more cornified than that between foliate papillae(Hou-Jensen, 1933). Fungiform papillae can be found onthe tops of these lateral rugae. Although human foliatepapillae were thought to be “rudiments” (in comparisonto the well developed rabbit foliate papillae), Mochizuki(1939) calculated an average of 1300 taste buds per tongue,which exceeds the number of fungiform taste buds [800buds: Braus, 1940; <1000 buds: Miller and Bartoshuk,1991]. Indeed, contiguous taste buds within the same cleftmay form a more functional unit, since they share accessto a common taste stimulus pool. Foliate papillae areinnervated by branches of the glossopharyngeal nerve (CNIX), but the more anterior portion also receives nerve fibersfrom the chorda tympani (Oakley, 1970; Pritchard, 1991).
29.2.2.4 Fungiform Papillae. Due to their mor-phological heterogeneity, fungiform papillae have beenvariously described as papillae clavatae, capitatae, lentic-ulares, obtusae, majores, and mediae (Haller, 1766;Sömmering, 1806). These papillae can be easily identified
Table 29.1 Distribution of taste bud numbers within the oral cavity of adult humans.
Vallate Papillae Fungiform Papillae Foliate Papillae Palate Larynx Author
– 20–30 ? 15–20 Hoffmann, 1875400 Wyss, 1870252±151* Arey, Tremaine, and Monzingo, 1935234±114* 1279 Mochizuki, 1939240±125* <1000 (total) – Miller and Bartoshuk, 1991
800 (total) Braus, 1940708–1328 Hou-Jensen, 1933
>2500 (neonate) Lalonde and Eglitis, 1961<25 (senile) Jowett and Shrestha, 1998
585 Cheng and Robinson, 19911120 Miller and Reedy, 1990b
*per papilla.

29.4 Salivary Glands of the Tongue 643
as pink elevations about 0.5 mm in diameter on the anteriorportion of the living human tongue. Notwithstanding itsconvenient location, the fungiform taste bud population hasbeen difficult to quantify since fungiform papillae vary inappearance and are distributed over a large area of tonguesurface. The anterior portion of the tongue extends from theline of vallate papillae to the tongue tip (Figures 29.4 and29.12). This region contains about 30 cm2 of surface area,depending on the size of the person, and the fungiformpapillae are spread unevenly over it. The number of tastebuds differs among fungiform papillae, and there are largevariations among human subjects in fungiform taste buddistribution. Most papillae on the 5 mm margin of thetongue tip are shorter than those on more posterior regions.Following the surface in a posterior direction from themidline of the tip toward the back of the tongue, fungiformpapillae become progressively larger in size, being largestin the more posterior lingual regions.
Small, rounded papillae are present on the margin.Some of them contain taste buds, while others are compa-rable in size to filiform papillae but lack fila. Some papillaeon the margin of the tip are elongated like conical papillae,and these, generally, lack taste buds. Fungiform papillaevary in size and shape: some are short and cuboidal,and others are tall with expanded heads like mushrooms.Among papillae on the margin of shorter height and smallerdiameter (<0.5 mm), the distinction between filiform andfungiform papillae becomes obscure. Fungiform papillaeoccasionally have projections on their apices like small filaon filiform papillae.
Taste buds have been quantified in terms of tongue sur-face areas, referred to as “taste bud density,” or numberof taste buds per cm2 of tongue surface. There are about145 gustatory (fungiform) papillae per lateral half of thetongue, with about 30 papillae per cm2 on the tip, but onlyabout 3 papillae per cm2 on the posterolateral area. Thereare about 30 large fungiform papillae in the posteromedialregion, for an estimated total of about 320 fungiform papil-lae per tongue (Miller and Reedy, 1990b; Shahbake et al.,2005). On 320 fungiform (gustatory) papillae, an averageof about 3.5 taste buds per papilla has been estimated, fora total of 1120 fungiform taste buds (Miller and Reedy,1990a, 1990b) (Table 29.1).
Most investigators who study human fungiform papil-lae report the existence of papillae without taste buds,which rarely occurs in other mammals (Mistretta andBaum, 1984). Studies in humans show, depending onthe methods used, that fungiform papillae lacking tastepores comprise 1–67% of the fungiform population persubject (Arvidson and Friberg, 1980; Cheng and Robinson,1991; Miller and Reedy, 1990a; Segovia et al., 2002).This wide range may reflect how investigators decidewhich papillae are “fungiform.” Children possess a higherdensity of fungiform papillae, which correlates well with
increased sensitivity for sucrose (Segovia et al., 2002).Modern non-invasive imaging techniques, for example,confocal laser microscopy, allow the in vivo visualizationof fungiform taste buds and their dynamics over a certainperiod of time, even buds that have not yet presented ataste pore (approx. 10%; Just et al., 2005; Srur et al., 2010)(Figure 29.8).
29.3 EXTRALINGUAL TASTE BUDS
There are “extralingual” taste buds in regions of the oral,pharyngeal, and laryngeal cavities. Interestingly, Magendie(1820) and Carus (1849) erroneously associated the teethwith taste perception. Verson (1868) described the first“goblet-like organs” within the dorsal epithelium of theepiglottis, and Davis (1877) and Wilson (1905) observedtaste buds in and the elicitation of taste perception from thehuman larynx. Lalonde and Eglitis (1961) counted morethan 2,500 taste buds on the epiglottis, soft palate, laryngealpharynx, and oral pharynx of one human neonate. Tastebuds are evident in the epiglottis of one neonatal specimen(Rabl, 1895), and esophagus in human fetuses (Ponzo,1907) and adults (Burkl, 1954; Schinkele, 1942). Tastebuds are also found near the openings of sublingual salivarygland ducts in some other primates (Hofer et al., 1979),and near the ducts of the molar glands in rodents (Iidaet al., 1983). In chickens and quail, taste buds in nonlingualparts of the oral cavity are almost always associated withsalivary gland ducts (Ganchrow and Ganchrow, 1987).Miller and Smith (1984) estimate that about 25% of thehamster’s total taste buds are extralingual, and Mistrettaand Baum (1984) accounted for a similar proportion ofextralingual taste buds in the rat. It is not known whetherextralingual taste buds are functionally different from thoseon the tongue. Taste buds of the epiglottis and/or uvulacould be involved in initiation of upper airway reflexes(Bradley et al., 1983) and in the pharyngolaryngeal waterresponse, possibly mediated by receptors signaling theabsence of chloride ions (reviewed by Lindemann, 1996).Similarly, taste buds of the larynx seem not to play a rolein gustation but detect chemicals that are not saline-like incomposition, for example, CO2 (Bradley, 2000; Nishijimaand Atoji, 2004). Apparently, structure and immunohisto-chemical properties of taste buds are remarkably conserveddespite their different locations and innervation patterns(Kinnamon, 2011).
29.4 SALIVARY GLANDSOF THE TONGUE
Lingua sicca non gustat (A dry tongue does not taste). Thisdeclaration of Haller (1766) refers to the dependence of

644 Chapter 29 Anatomy of the Tongue and Taste Buds
taste ability on solutions, within which tastants are dis-solved and transported to the taste bud. Extralingual salivais secreted by small, mostly mucous glands embeddedin the epithelium of the cheek and palate. More saliva isproduced by the serous parotid gland and the muco-seroussublingual and submandibular glands (Figure 29.3), whosesecretory ducts open at the tongue frenulum just underneathits tip. Intralingual saliva originates from the mucoserousanterior lingual glands [glands of Blandin and Nuhn,(Tandler et al., 1994), Figure 29.3], and deep posteriorserous salivary glands (Ebner) located in the submucousconnective tissue below the foliate and vallate papillae ofthe tongue (Ebner, 1873; Riva et al., 1999). Their excretoryducts lead to the deepest sites of the papillar furrows(Figure 29.6). The gland lobules lie deeply in large patchesof connective tissue which, in turn, are separated fromeach other by muscle fiber bundles. In addition, adjacentto Ebner glands in vallate papillae lie mucous (Weber’s)glands (Nagato et al., 1997), which in humans open intothe crypts of the lingual tonsils (Zimmermann, 1927).Neither Weber’s nor Blandin-Nuhn glands lie in closeproximity to taste buds and their particular significancefor taste perception is unknown because of the difficultiesin collecting saliva from these glands (Tandler et al.,1994).
There is biochemical and histochemical evidencethat the saliva of Ebner’s glands, as well as that of othernon-lingual salivary glands, has more functions than thatof a serious “washing solution.” Binding proteins such asEbnerin (Li and Snyder, 1995) are supposed to modulatesensations. Schmale et al. (1990) isolated a protein fromrat Ebner’s glands that is structurally similar to odorantbinding proteins in Bowman’s glands of the olfactorymucosa. The gland is under autonomic control (Fukamiand Bradley, 2005; Gurkan and Bradley, 1987). Interest-ingly, a recent report seems to point at a dependency of thehypoglossal nerve on sympathetic innervation of posteriorlingual glands in hamsters (Cheng et al., 2009). For reportson specific ligand-receptor interaction with taste qualities,see Azen et al. (1990), Schmale et al. (1993), Spielman,(1990), Toto et al. (1993), and Chapter 31 of this volume.
29.5 BLOOD SUPPLY TOGUSTATORY PAPILLAE
The mammalian tongue receives its blood supply fromthe lingual artery, which is usually a branch of the exter-nal carotid artery (Figures 29.3 and 29.4). Study of thetongue’s vascular system historically parallels that of thelingual papillae. For example, Albinus (1754), Sömmer-ing (1806) (Figure 29.4) and Arnold (1839) performedintravascular injections in order to visualize the papillarysurface. More recently, distribution of the blood supply
to different regions of the tongue and different types oflingual papillae has been described by Hellekant (1976).Each type of gustatory papillae is supplied by a character-istic capillary configuration (rat: Ohshima et al., 1990; cat,rabbit: Ojima et al., 1997a-c) and fine capillary networksare found adjacent to taste buds. The capillary loops oflarger papillae in rats and dogs often show a constriction,maybe sphincter-like structures, but rarely arteriovenousanastomoses (Hu et al., 1996; Selliseth and Selvig, 1993).Taste stimuli injected systemically elicit responses ingustatory nerves as the bolus passes through the tongue(Bradley, 1973).
29.6 SOLITARY CHEMOSENSORYCELLS
In addition to taste buds and free nerve endings, the solitarychemosensory cells (SCC) comprise another chemosen-sory system in vertebrates. They are not assembled inclusters, but are dispersed across the surface of the animal.SCC are related to taste bud cells in the sense that theformer are secondary sensory cells with a slender, bipolarphenotype (Finger, 1997). “Classical” SCC have beenstudied first in teleosts (Whitear, 1992). The evolutionarybenefits of these cells are still in question: In sea robins(Trigon), they are, beside taste buds, involved in findingfood; in rocklings (Gaidropsarus) they are assumed tobe important for predator avoidance (Kotrschal, 1996).Since the arginine-like receptor in catfish taste buds alsooccurs in SCC, Finger (1997) suggests that taste budsmight include SCC within them. During developmentof fish, SCC seem to precede the development of tastebuds. In mammals, however, SCC-like cells have beenobserved only transiently, during development. In new-born rats, single gustducin-immunopositive cells are seenin locations where later-developing vallate papillae willappear (Sbarbati et al., 1999). Individual slender cells,immunopositive for cytokeratin 20 (Witt and Kasper,1999), an intermediate filament protein that is exclusivelypresent in taste bud and epidermal Merkel cells (Moll,1993; Zhang and Oakley, 1996; Zhang et al., 1995), areseen occasionally during early ontogenesis of the humantongue. Gustatory epithelia sensu strictu of adult mammalshave not yet been reported to possess SCC. However,alpha-gustducin-immunoreactive SCC occur in the nasalmucosa of mice and have been interpreted as sentinels inthe anterior nasal air passages (Finger et al., 2003). Tastereceptors (Tas1R, Tas2R) in SCC along with their synapticconnectivity to CGRP-positive polymodal pain fibers ofthe trigeminal nerve indicate a role in detection of irritantsand foreign substances by triggering trigeminally-mediatedreflexes (Tizzano et al., 2011).

29.7 Cell Types of Vertebrate Taste Buds 645
29.7 CELL TYPES OF VERTEBRATETASTE BUDS
Peripheral taste organs differ in number, size, and shapein different vertebrate taxa, according to their importancefor the particular species. For the sake of brevity, thefollowing overview is restricted to some functionallywell-characterized vertebrate species. Details are availablein reviews by Reutter and Witt (1993) or Chaudhari andRoper (2010). Figure 29.9 shows a scheme representingthe organization of cells in fish, frog, and mammaliantaste buds.
29.7.1 Taste Buds of LowerVertebrates – Cell Types29.7.1.1 Fish. In fish, and especially in some teleoststhat are well-adapted to the dark, the taste organ is signifi-cantly more important for food intake than in amphibiansand mammals. Thus, these fish, like the Siluridae, pos-sess many more taste buds than representatives of thelatter classes (Atema, 1971; Finger et al., 1996; Millerand Bartoshuk, 1991). The fish taste bud is generallypear-shaped and similar to that of mammals (Figure 29.9).Electron microscopic studies of teleost taste buds have
Teleost Frog Rabbit
IIII IISm
wwSr
M
D
H
DL
DD
L∗
(a) (b) (c)
Figure 29.9 (a)–(c) Longitudinal sections of taste organs from representatives of three different vertebrate classes. (a) Fish (bullhead, Teleostei),(b) Amphibian (frog, Anura), (c) Mammal (rabbit). In the schematic drawing, each sensory epithelial cell type is represented once with a distinctgrey-step. The organs lie in squamous epithelium of different height, on top of dermal papillae which are also of different height. Each dermalpapilla contains nerve fibers and a capillary vessel. (Below) 3D reconstructions of the respective apical taste bud portions. In all species, glia-likecells (blue in teleost and rabbit; sepia in frog “wing” cells) enwrap sensory cells. Sensory cells (green; sepia in rabbit as type II receptor cell). Nervefibers entangle all cells, but synapses are seen only with type III cells in rabbit and microvillar (Sm) and rod-like receptor cells (Sr) in frog. (Seeplate section for color version.)

646 Chapter 29 Anatomy of the Tongue and Taste Buds
attempted, somewhat incompletely, to relate the ultrastruc-ture and functions of taste bud cell types (for reviews seeJakubowski and Whitear, 1986; Reutter, 1986; Reutter andWitt, 1993). This is also apparent in the nonuniform usageof nomenclatures. Most authors refer to elongated “light”and “dark” taste bud cells, as well as “intermediate” and“degenerative” cells (Connes et al., 1988; Desgranges,1965; Reutter, 1971, 1978; Welsch and Storch, 1969;Whitear, 1970). Generally, light cells are supposed tobe “sensory” (receptor) cells, while the dark cells areregarded as “supporting” or “sustentacular” cells (e.g.,Desgranges, 1965; Hirata, 1966; Whitear, 1970). However,Reutter (1992) considers the dark cells also as sensorybecause they exhibit synaptic contacts with nerve profilesor with basal cells. These observations, however, are notsupported by the work of Grover Johnson and Farbman(1976) and Jakubowski and Whitear (1986). Accordingto these authors the differentiation “light” and “dark” incombination with functional terms such as “supportingcells” and “sensory (receptor) cells” are misleading andshould be avoided. According to Hirata (1966), Merkel(1880) and Reutter (1973) the taste bud may also havemechanoreceptive functions, particularly in view of themorphology of basal cells (Reutter, 1971, 1986) or seroton-ergic Merkel-like cells (Zachar and Jonz, 2011). While theexistence of synaptic contacts of light and dark taste budcells has not yet been finally proven, the different lengthsof their microvilli perhaps suggest functional differences:The light taste bud cells have a single long microvillusreaching far into the mucous layer of the taste bud sur-face, the receptor area. By penetrating this layer, longermicrovilli may be exposed to quite different “perireceptorevents” (Getchell et al., 1984) than the small microvilli ofdark taste bud cells that do not penetrate the mucous layer(Reutter, 1980; Witt and Reutter, 1990).
In larvae of lampreys, representatives of fish-likejawless vertebrates (Petromyzontidae), taste bud cellswere characterized by Retzius (1893) using the Golgisilver impregnation technique. Recently, Barreiro-Iglesiaset al. (2010) reported three different cell types includingserotonergic bi-ciliated cells, basal cells, and sustentacularcells.
Ultrastructural (Reutter and Witt, 1993) and neu-rochemical (Ferrando et al., 2012) investigations innon-teleostean fish reveal clearly that taste buds differwithin the main vertebrate taxa, which is partly repre-sented in the differential expression of G protein-coupledreceptors (Oka and Korsching, 2011). Further, in differentsystematic groups of fish the taste buds do not follow onlyone structural design. Thus, taxon-specific taste buds orcell types do not exist. Similar differences among mam-malian taste buds point to a similar inevitable conclusionregarding particular phenotypes: There are only species-specific taste bud types, and a general “model” seems
difficult to find among vertebrates. This underlines thethesis that morphological phenotypes and the structuralorganization of taste bud cells do not necessarily reveal ageneral bauplan, but rather, reflect specific environmentalconditions and/or feeding behaviors (Reutter and Witt,1999).
The influence of environmental differences has beenstudied in two closely related teleosts, one of which issighted (Astynax mexicanus) and the other of which isa blind cave fish (Astynax jordani). Whereas taste budmorphology is rather similar, the cave fish compensatesfor blindness by significantly more gustatory axon profiles(Boudriot and Reutter, 2001) and an expanded expressionof Prox 1 gene in developing taste buds (Jeffery et al.,2000; Varatharasan et al., 2009).
29.7.1.2 Amphibians. The taste organs of post-metamorphotic Salientia (=Anura), unlike piscine andmammalian taste bud bulb-like formations, are rela-tively large disk-like epithelial differentiations of thedorsal lingual and palatal mucosa (Figure 29.9). Waller(1847, 1849) and Engelmann (1872) called these struc-tures “Geschmacksscheibe” or “taste disc.” In contrastto Salientia, the taste buds of the urodeles [= Caudata,e.g., mudpuppy (Necturus), newt (Triturus) or Axolotl(Ambystoma)] have a bulb-like shape (Cummings et al.,1987; Delay and Roper, 1988; Fährmann, 1967; Farbmanand Yonkers, 1971; Toyoshima et al., 1987; Toyoshimaand Shimamura, 1987). The cellular elements of thetaste disc, or “Endscheibe” (Merkel, 1880), of the frogwere subject to numerous investigations and receivedvarious designations: After Waller (1847, 1849) had firstdistinguished between papillae conicae (=P. filiformes)and papillae fungiformes, Fixsen (1857) described twodifferent cell types, the so-called “cellulae cylindricae”and “cellulae fusiformes,” the processes of which passthrough the whole sensory epithelium to reach the con-nective tissue core of the papilla. Engelmann (1872) andMerkel (1880) further developed the terminology: Merkeldistinguished between “cylindrical cells” (Cylinderzellen)situated on the epithelial surface and surrounding “wingcells” (Flügelzellen), the nuclei of which lie deeper inthe epithelium. After Graziadei and DeHan (1971) haddescribed only two cell types (“associate cells” and “sen-sory cells”) in electron microscopy, the close relationshipbetween “rod cells” (Stäbchenzellen) and cylindrical cells(Merkel, 1880) was recently re-introduced by von Düringand Andres (1976). In addition, the latter authors firstdescribed basal cells and Merkel cells of the frog tastedisc, which are the only cells that do not contact theepithelial surface. The taste disc in adult frogs containsup to eight cell types (Reutter and Witt, 1993): mucuscells, wing cells, two types of sensory cells (cylindricaland rod-like type), two types of basal cells [stem and

29.7 Cell Types of Vertebrate Taste Buds 647
Merkel cell-like basal cells (Zancanaro et al., 1995)],and marginal cells and ciliated cells (Toyoshima et al.,1999) (Figure 29.9). The cell types and the history ofthe nomenclature are described in detail by Jaeger andHillman (1976), Reutter and Witt (1993), Witt (1993),Osculati and Sbarbati (1995), and Li and Lindemann(2003).
Tadpoles possess so-called premetamorphic papillaewhich bear bud-like taste organs at their tops. Dur-ing metamorphosis, these structures wholly disappearand are replaced by fungiform papillae with large tastedisks (Nomura et al., 1979; Zuwała, 1997; Zuwała andJakubowski, 1991). Tadpole taste discs consist of sensoryand supporting cells, and basal cells are lacking.
Taste buds of the mudpuppy (Necturus, Urodela) aresimilar to those of fishes. They are composed of dark andlight cells and possess serotonergic Merkel cell-like basalcells which are synaptically connected with either nervefibers or dark and light cells (Delay et al., 1994, 1993; Delayand Roper, 1988).
29.7.1.3 Reptiles and Birds. Lingual taste buds inreptiles have been described in turtles (Iwasaki et al., 1996;Korte, 1979), tortoises (Pevzner and Tikhonova, 1980),and some lizards (Uchida, 1980). Lizards (Gekkonidaeor Anguidae), as well as snakes, have virtually no tastebuds on the tongue, but rather on the buccal floor andoral epithelia of the mandible and maxilla (Toubeau et al.,1994). In shape, reptilian taste buds resemble those ofmammals. There are up to five different types of tastebud cells, classified into light and dark cells and as types1,2,3,A,B, or as types I, II, II, and basal cells (Reutter andWitt, 1993).
Reptiles and birds belong to the same superclass,and one might expect a similar organization of aviantaste buds. However, the few examples of bird taste budsshow great variability among species. Buds in grain-eatingbirds appear in the posterior part of the tongue, near thepharynx, and in the distal palatal mucosa (Ganchrowet al., 1991; Ganchrow and Ganchrow, 1987; Saito,1966; Sprissler, 1994). Unlike mammals, avian tastebuds do not reside within lingual bud-bearing papil-lae. In addition, taste buds contain tubular-like channelscircumscribed by elongated cells grouped in a rosetteconfiguration, with the channel lumen continuous api-cally with the taste pore (Berkhoudt, 1985; Ganchrowet al., 1993). Taste buds in chicken are richly innervated(Ganchrow et al., 1986), contain gustducin in a subsetof taste cells (Kudo et al., 2010), and synapses are seenbetween all cell types (light cells and dark cells) andnerve fibers (Reutter and Witt, 1993; Sprissler, 1994).Stornelli et al. (2000) observed four cell types in duck tastebuds: light cells, dark cells, intermediate cells, and basalcells.
29.7.2 Mammalian TasteBuds – Cell TypesEarly histological investigators of mammalian taste budsdescribed two types of elongated, fusiform cells in tastebuds of human vallate papillae (Schwalbe, 1868): “support-ing” and “taste cells,” the latter divided into Stiftchenzellen(pin cells) and Stabzellen (rod cells) with differences incontrast and brightness (Figure 29.5).
Classification into “light” and “dark” taste bud cellswas also used in early electron microscopic analyses(Engström and Rytzner, 1956). Farbman (1965) considereddark, fusiform taste bud cells of human fungiform papillaeas sensory cells (type I), whereas other investigators suchas Paran et al. (1975) described a type II-cell that containsnumerous vacuoles and mitochondria, especially in apicalareas. This latter cell is not believed to be sensory. CottlerFox et al. (1987) speculated that the different electrondensities are due to irreproducible fixation artifacts. Ingeneral, ultrastructural and immunohistochemical criteriaare considered more important for the classification of cellsthan the evaluation of the electron density of the cytoplasm.Moreover, present understanding in mammalian taste budcytology leads to using a rather heterogeneous nomencla-ture, based on morphological and functional differencesacross species.
The basis for the current nomenclature was establishedby the Murrays and colleagues (Murray, 1986; Murray andMurray, 1967, 1971; Murray et al., 1969) in taste bud cellsof the rabbit foliate papillae. However, these cell typesdiffer in some respects from rodent taste buds (Kinnamonet al., 1985, 1993, 1994; Pumplin and Getschman, 2000;Pumplin et al., 1997, 1999). Thus, Royer and Kinnamon(1988) observed considerable deviations in the cytoarchi-tecture of murine foliate taste buds compared to that ofother mammals. For example, they did not find type IIIcells, and all bud cells had synaptic connections with nervefibers. Most current information on chemoreception oftaste stimuli and information transfer to primary afferentaxons have been established in mice (Dando and Roper,2009; Huang et al., 2009; Roper, 2006), but there aresignificant species differences (Ma et al., 2007), and theprecise organization of the taste bud in humans remains tobe determined (Azzali, 1996, 1997; Paran et al., 1975; Wittand Reutter, 1996). An example for a more generalizedtaste bud is depicted in Figure 29.10. Work of the lasttwo decades has revealed that taste stimuli are receivedby type II receptor cells (Tomchik et al., 2007), whiletype III (presynaptic) taste cells are the only ones thatcontain synaptic proteins and form synapses with nervefibers (Kinnamon et al., 1988). The information transferfrom type II (receptor) cells to type III (presynaptic) cellsis mediated through ATP and pannexin hemichannels(reviewed by Chaudhari and Roper, 2010) or directly

648 Chapter 29 Anatomy of the Tongue and Taste Buds
I II III
V
IV
Figure 29.10 Mammalian taste bud in longitudinal section,idealized schematic drawing according to electron microscopicalfindings. Each cell type is depicted once. Following Murray‘snomenclature, cells of type I, II and III are elongated and form thebuds sensory epithelium proper. Apically, these cells end with differenttypes of microvilli within the taste pit and may reach the taste pore.Type I cells are glia-like, type II cells comprise receptor cells for bitter,sweet, and umami stimuli. Synapses are only found at the bases of typeIII (presynaptic) cells. Type IV cells are basal cells, type V marginalcells. Nerve fibers within the short dermal papilla are slightlymyelinated, and within the taste bud they form an unmyelinatedplexus. Note the basal lamina between dermis (which contains acapillary) and the epithelium.
from type II cells to nerve fibers, as gustatory afferentsalso contain purinergic receptors P2X2 and P2X3 forthe interaction with ATP (Finger et al., 2005). A surveyrepresenting the most important substrates associated withtaste cell types in vertebrates is shown in Table 29.2.
29.7.2.1 Type I Cells (Glia-Like Cells). Thesecells are the most frequent. They are spindle-shaped andhave a basal process that envelops the axons in a Schwanncell-like manner. Type I cells protrude with brush-like, longmicrovilli (1–2 μm) into the taste pit. These cells ensheathtype II and type III cells with cellular protrusions and mayinsulate them (Figures 29.9 and 29.10). Apically, theycontain large granules, 100–400 nm in diameter (Murray,
1986). The nuclei of type I cells are irregularly shaped.According to Murray and Murray (1971) and Murray(1986) these cells have a secretory (supporting cells) andpossibly phagocytotic function, and probably produce theamorphous material of the taste pit (e.g., (Farbman, 1965;Menco, 1989; Ohmura et al., 1989; Witt, 1996). Type Icells are involved in neurotransmitter clearance and ionredistribution, similar to the function of glia cells in thecentral nervous system. They contain GLAST, a glutamatetransporter (Lawton et al., 2000), as well as enzymesdegrading extracellular ATP (Bartel et al., 2006). Sometype I cells possess the K+ channel ROMK that may elimi-nate accumulated extracellular potassium ions to retain theexcitability of type II and type III cells (Dvoryanchikovet al., 2009).
29.7.2.2 Type II Cells (Receptor Cells). Most ofthese cells are located in the periphery of the taste bud.They are fusiform, but do not possess enveloping processes,or granules. Their cytoplasm is moderately electron-dense,and nuclei are round to oval. Synapses are not observed(Farbman et al., 1985, 1987). Toyoshima and Tandler(1987) describe a modified endoplasmic reticulum withspecialized sub-surface cisterns adjacent nerve profiles. Inmice, vallate and foliate “light” (type II-) and “dark” (typeI- and type II-) taste bud cells exhibit synaptic contactswith nerve fibers, thus suggesting a gustatory function.However, type I- and type II- cells do not form synapseswith the same nerve fibers (Kinnamon et al., 1985, 1988;Royer and Kinnamon, 1988). These cells are equipped withG-protein coupled receptors and tuned to sweet, bitter, andumami, but not to sour and salty taste (Matsunami et al.,2000; Tomchik et al., 2007).
29.7.2.3 Type III Cells (Presynaptic Cells).Taste buds contain only 5–7% of type III cells. Theyhave unbranched basal and apical processes. Their apicalportion protrudes with a single large microvillus into thetaste pit and may reach the taste pore. Type III cells are theonly cells that have synaptic contacts with intragemmalnerve fibers. Near the cell nucleus there are numerousdense-cored vesicles (80–140 nm in diameter) which areinvolved in neurotransmitter synthesis (GABA, serotonin,norepinephrine) (Fujimoto et al., 1987; Hokfelt et al.,1980; Huang et al., 2005; Nada and Hirata, 1977). TypeIII cells contain several synaptic proteins and are thereforetermed “presynaptic cells” (DeFazio et al., 2006). Theirapical process apparently possesses channels for sour tastereception (Huang et al., 2008).
29.7.2.4 Type IV Cells (Basal Cells). Type IVcells (Murray, 1973; Murray et al., 1969) include basalcells (Nemetschek-Ganssler and Ferner, 1964) or “pre-gustatory cells” (Scalzi, 1967). These are relatively small

29.7 Cell Types of Vertebrate Taste Buds 649
Table 29.2 Survey of selected substances (transmitters, channel or transporter proteins) associated with certain types of tastebud cells in various species.
Taste Bud Cell Type Substance/Transmitter Species Reference
Basal Cells 5HT (serotonin) catfish Reutter, 1971basal cells 5Ht, Glu, GABA necturus Kim and Roper, 1995; Nagai et al., 1996basal cells 5HT frog Jain and Roper, 1991I Glutamate/aspartate transporter
(GLAST)mouse Lawton et al., 2000
I Ecto-ATPases mouse Bartel et al., 2006I Epithelial Na+ channel (ENAC) mouse Lin and Kinnamon, 1999; Vandenbeuch, Clapp,
and Kinnamon, 2008I K+ channel ROMK mouse Dvoryanchikov et al., 2009II 5HT, Serotonin transporter rat Ren et al., 1999II Acetylcholine mouse Dando and Roper, 2009II Acetylcholine transporter,
Choline-acetyltransferase(ChAT),
mouse Ogura et al., 2007
II Vasoactive intestinal peptide(VIP)
rat, carp Shen et al., 2005; Witt, 1995
II Cholecystokinin rat Herness et al., 2002II Neuropeptide Y rat Zhao et al., 2005II Type III IP3 receptor rat, mouse Clapp et al., 2001n.d. Gustducin chicken Kudo et al., 2010II Gustducin mouse, rat, hamster Boughter Jr. et al., 1997; Hoon et al., 1995;
McLaughlin, McKinnon, and Margolskee,1992; Ruiz-Avila et al., 1995
II TAS2R38 (bitter taste receptor) human Behrens et al., 2012II NGF, trkA mouse Yee, Bartel, and Finger, 2005II PLCbeta2 Mouse Clapp et al., 2004II PLCbeta2 Human Behrens et al., 2012II TRPM5 mouse Perez et al., 2002II T2R Matsunami, Montmayeur, and Buck, 2000II T1R Hoon et al., 1999II GPCRs signal molecules for
bitter, sweet and umamimouse DeFazio et al., 2006
II (III?) ATP release mouse Bartel et al., 2006; Finger et al., 2005II (Gustducin-neg.) PGP 9.5 rat Yee et al., 2001(II) Snap 25 rat Oike, Matsumoto, and Abe, 2006; Pumplin and
Getschman, 2000; Ueda et al., 2006(III) Snap 25 rat Yang et al., 2000III SNAP25, synapsin II,
NCAM,AADC.mouse DeFazio et al., 2006
II GABA, glutamate decarboxylase(GAD)
rat Cao et al., 2009
(II) GABA rat Obata et al., 1997II (Gustducin neg.) GABAB2 mouse Starostik et al., 2010III GAD, GABA-B Receptor mouse Starostik et al., 2010III GAD mouse DeFazio et al., 2006III 5HT mouse, rat, rabbit, monkey Fujimoto, Ueda, and Kagawa, 1987; Kaya et al.,
2004; Nada and Hirata, 1975; Nada and Hirata,1977
III 5HT mouse Huang et al., 2005III 5HT monkey, rb Fujimoto, Ueda, and Kagawa, 1987III NCAM rat Nelson and Finger, 1993III BDNF, trkB mouse Yee, Bartel, and Finger, 2005III (5HT-neg.) Protein gene product 9.5
(PGP 9.5)rat Yee et al., 2001

650 Chapter 29 Anatomy of the Tongue and Taste Buds
undifferentiated cells that lie at the taste bud’s base, whichdo not form processes that reach the pore. They containnumerous bundles of intermediate filaments (Royer andKinnamon, 1991), and differ from Merkel cell-like basalcells of fishes and amphibians. Type IV cells are consideredto be undifferentiated stem cells of their bud cell progeny(Chaudhari and Roper, 2010; Murray, 1973; Murray, 1986;Roper, 1989). In embryonic and adult vallate taste budsof mice, basal cells also express the transcription factorsHes6 and Mash-1, the latter possibly being involved inspecification of a type III lineage (Seta et al., 2003, 2011).Using genetic lineage tracing in murine taste buds, Miuraet al. (2014) found sonic hedgehog (shh) in most basalcells, which indicates a fate as postmitotic, immediateprecursors of all three cell types rather than a stem cell.
Some authors report on “intermediate cells” (e.g.,Kinnamon et al., 1985). It has been pointed out (Farbmanet al., 1985; Roper, 1989) that differences in the electrondensity of the cytoplasm could also reflect different stagesin the maturation of the same cell type which indicatedifferent states of function.
29.7.2.5 Type V- Cells (Marginal Cells).“Marginal cells” (also “perigemmal cells,” and, in exten-sion of Murray’s nomenclature, “type V cells”) havebeen described (Beidler and Smallman, 1965; Farbman,1980; Gurkan and Bradley, 1987; Reutter and Witt, 1993).However, they have nothing in common with the secretorymarginal cells of taste organs in fish and frog, and maypossibly be taste bud stem cells (Beidler and Smallman,1965; Farbman, 1980) which express particular non-tastereceptor proteins, for example, CD44 isoforms (Witt andKasper, 1998) during human taste bud ontogenesis.
29.7.3 Molecular Markers of TasteBud CellsOne of the most intriguing challenges for suggesting pos-sible functional properties of taste bud cells is to identifysubsets of these cells by morphological features as well asmolecular properties, many of which can be traced evenin enriched primary taste bud cell cultures (Kishi et al.,2001, 2002; Ozdener et al., 2006). Histochemical evidenceon the neurochemical nature of taste cells have identifiedthe panneuronal markers, neuron-specific enolase (NSE)and protein gene product 9.5 (PGP 9.5) (Astbäck et al.,1997; Montavon et al., 1996; Yee et al., 2001; Yoshie et al.,1988), carbohydrate- binding proteins, the lectins (Wittand Miller, 1992; Witt and Reutter, 1988). Immunoelec-tron microscopic studies have tried to match functionalparameters with those of conventional electron density.For example, cell adhesion molecules (Nolte and Martini,1992; Smith et al., 1993, 1994) and several blood-group
antigens (Pumplin et al., 1997, 1999; Smith et al., 1999)characterize subsets of type II taste bud cells. A subsetof the (light) type II cell contains partly the G proteingustducin (Menco et al., 1997; Ruiz-Avila et al., 1995),which is involved in the perception of sweet and bitter taste(Tomonari et al., 2012; Wong et al., 1996).
Choline acetyl transferase, an enzyme involved in thesynthesis of the neurotransmitter acetylcholine, has beenidentified in rat type II cells (Menco et al., 1997). Whereasthe putative neurotransmitter serotonin is confined to basalcells of fish taste buds (Reutter, 1971) and Merkel cell-likebasal cells of amphibian taste organs (Delay et al., 1997;Hamasaki et al., 1998; Toyoshima and Shimamura, 1987),serotonin in mammals has been described in type III cells inthe rabbit (Fujimoto et al., 1987; Kim and Roper, 1995) andhuman taste buds (Azzali, 1997). This led to the hypothesisthat these cell types were equivalent in both taxa (Kim andRoper, 1995). Lindemann (1996) suggests the term “sero-tonergic cells” instead of type III cells (<10% of all cells).
Generally, neuropeptides are located in intragemmalnerve fibers rather than in particular bud cells. An exceptionis vasoactive intestinal peptide (VIP) that has been detectedin a subset of rat type II cells (Herness, 1989; Shen et al.,2005), and in light taste bud cells in the carp (Witt, 1995)by electron microscopy. Though most of these markers areexpressed only in differentiated cells and are not evidentafter nerve dissection (Smith et al., 1993; Whitehead et al.,1998), their functional correlation with taste perceptiondata is mostly unknown.
To avoid the present Babylonian confusion of tongueswith regard to taste bud cell nomenclature, presentresearch directions try to associate electrophysiologically-characterized, isolated taste bud cells with a particular celltype based on its specific substrate expression. Modern cellbiological approaches, for example, introduction of greenfluorescent protein chimeras in vitro (Landin et al., 2005)or calcium imaging after application of specific stimuli(Caicedo et al., 2000), have contributed to a solution of thisproblem.
Several authors report on morphological and immuno-histochemical differences between vallate/foliate andfungiform taste buds within the same species. For example,mouse taste bud cells of fungiform papillae contain moresynapses and presynaptic vesicles than those of vallatepapillae (Kinnamon et al., 1993), and the number of tastebud cells containing group H blood antigen and gustducinis three times higher in vallate than in fungiform papillae(Smith et al., 1993). The coexpression pattern of tastereceptors (T2R and T3R) and gustducin differs betweenfungiform and vallate taste buds in mice (Kim et al., 2003).In rabbit, lectin carbohydrate profiles of both taste budpopulations differ as well (Witt and Miller, 1992). Thereasons and significance of these differences betweenfungiform and vallate/foliate taste cells are not clear, but

29.8 Development of the Human Peripheral Taste System 651
factors determining their varying phenotype could includea different local saliva composition (Schmale and Bam-berger, 1997; Schmale et al., 1990; Shatzman and Henkin,1981) or morphogenetic conditions of local epithelium(Smith et al., 1999).
Evidence for communication between taste cells, apartfrom purinergic transmission (Finger et al., 2005), includesthe presence of cell adhesion molecules (Nolte and Mar-tini, 1992; Smith et al., 1993), heparin-binding proteins(Wakisaka et al., 1998), and membrane receptors that influ-ence the intracellular signal transduction cascades. Forexample, the hyaluronan receptor, CD44, was identified ina subset of human fetal taste bud cells (type V, marginalcells) and most of adult human taste bud cells (Witt andKasper, 1998). This transmembrane protein is linked toa series of actin-associated microfilaments, for example,ezrin and ankyrin, which are located in microvilli of type Icells and might influence the function of ion-translocatingmembrane proteins (Höfer and Drenckhahn, 1999). In lightof efferent neural control, taste bud cell communicationmay be mediated via local axon reflexes between sensorycells (Caicedo et al., 2000, Reutter and Witt, 2004).
29.8 DEVELOPMENT OF THEHUMAN PERIPHERAL TASTESYSTEM
Morphogenesis of the mouth cavity is characterized by thedevelopment of the tongue anlage which appears prior to,and is a prerequisite of, the formation of gustatory papillae.
At the embryonic age of 4 weeks, the first structureof the tongue anlage to appear is the tuberculum imparwhich is situated between the first (mandibular) and second(hyoid) branchial arches (Figure 29.11). Then, anterolat-eral to the tuberculum impar, the paired lingual swellings(which derive from the medial parts of the mandibulararches) fuse with the tuberculum impar. The tongue’s baseis formed by the hypobranchial eminence (copula of His)forming within the third and fourth branchial arches. Theborder between the caudal part and the body of the tongueis demarcated by a V-shaped rim, the sulcus terminalis(Bradley, 1972; Witt and Reutter, 1997). The innervationpattern of cranial nerves, which later supplies particularlingual regions, reflects the early innervation of branchialarches (see Figure 29.11): The pretrematic nerve of thefirst branchial arch is the lingual nerve (from CN V3);that of the second arch, the chorda tympani (from theintermedio-facial nerve); and that of the third arch con-stitutes later lingual rami of the glossopharyngeal nerve(for details see textbooks on embryology, for example,Hinrichsen, 1990; Williams et al., 1989).
The first detailed developmental studies on the surfaceappearance of the tongue were carried out by Froriep
(1828), and continued histologically by Tuckerman (1889),Gråberg (1898), and Hellman (1922). Hermann (1885)described the stages of karyokinesis in developing tastebuds. It was unclear to Hermann if supporting, or neu-roepithelial, cells were being replaced. Vallate papillaestart to develop earlier than fungiform papillae, and beginwith the appearance of a central midline papilla justbehind the foramen caecum around the sixth postovulatoryweek. From week 7 on, there develop many hillock-likeepithelial elevations on the tongue’s dorsum, as seenwith scanning electron microscopy (Figure 29.12). Someof these elevations are precursors of fungiform papillaeand are especially densely distributed near the midlineand the lateral ridges of the tongue (Habermehl, 1952;Hersch and Ganchrow, 1980; Witt and Reutter, 1997).Analysis of serial sections of the tongue encompassingthis critical developmental age (weeks 6–8) demonstratesthat not every dermal elevation will be the target of nervefibers. First, around week 7–8, nerve fibers, migratingtowards the periphery, form a large intragemmal plexus.Our own studies show that there are no taste bud anlagenwithout approaching nerve fibers (Witt and Reutter, 1996;Witt and Kasper, 1998), but recent studies revealed thenerve-independent development of taste buds and theirpreforming papillae (Barlow, 2003; Ito and Nosrat, 2009;Nosrat et al., 2012; Stone et al., 1995; Thirumangalathuet al., 2009). Also, taste bud primordia without dermalpapillae are evident, as well as individual bipolar epithe-lial cells resembling solitary chemosensory cells. Theseindividual cells are immunopositive for cytokeratin 20, amarker for lingual taste bud cells (Witt and Kasper, 1999;Zhang and Oakley, 1996). Temporal correlation, whichwould suggest dependence of taste bud development onnerve ingrowth, has not as yet been seen.
Lingual taste bud primordia first occur around the 7thand 8th postovulatory weeks (Bradley, 1972; Bradley andStern, 1967). Taste pores, commonly acknowledged as asign of taste bud maturity, appear between the 10th and 14thweek. The presence of a taste pore is not always associ-ated with a fully mature taste bud because the bottoms ofearly taste pits may be covered by flat epithelial cells (Wittand Reutter, 1997). However, transmission electron micro-scopical studies show that early taste bud primordia (week8) synaptically contact nerve fibers, suggesting the poten-tial for neurotransmission precedes the exposure of sapidmolecules to the apical surface of the taste bud cell. At itsbase, the developing human taste bud (weeks 12–15) con-tains processes of dark and light cells, as well as processesresembling type III cells (exhibiting synapses with nervefibers). At their apical ends, taste bud cells cannot be distin-guished by their electron density (week 15, Figure 29.13).There are cells with long, slender microvilli, but, in contrastto adult taste buds (Azzali, 1997), there are no type I cells

652 Chapter 29 Anatomy of the Tongue and Taste Buds
1
2
I
II
III
IV
VII
IX
X
XII
V3
3
4
5
Figure 29.11 Development of the human tongue, 5th postovulatory week. The schematic drawing was done by compiling Hinrichsen‘s (1990) andour own data. – By horizontal section the floor of the forecoming buccal cavity of a human embryo is removed and viewed from dorsally. The floorrelief is derived from the branchial arches I (mandibular arch), II (hyoidal arch), III (3rd pharyngeal arch) and IV (4th pharyngeal arch), and by theirderivatives which are 1- the paired lingual swellings, 2- the impar tubercle and 4- the hypobranchial eminence. These three structures form thetongue anlage. Between 2 and 4, the anlage of the thyroid gland invaginates, and as its remnant the 3- foramen caecum is left. 5- is the anlage of theepiglottis. The branchial arches, as well as the tongue anlage, are innervated by the cranial nerves V3 (mandibular nerve), VII (facial nerve), IX(glossopharyngeal nerve) and X (vagal nerve). (CN XII -hypoglossal nerve- invades the tongue anlage as well, and innervates its muscular system).Later, the lingual nerve (from V3) and the chorda tympani (running with VII) join each other and to supply the anterior two- thirds of the tonguewith somatosensory and gustatory nerve fibers, whereas IX and X carry taste fibers for the posterior third of the tongue, the epiglottis andthe pharynx.
Figure 29.12 Scanning electron micrograph of a humanembryonic tongue, 7th postovulatory week. Fine dots in the dorsalsurface demarcate later fungiform papillae. Anlagen of vallatepapillae (arrows) lie in front of the sulcus terminalis. Short arrowindicates the median vallate papillae, which originates first. Scalebar: 0.5 mm.

29.9 Innervation of the Human Tongue and Taste Buds 653
TP
mC
mC
mC
BL
N
N
N
N N
Figure 29.13 Transmission electron micrograph of a humanfungiform taste bud during development (week 15). The taste pore(TP) is already open, and some elongated cells stick into the taste pit.However, differences between cell types in the apical portion of thetaste bud cannot be made yet. There is no mucus in the taste pit.Approximately two- thirds of the taste bud are filled with ramificationsof nerve fibers (N). A part of the basal portion is outlined with arectangle (see Figure 29.14). mC = marginal cell, BL = basal lamina;scale bar: 10 μm. From Witt and Reutter (1996), with permission ofWiley & Sons.
with typical dark granules believed to secrete the mucousmaterial that fills the taste pit (Witt and Reutter, 1996).
29.9 INNERVATION OF THEHUMAN TONGUE AND TASTE BUDS
The tongue is innervated by (1) motor nerve fibers con-stituting the hypoglossal nerve (CN XII), which suppliesthe inner (intrinsic) and the hyoidal tongue muscles; (2)somatosensory nerve fibers, composed of divisions ofthe trigeminal (CN V3) and glossopharnygeal (CN IX)nerves; (3) autonomic nerve fibers which stem from theintermedio-facial nerve (CN VII), the glossopharyngealnerve, and the vagus nerve (CN X); and (4) sensory (gusta-tory) nerve fibers that transmit taste information centrally
N
NdC
BLBL
∗
Figure 29.14 Basal portion of a developing human taste bud, week15 (detail from Figure 29.13). Numerous processes of anelectron-dense cell (dC) rich in dense cored vesicles surround nervefibers (N), but synapses and signs of release of the vesicular contentsare not observed. One cell process is filled with dense-cored and clearvesicles (asterisk). BL = basal lamina. Bar = 1 μm. From Witt andReutter (1996), with permission of John Wiley & Sons.
from taste buds, namely (a) the chorda tympani, a branchof the intermediate nerve (generally considered a part ofthe facial nerve, CN VII), (b) the greater (=superficial)petrosal nerve, also part of the intermedio-facial complex,(c) glossopharyngeal nerve (CN IX), and (d) vagus nerve(CN X) (summarized in Figure 29.15).
Classical research papers of the nineteenth centuryexplored the clinical consequences in taste perceptionassociated with diagnostic features of the cranial nerves.These papers have formed the bases for the neurologi-cal examination. Two prominent themes were explored:(1) Which cranial nerves are associated with functionalattributes of the taste system? (2) How is taste perceptionaffected by neurological diseases? In physiological exper-iments, Magendie (1820) dissected the lingual nerve ofliving, unanaesthetized animals and observed loss of taste,but intact movement and sensation of palate, gingival, andbuccal mucosa. Panizza (1834) performed dissection of theglossopharyngeal nerve resulting in loss of taste. Alcock(1836) described the role of the chorda tympani and thesphenopalatine (pterygopalatine) ganglion. Lussana (1869,1872) traced the target tissue of the chorda tympani nerveto the anterior two-thirds of the tongue. The dependenceof taste buds on nerve supply was experimentally shownby von Vintschgau and Hönigschmied (1877): 40 daysafter dissection of the glossopharyngeal nerve, the numberof taste buds dramatically decreased. In similar experi-ments, Ranvier (1888) observed that taste bud sensory cellsdegenerate, and supporting cells pushed through the poreto the tongue surface. During this process, he observedcellules migratrices [phagocytic fibroblasts? (Suzuki et al.,1996)] loaded with fed particles, which were “probablyresponsible for removal of old material.” Soon it becameevident that experiments based on vivisection were not

654 Chapter 29 Anatomy of the Tongue and Taste Buds
V1
V2
V3
V
pG
LN
CT
gG VII
IXX
XII
iG iG
sG
Figure 29.15 Innervation of the human tongue and the tastebud-bearing epithelia (hatched regions), compiled from(Feneis, 1985) and (Sobotta, 1993). The cranial nerves VII(which includes the intermediate nerve with its branches,greater petrosal nerve and chorda tympani), IX and X containsensory gustatory fibers (yellow). V, trigeminal nerve (blue)with its divisions V1, ophthalmic, V2, maxillary, and V3,mandibular nerves. XII, hypoglossal nerve (red), is the motornerve of the intrinsic tongue muscles. gG, geniculate ganglion;pG, pterygopalatine (sphenopalatine) ganglion; iG inferiorganglion of the glossopharyngeal (IX) and vagal (X) nerves,respectively; sG, submandibular ganglion with postganglionicautonomic nerve fibers of the chorda tympani to supply thesubmandibular and sublingual glands. (See plate section forcolor version.)
only painful for the animal (mostly dogs or cats), but alsounreliable in their results (Alcock, 1836; Wagner, 1837).Nevertheless, the overall conclusions derived from thesenineteenth century nerve dissection experiments (reviewedby Jägel, 1991; Parker, 1922) cannot deny their importfor current knowledge of cranial nerve supply and tastesensitivity.
An elegant review of the background of peripheraltaste pathways in humans was written by Lewis andDandy (1930). They examined both the neurological andneuroanatomical literature on gustatory pathways. Thesensory distribution of the facial nerve and its clinicalimportance was described by Hunt (1915). He disentan-gled the overlapping sensory fields of the facial nerve(including the chorda tympani) from the trigeminal nerveby documenting the distribution of herpes zoster inflamma-tion. The herpetic eruptions outlined the sensory fields ofthe geniculate ganglion on the tongue, soft palate, and ear.Another basis for evaluating the involvement of the chordatympani nerve with lingual taste buds came from patientswho had undergone middle ear surgery (Borg et al., 1967;Bull, 1965). Contemporary reviews of human (Norgren,1990; Prichard, 2011) and primate (Pritchard, 1991) tastepathways have incorporated observations from the secondhalf of the twentieth century, particularly those derivedfrom electrophysiological studies.
1. Chorda tympani and greater petrosal nerve: Distalto the intermediate nerve branch of the facial nerve,peripheral axons of some geniculate ganglion somata(the chorda tympani nerve) take a recurrent coursewithin the facial canal in the petrosal part of the tem-poral bone, pass through the middle ear, and exit theskull via the petrotympanic fissure to join the lingualdivision of the trigeminal nerve, the lingual nerve.
Both intermedio-facial (gustatory) and trigeminal(somatosensory) fibers run in the lingual nerve anddistribute to the fungiform papillae on the anterior twothirds of the tongue and may reach also the anteriorportion of the foliate papillae. Taste buds on the softpalate are innervated by the greater petrosal branchof the intermedio-facial nerve, whose somata also liewithin the geniculate ganglion (Harris, 1952; Millerand Spangler, 1982). Some chorda tympani fibers arereported to anastomose with the greater petrosal nervevia the otic ganglion (Pritchard, 1991; Schwartz andWeddell, 1938). Both the greater petrosal and chordatympani nerves also carry parasympathetic fibers totheir associated salivary glands: The greater petrosalnerve serves the palatine and the lacrimal glands, whilethe chorda tympani innervates the submandibular andsublingual glands via the submandibular ganglion.
2. Glossopharyngeal nerve: Axons of the glossopharyn-geal nerve originate from ganglion cells mainly in theinferior (petrosal) glossopharyngeal ganglion. Theseperipheral axons supply both taste buds and generalsensory innervation to the vallate and foliate papillae.Salivary glands (Ebner) are supplied by parasympa-thetic fibers via an intrinsic ganglion (Remak, 1852).Probably, the glossopharyngeal nerve also suppliestaste buds in the pharynx. Bourgery and Jacob (1839)and Doty et al. (2009) observed that some CN IXfibers extend into the middle part of the tongue tooverlap with innervation areas of the chorda tympani.However, it is unclear if they innervate taste buds.
3. Vagus nerve: Taste buds on the laryngeal surface ofthe epiglottis, larynx, and proximal portion of theesophagus are innervated by the superior laryngeal

References 655
branch of the vagus nerve, which has the perikaryaof its chemosensory neurons in the inferior (nodose)vagal ganglion.
4. Trigeminal nerve: The possible role of trigeminalnerve fibers in taste perception has been discussedfor two millenia, and there is no end in sight. Viathe lingual nerve, this nerve conveys somatosensoryand viscerosensory information from the tongue totrigeminal brain stem nuclei. In fact, most of the nervefibers entering the fungiform papillae are trigeminal,while a few fibers originate from the chorda tympani(25% in rat: Farbman and Hellekant, 1978).
Trigeminal fibers may respond to sapid stimuli, asrevealed by trigeminal transection experiments (Berridgeand Fentress, 1985), electrophysiology (Harada and Smith,1992), and trigeminal ganglion cell response (Liu andSimon, 1998). Taste receptors also occur in a variety ofnongustatory cells proper, outside the oral cavity (Tizzanoet al., 2011). Finally, taste qualities may be influenced bynonsapid stimuli, for example, temperature: approximatelyone-half of the nerve fibers involved in taste transductionrespond to temperature (Cruz and Green, 2000). Theinteraction of both gustatory and somatosensory qualitiesmay be as tight-knit as their anatomical proximity. Katzet al. (2000) suggest that gustation should be thought ofas an integral part of a distributed, interacting multimodalsystem.
In contrast to most other sensory systems, gusta-tory function is distributed among three different cranialnerves, which makes taste difficult to eliminate and,secondly, also difficult to distinguish from trigeminalperception (Prichard, 2011).
The observation that taste buds degenerate after dis-section of their sensory innervation and, subsequently,reappear after regeneration of their peripheral nerves hasbeen a major focus of research in the peripheral tastesystem. Nineteenth- and early twentieth-century literatureon taste bud degeneration, regeneration, and developmentwas reviewed comprehensively by Parker (1922). Olmsted(1920) proposed that trophic maintenance of fish tastebuds depended on the transmission of a putative trophicmaterial from nerve to epithelium. Cross reinnervation ofthe glossopharyngeal nerve to fungiform taste buds (whichare normally supplied by the chorda tympani) had no effecton the usual immunohistochemical properties of fungiformversus vallate taste buds (Smith et al., 1999). As a conse-quence, these authors believe that the protein expressionin and subsequent function of taste buds depend on theepithelium from which the cells arise, and not the buds’specific nerve supply. Meanwhile, Nosrat and Olson (1995)and Nosrat et al. (2000) detected mRNA of brain-derivedneurotrophic factor (BDNF) and neurotrophins in devel-oping anterior tongue epithelium, before nerve fibers were
observed. In BDNF-overexpressing mice, larger tastebuds and more taste cells have been observed (Nosratet al., 2012). This argues for the hypothesis that trophicfactors act as target-derived chemoattractants for the earlynerve fibers. These, in turn, initiate the formation of tastebuds. BDNF-null mutant mice fail to develop taste buds(Oakley et al., 1998). Sensory ganglia involved in tastebud innervation (see above) are reduced by 40% in volumecompared to about 20% of trigeminal ganglion under thesame condition (Mistretta et al., 1999). Taste buds do notdevelop after injection of ß-bungarotoxin into the amnioticfluid in fetal mice. This neurotoxin abolishes motor andsensory nerve development (Morris-Wiman et al., 1999).
Although nerve fibers are required to maintain tastebuds once the latter are formed and start to function (e.g.,Hosley et al., 1987; Yee et al., 2005), nerves seem not to benecessary to initiate taste bud development. Initial taste buddevelopment is nerve-independent, suggested by a seriesof studies in salamanders (Stone, 1940), axolotl (Barlowet al., 1996; Barlow and Northcutt, 1998a,b) and mouse(Mbiene and Roberts, 2003). Taste buds seem to developfrom local epithelium and not from neurogenic ectoderm[axolotl: Barlow and Northcutt (1997), mouse: Stone et al.(1995)]. It may be that mechanisms of differentiation of thesame receptor organ vary among vertebrate taxa (Barlow,2003). Growth factors other than BDNF may contribute tothe maintenance of gustatory papillae, for example, epi-dermal growth factor (EGF) supplied by salivary glands(Liu et al., 2008; Morris-Wiman et al., 2000). More detailedstudies on developmental aspects of the peripheral gusta-tory system including whether taste buds may develop with-out the stimulation of nerves are described later in this book(Chapter 33).
ACKNOWLEDGEMENTS
The authors are indebted to Dr. Inglis Miller, Jr. who wrotea previous version of this chapter in the first edition, andMihnea Nicolescu, who provided the schematic drawings.Drs. Judith and Donald Ganchrow helped with critical read-ing of an earlier version of the manuscript.
REFERENCES
Albinus (1754). Academicarum annotationum libri I-VIII. liber I, Tab.I,Lugdunum Bataviensis (Leiden).
Alcock, B. (1836). Determination of the question, which are the nerves oftaste. Dublin. J. Med. Chem. Sci. 10: 256–279.
Arey, L., Tremaine, M., and Monzingo, F. (1935). The numerical and topo-graphical relation of taste buds to human circumvallate papillae through-out the life span. Anat. Rec. 64: 9–25.
Arnold, F. (1839). Tabulae Anatomicae. Icones organum sensuum.Organon gustus. Fasc. secundus. Orellii, Fuesslini, Zürich.

656 Chapter 29 Anatomy of the Tongue and Taste Buds
Arvidson, K., and Friberg, U. (1980). Human taste: response and taste budnumber in fungiform papillae. Science 209: 807–808.
Astbäck, J., Arvidson, K., and Johansson, O. (1997). An immunohisto-chemical screening of neurochemical markers in fungiform papillae andtaste buds of the anterior rat tongue. Arch. Oral. Biol. 42(2): 137–147.
Atema, J. (1971). Structures and functions of the sense of taste in the catfish(Ictalurus natalis). Brain. Behav. Evol. 4(4): 273–294.
Azen, E. A., Hellekant, G., Sabatini, L. M., and Warner, T. F. (1990).mRNAs for PRPs, statherin, and histatins in von Ebner’s gland tissues.J. Dent. Res. 69: 1724–1730.
Azzali, G., Gennari, P. U., Maffei, G., and Ferri, T. (1996). Vallate, foliateand fungiform human papillae gustatory cells. An immunocytochemicaland ultrastructural study. Minerva Stomatol. 45(9): 363–379.
Azzali, G. (1997). Ultrastructure and immunocytochemistry of gustatorycells in man. Anat. Anz. 179(1): 37–44.
Barlow, L. A. (2003). Toward a unified model of vertebrate taste bud devel-opment. J. Comp. Neurol. 457(2): 107–110.
Barlow, L. A., Chien, C. B., and Northcutt, R. G. (1996). Embryonictaste buds develop in the absence of innervation. Development 122:1103–1111.
Barlow, L. A., and Northcutt, R. G. (1997). Taste buds developautonomously from endoderm without induction by cephalic neural crestor paraxial mesoderm. Development 124(5): 949–957.
Barlow, L. A., and Northcutt, R. G. (1998a). The role of innervation in thedevelopment of taste buds: insights from studies of amphibian embryos.Ann. N. Y. Acad. Sci. 855: 58–69.
Barlow, L. A., and Northcutt, R. G. (1998b). Vertebrate taste-bud devel-opment: are salamanders the model? Reply. Trends Neurosci. 21(8):338–338.
Barreiro-Iglesias, A., Anadon, R., and Rodicio, M. C. (2010). The gustatorysystem of lampreys. Brain Behav. Evol. 75(4): 241–250.
Bartel, D. L., Sullivan, S. L., Lavoie, E. G., et al.(2006). Nucleoside triphos-phate diphosphohydrolase-2 is the ecto-ATPase of type I cells in tastebuds. J. Comp. Neurol. 497(1): 1–12.
Behrens, M., Born, S., Redel, U., et al. (2012). ImmunohistochemicalDetection of TAS2R38 Protein in Human Taste Cells. PLoS One 7(7):e40304.
Beidler, L. M., and Smallman, R. L. (1965). Renewal of cells within tastebuds. J. Cell Biol. 27: 263–272.
Bellini, L. (1665). Gustus organum novissime deprehensum praemissisad faciliorem intelligentiam quibusdam de saporibus (Taste organsnewly observed; with certain premises about the senses given for easierunderstanding (translation from Latin into German by Jurisch (1922)).Mangetus Bibliotheca anat.2, Bologna.
Berkhoudt, H. (1985). Special sense organs: Structure and function of aviantaste receptors. In Form and function in birds, King, A. S. and McIelland,J. (Eds). Vol. 3, Academic Press, New York, pp. 463–496.
Berridge, K. C., and Fentress, J. C. (1985). Trigeminal-taste interaction inpalatability processing. Science 228(4700): 747–750.
Borg, G., Diamant, H., Oakley, B., et al. (1967). A comparative study ofneural and psychophysical responses to gustatory stimuli. In Olfactionand Taste 2, Hayashi, T. (Ed). Pergamon Press, Oxford, pp. 253–265.
Boudriot, F., and Reutter, K. (2001). Ultrastructure of the taste buds inthe blind cave fish Astyanax jordani (“Anoptichthys”) and the sightedriver fish Astyanax mexicanus (Teleostei, Characidae). J. Comp. Neurol.434(4): 428–444.
Boughter Jr.,, J. D., Pumplin, D. W., Yu, C., et al. (1997). DifferentialExpression of alpha—Gustducin in Taste Bud Populations of the Rat andHamster. J. Neurosci. 17(8): 2852–2858.
Bourgery, J. M., and Jacob, N. H. (1839). Organes de la digestion, de ladépuration urinaire et de la génération. Embryotomie. In Traité completde l’anatomie de l’homme, Delaunay, C. A. (Ed). Vol. 5, Jules DidotL’Ainé, Paris, pp.
Bradley, R. M. (1972). Development of the taste bud and gustatory papillaein human fetuses. In Third symposium on oral sensation and percep-tion, Bosma, J. F. (Ed). Charles C Thomas, Springfield, Ill., USA, pp.137–162.
Bradley, R. M. (1973). Electrophysiological investigations of intravasculartaste using perfused rat tongue. Am. J. Physiol. 224(2): 300–304.
Bradley, R. M. (2000). Sensory receptors of the larynx. Am. J. Med. 108Suppl 4a: 47S–50S.
Bradley, R. M., Stedman, H. M., and Mistretta, C. M. (1983). Supe-rior laryngeal nerve response patterns to chemical stimulation of sheepepiglottis. Brain Res. 276(1): 81–93.
Bradley, R. M., and Stern, I. B. (1967). The development of the human tastebud during the foetal period. J. Anat. 101: 743–752.
Braus, H. (1940). Handbuch der Mikroskopischen Anatomie des Menschen.Springer, Berlin.
Bull, T. R. (1965). Taste and the chorda tympani. J. Laryngol. Otol. 79:479–493.
Burkl, W. (1954). Über das Vorkommen von Geschmacksknospen im mit-tleren Drittel des Oesophagus. Anat. Anz. 100: 320–321.
Caicedo, A., Kim, K. N., and Roper, S. D. (2000). Glutamate-inducedcobalt uptake reveals non-NMDA receptors in rat taste cells. J. Comp.Neurol. 417(3): 315–324.
Cao, Y., Zhao, F. L., Kolli, T., et al. (2009). GABA expression in the mam-malian taste bud functions as a route of inhibitory cell-to-cell communi-cation. Proc. Natl. Acad. Sci. U.S.A. 106(10): 4006–4011.
Carus, C. G. (1849). System der Physiologie. 2nd ed. Brockhaus, Leipzig.
Casserius (1609). Penthaesteseion (cited after Jurisch, 1922).
Chaudhari, N., and Roper, S. D. (2010). The cell biology of taste. J. CellBiol. 190(3): 285–296.
Cheng, L. H., and Robinson, P. P. (1991). The distribution of fungiformpapillae and taste buds on the human tongue. Arch. Oral Biol. 36:583–589.
Cheng, S. J., Huang, C. F., Chen, Y. C., et al. (2009). Ultrastructural changesof posterior lingual glands after hypoglossal denervation in hamsters. J.Anat. 214(1): 163–170.
Clapp, T. R., Stone, L. M., Margolskee, R. F., and Kinnamon, S. C. (2001).Immunocytochemical evidence for co-expression of Type III IP3 recep-tor with signaling components of bitter taste transduction. BMC Neu-rosci. 2: 6.
Clapp, T. R., Yang, R., Stoick, C. L., et al. (2004). Morphologic char-acterization of rat taste receptor cells that express components of thephospholipase C signaling pathway. J. Comp. Neurol. 468(3): 311–321.
Connes, R., Granie-Prie, M., Diaz, J. P., and Paris, J. (1988). Ultrastructuredes bourgeons du gout du téléostéen marin Dicentrarchus labrax L. Can.J. Zool. 66: 2133–2142.
Cottler Fox, M., Arvidson, K., Hammarlund, E., and Friberg, U. (1987).Fixation and occurrence of dark and light cells in taste buds of fungiformpapillae. Scand. J. Dent. Res. 95: 417–427.
Cruz, A., and Green, B. G. (2000). Thermal stimulation of taste. Nature403(6772): 889–892.
Cummings, T. A., Delay, R. J., and Roper, S. D. (1987). Ultrastructure ofapical specializations of taste cells in the mudpuppy, Necturus maculo-sus. J. Comp. Neurol. 261(4): 604–615.
Dando, R., and Roper, S. D. (2009). Cell-to-cell communication inintact taste buds through ATP signalling from pannexin 1 gap junctionhemichannels. J. Physiol. 587(Pt 24): 5899–5906.

References 657
Davis, C. (1877). Die becherförmigen Organe des Kehlkopfs. Arch. mikr.Anat. 14: 158–167.
DeFazio, R. A., Dvoryanchikov, G., Maruyama, Y., et al. (2006). Separatepopulations of receptor cells and presynaptic cells in mouse taste buds.J. Neurosci. 26(15): 3971–3980.
Delay, R. J., Kinnamon, S. C., and Roper, S. D. (1997). Serotonin modu-lates voltage-dependent calcium current in Necturus taste cells. J. Neu-rophysiol. 77(5): 2515–2524.
Delay, R. J., Mackay Sim, A., and Roper, S. D. (1994). Membrane prop-erties of two types of basal cells in Necturus taste buds. J. Neurosci. 14:6132–6143.
Delay, R. J., and Roper, S. D. (1988). Ultrastructure of taste cells andsynapses in the mudpuppy Necturus maculosus. J. Comp. Neurol. 277:268–280.
Delay, R. J., Taylor, R., and Roper, S. D. (1993). Merkel-like basal cellsin Necturus taste buds contain serotonin. J. Comp. Neurol. 335(4):606–613.
Desgranges, J. C. (1965). Sur l’existence de plusieurs types de cellules sen-sorielles dans les bourgeons du gout des barbillons du Poisson-chat. C.R.Acad. Sc.(Paris) 261: 1095–1098.
Doty, R. L., Cummins, D. M., Shibanova, A., et al. (2009). Lingual distri-bution of the human glossopharyngeal nerve. Acta Otolaryngol. 129(1):52–56.
Dvoryanchikov, G., Sinclair, M. S., Perea-Martinez, I., et al. (2009). Inwardrectifier channel, ROMK, is localized to the apical tips of glial-like cellsin mouse taste buds. J. Comp. Neurol. 517(1): 1–14.
Ebner, V. (1873). Die acinösen Drüsen der Zunge und ihre Beziehungen zuden Geschmacksorganen. Leuschner & Lubensky, Graz.
Ehrenberg, C. G. (1833). Notwendigkeit einer feineren mechanischen Zer-legung des Gehirns und der Nerven vor der chemischen, dargestellt anBeobachtungen von C.G.Ehrenberg. Poggendorfs Annl. Physik. Chemie.XXVIII: 450.
Ehrlich, P. (1886). Über die Methylenblaureaktion der lebenden Nerven-substanz. Deutsche Med. Wochenschr. 12: 49–52.
Engelmann, T. W. (1872). Die Geschmacksorgane. In Handbuch der Lehrevon den Geweben des Menschen und der Thiere, Stricker, S. (Ed). Vol.2, Engelmann, Leipzig, pp. 822–838.
Engström, H., and Rytzner, C. (1956). The fine structure of taste buds andtaste fibers. 361–375 65.
Fährmann, W. (1967). [Light and electron microscopical studies on the tastebud of neotene axolotls (Siredon mexicanum Shaw)]. Z. Mikrosk. Anat.Forsch. 77: 117–152.
Farbman, A. I. (1965). Electron microscope study of the developing tastebud in rat fungiform papilla. Dev. Biol. 11: 110–135.
Farbman, A. I. (1980). Renewal of taste bud cells in rat circumvallate papil-lae. Cell Tissue Kinet. 13: 349–357.
Farbman, A. I., and Hellekant, G. (1978). Quantitative analyses of the fiberpopulation in rat chorda tympani nerves and fungiform papillae. Am. J.Anat. 153: 509–521.
Farbman, A. I., Hellekant, G., and Nelson, A. (1985). Structure of tastebuds in foliate papillae of the rhesus monkey, Macaca mulatta. Am. J.Anat. 172(1): 41–56.
Farbman, A. I., Ogden Ogle, C. K., Hellekant, G., et al. (1987). Labelingof sweet taste binding sites using a colloidal gold- labeled sweet protein,thaumatin. Scanning. Microsc. 1: 351–357.
Farbman, A. I., and Yonkers, J. D. (1971). Fine structure of the taste bud inthe mud puppy, Necturus maculosus. Am. J. Anat. 131: 353–369.
Feneis, H. (1985). Pocket Atlas of human anatomy. Thieme, New York.Ferrando, S., Gallus, L., Gambardella, C., et al. (2012). First detection of
taste buds in a chimaeroid fish (Chondrichthyes: Holocephali) and theirGalphai-like immunoreactivity. Neurosci. Lett. 517(2): 98–101.
Finger, T. E. (1997). Evolution of taste and solitary chemoreceptor cell sys-tems. Brain Behav. Evol. 50(4): 234–243.
Finger, T. E., Bottger, B., Hansen, A., et al. (2003). Solitary chemoreceptorcells in the nasal cavity serve as sentinels of respiration. Proc. Natl. Acad.Sci. U.S.A. 100(15): 8981–8986.
Finger, T. E., Bryant, B. P., Kalinoski, D. L., et al. (1996). Differentiallocalization of putative amino acid receptors in taste buds of the channelcatfish, Ictalurus punctatus. J. Comp. Neurol. 373(1): 129–138.
Finger, T. E., Danilova, V., Barrows, J., et al. (2005). ATP signaling iscrucial for communication from taste buds to gustatory nerves. Science310(5753): 1495–1499.
Fixsen, C. (1857). De linguae raninae textura disquisitiones microscopi-cae. Diss. inaug. Dorpati Livonor.
Froriep, R. (1828). De lingua anatomica quaedam et semiotica.
Fujimoto, S., Ueda, H., and Kagawa, H. (1987). Immunocytochemistry onthe localization of 5-hydroxytryptamine in monkey and rabbit taste buds.Acta Anat. 128: 80–83.
Fukami, H., and Bradley, R. M. (2005). Biophysical and morphologicalproperties of parasympathetic neurons controlling the parotid and vonEbner salivary glands in rats. J. Neurophysiol. 93(2): 678–686.
Ganchrow, D., Ganchrow, J. R., and Goldstein, R. S. (1991). Ultrastruc-ture of palatal taste buds in the perihatching chick. Am. J. Anat. 192(1):69–78.
Ganchrow, J. R., and Ganchrow, D. (1987). Taste bud development inchickens (Gallus gallus domesticus). Anat. Rec. 218: 88–93.
Ganchrow, J. R., Ganchrow, D., and Oppenheimer, M. (1986). Chorda tym-pani innervation of anterior mandibular taste buds in the chicken (Gallusgallus domesticus). Anat. Rec. 216(3): 434–439.
Ganchrow, J. R., Ganchrow, D., Royer, S. M., and Kinnamon, J. C. (1993).Aspects of vertebrate gustatory phylogeny: morphology and turnover ofchick taste bud cells. Microsc. Res. Tech. 26: 106–119.
Gerlach, J. (1858). Mikroskopische Studien aus dem Gebiet der men-schlichen Morphologie. F. Enke, Erlangen.
Getchell, T. V., Margolis, F. L., and Getchell, M. L. (1984). Perireceptor andreceptor events in vertebrate olfaction. Prog. Neurobiol. 23: 317–345.
Golgi, C. (1873). Sulla struttura della sostanza grigia del cervello (comuni-cazione preventiva). Gazzetta Medica Italiana-Lombardia 33: 244 –246.
Gråberg, J. (1898). Beiträge zur Genese des Geschmacksorgans des Men-schen. Morphol. Arb. 8: 117–134.
Graziadei, P. P., and DeHan, R. S. (1971). The ultrastructure of frogs’ tasteorgans. Acta Anat. (Basel) 80(4): 563–603.
Grover Johnson, N., and Farbman, A. I. (1976). Fine structure of taste budsin the barbel of the catfish, Ictalurus punctatus. Cell Tissue Res. 169:395–403.
Gurkan, S., and Bradley, R. M. (1987). Autonomic control of von Ebner’slingual salivary glands and implications for taste sensation. Brain Res.419: 287–293.
Habermehl, K. H. (1952). Über besondere Randpapillen an der Zungeneugeborener Säugetiere. Z. Anat. Entwickl. Gesch. 116: 355–372.
Haller, A. v. (1766). Gustus. In Elementa physiologiae Vol. IV, V, F.Grasset,Lausanne, pp. 99–124.
Hamasaki, K., Seta, Y., Yamada, K., and Toyoshima, K. (1998). Possiblerole of serotonin in Merkel-like basal cells of the taste buds of the frog,Rana nigromaculata. J. Anat. 193(4): 599–610.
Hannover, A. (1840). Die Chromsäure, ein vorzügliches Mittel beimikroskopischen Untersuchungen. Müllers Archiv: 549–558.
Harada, S., and Smith, D. V. (1992). Gustatory sensitivities of the hamster’ssoft palate. Chem. Senses 17: 37–51.
Harris, W. (1952). The fifth and seventh nerves in relation to the nervousmechanism of taste sensation: a new approach. Brit. Med. J. 1: 831–836.

658 Chapter 29 Anatomy of the Tongue and Taste Buds
Hellekant, G. (1976). The blood circulation of the tongue. In Frontiers ofOral Physiology, Kawamura, Y. (Ed). S. Karger, Basel, pp. 130–145.
Hellman, T. J. (1922). Die Genese der Zungenpapillen beim Menschen.Ups. Läkaref. Förh. 26,5–6: 1–69.
Helmholtz (1842). De fabrica systematis nervosi evertebratorum. Disser-tation, University of Berlin.
Hermann, F. (1885). Beittrag zur Entwicklungsgeschichte des Geschmack-sorgans beim Kaninchen. Arch. f. Mikroskop. Anat. Entwicklungsmech.24: 216–229.
Herness, M. S. (1989). Vasoactive intestinal peptide-like immunoreactivityin rodent taste cells. Neuroscience 33(2): 411–419.
Herness, S., Zhao, F. L., Lu, S. G., et al. (2002). Expression and physio-logical actions of cholecystokinin in rat taste receptor cells. J. Neurosci.22(22): 10018–10029.
Herrick, C. J. (1904). The organ and sense of taste in fishes. U.S. FishComm. Bull. 1902 (abstracted in J. Comp. Neurol. Psych. 277–278,1904): 237–272.
Hersch, M., and Ganchrow, D. (1980). Scanning electron microscopy onhuman embryonic and fetal tongue. Chem. Senses 5: 331–341.
Hinrichsen, K. V. (1990). Humanembryologie. Lehrbuch und Atlas dervorgeburtlichen Entwicklung des Menschen. (Hinrichesen, K. V., ed.)Berlin, Heidelberg, New York, Springer Verlag.
Hirata, Y. (1966). Fine structure of the terminal buds on the barbels of somefishes. Arch. Histol. Jpn. 26(5): 507–523.
Höfer, D., and Drenckhahn, D. (1999). Localisation of actin, villin, fimbrin,ezrin and ankyrin in rat taste receptor cells. Histochem. Cell Biol. 112(1):79–86.
Hofer, H., Meinel, W., and Rommel, C. (1979). Taste buds in the epithe-lium of the plica sublingualis of New World monkeys. Anat. Anz. 145(1):17–31.
Hoffmann, A. (1875). Ueber die Verbreitung der Geschmacksknospen beimMenschen. Arch. pathol. Anat. Physiol. klin. Med. 62: 516–530.
Hokfelt, T., Lundberg, J. M., Schultzberg, M., et al. (1980). Cellular local-ization of peptides in neural structures. Proc. R. Soc. Lond B. Biol. Sci.210(1178): 63–77.
Hönigschmied, J. (1873). Beiträge zur mikroskopischen Anatomie überdie Geschmacksorgane der Säugethiere. Zeitschr. Wiss. Zoologie 23:414–434.
Hoon, M. A., Adler, E., Lindemeier, J., et al. (1999). Putative mammaliantaste receptors: a class of taste-specific GPCRs with distinct topographicselectivity. Cell 96(4): 541–551.
Hoon, M. A., Northup, J. K., Margolskee, R. F., and Ryba, N. J. (1995).Functional expression of the taste specific G-protein, alpha-gustducin.Biochem. J. 309(Pt 2): 629–636.
Hosley, M. A., Hughes, S. E., and Oakley, B. (1987). Neural induction oftaste buds. J. Comp. Neurol. 260: 224–232.
Hou-Jensen, H. (1933). Die Papillae foliatae des Menschen. Z. Anat.Entwicklgesch. 102: 348–388.
Hu, Z. L., Masuko, S., and Katsuki, T. (1996). Distribution and originsof nitric oxide-producing nerve fibers in the dog tongue: correlatedNADPH-diaphorase histochemistry and immunohistochemistry for cal-citonin gene-related peptide using light and electron microscopy. Arch.Histol. Cytol. 59(5): 491–503.
Huang, Y.-J., Maruyama, Y., Lu, K.-S., et al. (2005). Mouse Taste BudsUse Serotonin as a Neurotransmitter. J. Neurosci. 25(4): 843–847.
Huang, Y. A., Dando, R., and Roper, S. D. (2009). Autocrine and paracrineroles for ATP and serotonin in mouse taste buds. J. Neurosci. 29(44):13909–13918.
Huang, Y. A., Maruyama, Y., Stimac, R., and Roper, S. D. (2008). Presy-naptic (Type III) cells in mouse taste buds sense sour (acid) taste. J.Physiol. 586(12): 2903–2912.
Hunt, J. R. (1915). The sensory field of the facial nerve: a further con-tribution to the symptomatology of the geniculate ganglion. Brain 38:418–446.
Iida, M., Yoshioka, I., and Muto, H. (1983). Taste bud papillae on the retro-molar mucosa of the rat, mouse and golden hamster. Acta Anat. Basel.117: 374–381.
Ito, A., and Nosrat, C. A. (2009). Gustatory papillae and taste bud develop-ment and maintenance in the absence of TrkB ligands BDNF and NT-4.Cell Tissue Res. 337(3): 349–359.
Iwasaki, S., Yoshizawa, H., and Kawahara, I. (1996). Three-dimensionalultrastructure of the surface of the tongue of the rat snake, Elaphe clima-cophora. Anat. Rec. 245(1): 9–12.
Jägel. (1991). Zur Geschichte der Anatomie und Physiologie desGeschmackssinnes. Diss. Univ. Kiel: 1–118.
Jaeger, C.B., Hillmann, D.E. (1976). Gustatory system. Morphology ofgustatory organs. In Frog neurobiology A Handbook, R. Llinás, andW. Precht, eds. (Berlin, Heidelberg, New York: Springer), pp. 588–606.
Jain, S., and Roper, S. D. (1991). Immunocytochemistry of gamma-amino-butyric acid, glutamate, serotonin, and histamine in Necturus taste buds.J. Comp. Neurol. 307(4): 675–682.
Jakubowski, and Whitear, M. (1986). Ultrastructure of taste buds in fishes.Folia Histochem. Cytobiol. 24: 310–311.
Jowett, A., and Shrestha, R. (1998). Mucosa and taste buds of the humanepiglottis. J. Anat. 193(4): 617–618.
Jeffery, W. R., Strickler, A. G., Guiney, S., et al. (2000). Prox 1 in eyedegeneration and sensory organ compensation during development andevolution of the cavefish Astyanax. Dev. Genes. Evol. 210(5): 223–230.
Jurisch, A. (1922). Studien über die Papillae vallatae beim Menschen. Z.Anat. Entwicklungsgesch. 66: 1–149.
Just, T., Pau, H. W., Bombor, I., et al. (2005). Confocal microscopy ofthe Peripheral Gustatory System: Comparison between Healthy Subjectsand Patients Suffering from Taste Disorders during Radiochemotherapy.Laryngoscope 115(12): 2178–2182.
Kaya, N., Shen, T., Lu, S.-g., et al. (2004). A paracrine signaling role forserotonin in rat taste buds: expression and localization of serotonin recep-tor subtypes. American Journal of Physiology-Regulatory, Integrativeand Comparative Physiology 286(4): R649–R658.
Katz, D. B., Nicolelis, M. A., and Simon, S. A. (2000). IV. There is more totaste than meets the tongue. Am. J. Physiol. Gastrointest Liver Physiol.278(1): G6–G9.
Kim, D. J., and Roper, S. D. (1995). Localization of serotonin in taste buds:a comparative study in four vertebrates. J. Comp. Neurol. 353: 364–370.
Kim, M. R., Kusakabe, Y., Miura, H., et al. (2003). Regional expressionpatterns of taste receptors and gustducin in the mouse tongue. Biochem.Biophys. Res. Commun. 312(2): 500–506.
Kinnamon, J. C., Henzler, D. M., and Royer, S. M. (1993). HVEM ultra-structural analysis of mouse fungiform taste buds, cell types, and asso-ciated synapses. Microsc. Res. Tech. 26: 142–156.
Kinnamon, J. C., McPheeters, M. M., and Kinnamon, S. C. (1994).Structure/Function correlates in taste buds. In: Olfaction and Taste(K.Kurihara, N Suzuki, H Ogawa, eds), Springer-Verlag, Tokyo, Berlin,Heidelberg, New York, London, Paris XI: 9–12.
Kinnamon, J. C., Sherman, T. A., and Roper, S. D. (1988). Ultrastructure ofmouse vallate taste buds: III. Patterns of synaptic connectivity. J. Comp.Neurol. 270: 1–10, 56.
Kinnamon, J. C., Taylor, B. J., Delay, R. J., and Roper, S. D. (1985). Ultra-structure of mouse vallate taste buds. I. Taste cells and their associatedsynapses. J. Comp. Neurol. 235: 48–60.
Kinnamon, S. C. (2011). Taste receptor signalling—from tongues to lungs.Acta Physiol. (Oxf) 204(2): 158–168.

References 659
Kishi, M., Emori, Y., Tsukamoto, Y., and Abe, K. (2001). Primary culture ofrat taste bud cells that retain molecular markers for taste buds and permitfunctional expression of foreign genes. Neuroscience 106(1): 217–225.
Kishi, M., Emori, Y., Tsukamoto, Y., and Abe, K. (2002). Changes in cellmorphology and cell-to-cell adhesion induced by extracellular Ca2+ incultured taste bud cells. Biosci. Biotechnol. Biochem. 66(2): 484–487.
Koelliker, R. A. (1844). Die Selbständigkeit und Abhängigkeit des sym-pathischen Nervensystems durch anatomische Beobachtungen bewiesen.Ein akademisches Programm. U.Zeller, Zürich.
Korte, G. E. (1979). Unusual association of ‘chloride cells’ with anothercell type in the skin of the glass catfish, Kryptopterus bicirrhis. TissueCell 11(1): 63–68.
Kotrschal, K. (1996). Solitary chemosensory cells: Why do primary aquaticvertebrates need another taste system. Trends Ecol.Evol. 11: 110–114.
Krause, R. (1911). Kursus der normalen Histologie. (American edition:Rebman Co., N.Y.) ed. Urban und Schwarzenberg, Berlin and Wien.
Kudo, K., Wakamatsu, K., Nishimura, S., and Tabata, S. (2010). Gustducinis expressed in the taste buds of the chicken. Anim. Sci. J. 81(6): 666–672.
Lalonde, E., and Eglitis, J. (1961). Number and distribution of taste buds onthe epiglottis, pharynx, larynx, soft palate and uvula in a human newborn.Anat.Rec. 140: 91–95.
Landin, A. M., Kim, J. W., and Chaudhari, N. (2005). Liposome-mediatedtransfection of mature taste cells. J. Neurobiol. 65(1): 12–21.
Lawton, D. M., Furness, D. N., Lindemann, B., and Hackney, C. M. (2000).Localization of the glutamate-aspartate transporter, GLAST, in rat tastebuds. Eur. J. Neurosci. 12(9): 3163–3171.
Lewis, D., and Dandy, W. E. (1930). The course of the nerve fibers trans-mitting sensation of taste. Arch.Surg. 21: 249–288.
Leydig, F. (1851). Über die Haut einiger Süßwasserfische. Z. Wiss. Zool. 3:1–12.
Li, J. H., and Lindemann, B. (2003). Multi-photon microscopy of cell typesin the viable taste disk of the frog. Cell Tissue Res. 313(1): 11–27.
Li, X. J., and Snyder, S. H. (1995). Molecular cloning of Ebnerin, a vonEbner’s gland protein associated with taste buds. J. Biol. Chem. 270(30):17674–17679.
Lin, W., and Kinnamon, S. C. (1999). Co-localization of epithelial sodiumchannels and glutamate receptors in single taste cells. Biol. SignalsRecept. 8(6): 360–365.
Lindemann, B. (1996). Taste reception. Physiol. Rev. 76: 719–766.
Linden, R. W. (1993). Taste. Br. Dent J. 175(7): 243–253.
Liu, H. X., Henson, B. S., Zhou, Y., et al. (2008). Fungiform papilla pat-tern: EGF regulates inter-papilla lingual epithelium and decreases papillanumber by means of PI3K/Akt, MEK/ERK, and p38 MAPK signaling.Dev. Dyn. 237(9): 2378–2393.
Liu, L., and Simon, S. A. (1998). Responses of cultured rat trigeminal gan-glion neurons to bitter tastants. Chem. Senses 23(2): 125–130.
Lovén, C. (1868). Beiträge zur Kenntnis vom Bau der Geschmack-swärzchen der Zunge. Arch. Mikrosk. Anat. IV: 96–110.
Lussana, F. (1869). Recherches expérimentales et observationspathologiques sur les nerfs du gout. Arch. phys. (Paris) 2: 197–210.
Lussana, F. (1872). Sur les nerfs du gout. Observations et expériences nou-velles. Arch. phys. (Paris) 4: 334–350.
Ma, H., Yang, R., Thomas, S. M., and Kinnamon, J. C. (2007). Qualitativeand quantitative differences between taste buds of the rat and mouse.BMC Neurosci. 8: 5.
Magendie, F. (1820). Grundriss der Physiologie (translated from Frenchby C.F. Heusinger). Baercke, Eisenach.
Malpighi, M. (1686). Exercitatio epistolica de lingua. ( 1664; Jo. AlphonsoBorellio). In Opera omnia, Malpighi, M. (Ed). R.Scott & G. Wells, Lon-dini, pp. 13–20.
Matsunami, H., Montmayeur, J. P., and Buck, L. B. (2000). A family of can-didate taste receptors in human and mouse. Nature 404(6778): 601–604.
Mbiene, J. P., and Roberts, J. D. (2003). Distribution of keratin 8-containingcell clusters in mouse embryonic tongue: evidence for a prepattern fortaste bud development. J. Comp. Neurol. 457(2): 111–122.
McLaughlin, S. K., McKinnon, P. J., and Margolskee, R. F. (1992). Gust-ducin is a taste-cell-specific G protein closely related to the transducins.Nature 357(6379): 563–569.
Menco, B. P. (1989). Olfactory and nasal respiratory epithelia, and foliatetaste buds visualized with rapid-freeze freeze-substitution and LowicrylK11M embedding. Ultrastructural and initial cytochemical studies.Scanning. Microsc. 3: 257–272.
Menco, B. P. M., Yankova, M. P., and Simon, S. A. (1997). Freeze- sub-stitution and postembedding immunocytochemistry on rat taste buds:G-proteins, calcitonin gene-related peptide, and choline acetyl trans-ferase. Microsc. Microanal. 3: 53–69.
Merkel, F. (1880). Ueber die Endigungen der sensiblen Nerven in der Hautder Wirbelthiere. Stiller, Rostock.
Miller, I. J., Jr., and Bartoshuk, L. M. (1991). Taste perception, taste buddistribution, and spatial relationships. 1 ed. In Smell and taste in healthand disease, Getchell, T. V., Bartoshuk, L. M., Doty, R. L. and Snow, J.B. J., Jr., (Eds). New York, Raven Press, pp. 205–233.
Miller, I. J., Jr., and Reedy, F. E., Jr., (1990a). Variations in human taste buddensity and taste intensity perception. Physiol. Behav. 47(6): 1213–1219.
Miller, I. J., Jr., and Reedy, F. E. J. (1990b). Quantification of fungiformpapillae and taste pores in living human subjects. Chem. Senses 15:281–294.
Miller, I. J., Jr., and Smith, D. V. (1984). Quantitative taste bud distributionin the hamster. Physiol.Behav. 32: 275–285.
Miller, I. J., Jr., and Spangler, K. M. (1982). Taste bud distribution andinnervation on the palate of the rat. Chem. Senses 7: 99–108.
Mistretta, C. M., and Baum, B. J. (1984). Quantitative study of taste budsin fungiform and circumvallate papillae of young and aged rats. J. Anat.138: 323–332.
Mistretta, C. M., Goosens, K. A., Farinas, I., and Reichardt, L. F. (1999).Alterations in size, number, and morphology of gustatory papillae andtaste buds in BDNF null mutant mice demonstrate neural dependence ofdeveloping taste organs. J. Comp. Neurol. 409(1): 13–24.
Miura, H., Scott, J. K., Harada, S., Barlow, L. A. (2014). Sonichedgehog-expressing basal cells are general post-mitotic precursors offunctional taste receptor cells. Dev. Dyn. 243: 1286–1297.
Mochizuki, Y. (1939). Studies of the papilla foliata of Japanese. OkajimasFolia Anat. Jpn. 18: 334–369.
Moll, R. (1993). Cytokeratins as markers of differentiation: Expression pro-files in epithelia and epithelial tumors. In Progress in Pathology, Seifert,G. (Ed). Vol. 142, G. Fischer, Stuttgart, Jena, New York, pp. 1–197.
Montavon, P., Hellekant, G., and Farbman, A. (1996). Immunohisto-chemical, electrophysiological, and electron microscopical study of ratfungiform taste buds after regeneration of chorda tympani through thenon-gustatory lingual nerve. J. Comp. Neurol. 367: 491–502.
Morris-Wiman, J., Basco, E., and Du, Y. (1999). The effects ofbeta-bungarotoxin on the morphogenesis of taste papillae and taste budsin the mouse. Chem. Senses 24(1): 7–17.
Morris-Wiman, J., Sego, R., Brinkley, L., and Dolce, C. (2000). The effectsof sialoadenectomy and exogenous EGF on taste bud morphology andmaintenance [In Process Citation]. Chem. Senses 25(1): 9–19.
Münch, F. (1896). Die Topographie der Papillen Zunge des Menschen undder Säugethiere. Morphol. Arb. Schwalbe 6: 605.
Murray, R. G. (1973). The ultrastructure of taste buds. In The Ultrastruc-ture of Sensory Organs, Friedmann, I. (Ed). North Holland publishingcompany, Amsterdam, London, pp. 1–81.

660 Chapter 29 Anatomy of the Tongue and Taste Buds
Murray, R. G. (1986). The mammalian taste bud type III cell: a criticalanalysis. J. Ultrastruct. Mol. Struct. Res. 95: 175–188.
Murray, R. G., and Murray, A. (1967). Fine structure of taste buds of rabbitfoliate papillae. J. Ultrastruct. Res. 19: 327–353.
Murray, R. G., and Murray, A. (1971). Relations and possible significanceof taste bud cells. Contrib. Sens. Physiol. 5: 47–95.
Murray, R. G., Murray, A., and Fujimoto, S. (1969). Fine structure of gus-tatory cells in rabbit taste buds. J.Ultrastruct.Res. 27: 444–461.
Nada, O., and Hirata, K. (1977). The monoamine-containing cell in thegustatory epithelium of some vertebrates. Arch. Histol. Jpn. 40 Suppl:197–206.
Nada, O., and Hirata, K. (1975). The occurrence of the cell type contain-ing a specific monoamine in the taste bud of the rabbit’s foliate papila.Histochemistry 43(3): 237–240.
Nagai, T., Kim, D. J., Delay, R. J., and Roper, S. D. (1996). Neuromodu-lation of transduction and signal processing in the end organs of taste.Chem. Senses 21(3): 353–365.
Nagato, T., Ren, X. Z., Toh, H., and Tandler, B. (1997). Ultrastructure ofWeber’s salivary glands of the root of the tongue in the rat. Anat. Rec.249(4): 435–440.
Nelson, G. M., and Finger, T. E. (1993). Immunolocalization of differentforms of neural cell adhesion molecule (NCAM) in rat taste buds. J.Comp. Neurol. 336:507–516.
Nemetschek-Ganssler, H., and Ferner, H. (1964). Über die Ultrastrukturder Geschmacksknospen. Z. Zellforsch. 63: 155–178.
Nishijima, K., and Atoji, Y. (2004). Taste buds and nerve fibers in the ratlarynx: an ultrastructural and immunohistochemical study. Arch. Histol.Cytol. 67(3): 195–209.
Nolte, C., and Martini, R. (1992). Immunocytochemical localization of theL1 and N-CAM cell adhesion molecules and their shared carbohydrateepitope L2/HNK-1 in the developing and differentiated gustatory papil-lae of the mouse tongue. J.Neurocytol. 21: 19–33.
Nomura, S., Shiba, Y., Muneoka, Y., and Kanno, Y. (1979). A scanning andtransmission electron microscope study of the premetamorphic papillae:possible chemoreceptive organs in the oral cavity of an anuran tadpole(Rana japonica). Arch. Histol. Jpn. 42(5): 507–516.
Norgren, R. (1990). Gustatory system. In The Human Nervous System, Pax-inos, G. (Ed). Academic Press, San Diego, pp. 845–861.
Nosrat, C. A., and Olson, L. (1995). Brain-derived neurotrophic factormRNA is expressed in the developing taste bud-bearing tongue papillaeof rat. J. Comp. Neurol. 360(4): 698–704.
Nosrat, I. V., Lindskog, S., Seiger, A., and Nosrat, C. A. (2000). LingualBDNF and NT-3 mRNA expression patterns and their relation to inner-vation in the human tongue: similarities and differences compared withrodents. J. Comp. Neurol. 417(2): 133–152.
Nosrat, I. V., Margolskee, R. F., and Nosrat, C. A. (2012). Targeted tastecell-specific overexpression of brain-derived neurotrophic factor in adulttaste buds elevates phosphorylated TrkB protein levels in taste cells,increases taste bud size, and promotes gustatory innervation. J. Biol.Chem. 287(20): 16791–16800.
Oakley, B. (1970). Reformation of taste buds by crossed sensory nerves inthe rat’s tongue. Acta. Physiol. Scand 79(1): 88–94.
Oakley, B., Brandemihl, A., Cooper, D., et al. (1998). The morpho-genesis of mouse vallate gustatory epithelium and taste buds requiresBDNF-dependent taste neurons. Dev. Brain Res. 105(1): 85–96.
Obata, H., Shimada, K., Sakai, N., and Saito, N. (1997). GABAergic neu-rotransmission in rat taste buds: immunocytochemical study for GABAand GABA transporter subtypes. Brain Res. Mol. Brain Res. 49(1–2):29–36.
Ogura, T., Margolskee, R. F., Tallini, Y. N., et al. (2007). Immuno- local-ization of vesicular acetylcholine transporter in mouse taste cells andadjacent nerve fibers: indication of acetylcholine release. Cell Tissue Res.330(1): 17–28.
Ohmura, S., Horimoto, S., and Fujita, K. (1989). Lectin cytochemistry ofthe dark granules in the type 1 cells of Syrian hamster circumvallate tastebuds. Arch. Oral Biol. 34: 161–166.
Ohshima, H., Yoshida, S., and Kobayashi, S. (1990). Blood vascular archi-tecture of the rat lingual papillae with special reference to their relationsto the connective tissue papillae and surface structures: a light and scan-ning electron microscope study. Acta. Anat. (Basel) 137(3): 213–221.
Oike, H., Matsumoto, I., and Abe, K. (2006). Group IIA phospholipaseA(2) is coexpressed with SNAP-25 in mature taste receptor cells of ratcircumvallate papillae. J. Comp. Neurol. 494(6): 876–886.
Ojima, K., Matsumoto, S., Takeda, M., et al. (1997a). Numerical variationand distributive pattern on microvascular cast specimens of vallate papil-lae in the crossbred Japanese cat tongue. Ann. Anat. 179(2): 117–126.
Ojima, K., Takahashi, T., Matsumoto, S., et al. (1997b). Angioarchitecturalstructure of the fungiform papillae on rabbit tongue anterodorsal surface.Ann. Anat. 179(4): 329–333.
Ojima, K., Takeda, M., Matsumoto, S., et al. (1997c). Functional role ofV form distribution seen in microvascular cast specimens of the filiformand fungiform papillae on the posterior central dorsal surface of the cattongue. Ann. Anat. 179(4): 321–327.
Oka, Y., and Korsching, S. I. (2011). Shared and unique G alpha proteins inthe zebrafish versus mammalian senses of taste and smell. Chem. Senses36(4): 357–365.
Olmsted, J. M. D. (1920). The results of cutting the seventh cranial nervein Amiurus nebulosus (Leseur). J.Exp.Zool. 31: 369–401.
Osculati, F., and Sbarbati, A. (1995). The frog taste disc: a prototype of thevertebrate gustatory organ. Prog Neurobiol 46, 351–399.
Ozdener, H., Yee, K. K., Cao, J., et al. (2006). Characterization andLong-Term Maintenance of Rat Taste Cells in Culture. Chem. Senses31(3): 279–290.
Panizza, B. (1834). Ricerche sperimentali sopra i nervi. Bizzoni, Pavia.
Paran, N., Mattern, C. F., and Henkin, R. I. (1975). Ultrastructure of thetaste bud of the human fungiform papilla. Cell Tissue Res. 161: 1–10.
Parker, G. H. (1922). Smell,Taste, and Allied Senses in the Vertebrates. Lip-pincott, London.
Perez, C. A., Huang, L., Rong, M., et al. (2002). A transient receptorpotential channel expressed in taste receptor cells. Nat. Neurosci. 5(11):1169–1176.
Pevzner, R. A., and Tikhonova, N. A. (1980). [Cytochemical demonstra-tion of cyclic nucleotide phosphodiesterases in the taste buds of Tes-tudo horsefieldi turtles upon exposure to flavored substances]. Zh. Evol.Biokhim Fiziol. 16(2): 133–136.
Ponzo, M. (1907). Sulla presenza di organi del gusto nella parte laringeadella faringe, nel tratto cervicale dellésofago e nel palato duro del fetoumano. Anat.Anz. 31: 570–575.
Prichard, T. C. (2011). Gustatory system. 3rd ed. In The Human NervousSystem, Mai, J. K. and Paxinos, G. (Eds). Elsevier, Amsterdam, Boston,Heidelberg, pp. 1187–1218.
Pritchard, T. C. (1991). The Primate Gustatory System. In Smell and Tastein Health and Disease, Getchell, T. V., Bartoshuk, L. M., Doty, R. L. andSnow, J. B. J., Jr., (Eds). Raven Press, New York, pp. 109–125.
Pumplin, D. W., and Getschman, E. (2000). Synaptic proteins in rattaste bud cells: appearance in the Golgi apparatus and relationship toalpha-gustducin and the Lewis(b) and A antigens. J. Comp. Neurol.427(2): 171–184.

References 661
Pumplin, D. W., Getschman, E., Boughter, J. D., Jr., et al. (1999). Differ-ential expression of carbohydrate blood-group antigens on rat taste-budcells: relation to the functional marker alpha-gustducin. J. Comp. Neurol.415(2): 230–239.
Pumplin, D. W., Yu, C., and Smith, D. V. (1997). Light and dark cells ofrat vallate taste buds are morphologically distinct cell types. J. Comp.Neurol. 378(3): 389–410.
Rabl, H. (1895). Notiz zur Morphologie der Geschmacksknospen auf derEpiglottis. Anat.Anz. 11: 153–156.
Ranvier, L. (1888). Technisches Lehrbuch der Histologie (German trans-lation by W.Nicati and H.v.Wyss). F.C.W.Vogel, Leipzig.
Remak, R. (1852). Ueber die Ganglien der Zunge bei Säugethierenund beim Menschen. In Archiv für Anatomie, Physiologie und Wis-senschaftliche Medicin, Müller, J. (Ed). Veit et Comp., Berlin, pp. 58–62.
Ren, Y., Shimada, K., Shirai, Y., et al. (1999). Immunocytochemical local-ization of serotonin and serotonin transporter (SET) in taste buds of rat.Brain Res. Mol. Brain Res. 74(1–2): 221–224.
Retzius, G. (1892). Die Nervenendigungen in dem Geschmacksorgan derSäugethiere und Amphibien. In Biologische Untersuchungen Neue FolgeIV , Samson & Wallin, Stockholm, pp. 26–32.
Retzius, G. (1893). Ueber Geschmacksknospen bei Petromyzon. In Biol-ogische Untersuchungen. Neue Folge Vol. V, Samson & Wallin, Stock-holm, pp. 69–70.
Reutter, K. (1971). The taste-buds of Amiurus nebulosus (Lesueur). Mor-phological and histochemical investigations. Z. Zellforsch. Mikrosk.Anat. 120(2): 280–308.
Reutter, K. (1973). [The types of taste buds in fishes. I. Morphological andneurohistochemical investigations on Xiphophorus helleri Heckel (Poe-ciliidae, Cyprinodontiformes, Teleostei) (author’s transl)]. Z. ZellforschMikrosk Anat. 143: 409–423.
Reutter, K. (1978). Taste organ in the bullhead (Teleostei). Adv. Anat.Embryol. Cell Biol. 55(1): 3–94.
Reutter, K. (1980). SEM- study of the mucus layer on the receptor-field offish taste buds. In Olfaction and Taste VII, van der Starre, H. (Ed). IRLPress, London, Washington DC, pp. 107.
Reutter, K. (1986). Chemoreceptors. In Biology of the Integument,Bereiter-Hahn, J., Matoltsy, A. G. and Richards, K. S. (Eds). Vol. 2,Springer, Berlin, Heidelberg, pp. 586–604.
Reutter, K. (1992). Structure of the peripheral gustatory organ, representedby the siluroid fish Plotosus lineatus (Thunberg). In Fish chemorecep-tion, Hara, T. J. (Ed). Chapman & Hall, London, pp. 60–78.
Reutter, K., and Witt, M. (1993). Morphology of vertebrate taste organsand their nerve supply. In Mechanisms of taste transduction, Simon, S.A. and Roper, S. D. (Eds). CRC Press, Boca Raton, Ann Arbor, London,Tokyo, pp. 29–82.
Reutter, K., and Witt, M. (1999). Comparative aspects of fish taste budultrastructure. In Advances in chemical signals in vertebrates, John-ston, E., Müller-Schwarze, D. and Sorensen, P. W. (Eds). KluwerAcademic/Plenum Publishers, New York, Boston, Dordrecht, London,Moscow, pp. 573–581.
Reutter, K., and Witt, M. (2004). Are there efferent synapses in fish tastebuds? J. Neurocytol. 33(6): 647–656.
Riva, A., Loffredo, F., Puxeddu, R., and Testa Riva, F. (1999). A scanningand transmission electron microscope study of the human minor salivaryglands. Arch. Oral Biol. 44 Suppl 1: S27–31.
Roper, S. D. (1989). The cell biology of vertebrate taste receptors. Annu.Rev. Neurosci. 12: 329–353.
Roper, S. D. (2006). Cell communication in taste buds. Cell Mol. Life Sci.63(13): 1494–1500.
Royer, S. M., and Kinnamon, J. C. (1988). Ultrastructure of mouse foliatetaste buds: synaptic and nonsynaptic interactions between taste cells andnerve fibers. J. Comp. Neurol. 270: 11–24, 58.
Royer, S. M., and Kinnamon, J. C. (1991). HVEM serial-section analysisof rabbit foliate taste buds: I. Type III cells and their synapses. J. Comp.Neurol. 306: 49–72.
Ruiz-Avila, L., McLaughlin, S. K., Wildman, D., et al. (1995). Coupling ofbitter receptor to phosphodiesterase through transducin in taste receptorcells. Nature 376: 80–85.
Saito, I. (1966). Comparative anatomical studies of the oral organs of thepoultry. V. Structures and distribution of taste buds of the fowl. (InJapanese). Bull Fac. Agric. Miyazahi. Univ. 13: 95–102.
Sbarbati, A., Crescimanno, C., Bernardi, P., and Osculati, F. (1999).Alpha-gustducin-immunoreactive solitary chemosensory cells in thedeveloping chemoreceptorial epithelium of the rat vallate papilla. Chem.Senses 24(5): 469–472.
Scalzi, H. A. (1967). The cytoarchitecture of gustatory receptors from therabbit foliate papillae. Z. Zellforsch Mikrosk Anat. 80(3): 413–435.
Schinkele, O. (1942). Über das Vorkommen von Geschmacksknospenim kranialen Drittel des Oesophagus. Z. Mikrosk.-anat. Forsch. 51:498–501.
Schmale, H., Ahlers, C., Blaker, M., et al. (1993). Perireceptor events intaste. Ciba Found Symp. 179: 167–80; discussion 180–185.
Schmale, H., and Bamberger, C. (1997). A novel protein with strong homol-ogy to the tumor suppressor p53 [In Process Citation]. Oncogene 15(11):1363–1367.
Schmale, H., Holtgreve-Grez, H., and Christiansen, H. (1990). Possible rolefor salivary gland protein in taste reception indicated by homology tolipophilic-ligand carrier proteins. Nature 343(6256): 366–369.
Schulze, F. E. (1863). Über die becherförmigen Organe der Fische. Z. Wiss.Zool. 12: 218–222.
Schwalbe, G. A. (1868). Über die Geschmacksorgane der Säugethiere unddes Menschen. Arch. Mikr. Anat. 4: 154–187.
Schwann, T. (1839). Mikroskopische Untersuchungen über die Überein-stimmung in der Struktur und dem Wachstum der Thiere und Pflanzen.Sander’sche Buchhandlung (Reimer), Berlin.
Schwartz, H. G., and Weddell, G. (1938). Observations on the pathwaystransmitting the sensation of taste. Brain 61: 99–115.
Segovia, C., Hutchinson, I., Laing, D. G., and Jinks, A. L. (2002). A quan-titative study of fungiform papillae and taste pore density in adults andchildren. Brain Res. Dev. Brain Res. 138(2): 135–146.
Selliseth, N. J., and Selvig, K. A. (1993). Microvasculature of the dorsumof the rat tongue: a scanning electron microscopic study using corrosioncasts. Scand J. Dent. Res. 101(6): 391–397.
Seta, Y., Oda, M., Kataoka, S., et al. (2011). Mash1 is required for the dif-ferentiation of AADC-positive type III cells in mouse taste buds. Dev.Dyn. 240(4): 775–784.
Seta, Y., Seta, C., and Barlow, L. A. (2003). Notch-associated gene expres-sion in embryonic and adult taste papillae and taste buds suggests a rolein taste cell lineage decisions. J. Comp. Neurol. 464(1): 49–61.
Shahbake, M., Hutchinson, I., Laing, D. G., and Jinks, A. L. (2005). Rapidquantitative assessment of fungiform papillae density in the humantongue. Brain Res. 1052(2): 196–201.
Shatzman, A. R., and Henkin, R. I. (1981). Gustin concentration changesrelative to salivary zinc and taste in humans. Proc. Natl. Acad Sci. U. S.A. 78(6): 3867–3871.
Shen, T., Kaya, N., Zhao, F.-L., et al. (2005). Co-expression patterns of theneuropeptides vasoactive intestinal peptide and cholecystokinin with thetransduction molecules [alpha]-gustducin and T1R2 in rat taste receptorcells. Neuroscience 130(1): 229–238.

662 Chapter 29 Anatomy of the Tongue and Taste Buds
Siegel, R. E. (1970). Galen on sense perception. His doctrines, observa-tions and experiments on vision, hearing, smell, taste, touch and pain,and their historical sources. S.Karger, Basel, New York.
Smith, D. V., Akeson, R. A., and Shipley, M. T. (1993). NCAM expressionby subsets of taste cells is dependent upon innervation. J. Comp. Neurol.336: 493–506.
Smith, D. V., Klevitsky, R., Akeson, R. A., and Shipley, M. T. (1994).Expression of the neural cell adhesion molecule (NCAM) and polysialicacid during taste bud degeneration and regeneration. J. Comp. Neurol.347: 187–196.
Smith, D. V., Som, J., Boughter, J. D., Jr,., et al. (1999). Cellular expres-sion of alpha-gustducin and the A blood group antigen in rat fungiformtaste buds cross-reinnervated by the IXth nerve. J. Comp. Neurol. 409(1):118–130.
Sobotta, J. (1993). Atlas der Anatomie des Menschen. Vol.1. Urban undSchwarzenberg, München.
Sömmering, S. T. (1806). Abbildungen der menschlichen Organe desGeschmackes und der Stimme. Varrentrapp und Wenner, Frankfurt.
Spielman, A. I. (1990). Interaction of saliva and taste. J. Dent. Res. 69(3):838–843.
Sprissler, C. (1994). Ultrastruktur der Geschmacksknospe der japanischenWachtel (Coturnix coturnix japonica). University of Tübingen.
Srur, E., Stachs, O., Guthoff, R., et al. (2010). Change of the human tastebud volume over time. Auris. Nasus. Larynx. 37(4): 449–455.
Starostik, M. R., Rebello, M. R., Cotter, K. A., et al. (2010). Expressionof GABAergic receptors in mouse taste receptor cells. PLoS One 5(10):e13639.
Stone, L. M., Finger, T. E., Tam, P. P., and Tan, S. S. (1995). Taste receptorcells arise from local epithelium, not neurogenic ectoderm. Proc. Natl.Acad. Sci. U.S.A. 92: 1916–1920.
Stone, L. S. (1940). The origin and development of taste organs in sala-manders observed in the living condition. J. Exp. Zool. 83: 481–506.
Stornelli, M. R., Lossi, L., and Giannessi, E. (2000). Localization, mor-phology and ultrastructure of taste buds in the domestic duck (Cai-rina moschata domestica L.) oral cavity. Ital. J. Anat. Embryol. 105(3):179–188.
Suzuki, Y., Takeda, M., Obara, N., and Nagai, Y. (1996). Phagocytic cellsin the taste buds of rat circumvallate papillae after denervation. Chem.Senses 21(4): 467–476.
Svejda, J., and Janota, M. (1974). Scanning electron microscopy of thepapillae foliatae of the human tongue. Oral Surg. Oral Med. Oral Pathol.37(2): 208–216.
Tandler, B., Pinkstaff, C. A., and Riva, A. (1994). Ultrastructure and histo-chemistry of human anterior lingual salivary glands (glands of Blandinand Nuhn). Anat. Rec. 240: 167–177.
Thirumangalathu, S., Harlow, D. E., Driskell, A. L., et al. (2009). Fatemapping of mammalian embryonic taste bud progenitors. Development136(9): 1519–1528.
Tizzano, M., Cristofoletti, M., Sbarbati, A., and Finger, T. E. (2011).Expression of taste receptors in solitary chemosensory cells of rodentairways. BMC Pulm. Med. 11: 3.
Tomchik, S. M., Berg, S., Kim, J. W., et al. (2007). Breadth of Tun-ing and Taste Coding in Mammalian Taste Buds. J. Neurosci. 27(40):10840–10848.
Tomonari, H., Miura, H., Nakayama, A., et al. (2012). Gα−gustducin IsExtensively Coexpressed with Sweet and Bitter Taste Receptors in boththe Soft Palate and Fungiform Papillae but Has a Different FunctionalSignificance. Chemical Senses 37(3): 241–251.
Toto, P. D., Nadimi, H., and Martinez, R. (1993). von Ebner’s gland. animmunohistochemical study. Ann. N. Y. Acad. Sci. 694: 322–324.
Toubeau, G., Cotman, C., and Bels, V. (1994). Morphological and kine-matic study of the tongue and buccal cavity in the lizard Anguis fragilis(Reptilia:Anguidae). Anat. Rec. 240: 423–433.
Toyoshima, K., Miyamoto, K., and Shimamura, A. (1987). Fine structureof taste buds in the tongue, palatal mucosa and gill arch of the axolotl,Ambystoma mexicanum. Okajimas Folia Anat. Jpn. 64(2–3): 99–109.
Toyoshima, K., Seta, Y., Toyono, T., and Takeda, S. (1999). Merkel cells areresponsible for the initiation of taste organ morphogenesis in the frog. J.Comp. Neurol. 406(1): 129–140.
Toyoshima, K., and Shimamura, A. (1987). Monoamine-containing basalcells in the taste buds of the newt Triturus pyrrhogaster. Arch. Oral Biol.32: 619–621.
Toyoshima, K., and Tandler, B. (1987). Modified smooth endoplasmicreticulum in type II cells of rabbit taste buds. J. Submicrosc. Cytol. 19:85–92.
Tuckerman, F. (1889). On the development of the taste-organs of man. J.Anat. Physiol. 23: 559–582.
Uchida, T. (1980). Ultrastructural and histochemical studies on the tastebuds in some reptiles. Arch. Histol. Jpn. 43: 459–478.
Ueda, K., Ichimori, Y., Okada, H., et al. (2006). Immunolocalization ofSNARE proteins in both type II and type III cells of rat taste buds. Arch.Histol. Cytol. 69(4): 289–296.
Vandenbeuch, A., Clapp, T. R., and Kinnamon, S. C. (2008).Amiloride-sensitive channels in type I fungiform taste cells in mouse.BMC Neurosci. 9: 1.
Varatharasan, N., Croll, R. P., and Franz-Odendaal, T. (2009). Taste buddevelopment and patterning in sighted and blind morphs of Astyanaxmexicanus. Dev. Dyn. 238(12): 3056–3064.
Verson, E. (1868). Beiträge zur Kenntnis des Kehlkopfes und der Tra-chea. Sitzungsber. Wiener Acad. Wissenschaft, Math.-naturwiss.Klasse57: 1093–1102.
von Düring, M. V., and Andres, K. H. (1976). The ultrastructure of tasteand touch receptors of the frog’s taste organ. Cell Tissue Res. 165(2):185–198.
von Vintschgau, M., and Hönigschmied, J. (1877). Nervus glossopharyn-geus und Schmeckbecher. Arch. Physiol. 14: 443–448.
Wagner, R. (1837). Bestätigung des Panizzaschen Lehrsatzes, dass das9te Nervenpaar (n.glossopharyngeus) der Geschmacksnerv ist. FroriepsNotizen 4: 129–131.
Wakisaka, S., Tabata, M. J., Maeda, T., et al. (1998). Immunohistochemicallocalization of pleiotrophin and midkine in the lingual epithelium of theadult rat. Arch. Histol. Cytol. 61(5): 475–480.
Waller, A. (1847). Microscopic examination of the papillae and nerves ofthe tongue of the frog, with observations on the mechanism of taste.London, Edinburgh, and Dublin Philosoph. Magazine J. Sci. Vol XXX:277–289.
Waller, A. (1849). Minute structure of the papillae and nerves ofthe tongue of the frog and the toad. Communicated by R. Owen.Philosoph.Transact.Royal Soc.London Pt.I: 139–149.
Welsch, U., and Storch, V. (1969). [Fine structure of the taste buds of catfish(Clarias batrachus (L) and Kryptopterus bicirrhis (Cuvier and Valenci-ennes)]. Z. Zellforsch Mikrosk. Anat. 100(4): 552–559.
Whitear, M. (1970). The skin surface of bony fishes. J.Zool.London 160:437–454.
Whitear, M. (1992). Solitary chemosensory cells. In Fish chemoreception,Hara, T. (Ed). Chapman & Hall, London, pp. 103–125.
Whitehead, M. C., Ganchrow, J. R., Ganchrow, D., and Yao, B. (1998).Neural cell adhesion molecule, neuron-specific enolase and calcitoningene-related peptide immunoreactivity in hamster taste buds after chordatympani lingual nerve denervation. Neuroscience 83(3): 843–856.

References 663
Williams, P. L., Warwick, R., Dyson, M., and Bannister, L. H. (1989).Gray’s Anatomy. Curchill Livingstone, Edinburgh London.
Wilson, J. G. (1905). The structure and function of the taste-buds of thelarynx. Brain 28: 339–351.
Witt, M. (1993). Ultrastructure of the taste disc in the red-bellied toadBombina orientalis (Discoglossidae, Salientia). Cell Tissue Res. 272:59–70.
Witt, M. (1995). Distribution of vasoactive intestinal peptide-likeimmunoreactivity in the taste organs of teleost fish and frog. Histochem.J. 27: 161–165.
Witt, M. (1996). Carbohydrate histochemistry of vertebrate taste organs.Progr. Histochem. Cytochem. 30/4: 1–172.
Witt, M., and Kasper, M. (1998). Immunohistochemical distribution ofCD44 and some of its isoforms during human taste bud development.Histochem. Cell Biol. 110: 95–113.
Witt, M., and Kasper, M. (1999). Distribution of cytokeratin filaments andvimentin in developing human taste buds. Anat. Embryol. 199: 291–299.
Witt, M., and Miller, I. J., Jr., (1992). Comparative lectin histochemistry ontaste buds in foliate, circumvallate and fungiform papillae of the rabbittongue. Histochemistry 98(3): 173–182.
Witt, M., and Reutter, K. (1988). Lectin histochemistry on mucous sub-stances of the taste buds and adjacent epithelia of different vertebrates.Histochemistry 88: 453–461.
Witt, M., and Reutter, K. (1990). Electron microscopic demonstration oflectin binding sites in the taste buds of the European catfish Silurus glanis(Teleostei). Histochemistry 94: 617–628.
Witt, M., and Reutter, K. (1996). Embryonic and early fetal developmentof human taste buds: a transmission electron microscopical study. Anat.Rec. 246(4): 507–523.
Witt, M., and Reutter, K. (1997). Scanning electron microscopical studiesof developing gustatory papillae in humans. Chem. Senses 22: 601–612.
Wong, G. T., Ruiz-Avila, L., Ming, D., et al. (1996). Biochemical and trans-genic analysis of gustducin’s role in bitter and sweet transduction. ColdSpring Harb. Symp. Quant. Biol. 61: 173–184.
Wyss, H. v. (1870). Die becherförmigen Organe der Zunge. Arch. mikrosk.Anat. 6: 237–260.
Yamasaki, F., and Takahashi, K. (1982). A description of the times ofappearance and regression of marginal lingual papillae in human fetusesand newborns. Anat.Rec. 204: 171–173.
Yang, R., Crowley, H. H., Rock, M. E., and Kinnamon, J. C. (2000). Tastecells with synapses in rat circumvallate papillae display SNAP-25-likeimmunoreactivity. J. Comp. Neurol. 424(2): 205–215.
Yee, C., Bartel, D. L., and Finger, T. E. (2005). Effects of glossopharyngealnerve section on the expression of neurotrophins and their receptors inlingual taste buds of adult mice. J. Comp. Neurol. 490(4): 371–390.
Yee, C. L., Yang, R., Böttger, B., et al. (2001). “Type III” cells ofrat taste buds: immunohistochemical and ultrastructural studies ofneuron-specific enolase, protein gene product 9.5, and serotonin. J.Comp. Neurol. 440(1): 97–108.
Yoshie, S., Wakasugi, C., Teraki, Y., et al. (1988). Immunocytochemicallocalizations of neuron-specific proteins in the taste bud of the guineapig. Arch. Histol. Cytol. 51: 379–384.
Zachar, P. C., and Jonz, M. G. (2011). Confocal imaging of Merkel–likebasal cells in the taste buds of zebrafish. Acta Histochem. 114(2):101–115.
Zancanaro, C., Sbarbati, A., Bolner, A., et al. (1995). Biogenic amines inthe taste organ. Chem.Senses 20: 329–335.
Zhang, C., and Oakley, B. (1996). The distribution and origin of ker-atin 20-containing taste buds in rat and human. Differentiation 61(2):121–127.
Zhang, C. X., Cotter, M., Lawton, A., et al. (1995). Keratin 18 is associatedwith a subset of older taste cells in the rat. Differentiation 59: 155–162.
Zhao, F. L., Shen, T., Kaya, N., et al. (2005). Expression, physiologicalaction, and coexpression patterns of neuropeptide Y in rat taste-bud cells.Proc. Natl. Acad. Sci. U.S.A. 102(31): 11100–11105.
Zimmermann, K. W. (1927). Die Speicheldrüsen der Mundhöhle und dieBauchspeicheldrüse. In Handbuch der Mikroskopischen Anatomie desMenschen, Möllendorf, W. v. (Ed). Vol. V, Part 1, Springer, Berlin, pp.61–244.
Zuwała, K. (1997). Ultrastructure of premetamorphic taste organs of theBombina variegata. Rocz. Akad. Med. Bialymst. 42 Suppl 2: 204–207.
Zuwała, K., and Jakubowski, M. (1991). Development of taste organsin Rana temporaria. Transmission and scanning electron microscopicstudy. Anat. Embryol. (Berl) 184(4): 363–369.