analysis of the brassica oleracea by b. campestris-oleracea...
TRANSCRIPT

Theor Appl Genet (1987) 74:758-766
D Springer-Verlag 1987
Analysis of the Brassica oleracea genome by the generation of B. campestris-oleracea chromosome addition lines: characterization by isozymes and rDNA genes
C. F.Quiros, O.Ochoa, S. F. Kianian and D. Douches
Department of Vegetable Crops, Umversily ofCalifomia. DavIs. CA 95616, USA
Received April 27, 1987: Accepted June 16, 1987 Communicated by C. S. Khush
Summary. This study aimed at generating chromosome <.tddltion hnes and disclosing genome specific markers in 8m.15I('(1. These stocks will be used 10 study genome evolutJvn in Bmss/('Ci ()ler(lCCo L. B, ('ampel/r!s Land the derived am phidi plOJd ,,,,,eCles B. napus L B. cam
pes/f1S-o!f/'aceo mono,om ic and dlsomic cli "omosome addition plants w.:re generated by crossing <lnd backcrosslng the natural amphidiploid B. napLl!> 10 lhe Jiplold parental species B, campeSlns. The pollen VI'
abtllty of the derived sesq uidiploid and hyperploid ranged from 63S, to l:I8~; . whIle the monosomic ;lnd disomic addition plants had an average pollen fCrlility of 94% and 9) %, respectively. The addition lines were genetically chari;lcteri/\'d by genome specific markers. The isozymes for 6PGD, LAP, PGI and PGM, and rONA Eel! RJ reslnClton fragments were found 10 posseSS lhe desired genome specificity. Duplicated loci for ,,~veral of these markers were observed in B. campeslris and R. o!UGCl'iI. <;upporling the hypothesis that these diplOid ,penes are actually secondary polyploids. A tolal of elghl monosom:c and eight disomlc addition rlants were identi fil'd and characterized on the basis of these markers. Another 51 plants remained uncharacterized due to the lack of addItional markers. rDNA genes were found to be distributed in more than one chr"mosome, diffl'ling in its restriction sites. In· tergenomlc recombination for some of the markers was detected at II eq uencies between 6~c. and 20',", revealing lhe feasibility of llllergenomic gene transfer.
Key words; Bml.>IC(/ - Cok crops - Cytogenetics - Genome - M 1rk..:rs
Jntrnduction
llle genus Bra.wca has been the subject of numer<)uS cytogenetJc 'itudies (Prakash and Hinata 1980). The large num ber of diploid species with t'L'oomic J) urn bers rangi ng from x. = 7 to x = 12. and derived am phidipJoids in lhe genus. mak.es it an attractive rese,Hch subject (Mlzushima 1980). Most of the cytogenic research in Bmss/c(I J1HS cen tered around the eI uCldation of the origm \)1' the culllvatcJ am phldi plaids. B. "opus L. B. carinll/a A. Be. and B. prnceo L. by Karpcchenko (1922), Morin"ba (1934). and U (1935). This early work was based on th..: synthesis of amphidiplolds by hybridi· zation or the three basic dIploid cuhiva led species B. nigra L. (x = 8, genome b). B. oferocea L. (X" 9. genome c)
and B. campevlris L. (x = 10. genome a). A result of this research was lhe poslUlaled tflangle of C, which IS a diagrammatic representation of dIploid and amphidip 10ld species relationshIp (U 1935). More recently research by Robbins and Vaugha n (1983) on Rubisco and by Palmer et al. (1983) <l nd Erickson et al. (1983) on chloroplast DNA not only confirmed the validity of the triangle of l' bu t also resolved lhe direclion of the crosses leading 10 two orlhe amphidiploids.
Very ll\lIe IS known about the evolution of the diplQld species and the origin of the different genomIc numbef$. On lhe basis of chromosome painng in haploids (Keller and Arm,trong 1983). di-gcllomic and {ri·genomic hybrids (Mlzushlma 1980; Prakash and Hinala 1980; Catcheside 1934. 1937; Sikh 1940). and l';'chy lene ka ryotypes (Robbclen 1960). II IS assumed lhal lhe baSIC genome of Brass/co is x = 6. Since no ,pecies Wllh 11115 genomic number has ever been reponed. they are presumed [0 be exllnct. Robbelen (1960) found SIX basIC lypes or chromosomes. designating them with the lellers A to F, on [he baSIS of helerochromallC knobs and centromere position. He proposed Iha1 lhe composilion or each genome in the culhvated dIploids was genome
a =AABCDDEFrr. b =AECDDEU and c'" ABBCCDEEF,

759
Two chromosomcs in each genome. namely A and C m B. nI
gra and B. olerclcea, and A in B. caml'eslris. were found 10 be involved in the organizalion of the nucleolus. Thus, the diplOId species are considered to be secondary polyploids, ,mce presumably they have some of the baSK: chromosome lypes forming part of their genome in duplicale or even m Irlplicate. The fact that chromosomes or the same lype wlthm or belween species are not identical. indicales lhal rearrangemcnl, have taken place during the evolution of the diplOid specIes.
In order to test these hypotheses based moslly on cytolo,2ical observations. cytogenetic slocks usd'ul to dissect each genome and genome specific chromoso'tle markers are reqoired. This approach has been very userul in Trilitum and relaled genera (Hart and Tuleen [983), and Allium (Pelney el aL 19l>5), Genome specific isozyme lOCI In the Cultivated Brass/en specIes have already been described (Cou;t!lan and Denford 1982: Aru sand Onon 1983. Arus 1984: Qui ros et a I, 1985).
In th is "udy. we rerNt the use Q( isozyme markers Jnu rDNA genes III 1:1;,; charactuiza lion of B, camfle(lris-o/eracea l11onosom IL' and disomic addition lines obla ined by crossl1lg and back crossing B, napll.\ 10 B. wmpeslris, TIlese lin es wdl permit studies of genome or,':aIlJzation and cvolu ti\Jo in Brassica.
1V13teri31s and methods
Plonl/?la/eria!
The lollowing acceSSIons were used: B napu:,. rapid cycling CrGC-05 and rapid cycling cytoplasmic male Slenle CrGC-14: R call1peslri.l, rapid cycling CrGe-0 I and rapId cychng cytoplasmic mal.. staile CrGC-I3: rapeseed cv "Torch': Chinese cabb~,~e cV 'K wan-Hoo Cho,' The rap,d cycling lines were obtainnl lJ'om Dr. Paul William, al the l.'niversity of Wisconsin. USA (Williams and Hill 1986) Also. the lurnip cv 'White Lady' was used to ,( ud I' the Hlhenta nce of the triose phospha Ie
isom erase JSozy me,.
De' "iopmenl o/addfllon {me"
111e amphldlplold species B nupus was crossed eilher as pistillate or poJleo parent 10 the diploid B, campeSlrls. Embryos were n:scued from developing 0' "les about 15 days afler pollJnJIIOn and cultured io sterile medium (Nitsch and Nitsch 1969). The resultmg sesqutdlpJold hybrids were backcrossed (0
B. (ampeSlfl5. Resulting progeny with more lhan 2n = 21 chromosomes were b~ckcrossed one or (wo more times to B. compeSl flS. Mooosom ic addi tion lines. 20 = 2 J. were selfed to obla I n d ison11 c udJ it,on Ii nes (2 n = 22) Amon 8- all the crosses a loul 0f ,',1)out 350 planls were generaled, The majorilY or these were S! udied cy lologlcally and eIeClrophoretically.
Chro(tlo\'ome ('(lunls
Flower buds were tixed [n propionic aeid: absolute ethanol (J : 3) 'iI,tll ferric chlond" added as a mordant (Swam lila than ct 3.1 1954) After 24 h the huds were rinsed and SlOred 111 70% tthanol Anlh<:,~ were d;~,ecled and squa~hed in a drop of I',. a<:etoeannme for chromosome COllnl~. 10 to 30 cells were exammed. PoJl;'n vlat>'''ly was determined on the basis of pollen stamabllity In l't acetoCo'T111lne, A ml1l1mum of 100 pollen grains was used for (his determination,
ISI):! me mark"r)
Horizon l(1I starch ge I el ecl [<J ph oreSI,' was u~ed to ~epara te the enzymes obtained from a crude extracl of y()ung leave,.., and
poJJen leachales (Weeden and GOlli,cb 1980), Details of 11m techmque have bt'c'n previously descnbed (Qujro~ and McHale 1985). TIle follOWing enzymes were assayed. 6phosphoglueonase deh yd rogen ase (6 PG D). ph 0sphogl uCOl$.Omerase (PG'). leuc ine UIn Inopeptidase (LA P). an d phosphoglucom ula se (PGM). Thc II1herllanee for lhe enzyme SVSlems LAP, PGI and PGM hJve been reported In B. o!erGl:e; by Arus and Orlon (1983) Although we did nol carry oul a formal genetic analysis In R C(llJlpCS I fI.\. the SlmllanlY between lhe zymo:;rams of both speCies mdlcak that they haw eq uivalenl loci c'oumg for lhese cnzyme~.
rDNA ;;en('s
Total genomic DNA was lsol.ued from !eaves of indiVidual plaots according to (he protocol of Saghal-Maroof et al. (1984) wjlh the following modlficallons: the leaf lissue was h'lIr,)gcnized With dry Ice in a coffee mIll (Mollhnex/Regal) (l<Jn<.lry and Michelmore 1985) After extraction. the DNA was dlgesled for 8 h with the endonuclease ErG Rl according 10 lhe manuraerUl," tBRl). The DNA fragments were separaled by h<Jrizoo tal ag" rose electro phoresls and transrered by Sou the rn blolling to Zeta-Probe membranes (Maniatis el al. 1982) Cloned DNA from wheat rONA, probes pTA71 (Saghal- Maroof et al. 1984) and pTA250-2 (Appels and Dvorak 1982) was nick tranSluted u.\tng standard lech mques (Mamatls el al. 1982) and hybfldiLcd 10 til, membranes. D!'-.\ fragments were Sized u~ing l:lmbd3 DNA as a refe.ence Radish (Raphanus SG/Il'US L.)
DNA was I~,ed as a control because lhe rONA genes of lhls close relalJve of Brassica are well characlem:ed (Oelseny et al. 1983),
Results
SeSlluidlploid hvtmd~'
111 f cross B. l1apus X B, campeslris and its reciprocal re,;ulted in seeds devoid of endosperm, A tOIJI of seven hyhrids were obtai ned after rescui ng and culturing embryos con (a Ined in these seeds. Four of the hybridS were male-sterile due to the use of Ilk cy(oplasmlc male-<;tcrile ~locks as pistillate parents. The absence of fertility re<;torer~ in the pollen parents made impractical the use of (hese hyhrids in the development of the addition lines. Therefore, our efforts were concentrated In the partially !CrtiJc hybrid. 85 B138, wi th 65 % pollen viability (Table I). A second partially fertile hybrid, 85 B137, derived
Table 1. Chromosome numbers and pollen Viability or Brassil'O napus x B. campesrris hybrid.,
Plant no. 20 % Pollen viability
85869-1 30 15 85666-2 29 ems' 85676-1 62 ems 85B146-1 29 ems 85B137-1 29 67 85B 138-1 29 65 85665-1 ' 29 ems
Reci rroca Icross b Cytopiasmic male Slerile
760
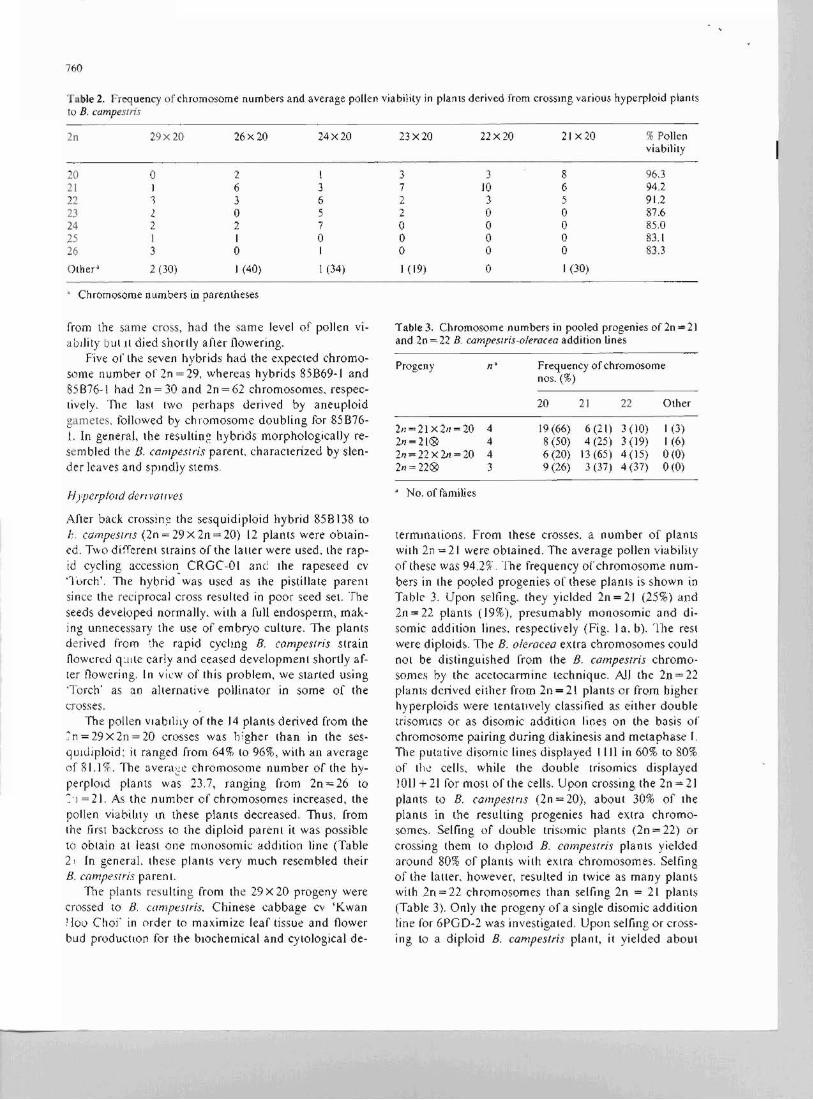
Table 2. Frequency of chromosome numbers and average pollen viability in plants derived from crossmg various hyperpJoid plants to B. campesrris
2n 29x20 26x20 24x20 23x20 22x20 21 x20 % Pollen viability
20 0 2 I 3 3 8 96.3 21 J 6 3 7 JO 6 94.2 22 1 3 6 2 3 5 91,2 23 2 0 5 2 0 0 87.6 24 2 2 7 0 0 0 85,0 25 I I 0 0 0 0 83.1 26 3 0 I 0 0 0 83.3
Other' 2 (30) J (40) 1 (34) 1(19) 0 I (30)
• Chromosome nllmbers in parentheses
from the same cro,s, had the same level of pollen vi Table 3. Chromosom e numbers in pooled progenies of 2n = 21 a bili ly but Jl died shorlly afler Dowering. and 2n = 22 B. campes/ris-o!eracea addition Lines
Five of the seven hybrids had the expected chromosome number of2n=29, whereas hybrids 85869-1 and 85 B76-1 had 2n = 30 and 2n = 62 chromosomes, respec
Progeny n' Frequency of chromosome nos. (%)
tively. 'llle last two perhaps derived by aneuploid 20 21 22 Olher gametes. iollowed by chromosome doubling for 85B76l. In general, the resultin~ hybrids morphologically resembled the B. campes/ris parenL characterized by slen
2,,=21X2n=20 4 2n=210 4 2n=22x2n=20 4
19 (66) 6 (21) 3 (10) I (3) 8 (50) 4 (25) 3 (19) 1(6) 6 (20) 13 (65) 4 (15) 0(0)
der leaves and spJOdly stems. 2n = 220 3 9 (26) 3 (37) 4 (37) 0(0)
HyperplOid def! VOI/VCS • No. of families
After back crossin" the sesq uidiploid hybrid 85 B138 to L campes/TIS (2n = 29 x 2n = 20) 12 plants were obtain terminations, From these crosses. a number of plants ed. Two different strains of the la Iter were used. the rap wi th 2n = 21 were obta ined. The average pollen viability id cycling accession. CRGe-OI ane: the rapeseed cv of these was 94,2 W The frequency of chromosome num.OJ vrch', llle hybrid was used as Ihe pistillate parent bers in the popled progenies of these plants is shown in since the reciprocal cross resulted in poor seed set. The Table 3. Upon selfing. they yielded 2n=21 (25%) and seeds developed normally. with a full endosperm, mak 2n = 22 plants (19%), presumably monosomic and diing unnecessary the use of embryo culture. The plants somic addition Jines. respectively (Fig. I a. b). The rest derived from '.he rapid cychng B. campeslris strain were diploids. The B. a/eroceo extra chromosomes could nowcred q'.llte carly and ceased development shortly af not be distinguished from the B. compeslris chromoler Dowering, In viL'W of this problem, we started using somes by the acetocarmine technique. All the 2n = 22 Torch' as an alternative pollinator in some of the plants derived either from 2n = 21 plants or from higher crosses. hyperploids were tentallvely dassi fied as either double
The pollen vlabilllY of the 14 plants derived from the trisomtcs or as disomic addition lines on the basis of ~ n = 29 x 2n = 20 crosses was 11 :gher than in the ses chromosome pairing during diakinesis and metaphase I, qUllllploid; it ranged from 64% to 96%, with an average The puta tive disomic lines displayed II Jl in 60% to 80% of 81.1%. The avera 2c chromosome number of the hy of thc cells, while the double trisomies displayed perplOld plants was 23,7, ranging from 2n=26 to 1011 + 21 for most of the cells. Upan crossing the 2n = 2I ~ ) = 21, As the number of ch romosomes increased, the plants to B. compes/TIS (2n = 20), about 30% of the pollen viability III these plants decreased. Thus. from plants in the resulting progenies had extra chromothe first backcross to the diploid parent it was possible somes. Selfing of double trisomic plants (2n = 22) or to obtain at least one monosomic addition line (Table crossing them to dJplOJd B. compeslris plants yielded 2, In generaL these plan ts very much resem bled their around 80% of plants with extra chromosomes. Selfing B. campeslris paren l. of the lalter. however, resulted in twice as many pJant~
The plants resulting from the 29 x 20 progeny were with 2n = 22 ch romosomes th,m selfing 2n = 21 plants crossed to B. Cilmpeslris. Chinese cabbage cv 'Kwan (Table 3). Only the progeny of a single disomic addition , 100 Choi' in order to maximize leaf tissue and /lower line lor 6PG 0-2 was investigated. Upon seJfing or crossbud productIOn for the biochemical and cytological de ing to a diploid B. compes/ris plant, it yielded aboul

761
50% and 30% of plants with extra chromosomes, respectively. About 50% of these har' ?n = 22 chromosomes.
Genome specific markers
The four enzyme systems listed in the "Materials and methods" section were satisfactory [or identifying the extra B. oferacea chromosomes present in the hyperploid plants. Other system~ were tried. but the overlapping or the complexity of the zymogTams precluded their use as reliable markers. The best diagnostic enzyme system was 6PGD. since it was found to be monomorphic for a number of accessions of B. o/eracea, B. CGlJlpeslris and B. napus (Fig. 2a). This monomorphism has meant that formal genetic tests have not been possible for the determination of the number of loci involved in the syntheSIS of these isozymes. The lY010grams of the diploids Showed clearly two activity rones, the more anodal one, named 6PGD-1. is composed of three equidistant bands. of which the most anodal band is the only one shared by both species (Fig. 1a, b). For the more cathodal zone, named 6PGD-2. most B. oferar:'a accessions display a three banded phenotype. whd;; B compeslris has only one band overlapping with the B. o/eracea band of slowest migration. Only the 6PGI)-1 isozymes persIsted in pollen leachates in both diplOids, indicating cytosolic location. Furthermore the lhro-'l: banded pa lIern observed In the leaves of B. o/eroceo was also retaliled in pollen leachates revealing durhcated loci for 6PGD-2 (Weeden and Gottlieb 1980). Conversely. the 6PGD-ll.;ozymes did not persist In pollen leachates indicating plastid location. The Isozymes from both diploid species were accounted for In B. napus. confirming the hybrid nature of this amphidiploid species. The multiple banded phenotypes for 6PGD-l and 6PG 0-2 bred true in selfed or sib progen ies for each of the three species confirming the existence of duphcated loci.
Duplicated loci We)~ also observed for the genes coding for tbe enzyme triose phosphate isomerase (Tpi2 and Tpi-2') In the two diploid cultivated species. B. oferacea and B. C(I 111pCSIris. non segregating multiple banded phenotypes were observed in leaves and in polkn leachates..hus indica ting cytosohc location. On the other h[lIiJ. the isozymes of the more anodal zone, TP1I were Jo(;a ted in p!;t,tlds. followlOg the criteria of Weeden and Gotlheb (1980). B. campeslris plants of the \;lrnip cv 'White Lady' heterozygous for ooe of the duplicat~d loci permitted the observalion of intergcnic heterodimers forming between the monomorphic locus and the two alleles of it~ polymorphic duplicate locus (Fig. 2c). A sib cross between two TPI-2"/TPI-2'2 heterozygotes segregated in the expected I: 2: 1 ratio, 10(2"): 20(2"12'2): 9(2'2). X2 = 0.03, P=0.99. For PGM and LAP. isozymes migrating closer to the calhode and
common in B. oleracea were diagnostic for extra B. o/eracca chromosomes (Fig. 3c).
rDNA genes were fOund (0 be extremely useful for the characterization of the hyperplOId hoes. Each diploid species has a lypical restriction pa llern with Eco RI, although a few fragments in common were a:so observed. The restriction pa ltern of B. napus had all the fragmlnts found in both diploid species (Fig. 3a). lhus. Ihe 6PGD and the rONA systems were very reliable for confirming the hybrid origin of B. napus. The res\r:ctlon pattern of the radish used as control W;LS a combinalion of those of B. oferacea and B. campeslris suggesting a close relatIOnship between the two genera. In B. o/eracea. the following fragments were observed: 6.0. 3.4. 1.8. 1.6, 1.4 and 0.8 Kb. while B. campeSlfIS displayed 5.3, 4.1. 2.6. 1.6 and 1.4 Kb fragments. Radish had a pallern similar to B. o/erucea, except for a 6.0 Kb fragment whtch IS replaced by the two 5.3 and 4.1 Kb fragments of B. w111peslris. All these flagments lighted
Fig. 1a, b. Pollen mother cells. a Mew.phase 11 for monosomic addition line (2n = 21); 10 chromosomes al the leli and II al the righl pole. b Metaphase I for a monosomic addition line (2n '" 21). Showing a trivalent association

Fig.2. a 6PGD ~ymograms for Brass/ca oleracea. B. comp"Slr;:, and B. napus B. oleracea and B campO'lm specific isozymes for 6PGD-1 and 6PGD-2 arc presenl lD lhe hybrid specIes B. napur. b InlerpretatIVe diagram for 6PGD-1. 6pgd-1 and 6pgd-1' are duplicated JOCL
SPGO 1 Jsozymes J01 (B. oleracea) and I<:p (B. camper/ris) at 6pgd-J have lhe SJ me migr31ions. Isozymes 201 and 2cp at 6pgd-r form helerodimers with lhe 6pgd.1 isozymes. T!lC
f. f'G02 amphidiploid B. Ilapus (second line from righl) and B. compeSlriS
101 oleracea addition lines (jar righl) diS
3pgd 1 1cp play an additIOnal hcterodimer
formed by the 2cp and 201 polypeptides. The inlenslty of 20 I de
2cp pends on lhe number of eXira Il Pgd}' 6PGD. B. oleracea chromosomes. c
Duplicated loci for the enzyme 11'[, Tpi-2 monomorphic for lhe Isozyme I and Tp/'}' polymorphic for the al
2cp/201 IOlymes 1 and 2. Firsr 16 lillt's from lef! show segregating progeny (1:2: I) for Tpi-1 allozymes I and 2. Heterozygous individuals for Tpr.2' (for example. line 2} form two helerodimers, one belween the Tpi-2 isozyme and the Tpi-2' allozyme I and another between the aJ)ozymes I and 2 ofTpi-2'. Homozygous-indi. vlduals for Tpi.2' for a single helerodimer wilh Isozyme I of TpI-]. (I.e. line J, TPI·2". line 7, TPI-2' L). Lines 17 to the end show the progeny of a double homozygous plant or phenolype TPl-]! and TPI-?'! breeding true. Ine band at the middle is lhe mterlOCIlS helerodlmer
Table 4. Distribution of B. oleracea-specilic alleles in B. campes/ris-o!eracea hyperploid plants and transmission 10 their progenies
Plant no. 2n PGI-I 6PGD-l 6PGD-2 LAP-I PGM-2 RA RB
85B224-1 86B25-3 86B25-8 86B29-2 86829-6
85B224-3 86B44-1
85B268-1 116B55-3 86B56-2
851:l268-2 86850- i
86B170-3 86B170-6
86B50-2 86850-3 86850-7
86B 183-6 858268-4
86BI55·' 85B268-7
86B I09-2
24 21
22
22 21 24 22 22 22 21 21 22 22 21 22 22 26 21 26 22
~:
" ~ ~ 'e
"' ~
* *
* '" ~: '" ~ "
" * " ~
* "
763
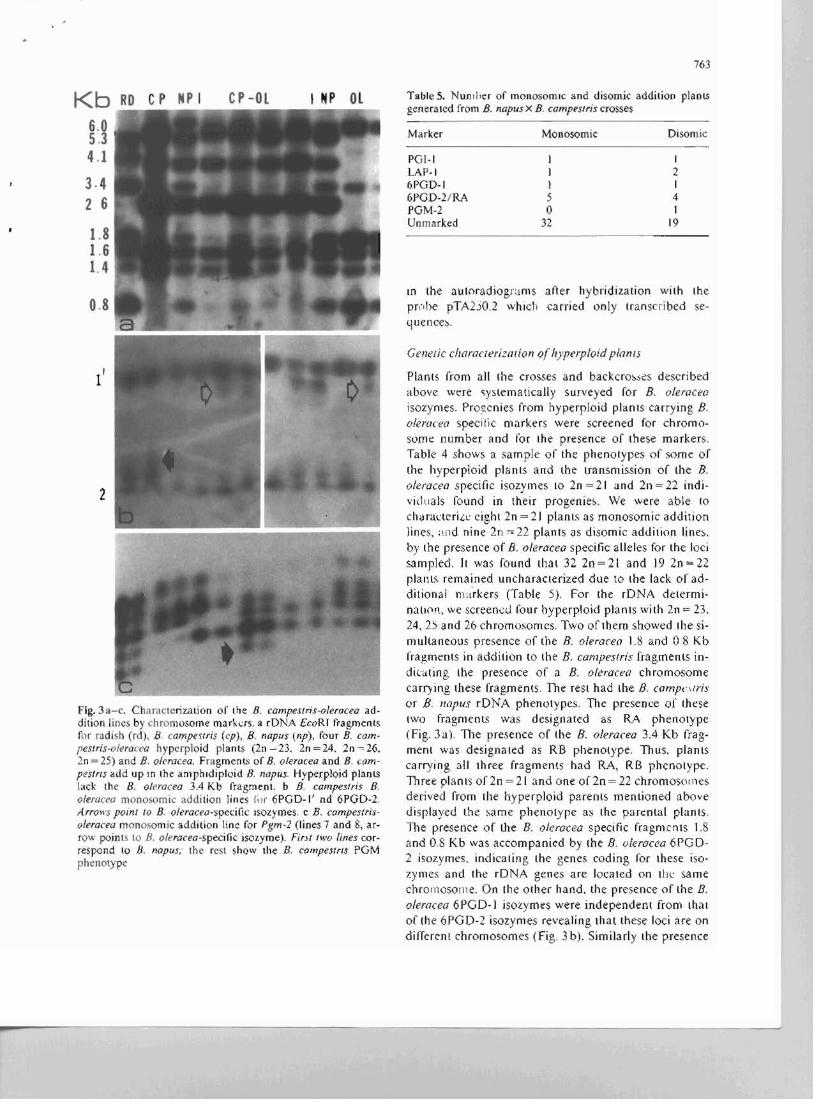
Kb RD CP NP I CP-OL I NP OL
6.0 5.3 4.1
3·4 2 6
1.8 1.6 1.4
0.8
l'
2
Fig.3a-c. Characterization of the B. campes/ris-oleracea addition lines by chromosome markas. a rDNA EcoRI fragments for radish (rd). S, campemis (cp), S, napus (np), four B. campesrris-o/eracea hyperploid plants (2n ~ 23. 2n = 24. 2n -~ 26, 2n = 25) and B, o/cracea, Fragmenl~ of D. oleracea and B, campes/r/S add up In the amphidiploid S, napus. Hyperploid plants lack the B. o/eracea 3.4 Kb fragment. b B, campes!ris B, oleracea mono omic addition lines Il,,. 6PGD-I' od 6PGD-2, Arrows poinl 10 S, o/eraceo-specific lsozymes, c B. wmpestrisolera£'eG monosomic addition line for Pgm-2 (lines 7 and 8, arrow points to D, a/eracea-specific isozyme), First two lines correspond to D. nopus; the rest show the D. C(Jmpestrls PGM phenotype
Table 5. Nun,r,er of monosomic and disomic addition plaoLS generated from S, napus X E, campes!ris crosses
Marker Monosomic Disomic
POI-I I I LAP-I I 2 6POD-1 1 I 6FGD-2/RA 5 4 PGM-2 0 1 Unmarked 32 19
10 the autoradiograms after hybridization with the pr",be pTAljO,2 which carried only transcribed sequence~.
Gene/it c!wrac/erizmion ofhyperploid plants
Plants from all (he crosses and backcro~.ses described above were ~ystematically surveyed for B. oleracea isozymes, Pro,genies from hyperploid plants carrying B. o!eroceo speciilc markers were screened for chromosome number and for lhe presence of these markers. Table 4 shows a sample of the phenotypes of some of the hyperploid plants and the transmission of the B. oleracea specific isozymes to 2n = 21 and 2n = 22 indio viduals found in lheir progenie~. We were able to clwradcriLL: eight 2n = 2J plants as monosomic addition lines, :\ild nine 2n "" 22 plants as disom ic addition lines, by the presence of B. o/erocco specific alleles for the loci sampled, II was found that 32 20 = 21 and 19 2n = 22 plants remained uncharaclerized due to the lack of additional m:t'rkers (Table S). For the rONA determi· na()0n, we screencJ four hyperploid plants with 2n "". 23, 24,25 and 26 chromosomes, Two oflhem showed the simultaneous presence of the B. o/Naceo 1.8 and 0 8 Kb fraj!.ments in addition to the B. compeslris fragments indicating (he preseoce of a B. oleracea chromosome car1)ing Ibese fragments, The rest had the B, campnlris or B, lIapus rONA phenotypes. The presence of Ihese two fragments was designated as RA phenotype (Fig. 3a), 111e presence oC (he B. oleracea 3A Kb fragment was designated as RB phenotype, Thus, plants carrying all three fragments had RA, RB phcnolype. Three plants of20 = 21 and one oC2n= 22 chromosomes derived Crom the hyperploid parents mentioned above displayed the same phenotype as the parental plan IS, The presence of lhe B. oleracea specific fragm cnts 1.8 and 0,8 Kb was accompanied by the S. oleracea 6PGD2 isozymes, indicaling the genes coding lor these isozymes and the rONA genes are located on tltt: same chromosome. On the other hand. the presence of lhe B, olera<:ea 6PGD-l isozymes were independent from thai oC the 6PGD-2 isozymes revealing that these loci are on diJTerent chromosomes (Fig, 3 b). Similarly the presence

764
Fig. 4. Progeny from hyperplQid B. campeslris-o(eracea plant carrying a PGI -2 8, oleracea-specific chromosome. Addition lmes are heterozygous (3 banded phenotype) and diploid B, ('ampeslris are homozygous ror the tsozymes of raster migra': Ill. The plant homozygous for lhe slower isozymc may have originated by intergenomlC reoombinallon
Table 6. Frequency of intcrgenomlC recom billanLS observed in the progenies of hyperplolds
Progeny n· Recombination Loci frequency (?OJ
85B 138-1 5 20.0 Pgl-l 86B ISO-I 10 10.0 Pgi-/ 86B150-7 12 8.3 6pgd-2 85B268-4 16 6.2 6pgd-f
. Progeny SJze
of LAP-I. PGI-I and PGM-2 B. oleracea specific ISOzymes were found to be Independent of each other.
Intergenomlc recombinants were observed in some of (he progenies from hyperploid plants. A few diploid plants showmg both B. o!eracea and B. campeslris isolvmes (Table 6) were delected. In addItIon, two plant., were homozygous for a B. oleracea POI Isozyme, lackmg the B. wmpt:5lns isozymes (Flg.4). The sporadic presence of multivalenls in some oC the addition lines explains the origin of these r.:com binan LS (Fig, I b).
In progenIes from hyperploId plants we detected eight self·compatible planls of either 2n =21 or 22 chromosomes derived from crosses not involving the Chinese cabbage ·Kwan-Hoo Chai'. a cultivar with a very relaxed self incompalJbility. [n addition. lWO diploid plants derived from the same hyperploid parents were also found to h,: self-compatible.
Discussion
Th e hIgh pollen viability in the sesq uidiploid hybrids and In the sub equent hyperplOid derivatives permits generation of a series of alien chromosome addItIon lines in Bmssiw, A good example of the high tolerance for exlra chromosomes In B. campesrris is shown by 2n = 26 plants (lJ,playing a p"lIen viability of 8]« TIle
tolerance for extra chromosomes might have evolved in this species as a step to JJ(oploldization in the generation of the hybrid polyploid B, napus. A similar situation occurs in wheat where a whole series of aneuploid stocks has been constructed (Sears 1969). The mO'lO
somic and disomic addition lines generaled in our study had a pollen viability of at least 90%. Furthermore, the transmission of the extra chromosome through Ihe ovules of :' I] = 21 plants was on the average 21 %. lile presence of disomic addition lines at a frequency of 10% indicates that the extra chromosome undergoes non-disjunction in Ihe female gametes, Selfing or monosomic addition plants increased the transmission of the exll':) chromosome resulling in 25% tri~omics and 19% tetrasomic plants In the progeny. This indicates that the extra chromosome may be transmitted through pollen.
The 6PG1) loci and lhe rDNA phenotypes confirm lJ'e Oflglll of B. 110pUS as a hybnd of B, ('ampes/ris and B. oleroao, Allhaugh the number of loci duplicated for 6PGD could not be pinpointed by genl'lic analysis du<: to the lack llf polymorphism. they can be eXlrapolalCd by Jnspection of the zymograms. Figure 2 b shows the interpretation for 6PG D I based 00 2 loci designated 6pgd-1 and 6pgd-J' for each of the diploid species, Locus 6pgd-1 seems to be monomorphIC for both speCies, carrying t:\e allozymes lo( and lep or identical nllgrallon, The duplicaled locus 6pgd-J' has genome speCIfiC aUozymes 201 and 2cp for B. oleracea and B, campesirls. respectively. 111e middle bands ror each dIploid are in(c;locus heterodimers. I n E. napus the allozymes 201 and 2cp f<11 m an additional Illtcrlocu.,>, interspecific heterodlmer right below the B. oleracea IOterlocus helerodilller. The same pallern .s observed in the monosomic addilion line~ for 6PGD-I. except thal the allozyme 201ls weak due to the presence or only one copy orlne B, oleroCf:o chromosome.
The distinct pattern of rONA fragmc:nts in R uleracea and B, campeslris is useful for the detection and characterization of the addition lines. The maio difference i~ the replacement in B. compeslris of the B. olerocea fragments 1.8 and 0,8 Kb by a 2.6 Kb fragmenl, resulting from the loss of a reslriction site. The sekctive loss of the rONA B, olerace(l 3.4 Kb fragment (RB
phenotype) and the simultaneous presence of the 1,8 and 0.8 Kb fragments in the addition lines indicates that more than one chromosome carry these genes tn Brass/c(l and that they have different restriction siles. One chromosome carries the 1.8 and lhe 0.8 Kb fragments while other carries the 3,4 Kb fragment. This dlffermce In restriction sites indicates initial duplication and subsequent divergence of these seq uenees. The hg;l ter intemi ty of Ihe 1.8 and 0.8 Kb Cragments in lhe addition lines is explained by the presence of a single copy or the RA B, oleTl1cea chromosome versus two coptes in B. /Jupt/s. In our limited SUn'ey for rONA phenotype~ we did not detect any addi:ion lines of only RB phenotype. Furlher testing of the unmarked additton hnes i~ expected to disclose individuals with this phenotype. The organization of the other fragments in these two chromosomes is oot known at thIS point.

765
An important observation was the presence of possible recombinants in some of the progenies, lntergenomic recombination has been reported between the B. oferaceG and S, campeslris genomes by Chiang and Crete (1983) after transferring a disease resistance gene from B. IWPUS to B, olerGcea. In the amphidiploid no evidence of recombination between the two genomes has been reported. 1l1is lack of recombination is likely due to the high diploidization of B. napus resulting in J911 in meiosis. After baekcrossing it 10 the diploid B.
CGl'llpeSlris parent. the loss of paiLng partners for some of the chromosomes migh t result in an increased change of multivalent formation and intergenomic recombinaUon, A possibility that needs further exploration is the prescilce or a paji I ng control mechanism sim ilar to tiwt reported in wheat (Riley et al. 1959: Altia and Robhe len I986). In any e\ en t, reco mbina tion open s th e possi bilit)' of exchanging genes among genomes, an im portant alternative for the lJrassica breeder. Recombination between th~ B. a/ero( ea and B. campeslris genomes supports the view that these species have anginated from a common ancestral genome by aneuploidy and chromosome repattern ing. 'I hese changes migh t have been expedited by translocations which not only rearrange the chromosomes in novel comhinations but also yield tertIary IflSomies by dlstu rballee, in chromosome disjunction (Stebbi ns In 1: Khus 1973).
The breakdown of self-mcompatlbllny In some of Ihe derivative, is also an !I1lerestmg findmg. Self-compalibilily could not b" c'«socI3ted with any speClfk B. oleracea marker. or even With Ih", eXira chromosomes. since IWO diploids were found to be compntible, 111e pOSSibility eXIsts. however. thal haVing the S locus in a trisomic condition might weaken its expressIOn, resulting in self.comrJtible plants, as occurs in the natural amphIdiploid species which are selr-compatible. In such a case, it might be possible to locate the S locus by using these addition lines,
Our work opens the r>ossibihty of generating useful cy logen dic stocks for tite genetic and cvolu tlOnary chamCierizallOn of Br05S!cO specie.,. 1l1c presence of duplicated locl ,n the B, (){eroceo and B. compeslris genomes agrees with the hypNhds of Robbelen (1960), which suggests that the bdS1C dLplolds are mdeed secondary polyploids. Olher evidence is the rema rka ble tol erance of an eu plOldy and Ihe high fertility of the aneuplOlds found m (Lis siudy, Generalion of addnion lines for each of Ihe dIplOid genomes, Including x= 7 genomes from several WIld speCie, IS underway. This WIll allow a com raral;ve study of the Rrass/C(I genomes, as additional markers are developed. A further step will be the generation of synlhelle amph,dlploids between diploids carrying agronom,eallv useful genes such as those determining disease res I' ia nee, pia nl arch i!eCIll re and presence of importa nt chemICal compounds, Generalion of addition lines from these will provide information on the loeation of genes deterrnllling til e va no us hOri'cu It IJ ral tra its observed in the diploids, i, e, curd III cauliflower. headmg in cabbage, root enlargement in turn! ps an d ax III 'Hy bud en largemen tin Brussels sprou ts.
kkno\<,'h'dgl:menrs. We are lIldepled to Charles Rick, Steve ranksley, Judy Greenlee and Margi Oard for reviewing the manuscript: to Vince D'Antonio. )anet Sutes and Mitch
McGru lh for tech meal assistance and to Jane Joh nson for typlllg the manusenpt. Supported by a USDA wmpditive granI86CRCR-I-1926.
References
Appels R, Dvorak J (1982) RelatJVe oaks of divergence of spacer and gene sequences within thc rONA rLgion of species in the Trillu:ae: implications for maintenance of homogeneity of the repeat"'d gene family. Theor Appl Genet 63: 361-365
Arus P (1984) B. oleroceo and B. nopus ,sozymes. Cruci fe rae Newsktt 9:64
Arus P. Orton TJ (1983) Isozyme and linkage relationships of Isozyme loc' in Brassica oleracea. J Hered 74:405--412
AllIa 1. Robbelen G (1986) Cytogenetic rehtionship withm cultivated Brass/co analyzed in amphihaploids from three diploid ancestors. Can j Genet CYlol 28: 323-329
Cateh eside DG (1934) The ch romosomaI relalionshlps 1Il the swede and turnip groups of BraSSica. Ann Bot 48: 60 I 633
CJ.lcheSlde DO (1937) Secondary pairmg In Brass/co olaoceo CytOIOglJ, Jubilee, pp 366-378
Chiang MS, Crete R (1983) Transfer of resistance 10 race 2 of PlasmodlOphora brassicae from Brossica "opus to cabbage (B. oleraceG ssp. COpi/OIG), V The inheritance of resiSlance. Eu ph ytlca 32: 479-483
Coulthart M, Denford KE (1982) Isozyme studies in BrassiCG. I. Electrophoretic techmques for leaf enzymes and cornpanson of B. napus. B, COmpl:SlflS and B. oleracea using phQsphoglucom utase. Can J Plan t Sci 62: 621-630
Delseny M. Cooke R. Penon P (1983) Sequence helerogenelty In radish nuclear ribosomal RNA genes. Plant SCI Lett 30: 107-109
Enckson LR, Siraus NA. Beve rsdorf WD (1983) ReslflctlOn pallerns reveal onglns of chloroplast genomes In Brass/fG amph id iploids, Theor Appl Ge net 65: 201- 206
!lart GE, Tuk~n NA {1983) Ch romosoma I location of ek ve n E/,/lrigio efongala (= Agropyron elongalum) isozyme structural genes, Genet Res 41: 181-202
KaflJechenkQ GD (1922) The number of chromosomes and the genetic correlation of cultivated Cruciferae. Bull Appl Bot Genet Plant Breed 13: 3-14
Keller Wk Armstrong KC (1983) Productton of haplOIds vIa anlher culture in BroSS/ca oleraceo var ilahc(l, Euphyuca 32:151-159
Khush GS (1973) Genetic; Qf aneuploids, Academic Press, New York, 301 pp
umdry BS. Michel more RW (1985) Selection of probes for restriction fragment length analysis from plant genomic clones. Plant Mol Bioi Rep 3: 174-179
Man,atlS T. Fritsch EF, Sam brook J (1982) Molecular cloning, a laboratory manual. Cold Springs Harbor Laboratory, Cold Springs Harbor, New York, 545 pp
Morinaga T (1934) On the chromosome number of Brassica jUlIC/:a and S, nopus, on Ihe hybrid between thc two. and on offspring line of the hybrid. Jpn J Genet 9' 161-163
Mlzu,hlma U (1980) Genome analysis in Bramca and allied genera, fn: Tsunoda S, Hinata K. Gomez-Campos C (eds) Bra,mco nops and wild all ie;, Japan SClen ti fie Sonelles Press, Tokyo. pp 89-106
Nitsch JP, "LISch C (1969) Haploid plants from pollen grains, Science 163: 85
Palmer JD, Shields CR, Cohen DB, Orton TJ {1983) Chloroplast DNA evolution and the Origin of amphidiploid BroIs/ca ;peCles. llleOf Aprl Genet 65: 18 1~ 189

766
Perney EB, Corgan IN, Horak KE, Tanbiey SD (InS) Electrophoretic analysis of AllIUm alien :"idJllon lJOes. Theor Appl Genet 71: J76- 184
Prakash S, Hinata K (1980) Taxonomy, cytogenetics and ongin of crop Brassicas, a review, Opera Bot 55: I-57
Quiros CF, McHale (1985, Genellc analysIs of Isozyme variants in diploid and tetraploid potatoes, Genetics 111:131-145
Quiros CF. Kianian SF. Ochoa 0, Douches 0 (/985) Genome evolution in Brussicu: use of molecular markers and cy· togenetic stocks, Cruci ferae Newsle II 10: 21-23
Riley K Chapman Y, Kimber G (1959) Genelle control of chromosome pairing in intcrgcaeric hybnds With wheal. Nature 183:1244-1245
Robbelen G (1960J Beitr1\ge zur Analyse des BraSSlcaGenomes. Chromosoma I): 205-228
Robbins MP, Vaughan 'JG (J 983) Rubisco in Brassicaceae, In: Jensen U. Fairbrothcrs DE (cds) Proteins and nucleiC acids In plant systematics. Spri'.'gcr. Berlin. pp 191-204
Saghai-Maroof MA. Sollman KM, Jorgensen RA Allard RW (1984) Ribosomal DNA spacer-length polymorphisms In
barley, lvkndelian inherilanct'. chromosomal locallQn, and population dynamiCS, PNAS lSA 81:80!4·-8018
Sear$ ER (1969) Wheal cytogenetics. Annu Rev Genet 3:451-468
Sikka SM (1940) Cytogenellcs of Bramca hybrids and species. J Genet 40: 441 - 509
Stebbins GL (1971) ChromosQmal evolution III hIgher plants Addison-Wesley, California
SW31lllnathan MS,' Magoon ML. Mehra KL (1954) A sImple propionlC.carmine PMC smcar method for plants with small chromosomes, Ind J G~r.et Plant Breed 14:87--88
U. N (1935) Genomic anaf"SIS JO Brassia} wtth spedal reference to the expenmental formatIOn of B, napils and peculiar mode QfferllhZ<l.lion.lpn J Genet 7:389-452
Weeden NF. Goltlieb LD (1980) Isolation of cytoplasmic enzymes from pollen, Planl PhysiQI 66: 4DO-403
Williams PH, Hill CB (1986) Rapid-cychllf populations of Brassiw, SCience 232: 1385-1389