an improved method for utilizing high-throughput amplicon ... · 1 an improved method for utilizing...
TRANSCRIPT

1
Animprovedmethodforutilizinghigh-throughputampliconsequencingtodeterminethe1
dietsofinsectivorousanimals2
3
Authors4
MichelleA.Jusino1++*,MarkT.Banik1++,JonathanM.Palmer1,AmyK.Wray2,3,LeiXiao4,Emma5
Pelton3,5,JesseR.Barber6,AkitoY.Kawahara4,ClaudioGratton3,M.ZachariahPeery2,and6
DanielL.Lindner1*7
8++indicatessharedfirstauthorshipbasedonequalcontributions9
*indicatescorrespondingauthors10
11
Affiliations12
(1) UnitedStatesForestService,NorthernResearchStation,CenterforForestMycology13
Research,OneGiffordPinchotDrive,Madison,Wisconsin,USA14
(2) UniversityofWisconsin-Madison,DepartmentofForestandWildlifeEcology,Madison,15
Wisconsin,USA16
(3) UniversityofWisconsin-Madison,DepartmentofEntomology,Madison,Wisconsin,USA17
(4) McGuireCenterforLepidopteraandBiodiversity,FloridaMuseumofNaturalHistory,18
UniversityofFlorida,Gainesville,USA19
(5) TheXercesSocietyforInvertebrateConservation,Portland,Oregon,USA20
(6) DepartmentofBiologicalSciences,GraduatePrograminEcology,Evolutionand21
Behavior,BoiseStateUniversity,Boise,Idaho,USA22
23
*MichelleA.Jusino:USFSNRSCFMR,OneGiffordPinchotDrive,Madison,WI,5372624
[email protected],[email protected]
*DanielL.Lindner:USFSNRSCFMR,OneGiffordPinchotDrive,Madison,WI,5372626
[email protected],[email protected]
28
Runningtitle:ImprovedHTSofinsectivorediets 29
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

2
Abstract30
DNAanalysisofpredatorfecesusinghigh-throughputampliconsequencing(HTS)31
enhancesourunderstandingofpredator-preyinteractions.However,conclusionsdrawnfrom32
thistechniqueareconstrainedbybiasesthatoccurinmultiplestepsoftheHTSworkflow.To33
bettercharacterizeinsectivorousanimaldiets,weusedDNAfromadiversesetofarthropodsto34
assessPCRbiasesofcommonlyusedandnovelprimerpairsforthemitochondrialgene,35
cytochromeoxidaseCsubunit1(CO1).WecompareddiversityrecoveredfromHTSofbat36
guanosamplesusingacommonlyusedprimerpair“ZBJ”toresultsusingthenovelprimerpair37
“ANML”.Toparameterizeourbioinformaticspipeline,wecreatedanarthropodmock38
communityconsistingofsingle-copy(cloned)CO1sequences.Toexaminebiasesassociated39
withbothPCRandHTS,mockcommunitymemberswerecombinedinequimolaramountsboth40
pre-andpost-PCR.Wevalidatedoursystemusingguanofrombatsfedknowndietsandusing41
compositesamplesofmorphologicallyidentifiedinsectscollectedinpitfalltraps.InPCRtests,42
theANMLprimerpairamplified58of59arthropodtaxa(98%)whereasZBJamplified24of5943
taxa(41%).Furthermore,inanHTScomparisonoffield-collectedsamples,theANMLprimers44
detectednearlyfour-foldmorearthropodtaxathantheZBJprimers.Theadditionalarthropods45
detectedincludemedicallyandeconomicallyrelevantinsectgroupssuchasmosquitoes.46
ResultsrevealedbiasesatboththePCRandsequencinglevels,demonstratingthepitfalls47
associatedwithusingHTSreadnumbersasproxiesforabundance.Theuseofanarthropod48
mockcommunityallowedforimprovedbioinformaticspipelineparameterization.49
50
51
Keywords:arthropodmockcommunity,batguano,dietaryanalysis,insectivore,next-52
generationsequencing,NGS53
54
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

3
Introduction55
High-throughputampliconsequencing(HTS)hasbecomethepreferredmethodforrapid56
molecularidentificationofmembersofmixedecologicalcommunities.HTSisnowalso57
increasinglyusedtoidentifythearthropoddietarycomponentsofawidetaxonomicrangeof58
animalsincludingmammals(Busscheetal.2016;Clareetal.2014a;Clareetal.2014b;Mallott59
etal.2015;Rydelletal.2016;Vesterinenetal.2016),birds(Crisol-Martínezetal.2016;Jedlicka60
etal.2016;Trevellineetal.2016),reptiles(Kartzinel&Pringle2015),fish(Harms-Tuohyetal.61
2016),andarthropods(Krehenwinkeletal.2016).IdentificationoftheDNAofdietary62
componentsisaccomplishedby“metabarcoding”,whichinvolvesextractingDNAfromfecal63
samples,amplifyingoneormorebarcodingloci,preparingDNAlibraries,andfinallysequencing,64
bioinformatics,anddataanalysis.Eachofthesestepsinvolvesdecisionsandassumptionsthat65
significantlyaffectresults.Forexample,biasesareunavoidablewhenamplifyingenvironmental66
DNAwithPCR-basedmethods(Brooksetal.2015)andcarefulconsiderationshouldbe67
exercisedwhenselectingaprimerpairforHTS.Thus,whileDNAmetabarcodingisapowerful68
toolforstudyingtrophicinteractions,conclusionsshouldtakeintoaccounttheshortcomings69
andparametersofthetechniques(e.g.:Brooksetal.2015;D’Amoreetal.2016;Lindahletal.70
2013;Nguyenetal.2015;Pompanonetal.2012).71
ThemitochondrialcytochromeoxidaseCsubunit1locus(CO1)isthemostfrequently72
usedbarcodinglocusforidentifyingawiderangeoftaxonomicgroups,includingarthropods.73
BecauseCO1hasthemostextensivereferencelibraryforarthropods(BOLDsystems,74
Ratnasignham&Hebert,2007),itisthemostcommonlyusedlocusfordietarystudiesof75
insectivorousanimals(Clarkeetal.2014).TheentireCO1barcodingregionisabout658base76
pairs(bp)andcurrentlytoolongtobeusedwithmostHTSplatforms.Thereforeitisnecessary77
tosequenceshorterregionsoftheCO1locus,whichhasprovenchallengingduetoalackof78
conservedprimingsiteswithintheCO1region(Deagleetal.2014).Therefore,novelprimer79
pairsshouldbetestedagainstasmanyexpectedtargetDNAsequencesaspossible.80
Zealeetal.(2011)developedtheZBJ-ArtF1c/ZBJ-ArtR2c(hereafterZBJ)primerpairfor81
detectingarthropodpreyDNAinbatguanobyamplifyinga157bpfragmentoftheCO1region.82
Intheinitialstudy,whichemployedcloningandsequencingratherthanHTS,theZBJprimers83
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

4
amplified37taxafrom13arthropodorders,butdidnotamplifybatCO1DNA.TheZBJprimers84
weredesignedtotargetashortfragmentinordertoamplifythepresumablydegradedDNA85
presentinguanoandcoincidentallythelengthoftheamplicongeneratediscompatiblewith86
manyHTSplatforms.Subsequently,numerousresearchershaveemployedtheZBJprimersin87
HTSstudiesthatanalyzedietsofinsectivorousanimals,includingbats(Busscheetal.2016;88
Clareetal.2014a;Clareetal.2014b;Rydelletal.2016;Vesterinenetal.2016)andbirds89
(Crisol-Martínezetal.2016;Jedlickaetal.2016;Trevellineetal.2016).AlthoughtheZBJ90
primershavebeenwidelyutilized,thereareindicationsthattheyhaveanarrowtaxonomic91
range(Brandon-Mongetal.2015;Clarkeetal.2014;Mallottetal.2015).92
TheassumptionsandparameterscommonlyemployedinHTSenvironmentalDNA93
analyseshavealargeimpactontheoperationaltaxonomicunits(OTUs)thatarerecovered.94
Bioinformaticsclusteringalgorithmscaninfluenceapparentdiversitywithinasample,oran95
entirelibraryofsamples,andtrimmingandfilteringparameterscanimpacttheresulting96
communitycomposition(Deagleetal.2013).Avalidationorcontrolisneededtoparameterize97
bioinformaticspipelines;therefore,theuseofmockcommunitiesaspositivecontrolsinHTSis98
increasinglybecomingcommon,especiallyamongresearcherswhoworkwithfungaland99
bacterialcommunities(Bokulich&Mills2013;Bokulichetal.2013;Nguyenetal.2015).Mock100
communitiescanbeusedtoexaminebiases,startingatthesamplingstepandendingatthe101
bioinformaticsandcommunityanalysissteps.102
Hereweusedareferenceinsectcommunitytoidentifyspecificamplificationbiases103
associatedwiththreecommonlyusedprimerpairs,includingZBJ,andtwonovelprimerpairs,104
LCO1-1490/CO1-CFMRa(hereafterANML)andLCO1490/CO1-CFMRb(hereafterCFMRb),for105
theCO1region(Table1).Tofurthertestprimers,wecomparedHTSresultsfromtheZBJ106
primerstoournovelANMLprimerpairusingfield-collectedbatguanosamples.Wedesigned107
anarthropodmockcommunitybasedonsingle-copy(cloned)mitochondrialCO1sequences,108
whichcanserveasastandardinHTSsequencingandtohelpparameterizeabioinformatics109
pipeline.Finally,wevalidatedtheaccuracyofoursystemofnovelprimers,themock110
communitycontrol,andourbioinformaticspipelinebyusingguanofrombatsfedknowninsect111
dietsandcompositesamplesofmorphologicallyidentifiedarthropodsfrompitfalltraps.112
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

5
MethodsandMaterials113
Testingofprimerpairsagainstknowninsectsamples114
DNAwasextractedfrom67arthropodtaxa(Table2)followingtheprotocolinLindner&115
Banik(2009)withmodificationsforinsects(SupportingInformation,AppendixS1).Briefly,DNA116
wasextractedfromexcisedlegmusclesoflargerinsectsor,forsmallerinsects,thethoraxwas117
puncturedandtheentireinsectwasusedforextraction.Legmusclesandsmallinsectswith118
puncturedthoraxeswereplacedin100μLoffilteredcelllysissolution(CLS;Lindner&Banik119
2009)andfrozenat-20°C,andtheextractionproceeded.FollowingDNAextraction,the120
effectivenessofthefollowingfiveprimerpairsinamplifyingthe67purifiedDNAswas121
evaluated:LCO1490/HCO2198(Folmeretal.1994;Hebertetal.2003;hereafterCO1L/H),ZBJ-122
ArtF1c/ZBJ-ArtR2c(Zealeetal.2011;ZBJ),LCO1-1490/CO1-CFMRa(ANML),LCO1490/CO1-123
CFMRb(CFMRb),andLepF1/mLepR(Hebertetal.2004;Smithetal.2006;hereafterLEP).The124
CO1-CFMRaandCO1-CFMRbprimersdesignedforthisstudywerederivedfromtheZBJ-ArtR2c125
primerandhadsequencesof5’-GGWACTAATCAATTTCCAAATCC-3’and5’-126
GGNACTAATCAATTHCCAAATCC-3’,respectively.TheCO1-CFMRaandCO1-CFMRbprimingsites127
arelocatedintheCO1geneapproximately180bpawayfromtheLCO1490primingsite128
(SupportingInformation,FigureS1).AlistoftheprimersusedispresentedinTable1.129
AmplificationoftheextractedDNAusingallprimerpairs,exceptZBJ,usedthefollowing130
reagentvolumesper15LμLreaction:7.88μLDNA-freemoleculargradewater,3μLGreen131
GoTaq5xbuffer(Promega),0.12μLof20mg/mLBSA,0.3μLof10μMdNTPs,0.3μLofeach10132
μMprimer,0.1uLof5u/μLGoTaqpolymerase(Promega),and3μLofextractedarthropod133
templateDNA.TheZBJprimerpairwasusedwithtwodifferentreagentregimes.One,termed134
themodifiedprotocol,wasthesameasaboveexcept1.0μLofeach10μMprimerwasadded135
andthesecondregimewasthatdescribedbyoriginalauthors(Zealeetal.2011).The136
thermocyclerparametersfortheCO1L/H,ANML,andCFMRbprimerpairswerethosedescribed137
byHebertetal.(2003)withonemodification:thefinalextensionat72°Cwasincreasedfrom5138
to7minutes.TheLepfF-1/mLepR-5amplificationparameterswerethoseofSmithetal.(2006),139
whiletheZBJprimerpairamplificationparameterswerethosedescribedbyZealeetal.(2011).140
Followingamplification,3μLofproductwasrunina2%agarosegelfor20minutesat110V,141
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

6
stainedwithethidiumbromideandvisualizedusingUVlight.Presenceorabsenceofbandswas142
recordedforeachprimerpairandDNAcombination.Toprovidereferencesequencesforeach143
speciestheLCO1490/HCO2198PCRproductswereSangersequencedwithABIPrismBigDye144
(AppliedBiosystems)sequencingfollowingthemethodofLindner&Banik(2009).Theresulting145
sequencesweresubjectedtoanNCBIBLASTsearchtoconfirmtheidentitiesoftheinsect146
speciesoforigin.147
HTSoffield-collectedguanosamplesusingtwodifferentprimerpairs148
ThearthropodDNApresentinthreefield-collectedbatguanosampleswasanalyzed149
usingtheANMLandZBJprimerpairs.DNAwasextractedfromthreeMyotislucifugusguano150
samplesfromthreedifferentlocationsinsouthernWisconsin(allcollectionswereapprovedby151
theWisconsinDepartmentofNaturalResources).Onesamplecontaining100mgofguano,152
approximately10pellets,wasextractedfromeachsiteusingQIAampDNAstoolMiniKit153
followingtheprocedureinAppendixS2oftheSupportingInformation.TheDNAwasthen154
amplifiedusingprimersmodifiedformetabarcodingbyaddinganIonTorrentXpresstrP1155
adaptersequenceonthereverseprimerandbarcodesequenceandIonTorrentXpressA156
adaptersequenceoneachforwardprimer(seeSupportingInformation,TableS1forbarcoded157
primersequences).AmplificationconditionsfortheANMLpairfollowedtheprotocolusedfor158
theprimerpairtestandconditionsfortheZBJpairfollowedthemodifiedprotocolforZBJ159
describedintheprimerpairtest.Followingamplification,eachoftheuniquelybarcodedPCR160
productswaspurifiedviasizeselectingE-GelCloneWellGels(Invitrogen)atapproximately161
180bp.Thesize-selectedproductswerethenquantifiedonanInvitrogenQubit2.0Fluorometer162
andbroughttoaconcentrationof2000pMusingDNA-free,moleculargradewater.Wethen163
combinedtheproductsinequalamountstoproducethesequencinglibrary.Thelibrarywas164
dilutedto13pMpriortotemplatingontoionsphereparticles(ISPs)withtheIonOneTouch2165
system(LifeTechnologies)andaPGMHi-QOT2templatingkit(ThermoFisher#A27739),166
accordingtothemanufacturer'srecommendations.ThetemplatedISPswerethenpurifiedand167
thetemplatedDNAwassequencedusingtheIonTorrentPersonalGenomeMachine(PGM;168
ThermoFisher)withtheIonPGMHi-QSequencingKit(ThermoFisher#A25592)accordingtothe169
manufacturer'sprotocolfor400bpsequencing.170
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

7
Bioinformatics171
HTSdatawereprocessedusingthe‘DADA2’methodviatheAMPtkpipeline172
(https://github.com/nextgenusfs/amptk).Briefly,theAMPtkpipelineprocesses(de-173
multiplexes)HTSampliconsequencingreadsby:1)identifyingavalidbarcodeindexineach174
read,2)identifyingforwardandreverseprimersequences,3)trimmingbarcodeandprimer175
sequences,4)renamingthereadbasedonbarcodeindex,and5)trimming/paddingthereads176
toasetlength.TheDADA2algorithm(Callahanetal.2016)isanalternativetowidelyused177
sequence-clusteringalgorithms(e.g.,UPARSE,UCLUST,nearest-neighbor,SWARM,etc.)and178
functionsto“denoise”HTSsequencingreads.DADA2hasbeenshowntobeveryaccurateand179
issensitivetosinglebasepairdifferencesbetweensequences(Callahanetal2016).AMPtk180
implementsamodifiedDADA2algorithmthatproducesthestandard“inferredsequences”181
outputofDADA2aswellasclustersthe“inferredsequences”intobiologicallyrelevantOTUs182
usingtheUCLUST(Edgar2010)algorithmemployedinVSEARCH(Rognesetal.2016).The183
resultingAMPtkOTUtablescanbefilteredbasedonspike-inmockcommunities(described184
below).TaxonomyformtCO1isassignedinAMPtkusingacombinationofglobalsequence185
alignment,UTAX(http://www.drive5.com/usearch/manual/utax_algo.html),andSINTAX(Edgar186
2016)usingaCO1referencedatabase.ThecurrentCO1databasedistributedwithAMPtkwas187
derivedfromcollatingsequencesfromrepresentativebarcodeindexnumbers(BIN)from188
chordatesandarthropodsintheBarcodeofLifev4database(BOLD;Ratnasingham&Hebert189
2007)andisavailableat:190
https://github.com/nextgenusfs/amptk/blob/master/docs/reference_databases.md.191
Developmentandtestingofanarthropodmockcommunity192
ToproduceamockcommunitytoserveasacontrolforHTSdataanalysis,43ofthe193
arthropodtaxausedintheprimerpairtestwerechosenascandidates(Table2).DNAfromeach194
arthropodwasamplifiedusingLCO1490/HCO2198primersasdescribedpreviously.Toremove195
intragenomicvariation(Songetal.2008),theresultingampliconswereclonedintoE.coliusing196
thePromegapGem-Tvectorsystemfollowingthemanufacturer’sinstructionswiththe197
modificationsusedbyLindner&Banik(2009).Threeclonesofeacharthropodtaxonwere198
subsequentlySangersequencedtoverifythepresenceoftheCO1insertsequence.Twoofthe199
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

8
clonedarthropodsproducedclonedsequencevariants,andthesevariants(3total)werealso200
includedinthemockcommunity,bringingourmockcommunitytotalto46.Plasmidswere201
purifiedusingstandardalkalinelysisandtheresultantDNAwasthenquantifiedonan202
InvitrogenQubit2.0Fluorometerandbroughttoaconcentrationof1500pMusingDNA-free,203
moleculargradewater.Plasmidswerethendiluted1:20usingDNA-freemoleculargradewater204
andindividuallyamplifiedusingtheionANMLprimerswiththesamebarcode.Theindividual205
PCRproductswerethenvisualizedona2%agarosegel,cleanedandsizeselectedat≥150bp206
usingZymoResearchSelect-A-SizeDNAClean&Concentratorspincolumns,quantifiedand207
equilibratedto2000pMasdescribedpreviously,andsubsequentlycombinedinequal208
amounts.Thisampliconmixtureisreferredtoasour“post-PCRcombinedmockcommunity”,209
whichservesasacontroltovalidatesequencingefficiencyofeachmockmember.Tomeasure210
initialPCRbiasandtoparameterizeourbioinformaticspipeline,wealsocreated“apre-PCR211
combinedmockcommunity”bycombiningour1500pMplasmidsinequalamounts.Thepre-212
PCRcombinedmockcommunitywasthendilutedtoa1:8000concentrationpriorto213
amplificationwithANMLbarcodedprimers.TheresultingbarcodedPCRproductwasthen214
visualized,sizeselected,quantified,andbroughtto2000pMasdescribedbefore.Theresulting215
barcodedPCRproductswerethenpreparedandsequencedonanIonTorrentPGManddata216
werebioinformaticallyprocessedasdescribedabove.217
Testingofknownmixedsampleswithmockcommunityandourpipeline218
TotestpreyDNArecoveryfrombatguano,twobats,oneEptesicusfuscusandone219
Lasiuruscinerus,werefedknowndietsofGalleriamellonella,TenebriomolitorandAntheraea220
polyphemusaloneandincombination(Table4).Thebatswerefedeachknowndietforone221
day,andguanopelletswerecollectedduringthefollowing24hours(approvedbyBoiseState222
UniversityInstitutionalAnimalCareandUseCommittee006-AC14-018).Weanalyzedthree223
knowndietcombinationsfromtheE.fuscusindividualandtwoknowndietcombinationsfrom224
theL.cinerusindividual.DNAwasextractedfromguanosamplesusingQiagenQIAampmini225
Stoolkits,followingthemodifiedprotocoldescribedinZealeetal.(2011).DNAfromtheknown226
dietsampleswasamplifiedwithbarcodedANMLprimers,andtheresultingPCRproductswere227
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

9
thenvisualized,sizeselectedat≥150bpusingZymoResearchSelect-A-SizeDNAcleanand228
concentratorspincolumns,quantified,andbroughtto2000pMasdescribedbefore.229
Totesttheeffectivenessofthemethodoncomplexinsectcommunities,fivesamples230
frompitfalltrapsfromtheSnakeRiverBirdsofPreyConservationAreainKuna,Idahowere231
analyzed.Eachpitfalltrapconsistedofaglassjarcontainingpropyleneglycol.Trapswereleft232
outsidefor2-3days,atwhichpointthecontentsofthetrapswererinsedwith100%ethanol233
andsubsequentlytransferredtoglassvialscontaining100%ethanolforstorageatroom234
temperature.AlltrapsamplesweresenttotheFloridaMuseumofNaturalHistoryin235
Gainesville,Floridaforvisualidentificationtoarthropodfamilyandlong-termstorageat-20°C.236
Initialidentitiesofthearthropodspresentintheinsecttrapsampleswereobtainedusing237
traditionalmorphologicalkeys,andmostwereidentifiedbyeyetothefamilylevel,withthe238
followingexceptions:allspringtailswereidentifiedtoorder(Collembola),centipedeswere239
identifiedtoclass(Chilopoda)andmiteswereidentifiedtosubclass(Acari).240
ThesamplesweresenttotheUnitedStatesForestService,NorthernResearchStation,241
CenterforForestMycologyResearchinMadison,Wisconsin,wheretheywereprocessedfor242
molecularanalysis.ArthropodsfromthetrapsampleswererinsedinDNA-freemoleculargrade243
waterandpreparedforDNAextractionintwoways:(1)theexcisedlegmusclesoflarger244
arthropods,andsmallerarthropodswithopenthoraxeswerecombinedandsubmersedinCLS245
andvortexed(dissectedsample),or(2)theintactarthropodswereaddedto15mLCLSand246
maceratedwithasterilepestleandvortexed(maceratedsample).DNAextractionfollowed247
detailsdescribedinAppendixS1oftheSupportingInformation;metabarcodingPCR,andHTS248
thenproceededaspreviouslydescribed.Datawerebioinformaticallyprocessedasdescribed249
before.250
Results251
Testingofprimerpairsagainstknowninsectsamples252
Fifty-eightofthe59taxa(98%)amplifiedwiththeANML(LCO1490/CO1-CFMRa)and253
CFMRb(LCO1490/CO1-CFMRb)primerpairs,withbothpairsfailingtoamplifythesamecarabid254
beetle(Table2).Fifty-twoof59taxa(89%)amplifiedwiththeCO1L/Hprimerpairand48of59255
(81%)amplifiedwiththeLEPprimerpair;theLEPpairamplified100%oftheLepidopteransand256
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

10
Dipteranstested(Table2).TheZBJprimerpairamplified24ofthe59(41%)taxatestedwiththe257
Zealeetal.(2011)protocoland27ofthe59(46%)taxausingourmodifiedprotocol.TheZBJ258
primerpairsuccessfullyamplifiedatleastonerepresentativefromeacharthropodordertested259
(Table2).260
HTSoffield-collectedguanosamplesusingtwodifferentprimerpairs261
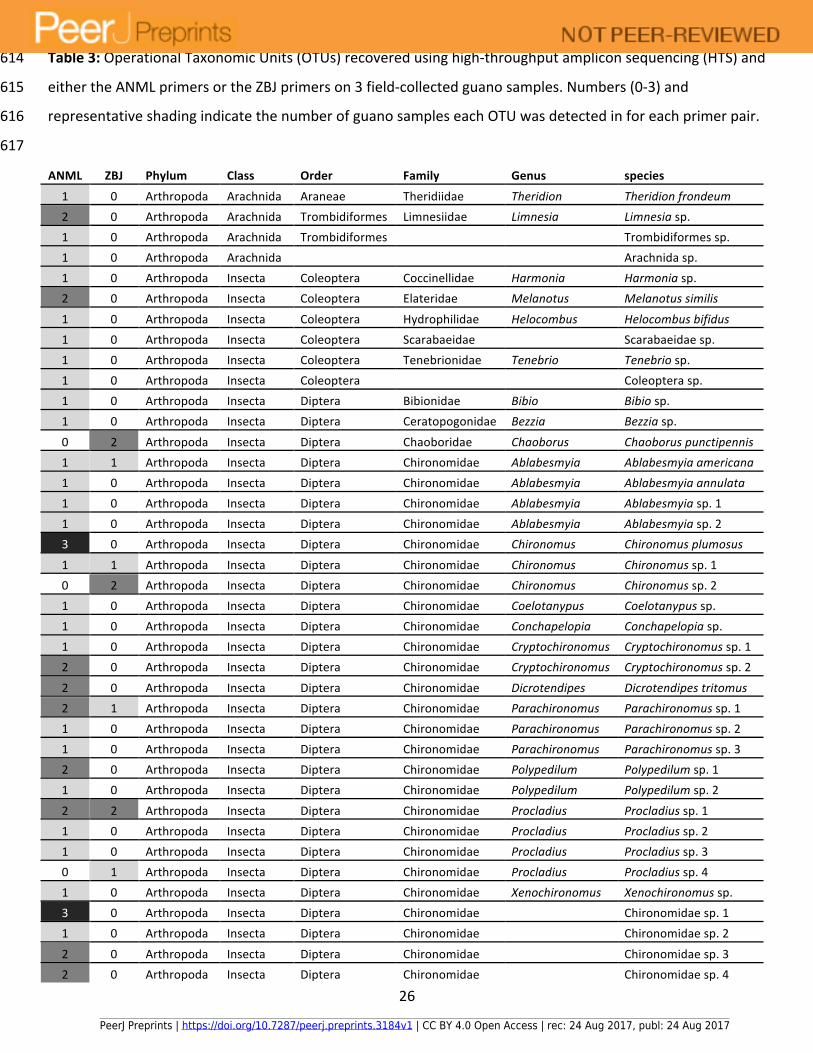
BoththeZBJandtheANMLprimersproducedanamplificationproductfromthethree262
Myotislucifugusguanosamples.Forbothprimersetscombined,atotalof64OTUs(Table3)263
weredetected,ofwhich59couldbeidentifiedtothefamilylevel,representing10orders264
comprisedof28families.TheANMLprimersdetected56OTUsandtheZBJprimersdetected15265
OTUs.Sevenofthe64totalOTUsweredetectedwithbothsetsofprimers,49weredetected266
onlywiththeANMLprimerswhile8wereonlydetectedwiththeZBJprimers.Representatives267
fromalltenordersand26familieswererecoveredusingtheANMLprimerpair,whiletheZBJ268
pairrecoveredrepresentativesfromthreeordersandeightfamilies.Themostoftendetected269
familywasthedipteranmidgefamilyChironomidae,with27OTUs,24ofwhichweredetected270
bytheANMLprimersand6byZBJ.Thesecondmostoftendetectedfamilyweremosquitoes271
(Family:Culicidae),with5OTUsdetectedbyANMLbutonlyonebyZBJ.Allbutoneofthe272
remainderofthefamilieswererepresentedbyonlyoneOTUeach.273
Developmentandtestingofanarthropodmockcommunity274
Theindividualplasmidcomponentsofourpost-PCRcombinedmockcommunity275
generatedreadcountsthatrangedfrom3740to4;themeanwas2119andstandarddeviation276
+/-799,with89%(41outof46)yieldinggreaterthan1500reads(Figure1;supplementaltable277
1).Allmockmembersinthepost-PCRcombinedcommunitywererecovered,although3278
generatedfinalreadcountsbelow100(range4to12).Incontrast,individualmembersofour279
pre-PCRcombinedmockcommunitygeneratedreadcountsthatrangedfrom10,577to0witha280
meanof2174andstandarddeviationof+/-2238,with54%(25of46)yieldingmorethan1500281
reads.Twoofourmockmembersdidnotgenerateanysequencesinthepre-PCRcombined282
communityandanadditional4generatedfinalreadcountsbelow100(range2to39).283
Testingofknownmixedsampleswithmockcommunityandourpipeline284
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

11
TheresultsoftheknowndietHTSsamplesaresummarizedinTable4.WedetectedDNA285
fromalloftheexpecteddietarycomponentsinall5oftheknowndietsamplestested.286
Additionally,wedetectedDNAfromtwopossibleaccidentaldietarycomponents(Empria287
takeuchiiandAgrotisipsilon)inbigbrownbat(Eptesicusfuscus)dietsamplesthatincluded288
bothGalleriaandTenebrioasdietarycomponents.WealsodetectedDNAfromaparasitoid289
wasp(Family:Ichneumonidae)in3of4(75%)dietsamplesthatincludedGallerialarvae.Finally,290
wedetectedbigbrownbat(E.fuscus)DNAin2ofthe3samplesfrombigbrownbats,and291
hoarybat(L.cinereus)DNAinboth(2of2)ofthesamplesfromhoarybats(Table4).Thesedata292
wereprocessedbioinformaticallywithDADA2,withandwithout97%clusteringappliedtothe293
inferredsequencetablethatresultedfromtheDADA2output.Withoutclustering,weobtained294
oneinferredsequenceforG.mellonella,Antheraeapolyphemus,E.takeuchii,A.ipsilonandE.295
fuscus,butobtained11inferredsequencesforTenebriomolitor,7fromIchneumonidae,and3296
forL.cinereus.Afterclusteringat97%,wemaintainedtheOTUnumberforalltaxathathadone297
OTUbeforeclustering,andobtained2OTUsforT.molitor,1OTUforIchneumonidae,and2298
OTUsforL.cinereus.299
Theresultsofthepitfalltrapsamplesaresummarizedbasedonpresenceorabsenceof300
familiesinTable5.Thereappearstobenosignificanteffectofthemethodinwhichthe301
communitieswereextracted(dissectedsamplesormaceratedsamples)ontheefficiencyof302
taxonrecovery.Overall,in5samples37familiesidentifiedusingconventionalmorphological303
methodswerealsorecoveredwithHTS,whileafurther18familiesmorphologicallyidentified304
werenotrecoveredwithHTSand16familieswereonlyrecoveredwithHTS.Ofthe18families305
missedbyHTS,9wereprobablyaresultofeitheramorphologicalorsequence306
misidentification,withtheremaining9mostlikelylostthroughsystembias.307
Discussion308
Throughanamplificationtestof5primerpairsagainstataxonomicallydiverse309
communityofarthropods,wedemonstratedthatourANMLandCFMRbprimerpairsamplified310
moretaxathanpreviouslydescribedprimerpairs(CO1L/H,ZBJ,andLEP)inastandardPCR.311
Throughadirectcomparisonoffield-collectedguanosamplessubjectedtoHTSwithtwoprimer312
pairs,ANMLandZBJ,wedemonstratedthattheANMLprimerpairamplifiedsubstantiallymore313
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

12
taxathantheZBJprimers,thecommonlyusedprimerpairforHTSstudiesexaminingthediets314
ofinsectivorousanimals.Whenweusedbothpairsonthesameenvironmentalsamples,the315
ANMLpairyieldedalmostfourtimesasmanyarthropodtaxathantheZBJpair.Wealso316
detectedchiropteran(bat)sequencesinfecalsamplesfrombatsusingtheANMLprimerpair,317
althoughthenumberofchiropteranDNAsequencesandOTUswasinsignificantcomparedto318
theoverallnumberofsequencesgenerated.Thus,theamplificationofchiropteranDNAdidnot319
significantlyimpacttherecoveryofarthropodDNA,afeaturethathelpsconfirmtheidentityof320
thebattargetspecies,aswellastheirdietarycomponents.ItislikelythattheCO1regionof321
othervertebratescouldalsobeamplifiedbytheANMLprimers,thushelpingtoconfirmthe322
identityoftheconsumerinarangeofsystems(e.g.othermammalspecies,reptiles,323
amphibians,andbirds).BecausetheyproducelongerPCRproducts,theANMLprimers(180bp324
product)alsoallowforbettertaxondelineationcomparedtotheZBJprimers(157bpproduct).325
Improveddetectionofpestspecies326
Insectivorousanimalsarevaluedasprovidersofpestcontrol;however,thetotal327
economicvalueofthisecosystemserviceisdifficulttoestimate(Boylesetal.2011;Clevelandet328
al.2006;Maine&Boyles2015;Williams-Guillénetal.2016).Determiningthefullvalueis329
dependentonthereliabledetectionofthepestspeciespresentinthedietsofinsectivorous330
animals.HTScanbeapowerfultoolforhelpingtobuildtheempiricalbasisnecessaryto331
estimateecosystemservices,butthesuccessofthisapproachdependsinpartonprimer332
efficacy.Basedonouranalyses,theANMLprimersareamajormethodologicalimprovement333
overexistingprimers,allowingforthedetectionofgreaterarthropoddiversityinthe334
environmentalsampleswetested,includingagreaterdiversityofknownpestssuchas335
mosquitoes(Family:Culicidae).Theprevalenceofmosquitoesisusuallyverylowinother336
molecularstudiesofbatguanothatrelyupontheZBJprimers(Clareetal.2014a;Clareetal.337
2014b;Gonsalvesetal.2013;Rolfeetal.2014),andsomehavegoneasfarastosaythat338
mosquitoesarenotimportantpreyitemsforbats(Fenton2012).Specifically,inourguano339
samples,theZBJpairwasonlyabletodetectAedesvexans,whiletheANMLpairdetectedA.340
vexansplusfourotherCulicidaespeciesinthesamesamples.Thus,theANMLprimersallowfor341
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

13
betterestimationoftheecosystemservicesofbats,andperhapsotherinsectivores,as342
predatorsofmosquitoesandothereconomicallyimportantpestspecies.343
Single-copyarthropodmockcommunity,sourcesofunexpectedvariationandsomesolutions344
WhilesomeauthorshavenotedthatHTSdataareunreliableasasourcetomeasure345
communitymemberabundance(Piñoletal.2015),manyHTSstudiesofenvironmentalsamples346
continuetouseabundancemetricsbasedonreadnumbers.Totestthevalidityofreadnumber347
asanestimateofrelativeabundance,wecombinedpre-andpost-PCRmockcommunitiesin348
equimolaramountspriortosequencing.Wepredictedthatiftheapproachisvalid,read349
numbersshouldbeequalacrosstaxa.Instead,eventhougheachmemberofthemock350
communityamplifiedwellinindividualPCRs,weobservedalargevariationinreadnumbersfor351
thepre-PCRcombinedmockcommunity,withsomemembersbeingabsent.Incontrast,the352
post-PCRcombinedmockwasfarlessvariable(Figure1).TheinitialPCRintroducedalarge353
amountoftaxonomicbiasbypreferentiallyamplifyingsometaxa,asinferredfromthe354
differenceinvariabilityinreadnumbersbetweenthepostandpre-PCRmixesofourarthropod355
mockcommunity.Sequencingitselfalsointroducedbiasresultingindifferencesinread356
numbersbetweenthemockmembersthatwerecombinedpost-PCR.Someofthevariationin357
readnumbersamongmockcommunitymemberswasprobablyinducedbymismatchesinthe358
primingsite,giventhatsomememberspossessingthreeormoreprimermismatches.While359
thisnumberofmismatchesdidnotinhibitamplificationinindividualPCRs,inacompetitive360
mixedPCRthemismatchescouldresultinanamplificationbias.Differencesinreadnumbers361
canalsobeattributedtosequencecharacteristicssuchashomopolymerregionsandGC362
content.Ourmockcommunitydatademonstratedthatusingreadnumbersasproxiesfor363
abundanceinenvironmentalsamplesisproblematic,especiallyincomplexsamples.364
Becauseourarthropodmockcommunityconsistsofsingle-copyclonedplasmids,we365
expectedtofindonlyoneOTUpermockmember,allowingtheconclusiveidentificationof366
spuriousorchimericsequencesgeneratedduringthesequencingprocess.Someofthese367
chimerasaretheresultofsimplebinningerrorsandothersaretruechimeras(i.e.,hybrid368
sequencesasaresultofPCRandsequencingerror).Acriticalcomponentofchimerafilteringis369
havingacurateddatabaseofreferencesequences.Weinitiallyattemptedtouseallavailable370
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

14
CO1sequencesinBOLD,butencounteredmanyinconsistencies;thus,wemanuallycurateda371
subsetofthosesequencestouseforreferencesequences.Thiscuratedreferencedatabaseis372
availableat373
https://github.com/nextgenusfs/amptk/blob/master/docs/reference_databases.md.As374
additionalwell-documentedsequencesareaddedtothedatabase,theabilitytoidentify375
chimericsequenceswillcontinuetoimprove,thusenhancingtheaccuracyofOTUidentification376
inHTSofCO1.377
Withouttheuseofamockcommunity,finalOTUcountsmaybegreatlyinflated378
becauseitisdifficulttoidentifyspuriousOTUs.SpuriousOTUsmayarisefromPCR-or379
sequencing-basedchimeraformationaswellaserrorsgeneratedbyclusteringalgorithms.380
Usingawidelyusedclusteringalgorithm(UPARSE;Edgar2013)andfine-tunedfiltering381
parameters,ourinitialOTUestimateforour46membersinglecopymockcommunitywas70,382
andthusinflatedbyatleast52%bythegenerationofspuriousOTUs.Throughmanual383
inspectionofthesequences,mostofthespuriousOTUsinthemockcommunitywerePCR-384
basedchimerasthatpassedthechimerafilterandwerenotobservedinanyothersample.385
Usingourmockcommunityasareference,wewereabletoassesstheefficacyofanalternative386
OTUpickingalgorithm,DADA2(Callahanetal.2016).UsingtheDADA2algorithmfollowedby387
97%UCLUSTclustering,wewereabletoreducethenumberofOTUsinourpre-PCRcombined388
mockcommunityfrom70to43.Thismethodisstillimperfect,asoneoftheOTUswas389
attributedtosequencingerrorandonewasachimera,thusreducingthefinalnumberto42.390
Twoofourmockmemberswerelostbecausetheydidnotsequencewell,andanadditional391
twowereintra-individualvariantsofothermockmembers(HarmoniaaxyridisandPhalangium392
opilio),andclusteredwiththeir“sibling”sequencesafterUCLUSTwasappliedtotheDADA2393
output.WhenweusedthecuratedreferencedatabaseforchimerafilteringwithUCHIMEin394
combinationwiththeDADA2algorithm,wewereabletoremoveallbutonespuriousOTUfrom395
ourmockcommunity,demonstratingthatclusteringalgorithmscanbefine-tunedtominimize396
spuriousOTUgenerationwiththeuseofsingle-copymockcommunities.397
Estimatesoftaxonomicrichnessmayalsobeinflatedbyintragenomicvariabilityin398
barcodingregions.Intragenomicvariabilityisknowninsomeofthemostcommonlyused399
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

15
barcodingregions,suchasthefungalITSregion(Lindner&Banik2011;Lindneretal.2013;400
Schochetal.2012),aswellasthemitochondrialCO1region(Songetal.2008).Therefore,401
intragenomicvariabilitycouldbeacommonissuewithmanyotherloci.Thepresenceofthis402
individual-levelvariationcanleadtotheinflationoftaxonnumbersbecauseintragenomic403
variantsareoftenmisclassifiedasseparateOTUs(Lindner&Banik2011;Songetal.2008).Two404
conditionsthatcancausethisapparentvariationintheCO1locus,specifically,are405
heteroplasmyandthepresenceofnuclearmitochondrialpseudogenes(numts),whichare406
piecesofmitochondrialDNAthathavebeenincorporatedintothegenome(Songetal.2008).407
Wedetectedintra-individualvariationintheCO1regioninHarmoniaaxyridisandPhalangium408
opilioviastandardcloningandsequencing,eventhoughalimitednumberofcloneswere409
sequenced(i.e.,twosequencevariantsweredetectedbysequencingonlythreeclonesfrom410
eachoftheseindividuals).Basedontheseobservationsfromaverylimitedsamplingof3clones411
perindividual,itseemslikelythatindividualsharbormanysuchvariantsandthatindividual-412
levelvariabilitycouldsignificantlyinflatediversityestimatesinHTSoftheCO1region.TheH.413
axyridisvariantsonlydifferedby2.1%(14of658bp)buttheP.opiliovariantsweremorethan414
threepercentdifferent(3.5%,or23of659bp).Manyofthesedifferencesoccurredinthe415
fragmentamplifiedbytheANMLprimersandthustraditionalclusteringwouldhaveidentified416
themasdistinctOTUs.BothvariantsofH.axyridisandP.opiliowereincludedinourarthropod417
mockcommunitytodetermineifourbioinformaticspipelinewouldbinthesequencevariants418
fromthesameindividualintoseparateinferredsequences.WhenweappliedtheDADA2419
algorithmwithoutclustering,thevariantsseparatedintoseparateOTUs.Afterweapplied97%420
clusteringtotheresultingDADA2inferredsequences,thevariantsweobservedinoursingle421
copymockcommunitybinnedtogether.Theuseofsingle-copyclonedplasmidDNAformock422
communitymembersiscrucialbecauseitremovescrypticsourcesofbiologicalvariationthat423
mightotherwiseoccurwithinthemockcommunity.424
ValidationoftheANMLprimerpairandmockcommunity425
WefurthervalidatedourprimersandHTSsystemusingtwotypesofsampleswith426
knowncomposition:(1)guanosamplesfrombatsfedknowndietsand(2)samplesfrominsect427
trapsthatwereidentifiedbymorphology.Fromtheguanosamples,werecoveredalltaxa428
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

16
includedintheknowndietsandalsorecoveredadditionalOTUs(Table4).TheinitialtotalOTU429
estimateforourfiveknowndietsampleswas42basedonUPARSEclustering,31inferred430
sequencesbasedonDADA2withoutclustering,and10OTUsbasedonDADA2with97%431
clustering.MuchofthetaxonomicreductionintheknowndietsamplesafterusingDADA2with432
clusteringcanbeattributedtosequencevariantsoftwotaxa,Tenebriomolitorand433
Ichneumonidaesp.Thesetwotaxayieldedupto11and7inferredsequencespersamplewith434
theDADA2algorithm,respectively,before97%clusteringwasapplied.However,after435
clusteringwasapplied,theyyieldeduptotwoOTUspersample.Thedegreetowhichthese436
variantsrepresentintra-individualsequencevariation,orvariantsamongindividuals,cannotbe437
determinedhere,butoffersaninterestingtopicforfutureinvestigation.Theestimatewith438
DADA2withclusteringismuchclosertotheexpectedrichnessof5OTUsthanotherestimates.439
SeveralOTUsdetectedfromtheknowndietsampleswereunexpected,butprobablyreal440
componentsofthebatdiet.TwooftheseOTUs,E.takeuchiiandAgrotisipsilon,arelikely441
contaminantsinthedietarycomponentsbecausetheirlarvalformsmayhavebeenmixedinto442
theG.mellonellalarvaethatcomprisedthediet.Wealsodetectedanichneumonidparasitoid443
wasp,whichwasperhapsparasitizingoneormoreoftheinsectsinthediet.Theunexpected444
taxacouldhavebeenanticipatedbysequencingasubsampleoftheknowndietarycomponents445
priortofeeding.446
HTSsuccessfullyrecoveredthemajorityofarthropodspresentinmixedsamplesfrom447
pitfalltraps(Table5).Aftertakingintoaccountprobablemorphologicalidentificationerrors,448
approximately80%ofthetaxaidentifiedbymorphologywerealsoidentifiedviaHTS.Those449
taxamissedbyHTSmayhavebeenmissedduetobiasesinthemolecularpipelinesuchasPCR450
biasesthataroseinthesecomplexcommunities,orperhapsthesetaxarequiremorespecific451
primers.TherewerealsotaxathatweredetectedwithHTSbutmissedbymorphological452
identification.Theseadditionaltaxamayhavebeenmayhavebeenconsumedbyorotherwise453
associatedwiththearthropodscollectedinthetraps,misidentifiedduringthemorphological454
identification,ormaybeDNAcontaminationofthetrapsorothercollectionequipment.455
Conclusion456
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

17
WedemonstratedthattheANMLprimerpairdetectsagreaternumberofarthropod457
taxathanotherfrequentlyusedCO1primerpairs.TheuseofHTSreadnumbersasameasure458
ofabundanceinenvironmentalsamplesisproblematicduetobiasesintroducedduringboth459
PCRandHTS.Thesebiasesmaybepartiallyalleviatedinthefuturebynon-PCRbased460
techniquessuchasshotgunmetagenomicsandtargetcapturetechniques.However,shotgun461
metagenomicsarecurrentlyfarmoreexpensivethanampliconsequencingandmaybecost-462
prohibitivetomostresearchers,andtargetcapturehasnotyetbeenthoroughlyevaluatedfor463
communitycharacterizationofenvironmentalsamples.Failingtouseappropriatepositive464
controlsforamplicon-basedstudiescanleadtoover-estimationofdiversity,andthe465
persistenceof“nonsensetaxa”.Thus,mockcommunitycontrolsarenecessarytoparameterize466
downstreambioinformatics,especiallyfordiversityandcommunitystructurerelatedquestions467
andweadvocatefortheinclusionofaspike-inmockcontrolineveryHTSrun.468
469
Acknowledgements470
FundingforthisworkwasprovidedbytheUSForestService,NorthernResearchStation471
andtheAgriculturalExperimentStationattheUniversityofWisconsin–MadisonviaHatch472
FormulaFunds,andNSFIOS-1121739toAYKandIOS-1121807toJRB.Wesincerelythank473
JamesSkeltonforinsightfuldiscussionthatimprovedpreviousversionsofthismanuscript,J.474
PaulWhiteandHeatherKaarakkaoftheWisconsinDepartmentofNaturalResourcesfor475
assistancewithcoordinatingfieldcollectionofguanosamples,MarceloFerreiradeMeloand476
ErinGreenforassistancewitharthropoddissections,andEliCintoMeijaandMitchLevenhagen477
forinsectcollectionfrompitfalltraps.478
479
References480
BokulichNA,MillsDA(2013)Improvedselectionofinternaltranscribedspacer-specificprimers481
enablesquantitative,ultra-high-throughputprofilingoffungalcommunities.Appliedand482
environmentalmicrobiology79,2519-2526.483
BokulichNA,SubramanianS,FaithJJ,etal.(2013)Quality-filteringvastlyimprovesdiversity484
estimatesfromIlluminaampliconsequencing.Naturemethods10,57-59.485
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

18
BoylesJG,CryanPM,McCrackenGF,KunzTH(2011)Economicimportanceofbatsin486
agriculture.Science332,41-42.487
Brandon-MongG-J,GanH-M,SingK-W,etal.(2015)DNAmetabarcodingofinsectsandallies:488
anevaluationofprimersandpipelines.Bulletinofentomologicalresearch105,717-727.489
BrooksJP,EdwardsDJ,HarwichMD,etal.(2015)Thetruthaboutmetagenomics:quantifying490
andcounteractingbiasin16SrRNAstudies.BMCmicrobiology15,1.491
BusscheRAVD,LeeDN,JudkinsME,etal.(2016)Moleculardietaryanalysisoftheendangered492
ozarkbig-earedbat(Corynorhinustownsendiiingens).ActaChiropterologica18,181-493
191.494
CallahanBJ,McMurdiePJ,RosenMJ,etal.(2016)DADA2:High-resolutionsampleinference495
fromIlluminaamplicondata.Naturemethods13,581-583.496
ClareEL,SymondsonWO,BrodersH,etal.(2014a)ThedietofMyotislucifugusacrossCanada:497
assessingforagingqualityanddietvariability.Molecularecology23,3618-3632.498
ClareEL,SymondsonWO,FentonMB(2014b)Aninordinatefondnessforbeetles?Variationin499
seasonaldietarypreferencesofnight-roostingbigbrownbats(Eptesicusfuscus).500
Molecularecology23,3633-3647.501
ClarkeLJ,SoubrierJ,WeyrichLS,CooperA(2014)Environmentalmetabarcodesforinsects:in502
silicoPCRrevealspotentialfortaxonomicbias.MolecularEcologyResources14,1160-503
1170.504
ClevelandCJ,BetkeM,FedericoP,etal.(2006)Economicvalueofthepestcontrolservice505
providedbyBrazilianfree-tailedbatsinsouth-centralTexas.FrontiersinEcologyandthe506
Environment4,238-243.507
Crisol-MartínezE,Moreno-MoyanoLT,WormingtonKR,BrownPH,StanleyD(2016)Usingnext-508
generationsequencingtocontrastthedietandexplorepest-reductionservicesof509
sympatricbirdspeciesinmacadamiaorchardsinAustralia.PloSone11,e0150159.510
D’AmoreR,IjazUZ,SchirmerM,etal.(2016)Acomprehensivebenchmarkingstudyofprotocols511
andsequencingplatformsfor16SrRNAcommunityprofiling.BMCgenomics17,1.512
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

19
DeagleBE,JarmanSN,CoissacE,PompanonF,TaberletP(2014)DNAmetabarcodingandthe513
cytochromecoxidasesubunitImarker:notaperfectmatch.Biologyletters10,514
20140562.515
DeagleBE,ThomasAC,ShafferAK,TritesAW,JarmanSN(2013)Quantifyingsequence516
proportionsinaDNA-baseddietstudyusingIonTorrentampliconsequencing:which517
countscount?MolecularEcologyResources13,620-633.518
EdgarR(2016)SINTAX:asimplenon-Bayesiantaxonomyclassifierfor16SandITSsequences.519
bioRxiv,074161.https://doi.org/10.1101/074161520
EdgarRC(2010)SearchandclusteringordersofmagnitudefasterthanBLAST.Bioinformatics521
26,2460-2461.522
EdgarRC(2013)UPARSE:highlyaccurateOTUsequencesfrommicrobialampliconreads.523
Naturemethods10,996-998.524
FentonMB(2012)Batsandwhite-nosesyndrome.ProceedingsoftheNationalAcademyof525
Sciences109,6794-6795.526
FolmerO,BlackM,HoehW,LutzR,VrijenhoekR(1994)DNAprimersforamplificationof527
mitochondrialcytochromecoxidaseSubunit1fromdiversemetazoaninvertebrates.528
MolecularMarineBiologyandBiotechnology3,294-299.529
GonsalvesL,BicknellB,LawB,WebbC,MonamyV(2013)Mosquitoconsumptionby530
insectivorousbats:doessizematter?PloSone8,e77183.531
Harms-TuohyCA,SchizasNV,AppeldoornRS(2016)UseofDNAmetabarcodingforstomach532
contentanalysisintheinvasivelionfishPteroisvolitansinPuertoRico.MarineEcology533
ProgressSeries558,181-191.534
HebertPD,CywinskaA,BallSL(2003)BiologicalidentificationsthroughDNAbarcodes.535
ProceedingsoftheRoyalSocietyofLondonB:BiologicalSciences270,313-321.536
HebertPD,PentonEH,BurnsJM,JanzenDH,HallwachsW(2004)Tenspeciesinone:DNA537
barcodingrevealscrypticspeciesintheneotropicalskipperbutterflyAstraptes538
fulgerator.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesof539
America101,14812-14817.540
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

20
JedlickaJA,VoA-TE,AlmeidaRP(2016)Molecularscatologyandhigh-throughputsequencing541
revealpredominatelyherbivorousinsectsinthedietsofadultandnestlingWestern542
Bluebirds(Sialiamexicana)inCaliforniavineyards.TheAuk134,116-127.543
KartzinelTR,PringleRM(2015)Moleculardetectionofinvertebratepreyinvertebratediets:544
trophicecologyofCaribbeanislandlizards.MolecularEcologyResources15,903-914.545
KrehenwinkelH,KennedyS,PekárS,GillespieRG(2016)Acost-efficientandsimpleprotocolto546
enrichpreyDNAfromextractionsofpredatoryarthropodsforlarge-scalegutcontent547
analysisbyIlluminasequencing.MethodsinEcologyandEvolution8,126-134.548
LindahlBD,NilssonRH,TedersooL,etal.(2013)Fungalcommunityanalysisbyhigh-throughput549
sequencingofamplifiedmarkers–auser'sguide.NewPhytologist199,288-299.550
LindnerDL,BanikMT(2009)Effectsofcloningandroot-tipsizeonobservationsoffungalITS551
sequencesfromPiceaglaucaroots.Mycologia101,157-165.552
LindnerDL,BanikMT(2011)IntragenomicvariationintheITSrDNAregionobscures553
phylogeneticrelationshipsandinflatesestimatesofoperationaltaxonomicunitsin554
genusLaetiporus.Mycologia103,731-740.555
LindnerDL,CarlsenT,HenrikNilssonR,etal.(2013)Employing454ampliconpyrosequencingto556
revealintragenomicdivergenceintheinternaltranscribedspacerrDNAregioninfungi.557
Ecologyandevolution3,1751-1764.558
MaineJJ,BoylesJG(2015)Batsinitiatevitalagroecologicalinteractionsincorn.Proceedingsof559
theNationalAcademyofSciences112,12438-12443.560
MallottE,MalhiR,GarberP(2015)High-throughputsequencingoffecalDNAtoidentifyinsects561
consumedbywildWeddell'ssaddlebacktamarins(Saguinusweddelli,Cebidae,562
Primates)inBolivia.Americanjournalofphysicalanthropology156,474-481.563
NguyenNH,SmithD,PeayK,KennedyP(2015)Parsingecologicalsignalfromnoiseinnext564
generationampliconsequencing.NewPhytologist205,1389-1393.565
PiñolJ,MirG,Gomez-PoloP,AgustíN(2015)Universalandblockingprimermismatcheslimit566
theuseofhigh-throughputDNAsequencingforthequantitativemetabarcodingof567
arthropods.MolecularEcologyResources15,819-830.568
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

21
PompanonF,DeagleBE,SymondsonWO,etal.(2012)Whoiseatingwhat:dietassessment569
usingnextgenerationsequencing.Molecularecology21,1931-1950.570
RatnasinghamS,HebertPD(2007)BOLD:TheBarcodeofLifeDataSystem(http://www/.571
barcodinglife.org).Molecularecologynotes7,355-364.572
RognesT,FlouriT,NicholsB,QuinceC,MahéF(2016)VSEARCH:aversatileopensourcetool573
formetagenomics.PeerJ4,e2584.574
RolfeAK,KurtaA,ClemansDL(2014)Species-levelanalysisofdietsoftwomormoopidbats575
fromPuertoRico.JournalofMammalogy95,587-596.576
RydellJ,BogdanowiczW,BoonmanA,etal.(2016)Batsmayeatdiurnalfliesthatrestonwind577
turbines.MammalianBiology-ZeitschriftfürSäugetierkunde81,331-339.578
SchochCL,SeifertKA,HuhndorfS,etal.(2012)Nuclearribosomalinternaltranscribedspacer579
(ITS)regionasauniversalDNAbarcodemarkerforFungi.ProceedingsoftheNational580
AcademyofSciences109,6241-6246.581
SmithMA,WoodleyNE,JanzenDH,HallwachsW,HebertPD(2006)DNAbarcodesreveal582
cryptichost-specificitywithinthepresumedpolyphagousmembersofagenusof583
parasitoidflies(Diptera:Tachinidae).ProceedingsoftheNationalAcademyofSciences584
oftheUnitedStatesofAmerica103,3657-3662.585
SongH,BuhayJE,WhitingMF,CrandallKA(2008)Manyspeciesinone:DNAbarcoding586
overestimatesthenumberofspecieswhennuclearmitochondrialpseudogenesare587
coamplified.ProceedingsoftheNationalAcademyofSciences105,13486-13491.588
TrevellineBK,LattaSC,MarshallLC,NuttleT,PorterBA(2016)Molecularanalysisofnestling589
dietinalong-distanceNeotropicalmigrant,theLouisianaWaterthrush(Parkesia590
motacilla).TheAuk133,415-428.591
VesterinenEJ,RuokolainenL,WahlbergN,etal.(2016)Whatyouneediswhatyoueat?Prey592
selectionbythebatMyotisdaubentonii.Molecularecology25,1581-1594.593
Williams-GuillénK,OlimpiE,MaasB,TaylorPJ,ArlettazR(2016)Batsintheanthropogenic594
matrix:challengesandopportunitiesfortheconservationofChiropteraandtheir595
ecosystemservicesinagriculturallandscapes.In:BatsintheAnthropocene:596
ConservationofBatsinaChangingWorld,pp.151-186.Springer.597
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

22
ZealeMR,ButlinRK,BarkerGL,LeesDC,JonesG(2011)Taxon-specificPCRforDNAbarcoding598
arthropodpreyinbatfaeces.MolecularEcologyResources11,236-244.599
600
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

23
TablesandFigures601
602
Table1:Sequencesandreferences,andprimerpairnamesfortheprimerstestedagainst603
knownarthropodsamples.604
605
Primername Primersequence Reference PairnameLCO1490 5’-GGTCAACAAATCATAAAGATATTGG-3’ Folmeretal.1994 CO1L/HHCO2198 5’-TAAACTTCAGGGTGACCAAAAAATCA-3’ Folmeretal.2003 LEPF1 5’-ATTCAACCAATCATAAAGATATTGG-3’ Hebertetal.2004 LEPmLEPR 5’-CTTGTTCCAGCTCCATTTT-3’ Smithetal.2006 ZBJ-ArtF1c 5’-AGATATTGGAACWTTATATTTTATTTTTGG-3’ Zealeetal.2011 ZBJZBJ-ArtR2c 5’-WACTAATCAATTWCCAAATCCTCC-3’ Zealeetal.2011 LCO1490 5’-GGTCAACAAATCATAAAGATATTGG-3’ Folmeretal.1994 ANMLCO1-CFMRa 5’-GGWACTAATCAATTTCCAAATCC-3’ Thisstudy LCO1490 5’-GGTCAACAAATCATAAAGATATTGG-3’ Folmeretal.1994 CFMRbCO1-CFMRb 5’-GGNACTAATCAATTHCCAAATCC-3’ Thisstudy
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

24
Table2:Resultsfromtestingthe5primerpairslistedinTable1onknowninsectsamples.Shadingandavalue606
of1indicateamplification;noshadingandavalueofzeroindicatenoamplification.Amplificationwas607
attemptedonavarietyofDNAconcentrationsforeachtemplateDNAsamplebeforeassigningavalueofzero608
(noamplification).ArthropodmockcommunitymembersareindicatedwiththesuperscriptIM.Anasterisk609
indicatesthattwodifferentclonedsequencevariantsofanindividualwereaddedtothearthropodmock610
community.**Indicatesthatthreedifferentclonedsequencevariantsofanindividualwereaddedtothe611
arthropodmockcommunity.612
Order Family Identity
ANML
CFMRb
CO1L/H
ZBJZealeetal.2011protocol
ZBJmodifiedprotocol LEP
Blattodea Blattidae PeriplanetafuliginosaIM 1 1 1 1 1 1Blattodea Ectobiidae SupellalongipalpaIM 1 1 1 0 0 1Coleoptera Cantharidae Chauliognathuspennsylvanicus 1 1 1 1 1 1Coleoptera Carabidae Carabidaesp. 0 0 0 0 0 0Coleoptera Cerambycidae Tetraopessp. 1 1 1 1 1 1Coleoptera Chrysomelidae PariafragariaeIM 1 1 1 1 1 1Coleoptera Coccinellidae HarmoniaaxyridisIM** 1 1 1 0 0 0Coleoptera Hydrophilidae Hydrophilidaesp.IM 1 1 1 0 1 1Coleoptera Meloidae Epicautasp. 1 1 1 0 0 1Coleoptera Scarabaeidae EuphoriafulgidaIM 1 1 1 0 0 1Coleoptera Tenebrionidae Tenebriomolitor 1 1 1 1 0 1Coleoptera unk.Coleoptera Polyphagasp.IM 1 1 1 0 0 1Dermaptera Forficulidae Forficulidaesp. 1 1 0 0 0 1Diptera Anthomyiidae DeliaplaturaIM 1 1 1 1 1 1Diptera Bombyliidae LepidophoraluteaIM 1 1 1 1 1 1Diptera Chironomidae Dicrotendipessp.IM 1 1 0 0 0 1Diptera Chironomidae Procladiussp.IM 1 1 1 1 1 1Diptera Culicidae AedesAlbopictusIM 1 1 1 0 0 1Diptera Culicidae AedesvexansIM 1 1 1 1 1 1Diptera Leptoceridae Oecetisinconspicua 1 1 1 0 0 1Diptera Tipulidae NephrotomaferrugineaIM 1 1 1 0 1 1Ephemeroptera Ephemeridae Hexagenialimbata1 1 1 1 0 0 1Ephemeroptera Ephemeridae Hexagenialimbata2IM 1 1 1 0 0 1Ephemeroptera Heptageniidae LeucrocutamaculipennisIM 1 1 1 1 1 1Ephemeroptera unk.Ephemeropotera Ephemeropterasp. 1 1 0 0 0 0Hemiptera Aphididae AphishelianthiIM 1 1 1 0 0 0Hemiptera Cicadellidae Osbornellusauronitens 1 1 1 0 0 0Hemiptera Cicadidae Cicadidaesp. 1 1 1 0 1 1Hemiptera Corixidae Corixidaesp. 1 1 0 0 0 1Hemiptera Corixidae SigaraalternataIM 1 1 1 1 1 1Hemiptera Pentatomidae AcrosternumhilareIM 1 1 1 0 0 0
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

25
Hymenoptera Apidae ApismelliferaIM 1 1 1 0 0 0Hymenoptera Crabonidae Spheciusconvallis 1 1 1 0 0 1Hymenoptera Eucharitidae Eucharitidaesp.IM 1 1 1 1 1 1Hymenoptera Formicidae Formicafusca 1 1 1 0 0 0Hymenoptera Formicidae Formicasp.IM 1 1 1 0 0 0Hymenoptera Tenthredinidae EmpriatakeuchiiIM 1 1 1 1 1 0Lepidoptera Crambidae CrambusagitatellusIM 1 1 1 1 0 1Lepidoptera Crambidae ElophilaobliteralisIM 1 1 1 0 1 1Lepidoptera Crambidae UdearubigalisIM 1 1 1 1 1 1Lepidoptera Depressariidae DepressariapastinacellaIM 1 1 1 1 1 1Lepidoptera Erebidae HypenascabraIM 1 1 1 1 1 1Lepidoptera Erebidae HyphantriacuneaIM 1 1 1 0 0 1Lepidoptera Erebidae IdiaaemulaIM 1 1 1 1 1 1Lepidoptera Erebidae ReniafactiosalisIM 1 1 1 1 1 1Lepidoptera Geometridae HaematopisgratariaIM 1 1 1 1 1 1Lepidoptera Noctuidae AgrotisipsilonIM 1 1 1 1 1 1Lepidoptera Tortricidae ChoristoneurarosaceanaIM 1 1 1 1 1 1Neuroptera Chrysopidae ChrysopaoculataIM 1 1 1 0 0 1Neuroptera Mantispidae Mantispidaesp.IM 1 1 1 0 1 1Opiliones Phalangiidae PhalangiumopilioIM* 1 1 1 0 0 1Orthoptera Acrididae MelanoplusfemurrubrumIM 1 1 1 0 0 1Orthoptera Tettigoniidae ScudderiacurvicaudaIM 1 1 1 0 0 1Orthoptera Tettigoniidae Tettigoniidaesp.IM 1 1 1 1 1 1Trichoptera Hydropsychidae Potamyiaflava 1 1 0 0 0 1Trichoptera Hydroptilidae Orthotrichiasp.IM 1 1 1 0 0 1Trichoptera Leptoceridae CeracleamaculataIM 1 1 1 1 1 1Trichoptera Leptoceridae LeptocerusamericanusIM 1 1 1 0 0 1Trichoptera unk.Trichoptera Trichopterasp. 1 1 0 0 0 0
Negativecontrol 0 0 0 0 0 0
Total 58 58 52 24 27 48
%Total 98.31 98.31 88.14 40.68 45.76 81.36
613
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017
26
Table3:OperationalTaxonomicUnits(OTUs)recoveredusinghigh-throughputampliconsequencing(HTS)and614
eithertheANMLprimersortheZBJprimerson3field-collectedguanosamples.Numbers(0-3)and615
representativeshadingindicatethenumberofguanosampleseachOTUwasdetectedinforeachprimerpair.616
617
ANML ZBJ Phylum Class Order Family Genus species1 0 Arthropoda Arachnida Araneae Theridiidae Theridion Theridionfrondeum2 0 Arthropoda Arachnida Trombidiformes Limnesiidae Limnesia Limnesiasp.1 0 Arthropoda Arachnida Trombidiformes
Trombidiformessp.
1 0 Arthropoda Arachnida
Arachnidasp.1 0 Arthropoda Insecta Coleoptera Coccinellidae Harmonia Harmoniasp.2 0 Arthropoda Insecta Coleoptera Elateridae Melanotus Melanotussimilis1 0 Arthropoda Insecta Coleoptera Hydrophilidae Helocombus Helocombusbifidus1 0 Arthropoda Insecta Coleoptera Scarabaeidae
Scarabaeidaesp.
1 0 Arthropoda Insecta Coleoptera Tenebrionidae Tenebrio Tenebriosp.1 0 Arthropoda Insecta Coleoptera
Coleopterasp.
1 0 Arthropoda Insecta Diptera Bibionidae Bibio Bibiosp.1 0 Arthropoda Insecta Diptera Ceratopogonidae Bezzia Bezziasp.0 2 Arthropoda Insecta Diptera Chaoboridae Chaoborus Chaoboruspunctipennis1 1 Arthropoda Insecta Diptera Chironomidae Ablabesmyia Ablabesmyiaamericana1 0 Arthropoda Insecta Diptera Chironomidae Ablabesmyia Ablabesmyiaannulata1 0 Arthropoda Insecta Diptera Chironomidae Ablabesmyia Ablabesmyiasp.11 0 Arthropoda Insecta Diptera Chironomidae Ablabesmyia Ablabesmyiasp.23 0 Arthropoda Insecta Diptera Chironomidae Chironomus Chironomusplumosus1 1 Arthropoda Insecta Diptera Chironomidae Chironomus Chironomussp.10 2 Arthropoda Insecta Diptera Chironomidae Chironomus Chironomussp.21 0 Arthropoda Insecta Diptera Chironomidae Coelotanypus Coelotanypussp.1 0 Arthropoda Insecta Diptera Chironomidae Conchapelopia Conchapelopiasp.1 0 Arthropoda Insecta Diptera Chironomidae Cryptochironomus Cryptochironomussp.12 0 Arthropoda Insecta Diptera Chironomidae Cryptochironomus Cryptochironomussp.22 0 Arthropoda Insecta Diptera Chironomidae Dicrotendipes Dicrotendipestritomus2 1 Arthropoda Insecta Diptera Chironomidae Parachironomus Parachironomussp.11 0 Arthropoda Insecta Diptera Chironomidae Parachironomus Parachironomussp.21 0 Arthropoda Insecta Diptera Chironomidae Parachironomus Parachironomussp.32 0 Arthropoda Insecta Diptera Chironomidae Polypedilum Polypedilumsp.11 0 Arthropoda Insecta Diptera Chironomidae Polypedilum Polypedilumsp.22 2 Arthropoda Insecta Diptera Chironomidae Procladius Procladiussp.11 0 Arthropoda Insecta Diptera Chironomidae Procladius Procladiussp.21 0 Arthropoda Insecta Diptera Chironomidae Procladius Procladiussp.30 1 Arthropoda Insecta Diptera Chironomidae Procladius Procladiussp.41 0 Arthropoda Insecta Diptera Chironomidae Xenochironomus Xenochironomussp.3 0 Arthropoda Insecta Diptera Chironomidae
Chironomidaesp.1
1 0 Arthropoda Insecta Diptera Chironomidae
Chironomidaesp.22 0 Arthropoda Insecta Diptera Chironomidae
Chironomidaesp.3
2 0 Arthropoda Insecta Diptera Chironomidae
Chironomidaesp.4
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

27
0 1 Arthropoda Insecta Diptera Chironomidae
Chironomidaesp.51 0 Arthropoda Insecta Diptera Culicidae Aedes Aedesabserratus1 0 Arthropoda Insecta Diptera Culicidae Aedes Aedesexcrucians1 0 Arthropoda Insecta Diptera Culicidae Aedes Aedesprovocans1 1 Arthropoda Insecta Diptera Culicidae Aedes Aedesvexans1 0 Arthropoda Insecta Diptera Culicidae Culiseta Culisetamelanura1 0 Arthropoda Insecta Diptera Hybotidae Platypalpus Platypalpussp.0 1 Arthropoda Insecta Diptera Limoniidae Shannonomyia Shannonomyialenta1 0 Arthropoda Insecta Diptera Psychodidae Psychoda Psychodaalternata1 1 Arthropoda Insecta Diptera Tachinidae
Tachinidaesp.
1 0 Arthropoda Insecta Diptera Tipulidae Nephrotoma Nephrotomaferruginea0 1 Arthropoda Insecta Diptera Tipulidae Tipula Tipulakennicotti1 1 Arthropoda Insecta Ephemeroptera Caenidae Caenis Caenisyoungi1 0 Arthropoda Insecta Ephemeroptera Palingeniidae Pentagenia Pentageniavittigera1 0 Arthropoda Insecta Ephemeroptera Siphlonuridae Siphlonurus Siphlonurustypicus1 0 Arthropoda Insecta Hemiptera Corixidae Trichocorixa Trichocorixaborealis1 0 Arthropoda Insecta Hemiptera Miridae Lygus Lyguslineolaris0 1 Arthropoda Insecta Lepidoptera Blastobasidae Blastobasis Blastobasisglandulella1 0 Arthropoda Insecta Lepidoptera Depressariidae Antaeotricha Antaeotrichaleucillana
1 0 Arthropoda Insecta Lepidoptera Tortricidae ArgyrotaeniaArgyrotaeniapinatubana
1 0 Arthropoda Insecta Lepidoptera
Lepidopterasp.1 0 Arthropoda Insecta Megaloptera Corydalidae Chauliodes Chauliodesrastricornis1 0 Arthropoda Insecta Trichoptera Hydroptilidae Oxyethira Oxyethiraserrata0 2 Arthropoda Insecta
Insectasp.
3 0 Chordata Mammalia Chiroptera Vespertilionidae Myotis Myotislucifugus 618
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

28
Table4:NumberofOTUsidentifiedfrombatsfedknowninsectdiets,brokendownbyexpecteddietarycomponents,possible619
accidentaldietarycomponents,andbatDNA.Blanksarezeros.DADA2isdatafromDADA2,withoutclustering.DADA297%clusteris620
datafromDADA2with97%clusteringappliedtotheOTUtable.Shadedcellsaredietarycomponentsthatwereexpected(i.e.known621
tobefedtothebat).EPFU1,EPFU2,andEPFU3arefromBigBrownBats(Eptesicusfuscus),andLACI1andLACI2arefromHoaryBats622
(Lasiuruscinereus).623
624
Expecteddietarycomponents Possibleaccidentaldietarycomponents BatDNA
Galleriamellonella
Tenebriomolitor
Antheraeapolyphemus
Empriatakeuchii
Agrotisipsilon Ichneumonidaesp.
Eptesicusfuscus
Lasiuruscinereus
DADA2 DADA297%
cluster
DADA2 DADA297%
cluster
DADA2 DADA297%
cluster
DADA2 DADA297%
cluster
DADA2 DADA297%
cluster
DADA2 DADA297%
cluster
DADA2
DADA297%
cluster
DADA2 DADA297%
clusterEPFU1 1 1 8 1 1 1 1 1 1 1 EPFU2 1 1 1 1 1 1 1 1 1 1 7 1 1 1 EPFU3 1 1 1 1 LACI1 1 1 7 1 3 2LACI2 11 2 3 2
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

29
Table5:ComparisonofmorphologicalandHTSfamily-levelidentificationsofarthropods625
collectedfrom5pitfalltraps.Arthropodsfromtraps1and4weredissectedpre-extraction,and626
arthropodsfromtraps2,3,and5weremaceratedpre-extraction.A“+”indicatespresenceofa627
family.628
629
Trap1 Trap2 Trap3 Trap4 Trap5
Key NGS Key NGS Key NGS Key NGS Key NGS
TotalTaxa 7 10 17 13 17 19 4 5 10 7
Class Order/subclass Family
Insecta Blattodea Ectobiidae + Insecta Coleoptera Carabidae + + Insecta Coleoptera Elateridae + Insecta Coleoptera Melyridae + + + +
InsectaColeoptera
Ptinidae/
+
Anobiidae
Insecta Coleoptera Scarabaeidae + Insecta Coleoptera Silphidae + + Insecta Coleoptera Tenebrionidae + + Entognatha Collembola + + Insecta Diptera Anthomyiidae + + + + Insecta Diptera Bombyliidae + +
Insecta Diptera Calliphoridae + + + Insecta Diptera Cecidomyiidae + Insecta Diptera Culicidae + Insecta Diptera Dipterasp. + Insecta Diptera Heleomyzidae + Insecta Diptera Phoridae + + + Insecta Diptera Scathophagidae + + Insecta Diptera Sciaridae + Insecta Diptera Syrphidae + +
Insecta Hemiptera Aphididae + + + +
Insecta Hemiptera Cicadidae + + Insecta Hemiptera Cicadellidae + + + + + Insecta Hemiptera Geocoridae + Insecta Hemiptera Miridae + + Insecta Hemiptera Pentatomidae + + Insecta Hemiptera Psyllidae + + Insecta Hymenoptera Formicidae + + + + + + + +
Insecta Hymenoptera Braconidae +
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

30
Insecta Hymenoptera Ceraphronidae + Insecta Hymenoptera Chalcidoidea + Insecta Hymenoptera Crabronidae + Insecta Hymenoptera Dryinidae + Insecta Hymenoptera Halictidae + +
Insecta Hymenoptera Hymenopterasp. + Insecta Hymenoptera Ichneumonidae + + + Insecta Hymenoptera Pompilidae + + Insecta Lepidoptera Gelechiidae + + Insecta Lepidoptera Tortricidae + +
Insecta Neuroptera Chrysopidae + Insecta Orthoptera Acrididae + Insecta Orthoptera Tettigoniidae + Insecta Thysanoptera Thripidae + Insecta Thysanoptera Hydroptillidae + Arachnida Acari + + + + + + + + Arachnida Araneae Araneaesp. + Arachnida Araneae Gnaphosidae + + + + Arachnida Araneae Lycosidae + + Arachnida Araneae Pisauridae + Arachnida Araneae Salticidae + + + + Arachnida Araneae Thomisidae + Chilopoda + +
630
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

31
Figure1:Heatmapofthehigh-throughputampliconsequencingreadnumbersofthe631
arthropodmockcommunity,equilibratedandcombinedbothpre-andpost-PCR.Thepost-PCR632
combinedmockcommunitywasfarmoreevenandrepresentativeoftheequalamountsof633
DNAaddedforeachmockmemberthanthepre-PCRcombinedmockcommunity.634
635
636
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017

32
DataAccessibility637
ThecorrespondingdataforthispaperwasdepositedintheNCBISRA(SRAstudySRP102878;638
BioProjectPRJNA380665),andbarcodedprimerinformationisprovidedinthesupplemental639
information.640
641
AuthorContributions642
MAJ,MTB,andJMPwrotethepaper;MAJ,MTB,JMP,andDLLdesignedresearch;MAJ,MTB,643
JMP,AKW,andEPperformedresearch;AYK,LX,JRB,CG,andMZPcontributedsamples;MAJ,644
MTB,JMPanalyzedthedata;andMAJ,MTB,JMP,AKW,JRB,AYK,CG,MZP,andDLLedited645
draftsofthepaper.646
647
SupportingInformation648
AppendixS1–DNAextractiondetailsfortheCLSextractionforarthropods649
AppendixS2–DNAextractiondetailsforthemodifiedbatguanoextraction650
TableS1–SequencesforIonbarcodedprimers651
FigureS1–MitochondrialcytochromeoxidaseCsubunit1locus(CO1)primermap652
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.3184v1 | CC BY 4.0 Open Access | rec: 24 Aug 2017, publ: 24 Aug 2017