amidoximereductasesystemcontainingcytochrome b5 typeb ... ·...
TRANSCRIPT

Amidoxime Reductase System Containing Cytochrome b5Type B (CYB5B) and MOSC2 Is of Importance for LipidSynthesis in Adipocyte Mitochondria*□S
Received for publication, December 7, 2011, and in revised form, December 21, 2011 Published, JBC Papers in Press, December 27, 2011, DOI 10.1074/jbc.M111.328237
Etienne P. A. Neve‡1, Åsa Nordling‡, Tommy B. Andersson‡§, Ulf Hellman¶, Ulf Diczfalusy�, Inger Johansson‡2,and Magnus Ingelman-Sundberg‡2
From the ‡Section of Pharmacogenetics, Department of Physiology and Pharmacology, Karolinska Institutet, SE-17177 Stockholm,§AstraZeneca Research and Development, Pepparedsleden 1, SE-431 83 Mölndal, the ¶Ludwig Institute for Cancer Research, Ltd.,Uppsala Branch, SE-75123 Uppsala, and the �Department of Laboratory Medicine, Division of Clinical Chemistry, KarolinskaInstitutet, Karolinska University Hospital Huddinge, SE-141 86 Huddinge, Sweden
Background:High amidoxime reductase activity is found in liver and fat tissue, although its composition in cells is unknown.Results: Amidoxime reductase is up-regulated during adipogenesis and requires MOSC2 and CYB5B.Conclusion:MOSC2 and CYB5B are essential components of the mitochondrial amidoxime reductase, and MOSC2 isimportant for lipogenesis.Significance:We suggest a role of MOSC2 in adipogenesis and fatty acid synthesis and as a potential novel drug target.
Reduction of hydroxylamines and amidoximes is importantfor drug activation and detoxification of aromatic and heterocy-clic amines. Such a reductase systemwas previously found to beof high activity in adipose tissue and liver, and furthermore, invitro studies using recombinant truncated and purifiedenzymes suggested theparticipationof cytochromeb5 reductase(CYB5R), cytochromeb5 (CYB5), andmolybdenumcofactor sul-furase C-terminal containing 1 and 2 (MOSC1 and -2). Here, weshow that purified rat liver outermitochondrialmembrane con-tains high amidoxime reductase activity and that MOSC2 isexclusively localized to these membranes. Moreover, using thesame membrane fraction, we could show direct binding of aradiolabeled benzamidoxime substrate to MOSC2. Followingdifferentiation of murine 3T3-L1 cells into mature adipocytes,the MOSC2 levels as well as the amidoxime reductase activitywere increased, indicating that the enzyme is highly regulatedunder lipogenic conditions. siRNA-mediated down-regulationofMOSC2 and themitochondrial form of cytochrome b5 type B(CYB5B) significantly inhibited the reductase activity in the dif-ferentiated adipocytes, whereas down-regulation of MOSC1,cytochrome b5 type A (CYB5A), CYB5R1, CYB5R2, or CYB5R3had no effect. Down-regulation of MOSC2 caused impairedlipid synthesis. These results demonstrate for the first time thedirect involvement of MOSC2 and CYB5B in the amidoximereductase activity in an intact cell system.Wepostulate thepres-ence of a novel reductive enzyme system of importance for lipidsynthesis that is exclusively localized to the outermitochondrialmembrane and is composed of CYB5B, MOSC2, and a thirdunknown component (a CYB5B reductase).
An increasing number of drugs and drug candidates containbasic functional groups such as amidines and guanidines thatdecrease their oral bioavailability by preventing efficient gas-trointestinal adsorption as these groups are protonated underphysiological pH. Instead, prodrugs have been developed byN-hydroxylation of these amidine groups, converting them intoamidoximes to improve their oral bioavailability as exemplifiedby the direct thrombin inhibitor ximelagatran (1, 2), its fol-low-up compound AZD0837 (3), and a platelet inhibitorsibrafidan (4). Once these prodrugs are absorbed into the body,they are reduced back to their bioactive amidines by a reductiveenzyme system believed to be composed of cytochrome b5(CYB5),3 cytochrome b5 reductase (CYB5R), and a thirdunknown component and is associated with both the micro-somal fraction (5) as well as the mitochondrial fraction (6, 7).The reductivemetabolism of nitrogen-containing groups is notonly of importance for the activation of prodrugs but also forreduction of hydroxylamines and for detoxification of carcino-genic N-hydroxylated aromatic and heterocyclic amines (8, 9).The endogenous role of this activity, however, is still unknown.In a previous study we found high amidoxime reductase
activity preferentially in outer mitochondrial membranes(OMM) associated with adipose tissue, liver, and kidney (6).Furthermore, the reductase activity was dependent on thecofactor NADH and inhibited by potassium cyanide (KCN),whereas typical CYP inhibitors, such as carbonmonoxide, wereineffective (6) indicating that the reductase is unlikely to be acytochrome P450 as was suggested previously (10). Severalstudies have attempted to identify the third component, and theinvolvement of stearoyl-CoA desaturase (11) and cytochrome
* This work was supported by grants from AstraZeneca and from the SwedishResearch Council.
□S This article contains supplemental “Experimental Procedures,” Figs. S1–S7,and Table S1.
1 To whom correspondence should be addressed. Tel.: 46-8-52487711; Fax:46-8-337327; E-mail: [email protected].
2 Both authors contributed equally to this work.
3 The abbreviations used are: CYB5, cytochrome b5; CYB5A, cytochrome b5
type A; CYB5B, cytochrome b5 type B; MOSC1, molybdenum cofactorsulfurase C terminus containing 1; MOSC2, molybdenum cofactor sul-furase C terminus containing 2; CYB5R, cytochrome b5 reductase; OMM,outer mitochondrial membrane; PTU, 6-propyl-2-thiouracil; Tricine,N-[2-hydroxy-1,1-bis(hydroxymethyl)ethyl]glycine; IMM, inner mito-chondrial membrane.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 9, pp. 6307–6317, February 24, 2012© 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6307
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

P450 isoenzyme CYP2D (10) in themicrosomal fraction as wellas a novel molybdenum-containing enzyme in the mitochon-drial fraction (12) was suggested. In subsequent studies, twohighly homologous molybdenum-containing proteins, namelymolybdenum cofactor sulfurase C-terminal containing 1 and 2(MOSC1 and MOSC2, respectively, referred to as mARC1 andmARC2), were shown to display amidoxime reductase activityin vitro when incubated in a reconstituted system with CYB5and CYB5R3 (cytochrome b5 reductase 3) (13, 14). In addition,Kurian et al. (15) showed that in a reconstituted systempurifiedsoluble CYB5 and purified soluble CYB5R were able to reduceamidoximes to amidines without the involvement of a thirdcomponent. However, all studies mentioned above were per-formed using reconstituted systems consisting of purified andsoluble truncated components, without showing the directinvolvement of these components in more integrated physio-logical systems.In this study, we demonstrate using a cell model commonly
used to study adipogenesis and siRNAknockdown experimentsthat MOSC2, but not MOSC1, is critically involved in the ami-doxime reductase activity in differentiated adipocytes. In addi-tion, the mitochondrial form of cytochrome b5 type B (CYB5B)is also shown to be essential for the reductase activity in thisintact cell system, which appears to be of importance for lipidformation. In contrast, the microsomal form of cytochrome b5type A (CYB5A) and CYB5R3 as well as its homologs cyto-chrome b5 reductase 1 and 2 (CYB5R1 and CYB5R2) are notinvolved in the reductase activity in the adipocyte cell system.
EXPERIMENTAL PROCEDURES
Reagents—Dulbecco’s modified Eagle’s medium (DMEM),fetal bovine serum (FBS), sodiumpyruvate, penicillin and strep-tomycin, insulin, and OptiMEM were obtained from Invitro-gen. Bovine calf serum was from American Type Culture Collec-tion (LGC Standards AB, Borås, Sweden). Cell culture flasks andplates were from Corning Life Science and Sarstedt (VWR Inter-national AB, Stockholm, Sweden). DharmaFECT Duo and ON-TARGETplus SMARTpool siRNA constructs (mouse MOSC1L-044395-01, mouse MOSC2 L-044395-01, mouse CYB5R3 L-058184-01, mouse CYB5R1 L-046143-01, mouse CYB5R2L-056941-01,mouseCYB5L-050436-01 (CYB5A),mouseCYB5BL-059461-01, and Nontargeting Pool D-001810-10) were pur-chased fromThermo Scientific (VWR International AB). Dexam-ethasone, 3-isobutylmethylxanthine, benzamidoxime, and 6-pro-pyl-2-thiouracil (PTU) were from Sigma. Complete proteaseinhibitor mixture was from Roche Applied Science. Rabbit anti-MOSC2 and anti-CYB5B antibodies were from Atlas Antibodies(StockholmSweden);mouse anti-mHSP70and rabbit anti-VDACwere from Affinity Bioreagents (Thermo Scientific); and rabbitanti-CYB5 was from Santa Cruz Biotechnology (AH DiagnosticsAB, Skärholmen, Sweden). Rabbit anti-CYB5R3 (16), rabbit anti-ERp27 (17), and rabbit anti-calnexin (18) were previouslydescribed. Secondary horseradish peroxidase-coupled anti-rabbit and anti-mouse antibodies were fromDako (Dako SwedenAB, Stockholm, Sweden). AR-H069637, AR-H069927, andAZ13228184-14Cwere fromAstraZenecaR&DMölndal, Sweden.
Purification of Outer Mitochondrial Membranes from RatLiver—OMM were purified as described by Hovius et al. (19)and deKroon et al. (20). PurifiedOMMvesicles were aliquoted,snap-frozen in liquid nitrogen, and stored at �70 °C. Also col-lected during fractionation were the homogenate (600 � gsupernatant), P10 (10,000� g pellet, containingmitochondria),the inner mitochondrial membrane (IMM)/mitochondrialmatrix fraction, and themicrosomes (100,000� g supernatant).Protein Concentration Determination—Protein concentra-
tions were determined according to the method of Lowry et al.(21) using bovine serum albumin as standard.Western Blot Analysis—Subcellular fractions isolated from
rat liver and cell lysates from adipocytes and preadipocyteswere subjected toWestern blot analysis as described previously(22).AmidoximeReductionAssay—Amidoxime reductase activity
was determined following the benzamidoxime reduction tobenzamidine as described previously (6). The amidoximereductase activity was also monitored by the reduction of theintermediatemetabolite of AZD0837, theN-hydroxylated ami-dine AR-H069927 to the corresponding amidine AR-H067637(supplemental Fig. S1) (23). Incubations were composed of 0.25�g of OMM and 20 �M AR-H069927 in 50 mM phosphatebuffer, pH 6.3, in a volume of 100 �l. After a 2-min preincuba-tion at 37 °C, the reaction was started by the addition of 250 �M
NADH, and samples were incubated for 20 min at 37 °C. Reac-tions were terminated by the addition of 10 �l of 6 M formicacid, and precipitated proteins were settled by centrifugation17,000 � g at 4 °C for 15 min. The supernatant was mixed withan equal volume of acetonitrile containing 1% (v/v) acetic acid.The formation of AR-H067637 was monitored by HPLC anal-ysis using aVarian ProStarmodel 410 autosampler, Varian Pro-Star 310UV-visible detector, andVarian ProStarmodel 240 sol-vent delivery module (Agilent Technologies Sweden AB, Kista,Sweden). Samples were separated on a LiChrospher� 60 RP-select B (5�m) column (Merck) using an isocraticmobile phasecomposed of 0.1% (v/v) acetic acid and 3.8mM ammonium ace-tate containing 18% acetonitrile. Metabolite and parent com-pound were detected at 229 nm and quantified using purifiedstandards.Cross-linking Studies—Purified OMM (0.5 mg) was incu-
bated with a radiolabeled (carbon-14) and cross-linkable(azide) benzamidoxime analog (AZ13228184-14C, supplemen-tal Fig. S2) to identify putative components of the amidoximereductase complex. OMM were diluted in phosphate-bufferedsaline (PBS) to a concentration of 1 mg/ml and incubated with25�M (1.42�Ci/ml or 52.5 kBq/ml) [14C]benzamidoxime azidein the presence and absence of 250�MNADH for 2min at 37 °Cin the dark, and sampleswere cross-linked on ice by exposure toUV light for 2.5 min. The cross-linked samples were subjectedto sequential detergent extraction using Triton X-100 followedby Zwittergent 3-14 (Calbiochem and VWR International AB)as described in the supplemental Experimental Procedures andsupplemental Fig. S3. The detergent-extracted samples werediluted in Laemmli sample buffer, boiled. and separated bySDS-PAGE using a 10% Tris-Tricine gel. The gel was stainedwith Coomassie Blue R-250 and dried, and protein-[14C]benz-
Mitochondrial Amidoxime Reductase Enzyme System
6308 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 9 • FEBRUARY 24, 2012
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

amidoxime complexes were visualized by autoradiography(Fuji-BAS 1800, FujiFilm, Stockholm, Sweden).Analysis of Proteins by Mass Spectrometry—Coomassie-
stained protein bands with an apparent molecular mass of30–40 kDa,whichwere identified by cross-linking to the radio-labeled benzamidoxime analog, were excised from the geland processed for mass spectrometer analysis essentially asdescribed previously (24). In brief, the gel pieces were destainedand dried, and trypsin (porcine, modified, sequence grade fromPromega Biotech AB, Nacka, Sweden) was allowed to soak intothe swelling gel pieces on ice. After overnight incubation at30 °C and acidification, the proteolytic peptides were subjectedto mass analysis by matrix-assisted laser desorption/ioniza-tion time of flight mass spectrometry on a Bruker Ultraflex IIITOF/TOF instrument from Bruker (Stockholm, Sweden),applying the manufacturer’s recommendations. �-Cyano-4-hydroxycinnamic acid was used asmatrix, and the spectra wereexternally calibrated using a 7-peptide mixture. The generatedpeptide mass lists were used to scan the current NCBInrsequence data base for protein identities, employing the searchengine ProFound.Immunoprecipitation Analysis—[14C]Benzamidoxime azide
was incubated and cross-linked toOMM (700�g) as described,and cross-linked samples were solubilized in 1.2% digitonin(Sigma) on ice for 30 min, conditions known to solubilize theMOSC2 protein from the OMM (data not shown). Nonsolubi-lized material was settled by centrifugation for 15 min at17,000� g, and supernatant was transferred to fresh Eppendorftubes. The precleared supernatant was subjected to immuno-precipitation using anti-MOSC2 antibodies (0.5 �g) overnightat 4 °C. The next day protein complexes were captured by incu-bation with 25 �l of protein A/G gel (Thermo Scientific) for1.5 h at 4 °C and washed two times in PBS containing 1.2%digitonin and two times in PBS. The collected protein com-plexes were eluted by boiling in Laemmli sample buffer, sepa-rated by SDS-PAGE, and analyzed by Western blotting forMOSC2 and subsequent autoradiography of the same nitrocel-lulose membrane to detect radiolabeled MOSC2.Culture andDifferentiation of 3T3-L1Cells—Murine 3T3-L1
preadipocytes were obtained from American Type CultureCollection and cultured in DMEM with high glucose contain-ing 10% bovine calf serum, 1% sodium pyruvate, and 100 IU/mlpenicillin, and 100 �g/ml streptomycin and subcultured beforethey reached 80% confluency. For differentiation, preadi-pocytes were cultured in completemedium supplemented with10% FCS, and fully differentiated adipocytes were obtained bytreating 3-day confluent cells with a standard MDI mixture of3-isobutylmethylxanthine (0.5 mM), dexamethasone (0.5 �M),and insulin (1.7 �M) for 2 days, followed by treatment withcompletemediumcontaining only insulin for another 5–7 days.Medium was changed every 2 days. Cells were cultured anddifferentiated in 6-well cluster plates or 10-cm dishes for prep-aration of subcellular fractions.Subcellular Fractionation of 3T3-L1 Cells—Preadipocytes or
differentiated adipocytes were washed twice with PBS, scrapedinto 1 ml of PBS, and centrifuged for 5 min at 750 � g and 4 °C.The cell pellet was resuspended in buffer A (10 mM Tris-HCl,pH 7.4, containing 20% (w/v) glycerol, 1 mM EDTA, and prote-
ase inhibitor mixture) and sonicated on ice with 20 times 1-sbursts. Cell lysates were centrifuged at 800 � g and 4 °C for 10min, and supernatantswere transferred and further centrifugedat 6,500� g and 4 °C for 15min to obtainmitochondrial pellets,which were resuspended in mitochondrial buffer (10 mM Trisacetate, pH 7.4, 0.25 M sucrose, 0.5 mM EDTA) and the 6,500 �g supernatant containing the microsomes and cytosol.siRNA Transfection of 3T3-L1 Adipocytes—Transfection of
differentiated 3T3-L1 adipocytes with siRNAwas adapted fromKilroy et al. (25). Three-day confluent 3T3-L1 preadipocyteswere differentiated using the standardMDI protocol for 5 days,after which they were trypsinized and resuspended in DMEMwith high glucose containing 10% FCS and insulin at 6 105 cells/ml. Transfections were performed in 6-well cluster plates, andthe transfectionmix for onewell (9.6 cm2)was composed of 200pmol of siRNA and 14 �l of DharmaFect Duo diluted in 400 �lofOptiMEM towhich 12 105 cells (2ml) were added. Cells wereanalyzed for reductase activity and protein expression 5 dayspost-transfection (10 days post-differentiation).Real Time PCR—The mRNA expression levels of several
genes in siRNA-treated or control adipocytes was determinedby ready to use TaqMan gene expression assays according tothe manufacturer’s instructions (Applied Biosystems, Stock-holm, Sweden) using �-actin as a housekeeping gene.Cytochrome b5 Reductase Assay—The activity of NADH
CYB5R was determined by monitoring the reduction of theartificial electron acceptor 2,6-dichlorophenolindophenol at590 nm. The incubation mixture contained 1 mM NADH, 3mg/ml 2,6-dichlorophenolindophenol, and 15 �g of rat livermicrosomes in a 100mM phosphate buffer, pH 7.4. The CYB5Ractivity was determined without and in the presence of theCYB5R inhibitor PTU.Oil Red O Staining—Intracellular lipids were measured
through staining with Oil Red O (Sigma). Cells were fixed with10% formalin for 10 min and incubated in fresh formalin for atleast 1 h. After washing twice with double distilled H2O and60% isopropyl alcohol, cells were stained for 10 min in freshlydiluted Oil Red O solution (5.2 mM). Oil Red O was eluted byadding isopropyl alcohol for 10 min, and the extracted dye wasmeasured at 510 nm on a Varian spectrophotometer (AgilentTechnologies).Fatty AcidDetermination—Cell homogenates were prepared
frommature adipocytes collected 5 days after transfection withcontrol or MOSC2 siRNA. To homogenates corresponding to0.36 mg of cell protein, nonadecanoic acid (C19:0, 20 mg) wasadded as an internal standard. After Folch extraction, theextracts (500 ml) were transmethylated by addition of 0.5 ml of0.5 M sodium methoxide in methanol. Following incubation at56 °C for 15min, 1 ml of 1 MHCl/methanol was added to ester-ify free fatty acids. After incubation at 56 °C for 15 min, 2 ml ofwater was added, and the samples were extracted three timeswith 3 ml of hexane. The samples were dried under a gentlestream of argon and redissolved in 500 ml of hexane. The rela-tive contents of fatty acids were determined on a Hewlett Pack-ardHP 6890 gas chromatograph connected to anHP 5977massselective detector. The gas chromatographwas equippedwith aHP-5MS column (30 m � 0.25 mm, 0.25-mm film thickness).Ratios of fatty acids (C14:0, C16:0, C16:1, C18:0, and C18:1) to
Mitochondrial Amidoxime Reductase Enzyme System
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6309
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from
the internal standard (C19:0) were calculated based on peakarea. Authentic fatty acids were used to establish retentiontimes, and mass spectra were recorded for the different fattyacids to ensure identity.
RESULTS
Enzyme Kinetics of the Amidoxime Reductase—The enzymekinetics of the amidoxime reductase enzyme systemwere char-acterized using subcellular fractions isolated from rat andhuman liver in the absence and presence of the inhibitor potas-sium cyanide (KCN) (6). The reductase activity observed in thepresence of KCN is considered to be the nonspecific reductaseactivity, especially at higher benzamidoxime concentrations.The kinetics of benzamidine formation by rat liver microsomesandmitochondria as well as human livermicrosomes displayedMichaelis-Menten kinetics (supplemental Fig. S4), and theapparent Km and Vmax values are listed in Table 1. The KCN-inhibitable Km values for the rat liver microsomes and mito-chondria are very similar, 24.4 and 24.6 �M, respectively,although the observedKm value in humanmicrosomes is muchhigher, namely about 430 �M. The highest Vmax value wasdetected in rat liver mitochondria 3.2 nmol/min/mg protein,being 2–3-fold higher than those observed in rat liver micro-somes and human liver microsomes.MOSC2 Is Localized in the Outer Mitochondrial Membrane—
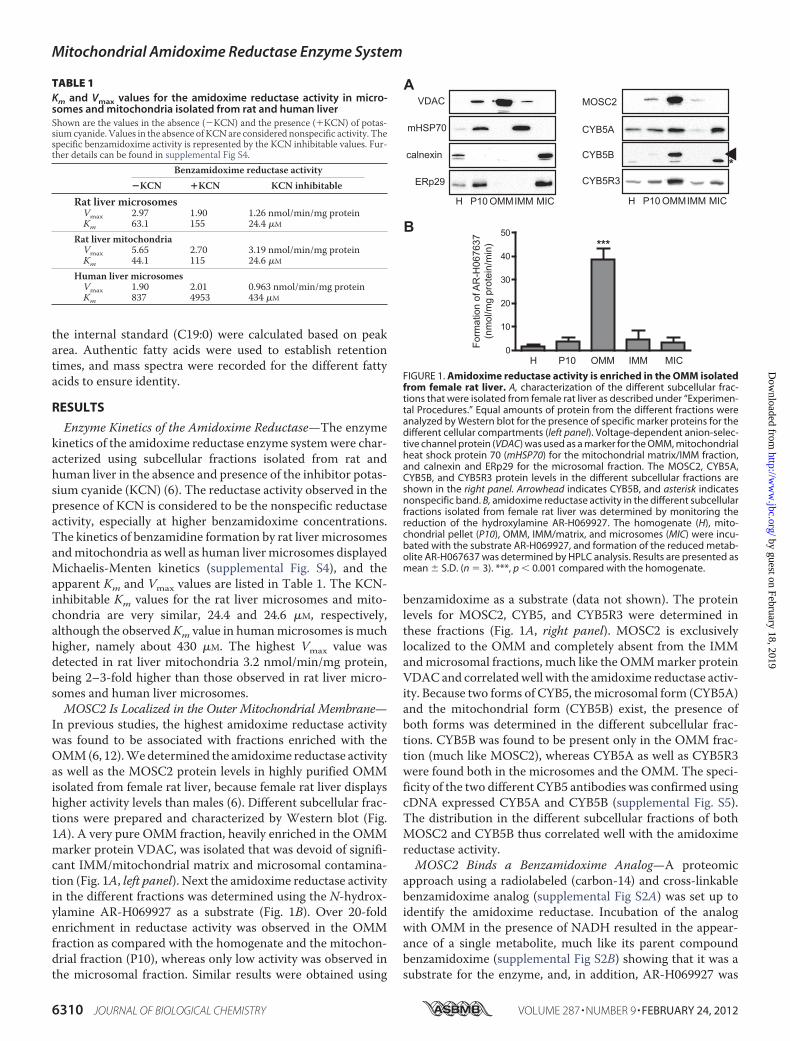
In previous studies, the highest amidoxime reductase activitywas found to be associated with fractions enriched with theOMM(6, 12).Wedetermined the amidoxime reductase activityas well as the MOSC2 protein levels in highly purified OMMisolated from female rat liver, because female rat liver displayshigher activity levels than males (6). Different subcellular frac-tions were prepared and characterized by Western blot (Fig.1A). A very pure OMM fraction, heavily enriched in the OMMmarker protein VDAC, was isolated that was devoid of signifi-cant IMM/mitochondrial matrix and microsomal contamina-tion (Fig. 1A, left panel). Next the amidoxime reductase activityin the different fractions was determined using the N-hydrox-ylamine AR-H069927 as a substrate (Fig. 1B). Over 20-foldenrichment in reductase activity was observed in the OMMfraction as compared with the homogenate and the mitochon-drial fraction (P10), whereas only low activity was observed inthe microsomal fraction. Similar results were obtained using
benzamidoxime as a substrate (data not shown). The proteinlevels for MOSC2, CYB5, and CYB5R3 were determined inthese fractions (Fig. 1A, right panel). MOSC2 is exclusivelylocalized to the OMM and completely absent from the IMMandmicrosomal fractions, much like the OMMmarker proteinVDACand correlatedwell with the amidoxime reductase activ-ity. Because two forms of CYB5, themicrosomal form (CYB5A)and the mitochondrial form (CYB5B) exist, the presence ofboth forms was determined in the different subcellular frac-tions. CYB5B was found to be present only in the OMM frac-tion (much like MOSC2), whereas CYB5A as well as CYB5R3were found both in the microsomes and the OMM. The speci-ficity of the two different CYB5 antibodies was confirmed usingcDNA expressed CYB5A and CYB5B (supplemental Fig. S5).The distribution in the different subcellular fractions of bothMOSC2 and CYB5B thus correlated well with the amidoximereductase activity.MOSC2 Binds a Benzamidoxime Analog—A proteomic
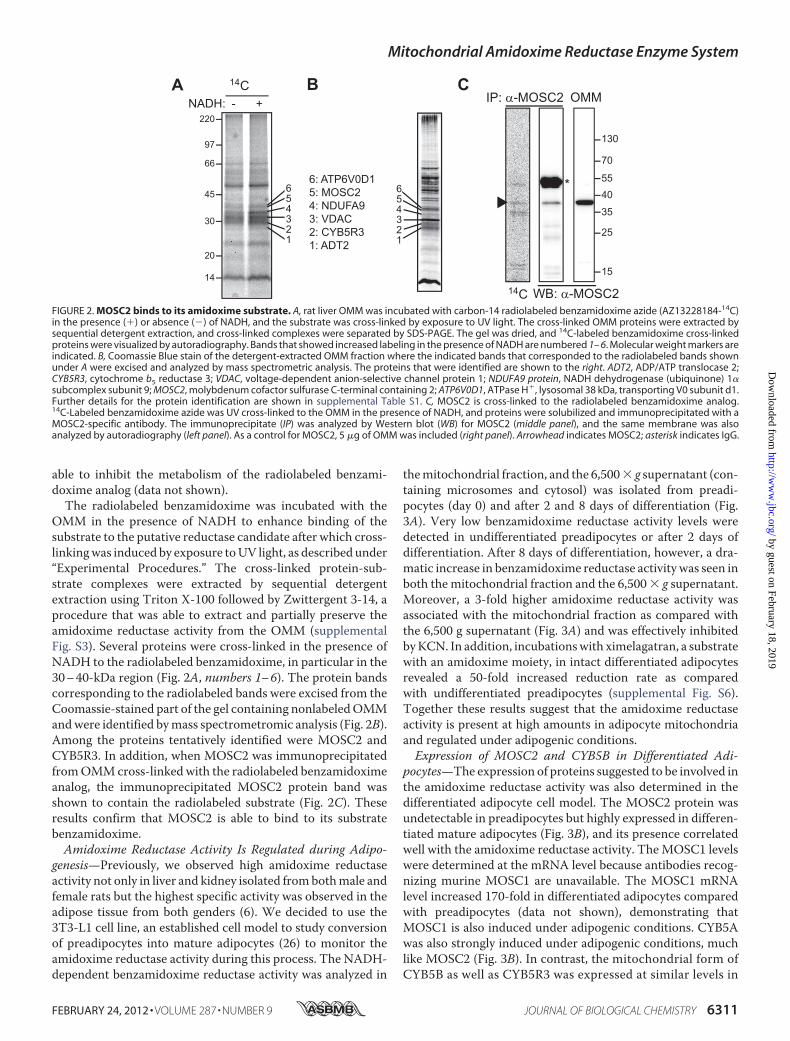
approach using a radiolabeled (carbon-14) and cross-linkablebenzamidoxime analog (supplemental Fig S2A) was set up toidentify the amidoxime reductase. Incubation of the analogwith OMM in the presence of NADH resulted in the appear-ance of a single metabolite, much like its parent compoundbenzamidoxime (supplemental Fig S2B) showing that it was asubstrate for the enzyme, and, in addition, AR-H069927 was
TABLE 1Km and Vmax values for the amidoxime reductase activity in micro-somes and mitochondria isolated from rat and human liverShown are the values in the absence (�KCN) and the presence (�KCN) of potas-sium cyanide. Values in the absence of KCNare considered nonspecific activity. Thespecific benzamidoxime activity is represented by the KCN inhibitable values. Fur-ther details can be found in supplemental Fig S4.
Benzamidoxime reductase activity�KCN �KCN KCN inhibitable
Rat liver microsomesVmax 2.97 1.90 1.26 nmol/min/mg proteinKm 63.1 155 24.4 �M
Rat liver mitochondriaVmax 5.65 2.70 3.19 nmol/min/mg proteinKm 44.1 115 24.6 �M
Human liver microsomesVmax 1.90 2.01 0.963 nmol/min/mg proteinKm 837 4953 434 �M
FIGURE 1. Amidoxime reductase activity is enriched in the OMM isolatedfrom female rat liver. A, characterization of the different subcellular frac-tions that were isolated from female rat liver as described under “Experimen-tal Procedures.” Equal amounts of protein from the different fractions wereanalyzed by Western blot for the presence of specific marker proteins for thedifferent cellular compartments (left panel). Voltage-dependent anion-selec-tive channel protein (VDAC) was used as a marker for the OMM, mitochondrialheat shock protein 70 (mHSP70) for the mitochondrial matrix/IMM fraction,and calnexin and ERp29 for the microsomal fraction. The MOSC2, CYB5A,CYB5B, and CYB5R3 protein levels in the different subcellular fractions areshown in the right panel. Arrowhead indicates CYB5B, and asterisk indicatesnonspecific band. B, amidoxime reductase activity in the different subcellularfractions isolated from female rat liver was determined by monitoring thereduction of the hydroxylamine AR-H069927. The homogenate (H), mito-chondrial pellet (P10), OMM, IMM/matrix, and microsomes (MIC) were incu-bated with the substrate AR-H069927, and formation of the reduced metab-olite AR-H067637 was determined by HPLC analysis. Results are presented asmean � S.D. (n � 3). ***, p � 0.001 compared with the homogenate.
Mitochondrial Amidoxime Reductase Enzyme System
6310 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 9 • FEBRUARY 24, 2012
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from
able to inhibit the metabolism of the radiolabeled benzami-doxime analog (data not shown).The radiolabeled benzamidoxime was incubated with the
OMM in the presence of NADH to enhance binding of thesubstrate to the putative reductase candidate after which cross-linkingwas induced by exposure toUV light, as described under“Experimental Procedures.” The cross-linked protein-sub-strate complexes were extracted by sequential detergentextraction using Triton X-100 followed by Zwittergent 3-14, aprocedure that was able to extract and partially preserve theamidoxime reductase activity from the OMM (supplementalFig. S3). Several proteins were cross-linked in the presence ofNADH to the radiolabeled benzamidoxime, in particular in the30–40-kDa region (Fig. 2A, numbers 1–6). The protein bandscorresponding to the radiolabeled bands were excised from theCoomassie-stained part of the gel containing nonlabeledOMMandwere identified bymass spectrometromic analysis (Fig. 2B).Among the proteins tentatively identified were MOSC2 andCYB5R3. In addition, when MOSC2 was immunoprecipitatedfromOMMcross-linked with the radiolabeled benzamidoximeanalog, the immunoprecipitated MOSC2 protein band wasshown to contain the radiolabeled substrate (Fig. 2C). Theseresults confirm that MOSC2 is able to bind to its substratebenzamidoxime.Amidoxime Reductase Activity Is Regulated during Adipo-
genesis—Previously, we observed high amidoxime reductaseactivity not only in liver and kidney isolated frombothmale andfemale rats but the highest specific activity was observed in theadipose tissue from both genders (6). We decided to use the3T3-L1 cell line, an established cell model to study conversionof preadipocytes into mature adipocytes (26) to monitor theamidoxime reductase activity during this process. The NADH-dependent benzamidoxime reductase activity was analyzed in
themitochondrial fraction, and the 6,500� g supernatant (con-taining microsomes and cytosol) was isolated from preadi-pocytes (day 0) and after 2 and 8 days of differentiation (Fig.3A). Very low benzamidoxime reductase activity levels weredetected in undifferentiated preadipocytes or after 2 days ofdifferentiation. After 8 days of differentiation, however, a dra-matic increase in benzamidoxime reductase activity was seen inboth themitochondrial fraction and the 6,500 � g supernatant.Moreover, a 3-fold higher amidoxime reductase activity wasassociated with the mitochondrial fraction as compared withthe 6,500 g supernatant (Fig. 3A) and was effectively inhibitedbyKCN. In addition, incubationswith ximelagatran, a substratewith an amidoxime moiety, in intact differentiated adipocytesrevealed a 50-fold increased reduction rate as comparedwith undifferentiated preadipocytes (supplemental Fig. S6).Together these results suggest that the amidoxime reductaseactivity is present at high amounts in adipocyte mitochondriaand regulated under adipogenic conditions.Expression of MOSC2 and CYB5B in Differentiated Adi-
pocytes—The expression of proteins suggested to be involved inthe amidoxime reductase activity was also determined in thedifferentiated adipocyte cell model. The MOSC2 protein wasundetectable in preadipocytes but highly expressed in differen-tiated mature adipocytes (Fig. 3B), and its presence correlatedwell with the amidoxime reductase activity. TheMOSC1 levelswere determined at the mRNA level because antibodies recog-nizing murine MOSC1 are unavailable. The MOSC1 mRNAlevel increased 170-fold in differentiated adipocytes comparedwith preadipocytes (data not shown), demonstrating thatMOSC1 is also induced under adipogenic conditions. CYB5Awas also strongly induced under adipogenic conditions, muchlike MOSC2 (Fig. 3B). In contrast, the mitochondrial form ofCYB5B as well as CYB5R3 was expressed at similar levels in
FIGURE 2. MOSC2 binds to its amidoxime substrate. A, rat liver OMM was incubated with carbon-14 radiolabeled benzamidoxime azide (AZ13228184-14C)in the presence (�) or absence (�) of NADH, and the substrate was cross-linked by exposure to UV light. The cross-linked OMM proteins were extracted bysequential detergent extraction, and cross-linked complexes were separated by SDS-PAGE. The gel was dried, and 14C-labeled benzamidoxime cross-linkedproteins were visualized by autoradiography. Bands that showed increased labeling in the presence of NADH are numbered 1– 6. Molecular weight markers areindicated. B, Coomassie Blue stain of the detergent-extracted OMM fraction where the indicated bands that corresponded to the radiolabeled bands shownunder A were excised and analyzed by mass spectrometric analysis. The proteins that were identified are shown to the right. ADT2, ADP/ATP translocase 2;CYB5R3, cytochrome b5 reductase 3; VDAC, voltage-dependent anion-selective channel protein 1; NDUFA9 protein, NADH dehydrogenase (ubiquinone) 1�subcomplex subunit 9; MOSC2, molybdenum cofactor sulfurase C-terminal containing 2; ATP6V0D1, ATPase H�, lysosomal 38 kDa, transporting V0 subunit d1.Further details for the protein identification are shown in supplemental Table S1. C, MOSC2 is cross-linked to the radiolabeled benzamidoxime analog.14C-Labeled benzamidoxime azide was UV cross-linked to the OMM in the presence of NADH, and proteins were solubilized and immunoprecipitated with aMOSC2-specific antibody. The immunoprecipitate (IP) was analyzed by Western blot (WB) for MOSC2 (middle panel), and the same membrane was alsoanalyzed by autoradiography (left panel). As a control for MOSC2, 5 �g of OMM was included (right panel). Arrowhead indicates MOSC2; asterisk indicates IgG.
Mitochondrial Amidoxime Reductase Enzyme System
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6311
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from
both preadipocytes and mature adipocytes and appeared notto be regulated during adipogenesis (Fig. 3B). BesidesCYB5R3, two homologous cytochrome b5 reductases exist inthe mammalian genome, namely CYB5R1 and CYB5R2.Because no commercial antibodies were available that rec-ognize these proteins, their mRNA levels were determined.Both genes were induced in mature adipocytes at the mRNAlevel 140- and 21-fold for CYB5R1 and CYB5R2, respectively(data not shown). Stearoyl-CoA desaturase (SCD1) wasincluded as a positive control for adipocyte differentiationand, as expected, was only expressed in mature adipocytes(Fig. 3B).
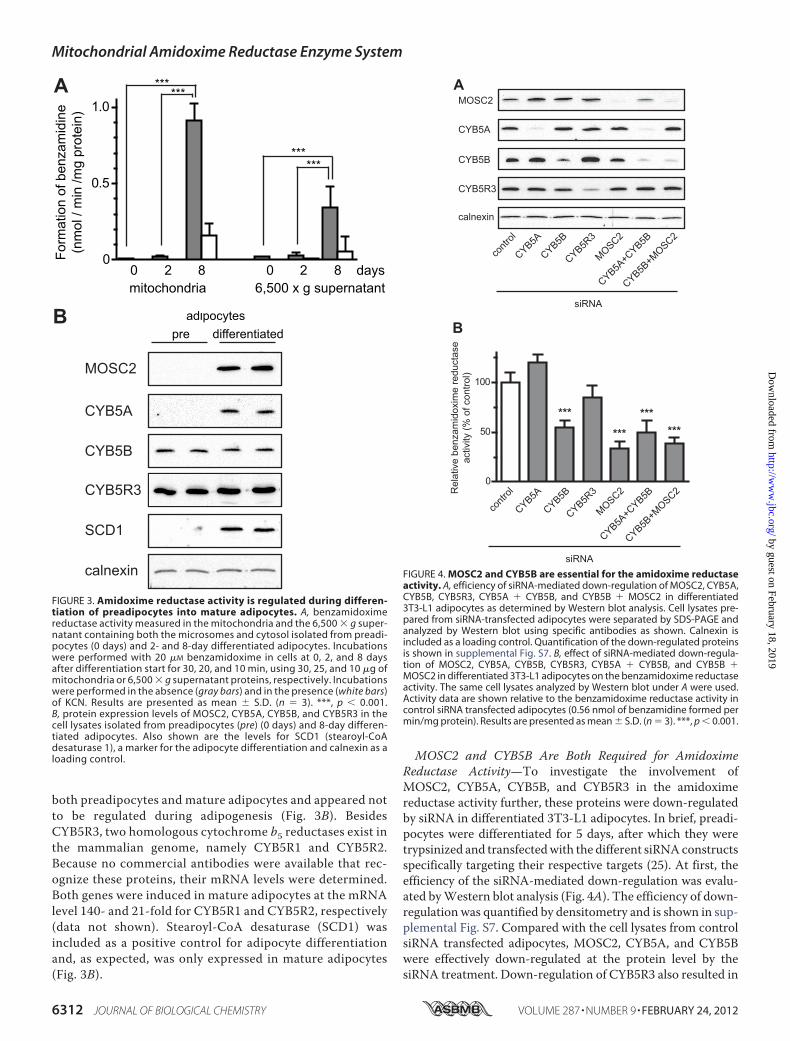
MOSC2 and CYB5B Are Both Required for AmidoximeReductase Activity—To investigate the involvement ofMOSC2, CYB5A, CYB5B, and CYB5R3 in the amidoximereductase activity further, these proteins were down-regulatedby siRNA in differentiated 3T3-L1 adipocytes. In brief, preadi-pocytes were differentiated for 5 days, after which they weretrypsinized and transfectedwith the different siRNAconstructsspecifically targeting their respective targets (25). At first, theefficiency of the siRNA-mediated down-regulation was evalu-ated byWestern blot analysis (Fig. 4A). The efficiency of down-regulation was quantified by densitometry and is shown in sup-plemental Fig. S7. Compared with the cell lysates from controlsiRNA transfected adipocytes, MOSC2, CYB5A, and CYB5Bwere effectively down-regulated at the protein level by thesiRNA treatment. Down-regulation of CYB5R3 also resulted in
FIGURE 3. Amidoxime reductase activity is regulated during differen-tiation of preadipocytes into mature adipocytes. A, benzamidoximereductase activity measured in the mitochondria and the 6,500 � g super-natant containing both the microsomes and cytosol isolated from preadi-pocytes (0 days) and 2- and 8-day differentiated adipocytes. Incubationswere performed with 20 �M benzamidoxime in cells at 0, 2, and 8 daysafter differentiation start for 30, 20, and 10 min, using 30, 25, and 10 �g ofmitochondria or 6,500 � g supernatant proteins, respectively. Incubationswere performed in the absence (gray bars) and in the presence (white bars)of KCN. Results are presented as mean � S.D. (n � 3). ***, p � 0.001.B, protein expression levels of MOSC2, CYB5A, CYB5B, and CYB5R3 in thecell lysates isolated from preadipocytes (pre) (0 days) and 8-day differen-tiated adipocytes. Also shown are the levels for SCD1 (stearoyl-CoAdesaturase 1), a marker for the adipocyte differentiation and calnexin as aloading control.
FIGURE 4. MOSC2 and CYB5B are essential for the amidoxime reductaseactivity. A, efficiency of siRNA-mediated down-regulation of MOSC2, CYB5A,CYB5B, CYB5R3, CYB5A � CYB5B, and CYB5B � MOSC2 in differentiated3T3-L1 adipocytes as determined by Western blot analysis. Cell lysates pre-pared from siRNA-transfected adipocytes were separated by SDS-PAGE andanalyzed by Western blot using specific antibodies as shown. Calnexin isincluded as a loading control. Quantification of the down-regulated proteinsis shown in supplemental Fig. S7. B, effect of siRNA-mediated down-regula-tion of MOSC2, CYB5A, CYB5B, CYB5R3, CYB5A � CYB5B, and CYB5B �MOSC2 in differentiated 3T3-L1 adipocytes on the benzamidoxime reductaseactivity. The same cell lysates analyzed by Western blot under A were used.Activity data are shown relative to the benzamidoxime reductase activity incontrol siRNA transfected adipocytes (0.56 nmol of benzamidine formed permin/mg protein). Results are presented as mean � S.D. (n � 3). ***, p � 0.001.
Mitochondrial Amidoxime Reductase Enzyme System
6312 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 9 • FEBRUARY 24, 2012
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

significant decrease in both CYB5R3 mRNA and protein levelswith �21% of the protein remaining after siRNA transfection,and at the mRNA level 25% remained (Figs. 4A and 5A andsupplemental Fig. S7). Simultaneous down-regulation ofCYB5A together with CYB5B orMOSC2 together with CYB5Balso resulted in effective down-regulation of both of the tar-geted proteins (Fig. 4A and supplemental Fig. S7).
Down-regulation ofMOSC2 alone resulted in a 65%decreasein the amidoxime reductase activity as compared with controltransfected cells (Fig. 4B), although the protein levels weredown-regulated to 6% of those present in control transfectedcells (Fig. 4A and supplemental Fig. S7). Down-regulation ofCYB5A had a slightly simulating effect on the reductase activ-ity, and although reproducible this was not statistically signifi-cant (Fig. 4B), although protein levels were down-regulated to6% of the levels present in control cells (Fig. 4A and supplemen-tal Fig. S7). In contrast, down-regulation CYB5B resulted in asignificant decrease in activity of about 50% (Fig. 4B), with 31%of the protein remaining (Fig. 4A and supplemental Fig. S7).Also, when both forms of CYB5 (CYB5A � CYB5B) weresimultaneously knocked down, activity was decreased by about50% (Fig. 4B) with 10% or less of both proteins remaining (Fig.4A and supplemental Fig. S7). These results clearly indicatedthat MOSC2 and the mitochondrial form of cytochrome b5(CYB5B), but not the microsomal CYB5A, are involved in theamidoxime reductase reaction.Simultaneous down-regulation of MOSC2 and CYB5B did
not further decrease the reductase activity as compared withdown-regulation of the individual proteins (Fig. 4B). Down-regulation of CYB5R3 did not significantly alter the reductaseactivity, even though the observed activity was slightly lowerthan control (Fig. 4B). All activities were efficiently inhibited byKCN indicating that the reductase activity was indeed specific(data not shown). Together, these data clearly demonstrate thatMOSC2 as well as CYB5B, but not CYB5A and CYB5R3, areimportant components of the amidoxime reductase enzymecomplex in differentiated adipocytes.MOSC1, CYB5R1, and CYB5R2 Are Not Involved in the Ami-
doxime Reductase Activity—MOSC1 has been shown to be ableto catalyze the reduction of amidoximes in reconstituted sys-tems containing soluble and truncated forms ofMOSC1,CYB5,and CYB5R3 (13, 14). The involvement of MOSC1 in the ami-doxime reductase activity in mature adipocytes was studied
also by down-regulation of MOSC1 in these cells. As shown inFig. 5A,MOSC1 siRNA down-regulated MOSC1 mRNA levelsto about 20% of the control levels. The amidoxime reductaseactivity in MOSC1 down-regulated adipocytes was not signifi-cantly affected (Fig. 5B).The involvement ofCYB5R1 andCYB3R2was also examined
by siRNA-mediated down-regulation in the mature adipocytecell model. Both CYB5R1 and CYB5R2 are efficiently down-regulated at the mRNA level with 20% or less of the levelsobserved in control transfected cells (Fig. 5A). The amidoximereductase activity in the CYB5R1 and CYB5R2 down-regulatedadipocytes was unaffected as compared with control cells (Fig.5B). In addition, simultaneous down-regulation of CYB5R1,CYB5R2, andCYB5R3 did not inhibit the amidoxime reductaseactivity either (data not shown).Finally, the involvement of CYB5R in the amidoxime reduc-
tase activity was studied using the specific CYB5R inhibitorPTU (27). Although PTU was able to efficiently inhibit theCYB5R activity with almost 90% efficiency (Fig. 6B), it was notable to significantly inhibit the benzamidoxime reductase activ-ity in theOMMfraction (Fig. 6A), again confirming thatCYB5Ris not an essential component of the amidoxime reductaseenzyme system in these cells.Down-regulation of MOSC2 Affects Intracellular Lipid
Levels—Because MOSC2 is up-regulated during adipogenesis,we investigated the effect of down-regulation ofMOSC2 on theintracellular lipid content in mature adipocytes. Adipocyteswere transfected with MOSC2 siRNA, and 5 days after trans-fection cells were stained for intracellular lipids usingOil RedO(Fig. 7A). Down-regulation of MOSC2 resulted in a significantdecrease (17%, n � 6; p � 0.001) of the intracellular lipid levelsas compared with control transfected cells. The intracellularfatty acids were also analyzed by GC/MS, and the amounts forC14:0, C16:0, C16:1, C18:0, andC18:1 were determined relativeto the internal standard (C19:0) (Fig. 7B). Down-regulation ofMOSC2 resulted in decreased levels of these fatty acids, thereduction being most pronounced for C16:0 and C16:1 withabout a 28% decrease.
DISCUSSION
In this study, we demonstrate for the first time in an intactcell system (mature adipocytes) that the mitochondrial ami-doxime activity is dependent on the expression of MOSC2 and
FIGURE 5. MOSC1 is not involved in the amidoxime reductase activity. A, efficiency of the siRNA-mediated down-regulation of MOSC1, CYB5R1, CYB5R2,and CYB5R3 in differentiated adipocytes as determined by RT-PCR. nd means not detected. B, effect of the siRNA-mediated down-regulation of MOSC1,CYB5R1, and CYB5R2 on the benzamidoxime reductase activity in differentiated adipocytes. Activity data are shown relative to the benzamidoxime reductaseactivity in control siRNA-transfected adipocytes. Results are presented as mean � S.D. (n � 3). *, p � 0.05.
Mitochondrial Amidoxime Reductase Enzyme System
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6313
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

CYB5B. CYB5A, the microsomal form of cytochrome b5,although present in the OMM, is not critically involved in themitochondrial amidoxime reductase activity. Furthermore,MOSC1 and CYB5R1, CYB5R2, or CYB5R3 were found not tobe crucial components of themitochondrial amidoxime reduc-tase enzyme system in mature adipocytes. In addition, our datashow that MOSC2 affects the fatty acid levels in adipocytessuggesting a role of MOSC2 in lipogenesis, a finding consistentwith its lipogenic regulation in the adipocytes.Further characterization established that MOSC2 is exclu-
sively localized to the OMM isolated from rat liver, and itsexpression correlated well with the amidoxime reductase activ-ity. Using a cross-linkable benzamidoxime analog and immu-noprecipitation, we could show direct binding of a benzami-doxime substrate to MOSC2. Differentiation of preadipocytesinto mature adipocytes resulted in an over 100-fold increase inmitochondrial associated amidoxime reductase activity andwas paralleled by an increase in MOSC2 expression. MoreoversiRNA-mediated down-regulation of MOSC2 attenuated theamidoxime reductase activity. An interesting observation wasthat although CYB5A was regulated under adipogenic condi-tions and CYB5B was not, down-regulation of CYB5B and notCYB5A inhibited the reductase activity, showing the involve-
ment of the mitochondrial form but not the microsomal formof cytochrome b5 in the amidoxime reductase activity. In ouradipocyte cell model, we were unable to demonstrate theinvolvement of MOSC1 in the reductase activity. MOSC1 waslike its homolog MOSC2 up-regulated in differentiated adi-pocytes, but down-regulation of MOSC1 in these cells did notaffect the amidoxime reductase activity.MOSC1 has previouslybeen shown to be able to reduce amidoximes to amidines invitro (13, 14). The reason for this discrepancy is unclear, but inthe in vitro system the amidoxime reductase activity was recon-stituted with recombinantly expressed truncated and solublecomponents CYB5B and CYB5R3 and MOSC1 or MOSC2.Although soluble CYB5R3 can support the amidoxime
reductase activity in vitro together with truncated solubleMOSC2 and CYB5B (13, 14), we were unable to confirm itsinvolvement in the adipocyte cell system.This is despite the factthatwe identifiedCYB5R3 as one of the proteins thatwas cross-linked by the radiolabeled substrate in the OMM and mightpossibly reflect not direct binding of the substrate to CYB5R3but background caused by nonspecific cross-linking back-ground. The closely related genes CYB5R1 and CYB5R2 werealso found not to be crucially involved in the reductase activity,although they were up-regulated in mature adipocytes. More-over, the selective CYB5R inhibitor PTU (27) was unable toinhibit the amidoxime reductase activity in the OMM.Together, these data suggest that another presently unknownreductase is involved in the NADH-supported mitochondrialamidoxime reductase activity. Because the other two compo-nentsMOSC2 andCYB5B are shown to be exclusively localizedto the mitochondrial outer membrane, it is suggested that theunknown reductase is also present in this compartment.CYB5B and CYB5A share considerable homology with each
other (28), and although much is known about the function ofCYB5A not much is known about the function of CYB5B.CYB5B has been suggested to be involved in the semi-dehy-droascorbic acid reductase activity (29) and is thought to func-tion as an activator of androgen synthesis by CYP17A1 (30).More recently, increased CYB5B levels were observed inHodg-kin and aggressive non-Hodgkin lymphomas (31) and luteiniz-ing hormone stimulated prostate cancer cell lines (32),although the exact role of elevated CYB5B levels in these can-
FIGURE 6. Inhibition of the CYB5R activity by PTU does not affect the amidoxime reductase activity. A, increasing concentrations of PTU do not inhibit thebenzamidoxime reductase activity. OMM was incubated with benzamidoxime in the absence (control) and presence of 0.05, 0.5, and 5.0 mM PTU, and thebenzamidoxime reductase activity was determined. Activity data are shown relative to the benzamidoxime activity in control OMM. B, efficiency of the CYB5Rinhibitor PTU was evaluated in rat liver microsomes by monitoring the reduction of 2,6-dichlorophenolindophenol in the absence (control) and the presenceof 0.05, 0.5, and 5.0 mM PTU. Activity data are shown relative to control microsomes. Results are presented as mean � S.D. (n � 3). **, p � 0.005.
FIGURE 7. MOSC2 levels affect the intracellular lipid content. A, matureadipocytes were transfected with control or MOSC2 siRNA and 5 days post-transfection were stained with Oil Red O. The dye was eluted in isopropylalcohol and quantified spectrophotometrically at 510 nm. Results are pre-sented as mean � S.D. (n � 6). ***, p � 0.001. B, relative amounts of fatty acidsin control and MOSC2 siRNA-transfected adipocytes. Transfected adipocyteswere harvested 5 days post-transfection, and cellular fatty acids wereextracted, converted into methyl esters, and analyzed by GC-MS. An internalstandard (IS), nonadecanoic acid (C19:0), was added to the cells before extrac-tion as described under “Experimental Procedures.”
Mitochondrial Amidoxime Reductase Enzyme System
6314 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 9 • FEBRUARY 24, 2012
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

cers remains unclear. Here, we show that CYB5B is exclusivelyassociated with the OMM, although it has been suggested thatCYB5B was present in both mitochondria and microsomes(30). In contrast, CYB5A and CYB5R3 display a less definedlocalization and are found to be associatedwith both themicro-somal membrane and the OMM, confirming previous observa-tions (16, 33). Based on the findings presented here, we proposea novel function for CYB5B, namely in the reduction of N-hy-droxylated amines together with MOSC2.The amidoxime reductase enzyme kinetic parameters dif-
fered considerably between human liver microsomes and ratliver microsomes and mitochondria, with rat liver mitochon-dria being themost efficient. The amidoxime reductase activityin isolated rat liver OMM (data not shown) and in mitochon-dria isolated from differentiated adipocytes was effectivelyinhibited byKCN.Cyanide has been shown to be an inhibitor ofthe reduced form of molybdenum-containing proteins such asxanthine oxidase, aldehyde oxidase, and sulfite oxidase (34).The inhibitory effect of KCN on the amidoxime reductaseactivity in our system is in good agreement with these observa-tions. It was recently reported that an in vitro recombinantsystem using truncated MOSC1 and MOSC2 that cyanide wasnot able to inhibit the reductase activity, and it was subse-quently concluded that MOSC1 and MOSC2 do not contain aterminal sulfide ligand on the molybdenum as is the case inxanthine oxidase, for example (14). One explanation for thisinconsistency might be that we included the cyanide directly inthe incubation mixture containing OMMor the mitochondrialfraction from adipocytes together with cofactor and substrateunder aerobic conditions, although Wahl et al. (14) incubatedthe recombinant proteins with cyanide under anaerobic condi-tions prior to the incubation with cofactor and substrate.Molybdenum is an essential trace element for nearly all
organisms, and molybdenum-dependent enzymes, such as sul-fite oxidase and xanthine oxidase, are conserved in all threedomains of life and usually catalyze redox reactions on carbon,nitrogen, or sulfur centers (35, 36). The MOSC (molybdenumcofactor sulfurase C-terminal) family of proteins is conservedin most eukaryotes and is responsible for the sulfuration of themolybdenum cofactor present in xanthine dehydrogenase andaldehyde oxidase, a modification essential for their catalyticactivity (37). In eukaryotes, this modification is catalyzed bymolybdenum cofactor sulfurase, a protein consisting of the fol-lowing two domains: an N-terminal NifS domain, which pos-sesses cysteine desulfurase activity, and a C-terminal MOSCdomain responsible for the transfer of the sulfur to the molyb-
denum cofactor (35, 37). Two other conserved members of theMOSC family only possess the MOSC domain and are calledMOSC1 and MOSC2. The physiological role of these two pro-teins is at present not known, but MOSC2 or CDK7 (candidatediabetes-associated kidney gene 7) was shown to be up-regu-lated in the kidney in a type II diabetes ratmodel and by glucosein human kidney cells (38). In Escherichia coli, hypersensitivitytoward N-hydroxylated derivatives of purines and pyrimidineswas shown to be caused by inactivatingmutations of twomem-bers of the MOSC superfamily, namely YcbX and YiiM (39).Moreover, it was suggested that both proteins were involved inthe detoxification of these N-hydroxylated base analogs byreducing them to their corresponding amines. This findingtogether with ours presented here suggests that the MOSCdomain-containing proteins are capable of catalyzing reduc-tion ofN-hydroxylamines and that this activity could be one ofthe physiological functions of these proteins. In a recent inte-grated proteomic and transcriptomic high throughput study,MOSC2 mRNA and protein levels were found to be down-reg-ulated in colon cancer, and it was suggested thatMOSC2 couldbe used as a colon tumor biomarker (40).Down-regulation of MOSC2 in mature adipocytes not only
affected the amidoxime activity but also decreased the intracel-lular lipid levels as shown by Oil Red O staining, and also thefatty acid levels were decreased in these MOSC2 down-regu-lated adipocytes. Together with the fact that high amidoximereductase activity was found in the adipose tissue in rats (6),these results imply that this activity is the result of a lipogenicenzyme. In contrast, the amidoxime reductase activity in liverfractions from starvation-treated rats was decreased by 75% ascompared with liver fractions from rats that received food adlibitum suggesting that during lipolytic conditions the activityis down-regulated (data not shown). Recently, a genome-wideassociation study identified a single nucleotide polymorphismlinked to the MOSC1 locus that was associated with alteredplasma concentrations of total cholesterol and low density lipo-protein cholesterol, two important risk factors for coronaryartery disease, suggesting a possible role of MOSC in lipopro-tein metabolism (41). It must be noted that in the 3T3-L1 adi-pocyte cell model, the up-regulation of the reductase activityalso coincided with that of the adipocyte differentiationmarkerSCD1 suggesting its regulation during adipogenesis. The exactrole of the outer mitochondrial electron transport chain andMOSC2 in lipid synthesis requires further research.In conclusion, we demonstrate using an intact cell system
thatMOSC2 and CYB5B are essential components of themito-
FIGURE 8. Proposed reaction scheme for the mitochondrial amidoxime reductase enzyme system. For more details see under “Discussion.” FAD, flavinadenine dinucleotide (oxidized); FADH2, flavin adenine dinucleotide (reduced).
Mitochondrial Amidoxime Reductase Enzyme System
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6315
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

chondrial amidoxime reductase enzyme system. The identifi-cation of MOSC2 in the amidoxime reductase system is inaccordance with results obtained with soluble protein compo-nents in a reconstituted system (13, 14). Our findings are sche-matically summarized in Fig. 8. Electrons from NADH aretransferred via an as yet unidentified reductase (FAD) toCYB5B, which in its turn will reduce the MOSC2-benzami-doxime complex and produce the reduced product benzami-dine. As both MOSC2 and CYB5B are exclusively localized inthe OMM, it is hypothesized that all components of the ami-doxime reductase enzyme system are embedded in the outermitochondrial membrane. Moreover, the reductase activity isregulated under adipogenic conditions, and down-regulation ofthe terminal component MOSC2 resulted in decreased lipidsynthesis, suggesting a possible physiological role of thisenzyme system and its component MOSC2 in lipogenesis.
Acknowledgments—We thank Prof. N. Borgese (Milan, Italy) and Dr.S. Mkrtchian (Stockholm, Sweden) for the gift of antibodies. We alsothank Malin Lemurell (Lead Generation, AstraZeneca R&D) for theidea to synthesize the radiolabeled azide compound; the isotope groupat AstraZeneca R&D, Mölndal, Sweden, for synthesizing the labeledazide used in the study, and Stefan C. Carlsson, AstraZeneca, forinitial support.
REFERENCES1. Eriksson, U. G., Bredberg, U., Hoffmann, K. J., Thuresson, A., Gabrielsson,
M., Ericsson, H., Ahnoff, M., Gislén, K., Fager, G., and Gustafsson, D.(2003) Absorption, distribution, metabolism, and excretion of ximelaga-tran, an oral direct thrombin inhibitor, in rats, dogs, and humans. DrugMetab. Dispos. 31, 294–305
2. Gustafsson, D., Nyström, J., Carlsson, S., Bredberg, U., Eriksson, U.,Gyzander, E., Elg, M., Antonsson, T., Hoffmann, K., Ungell, A., Sörensen,H., Någård, S., Abrahamsson, A., and Bylund, R. (2001) The direct throm-bin inhibitor melagatran and its oral prodrug H 376/95. Intestinal absorp-tion properties, biochemical and pharmacodynamic effects. Thromb. Res.101, 171–181
3. Deinum, J.,Mattsson, C., Inghardt, T., and Elg,M. (2009) Biochemical andpharmacological effects of the direct thrombin inhibitor AR-H067637.Thromb. Haemost. 101, 1051–1059
4. Timm, U., Zumbrunnen, R., Erdin, R., Singer, M., and Steiner, B. (1997)Oral platelet aggregation inhibitor Ro 48-3657. Determination of the ac-tive metabolite and its prodrug in plasma and urine by high performanceliquid chromatography using automated column switching. J. Chro-matogr. B Biomed. Sci. Appl. 691, 397–407
5. Kadlubar, F. F., and Ziegler, D. M. (1974) Properties of an NADH-depen-dentN-hydroxyamine reductase isolated frompig livermicrosomes.Arch.Biochem. Biophys. 162, 83–92
6. Andersson, S., Hofmann, Y., Nordling, A., Li, X. Q., Nivelius, S., Ander-sson, T. B., Ingelman-Sundberg, M., and Johansson, I. (2005) Character-ization and partial purification of the rat and human enzyme systemsactive in the reduction of N-hydroxymelagatran and benzamidoxime.Drug Metab. Dispos. 33, 570–578
7. Bernheim,M. L., andHochstein, P. (1968) Reduction of hydroxylamine byrat liver mitochondria. Arch. Biochem. Biophys. 124, 436–442
8. King, R. S., Teitel, C. H., Shaddock, J. G., Casciano, D. A., and Kadlubar,F. F. (1999) Detoxification of carcinogenic aromatic and heterocyclicamines by enzymatic reduction of the N-hydroxy derivative. Cancer Lett.143, 167–171
9. Kurian, J. R., Chin, N. A., Longlais, B. J., Hayes, K. L., and Trepanier, L. A.(2006) Reductive detoxification of arylhydroxylamine carcinogens by hu-man NADH cytochrome b5 reductase and cytochrome b5. Chem. Res.Toxicol. 19, 1366–1373
10. Clement, B., Lomb, R., and Möller, W. (1997) Isolation and characteriza-tion of the protein components of the liver microsomal O2-insensitiveNADH-benzamidoxime reductase. J. Biol. Chem. 272, 19615–19620
11. Reh, R., Ozols, J., and Clement, B. (2008) Involvement of stearoyl-CoAdesaturase in the reduction of amidoxime prodrugs. Xenobiotica 38,1177–1190
12. Havemeyer, A., Bittner, F., Wollers, S., Mendel, R., Kunze, T., and Clem-ent, B. (2006) Identification of the missing component in the mitochon-drial benzamidoxime prodrug-converting system as a novel molybdenumenzyme. J. Biol. Chem. 281, 34796–34802
13. Gruenewald, S.,Wahl, B., Bittner, F., Hungeling, H., Kanzow, S., Kotthaus,J., Schwering, U., Mendel, R. R., and Clement, B. (2008) The fourth mo-lybdenum containing enzyme mARC. Cloning and involvement in theactivation of N-hydroxylated prodrugs. J. Med. Chem. 51, 8173–8177
14. Wahl, B., Reichmann, D., Niks, D., Krompholz, N., Havemeyer, A., Clem-ent, B., Messerschmidt, T., Rothkegel, M., Biester, H., Hille, R., Mendel,R. R., and Bittner, F. (2010) Biochemical and spectroscopic characteriza-tion of the human mitochondrial amidoxime reducing componentshmARC-1 and hmARC-2 suggests the existence of a new molybdenumenzyme family in eukaryotes. J. Biol. Chem. 285, 37847–37859
15. Kurian, J. R., Bajad, S. U., Miller, J. L., Chin, N. A., and Trepanier, L. A.(2004) NADH cytochrome b5 reductase and cytochrome b5 catalyze themicrosomal reduction of xenobiotic hydroxylamines and amidoximes inhumans. J. Pharmacol. Exp. Ther. 311, 1171–1178
16. Borgese, N., Aggujaro, D., Carrera, P., Pietrini, G., and Bassetti, M. (1996)A role for N-myristoylation in protein targeting. NADH-cytochrome b5reductase requires myristic acid for association with outer mitochondrialbut not ER membranes. J. Cell Biol. 135, 1501–1513
17. Mkrtchian, S., Fang, C., Hellman, U., and Ingelman-Sundberg, M. (1998)A stress-inducible rat liver endoplasmic reticulum protein, ERp29. Eur.J. Biochem. 251, 304–313
18. Andersson, A.M., and Pettersson, R. F. (1998) Targeting of a short peptidederived from the cytoplasmic tail of the G1 membrane glycoprotein ofUukuniemi virus (Bunyaviridae) to the Golgi complex. J. Virol. 72,9585–9596
19. Hovius, R., Lambrechts, H., Nicolay, K., and de Kruijff, B. (1990) Improvedmethods to isolate and subfractionate rat liver mitochondria. Lipid com-position of the inner and outer membrane. Biochim. Biophys. Acta 1021,217–226
20. de Kroon, A. I., Dolis, D., Mayer, A., Lill, R., and de Kruijff, B. (1997)Phospholipid composition of highly purified mitochondrial outer mem-branes of rat liver and Neurospora crassa. Is cardiolipin present in themitochondrial outer membrane? Biochim. Biophys. Acta 1325, 108–116
21. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951)Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193,265–275
22. Sivertsson, L., Ek,M.,Darnell,M., Edebert, I., Ingelman-Sundberg,M., andNeve, E. P. (2010) CYP3A4 catalytic activity is induced in confluent Huh7hepatoma cells. Drug Metab. Dispos. 38, 995–1002
23. Johansson, S., Cullberg, M., Eriksson, U. G., Elg, M., Dunér, K., Jensen, E.,Wollbratt, M., and Wåhlander, K. (2011) Single-dose pharmacokinetics,pharmacodynamics, and safety of AZD0837, a novel oral direct thrombininhibitor, in young healthymale subjects. Int. J. Clin. Pharmacol. Ther. 49,258–267
24. Hellman, U. (2000) in Proteomics in Functional Genomics. Protein Struc-ture Analysis (Jollès, P., and Jörnvall, H., eds) pp. 43–54, Birkhauser VerlagAG, Basel
25. Kilroy, G., Burk, D. H., and Floyd, Z. E. (2009) High efficiency lipid-basedsiRNA transfection of adipocytes in suspension. PLoS One 4, e6940
26. Ntambi, J. M., and Young-Cheul, K. (2000) Adipocyte differentiation andgene expression. J. Nutr. 130, 3122S-3126S
27. Kariya, K., Lee, E., Yamaoka, M., and Ishikawa, H. (1984) Selective induc-tion of cytochrome b5 and NADH cytochrome b5 reductase by propylth-iouracil. Life Sci. 35, 2327–2334
28. Altuve, A., Wang, L., Benson, D. R., and Rivera, M. (2004) Mammalianmitochondrial and microsomal cytochromes b5 exhibit divergent struc-tural and biophysical characteristics. Biochem. Biophys. Res. Commun.314, 602–609
Mitochondrial Amidoxime Reductase Enzyme System
6316 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 9 • FEBRUARY 24, 2012
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

29. Ito, A., Hayashi, S., and Yoshida, T. (1981) Participation of a cytochromeb5-like hemoprotein of outer mitochondrial membrane (OM cytochromeb) in NADH-semidehydroascorbic acid reductase activity of rat liver.Biochem. Biophys. Res. Commun. 101, 591–598
30. Ogishima, T., Kinoshita, J. Y., Mitani, F., Suematsu, M., and Ito, A. (2003)Identification of outermitochondrial membrane cytochrome b5 as amod-ulator for androgen synthesis in Leydig cells. J. Biol. Chem. 278,21204–21211
31. Murphy, D., Parker, J., Zhou, M., Fadlelmola, F. M., Steidl, C., Karsan, A.,Gascoyne, R. D., Chen, H., and Banerjee, D. (2010) Constitutively overex-pressed 21-kDa protein in Hodgkin lymphoma and aggressive non-Hodg-kin lymphomas identified as cytochrome Bb5 (CYB5B).Mol. Cancer 9, 14
32. Pinski, J., Xiong, S.,Wang,Q., Stanczyk, F., Hawes, D., and Liu, S. V. (2011)Effect of luteinizing hormone on the steroidogenic pathway in prostatecancer. Prostate 71, 892–898
33. Carlsen, J., and Christiansen, K. (1995) The subcellular localization ofnewly synthesized cytochrome b5. Cell Biol. Int. 19, 759–767
34. Coughlan,M. P., Johnson, J. L., and Rajagopalan, K. V. (1980)Mechanismsof inactivation of molybdoenzymes by cyanide. J. Biol. Chem. 255,2694–2699
35. Schwarz, G., Mendel, R. R., and Ribbe, M. W. (2009) Molybdenum cofac-tors, enzymes, and pathways. Nature 460, 839–847
36. Zhang, Y., and Gladyshev, V. N. (2008) Molybdoproteomes and evolutionof molybdenum utilization. J. Mol. Biol. 379, 881–899
37. Anantharaman, V., and Aravind, L. (2002) MOSC domains. Ancient, pre-dicted sulfur-carrier domains, present in diverse metal-sulfur cluster bio-synthesis proteins, including molybdenum cofactor sulfurases. FEMSMi-crobiol. Lett. 207, 55–61
38. Malik, A. N., Rossios, C., Al-Kafaji, G., Shah, A., and Page, R. A. (2007)Glucose regulation of CDK7, a putative thiol related gene, in experimentaldiabetic nephropathy. Biochem. Biophys. Res. Commun. 357, 237–244
39. Kozmin, S. G., Leroy, P., Pavlov, Y. I., and Schaaper, R.M. (2008) YcbX andyiiM, two novel determinants for resistance of Escherichia coli to N-hy-droxylated base analogs.Mol. Microbiol. 68, 51–65
40. Mikula, M., Rubel, T., Karczmarski, J., Goryca, K., Dadlez, M., and Os-trowski, J. (2010) Integrating proteomic and transcriptomic high through-put surveys for search of new biomarkers of colon tumors. Funct. Integr.Genomics 11, 215–224
41. Teslovich, T. M., Musunuru, K., Smith, A. V., Edmondson, A. C., Styl-ianou, I.M., Koseki,M., Pirruccello, J. P., Ripatti, S., Chasman, D. I.,Willer,
C. J., Johansen, C. T., Fouchier, S. W., Isaacs, A., Peloso, G. M., Barbalic,M., Ricketts, S. L., Bis, J. C., Aulchenko, Y. S., Thorleifsson, G., Feitosa,M. F., Chambers, J., Orho-Melander,M.,Melander, O., Johnson, T., Li, X.,Guo, X., Li, M., Shin Cho, Y., Jin Go, M., Jin Kim, Y., Lee, J. Y., Park, T.,Kim, K., Sim, X., Twee-Hee Ong, R., Croteau-Chonka, D. C., Lange, L. A.,Smith, J. D., Song, K., Hua Zhao, J., Yuan, X., Luan, J., Lamina, C., Ziegler,A., Zhang, W., Zee, R. Y., Wright, A. F., Witteman, J. C., Wilson, J. F.,Willemsen, G., Wichmann, H. E., Whitfield, J. B., Waterworth, D. M.,Wareham, N. J., Waeber, G., Vollenweider, P., Voight, B. F., Vitart, V.,Uitterlinden, A. G., Uda, M., Tuomilehto, J., Thompson, J. R., Tanaka, T.,Surakka, I., Stringham, H. M., Spector, T. D., Soranzo, N., Smit, J. H.,Sinisalo, J., Silander, K., Sijbrands, E. J., Scuteri, A., Scott, J., Schlessinger,D., Sanna, S., Salomaa, V., Saharinen, J., Sabatti, C., Ruokonen, A., Rudan,I., Rose, L. M., Roberts, R., Rieder, M., Psaty, B. M., Pramstaller, P. P.,Pichler, I., Perola, M., Penninx, B. W., Pedersen, N. L., Pattaro, C., Parker,A. N., Pare, G., Oostra, B. A., O’Donnell, C. J., Nieminen,M. S., Nickerson,D. A.,Montgomery, G.W.,Meitinger, T.,McPherson, R.,McCarthy,M. I.,McArdle, W., Masson, D., Martin, N. G., Marroni, F., Mangino, M., Mag-nusson, P. K., Lucas, G., Luben, R., Loos, R. J., Lokki, M. L., Lettre, G.,Langenberg, C., Launer, L. J., Lakatta, E. G., Laaksonen, R., Kyvik, K. O.,Kronenberg, F., König, I. R., Khaw, K. T., Kaprio, J., Kaplan, L. M., Johans-son, A., Jarvelin,M. R., Janssens, A. C., Ingelsson, E., Igl,W., KeesHovingh,G., Hottenga, J. J., Hofman, A., Hicks, A. A., Hengstenberg, C., Heid, I. M.,Hayward, C., Havulinna, A. S., Hastie, N. D., Harris, T. B., Haritunians, T.,Hall, A. S., Gyllensten, U., Guiducci, C., Groop, L. C., Gonzalez, E., Gieger,C., Freimer, N. B., Ferrucci, L., Erdmann, J., Elliott, P., Ejebe, K. G., Döring,A., Dominiczak, A. F., Demissie, S., Deloukas, P., deGeus, E. J., de Faire, U.,Crawford, G., Collins, F. S., Chen, Y. D., Caulfield, M. J., Campbell, H.,Burtt, N. P., Bonnycastle, L. L., Boomsma,D. I., Boekholdt, S.M., Bergman,R. N., Barroso, I., Bandinelli, S., Ballantyne, C.M., Assimes, T. L., Querter-mous, T., Altshuler, D., Seielstad, M.,Wong, T. Y., Tai, E. S., Feranil, A. B.,Kuzawa, C. W., Adair, L. S., Taylor, H. A., Jr., Borecki, I. B., Gabriel, S. B.,Wilson, J. G., Holm, H., Thorsteinsdottir, U., Gudnason, V., Krauss, R.M.,Mohlke, K. L., Ordovas, J. M., Munroe, P. B., Kooner, J. S., Tall, A. R.,Hegele, R. A., Kastelein, J. J., Schadt, E. E., Rotter, J. I., Boerwinkle, E.,Strachan, D. P., Mooser, V., Stefansson, K., Reilly, M. P., Samani, N. J.,Schunkert, H., Cupples, L. A., Sandhu,M. S., Ridker, P.M., Rader, D. J., vanDuijn, C.M., Peltonen, L., Abecasis, G. R., Boehnke,M., and Kathiresan, S.(2010) Biological, clinical and population relevance of 95 loci for bloodlipids. Nature 466, 707–713
Mitochondrial Amidoxime Reductase Enzyme System
FEBRUARY 24, 2012 • VOLUME 287 • NUMBER 9 JOURNAL OF BIOLOGICAL CHEMISTRY 6317
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from

Inger Johansson and Magnus Ingelman-SundbergEtienne P. A. Neve, Åsa Nordling, Tommy B. Andersson, Ulf Hellman, Ulf Diczfalusy,
MOSC2 Is of Importance for Lipid Synthesis in Adipocyte Mitochondria Type B (CYB5B) and5bAmidoxime Reductase System Containing Cytochrome
doi: 10.1074/jbc.M111.328237 originally published online December 27, 20112012, 287:6307-6317.J. Biol. Chem.
10.1074/jbc.M111.328237Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2011/12/27/M111.328237.DC1
http://www.jbc.org/content/287/9/6307.full.html#ref-list-1
This article cites 40 references, 13 of which can be accessed free at
by guest on February 18, 2019http://w
ww
.jbc.org/D
ownloaded from