agi phd 3 printelni vegso x - sci.u-szeged.hu
TRANSCRIPT

Az idiopathiás scoliosis molekuláris biológiai vizsgálata
Ph.D. értekezés
Czibula Ágnes
Témavezetı: Dr. Raskó István
MTA Szegedi Biológiai Központ Genetikai Intézet
Biológiai Ph.D. Iskola, SZTE TTIK
SZEGED
2010

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
2
TARTALOMJEGYZÉK
RÖVIDÍTÉSEK JEGYZÉKE ............................................................................................................................. 4
IRODALMI ÁTTEKINTÉS ................................................................................................................................ 6
GENETIKAI MEGKÖZELÍTÉSEK AZ EMBERI BETEGSÉGEK ÖRÖKLETES TÉNYEZİINEK FELTÁRÁSÁRA .................... 6
AZ IDIOPATHIÁS SCOLIOSIS DEFINÍCIÓJA ............................................................................................................. 9
Az AIS klinikai tünetei .................................................................................................................................. 10
Cobb fok meghatározása .............................................................................................................................. 11
A gerincferdülés terápiája ........................................................................................................................... 12
A gerincferdülés epidemiológiája ................................................................................................................ 13
Az idiopathiás scoliosis etiológiája .............................................................................................................. 13
Strukturális eltérések ................................................................................................................................................. 14
Neurológia rendellenességek ..................................................................................................................................... 16
Növekedési és biomechanikai hatások ...................................................................................................................... 16
Vérlemezkék rendellenességei .................................................................................................................................. 17
Hormonok szerepe az AIS-ben .................................................................................................................................. 18
Genetikai tényezık .................................................................................................................................................... 19
Az etiológiai teóriák összegzése ................................................................................................................... 22
AZ ÉRTEKEZÉS SZEMPONTJÁBÓL KIEMELTEN FONTOS ETIOLÓGIAI FAKTOROK .................................................. 23
Leptin ........................................................................................................................................................... 23
Leptin szerepe a csonttömeg szabályozásban ............................................................................................................ 24
Leptinszint nemek közötti különbsége ...................................................................................................................... 26
Polimorfikus helyek vizsgálat a LEP gén szabályozó régiójában .............................................................................. 26
Leptin és az AIS ........................................................................................................................................................ 28
Az interleukin 6 ............................................................................................................................................ 28
Az IL6 szerepe a csontátépítésben............................................................................................................................. 29
Polimorfikus helyek vizsgálat az IL6 gén szabályozó régiójában ............................................................................. 30
Az IL6 és az AIS ....................................................................................................................................................... 31
CÉLKITŐZÉSEK .............................................................................................................................................. 33
ANYAGOK ÉS MÓDSZEREK......................................................................................................................... 34
KÍSÉRLETBE BEVONT SZEMÉLYEK ADATAI ÉS MINTÁI ....................................................................................... 34
DNS ALAPÚ MUNKAFOLYAMATOK ................................................................................................................... 35
Genomikus DNS tisztítása ............................................................................................................................ 35
SNP-k kimutatása PCR-RFLP módszerrel ................................................................................................... 35
RNS ALAPÚ MUNKAFOLYAMATOK ................................................................................................................... 37
RNS izolálás ................................................................................................................................................. 37
Northern gél elektroforézis .......................................................................................................................... 37

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
3
RNS mintacsoportok tisztítása ..................................................................................................................... 38
Fluoreszcensen jelölt próba készítése a microarray hibridizáláshoz........................................................... 39
A cDNS és a 3DNA hibridizációja a CHIP-hez ........................................................................................... 40
A hibridizáció utáni kiértékelés .................................................................................................................... 41
Valós idejő kvantitatív polimeráz láncreakció (QRT-PCR) ......................................................................... 42
STATISZTIKAI ANALÍZIS .................................................................................................................................... 43
A MUNKA SORÁN HASZNÁLT FONTOSABB ONLINE BIOINFORMATIKAI ESZKÖZÖK ÉS ADATBÁZISOK INTERNET
CÍMEI: ............................................................................................................................................................... 44
EREDMÉNYEK ................................................................................................................................................. 45
PARAVERTEBRÁLIS IZMOK RNS KIFEJEZİDÉSI MINTÁZATÁNAK VIZSGÁLATA CHIP MICROARRAY-VEL .......... 45
EGÉSZSÉGES ÉS SCOLIOSISOS BETEGEK IZOMBIOPSZIÁJÁNAK GÉNKIFEJEZİDÉSBELI ÖSSZEHASONLÍTÁSA ........ 45
SCOLIOSISOS BETEGEKBEN A GERINCOSZLOP KÉT OLDALÁRÓL SZÁRMAZÓ IZOMBIOPSZIÁK
GÉNKIFEJEZİDÉSBELI ÖSSZEHASONLÍTÁSA ....................................................................................................... 49
PARAVERTEBRÁLIS IZMOKBAN KIFEJEZİDİ MRNS-EK VIZSGÁLATA QRT-PCR-REL ....................................... 53
POLIMORFIKUS HELYEK VIZSGÁLATA ............................................................................................................... 55
Leptin promóter G-2548A polimorfizmusa .................................................................................................. 55
IL6 promóter C-174G polimorfizmusa ........................................................................................................ 57
A két polimorfizmus IS kialakulására hajlamosító együttes hatásának vizsgálata ...................................... 58
A polimorfizmusok hatása az IS súlyosbodására ......................................................................................... 60
EREDMÉNYEK MEGVITATÁSA .................................................................................................................. 61
IRODALOMJEGYZÉK .................................................................................................................................... 67
KÖSZÖNETNYILVÁNÍTÁS ............................................................................................................................ 78
ÖSSZEFOGLALÁS ........................................................................................................................................... 79
SUMMARY ......................................................................................................................................................... 83
KÖZLEMÉNYEK LISTÁJA ............................................................................................................................ 87

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
4
RÖVIDÍTÉSEK JEGYZÉKE
ACAN agrecan AIS adolescens idiopathiás scoliosis, serdülıkori, ismert ok nékül kialakuló
gerincferdülés BMP4 bone morphogenetic protein 4 C/EBP CCAAT/enhancer binding protein CART cocaine- and amphetamine-regulated transcript CDK5 cyclin-dependent kinase 5 CHD7 chromodomain helicase DNA binding protein 7 CHRDL1 chordin-like 1 CI confidence interval, megbízhatósági mérettartomány COL collagen DEPC diethylpyrocarbonate DST dystonin EDTA ethylenediaminetetraacetate EEG elektro-enkefalográf ER estrogen receptor EST expressed sequence tag FBN fibrillin FGF fibroblast growth factor FOXO 1 forkhead box O1 GDF growth differentiation factor GHR growth hormone receptor GLM Generalizált Lineáris Model GO génontológia GPC glypican HIF-1 hypoxia inducible factor 1 HOX7 msh homeobox 1 HPRT hypoxanthine phosphoribosyltransferase 1 HS3ST3 heparan sulfate (glucosamine) 3-O-sulfotransferase, HWE Hardy-Weinberg egyensúly IGF insulin-like growth factor receptor IL4R interleukin 4 receptor IL6 interleukin-6 IRX iroquois homeobox IS idiopathiás scoliosis, ismert ok nékül kialakuló gerincferdülés JAK1 Janus kinase 1 KCNA potassium voltage-gated channel, shaker-related subfamily LD linkage disequilibrium , kapcsoltsági kiegyenlítetlenség LEP leptin LMNA lamin A MATN matrilin MDR Multifaktoros Dimenzionalitás Redukció

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
5
MFAP microfibrillar associated protein ML maximum likelihood MOPS 3-(N-morpholino)propanesulfonic acid MTNR melatonin receptor NLRP12 NLR family, pyrin domain containing 12 NOS nitric oxide synthase NP nucleus pulposus NTF neurotrophin OMIM Online Mendelian Inheritance in Man OR odds ratio, esélyhányados PCR polymerase chain reaction, polimeráz láncreakció PCR-RFLP polymerase chain reaction based restriction fragment length polymorphism POSTN periostin PRKAG2 protein kinase, AMP-activated, gamma 2 non-catalytic subunit alfa 1 QRT-PCR quantitative real time polymerase chain reaction, valós idejő kvantitatív
polimeráz láncreakció RANK NF-κβ activator RANKL NF-κβ receptor activator ligand SAGE Serial Analysis of Gene Expression SDS Sodium dodecyl sulfate SH3GL1 SH3-domain GRB2-like 1 SLC2A1 solute carrier family 2 (facilitated glucose transporter), member 1 SNP single nucletide polimorphism, egypontos nukleotid-polimorfizmus SNS sympathetic nervous system SNTG syntrophin SRS Scoliosis Research Society SSC saline-sodium citrate TNS1 tensin 1 UTR untranslated region, nem transzlálódó régió VDR vitamin D receptor

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
6
IRODALMI ÁTTEKINTÉS
Az orvostudomány egyik nagy és napjainkban egyre nagyobb jelentıségő feladata a
betegségek hátterében álló örökletes tényezık felderítése. A tudás birtokában lehetıvé válik a
betegség kialakításáért felelıs molekuláris mechanizmusok megismerése és ezáltal célzott
terápia kialakítása. Természetesen nagyon hosszú az út a labortól a betegágyig; több
tudományterületen dolgozó sok kutató és sok orvos munkájára van szükség ahhoz, hogy egy
terápia vagy gyógyszer a betegségben szenvedık gyógyulását jelenthesse. A személyre
szabott orvosláshoz az elsı lépés a betegség pathomechanizmusának feltárása és ebben
sarkalatos pont a betegséget okozó, vagy az arra hajlamosító gének leírása.
Genetikai megközelítések az emberi betegségek örökletes tényezıinek
feltárására
A humángenetika alapjait jelentı öröklıdési mechanizmusok egy részét már jóval a
gének természetének megismerése elıtt leírták. 1866-ban Gregor Mendel által publikált
öröklıdési törvények az emberi betegségek egy részének az öröklıdésére is igazak; a mai
napig az egyetlen gén hibájából kialakuló betegségeket mendeli öröklıdéső betegségeknek
nevezik a szakirodalomban. 1911-ben kezdıdött meg az emberi kapcsoltsági csoportok
leírása annak a megfigyelésével, hogy színtévesztés ugyanúgy öröklıdik, mint az X
kromoszóma. Az 1970-es években kifejlesztették a kromoszóma festés módszerét, ami
lehetıvé tette az autoszomális kromoszómán elhelyezkedı gének öröklıdésének követését.
Ugyanebben az évtizedben klónozták az elsı humán gént, a β-globint, és kimutatták, hogy a
β-globin lánc 6. aminosavának cseréje okozza a sarlóssejtes vérszegénységet. Ez volt az elsı
humán betegség, aminek meghatározták a genetikai okát. 1976-ban pedig már rendelkezésre
álltak azok a DNS próbák, melyekkel prenatális diagnosztikát lehetett végezni a
vérszegénység kimutatására. 1994-ben 600, 2001-ben 1112, napjainkban pedig az OMIM
adatbázis 2010. márciusi adata alapján, 3063 mendeli módon öröklıdı betegség génje ismert
(Peltonen and McKusick, 2001; Rasko, 1994).
Az új évszázadban a monogénes betegségek kutatásáról a bonyolultabb, nem mendeli
öröklıdésmenető betegségek felé fordult a figyelem, amiket komplex betegségeknek

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
7
neveznek, mivel több gén a környezeti hatásokkal kölcsönhatásban alakítja ki ıket. A
komplex betegségek a társadalom nagyobb részét érintik, mint a ritkán elıforduló monogénes
betegségek; ide tartoznak többek között a szív- és érrendszeri betegségek, a cukorbetegség és
a pszichiátriai betegségek nagy része.
A betegségeket okozó gének felderítésére alapvetıen két módszert használnak: a
kapcsoltsági analízist és az asszociációs vizsgálatokat. A kettı között a lényegi különbség az,
hogy a kapcsoltsági analízis a családfa elemzésekkel lokuszokat kapcsol a betegséghez, míg
az asszociációs vizsgálatokban populációk összehasonlításával egy gén valamelyik allélját
kötik az adott betegségekhez. Mindkét analízis használható mind a „reverz” mind „forward”
genetikai megközelítéssel. A „reverz” genetikai megközelítés szerint véletlenszerő, anonim
markerekkel végzik a vizsgálatot és utána keresik meg a jelölt markerrel fizikai kapcsolatban
lévı, betegséget okozó lokuszt, míg a „forward” genetikával a betegséggel valamilyen ok
alapján kapcsolatba hozható gén (kandidáns gén) polimorfizmusát vizsgálják.
A kapcsoltsági analízist a mendeli öröklıdéső betegségek vizsgálatára használták nagy
sikerrel, komplex betegségek esetén kevésbé alkalmazható ez a módszer, mivel csak a nagy
hatású géneket tudták vele azonosítani. Ennek oka a komplex betegségek jellegzetességeiben
rejlik. Elıször is a betegség fenotípusos megjelenése, kialakulásának idıpontja, súlyossága
nagyon eltérı, ami mögött a hasonló tüneteket okozó, de molekuláris folyamataiban
különbözı pathomechamizmus állhat. Másrészrıl a komplex betegségeket nem egy, hanem
több, alacsony penetranciájú, viszonylag kis kockázatú gén okozza, ráadásul a fenotípus
kialakulását még környezeti tényezık is befolyásolják.
Az asszociációs vizsgálatok használata a humángenetikusok leggyakrabban alkalmazott
módszerévé vált, amit a Human Genom Program befejezıdése után beindult HapMap projekt
adatainak felhasználása tesz alkalmassá a komplex betegségek vizsgálatára. A kapcsoltsági
kiegyenlítetlenségen alapuló térképezés (linkage disequilibrium maping, LD) az asszociációs
vizsgálatok „reverz” genetikai megközelítése. Azon a feltételezésen alapul, hogy betegségre
hajlamosító allél a közvetlen szomszédságában lévı polimorfikus helyekkel együtt öröklıdik,
míg a tıle távolabb lévı polimorfizmusok a betegséget okozó mutáció kialakulása óta eltelt
idıben, a rekombináció miatt, kapcsoltsági egyensúlyba kerülnek. A kapcsoltsági
kiegyenlítettlenséget statisztikai eszközökkel mutatják ki. A polimorfizmusok közül az
asszociációs vizsgálatra a legalkalmasabbak az egypontos nukleotid-polimorfizmusok (SNP),
melyek az emberi genetikai variációk 90%-át jelentik és átlagosan minden 300 bp-nyi DNS
szakaszon egy elıfordul belılük. Az LD térképezés alkalmas a kandidáns gén kiválasztására,

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
8
amit aztán további asszociációs vizsgálatokban az immár a „forward” genetikai megközelítés
szerint vizsgálhatnak.
A komplex betegségek epidemiológiai kutatásaiban a patológiás eltéréseket kísérı
génexpressziós változások vizsgálata a technikai tudásunk fejlıdése után válhatott a jelölt
gének kiválasztásának új módszerévé napjainkban. A századforduló környékén a
hibridizáláson és a szekvenáláson alapuló génexpressziós vizsgálatok vonatkozásában is átütı
technológiai újítások történtek. Egyrészrıl a DNS microarray vagy DNS chip technika
elterjedése, amely során egyetlen hibridizálási lépésben egy tárgylemeznyi területen több
tízezer mRNS analizálható egyszerre (1. ábra) (DeRisi et al., 1996), másrészrıl a
szekvenáláson alapuló nagy kapacitású módszerek kifejlesztése (Serial Analysis of Gene
Expression-SAGE, Next Generation Sequencing) adtak lehetıséget a transzkriptoma szintjén
történı változások detektálására. Bioinformatikai adatfeldolgozás és körültekintı statisztikai
elemzések után lehet olyan géneket kiválasztani, amelyek további vizsgálatokra érdemesek.
A kandidáns gének alléljainak elıfordulási gyakoriságát a beteg és az egészséges
kontroll populációban az ún. eset-kontroll tanulmányokban hasonlítják össze. Amennyiben
valamely allél elıfordulási gyakorisága szignifikánsan magasabb a beteg populációban, akkor
az adott gén, adott allélját a betegségre hajlamosítónak tekintik.
1. ábra cDNS-chipek alkalmazása génexpressziós mintázatok nyomonkövetésére
Jelen dolgozat egy komplex betegség, az idiopathiás scoliosis genetikai hátterét kutatja
a DNS chip technikán alapuló kandidáns gén kiválasztása után, „forward” genetikai
megközelítéssel, eset-kontroll asszociációs tanulmányban.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
9
Az idiopathiás scoliosis definíciója
A gerincoszlop betegségei a két lábon járás óta érintik az emberiséget. A gerinc
oldalirányú elhajlását Hippocrates írja le elıször, melyet izomproblémának tekint. Celsus -a
Krisztus utáni I. században- a gerincdeformitások kezelésére a tornáztatást javasolja. Az
orvostudományban a gerincnek a tér mindhárom irányába történı elferdülésére Galenus a II.
században a scoliosis nevet adta, ami a görögül azt jelenti, hogy görbe, ferde. A XIX. század
közepén Bauer írja le elıször a betegség súlyos tünetekkel járó, fiatalkorban manifesztálódó
formáját, és idiopathiás scoliosisnak nevezi. 1966-ban megalakul a gerincferdülésekkel
foglalkozó orvosokat, kutatókat nemzetközi szinten összefogó szervezet, a Scoliosis Research
Society (SRS) és definíciója alapján az idiopathiás scoliosis (IS) a gerinc ismeretlen okból
létrejövı strukturális laterális elhajlása minimum 10°-os Cobb szöggel (normális görbülettıl
való eltérés fokban kifejezve) és megegyezı csigolya rotációval.
Raffaello: A három grácia, (1498 körül). A középen álló grácián jobbra konvex
scoliosis látszik (Musée Condé, Chantilly)

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
10
A gerincferdülések egy része nem stukturális (tartási, kompenzációs scoliosis), vagy csak
átmenetileg (pl. gyulladás miatt) kialakuló strukturális gerincdeformitás, nagyobb részüket
azonban szerkezeti eltérések jellemzik, ezért strukturálisak. A strukturális scoliosisok egy
része veleszületett rendellenesség, más részük valamilyen más szindrómához társultan
jelentkezik (pl. neuromuszkuláris betegségekhez, kötıszöveti betegségekhez,
neurofibromatosishoz) és nagy részük kialakulásának az oka nem ismert.
A gerincferdülések 70-80%-át a strukturális idiopathiás scoliosis adja, melynek a megjelenés
ideje alapján három különbözı típusa különíthetı el: infantilis (0-3 életév), juvenilis (4-9 év),
adolescens (10-18 év). A zárójelben lévı évszámok a betegség kezdetét jelzik, de a
csontosodás állapotától függıen kissé eltérhetnek a megadott értéktıl.
Vizsgálataink során a leggyakrabban elıforduló, és ezért a kutatások homlokterében
álló adolescens idiopathiás scoliosisra (AIS) fókuszáltunk.
Az AIS klinikai tünetei
Gyermekkorban a deformitás az egyetlen tünet, ekkor fájdalom
még nem jelentkezik. Aszimmetrikus a nyak-váll vonal, a
lapockák vonala, a bordák és a csípılapátok elhelyezkedése. A
konvex oldalon a nyak-váll vonal és a lapocka magasabban áll,
utóbbi a törzstıl elemelkedik.
2. ábra Scoliosisos nıbeteg

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
11
A deformitás a gerinc több régióját is érintheti. A háti görbület a leggyakrabban elıforduló,
míg a nyaki vagy a medencei régió ritkán deformálódik. Az esetek 90%-ban a jobbra konvex
dorzális (háti) scoliosis fordul elı (2. ábra).
Cobb fok meghatározása
A klinikai vizsgálatok során nagyon fontos a mellkas röntgen
felvétele és az alapján az ún. Cobb fok meghatározása. A Cobb˚
a gerinc görbületének mértékét adja meg és fontos a további
diagnosztikában, illetve a megfelelı terápia kiválasztásában.
A mellkas anteroposterior felvételén láthatóvá válik a frontális
síkú elhajlás mértéke. A Cobb˚meghatározásakor az alsó és felsı
végcsigolyák görbület csúcsától távolabb esı zárólemezével
párhuzamos vonalra egy-egy merılegest húzunk, és az általuk
bezárt szög adja a görbület mértékét (3. ábra).
Nagyon fontos a görbület mértékének meghatározása a scoliosis megjelenésekor, mert a
betegség mindaddig progrediálhat, amíg a csontosodás be nem fejezıdik, tehát minél
fiatalabb a gyerek és minél nagyobb a növekedési potenciálja, valamint minél súlyosabb a
deformitás az észlelés pillanatában, annál nagyobb eséllyel fog tovább romlani a görbület
mértéke.
A korai iskolai szőréseken az Adam tesztet használják a gerincferdülés korai stádiumban
történı felfedezéséhez, melynek során a vizsgáló iskolaorvos megkéri a gyermeket, hogy
törzsben hajoljon elıre, térdben ne hajlítson, a karjait engedje szabadon lógni, és
megvizsgálja, hogy nincs e borda- vagy paravertebrális izom elıemelkedés. Hazánkban
évente mintegy 450 új esettel kell számolni, melyek közül 150 gyermeknek van szüksége
orthopédiai kezelésre.
3. ábra A röntgen felvételen
egy 62 Cobb˚-os scoliosisos
gerincoszlop látszik

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
12
A gerincferdülés terápiája
A scoliosisos beteg kezelését befolyásolja a beteg
életkora -biológiai és csontkor-, a görbület típusa és
lokalizációja.
A konzervatív kezelés célja a progresszió lassítása.
Ebbıl a célból gyógytorna, ill. főzı viselése javasolt.
Speciális gyógytorna elsajátítása gyógytornász
segítségével, melyet a beteg késıbb otthonában
végezhet, lehetıleg mindennap. Ajánlott kiegészítı
testmozgás az úszás, mely a gerincoszlop csökkent
gravitációs terhelése mellett mindkét testfél izomzatát
egyformán terheli. Az iskolai tornaóra alól csak a
súlyosabb esetekben szükséges felmentés, enyhébb
formáknál csak a kritikus feladatokat kell mellızni. Ma
már csak az esztétikailag elfogadható alacsony profilú
főzıket alkalmazzák. Főzı viselése javasolt 15-40
Cobb˚ között, és ha a nagyfokú görbület ellenére a
mőtét valamely okból ellenzett.
Mőtéti kezelés szükséges, ha az ágyéki görbület meghaladja a 30 Cobb fokot, vagy a háti
görbület a 40 Cobb fokot. Hatalmas elırelépést jelentett, és alapvetıen új lehetıségeket
teremtett a gerincdeformitások kezelésében Yves Cotrel és Jean Dubousset munkássága, akik
1983-ban új megközelítéső multiszegmentális rögzítést biztosító korrekciós eszközt vezettek
be a kezelésbe. A mőtéti technika forradalmasította a gerinc deformitások sebészetét. A
technika alapja, hogy a deformált gerincet több helyen horgokkal rögzítik a kívánt fiziológiás
görbületeknek megfelelıen meghajlított rúdhoz, majd a mőtıasztalon a rúd 90°-os
elforgatásával korrigálják az oldalirányú deformitást, így kialakulnak a fiziológiás
görbületek. Kb. 3 hónappal a mőtét után az elmerevített csontos gerincszakasz átveszi a
terhelést a fém rúdtól.
4. ábra Nicolas Andry ortopédiai
szakkönyv illusztrációja, Párizs 1741

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
13
A gerincferdülés epidemiológiája
A kismértékő (Cobb˚<10) gerincferdülés elıfordulási gyakorisága 1.5-3%, a 20 Cobb˚
értéket meghaladóé 0.3-0.5% és a nagymértékő (Cobb˚>30) gerincdeformitásé 0.2-0.3%. A
10-16 évesek körében a legmagasabb az idiopathiás scoliosis elıfordulási gyakorisága, a
gyerekek 2-4%-ánál figyelhetı meg. Az alacsony 10 Cobb˚ körüli deformitás esetén a fiúk és
a lányok aránya egyenlı, ám a 30 Cobb˚ feletti görbületeknél a lányok aránya tízszeres
(Roach, 1999). Magyarországon az 1970-es években Dr. Bellyei Árpád határozta meg az AIS
prevalenciáját, a 14 éven felüliekben 0.51-0.54 ±0.05%-ban fordult elı a gerincferdülés és a
10 Cobb˚ görbületet meghaladó idiopathiás scoliosisos betegek 0.29 ± 0.035%-ban voltak
jelen hazánkban (Bellyei et al., 1977). További vizsgálatok kimutatták, hogy a lányok aránya
3.4-szer magasabb a fiúkénál a 10 Cobb˚ görbületet meghaladó esetekben. A betegek
családját is megvizsgálva azt találták, hogy az elsıfokú rokonok 8.34±0.37%-ban érintettek,
másod és harmadfokú rokonok pedig 2.0±0.17% illetve 0.87±0.07%-ban. Amennyiben
mindkét szülı érintett volt, akkor a gyerekek 40%-ánál, ha csak az egyikük volt az, akkor
pedig 29.3 %-uknál jelent meg az AIS (Czeizel et al., 1978).
Az ikertanulmányok eredményei azt mutatták, hogy a egypetéjő ikrek között 73%, a
kétpetéjőek között 36%-os az együttes elıfordulás (konkordancia) (Kesling and Reinker,
1997).
Az idiopathiás scoliosis etiológiája
Az idiopathiás scoliosis etiológiájának tisztázására az elmúlt 40 év alatt sok próbálkozás
történt, számos területen keresték a deformitás kialakulásának okát. A fenotípusos megjelenés
is heterogén, eltérı például a kialakulás ideje, a progresszió mértéke és sebessége,
valószínőleg több különbözı patomechanizmus is eredményezheti ugyanazt a klinikai képet.
Világossá vált, hogy egy komplex és valószínőleg multifaktoriális folyamatról van szó. A
betegség kialakulásának okai között a genetikai faktor jelenléte általánosan elfogadott, bár az
öröklıdés módja kétséges.
Mivel az esetek döntı többségében a gerincferdülés kialakulása a serdülıkorra tehetı, a
növekedéssel kapcsolatos hormonális, metabolikus és biomechanikai változások is szerepet
játszhatnak az etiopathogenézisben, különösen a lányoknál a rövidebb idı alatt és gyorsabban
lezajló fejlıdés miatt. Az etiológiai kutatások az alábbi területen folynak:

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
14
1) Strukturális eltérések
A. Csontszöveti eltérések
B. Kötıszöveti eltérések
C. Harántcsíkolt izom szerkezeti eltérései
2) Neurológia rendellenességek
3) Növekedés és biomechanikai hatások
4) Vérlemezkék eltérései
5) Hormon háztartás
6) Genetikai tényezık
Strukturális eltérések
A. Csontszövet rendellenességek
Elıször 1982-ben Burner munkacsoportja talált összefüggést a csökkent csontsőrőség és az
idiopathiás scoliosis között és azóta több kutatócsoport is végzett hasonló méréseket (Burner,
III et al., 1982). A csontsőrőség mérési adatok összességében azt mutatják, hogy csont
ásványsőrősége csökkent az IS-es betegekben a korban egyeztetett kontroll populációhoz
képest, de nem bizonyítja, hogy a gyengébb csontminıség a kiváltó oka vagy okozata-e a
gerincferdülésnek (Li et al., 2008). Csigolya-tövisnyúlványból és a csípıcsontból származó
csontmintákon végzett szövettani kísérletekben azonban azt tapasztalták, hogy az IS
betegekben az oszteoklasztok száma csökkent, miáltal a csontátépítés dinamikája is csökkent.
Az abnormális csont metabolizmus megzavarhatja a növekedést így okozója lehet az AIS-es
betegek 20-38%-ában megfigyelt csontritkulásnak, ami akár elsıdleges kiváltó oka is lehet a
scoliosisnak (Cheng et al., 2001).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
15
B. Kötıszöveti rendellenességek
A biomechanikai egységként értelmezett gerincoszlop funkcionális stabilitásáért a
kollagénekbıl, proteoglikánokból, elasztikus rostokból és egyéb extracelluláris mátrix
elemekbıl álló kötıszöveti elemek is felelısek. Ebbe a csoportba tartoznak a csigolyák
közötti porckorongok és a csigolyákat összefőzı hosszanti szalagpántok. Mivel több olyan
kötıszöveti rendellenesség (pl. Marfan szindróma) mellé is társulhat gerincferdülés,
feltételezhetı, hogy e szövetek eltérései az AIS kiváltó okaként is szerepelhetnek. Az AIS-es
betegek 82%-ában valóban találtak eltéréseket a ligamentum flavum fibrillin
rostszerkezetében, azonban nem igazolt, hogy az idiopathiás scoliosisos betegeknél
generalizált kötıszöveti rendellenesség állna fent (Kouwenhoven and Castelein, 2008). A
porckorongokat illetıen is számos tanulmány jelent meg , ám eredményeik nem egyöntetőek.
Carl Nicoladoni német sebész professzor már 1904-ben felfigyelt arra, hogy a porckorong
alakja a scoliosis korai fázisában, még a csigolyák alakjának változása elıtt ék alakúvá válik.
Többen leírták, hogy a ferde gerincbıl származó porckorong kocsonyamagja a görbület
konvex oldalára tolódik és belsı szerkezete is megváltozik, valamint proteoglikán és kollagén
tartalma is eltérıvé válik (Pedrini et al., 1973; Stokes and Aronsson, 2001). Egyes
kutatócsoportok ezeket az eredményeket vitatják, mások megerısítik, de az általánosan
elfogadott vélemény szerint a porckorongokban megfigyelt eltérések inkább a görbült
gerincben fellépı egyenetlen terhelés következményei és nem a gerincferdülés
kialakulásának okai (Oegema, Jr. et al., 1983; Roberts et al., 1993; Cheung et al., 2008).
C. Harántcsíkolt izom eltérések
Mivel a paravertebrális izmok ereje tartja a gerincoszlopot függıleges állapotban és több,
az izmok felépítését és az izomfunkciót érintı betegségben másodlagosan kialakul a
gerincferdülés, igen korán, már a XIX. században felmerült a kérdés, hogy neuromuszkuláris
elégtelenség okozza-e a gerincdeformitást (Kouwenhoven and Castelein, 2008). Hisztokémiai
vizsgálatok alapján Spencer és Eccles írta le elıször, hogy az IS-es betegek paravertebrális
izmaiban a gerincoszlop két oldalán eltérı a különbözı izomrostok (I-es típusú-lassú és II-es
típusú- gyors) aránya, oly módon, hogy a II-típusú izomrostok száma lecsökkent a konvex
oldalon (Spencer and Eccles, 1976). További megfigyelések igazolták, hogy nem csak a
mélyizmokban, hanem a felszíni izmokban is, a konvex oldalon az I-es típusú izomrostok
aránya magasabb a konvex oldalon, így a konvex oldalon megnövekedett izomaktivitás

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
16
szerepet játszhat a gerincferdülés kialakításában. Az elmúlt évek során elektromyográfiás
aktivitásmérésekkel is alátámasztották a konvex oldali dominanciát és kimutatták, hogy a
görbület progresszivitása is összefügg az izomaktivitással (Cheung et al., 2005a). Az
izomrendellenességek gerincferdülés kialakulásában játszott esetleges szerepe még távolról
sem tisztázott és annak ellenére, hogy a legtöbb kutató úgy gondolja, hogy az izomerı
egyenetlenség a görbült gerinc kompenzálása miatt alakult ki és következménye nem pedig
oka a gerincferdülésnek, további vizsgálatok szükségesek a kérdés megnyugtató lezárásához.
Neurológia rendellenességek
A gerincferdülések egy része neuromuszkuláris rendellenesség fennállása során fejlıdik
ki a szindrómás scoliosisos betegekben, ami felveti az esetleges idegrendszeri hibát az AIS
vonatkozásában is. A mai napig nem tisztázott, hogy az AIS-sel együtt megjelenı neurológiai
rendellenességek szerepet játszanak-e az idiopathiás scoliosis kialakulásában. Az IS-es
gyerekekben enyhe eltérések mutathatók ki az EEG aktivitásban, a testhelyzet uralásában
valamint a szomatoszenzoros neurológiai funkciókban (Kouwenhoven and Castelein, 2008).
Legújabb kutatások szerint, amelyekben 3 dimenziós mágneses rezonancia képalkotó
eljárások segítségével 99 neuroanatómiai agyterület nagyságát hasonlították össze 20 AIS-es
és 26 korban, nemben egyeztetett kontroll személy analízisével, a vizsgált agyterületek
mintegy negyedénél eltéréseket találtak a két csoport között (Liu et al., 2008). A
gerincferdülés kialakulásának lehetséges okát a Roth, illetve Porter által megalkotott
koncepció szerint, a gerincoszlop és a gerincvelı növekedési ütemének eltérése jelenti; ezek
nincsenek egymással szinkronban és a rövidebb velı magával húzza az egyébként normális
morfológiájú csontos szerkezetet, ezáltal alakítva ki gerincdeformitást (Roth, 1968; Porter,
2001a).
Növekedési és biomechanikai hatások
Az AIS kialakulása és progressziója egyértelmően a serdülıkorhoz köthetı, amikor a
növekedés a legintenzívebb. A lányokban a felgyorsult gerincnövekedés egy évvel korábban
kezdıdik, mint a fiúknál és gerincferdülésük is hamarabb alakul ki, a gerincferdülés
progressziója pedig gyorsabb és nagyobb fokú. Azt is megfigyelték, hogy a nagyobb
görbülettel rendelkezı gyermekek magasabbak mind az egészséges, mind a kisebb
görbülettel bíró társaiknál (Shohat et al., 1988). Ezekbıl a megfigyelésekbıl következıen a
gerincferdülést kiváltó okot eltérıen ítélik meg; Gou és mtsai a lányok csigolyáinak

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
17
karcsúságában és nagyobb magasságában látja az okot, Lowe szerint a gerincoszlop elülsı
részeinek a hátsó struktúrákhoz képest történı relatív túlnövekvése eredményezi a
gerincoszlop kezdeti elhajlását, Sevastin pedig a bordák aszimmetrikus növekedését tartja
fontosnak (Guo et al., 2003; Lowe et al., 2000; Sevastik et al., 2003). Mindezek az egyes
csigolyákra ható erık egyenetlen eloszlását okozzák. Mivel a gerincoszlopban a csigolyák
szorosan össze vannak kötve, a felsı csigolyákra ható hajlító erık az alsóbb részeken a
gerincoszlop oldalirányú elhajlását eredményezik a biomechanikai modell szerint (Stokes and
Laible, 1990).
Vérlemezkék rendellenességei
A könnyen izolálható vérlemezkék vonzó tárgyai a scoliosis kutatásának, mivel
kontraktilis rendszerük nagymértékben hasonlít a harántcsíkolt izmokéhoz, így egy, a
vázizmokat érintı rendellenesség kitőnıen vizsgálható bennük. Ezen felül további nagy
elınyük, hogy a gerincferdülés okozta másodlagos hatások nem károsítják ezt a rendszert.
Több tanulmány is megjelent, melyben az IS-es betegek vérlemezkéinek abnormális
szerkezetét és megváltozott funkcionális mőködését írták le, és az eltérések nagy része a
megváltozott Ca2+ transzportra utalt. A kalmodulin, amely egy Ca2+-függı receptorfehérje az
izomsejtekben és a trombocitákban kölcsönhat az aktinnal és a miozinnal és hatására megnı a
szarkoplazmatikus retikulumból történı Ca2+ kiáramlás. Kindsfater és mtsai megemelkedett
kalmodulin szintet mértek azon IS-es betegek vérlemezkéikben, ahol a görbület progresszív
volt (Kindsfater et al., 1994). A kalmodulin, mint neurotranszmitter a melatonin szekréció
szabályozásában is szerepet játszik, így a megemelkedett kalmodulin aktivitás a csökkent
melatonin termelıdéssel összefüggésben lehet (Machida et al., 1996).
Acaroglu és mtsai a kalmodulin és a melatonin szintjét is megvizsgálták mind a
trombocitákban, mind a paravertebrális izmokban az egészséges, trauma miatt operált
esetekben és a gerincferdülés korrigálása miatt mőtött IS-es betegekben is. Nem találtak
különbséget a melatonin szintben sem a beteg-kontroll viszonylatban sem pedig a jobb vagy
bal oldalról származó izombiopsziák között. Az IS-es betegeknél azonban a kalmodulin szint
magasabb volt a gerincoszlop konvex oldaláról származó izombiopsziában (Acaroglu et al.,
2009). Az eddigi kísérleti eredmények ugyan még nem elegendıek annak pontos eldöntésére,
hogy mi a szerepe a kalmodulinnak az IS etiológiájában, mégis úgy tőnik a Ca2+-függı
kalmodulin köthetı az IS súlyosbodásához.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
18
Hormonok szerepe az AIS-ben
Az a tény, hogy az AIS kialakulása és progressziója a serdülıkori gyors növekedési
szakaszhoz társul, felveti a növekedés komplex folyamatában résztvevı hormonok szerepét.
Feltételezték a növekedési hormon és a IGF-1 szerepét, ám a vérszérum szintben nem találtak
eltérést az IS-es betegeknél (Misol et al., 1971). Mivel a nagyobb Cobb fokú görbülettel bíró
eseteknél a lányok aránya magasabb, Esposito és mtsai a nıi nemi hormonok esetleges
szerepét vizsgálták és azt találták, hogy mind az ösztrogén, mind a progeszteron
vérszérumbeli szintje csökkent volt az IS-es lányokban az egészséges, korban egyezı
társaikéhoz képest (Esposito et al. 56-60). A serdülı korú lányok antropometriás vizsgálata
alapján az IS-es lányok magasabbaknak és vékonyabbaknak bizonyultak, mint a kontrollok,
az alacsonyabb testsúlyuk miatt egy, a zsírszövet által termelt hormon, a leptin szerepe is
felmerülhet a patogenezisben (Lee et al. 1024-35;Siu King et al. 2152-57). Mivel a dolgozat
egyik célja, ennek az etiológiai elemnek a vizsgálata, részletesebben egy késıbbi, külön
fejezetben fogom tárgyalni a leptin és az AIS kapcsolatát.
A tobozmirigy által termelt hormon, a melatonin etiológiai tényezıként azután került az
érdeklıdés középpontjába, miután 1959-ben Thillard munkacsoportja arról számolt be, hogy
a csirkében (amit a gerincferdülés modellállatának tartanak), a tobozmirigy kimetszése
(pinealectomia) után rendszeresen kialakul a scoliosis. Megfigyelésüket késıbb többen
megerısítették a csirkékkel és a két lábon járásra kényszerített patkányokkal végzett
kísérletekben is (THILLARD, 1959; Machida et al., 1999; Machida et al., 1993). Az újabb
kutatások szerint azonban pinealectomia hatására nem alakult ki gerincdeformitás az
emberhez evolúciósan közelebb álló, kétlábon járó rhesus majmokban (Cheung et al., 2005b).
Dubousset és Machida 30 IS-os serdülı betegben kimutatta, hogy a progresszív formákban
(melyekben az utóbbi évben több, mint 10 fokot nıtt a Cobb szög mértéke) az éjszakai
szérum melatonin szint 35%-kal csökkent a stabil, nem progrediáló gerincferdüléses
betegekkel és 50 kontroll egyénnel összevetve, ám ezt az eredményt több ezt követı
tanulmányban sem sikerült megerısíteni (Machida et al., 1996; Bagnall et al., 1996; Fagan et
al., 1998).
A fentiek alapján a melatonin szerepe az IS kialakításában, vagy a gerincferdülés
súlyosbodásában nem meggyızıen bizonyított, viszont a melatoninnal kapcsolatos kutatások
napjainkban újra az érdeklıdés középpontjába kerültek a melatonin csontnövekedés
szabályozásában játszott szerepe miatt (Roth et al., 1999; Satomura et al., 2007). In vitro
kísérletekben AIS betegekbıl származó primer oszteoblaszt sejtvonalakban a melatonin

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
19
szignáltranszdukciós útvonal nem megfelelı mőködését mutatta ki Moreau munkacsoportja
(Moreau et al., 2004). További megerısítést jelenthetnek a genetikai asszociációs vizsgálatok
is, amit a következı fejezetben írok le.
Genetikai tényezık
A genetikai tényezık szerepére kevés figyelem irányult az idiopathiás scoliosis
kutatásának kezdetén, mivel a betegség többnyire szórványosan lépett fel, és gyakran a
családon belül halmozódó esetek sem mutattak egyértelmő öröklıdésmenetet. A genetikai
háttér kutatása a hetvenes évek óta folyik, s mára már a legfontosabb és leginkább kutatott
területté vált.
Az epidemiológiai részben leírt elıfordulási gyakoriságok, különösen a betegek
családjában elıforduló magasabb incidencia és az ikerkutatások során kapott konkordancia
adatok arra utaltak, hogy létezik örökletes faktor. A külföldi populációs tanulmányok az elsı,
másod és harmadfokú rokonok között rendre 11%-os, 2.4%-os és 1.4%-os érintettséget
találtak, ami jó egyezést mutat a hazai adatokkal (Riseborough and Wynne-Davies, 1973).
Bellyeiék a magyar populációs adatokból poligénes, multifaktoriális öröklıdésmenetre
következtettek, amit megerısíteni látszik Wyne-Davies és Risenborough közös tanulmánya,
melyet 2869 önként jelentkezın, mindegyikük elsıfokú rokonain illetve 81%-uk másod és
harmadfokú rokonain végeztek (Czeizel et al., 1978; Riseborough and Wynne-Davies, 1973).
Wynne-Davies 2000 önként jelentkezın és elsıfokú rokonain végzett radiográfiás
vizsgálatokat, eredményeik domináns öröklıdésmenetre engedtek következtetni (Wynne-
Davies, 1968). Aksenovich komplex szegregációanalízissel 90 család 283 tagját vizsgálta és
eredményeik alapján egy autoszomális domináns, de nem teljes penetranciával öröklıdı
fontos gén jelenlétét valószínősítik, legalábbis a súlyosabb idiopáthiás scoliosisos esetekben
(Aksenovich et al., 1988). Cowell és mtsai 17 családot megvizsgálva nem találtak apáról-
fiúra történı öröklıdést, ezért X-hez kapcsolt, domináns öröklıdésmenetet véleményezetek
csakúgy, mint Miller és mtsai, akik szintén családfa analízissel jutottak ugyanerre a
következtetésre (Cowell et al., 1972; Justice et al., 2003).
Az etiológiai kutatások során feltárt eltérések alapján érintettnek gondolt gének
mutációinak célzott vizsgálatával kezdıdtek a molekuláris genetikai vizsgálatok. A
kötıszövet felépítésében fontos strukturális géneket, mint a kollagének (COL1A1, COL1A2,
COL2A1), fibrillinek (FBN1, FBN2), elasztin (ELN) és a heparán-szulfát glükózamin 3-O-
szulfotranszferázok (HS3ST3A1, HS3ST3B1), nem találták érintettnek az AIS-ben (Carr et

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
20
al., 1992; Miller et al., 1996; Salehi et al., 2002). A porcspecifikus mátrixmolekulák közül az
aggrekán (ACAN) nem mutatott kapcsoltságot az AIS-sel, míg a matrilin-1 (MATN1) gén
3’UTR régiójában elhelyezkedı mikroszatellita marker által jelölt, 103 bp-os allélje
asszociációt mutat az AIS-sel (Montanaro et al., 2006; Marosy et al., 2006). Mivel a
növekedés és a gerincferdülés között szoros kapcsolat áll fenn, a növekedési hormon receptor
(GHR) alléleloszlását is vizsgálták az AIS betegekben 5 SNP alapján, de nem találtak eltérést
a genotípus gyakoriságban a kontrollhoz viszonyítva (Qiu et al., 2007b). Hasonló kísérleti
elrendezéső vizsgálatban az inzulinszerő növekedési faktor (IGF1) promóterének SNP
analízise során sem találtak különbséget a genotípus eloszlásban, de az egyik homozigóta
genotípushoz magasabb Cobb˚ tartozott, ami alapján a szerzık az IGF1-et az AIS-t módosító,
progressziós faktornak tekintik az IGF1-et (Yeung et al., 2006). Az ösztrogén receptor β gén
(ERβ) polimorfikus helyeinek vizsgálatakor találtak egy olyan SNP-t, aminek egyik allélje
gyakrabban fordul elı a scoliosisos lányokban, mint a kontrollokban, ezért ezt a gént az AIS-
re hajlamosítónak gondolják a lányok esetén (Zhang et al., 2009). A melatonin metabolizmus
zavarát feltételezik az AIS kialakulásának egyik lehetséges okaként, ezért kínai kutatók
megvizsgálták a melatonin receptorainak polimorfikus helyeit asszociációt keresve a
gerincferdüléssel. Az oszteoblasztokban is kifejezıdı melatonin 1B receptor (MTNR1B) gén
promóterében olyan SNP polimorfizmust írtak le a-1193 pozícióban (C-1193 T, rs4753426),
melynek C allélja asszociál a gerincferdüléssel és azon egyéneknél, akik homozigóta
formában hordozzák a C allélt a gerincferdülés kialakulásának kockázata magasabb, mint a T
allélt is hordozóké (OR:1.29) a kínai populációban (Qiu et al., 2007a). Saját vizsgálataink
nem támasztják alá ezt az eredményt, a genotípus gyakoriságok nem mutattak eltérést az AIS-
es beteg és a kontroll populáció közt (közlés alatt). Megvizsgálták a másik melatonin receptor
(MTNR1A) egyik promóter polimorfizmusát is, de asszociációt nem tudtak kimutatni (Qiu et
al., 2008).
Az asszociációs vizsgálatokkal párhuzamosan kapcsoltsági analízist is végeztek
számos laboratóriumban és az elmúlt tíz évben számos kromoszóma szakaszt azonosítottak
az AIS-sel összefüggésben. Mind Chan, és mtsai, mind Alden munkacsoportja, egymással
rokoni kapcsolatban nem álló családokat elemezve a 19p13.3 régióban az AIS-hoz kapcsolt
lokuszt írtak le (Alden et al., 2006; Chan et al., 2002). Ocaka és mtsai is megerısítették
Miller munkacsoportja által jelzett 9q31.1-q34.2 régió kapcsoltságát az AIS-hez (Ocaka et
al., 2008; Miller et al., 2005). Salehi és mtsai egy háromgenerációs olasz családot vizsgáltak,
ahol a 17p11 kromoszóma régiót találták kapcsoltnak 11 érintett személy kapcsán, amit
Miller munkacsoportja is igazolt egy 50 családos mintán (Salehi et al., 2002; Miller et al.,

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
21
2005). Az 1. táblázat tartalmazza a kapcsoltsági analízis eredményeként kapott kromoszóma
régiókat és az ott elhelyezkedı, a szerzık által az AIS-ben potenciálisan érintettnek jelölt
géneket.
A kapcsoltsági analízissel jelölt gének közül a γ1 syntrophint (SNTG1) és a
kromodomént tartalmazó helikáz, DNS-kötı fehérje 7-t (CHD7) vizsgálták részletesen. A
SNTG1 génjében talált mutációk csak a vizsgált AIS-es betegek kis százalékában voltak
megfigyelhetık és a családi halmozódást mutató esetekben sem szegregálódott együtt az AIS
az SNTG1 mutációival. (Bashiardes et al., 2004) A CHD7 gén esetén Wise munkacsoportja
egy potenciálisan funkcióval rendelkezı polimorfizmust (rs4738824) kapcsoltnak talált az
AIS-sel beteg+szülık hármasát vizsgálva (Gao et al., 2007).
A jelenleg elfogadott etiológiai modell alapján a betegség kialakulása két szakaszra
osztható, egy iniciációs és egy progressziós fázisra. Mindkét fázisban több gén is szerepet
játszhat és eddig ismeretlen környezeti faktorok hathatnak a fázisokra külön-külön is. A
modell szerint az AIS komplex genetikai betegség, ahol egy vagy több gén egymással és a
környezeti tényezıkkel együtthatva alakítja ki a gerincdeformitást (Cheung et al., 2008).
1. táblázat A kapcsoltsági analízisek eredményeként az AIS-sel kapcsolatba hozható
kromoszóma szakaszok és jelölt gének
résztvevık száma család/egyén
kromoszóma lokalizáció
jelölt gének referencia
1/14 6q
10q 18q
(Wise et al., 2000)
1/17 17p11 HS3ST3A1 HS3ST3B1
(Salehi et al., 2002)
7/52 19p13.3 NLRP12, SH3GL1
(Chan et al., 2002)
202/1198 Xq23 Xq26.1
CHRDL1 COL4A5, 6
(Justice et al., 2003)
202/1198
6p22-p25 6q13-q16 9q31.1-q34.2
16p13.13-q13.2 17p12-q11.2
(Miller et al., 2005)
202/1198
5p13 13q13.3 13q32
IRX1, IRX2, IRX4 POSTN,FOXO1 GPC5
(Miller et al., 2006)
202/1198 19p13 (Alden et al., 2006)
52/123 1p 8q
10q
- SNTG1, CHD7
- (Gao et al., 2007)
25/116 9q31.2-q34.2
17q25.3-qtel (Ocaka et al., 2008)
7/54 12p3 NTF3,FGF3,GDF3 KCNA1,5,6,MFAP5
(Raggio et al., 2009)

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
22
Az etiológiai teóriák összegzése
Az elızı fejezetekben felsorolt rendellenességek leírása után minden szerzı kialakította a
saját megfigyelése alapján a saját teóriáját, így mára közel 20 elmélet létezik az AIS
kórfejlıdésére.
A legfontosabbak listaszerő felsorolása:
1. Genetikai betegség multifaktoriális öröklıdéssel (Kouwenhoven and Castelein,
2008; Miller et al., 2005)
2. A gerincoszlop növekedésének aránytalansága, miszerint az anterior rész
gyorsabban nı (Guo et al., 2003)
3. A gerincoszlop és a gerincvelı növekedésének aszinkronja (Roth, 1968; Porter,
2001b)
4. Tartási rendellenesség, ami kialakulhat a hátsó agy diszfunkciója miatt,
neuromuszkuláris eltérések miatt, vagy az idegrendszer test-sémájának megkésett
fejlıdése miatt (Burwell et al., 2008a; Lao et al., 2008; Veldhuizen et al., 2000)
5. Melatonin hiány, vagy a melatonin szignálút nem megfelelı mőködése (Machida et
al., 1996)
6. A vérlemezkékben talált kalmodulin diszfunkció szisztémás érvényesülése (Lowe
et al., 2000)
7. NOTOM (normal neuro-osseous timing of maturation) modell, ami több teória
egyesítésével jött létre (Burwell et al., 2008b)
8. LHS (leptin, hipotalamusz, szimpatikus idegrendszer) (Burwell et al., 2009)
Eddig egyetlen elmélet sem tudta az AIS-ben megfigyelt eltérések mindegyikét
beilleszteni a kórfejlıdésbe, ami azt sugallja, hogy az AIS, mint a legtöbb komplex betegség
pathomechanizmusa összetett. Egyre több adat utal arra, hogy a hormonháztartás nem
megfelelı mőködése fontos etiológiai tényezıje az AIS-nek. Saját kísérleti eredményeink is
arra utaltak, hogy fontos lenne az endokrin rendszer részének tekinthetı zsírszövet által
termelt fehérjéket, az adipokineket tanulmányozni. Eddig több mint 50 adipokint írtak le,
amelyek számos szerv mőködését befolyásolják,mint például a máj, az izmok, a csontok, a
szaporító szervek (Trayhurn and Wood, 2004). Az adipokinek közül részletesen a leptinnel és
az interleukin 6-tal foglalkozom, mivel mindkettınek fontos szerepe van a csonttömeg
átalakítás szabályozásában és szerepük valószínősíthetı az AIS kialakításában.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
23
Az értekezés szempontjából kiemelten fontos etiológiai faktorok
Leptin
A leptin egy, a citokinek családjába tartozó hormon, melyet 1994-es felfedezésekor az
energiaegyensúly fenntartásában és a táplálékfelvétel szabályozásában tartottak fontosnak
(Zhang et al., 1994). Elsıdleges feladatának a kövérség megelızését gondolták, erre utal az
elnevezése is, amely a görög leptos (jelentése vékony) szóból ered. Az elmúlt 15 évben
folytatott szerteágazó kutatások nyomán mára már elfogadott, hogy a leptin befolyásolja a
glükóz- és lipidmetabolizmust, fontos szerepe van neuroendokrin rendszer szabályozásában,
az immunrendszerben, a csontszerkezet átszervezıdésben.
A leptin gént (LEP) Friedman munkacsoportjának extrém kövér egértörzsek (ob/ob)
keresztezését követı pozícionális klónozással sikerült azonosítani (Zhang et al., 1994). Ezt
követıen emberben is azonosították a leptin génjét és megállapították, hogy aminosav szinten
84%-os a homológia a két faj leptin szekvenciája között, ami arra utal, hogy egy
funkcionálisan erısen konzervált fehérjérıl van szó.
A LEP gén az emberben a 7-es kromoszóma 7q31.3 régiójában található, ~16.5 kb
hosszúságú és 3 exonból valamint 2 intronból áll, a kódoló régiót a 2. és 3. exon tartalmazza
(Gong et al., 1996). A promóter régiója tanulmányozásakor C/EBP, SP-1, HIF-1 reszponzív
elemeket sikerült azonosítani (Ambrosini et al., 2002; Mason et al., 1998). A hozzájuk kötıdı
transzkripciós faktorok egy alapszintő átíráson túl indukálható leptin kifejezıdést is lehetıvé
tesznek. A leptin elsısorban a fehér zsírszövetben termelıdik, de kimutatták alacsony szintő,
de indukálható kifejezıdését a vázizmokban is (Ambrosini et al., 2002; Wang et al., 1998).
Termelıdését az inzulin és a glükokortikoidok fokozzák, a tesztoszteron pedig gátolja
(Saladin et al., 1995; Blum et al., 1997). A fenti hormonokon kívül még gyulladásos
citokinek is befolyásolják a termelıdését (Mantzoros and Moschos, 1998).
A leptin molekula egy glikozilálatlan 16 kDa molekulasúlyú, 4 alfa hélixet tartalmazó
polipeptid, amely a strukturális vizsgálatok alapján az IL6 citokin család tagjaival mutat
hasonlóságot (Zhang et al., 1997).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
24
Leptin szerepe a csonttömeg szabályozásban
Az elmúlt években több tanulmány jelent meg, amelyek rávilágítanak arra, milyen
bonyolult kapcsolatrendszer áll fenn a leptin, a központi idegrendszer és a csontszerkezet
átalakulása között. Az elsı arra utaló jel, hogy a leptin fontos szerepet játszik a megfelelı
csonttömeg kialakításában, abból két a megfigyelésbıl adódott, hogy a tartós energiahiánnyal
párosuló alacsony leptin szérumszint összefügg az alacsony csonttömeggel (pl. az anorexia
nervosa esetén) és, hogy a csontritkulás kevésbé fenyegeti az elhízott embereket (Soyka et al.,
1999; De Laet et al., 2005). Bár a pontos mechanizmusok még nem teljesen ismertek, úgy
tőnik, hogy a leptin két teljesen különbözı csonttömeg szabályozó folyamatban is részt vesz
(5. ábra piros nyilak). Az elsı, egy a hipotalamuszon keresztüli indirekt út, két egymással
ellentétes hatású folyamatból áll. Az egyik folyamatot Ducy és munkatársai írták le, miszerint
a leptin a szimpatikus idegrendszeren keresztül hat és hatására az oszteoklasztok
differenciációja fokozódik, ami a csontbontást erısíti (Ducy et al., 2000). A másik,
csontbontást gátló folyamatra Elefteriou csoportjának munkája során derült fény, melyben a
leptin egy hipotalamuszban kifejezıdı neuropeptid, a CART (kokain amfetamin által
szabályozott transzkript) expresszióját fokozza, amely az oszteoklaszt-differenciáció
gátlásához vezet (Elefteriou et al., 2005). A közvetlen úton keresztül a leptin a csontvelıi
ıssejtekre hat, mégpedig a csontépülésért felelıs oszteoblasztok differenciációját segíti és az
oszteoklasztok differenciációját gátolja (Thomas et al., 1999; Holloway et al., 2002). A fenti
mechanizmusok bonyolult kölcsönhatásaként a leptin szérumszintje határozza meg, hogy a
csonttömeg átalakulás egyensúlyi folyamatai közül melyik kerül túlsúlyba A szérum
leptinszint a különbözı koncentráció tartományokban a csontépítést és a csontbontást is
befolyásolhatja, de a koncentrációjától függıen ellentétesen (hormesis) (Martin et al., 2007).
Természetesen a leptin nem egymaga határozza meg a csonttömeg átalakulást, hanem számos
egyéb szabályozó úttal kölcsönhatásban, ami kielégítı magyarázatot szolgáltat arra, hogy
miért olyan ellentmondóak azok a klinikai adatok, amelyek a szérum leptinszint és a
csonttömeg közötti összefüggés vizsgáltával születtek.
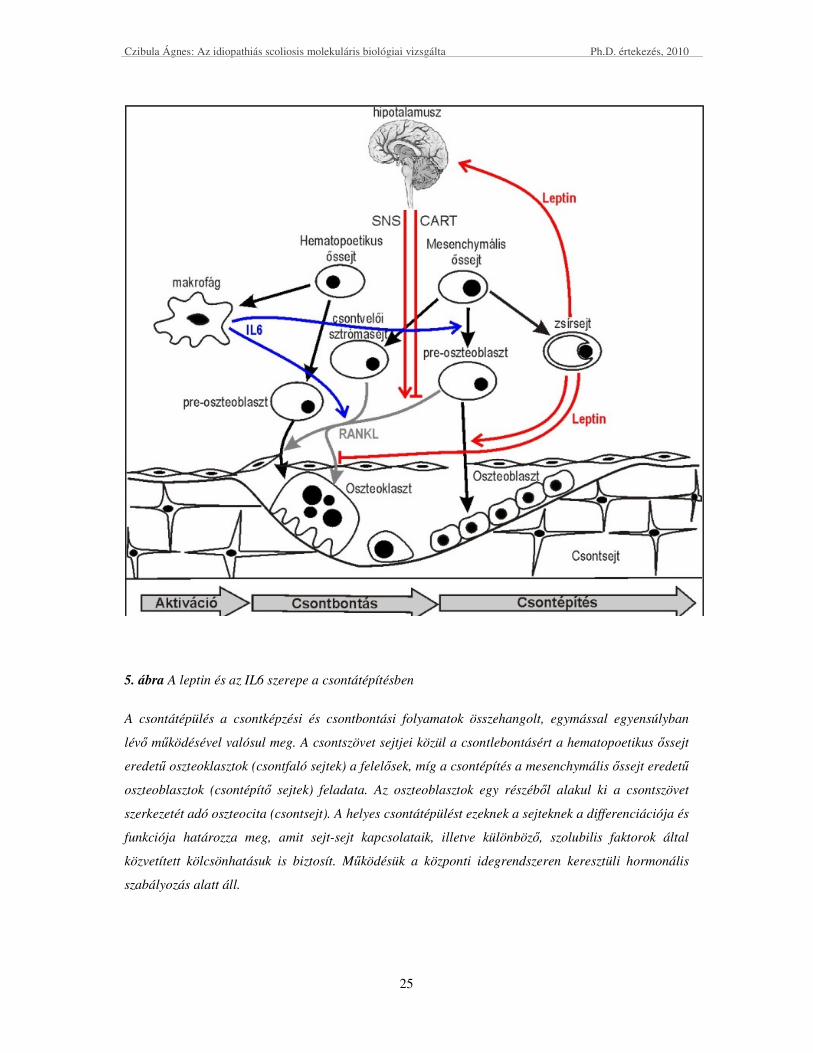
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
25
5. ábra A leptin és az IL6 szerepe a csontátépítésben
A csontátépülés a csontképzési és csontbontási folyamatok összehangolt, egymással egyensúlyban
lévı mőködésével valósul meg. A csontszövet sejtjei közül a csontlebontásért a hematopoetikus ıssejt
eredető oszteoklasztok (csontfaló sejtek) a felelısek, míg a csontépítés a mesenchymális ıssejt eredető
oszteoblasztok (csontépítı sejtek) feladata. Az oszteoblasztok egy részébıl alakul ki a csontszövet
szerkezetét adó oszteocita (csontsejt). A helyes csontátépülést ezeknek a sejteknek a differenciációja és
funkciója határozza meg, amit sejt-sejt kapcsolataik, illetve különbözı, szolubilis faktorok által
közvetített kölcsönhatásuk is biztosít. Mőködésük a központi idegrendszeren keresztüli hormonális
szabályozás alatt áll.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
26
Leptinszint nemek közötti különbsége
A leptin felfedezése után elıször részletesen a test zsírtartalma és a leptin szérumszint
összefüggését vizsgálták, és megállapították, hogy a nık leptin szintje még azonos testtömeg
index mellett is 2-3-szor magasabb mint a férfiaké (Ostlund, Jr. et al., 1996). Mivel a nıkben
magasabb a zsír százalékos aránya, nem meglepı, hogy a zsírsejtek által termelt leptin szintje
is különbözik. Ez az eltérés a pubertás korban válik kifejezetté, ami azt mutatja hogy a
zsírszövet mennyisége mellett a nemi hormonok is befolyásolják a leptin szérumszintjét. A
pubertás kezdetén mindkét nemben megnı a leptin szérumszintje, ami azt jelzi, hogy a
leptinnek fontos szerepe van a hipotalamusz-hipofízis-gonád tengely aktiválásában, azáltal,
hogy jelet szolgáltat az agynak arról, hogy megfelelı energiatartalékkal rendelkezik-e a
szervezet a reprodukcióhoz. Ezt alátámasztja az a megfigyelés, hogy alacsony leptinszinttel
rendelkezı élsportoló tornászoknál a serdülıkor kezdete késik és a sovány atlétanık
rendszerint amenorrheások (Weimann et al., 1999). További bizonyítékot jelent, hogy
azokban az esetekben, ahol a leptin génjében elıforduló misszensz mutáció következtében
nem termelıdik fehérje, a betegek az elhízás mellett hipogonadizmusban szenvednek (Strobel
et al., 1998). A serdülıkor késıbbi fázisában a leptin szérumszintje a lányokban tovább
emelkedik, szemben a fiúkkal akiknél inkább csökken kialakítva ezáltal a két nem közötti, az
élet további részében is fennmaradó leptinszintbeli különbséget (Blum et al., 1997).
Polimorfikus helyek vizsgálat a LEP gén szabályozó régiójában
Egy gén esetleges szerepét valamely fenotípus kialakításában leggyakrabban az adott gén
vagy vele genetikai kapcsoltságban lévı DNS szakaszon lévı polimorfikus helyek
vizsgálatával próbálják meg tisztázni az asszociációs vizsgálatokban. Eltérıen a LEP gén
kódoló szakaszától, ahol csak elvétve akad polimorfikus hely, az 5’végi szabályozó régióban
számos fellelhetı, közöttük olyanok is amelyek asszociációt mutattak az elhízással és a
testtömegindex-szel (a testtömegindex a testsúly és a testmagasság négyzetének a hányadosa,
amellyel a test zsírszövettartalmát lehet megbecsülni) (Carlsson et al., 1997). Több egymással
részben ellentmondó tanulmányban számos SNP-t azonosítottak, amelynek valamely allélja
kapcsoltságot mutat az alacsonyabb leptinszinttel vagy az elhízással.
A leggyakrabban vizsgált SNP, egy általánosan elıforduló polimorfizmus, amely a
transzkripciós starthelytıl 5’ irányban 2548 bázispárnyira helyezkedik el és egy G/A

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
27
tranzíciót jelent; jelölése G-2548A. Funkciójáról egyenlıre nincs adatunk, de számítógépes
elemzések azt mutatják, hogy evolúciósan nem konzervált régióban helyezkedik el és egyik
(eddig ismert) transzkripciós faktor kötıhely sem térképezhetı erre a pontra. Mivel azonban
számos vizsgálatban erıs asszociációt mutat az elhízással egy függetlenül ható, funkcionális
SNP-nek tekintik.
A vérszérum leptinkoncentrációja a szervezetben tárolt energia mennyiségérıl ad információt
és a test zsírszázalékával egyenesen arányos, bár az egyéni szérum leptinszintek azonos
testtömegindex mellett is nagy eltéréseket mutathatnak. Több tanulmány is foglalkozott azzal,
hogy a szérum leptinszintje és a G-2548A SNP alléljei közötti összefüggést feltárja, de a
kapott adatok ellentmondóak. Két kutatócsoport számolt be olyan eredményrıl, hogy a -2548
A/A homozigóta genotípus a magasabb szérum leptinszinttel hozható összefüggésbe. Az
egyik közlemény csak férfiakban tudta kimutatni ezt az összefüggést, a másik pedig, nem
elhízott felnıtt nıkben találta a magasabb leptin szérumszintet az A/A homozigótákkal
kapcsoltnak (Mammes et al., 2000; Hoffstedt et al., 2002). Több ellentétes értelmő publikáció
is megjelent, amikben a -2548 A/A genotípusú testesebb fiatal lányokban 20 %-kal
alacsonyabb szérum leptinszintet mutattak ki, mint a G/A vagy G/G genotípusúakban, illetve
50%-kal alacsonyabb receptorhoz kötött szérum leptinszintet A/A genotípusú normál súlyú
lányokban (Le Stunff et al., 2000; Yiannakouris et al., 2003). A legújabb témával foglalkozó
cikk az eddigiektıl eltérıen nem európai populációkat, hanem brazil nıket vizsgálva jutott
arra a következtetésre, hogy a -2548 G allél jelenléte összefüggésben van az emelkedett
szérum leptinszinttel (Hinuy et al., 2008). Meg kell említeni azonban, hogy olyan publikációk
is megjelentek, amelyek nem tudtak összefüggést kimutatni a LEP -2548 G/A allélok és a
szérum leptinszintje között (Bienertova-Vasku et al., 2008; Wang et al., 2006). Az
ellentmondó eredmények hátterében több jelenség is állhat. Egyik ilyen különbséghez vezetı
ok lehet a G-2548A polimorfizmussal kölcsönható másik allél jelenléte a leptin vagy a leptin
receptorának a génjében. Szintén eltérı eredmények megszületéséhez vezethetnek a
vizsgálatokba bevont populációk nagyságában, illetve nemi és testfelépítési paramétereiben
megnyilvánuló különbségek, valamint az eltérı statisztikai modellek használata.
A leptin rákos folyamatokban betöltött lehetséges részvételére elıször azon in vitro
eredmények hívták fel a figyelmet, amelyek igazolták a sejtosztódásban és a véredények
kialakulásában betöltött szerepét. E kísérletek nyomán a G-2548A polimorfizmust nemcsak
az elhízással kapcsolatban, hanem számos rákos elváltozással összefüggésben is
megvizsgálták (Dieudonne et al., 2002). Több, eddig vizsgált tumor kialakulása vagy
malignitásának vonatkozásában igazolást nyert, hogy a 2548 A/A homozigóta állapot

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
28
tekinthetı a magasabb kockázati tényezınek. A kutatások során tapasztalt, esélyhányados
értékük (OR) a mellrák esetén 3.17, nem kissejtes tüdıráknál 1.97, szájnyálkahártya tumor
esetén 1,56, non-Hodgkin limfóma esetén 2,3, és a prosztataráknál 2,93 (Ribeiro et al., 2004;
Ribeiro et al., 2006; Skibola et al., 2004; Snoussi et al., 2006; Yapijakis et al., 2009).
Leptin és az AIS
Mára már széleskörben elfogadott nézet, hogy az AIS kialakulása és súlyosbodása a
növekedéssel áll kapcsolatban. Az AIS-es betegeknek nemcsak alacsonyabb a testsúlyuk és a
testtömegindexük, hanem csonttömegük is és csontsőrőségük is csökkent (Cheng et al., 2000;
Sadat-Ali et al., 2008). Ez az állapot felnıttkorban is fennmarad, és a betegek további
életútját követı kísérletekbıl az is kiderült, hogy a gerincferdülésben szenvedı betegek 50%-
ában csontritkulásos kórkép alakult ki (Cheng et al., 1999; Thomas et al., 1992). Mivel az
elızı fejezetekben már részletesen leírtam, milyen szerepet tulajdonítanak a leptinnek a test-
és a csonttömeg szabályozásában logikusan merült fel a kérdés, van-e szerepe a leptinnek az
AIS kialakításában. Eddig a szakirodalomban egyetlen tanulmány jelent meg, ami a leptin
szérumszintjét vizsgálja a gerincferdüléssel összefüggésben (Qiu et al., 2007c). A kínai
kutatócsoport 120 AIS-es és 80 korban illesztett egészséges lány leptinszintjét vizsgálta meg
és azt találta, hogy a kontrollhoz képest mintegy a felére csökkent a szérum leptinszint az
AIS-es betegekben, 14.9 µl/L-rıl 7.2 µl/L-re függetlenül a görbület nagyságától.
Az interleukin 6
Az interleukin 6–ot (IL6) eredetileg mint makrofág differenciációs markert és hepatocita, B
sejt stimuláló faktort írták le, mára már igazolást nyert szerteágazó pleiotróp hatása az
immunrendszert, központi idegrendszert, sıt a csontrendszert érintı számos biológiai
folyamatban.
Az IL6 génje a 7-es humán kromoszóma 7p21 régiójában található és 6.1 kb hosszúságú. A
gén 5 exont tartalmaz és eddig 9 különbözı mRNS variánsát írták le. Az IL6 fehérje 212
aminosavból áll és különbözı poszttranszlációs módosulások jellemzik, mint az O- és N-
glikoziláció valamint a szerin és tirozin aminosavakon történı foszforiláció. A vérszérumban
jelenlévı 26 kDa-os fehérje mintegy 30%-a a zsírszövetben termelıdik, de az adipocitákon
kívül termelik még az oda bevándorló makrofágok is. A mőködı vázizom is nagy
mennyiségő IL6-ot termel, a maratoni táv lefutása után például a vérplazma IL6 szintje

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
29
mintegy 100-szorosára emelkedik, melynek mértéke hasonló, mint amekkorát egy komoly
fertızésben szenvedı betegnél tapasztalnak (Ostrowski et al., 1999). A csontfejlıdésben
parakrin módon szerepet játszó IL6-ot elsısorban az oszteoblasztok és a csontvelıi sztróma
sejtek termelik (Holt et al., 1996).
Az IL6 az antigénspecifikus immunválasz szabályozásában szintén meghatározó szerepet
játszik. A B sejtek plazmasejtté történı érését és ellenanyagtermelı képességét szabályozza,
indukálja a T sejtek növekedését és a citotoxikus T sejt differenciációt. Májsejtekben az akut
fázis proteinek, elsısorban a C reaktiv-protein termelıdését közvetlenül indukálja. Az akut
fázis válasz során az izom- és zsírszövetekben energiát mobilizál, ezáltal segítve elı a láz
kialakulását, illetve hatására alakul ki a gyulladás során megfigyelhetı súlyvesztés (Leon et
al., 1998).
Dolgozatom szempontjából talán a legérdekesebb megfigyelés, hogy az IL6 érintett több
elhízással járó megbetegedésben (2-es típusú diabetes, a szív- és érrendszeri
megbetegedések), illetve a csontritkulás kialakulásában is (Thalmann and Meier, 2007; Zhao
et al., 2008).
Az IL6 szerepe a csontátépítésben
A szervezetben lejátszódó csontátépítési esemény a csontbontás, illetve csontépítés
egymással ellentétes, viszont térben és idıben egymással szorosan kapcsolódó, egymással
összehangolt folyamatai által valósul meg, melyben a résztvevı oszteoklasztok és
oszteoblasztok egymással kölcsönhatásban szabályozzák differerenciációjukat és
mőködésüket. Ebben a folyamatban a legjobban vizsgált pont az oszteoblasztok és a
csontvelıi sztróma sejtek által termelt citokinek hatása az oszteoklaszt differenciációra. A
legnagyobb szerepet az oszteoblaszt/oszteoklaszt kommunikációban a RANKL (NF-κβ
receptor aktivátor ligandja) szolubilis citokinnek tulajdonítanak (5. ábrán szürke nyilak). A
RANKL kötıdése receptorához, a RANK-hoz (NF-κβ receptor aktivátor) stimulálja a pre-
oszteoklasztok differenciációját és aktiválja az érett oszteoklasztokat. Számos egyéb
gyulladáskeltı citokin, köztük az IL6 is befolyásolja a RANKL funkcionális hatását.
Legújabb adatok szerint az IL6 típusú citokinek hatására az oszteoblasztokban indukálódik a
RANKL expressziója, mely az oszteoklasztokra parakrin, az oszteoblasztokra autokrin
módon képes hatást gyakorolni, tovább stimulálva ez által a parakrin faktorok kifejezıdését
(5. ábra kék nyilak). A kaszkád eredményeképp a gyulladáskeltı citokinek végsı soron a
csontbontást segítik (Kwan et al., 2004). In vitro adatok szerint az IL6 típusú citokinek a

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
30
mesenchymális ıssejtek oszteoblaszt irányba történı differenciációját is segítik és védik az
oszteoblaszt sejteket az apoptózistól, ami pozitívan befolyásolja a csontépítés folyamatát
(Taguchi et al., 1998). Az in vitro kísérletekkel párhuzamosan különbözı transzgenikus és
génkiütött egerek létrehozásával in vivo is vizsgálták az IL6 típusú citokinek csontképzıdésre
kifejtett hatását. Az IL6 hiányos egerekben a csontépítés mértéke nıtt, míg az IL6-ot
túlzottan kifejezı transzgenikus egerekben mind az oszteoblasztok, mind az oszteoklasztok
csökkenı száma mellett, a csontbontás folyamata dominált. Összeségében úgy tőnik, hogy a
csontbontásból és csontépítésbıl álló csontátalakulási folyamat egészét szupresszálja az IL6
(Blanchard et al., 2009).
Polimorfikus helyek vizsgálat az IL6 gén szabályozó régiójában
Az IL6 vérszérum koncentrációja egy adott populáción belül lényeges eltéréseket mutat az
egyedek között. Különösen a gyulladásos folyamatokra jellemzı IL6 szérumszint emelkedés
mértéke mutat nagy egyedi eltérést. A különbözı immunstimulusra kialakuló magas IL6
szérumkoncentráció sokáig fennmarad, így jó molekuláris markere számos klinikai
állapotnak, mint pl. a hasi fertızéseknek, áttétes mellráknak, szív- és érrendszeri
megbetegedéseknek. Idıvel a vizsgálatok egyik lényeges kérdésévé az vált, hogy az egyén
genetikai háttere hogyan befolyásolja az IL6 szérumkoncentrációt egészséges és pathológiás
állapotokban. Mivel az IL6 gén kódoló szekvenciája csak néhány nagyon ritkán elıforduló
polimorfikus helyet tartalmaz, a gén szabályozó régiójában található és a génkifejezıdésre
ható variánsokat kezdték el vizsgálni. A leggyakrabban vizsgált SNP a transzkripciós
starthelytıl -174 bázispárnyira elhelyezkedı G>C transzverzió(G-174C, rs1800795), amely
az IL6 promóter egyik negatív regulációs doménjében helyezkedik el és in vitro kísérletek
alapján funkcionális variánsnak tekinthetı. HeLa sejteken végzett „reporter assay”
vizsgálatokban a -174C allélt tartalmazó konstrukciók luciferáz expressziója 0.624±0.15-ször
volt alacsonyabb a -174G allélt hordozó vektorénál. IL1 stimulust követıen a -174C reporter
gén aktivitása nem változott, míg a -174G allélt tartalmazó konstrukciók esetében az aktivitás
3.60±0.26-szorosára nıtt (Fishman et al., 1998). Számos tanulmány foglalkozott a G-174C
polimorfizmus és a vérszérum IL6 szintjének összefüggésével, de a kapott eredmények
ellentmondásosak. A G-174C polimorfizmust felfedezı, az elıbbiekben ismertetett in vitro
eredményeket közlı csoport egészséges emberek vizsgálatakor azt találta, hogy
összehasonlítva a G allélt is hordozó egyedeknél mért értékekkel, a C allélt homozigóta
formában hordozóknál szignifikánsan alacsonyabb az IL6 szérumszint (Fishman et al., 1998).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
31
A késıbbiekben ennek épp az ellenkezıjét megállapító tanulmányok is megjelentek (Boiardi
et al., 2006; Ravaglia et al., 2005). Az ellentmondásos eredményekre egy lehetséges
magyarázattal szolgálhat az IL6 kifejezıdését szabályozó, egy másik régióban jelenlévı
polimorfizmus, amelyet Smith és mtsai le is írtak a közelmúltban (Smith et al., 2008).
A G-174C polimorfizmust a felfedezése óta eltelt tíz évben számtalan olyan betegséggel
kapcsolatban megvizsgálták, ahol az IL6 szerepe felmerülhetett. Ezek:
- a fiatalkori ízületi gyulladásban a -174C/C genotípus ritkábban fordul elı (Fishman
et al., 1998)
- a 2-es típusú diabetesben a kezdeti eredmények alapján a -174G allél, míg az
elhízással kapcsolatban a -174C allél jelenlétét tekintették a magasabb kockázati
tényezınek, ám ezeket az eredményeket a meta-analízisek már nem támasztják alá (Qi
et al., 2006) (Qi et al., 2007)
- a -174C/C genotípus hordozóinak a méhnyakrák kialakulására nagyobb a kockázata
(OR=3.16; p=0.014) míg a nyelıcsı tumor esetén a -174G/G genotípusú homozigóta
egyéneknek magasabb a esélyhányadosa (OR 2.28; P=0.001) (Gangwar et al., 2009;
Upadhyay et al., 2008)
- a csontritkulással kapcsolatos vizsgálatban azt találták, hogy a -174C/C
homozigótákban magasabb a csontsőrőség, mint a -174G/G homozigótákban (Xiao et
al., 2005)
Az IL6 és az AIS
A porckorongokat érı abnormális terhelés miatt a gerincferdülésben a porckorong alakja
megváltozik, kiékelıdik és ezt a változást követi a porckorong belsı kocsonyás magjának
(nucleus pulposus, NP) szerkezeti és metabolikus módosulása is (Oegema, Jr. et al., 1983;
Roberts et al., 1993; Taylor et al., 1981). Burke és mtsai kimutatták, hogy gyulladáskeltı
stimulusra a scoliotikus porckorong NP sejtjei megnövekedett IL6 szekrécióval válaszolnak,
ugyanakkor alapállapotban nincs eltérés az egészséges és scoliotikus sejtek IL6 termelésben
(Burke et al., 2003). A fenti eredmények ismeretében Logroscino munkacsoportja
megvizsgálta, hogy az IL6 promóterében lévı G-174C SNP eloszlása mutat-e eltérést az
egészséges és kontroll populáció között (Aulisa et al., 2007). A vizsgálatba Olaszország
középsı és déli részérıl származó 53 IS-es és 202 kontroll egyedet vontak be és eset-kontroll

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
32
asszociációs analízist végeztek. A kapott genotípus eloszlás mind a kontroll, mind a beteg
csoportban Hardy-Weinberg egyensúlyban volt és a kontroll minták alléleloszlása nem
mutatott eltérést a más populációt érintı irodalmi adatokhoz képest. A gerincferdülésben
szenvedı betegek között azonban kétszer olyan gyakran fordult elı a -174 G/G genotípus,
mint a kontroll csoportban (52.8% , illetve 26.2% p<0.001) míg a -174 C/C genotípus
gyakorisága szignifikánsan magasabb volt a kontroll csoportban (30,1%), mint a scoliosisos
betegek között (5.7%). Az elvégzett statisztikai elemzés szerint az IL6 -174 G/G homozigóta
genotípusú egyedek esélye a gerincferdülés kialakulására 10.54-szer, a G/C heterozigóta
genotípusú személyeké pedig 4.84-szer magasabb, mint a C/C homozigóta genotípusú
embereké, bár szignifikáns különbséget csak a két homozigóta pár között lehet kimutatni. Az
eredmények alapján a szerzık úgy gondolják, hogy az IL6 gén promóterének polimorfizmusa
fontos genetikai faktora lehet a gerincferdülésre való hajlamnak. Ugyanakkor a vizsgálatba
bevont 53 gerincferdülésben szenvedı beteg mintaszáma alacsony ahhoz, hogy a kapott
eredményeket maradéktalanul elfogadhassuk.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
33
CÉLKITŐZÉSEK
Az idiopathiás scoliosis kialakulásának okát évtizedek óta széles körben kutatták és kutatják a
mai napig. Az erıfeszítések ellenére azonban eddig nem sikerült olyan etiolóiai teóriát
kialakítani, ami minden, az AIS-ben megfigyelhetı jellegzetességet, rendellenességet, eltérést
be tudna illeszteni egy egységes rendszerbe. A munkánk során célul tőztük ki, hogy
molekuláris szintő vizsgálatokban, két megközelítéssel az AIS-re jellemzı további eltéréseket
tárjunk fel és az eltérések hátterében álló genetikai tényezıket vizsgáljuk.
A. Az elsı megközelítésben a gerincoszlop stabilitásáért felelıs paravertebrális izmok
génexpressziós mintázatát kívántuk megvizsgálni. A teljes izom transzkriptoma egyidejő
analízisével, a CHIP microarray módszerrel összehasonlítottuk az:
I. Egészséges és scoliotikus paravertebrális izmok génkifejezıdését
II. Scoliotikus izmokban a görbület konvex és konkáv oldali génkifejezıdését
Azokat a géneket vizsgáltuk tovább, amelyek kandidánsként szóbajöhetnek az alábbi
szempontok alapján:
1. Bioinformatikai módszerekkel meghatározott, a háttér eloszlástól eltérı
génontológiai kategóriába esnek
2. Közös az AIS szempontjából érdekes, biológiai, metabolikus folyamatban vesznek
részt
3. Az AIS vonatkozásában elvégzett kapcsoltsági analízisek által jelölt
kromoszómaszakaszra lokalizálódnak
4. Az általuk termelt fehérje a funkciója alapján beilleszthetı az AIS etiológiájába
B. Második lépésként a génexpressziós vizsgálatok által jelölt gén/gének SNP
polimorfizmusának asszociációs analízisét eset-kontroll tanulmányban terveztük vizsgálni. A
genotipizálás eredményeit különbözı, a gének közötti interakciók kimutatására alkalmas
statisztikai módszerekkel szándékoztuk kiértékelni. A genotípus adatokat a klinikai adatokkal
összevetve a statisztikai analízis lehetıséget ad annak a meghatározására, hogy a gén/gének
adott allélja a gerincferdülés kialakulásához vagy súlyosbodásához köthetı-e.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
34
ANYAGOK ÉS MÓDSZEREK
Kísérletbe bevont személyek adatai és mintái
Gerincferdülésben szenvedı betegek mintái
Az AIS-es betegek mintáit a Pécsi Tudományegyetem Általános Orvosi Kar Ortopédiai
Klinikájáról Prof. Illés Tamástól kaptuk. A vizsgálatba bevont személyeknél a kiválasztásuk
elıtt ortopédiai vizsgálat, gerincröntgen, laboratóriumi vizsgálat (szérum kalcium, foszfor,
alkalikus foszfatáz szint és vizelet kalcium és foszforszint mérés) történt. Teljes vázrendszeri
vizsgálat készült a lehetséges diszpláziák kiszőrésére. A betegeket és elsı fokú rokonaikat
egy genetikus vizsgálta meg a lehetséges genetikai szindrómák szempontjából. Az idiopathiás
scoliosis diagnózisát bizonyított genetikai szindróma, metabolikus, neurológiai-, kollagén-
vagy izombetegség hiányában állapították meg.
DNS minták egyrészt a mőtétek során nyert izom biopsziákból, másrészt IS-es betegek
perifériás vérébıl nyert fehérvérsejtjeibıl preparáltuk, összesen 126-ot.
RNS mintákat a korrekciós mőtéten átesett betegekbıl az operáció során nyert bilaterális
musculus latissimus dorsi izombiopsziából preparáltuk összesen 32-öt.
A betegeket minden esetben tájékoztattuk a kísérletbe történı bevonásukról és a
beleegyezésükkel történtek a mintavételek.
Kontroll személyek mintái
A kontroll személyek mintái több forrásból származnak, de a kiválasztásnál mindig szem
elıtt tartottuk a dél alföldi régióból való származást. A kontroll minták mintegy felét (112
minta) a Szegedi Tudományegyetem Gyerekgyógyászati Klinikájáról, Dr Endreffy Emıkétıl
kaptuk, a másik részét a munkacsoportunkban folytatott másik populációgenetikai
vizsgálathoz győjtöttük, fıleg az SZBK Genetika Intézet dolgozóitól. A kontroll személyek
sem egymással sem az AIS betegekkel nem állnak rokonságban.
DNS mintákat perifériás vérbıl kinyert fehérvérsejtekbıl, szájnyálkahártya kenetbıl és
hajszálak végén lévı hajhagymákból készítettük.
A microarray analízishez használt kontroll RNS mintákat trauma miatt operált betegek
bilaterális musculus latissimus dorsi biopsziából készítettük.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
35
DNS alapú munkafolyamatok
Genomikus DNS tisztítása
A genomikus DNS tisztítása perifériás vérbıl, izombiopsziából a hagyományos fenolos
extrakcióval (Short Protocols in Molecular Biology 1992), száj nyálkahártya kenetbıl és
hajszálakból pedig a Walsh és munkatársai által kidolgozott Chelex-alapú módszerrel történt
(Walsh et al., 1991).
SNP-k kimutatása PCR-RFLP módszerrel
A PCR-RFLP módszeren alapuló genotipizálás során elıször PCR-rel felszaporítjuk a
vizsgálni kívánt SNP-t tartalmazó DNS szakaszt, majd olyan restrikciós enzimmel emésztjük
az amplikont, melynek felismerı helyét a polimorfizmus érinti.
A leptin promóter G-2548A polimorfizmusát ezzel a módszerrel már többen is vizsgálták, mi
Yapijakis és kollégái által leírt módszer alapján végeztük el a genotipizálást. (Yapijakis et al.,
2009) Az IL6 promóter C-174G polimorfizmusát pedig Aulisa és munkatársai által használt
protokoll szerint vizsgáltuk. (Aulisa et al., 2007) A következı primereket (Merck Kft)
használtuk a PCR-hez:
Leptin (rs7799039) F:5`-TTTCCTGTAATTTTCCCGTGAG-3`
R:5`-AAAGCAAAGACAGGCATAAA-3`
IL6 (rs1800795) F:5’- TGACTTCAGCTTTACTCTTTgT-3’
R:5’- CTGATTGGAAACCTTATTAGG-3’
A PCR-t MJ Research PTC 200-as PCR készülékben végeztük GOTaq (Promega) DNS
polimerázzal a következık szerint:
� 25 µl össztérfogatban a PCR reakciót az alábbiak alkották: 100 ng DNS templát,
200 µM ΣdNTP, 2 mM MgCl2, 1 µM forward és 1 µM reverz primer, 1x GOTaq puffer
(Promega), 0.25 U GOTaq DNS polimeráz.
� A reakcióelegyet 94 oC–on 5 percig tartottuk a Taq polimeráz aktiválásához majd
35 ciklusban végeztük az amplifikálást: denaturálás 94 oC–on 30 másodpercig, primer
kapcsolódás (annealing) 52 oC-on 30 másodpercig, átírás 72 oC–on 30 másodpercig.
Végezetül a szál lezárása 72 oC–on 5 percig zajlott.
� A PCR termék restrikciós emésztését a következık szerint végeztük: a 20 µl
össztérfogatú reakcióelegyben: 8 µl PCR termék, a gyártó által szolgáltatott megfelelı 1x

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
36
puffer, 3 U HhaI (Fermentas), illetve 5 U NlaIII (New England Biolabs). A reakciót 37 oC–on
egész éjszakán át inkubáltuk. A restrikciós emésztéseknél mindig használtunk kontroll DNS
mintát, hogy ellenırizhessük az emésztés eredményességét.
� A PCR termékek kimutatására a Mini-PROTEAN III Electrophoresis System (BioRad)
gélelektroforézis rendszert használtunk. 8%-os natív poliakrilamid gélen 1x TBE futtató
puffer jelenlétében. A vizualizációhoz festettük a gélt 1 µg/ml etidium-bromidos vízben 10
percig, majd a GelBase (UVP) géldokumentációs rendszerrel archiváltuk és értékeltük a
géleket (6. ábra).
6.ábra A. A leptin G-2548A, illetve az IL6 C-174G SNP által meghatározott genotipusokhoz
tartozó restrikciós fragmentumok hossza
B. Gélfotó, amely a leptin G-2548A SNP meghatározását mutatja
C. Gélfotó, amely az IL6 C-174G SNP meghatározását mutatja

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
37
RNS alapú munkafolyamatok
RNS izolálás
� A hátizom biopsziából kb. 50-100 mg szövetet folyékony nitrogén alatt dörzsmozsárban
porítottuk, ehhez 1 ml TRIzol (Invitrogen) reagenst adtunk és 5 percig szobahımérsékleten
inkubáltuk a nukleoprotein komplexek disszociációja végett. A mintát nagy proteoglikán, zsír
és poliszacharid tartalma miatt 4 oC-on, 12000 g-vel, 10 percig centrifugáltuk.
� A felülúszót új Eppendorf csıbe helyeztük s ahhoz 200 µl kloroformot adtunk. 15
másodpercig összeráztuk, majd 15 perces inkubáció után 4 oC-on, 12000 g-n, 15 percig
centrifugáltuk. A kialakult három fázis közül az RNS-t is tartalmazó vizes óvatosan
eltávolítottuk, ügyelve, hogy az interfázistól elválasszuk.
� A vizes fázishoz hozzámértünk 0.5 ml izopropilalkoholt, majd 15 másodperc összerázás
után 10 percig szobahımérsékleten inkubáltuk. A 4 oC-on, 12000 g-vel, 10 percig tartó
centrifugálás hatására az Eppendorf csı aljára kicsapódott az RNS. A felülúszót leszívtuk, 1
ml 70%-os etil-alkohollal mostuk a csapadékot vortexelés mellett. 7500 g-s 5 perces
centrifugálás után az etil-alkoholt eltávolítottuk.
� Az RNS-t szobahımérsékleten szárítottuk, majd 50 µl RN-áz mentes, DEPC-s vizet
(Fermentas) mértünk a csapadékra. Az RNS oldódását az 55-60 oC-on történı 10 perces
inkubáció segítette.
Northern gél elektroforézis
Az RNS minták minıségének ellenırzésére használtuk.
� Agaróz gél összetétele: 1% agaróz (GIBCO), 1x MOPS pH:7, 0.65% formaldehid.
� A futtató puffer: 1x MOPS ph:7, 0.65%-os formaldehid.
� RNS minták gélfuttatás elıtti kezelése az alábbiak szerint történt: 3.5 µl RNS
oldathoz hozzámértünk 10 µl formamidot, 3.5 µl 35%-os formaldehidet, 2 µl 10x
MOPS pH:7 oldatot, 1 µl 1 mg/ml etídium-bromidot, majd 70 oC-on 10 percig
inkubáltuk.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
38
� A mintákhoz a gélre való feltöltés elıtt 0.1 térfogat Blue Juice oldatot (50%
glicerin, 0.4% bróm-fenolkék, 0.4% xilén-cianol, 1mM EDTA)adtunk.
� A gél futtatása 80 V-on kb. 2 órán keresztül történt.
� A vizualizációhoz és a dokumentációhoz a GelBase géldokumentációs rendszert
(UVP) használtuk.
7.ábra Northern agaróz gélelektroforézis
Az RNS mintákat akkor fogadtuk el megfelelı minıségőnek, amennyiben a 28S nagy
molekulasúlyú riboszomális RNS-ek is erısen látszottak, mint az a 7.ábrán is látható.
RNS mintacsoportok tisztítása
Az egyéni RNS mintákat egyenlı arányban összekevertük, létrehozva ezzel azokat a
mintacsoportokat (pool-okat), amelyekkel a továbbiakban dolgozni szándékoztunk. A további
munkafolyamatok elıtt a kialakított RNS oldatokat tovább tisztítottuk Macherey-Nagel
Nucleospin RNAII kitjével a gyártó leírása alapján:
� Az RNS oldathoz 450 µl RA1 oldat és 7 µl β-merkaptoetanol keverékét adtuk, majd
összeráztuk és 350 µl 70%-os etanolt hozzáadása után nukleinsav-kötı oszlopra
pipettáztuk. 2 percig centrifugáltuk 1000 RPM-mel.
� A membránhoz kötött RNS mosása után 90 µl 25 mM-os DN-áz I hozzáadásával 15
percig szobahımérsékleten inkubáltuk, majd hozzámértünk 200 µl RA2-t (DNase stop
solution buffer). 2 perces centrifugálás-1000 RPM-mel.
28S RNS
18S RNS
5S RNS

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
39
� A DN-áz tökéletes eltávolítása érdekében további mosásokkal tisztítottuk a
membránhoz kötött RNS-t: 300 µl RA3 puffer hozzáadása után 2 perc centrifugálás
1000 RPM-mel, majd 125 µl RA3 pufferrel történı mosás után 4 perc centrifugálás
1000 RPM-mel.
� A nukleinsav-kötı mosást követıen az oszlopot áttettük egy steril Epp.csıbe, 60 µl
DEPC H2O-t (Fermentas) mértünk rá és szobahımérséklet 10 percig állni hagytuk,
majd az RNS-t 4 perces 1000RPM-mel történı centrifugálással oldottuk le a
membránról.
� Amennyiben szükséges volt akkor az RNS oldatokat koncentráltuk Amicon
Microcon centrifugacsıvel (Millipore) 11000 RPM-mel 20 percig történı
centrifugálás után 8 µl DEPC H2O-t mértünk a nukleinsav-kötı oszlopra, majd a
csövet megfordítva 1 percig centrifugáltuk.
Fluoreszcensen jelölt próba készítése a microarray hibridizáláshoz
A próba készítéséhez a Cy5 fluoreszcens festékkel kapcsolt 3DNA Array 900MPX Kitet
(GeniSphere) használtuk a gyártó leírása alapján:
1. A reverz transzkripciót a RevertAid™ First Strand cDNA Synthesis Kittel (Fermentas)
végeztük a gyártó leírása alapján. Röviden:
� A mintacsoportonként 2.5 µg RNS-hez 5µl random hexamer primert és 1 µl oligo
dT primert adtunk 13 µl térfogatban. 10 percig 80 oC-on inkubáltuk, majd rögtön jégre
tettük.
� Hozzáadtunk 4 µl 5x Fermentas puffert, 1 µl 10mM ΣdNTP-t, 1 µl ribonukleáz
inhibitort és 1 µl Fermentas MMLV reverz transzkriptázt. Inkubáltuk 2 órán át 42 oC-
on.
� A mRNS-t hidrolizáltuk 3.5 µl 0.5M NaOH/50mM EDTA oldat hozzáadása után 65 oC-on 15 percig, majd a cDNS oldatot neutralizáltuk 5 µl TRIS-t pH:7.5
hozzáadásával. A mintákat -20 oC-on tároltuk másnapig.
2. A cDNS tisztítása MinElute Spin column 50 kittel (Qiagen) történt a gyártó leírása alapján:

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
40
� 50 µl cDNS-hez 250 µl PB puffert adtunk, és az oldatot a MinElute oszlopra
pipettáztuk, majd 1 percig 13000 RPM fordulatszámmal centrifugáltuk.
� Az átfolyt PB mosópuffer eltávolítása után 750 µl PE puffert mértünk az oszlopra és
1 percig centrifugáltuk majd az átfolyó puffert kiöntve újabb egy perces centrifugálás
történt a maradék etanol eltávolítása érdekében.
� A cDNS visszanyerésénél az oszlopot egy steril csıbe helyeztük, a nukleinsav-kötı
membránra 10 µl EB puffert pipettáztunk, 2 percig szobahımérsékleten inkubáltuk,
majd 2 perc centrifugálás után az oszlopot eltávolítottuk.
3. Címkézés
� A cDNS-hez 6.5 µl nukleázmentes desztillált vizet adtunk. A 16,5 µl-nyi mintát
80oC-on 10 percig melegítettük, majd rögtön jégre helyeztük.
� Hozzámértünk 2.5 µl 10x címkézı puffert, 4 µl 10 mM dTTP-t és 2 µl terminális
deoxynukleotidil transzferázt, 37 oC-on 30 percig, majd 65 oC-on 15 percig
inkubáltuk.
4. Ligálás
� A 25 µl mintát 10 percre 95 oC-ra helyeztük, majd rögtön jégre tettük.
� Hozzáadtunk 5 µl 6x Ligation Mix-et (Cy5/Alexa 647) és 2 µl T4 DNS ligázt.
Finoman összeráztuk, szobahımérsékleten fél óráig inkubáltuk.
� A reakciót leállítottuk 3.5 µl 0.5M EDTA oldat hozzáadásával és 10 másodpercig
vortex-szel összekevertük. 14.5µl TE8 hozzáadásával értük el az 50 µl-es
végtérfogatot.
5. A jelölt cDNS-t tisztítottuk, a fentebb már említett módon, Qiagen MinEluteTM Spin
columns 50 kittel.
A cDNS és a 3DNA hibridizációja a CHIP-hez
A Genishere jelölı kithez tartozó protokollt követtük a továbbiakban is a hibridizációs mix
elkészítéséhez.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
41
� DNS hibridizációs mixet állítottunk össze a következıképpen: 10 µl cDNS, 2 µl
LNA dT Blocker, 3 µl nukleáz mentes víz, 15 µl 2x hibridizációs puffer.
� A mixet összekevertük vortex-szel, röviden centrifugáltuk, 75-80 oC-on inkubáltuk
10 percig, majd 55-65 oC között tartottuk a további használatig.
� Rövid vortexelés és centrifugálás után a mixet az elımelegített CHIP lemezre
tettük.
A hibridizáció a Ventana hibridizáló állomásban (Ventana Discovery) „antibody” protokol
szerint zajlott.
� Az elsı hibridizálási lépés (cDNS hibridizálás) 42 oC-on 18 órán keresztül a
következı hibridizáló pufferben történt: 10x Denhart oldat, 0.25 M Na foszfát puffer
pH 7.0, 1 mM EDTA, 1x SSC, 0.5% SDS.
� A második hibridizálási lépés (3DNA hibridizálás) 2.5 µl Cy5 fluorofort tartalmazó
„elıhívó” reagens jelenlétében 200 µl „Chiphyb” hibridizáló pufferben (Ventana)
történt 42 oC-on 2 óráig.
� A hibridizálás után a lemezeket 0.2X SSC oldatban kétszer mostuk.
A hibridizáció utáni kiértékelés
� A lemezek leolvasása konfokális lézer szkennerrel (ScanArray Lite, GSI Luminics)
történt.
� A pontok kiértékelése GenePix analizáló programmal történt.
� A számszerősített adatokat Misrosoft Excell program segítségével, minıségi
paraméterek figyelembe vételével értékeltük és normalizáltuk. Azok a pontok,
melyeknél a minta és a kontroll fluoreszcencia intenzitás értékekbıl képzett,
normalizált hányados -ami a gén relatív kifejezıdési szintjét mutatja- 2 felett illetve
0,5 alatt van, olyan gént jelöl, amelynek kifejezıdése eltérı a kontrollból és betegbıl,
illetve a jobb és a bal oldalról származó izombiopsziákban.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
42
Valós idejő kvantitatív polimeráz láncreakció (QRT-PCR)
A génspecifikus primereket a Primer Express programmal (Applied Biosystem)
terveztük a szoftver által javasolt beállításokat követve. A primerek specifitását az NCBI
BLAST szerverének a primerek tesztelésére alkalmas programjával (“Search for Short,
Nearly Exact Matches”) ellenıriztük. A primerek szintetizálását az SZBK Nukleinsav
Szintézis Központi Laboratóriumában végezték el. A primer szekvenciák a következıek
voltak:
LMNA F:5’ GAGCTGCCTTCCCTAGCTTTAGA 3’
R:5’ CTCCCAGCACCCCACTTG 3’
VDR F: 5’ GATATGGCAGCGAAAGGATGTAA 3’
R: 5’ TGACAAACTTCTGGCAAACTTTTT3’
COL15A1 F:5’ CATCCCAACATAGGTTAAGAGCAA 3’
R:5’ 3ACACTCAACAACAATGGTTTAAAAGG’
JAK1 F: 5’ TGCCCAGAAGCAGTTCAAGA 3’
R: 5’ TCCGAACCGTGCAGACTGTA 3’
BMP4 F:5’ CAAGCGTAGCCCTAAGCATCA 3’
R:5’ GGCGCCGGCAGTTCTT 3’
IL4R F: 5’ GCACCTCGACTTGTGAACGA 3’
R: 5’ GGACAGTCTGCTGCAGAAGCT 3’
NTF3 F:5’ TCCAGCCGGTGATTGCA 3’
R:5’ CGGTGAGTTGTAGCGTCTCTGT 3’
LEP F:5’ TTGTCACCAGGATCAATGACA 3’
R:5’ GTCCAAACCGGTGACTTTCT 3’
CDK5 F:5’ TTAATTTCCTGAGTGACCAGCAT 3’
R:5’ AGCCTCCTTTGGAAAGGTCT 3’
HOX7 F:5’ ACATCGAGCCTTCAACGTG 3’
R:5’ ACTTTTTGCAAATGAAGAAACTAAGA 3’
NOS1 F:5’ CCTCCTGGGTTTACTCCTTGAGT 3’
R:5’ AGTTTTGTAAGAGCCACTGCAGATT 3’
NOS2 F:5’ CCTCAAGTCTTATTTCCTCAACGTT 3’
R:5’ CCGATCAATCCAGGGTGCTA 3’
NOS3 F:5’ GCGGCTGCATGACATTGAG 3’
R:5’ TCGCGGTAGAGATGGTCAAGT 3’
HPRT F:5’ CACTGGCAAAACAATGCAGACT 3’
R:5’ CGACCTTGACCATCTTTGGATT 3’
β-AKTIN F:5 ’CCAACCGCGAGAAGATGA 3’
R:5 ’CCAGAGGCGTACAGGGATAG 3’

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
43
A reverz transzkripciót oligoT és random hexamer primerek felhasználásával a RevertAid™
First Strand cDNA Synthesis Kittel (Fermentas) végeztük a gyártó leírása alapján, amint azt a
microarray próba készítésénél már röviden leírtam.
A valós idejő PCR során a Rotor-Gene 3000 (Corbett Research) PCR készüléket használtuk a
PCR termék amplifikálására és kimutatására, ami a SYBR-Green festésen alapuló detekciós
eljárást használtuk a keletkezı termék nyomon követésére. Röviden:
� 20 µl össztérfogatban a PCR reakciót az alábbiak alkották: 5 µl ötszörösére hígított
cDNS, 0.5 µl 10 µM-os forward primer, 0.5 µl 10 µM-os reverz primer, 4 µl nukleáz mentes
víz (Fermentas) és 10 µl ABsolute QPCR SYBR Green Mix (ABgene).
� A reakcióelegyet elıször 94 oC–on 10 percig tartottuk, a DNS polimeráz
aktiválásához, majd 50 ciklusban amplifikáltuk a következık szerint: denaturálás 94 oC–on
25 másodpercig, primer kapcsolódás 61 oC-on 25 másodpercig, átírás 15 másodpercig.
� Minden reakció után elvégeztük az olvadáspont analízist, amihez a PCR reakció
hımérsékletét 55 oC–ról 93 oC–ra emeltünk és folyamatosan, fél percenként mértük a SYBR-
Green festék floureszcencia jelét. A PCR termékre jellemzı olvadáspont meghatározásával a
keletkezett amplikont ellenıriztük. Minden primerpárhoz használtunk egy negatív kontrollt,
ahol a cDNS helyett vizet tettünk a reakcióelegybe, ellenırizve a primer dimer képzıdést.
Statisztikai analízis
A beteg-kontroll asszociációs vizsgálatoknál alapvetı fontosságú, hogy a biallélikus
polimorfikus pontok által meghatározott allél- és genotípus gyakoriság eloszlások Hardy-
Weinberg egyensúlyban (HWE) legyenek. A HWE fennállását a „Hardy-Weinberg
equilibrium calculator including analysis for ascertainment bias” web program
segítségével igazoltuk (http://www.oege.org/software/hwe-mr-calc.shtml) (Rodriguez et al.,
2009).
Pearson-féle χ 2 vagy Fisher egzakt teszt segítségével hasonlítottuk össze a LEP G-2548A és
az IL6 G-174C polimorfizmusok elıfordulási gyakoriságát a beteg és a kontroll
populációban, meghatároztuk az egyes genotípusokhoz tartozó esélyhányadosokat (OR, odds
ratio) és a hozzájuk tartozó 95%-os megbízhatósági tartományt (CI, confidence intervals).
A polimorfikus SNP-k közötti kölcsönhatások vizsgálatára két módszert használtunk. Az
MDR (Multifaktoros Dimenzionalitás Redukció) analízis egy genetikai modell-mentes, nem

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
44
paraméteres eljárás amihez Jason Moore Számítástechnikai Genetika Laborja által fejlesztett,
szabadon felhasználható szoftvercsomagot (MDR 2.0 Beta 6) használtuk és a következı
webcímrıl töltöttük le: http://www.multifactordimensionalityreduction.org (Hahn et al.,
2003b).
A másik módszer egy paraméteres eljárás, ami a génkombinációk hatásának ML (maximum
likelihood) becslését alkalmazza annak a kimutatására, hogy az adott génkombináció milyen
mértékben emeli meg a betegség kialakulásának esélyét a legkevésbé hajlamosító genotípus
kombinációhoz viszonyítva (Yamada et al., 2001). A relatív esélyhányadosok meghatározását
és kereszttáblába történı rendezését Hirose és mtsai közleménye alapján végeztük el (Hirose
et al., 2008).
A GLM (Generalizált Lineáris Model) analízist használtuk arra, hogy kimutassuk létezik-e
valamilyen összefüggés a Cobb˚ nagysága és a vizsgált két funkcionális SNP által
meghatározott genotípus, vagy kettıjük genotípus kombinációi között.
Az elızıekben említetteken kívül a statisztikai elemzésekhez az SPSS 16.0 for Windows
szoftvercsomagot használtuk. A p<0,05 statisztikai teszteredményeket tekintettük
szignifikánsnak.
A munka során használt fontosabb online bioinformatikai eszközök és
adatbázisok internet címei:
• DAVID: http://niaid.abcc.ncifcrf.gov/
• GeneCards: http://www.genecards.org/
• KEGG PATHWAY Database: http://www.genome.jp/kegg/pathway.html
• MDR 2.0: http://www.multifactordimensionalityreduction.org
• NCBI: http://www.ncbi.nlm.nih.gov/
• Scoliosis Research Society: www.srs.org
• STRING: http://string.embl.de/

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
45
EREDMÉNYEK
Az idiopathiás scoliosis etiológiájának genetikai komponensét két megközelítéssel
vizsgáltuk. Az elsıben a gerincoszlop tartásában szereppel bíró paravertebrális izmok
génexpresszió szintő vizsgálata volt a cél, a másodikban pedig a génkifejezıdésben
szabályozó szerepet játszó promóter polimorfizmusok asszociációját néztük eset-kontroll
tanulmányban.
Paravertebrális izmok RNS kifejezıdési mintázatának vizsgálata CHIP
microarray-vel
Az elsı megközelítés a kontroll és az IS-es betegek paravertebrális izmainak génexpressziós
analízise volt és abból a feltevésbıl indult ki, hogy az idiopathiás scoliosisban jelenlévı
strukturális változások tükrözıdnek a génexpresszió szintjén is. A PTE ÁOK Ortopédiai
Klinikától 95 beteg mőtéti anyagából bilaterális musculus latissimus dorsi biopsziákat
kaptunk, amibıl teljes RNS preparátumot készítettünk. Az SZBK Funkcionális Genomika
munkacsoportjával (CHIP Labor) közösen két microarray analízist végeztünk el. Az elsıben
gerincferdüléses és egészséges személyekbıl származó mintákat, a másodikban pedig a
gerincferdüléses személyeknél a gerincoszlop két oldaláról vett mintákat hasonlítottunk
össze.
Egészséges és scoliosisos betegek izombiopsziájának génkifejezıdésbeli
összehasonlítása
A CHIP kísérlet során két párhuzamos pool-t készítettünk (IS I és IS II), melyek esetében
egyenként 6-6 korban, nemben és a scoliosisos görbülete fokában közel megegyezı személy
RNS mintáit kevertük össze. Ez a ún. ”pool„ kialakítás arra szolgál, hogy az egyedi
különbségeket elfedjük és a közös, vizsgálni kívánt jelenséghez kapcsolható változás
hangsúlyozottabban jelenhessen meg. A választásnál figyelembe vettük a közel azonos
életkort és a Cobb fokot. (2. táblázat) A kontroll csoport 4 traumás eredető és ékcsigolya
miatt mőtött egyén mintáit tartalmazta.
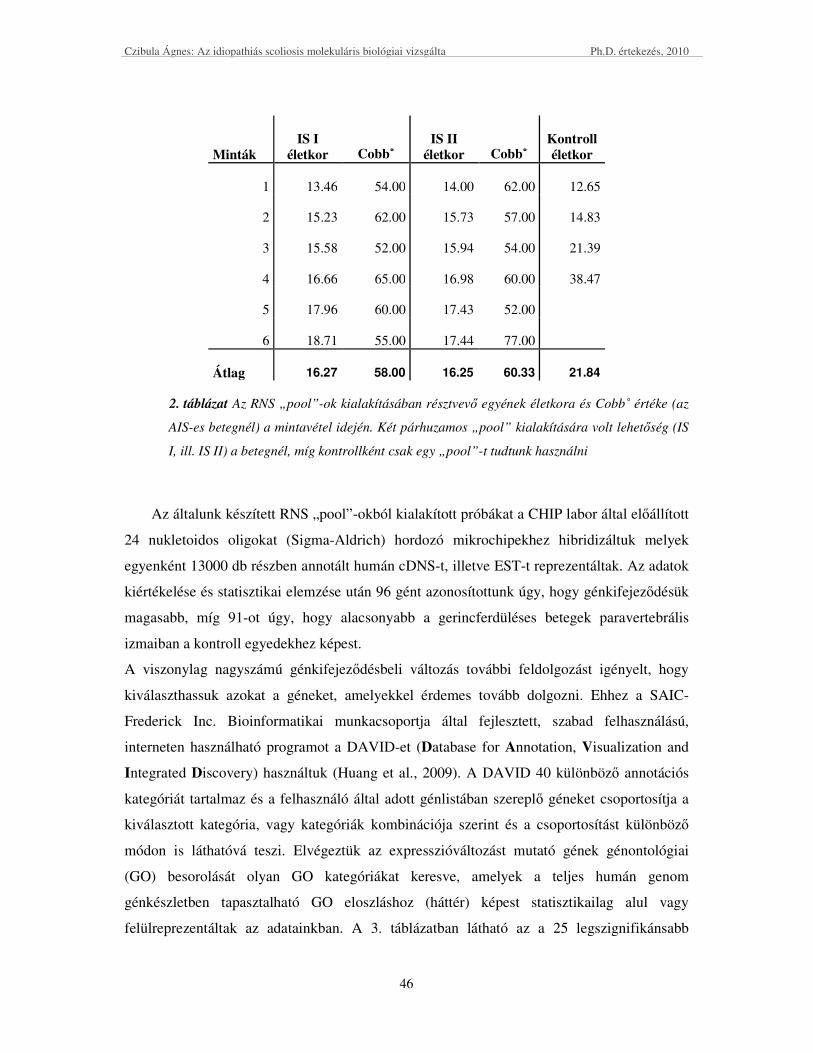
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
46
Minták IS I
életkor Cobb˚ IS II
életkor Cobb˚ Kontroll életkor
1 13.46 54.00 14.00 62.00 12.65
2 15.23 62.00 15.73 57.00 14.83
3 15.58 52.00 15.94 54.00 21.39
4 16.66 65.00 16.98 60.00 38.47
5 17.96 60.00 17.43 52.00
6 18.71 55.00 17.44 77.00
Átlag 16.27 58.00 16.25 60.33 21.84
2. táblázat Az RNS „pool”-ok kialakításában résztvevı egyének életkora és Cobb˚ értéke (az
AIS-es betegnél) a mintavétel idején. Két párhuzamos „pool” kialakítására volt lehetıség (IS
I, ill. IS II) a betegnél, míg kontrollként csak egy „pool”-t tudtunk használni
Az általunk készített RNS „pool”-okból kialakított próbákat a CHIP labor által elıállított
24 nukletoidos oligokat (Sigma-Aldrich) hordozó mikrochipekhez hibridizáltuk melyek
egyenként 13000 db részben annotált humán cDNS-t, illetve EST-t reprezentáltak. Az adatok
kiértékelése és statisztikai elemzése után 96 gént azonosítottunk úgy, hogy génkifejezıdésük
magasabb, míg 91-ot úgy, hogy alacsonyabb a gerincferdüléses betegek paravertebrális
izmaiban a kontroll egyedekhez képest.
A viszonylag nagyszámú génkifejezıdésbeli változás további feldolgozást igényelt, hogy
kiválaszthassuk azokat a géneket, amelyekkel érdemes tovább dolgozni. Ehhez a SAIC-
Frederick Inc. Bioinformatikai munkacsoportja által fejlesztett, szabad felhasználású,
interneten használható programot a DAVID-et (Database for Annotation, Visualization and
Integrated Discovery) használtuk (Huang et al., 2009). A DAVID 40 különbözı annotációs
kategóriát tartalmaz és a felhasználó által adott génlistában szereplı géneket csoportosítja a
kiválasztott kategória, vagy kategóriák kombinációja szerint és a csoportosítást különbözı
módon is láthatóvá teszi. Elvégeztük az expresszióváltozást mutató gének génontológiai
(GO) besorolását olyan GO kategóriákat keresve, amelyek a teljes humán genom
génkészletben tapasztalható GO eloszláshoz (háttér) képest statisztikailag alul vagy
felülreprezentáltak az adatainkban. A 3. táblázatban látható az a 25 legszignifikánsabb
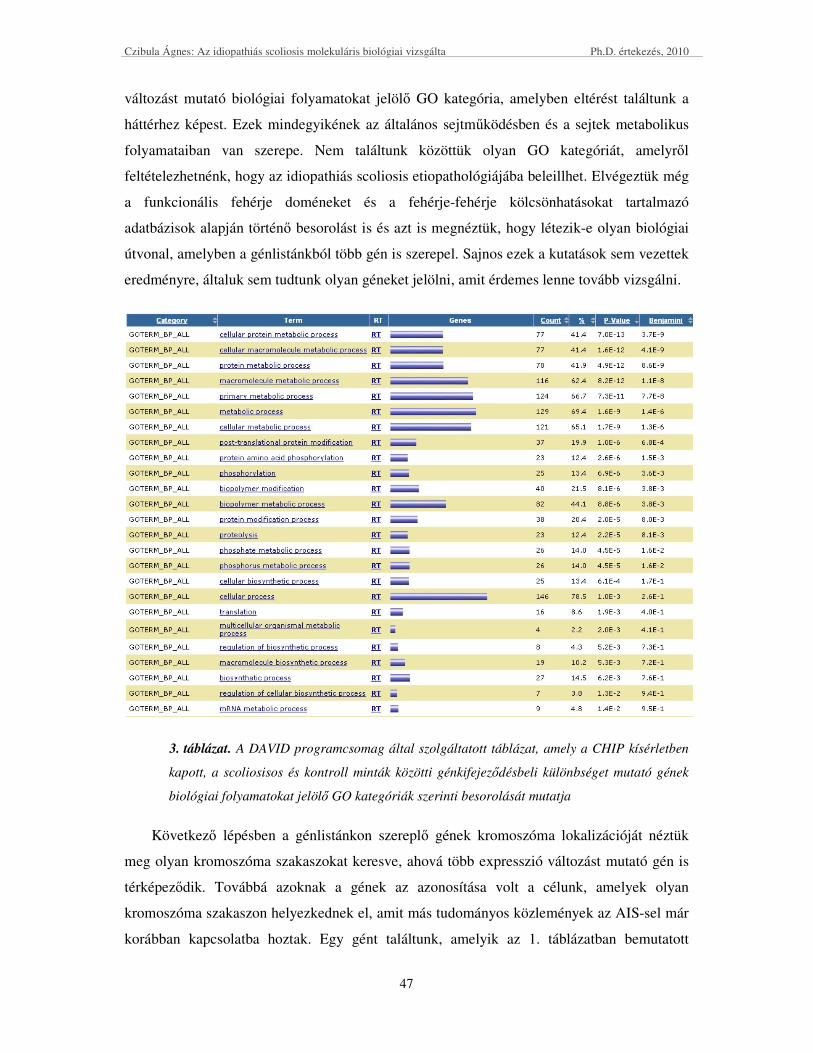
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
47
változást mutató biológiai folyamatokat jelölı GO kategória, amelyben eltérést találtunk a
háttérhez képest. Ezek mindegyikének az általános sejtmőködésben és a sejtek metabolikus
folyamataiban van szerepe. Nem találtunk közöttük olyan GO kategóriát, amelyrıl
feltételezhetnénk, hogy az idiopathiás scoliosis etiopathológiájába beleillhet. Elvégeztük még
a funkcionális fehérje doméneket és a fehérje-fehérje kölcsönhatásokat tartalmazó
adatbázisok alapján történı besorolást is és azt is megnéztük, hogy létezik-e olyan biológiai
útvonal, amelyben a génlistánkból több gén is szerepel. Sajnos ezek a kutatások sem vezettek
eredményre, általuk sem tudtunk olyan géneket jelölni, amit érdemes lenne tovább vizsgálni.
3. táblázat. A DAVID programcsomag által szolgáltatott táblázat, amely a CHIP kísérletben
kapott, a scoliosisos és kontroll minták közötti génkifejezıdésbeli különbséget mutató gének
biológiai folyamatokat jelölı GO kategóriák szerinti besorolását mutatja
Következı lépésben a génlistánkon szereplı gének kromoszóma lokalizációját néztük
meg olyan kromoszóma szakaszokat keresve, ahová több expresszió változást mutató gén is
térképezıdik. Továbbá azoknak a gének az azonosítása volt a célunk, amelyek olyan
kromoszóma szakaszon helyezkednek el, amit más tudományos közlemények az AIS-sel már
korábban kapcsolatba hoztak. Egy gént találtunk, amelyik az 1. táblázatban bemutatott

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
48
kapcsoltsági analízisben jelöltként szerepelt, ez a 12 p13-as kromoszóma szakaszon található
NTF3 (neurotrofin 3) gén volt, ami egy neuronokra ható növekedési faktor (8. ábra). A
továbbiakban irodalmi kutatásokat végeztünk a microarray kísérletben szignifikáns
eredményt adó génekkel kapcsolatban, majd a vonatkozó GeneCards leírások és számos
publikáció elolvasása után 5 gént kiválasztottunk a további célzott vizsgálatok célpontjainak.
A kiválasztott gének a következık voltak: strukturális alkotókat kódoló gének: LMNA (lamin
A), COL15A1 (XV-ös típusú kollagén alfa 1 lánc); a VDR (D vitamin receptor); egy a
másodlagos jeltovábbításban szereplı kináz JAK1 (Janus kináz 1); sejtmembrán receptor;
IL4R (interleukin 4 receptor); a csontfejlıdésben szerepet játszó differenciációs faktor BMP4
(csont morfogenetikus protein 4); a Ca2+-szignálútban a kalmodulinnal kapcsolatban lévı
NOS1 (nitrogénmonoxid szintetáz 1). Ezeknek a géneknek a további, QRT-PCR-rel történı
vizsgálatára a fejezet egy késıbbi szakaszában még visszatérek.
8. ábra. Az ENSEMBL Genome browser által készített kariogram, ahol a CHIP kísérletben
kapott, a scoliosisos és kontroll minták közötti génkifejezıdésbeli különbséget mutató gének
kromoszóma lokalizációja látható grafikus ábrázolással. Piros nyíllal jelöltem azt a régiót,
amit az AIS-sel már korábban kapcsolatba hoztak.
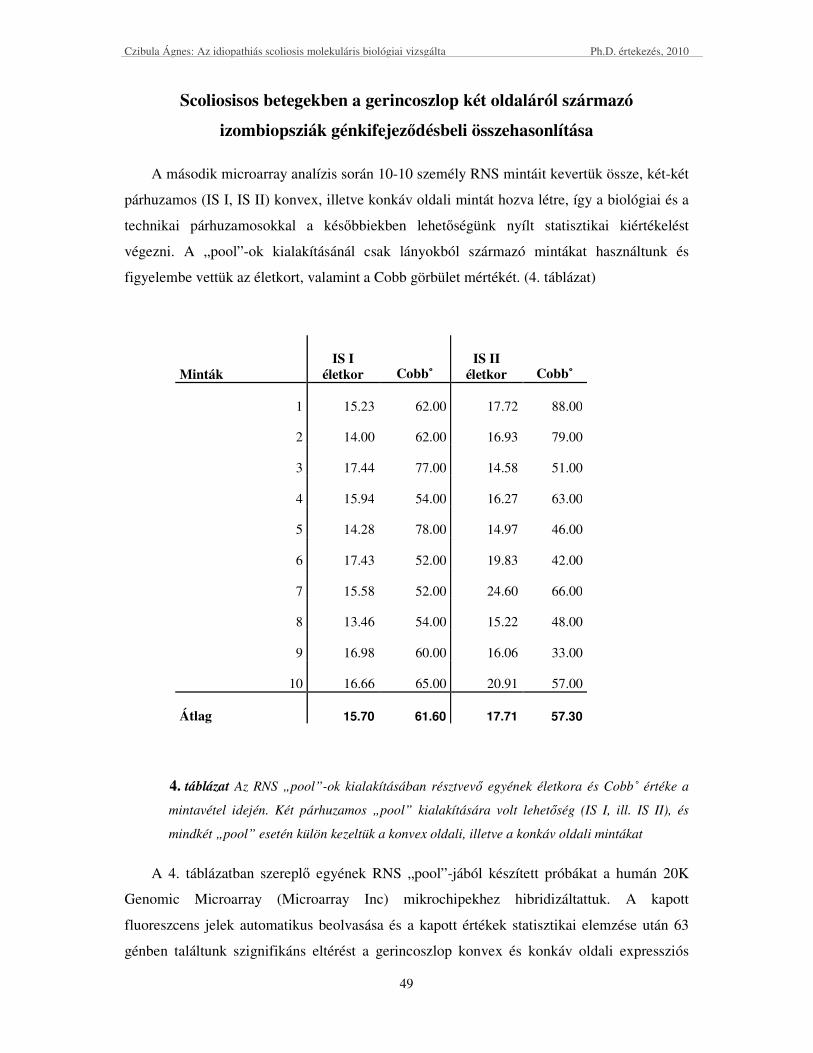
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
49
Scoliosisos betegekben a gerincoszlop két oldaláról származó
izombiopsziák génkifejezıdésbeli összehasonlítása
A második microarray analízis során 10-10 személy RNS mintáit kevertük össze, két-két
párhuzamos (IS I, IS II) konvex, illetve konkáv oldali mintát hozva létre, így a biológiai és a
technikai párhuzamosokkal a késıbbiekben lehetıségünk nyílt statisztikai kiértékelést
végezni. A „pool”-ok kialakításánál csak lányokból származó mintákat használtunk és
figyelembe vettük az életkort, valamint a Cobb görbület mértékét. (4. táblázat)
Minták IS I
életkor Cobb˚ IS II
életkor Cobb˚
1 15.23 62.00 17.72 88.00
2 14.00 62.00 16.93 79.00
3 17.44 77.00 14.58 51.00
4 15.94 54.00 16.27 63.00
5 14.28 78.00 14.97 46.00
6 17.43 52.00 19.83 42.00
7 15.58 52.00 24.60 66.00
8 13.46 54.00 15.22 48.00
9 16.98 60.00 16.06 33.00
10 16.66 65.00 20.91 57.00
Átlag 15.70 61.60 17.71 57.30
4. táblázat Az RNS „pool”-ok kialakításában résztvevı egyének életkora és Cobb˚ értéke a
mintavétel idején. Két párhuzamos „pool” kialakítására volt lehetıség (IS I, ill. IS II), és
mindkét „pool” esetén külön kezeltük a konvex oldali, illetve a konkáv oldali mintákat
A 4. táblázatban szereplı egyének RNS „pool”-jából készített próbákat a humán 20K
Genomic Microarray (Microarray Inc) mikrochipekhez hibridizáltattuk. A kapott
fluoreszcens jelek automatikus beolvasása és a kapott értékek statisztikai elemzése után 63
génben találtunk szignifikáns eltérést a gerincoszlop konvex és konkáv oldali expressziós
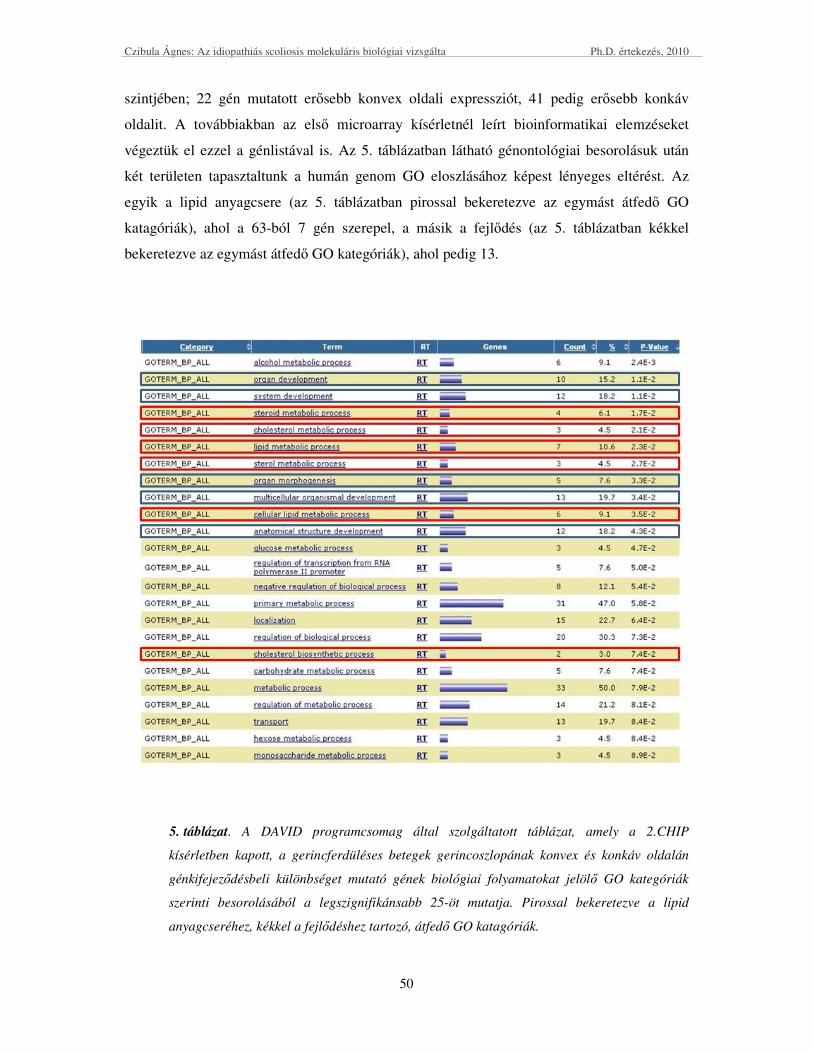
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
50
szintjében; 22 gén mutatott erısebb konvex oldali expressziót, 41 pedig erısebb konkáv
oldalit. A továbbiakban az elsı microarray kísérletnél leírt bioinformatikai elemzéseket
végeztük el ezzel a génlistával is. Az 5. táblázatban látható génontológiai besorolásuk után
két területen tapasztaltunk a humán genom GO eloszlásához képest lényeges eltérést. Az
egyik a lipid anyagcsere (az 5. táblázatban pirossal bekeretezve az egymást átfedı GO
katagóriák), ahol a 63-ból 7 gén szerepel, a másik a fejlıdés (az 5. táblázatban kékkel
bekeretezve az egymást átfedı GO kategóriák), ahol pedig 13.
5. táblázat. A DAVID programcsomag által szolgáltatott táblázat, amely a 2.CHIP
kísérletben kapott, a gerincferdüléses betegek gerincoszlopának konvex és konkáv oldalán
génkifejezıdésbeli különbséget mutató gének biológiai folyamatokat jelölı GO kategóriák
szerinti besorolásából a legszignifikánsabb 25-öt mutatja. Pirossal bekeretezve a lipid
anyagcseréhez, kékkel a fejlıdéshez tartozó, átfedı GO katagóriák.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
51
Az elemzések egy a KEGG Pathway adatbázisban szereplı biológiai útvonalat is a
felszínre hoztak, amelyben a génlistánkból 3 gén is szerepel. A KEGG Pathway (Kyoto
Encyclopedia of Genes and Genomes) adatbázis a Japán Humán Genom Program részeként
jött létre és a sejten belüli kölcsönhatások olyan győjteménye, amelyet folyamatosan
bıvítenek és magas szinten annotálnak így adatai a aktuális tudásunknak leginkább megfelelı
naprakész információkat hordozzák (Kanehisa et al., 2010). A 9. ábrán látható az adipocita
szignálút (KEGG_PATHWAY:hsa04920), aminek három tagja: LEP (Leptin), PRKAG2
(alfa 1 AMP-aktivált protein kináz katalitikus alegysége), SLC2A1 (szolubilis karrier család
2 4. tagja (facilitált glükóz transzporter)) mutatott szignifikáns génexpressziós eltérést a
gerincoszlop két oldalán. Mindhárom gén esetében a gerincferdülés konvex oldalán
magasabb génexpressziós szintet észleltünk, mint a konkávon.
9. ábra. A KEGG_PATHWAY:hsa04920 adipocita szignálút folyamatábrája. Piros csillag
jelöli azokat a géneket, amelyek expressziója eltérést mutat a gerincoszlop két oldala között a
gerincferdüléses betegekben.
Az elızı elemzésünkben, ahol a biológiai folyamatokat jelölı GO kategóriákba soroltuk a
génlistán lévı géneket (5. táblázat), a szignifikánsan (P<0.05) megjelenı génontológiai
kategóriák két fı csoportot határoztak meg, nevezetesen a lipid metabolizmust és a fejlıdési
folyamatokat. A leptin gén mindkét megnevezett biológiai folyamatban szerepet játszik. Az
irodalmi bevezetıben is ismertettem a leptin lehetséges kapcsolódását a gerincferdülés
kialakulásához, így ezt a gént mindenféleképp további vizsgálatra érdemesnek tartottuk.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
52
Irodalmi adatok alapján a fejlıdéssel kapcsolatba hozható GO kategóriába tartozó további
két, a microarray vizsgálatban szignifikánsnak mutatkozó gént, a HOX7-et (msh homeobox
1) és a CDK5-öt (ciklin-dependens kináz 5) is kiválasztottuk még célzott vizsgálatok
elvégzésére. A kromoszóma lokalizációs elemzéseket elvégezve hét olyan gént találtunk,
melyek olyan kromoszóma szakaszra esnek, amit más laboratóriumok kapcsoltsági
analízisben az AIS-sel összefüggésbe hoztak (ld. 1. táblázat). Ezek a gének a következık
voltak: GPR107 (G-proteinhez kapcsolódó receptor 107)-9q34.11, ATAD1 (ATP-áz családba
tartozó AAA domént tartalmazó fehérje 1)-10q23.2, APC2 (vastagbél adenomatózus
polipózis 2)-19p13.3, APLN (apelin)-Xq25, CDK5-7q36.1, ARMC7 (armadilló ismétlıdést
tartalmazó fehérje 7)-17q25.1, CPN3 (copine 3)-8q21.3. (10. ábra) Ezen felül a microarray
kísérletben szignifikáns eltérést mutató gének listáján szereplı összes gén GeneCards leírását
feldolgozva összefüggést próbáltunk találni a gének jelenleg ismert funkciója és az AIS
pathomechanizmusára vonatkozó jelenleg ismert modellek között. Két gént találtunk
érdekesnek, az izomerı meghatározásában szereplı a disztonint (DST) (Dalpe et al., 1999a)
és a sérült izmok regenerálódásában fontos tenzint (TNS1)(Ishii and Lo, 2001).
10. ábra. Az ENSEMBL Genome browser által készített kariogram, ahol a második CHIP
kísérletben kapott, a scoliosisos és kontroll minták közötti génkifejezıdésbeli különbséget mutató
gének kromoszóma lokalizációja látható grafikus ábrázolással. Piros nyíllal jelöltem azokat a
kromoszóma szakaszokat, amelyek korábbi a kapcsoltsági analízis során már kandidáns génként
felmerültek (1. táblázat).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
53
A microarray kísérletek mindig kiváló kiindulási alapként szolgálnak a génexpressziós
vizsgálatokban, mivel látótérbe hoznak a vizsgált jelenséggel funkcionális kapcsolatban álló,
de eddig az adott kontextusban még le nem írt géneket. Második lépésként azonban
mindenképp szükséges ellenırizni egy független módszerrel, hogy valóban fennáll-e a
megfigyelt génkifejezıdésbeli különbség. Ez a független módszer napjainkban általában
QRT-PCR.
Paravertebrális izmokban kifejezıdı mRNS-ek vizsgálata QRT-PCR-rel
A microarray vizsgálatok után az AIS-es betegek konvex és a konkáv oldali paravertebrális
izmainak génexpressziós analízisét elvégeztük kvantitatív valós idejő polimeráz láncreakció
(QRT-PCR) módszerrel is. Mivel minden egyes génhez külön-külön kell PCR reakciót
végezni ez technika már nem alkalmas nagyszámú gén vizsgálatára, így az elızıekben
ismertetett CHIP analízisekben a szignifikáns expresszióbeli különbséget mutató és az
irodalmi adatok alapján kiválasztott gének közül 13-t analizáltam tovább. Ezek a gének a
LMNA, VDR, COL15A1, JAK1, BMP4, IL4R, NTF3, LEP, CDK5, HOX7, NOS1, illetve a
NOS2 és a NOS3.
A QRT-PCR során a cDNS mintákat 4-6 személy RNS-ének elegyébıl állítottam elı. A
QRT-PCR során lehetıség van az amplifikációval egy idıben a PCR kinetikai-görbéjének
felvételére és a kiindulási mRNS mennyiségével arányos áttörési ciklusszám (threshold cycle,
Ct) meghatározására (Higuchi et al., 1993). A vizsgálni kívánt target gének expressziós
szintjét egy adott referencia gén, esetünkben az aktin, illetve a HPRT (hipoxantin
foszforibozil-transferáz 1) expressziós szintjére vonatkoztatjuk azért, hogy
összehasonlíthassuk a gerincoszlop két oldalán a target gén kifejezıdést. A komparatív ∆Ct-
módszer alkalmazásával meghatároztuk azokat a géneket, ahol legalább kétszeres különbség
van a két oldali izombiopszia közt a relatív mRNS szint tekintetében, vagyis a ∆Ctkonvex-
∆Ctkonkáv 1-nél nagyobb, vagy -1-nél kisebb. A 13 gén közül egyedül a leptin esetén tudtunk
kimutatni eltérést, a konvex oldalon a leptin relatív mRNS mennyisége 2,5-szerese a
konkávénak. A 11. ábrán pirossal jelölve láthatók a leptinnél kapott értékek.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta
Ct átlag konvex
LMNA VDR COL15A1 AKT JAK1 19.615HPRT BMP4 21.685IL4R 22.325NTF3 26.275HPRT 21.505LEP CDK5 HOX7 HPRT NOS1 NOS2 25.795NOS3 21.945HPRT 20.045
11. ábra. Konvex és konkáv oldal közötti RNS expresszió
alkalmazásával 13 gén esetén. Az
ciklusszámok átlagát (Ct), a belı
ciklusszámot (∆ Ct) és a konvex és konkáv oldal közötti ci
Az ábra B. részén az A. rész
szemléletesen látható, hogy csak a LEP gén esetén található több, mint egy ciklusszám különbség,
amit konvencionálisan valós különbségnek lehet tekinteni a QRT
LMNA
VDR
COL15A1
JAK1
-1
-0.5
0
0.5
1
1.5
∆ ∆
Ct
konv
ex-k
onká
v
A
B
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
54
Ct átlag konvex
Ct átlag konkáv
∆ Ct konvex
∆ Ct konkáv
∆∆Ct konvex-konkáv
18.86 19.3 2.095 2.705 -0.61 24.52 24.37 7.755 7.775 -0.02 17.89 17.99 1.12 1.395 -0.275 16.77 16.59 0 0 0
19.615 19.49 -2.165 -2.405 0.24 21.78 21.895 0 0 0
21.685 22.56 0.18 0.82 -0.64 22.325 22.9 0.82 1.16 -0.34 26.275 26.035 4.77 4.295 0.475 21.505 21.74 0 0 0 21.53 20.16 1.69 0.37 1.32 21.33 21.55 1.49 1.76 -0.27
27.8 28.33 7.96 8.54 -0.58 19.84 19.79 0 0 0 21.45 21.37 1.405 0.97 0.435
25.795 26.25 5.75 5.85 -0.1 21.945 22.345 1.9 1.945 -0.13 20.045 20.4 0 0 0
Konvex és konkáv oldal közötti RNS expresszió különbség meghatározása a
Az ábra A. részén két párhuzamos QRT-PCR-bıl származó kitörési
(Ct), a belılük számított, aktin, illetve HPRT referenciagénhez normált
és a konvex és konkáv oldal közötti ciklusszám különbséget (∆∆ Ct) adtam meg
utolsó oszlopában lévı értékeket ábrázoltam grafikusan, így
szemléletesen látható, hogy csak a LEP gén esetén található több, mint egy ciklusszám különbség,
amit konvencionálisan valós különbségnek lehet tekinteni a QRT-PCR használatakor
COL15A1
JAK1
BMP4
IL4R
NTF3
LEP
CDK5
HOX7
NOS1
NOS2 NOS3
Ph.D. értekezés, 2010
a ∆Ct-módszer
l származó kitörési
HPRT referenciagénhez normált
Ct) adtam meg.
értékeket ábrázoltam grafikusan, így
szemléletesen látható, hogy csak a LEP gén esetén található több, mint egy ciklusszám különbség,
NOS3

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
55
Polimorfikus helyek vizsgálata
Annak ellenére, hogy több tanulmány is megjelent a paravertebrális izmok vizsgálatáról,
amelyekben a két oldal között eltérı strukturális, fehérjeszintbeli és génkifejezıdési
mintázatot írtak le, a tudományos közvélemény inkább a kialakult gerincferdülés
következményének, nem pedig az okának tartja a megfigyelt különbségeket. Amennyiben a
megfigyelt eltérések genetikai okaira sikerül fényt deríteni, akkor jelenthetı ki, hogy a
betegség kóroki tényezıi, nem pedig a gerincferdülés következtében kialakult eltérések a
megfigyelt jelenségek. Több strukturális gén kódoló szekvenciájában lévı mutációt
vizsgáltak eddig, de nem találtak eltérést az AIS-es betegekben. Egyre több humán betegség
kapcsán derül ki, hogy a gének szabályozó régióiban lévı nukleotid eltéréseknek szerepe
lehet az adott betegség kialakításában, így mindenképp indokoltnak láttuk, hogy a leptin gén
esetén megvizsgáljuk annak az SNP-nek az elıfordulási gyakoriságát, amelynek szerepe lehet
a leptin gén szabályozásában. Egy 2007-ben megjelent közleményben bemutatták, hogy az
IL6 promóter régiójában - a transzkripció szabályozásban érintett - polimorfikus hely
asszociációt mutat a gerincferdüléssel, amit mi is megpróbálunk igazolni (Aulisa et al., 2007).
Mivel mindkét vizsgálni kívánt SNP olyan gének szabályozásában vehet részt, amelyek az
AIS-sel kapcsoltba hozható, zsírszövetben termelıdı adipokint kódolnak, szükségesnek
láttuk a két polimorfikus hely kombinatorikus analízisét is elvégezni.
Leptin promóter G-2548A polimorfizmusa
Eset-kontroll asszociációs vizsgálatainkban 126 idiopathiás scoliosisos beteg és 197
egészséges, egymással nem rokon személy esetében határoztuk meg a leptin gén G-2548A
(rs7799039) SNP genotípus eloszlását PCR-RFLP módszerrel. A genotípus eloszlás alapján
mindkét vizsgált csoport Hardy-Weinberg egyensúlyban volt. A genotípus eloszlást a 6.
táblázat A. része mutatja. Mivel a nagyfokú görbülettel rendelkezı AIS-es betegek nagyrésze
lány volt - ami egyezik az irodalmi adatokkal – külön vizsgáltuk a genotípus és allél eloszlást
ebben a csoportban is ( 6. táblázat B. része). A Hardy-Weinberg egyensúly továbbra is
fennállt. Az A allél elıfordulási gyakoriságának értéke magasabb a beteg csoportban (0.48
vs. 0.42), de a statisztikai értékelés azt mutatta, hogy sem az allélgyakoriság, sem a genotípus
gyakoriság szignifikánsan nem tér el a beteg és a kontroll csoport között (χ2=1.705, p=0.192).
A relatív kockázatbecslést elvégezve azt kaptuk, hogy az A allél jelenléte hajlamosít a
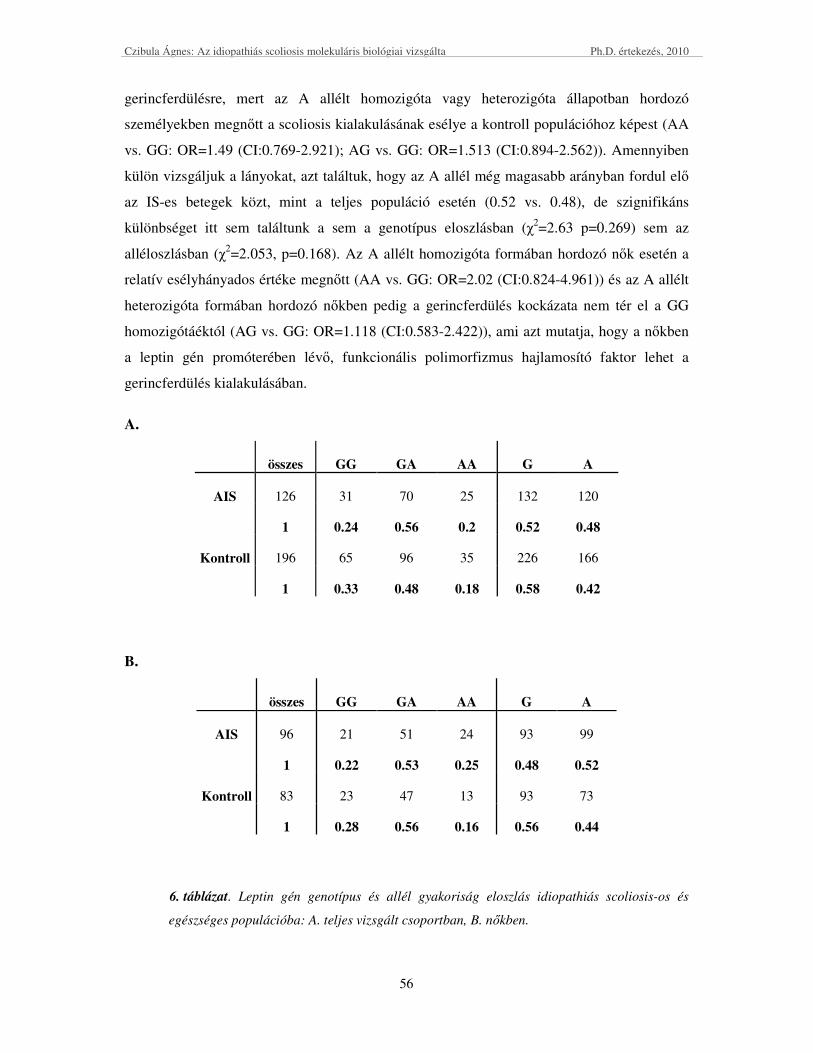
Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
56
gerincferdülésre, mert az A allélt homozigóta vagy heterozigóta állapotban hordozó
személyekben megnıtt a scoliosis kialakulásának esélye a kontroll populációhoz képest (AA
vs. GG: OR=1.49 (CI:0.769-2.921); AG vs. GG: OR=1.513 (CI:0.894-2.562)). Amennyiben
külön vizsgáljuk a lányokat, azt találtuk, hogy az A allél még magasabb arányban fordul elı
az IS-es betegek közt, mint a teljes populáció esetén (0.52 vs. 0.48), de szignifikáns
különbséget itt sem találtunk a sem a genotípus eloszlásban (χ2=2.63 p=0.269) sem az
alléloszlásban (χ2=2.053, p=0.168). Az A allélt homozigóta formában hordozó nık esetén a
relatív esélyhányados értéke megnıtt (AA vs. GG: OR=2.02 (CI:0.824-4.961)) és az A allélt
heterozigóta formában hordozó nıkben pedig a gerincferdülés kockázata nem tér el a GG
homozigótáéktól (AG vs. GG: OR=1.118 (CI:0.583-2.422)), ami azt mutatja, hogy a nıkben
a leptin gén promóterében lévı, funkcionális polimorfizmus hajlamosító faktor lehet a
gerincferdülés kialakulásában.
A.
összes GG GA AA G A
AIS 126 31 70 25 132 120
1 0.24 0.56 0.2 0.52 0.48
Kontroll 196 65 96 35 226 166
1 0.33 0.48 0.18 0.58 0.42
B.
összes GG GA AA G A
AIS 96 21 51 24 93 99
1 0.22 0.53 0.25 0.48 0.52
Kontroll 83 23 47 13 93 73
1 0.28 0.56 0.16 0.56 0.44
6. táblázat. Leptin gén genotípus és allél gyakoriság eloszlás idiopathiás scoliosis-os és
egészséges populációba: A. teljes vizsgált csoportban, B. nıkben.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
57
IL6 promóter C-174G polimorfizmusa
Szintén PCR-RFLP módszerrel határoztuk meg az IL6 promóter régiójában lévı,
funkcionális SNP allélgyakoriságát is. A genotípusok eloszlása szerint fennállt a Hardy-
Weinberg egyensúly. Sem az allél, sem a genotípus eloszlásban nem találtunk szignifikáns
eltérést a gerincferdüléssel élık és az egészséges kontroll személyek között (χ2=0.779,
p=0.378) (7. táblázat). A relatív kockázatbecsléseket elvégezve a különbözı genotípusok
vonatkozásában az esélyhányados 1 körüli értékeket mutatott, az 1-tıl történı legnagyobb
eltérés a homozigóta párok összehasonlításakor mutatkozott GG vs. CC: OR=0.765 (CI:
0.396-1.468), ami alapján a GG genotípus protektívnek mondható. Külön értékelve a lányok
szubpopulációját nem kaptunk eltérést a teljes populációhoz képest, sem az
allélgyakoriságban, sem a relatív esélyhányados mértékében (GG vs. CC: OR=0.594 (CI:
0.242-1.457)).
A.
összes CC GC GG C G
AIS 126 25 67 34 117 135
1 0.2 0.53 0.27 0.46 0.54
Kontroll 197 36 97 64 169 225
1 0.18 0.49 0.33 0.43 0.57
B.
összes CC GC GG C G
AIS 96 21 48 27 90 102
1 0.22 0.5 0.28 0.47 0.53
Kontroll 83 12 46 25 70 96
1 0.15 0.55 0.3 0.42 0.58
7. táblázat. IL6 gén genotípus és allél gyakoriság eloszlása idiopathiás scoliosis-os és
egészséges populációban: A. teljes vizsgált csoportban, B. nıkben.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
58
12. ábra. Az MDR analízis az összes genotípus
kombinációt besorolja a betegség kialakulása
szempontjából alacsony (világosszürke hátterő)
vagy magas (sötétszürke hátterő) kockázatot jelentı
kategóriába. Minden egyes négyzetben a bal oldali
oszlop a scoliosisos a jobb oldali pedig a kontroll
személyek eloszlását mutatja.
A két polimorfizmus IS kialakulására hajlamosító együttes hatásának vizsgálata
Multifaktoriális öröklıdéső betegségek esetén több gén eltérı hozzájárulással a
környezettel kölcsönhatásban vesz részt a legtöbbször heterogén fenotípus kialakításában, így
2-3 gén egy adott allélkombinációja, legalábbis az esetek egy részében, hozzájárulhat a
betegség etiológiájához. A gén-gén kölcsönhatás feltárásához két eljárást is használtunk, az
elsı az MDR analízis, amely kisszámú mintaelem esetén is kimutatja az episztázist és a
módszer kidolgozói rámutattak, hogy akkor is alkalmas a gének közötti szinergizmus
kimutatására, ha az allél eloszlásban nem mutatkozik eltérés a beteg-kontroll vizsgálat során
(Hahn et al., 2003a). A génkombinációk hatásának maximum likelihood becslését alkalmazza
a másik statisztikai módszer és
megadja, hogy egy adott
génkombináció milyen mértékben
emeli meg a betegség kialakulásának
esélyét egy másik
génkombinációhoz viszonyítva.
Az MDR analízis a LEP promóter
SNP és az IL6 promóter SNP között
szinergiát jelzett. A 12. ábrán a
kereszttáblán összerendezett
genotípus kombinációk láthatók,
minden egyes négyzetben bal oldali
oszlop a scoliosisos jobb oldali pedig
a kontroll személyek eloszlását
mutatja. Az MDR analízis
végeredményként minden genotípus
kombinációt besorol vagy alacsony,
vagy magas kockázatot jelentı
csoportba. A grafikus ábrázolás
szerint világosszürke kitöltéssel az
alacsony, míg sötétszürke háttérrel a
magas rizikójúakat jelölve.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
59
Mivel az MDR analízis arra alkalmas, hogy felfedje az episztázist és megadja azokat a
genotípus kombinációkat, amelyek hordozása kockázatot jelent az adott betegség
kialakulására, további vizsgálatot végeztünk annak a megállapítására, hogy milyen mértékő
kockázatot jelentenek az egyes genotípus kombinációk a gerincferdülés esetén. Ehhez a LEP
G-2548A és az IL6 C-174G polimorfizmus genotípusait páronként összerendeztük és relatív
kockázat közelítı becslését végeztük el a legkevésbé hajlamosító genotípus kombinációhoz
képest adva meg az esélyhányadosokat. Az elızı fejezetekben leírtak alapján a LEP gén
esetében az A allél hordozása jelentett magasabb kockázatot, ezért a GG homozigóta lehet a
legkevésbé hajlamosító genotípus, míg az IL6 esetén a GG genotípus protektívnek bizonyult.
Így a LEP GG-IL6 GG genotípus kombináció lehet a legkevésbé hajlamosító, amit alátámaszt
az MDR analízis is, mivel alacsony kockázattal bírónak jelölte ezt a genotípus kombinációt.
A relatív esélyhányadosok kiszámolásánál ez volt a referencia, amit 1.00-es szám jelöl a 8.
táblázatban. A LEP GG-IL6 GG genotípus kombinációhoz viszonyítva a leginkább
hajlamosító kombináció az IL6 CC-LEP AA, ami azt jelenti, hogy a hajlamosító genotípust
hordozó egyénnél szignifikánsan (χ2=5.511, p=0.019), közel ötször nagyobb (OR=4.667
(CI:1.237-17.6)) az esélye a gerincferdülés kialakulásának, mint a referenciának tekintett
genotípussal bíró személyé.
Az MDR analízis és a ML becslésen alapuló kombinációs hatás vizsgálat eredményei jól
összeegyeztethetık, mivel azokat a genotípus kombinációkat jelölte magas rizikót jelentınek
az elsı vizsgálat, amelyekben a relatív OR értéke magasabb volt 1.5-nél.
8. táblázat Az allélkombinációk kombinatorikus hatásának kimutatása a relatív
esélyhányadosok megadásával
LEP G-2548A
GG GA AA
CC 0.848 (0.212-3.391) 1.283 (0.439-3.752) 4.667 (1.237-17.600)
GC 1.273 (0.483-3.355) 1.887 (0.775-4.595) 1.51 (0.508-4.484)
GG 1.00 1.633 (0.626-4.264) 0.787 (0.183-2.814) IL6
C-1
74G

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
60
Amennyiben a relatív esélyhányadost meghatározzuk csak a lányok esetén, akkor azt kapjuk,
hogy a leginkább hajlamosító allélkombináció (IL6 CC-LEP AA) hordozása még nagyobb
kockázatot jelent a gerincferdülés kialakulásában (OR= 6.667 (CI:0.597-74.506). A relatív
esélyhányadoshoz tartozó konfidencia intervallum elég széles, ami az esélyhányados
értékének megbízhatóságát rontja és ez egyértelmően az alacsony mintaszámnak tudható be.
A polimorfizmusok hatása az IS súlyosbodására
További statisztikai elemzéseket végeztünk annak felderítésére, hogy a LEP promóter G-
2548A polimorfizmusa, illetve az IL6 promóter C-174T polimorfizmusa szerepet játszik-e a
gerincferdülés progressziójában. Nem találtunk szignifikáns összefüggést sem a LEP -
p=0.712 (ANOVA) -, sem az IL6 - p=0.586 (ANOVA) – polimorfizmusa és a Cobb˚ között.
Meghatároztuk minden egyes genotípus kombinációhoz tartozó átlagos Cobb˚-ot és nem
találtunk olyan genotípus kombinációt, amelyhez az összes adatból számolt átlagtól
szignifikánsan eltérı Cobb˚ társult volna. A fentiek alapján úgy gondoljuk, hogy a LEP AA-
IL6 CC genotípus kombináció a betegség kialakításában inkább hajlamosító, nem az AIS
súlyosbodásában vehet részt.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
61
EREDMÉNYEK MEGVITATÁSA
A komplex betegségek etiológiájának feltárása a XXI. század kutatóinak egyik
legnagyobb kihívása. A mendeli öröklıdésmenetet követı monogénes betegségek genetikai
hátterét a rendelkezésre álló módszerekkel jórészt feltárták, vagy nagy lépésekkel haladnak e
cél elérése felé. Ezzel ellentétben az emberek többségét érintı multifaktoriális, poligénes
betegségek genetikai elemeinek azonosítása eddig nem hozott áttörı eredményt.
Természetesen tudtak azonosítani hajlamosító allélokat különbözı betegségekben, de csak
azoknál, ahol az adott allél jelenléte nagymértékben hozzájárult a betegség kialakulásához.
Jelen dolgozatban a serdülıkori populáció 10%-át érintı, ismert ok nélkül kialakuló
gerincferdülés genetikai hátterében álló elemek felkutatására vonatkozó vizsgálataimat írtam
le. Két megközelítést használtunk a munkánk során. Az elsı megközelítésünk szerint az AIS
etiológiájában szerepet játszó gének (kandidáns gén) kiválasztása a transzkriptoma szintjén
jelenlévı eltéréseken alapult. Vizsgálatainkra a gerincoszlop két oldalán elhelyezkedı izmok
RNS mintáit választottuk, amit a korrekciós mőtét során kivett izombiopsziákból nyertünk. A
paravertebrális izmok génexpressziós változásait vizsgáltuk microarray analízissel két
különbözı összehasonlítással. Az elsı CHIP kísérletben egészséges és AIS-es betegek külön-
külön „pool”-ozott mintáit vetettük össze és 187 génben tudtunk génexpresszióbeli eltérést
detektálni. A másodikban a kontroll minták kihagyásával csak az AIS-es betegek konvex és
konkáv oldali izmainak expressziós szintő összehasonlítását végeztük el és 63 génben
tudtunk különbséget kimutatni. A microarray kísérletek az elsı szőrıt jelentették a jelölt gén
kiválasztásában, a másodikat bioinformatikai adatelemzéssel végeztük el.
Meghatároztuk a génlistáinkon lévı gének kromoszóma lokalizációját és két szempont
szerint is átnéztük a gének elhelyezkedését. Egyrészrıl kerestünk olyan kromoszóma
szakaszokat, ahová több gén is térképezıdik közel egymáshoz, mert egy helyen lévı és
egyforma expressziós eltérést mutató gének esetén gondolhatunk valamiféle genomszintő
átrendezıdésre az adott régióban (delécióra vagy inszercióra). Ilyet nem tudtunk kimutatni.
Másrészrıl, azokat a géneket a késıbbiekben mindenféleképpen érdemes tovább vizsgálni,
amelyek olyan kromoszóma szakaszra esnek, amit családokon kapcsoltsági analízissel az
AIS-hez kötöttek (1. táblázat). Nyolc ilyen gént találtunk, amelyek közül kettınek (NTF3 és
CDK5) meghatároztuk a relatív mRNS mennyiségét az AIS-es betegek gerincoszlopának
konvex és konkáv oldalán QRT-PCR-rel és nem tudtunk különbséget kimutatni a két oldal

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
62
között. A többi 6 gén további vizsgálata még az elkövetkezendı idıszak hátralévı feladatai
közé tartozik.
A két elvégzett microarray egy 187, illetve egy 63 tagból álló génlistát eredményezett,
melyek tagjait a génontológiai kategóriákhoz rendeltük, olyan GO kategóriákat keresve,
amelyekben a génjeink a humán genom átlagos GO eloszlásához képest magasabb arányban
reprezentáltak és szerepük lehet az AIS kialakításában. Az elsı CHIP kísérletbıl nem tudtunk
ilyen kategóriát kiemelni, de a másodikból a lipid anyagcsere (GO:0006629) és a
morfogenezis (GO:0007275) is felvetıdött; ebbe a két katagóriába a 63 génbıl összesen 18-at
tudtunk besorolni. A lipid anyagcsere érintettségét az AIS-ben alátámaszthatja az a tény is,
hogy az adipociták szignálútjának (KEGG_Pathway: hsa04920) több elemében is (LEP,
PRKAG2, SLC2A1) ki tudtuk mutatni a génexpressziós különbséget a második microarray
analízisben, amikor a konvex és konkáv oldali paraspinális izmok transzkriptomáit
hasonlítottuk össze. A leptin szerepének további vizsgálatához elıször egy független
módszerrel, a QRT-PCR-rel meghatároztuk az AIS betegekre jellemzı génexpressziós
szintet. Az így kapott eredmény megerısítette a microarray analízisben megfigyelt értéket,
vagyis a LEP génrıl az izmokban termelıdı mRNS mennyisége a konvex oldalon 2.5-
szerese a konkávénak. Ahhoz, hogy minél kevesebb információt veszítsünk el a microarray
adatokból egy a tudományos szakirodalmi adatokon alapuló szőrıt is beépítettünk
vizsgálatainkba, azaz a listán szereplı gének közül az adatbázisokban és a rendelkezésre álló
publikációs forrásokban fellelhetı adatok tanulmányozása után kiemeltük azokat,
amelyeknek szerepük lehet az AIS etiológiájában. Egy részüket ezidáig már sikerült tovább
célzott vizsgáltnak alávetni arra vonatkozóan, hogy megerısíthetıek-e a microarray
analízisben kapott eredmények. A génlistánk másik részének esetében azonban ezek a
vizsgálatok még elıttünk állnak. QRT-PCR-módszert használtuk, annak a kimutatására, hogy
az AIS-es betegek paravertebrális izmaiban tapasztalható-e különbség az adott gének
expressziójában attól függıen, hogy a konvex vagy a konkáv oldalról származtak-e. A már az
elızıekben említett leptin kivitelével nem találtunk több gént, ahol szignifikáns expressziós
különbség állt volna fenn. Az irodalomban eddig egy közlemény jelent meg a paravertebrális
izmok molekuláris biológiai vizsgálatáról, ahol Western-blottinggal a kalmodulin és a NOS1
fehérje mennyiségét vizsgálták a görbült gerincoszlop két oldaláról származó sejtlizátumban.
Mind az egészséges kontrollhoz, mind pedig a konkáv oldalhoz képest csökkent fehérje
mennyiségeket találtak a konvex oldalon (Zhao and Qiu, 2004). Ennek az eredménynek a
kapcsán vizsgáltuk meg mindhárom nitrogénmonoxid szintetáz mRNS mennyiségét, mivel az
elsı microarray analíziskor mi is azt tapasztaltuk, hogy a kontrollhoz képest csökkent a

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
63
NOS1mRNS mennyisége az AIS-es betegben. Ezt az eltérést azonban nem tudtuk kimutatni
az AIS-es betegekben a konvex és konkáv oldali génexpresszió vonatkozásában.
A microarray analízisben kapott és az AIS kialakulásának lehetséges okai közé
beilleszthetı gének közül további vizsgálatra érdemesek azok a gének, amelyek a
kapcsoltsági vizsgálatok által jelölt kromoszóma szakaszra esnek. Mivel a paraspinális izmok
megfelelı izomereje nagyon fontos a gerincoszlop támasztásában és az AIS-es betegekben
valóban kimutattak eltéréseket a konvex és a konkáv oldali izomaktivitásban, azok a gének,
amelyek az izmok izomerejének kialakításában vesznek részt jelöltek lehetnek a betegség
súlyosbodásáért felelıs progressziós génként az AIS etiológiájában (Cheung et al., 2005a).
Az irodalmi adatok alapján a disztonin kandidáns lehet az AIS progressziós génjeként, mivel
a desmin és az aktin szálak keresztkötésével az izomsejtek struktúrájának stabilizálásában
van fontos szerepe. A disztonin hiányos egerek izomgyengeséget mutatnak (Dalpe et al.,
1999b). Az AIS-es betegek paravertebrális konvex és konkáv oldali izmainak expresszióját
összehasonlítva a disztonin gén mRNS mennyisége kétszer magasabb a konvex oldalon, mint
a konkávon a CHIP hibridizálás eredménye szerint, viszont ezt az eredményt független
módszerrel a késıbbiekben igazolni kell.
A génkifejezıdés szabályozása az egyik nagyon sokat tanulmányozott területe a molekuláris
biológiának. A transzkripciós faktorok, a cisz regulációs elemek és a kromatin átszervezıdés
génszabályozásban betöltött szerepe régóta a sejt és molekuláris biológia egyik
legintenzívebben kutatott területe. A komplex betegségek vonatkozásában az elmúlt
idıszakban egyre nagyobb figyelem fordul a génkifejezıdés módosulatai mögött álló
allélspecifikus transzkripció felé. Többen is kimutatták, hogy a humán gének 20 %-ában a
gén két alléljáról nem egyforma mértékben fejezıdnek ki a géntermékek, aminek az egyik
oka a gének promóter régiójában található, funkcionális polimorfizmus lehet (Lo et al., 2003).
A leptin gén promóter régiójában is található olyan SNP, aminek hatása lehet a
génkifejezıdésre, amint részletesen kifejtve a dolgozat bevezetıjében leírtam. A dolgozatban
bemutatott munka során megvizsgáltuk, hogy az általunk detektált génexpressziós
különbségben szerepet játszik-e a LEP G-2548A polimorfizmus. Az irodalmi adatok alapján
az összefüggés a LEP gén alléljei és a vérszérumban mért leptin szint között ellentmondó,
valószínőleg a köztük lévı kapcsolatot más tényezık is befolyásolják (Yiannakouris et al.,
2003). Egy eset-kontroll tanulmány során meghatároztuk a LEP genotípusok elıfordulási
gyakoriságát mindkét populációban és kiszámoltuk a különbözı genotípusok
esélyhányadosait. Nem kaptunk szignifikáns eltérést a két populáció között a genotípus
gyakoriságok tekintetében, de az esélyhányadosok meghatározása után úgy tőnik, hogy az A

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
64
allél jelenléte lehet hajlamosító faktor az AIS-ben. Vizsgálatainkban az AA vs. GG:
OR=1.513(CI:0.894-2.562), ami komplex betegségek esetén a hajlamosító allélokra jellemzı
értéknek tekinthetı (Baron, 2001).
Kíváncsiak voltunk arra, milyen más tényezı befolyásolja a LEP -2548A allél hatását.
Elıször a szérum leptinszint nemek közti különbségébıl kiindulva megvizsgáltuk a csak
külön a lányokat tartalmazó vizsgálati csoport esetében a LEP genotípus gyakoriságát, de
ebben a szubpopulációban sem kaptunk szignifikáns eltérést. Az esélyhányados értéke
megnıtt (AA vs. GG: OR=2.02(CI:0.824-4.961)), ami azt jelzi, hogy a leptinnek a lányokban
nagyobb szerepe lehet az AIS kialakításában. A komplex betegségekben a gének egymással
bonyolult kölcsönhatásban alakítják ki a fenotípust, ezért a leptinnel funkcionális
kölcsönhatásban lévı gének vizsgálatával is értékes információhoz juthatunk az AIS
etiológiáját illetıen. Bioinformatikai eszközök használatával jelenítettük meg az
interakciókat. Ehhez a STRING 8.2 adatbázist használtuk, ami az ismert és prediktált, fizikai
és funkcionális fehérje kölcsönhatásokat több forrásból összegyőjtve tartalmazza (Jensen et
al., 2009). Ez az adatbázis jelenleg 630 élılény 2,590,259 fehérjéjét tartalmazza. A 13. ábrán
látható a leptinre jellemzı hálózat ábrázolása. A leptin és a leptin receptora körül két csoport
rajzolódik ki, melyek több különbözı szempont alapján összegyőjtött kölcsönható fehérjéket
tartalmaznak. A 13. ábrán látható táblázatból további elemzésre az IL6-ot választottuk ki,
mert egy nemrég megjelent közleményben már vizsgálták az AIS-sel összefüggésben (Aulisa
et al., 2007).
13. ábra. A STRING
programcsomag által
készített hálózat, amely a leptin interakciós partnereit
jeleníti meg.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
65
Az IL6 G-174C SNP GG genotípusát szignifikánsan magasabb elıfordulási gyakorisággal
találták meg egy olasz eset-kontroll tanulmányban, ahol Aulisa és mtsai 53 AIS-es beteg és
206 egészséges kontroll egyed genotípusát határozták meg. A GG genotípus kétszer olyan
gyakran fordult elı az AIS-esek között, mint a kontroll populációban (0.52 vs. 0.26) és az
esélyhányados OR= 10.54-nak adódott, ami szokatlanul magas egy multifaktoriális betegség
esetén (Aulisa et al., 2007). Elvégezve a genotipizálást nem tudtunk kimutatni semmilyen
eltérést a genotípus eloszlásban, az OR értékének egytıl való legnagyobb eltérése a két
homozigóta pár összehasonlításánál adódott GG vs. CC: OR=0.765 (CI: 0.396-1.478), ami a
G allél protektív tulajdonságát jelzi, és ellentmond az olasz kutatócsoport által tett
megfigyelésnek. A komplex betegségek asszociációs vizsgálatánál a reprodukálhatóság kicsi,
aminek okát több összefoglaló cikk is taglalja (Hirschhorn et al., 2002; Newton-Cheh and
Hirschhorn, 2005). A megismételhetıség hiányának fıbb okait a nem megfelelı kísérleti
elrendezésben, a kicsi statisztikai erıben vagy a populáció rétegzıdésében jelölik meg.
Kiemelik, hogy a multifaktoriális betegségek esetén a gének és a környezeti tényezık közötti
komplex kölcsönhatás figyelembe vétele is fontos lehet. Még több, megfelelı statisztikai
erıvel rendelkezı, független populáción elvégzett eset-kontroll tanulmány elvégzése
szükséges ahhoz, hogy eldönthetı legyen az IL6 promóter polimorfizmusának asszociációja a
gerincferdüléshez.
A LEP és az IL6 gének közötti episztázis analízist két független statisztikai módszerrel is
elvégeztük, mindkettı szerint létezik az alléljeik között kölcsönhatás és a LEP AA-IL6 CC
genotípus kombináció hordozása szignifikánsan magasabb kockázatot jelent az AIS
kialakulásában (OR=4.667 (CI:1.237-17.6). Mivel a Cobb˚ és a genotípusok között nem
találtunk összefüggést úgy gondoljuk, hogy a LEP AA-IL6 CC genotípus kombináció az AIS
kialakulására hajlamosít, nem pedig a progressziójában játszhat szerepet.
Az AIS etiológiájában fontos szerepe van az örökletes tényezıknek, de a kiterjedt kutatások
ellenére a mai napig nem sikerült olyan gént/géneket találni, melyek hibái az AIS
kialakulásáért lennének felelısek. A sikertelenségnek több oka is lehet, és ezek a komplex
betegségekre általánosan jellemzı tulajdonságokból erednek. Egyrészrıl a komplex
betegségek multigénes öröklıdésőek, ahol egy-egy génhiba hozzájárulása a fenotípus
kialakulásához nem minden esetben egyforma, sıt még a környezeti tényezık is
befolyásolhatják azt. Másrészrıl egy adott betegség megjelenési formája mögött teljesen más
pathomechanizmus is állhat, mivel a komplex betegségek gyakran genetikailag heterogének.
Az AIS genetikai modellje szerint a betegség kialakulása két szakaszra osztható, egy
iniciációs és egy progressziós fázisra (14. ábra). Feltételezések szerint más-más gének hibái

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
66
felelısek a hajlamosításra és a súlyosbodásra. Ezért nem csak a prediszponáló, hanem a
progressziós gének azonosítása is fontos. Amennyiben bizonyos génhibák, vagy
polimorfizmusok ismeretében megjósolható a súlyosbodás mértéke, akkor a megfelelı
terápiás kezelések kerülhetnek alkalmazásra. A legújabb kutatási eredmények szerint
kifejlesztettek olyan genetikai tesztet, amely alkalmas annak az eldöntésére, hogy mennyi a
kockázata a görbület további súlyosbodásának (Ogilvie, 2009).
14.ábra A jelenleg elfogadott genetikai modell az AIS kialakulására
Környezeti faktorok
Progressziós gének
Normál pubertás
Az AIS kezdete
Iniciációs gének
A görbület progressziója a
csontosodás befejezıdéséig

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
67
IRODALOMJEGYZÉK
Acaroglu,E., I.Akel, A.Alanay, M.Yazici and R.Marcucio: Comparison of the melatonin and calmodulin in paravertebral muscle and platelets of patients with or without adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 34, E659-E663(2009).
Aksenovich,T.I., I.R.Semenov, E.K.Ginzburg and A.M.Zaidman: [Preliminary analysis of inheritance of scoliosis]. Genetika Dev. 24, 2056-2063 (1988).
Alden,K.J., B.Marosy, N.Nzegwu, C.M.Justice, A.F.Wilson and N.H.Miller: Idiopathic scoliosis: identification of candidate regions on chromosome 19p13. Spine (Phila Pa 1976.) Dev. 31, 1815-1819 (2006).
Ambrosini,G., A.K.Nath, M.R.Sierra-Honigmann and J.Flores-Riveros: Transcriptional activation of the human leptin gene in response to hypoxia. Involvement of hypoxia-inducible factor 1. J.Biol.Chem. Dev. 277, 34601-34609 (2002).
Aulisa,L., P.Papaleo, E.Pola, F.Angelini, A.G.Aulisa, F.C.Tamburrelli, P.Pola and C.A.Logroscino: Association between IL-6 and MMP-3 gene polymorphisms and adolescent idiopathic scoliosis: a case-control study. Spine (Phila Pa 1976.) Dev. 32, 2700-2702 (2007).
Bagnall,K.M., V.J.Raso, D.L.Hill, M.Moreau, J.K.Mahood, H.Jiang, G.Russell, M.Bering and G.R.Buzzell: Melatonin levels in idiopathic scoliosis. Diurnal and nocturnal serum melatonin levels in girls with adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 21, 1974-1978 (1996).
Baron,M.: The search for complex disease genes: fault by linkage or fault by association? Mol.Psychiatry Dev. 6, 143-149 (2001).
Bashiardes,S., R.Veile, M.Allen, C.A.Wise, M.Dobbs, J.A.Morcuende, L.Szappanos, J.A.Herring, A.M.Bowcock and M.Lovett: SNTG1, the gene encoding gamma1-syntrophin: a candidate gene for idiopathic scoliosis. Hum.Genet. Dev. 115, 81-89 (2004).
Bellyei,A., A.Czeizel, O.Barta, T.Magda and L.Molnar: Prevalence of adolescent idiopathic scoliosis in Hungary. Acta Orthop.Scand. Dev. 48, 177-180 (1977).
Bienertova-Vasku,J., P.Bienert, J.Tomandl, M.Forejt, M.Vavrina, J.Kudelkova and A.Vasku: No association of defined variability in leptin, leptin receptor, adiponectin, proopiomelanocortin and ghrelin gene with food preferences in the Czech population. Nutr.Neurosci. Dev. 11, 2-8 (2008).
Blanchard,F., L.Duplomb, M.Baud'huin and B.Brounais: The dual role of IL-6-type cytokines on bone remodeling and bone tumors. Cytokine Growth Factor Rev. Dev. 20, 19-28 (2009).
Blum,W.F., P.Englaro, S.Hanitsch, A.Juul, N.T.Hertel, J.Muller, N.E.Skakkebaek, M.L.Heiman, M.Birkett, A.M.Attanasio, W.Kiess and W.Rascher: Plasma leptin levels in healthy children and adolescents: dependence on body mass index, body fat mass, gender, pubertal stage, and testosterone. J.Clin.Endocrinol.Metab Dev. 82, 2904-2910 (1997).
Boiardi,L., B.Casali, E.Farnetti, N.Pipitone, D.Nicoli, F.Cantini, P.Macchioni, G.Bajocchi, M.G.Catanoso, L.Pulsatelli, D.Consonni and C.Salvarani: Relationship between interleukin 6

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
68
promoter polymorphism at position -174, IL-6 serum levels, and the risk of relapse/recurrence in polymyalgia rheumatica. J.Rheumatol. Dev. 33, 703-708 (2006).
Burke,J.G., G.W.RW, D.Conhyea, D.McCormack, F.E.Dowling, M.G.Walsh and J.M.Fitzpatrick: Human nucleus pulposis can respond to a pro-inflammatory stimulus. Spine (Phila Pa 1976.) Dev. 28, 2685-2693 (2003).
Burner,W.L., III, V.M.Badger and F.C.Sherman: Osteoporosis and acquired back deformities. J.Pediatr.Orthop. Dev. 2, 383-385 (1982).
Burwell,R.G., R.K.Aujla, M.P.Grevitt, P.H.Dangerfield, A.Moulton, T.L.Randell and S.I.Anderson: Pathogenesis of adolescent idiopathic scoliosis in girls - a double neuro-osseous theory involving disharmony between two nervous systems, somatic and autonomic expressed in the spine and trunk: possible dependency on sympathetic nervous system and hormones with implications for medical therapy. Scoliosis. Dev. 4, 24(2009).
Burwell,R.G., P.H.Dangerfield and B.J.Freeman: Concepts on the pathogenesis of adolescent idiopathic scoliosis. Bone growth and mass, vertebral column, spinal cord, brain, skull, extra-spinal left-right skeletal length asymmetries, disproportions and molecular pathogenesis. Stud.Health Technol.Inform. Dev. 135, 3-52 (2008a).
Burwell,R.G., P.H.Dangerfield and B.J.Freeman: Etiologic theories of idiopathic scoliosis. Somatic nervous system and the NOTOM escalator concept as one component in the pathogenesis of adolescent idiopathic scoliosis. Stud.Health Technol.Inform. Dev. 140, 208-217 (2008b).
Carlsson,B., K.Lindell, B.Gabrielsson, C.Karlsson, R.Bjarnason, O.Westphal, U.Karlsson, L.Sjostrom and L.M.Carlsson: Obese (ob) gene defects are rare in human obesity. Obes.Res. Dev. 5, 30-35 (1997).
Carr,A.J., D.J.Ogilvie, B.P.Wordsworth, L.M.Priestly, R.Smith and B.Sykes: Segregation of structural collagen genes in adolescent idiopathic scoliosis. Clin.Orthop.Relat Res. Dev. 305-310 (1992).
Chan,V., G.C.Fong, K.D.Luk, B.Yip, M.K.Lee, M.S.Wong, D.D.Lu and T.K.Chan: A genetic locus for adolescent idiopathic scoliosis linked to chromosome 19p13.3. Am.J.Hum.Genet. Dev. 71, 401-406 (2002).
Cheng,J.C., X.Guo and A.H.Sher: Persistent osteopenia in adolescent idiopathic scoliosis. A longitudinal follow up study. Spine Dev. 24, 1218-1222 (1999).
Cheng,J.C., L.Qin, C.S.Cheung, A.H.Sher, K.M.Lee, S.W.Ng and X.Guo: Generalized low areal and volumetric bone mineral density in adolescent idiopathic scoliosis. J.Bone Miner.Res. Dev. 15, 1587-1595 (2000).
Cheng,J.C., S.P.Tang, X.Guo, C.W.Chan and L.Qin: Osteopenia in adolescent idiopathic scoliosis: a histomorphometric study. Spine (Phila Pa 1976.) Dev. 26, E19-E23(2001).
Cheung,J., J.P.Halbertsma, A.G.Veldhuizen, W.J.Sluiter, N.M.Maurits, J.C.Cool and J.R.van Horn: A preliminary study on electromyographic analysis of the paraspinal musculature in idiopathic scoliosis. Eur.Spine J. Dev. 14, 130-137 (2005a).
Cheung,K.M., T.Wang, A.M.Poon, A.Carl, B.Tranmer, Y.Hu, K.D.Luk and J.C.Leong: The effect of pinealectomy on scoliosis development in young nonhuman primates. Spine (Phila Pa 1976.) Dev. 30, 2009-2013 (2005b).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
69
Cheung,K.M., T.Wang, G.X.Qiu and K.D.Luk: Recent advances in the aetiology of adolescent idiopathic scoliosis. Int.Orthop. Dev. 32, 729-734 (2008).
Cowell,H.R., J.N.Hall and G.D.MacEwen: Genetic aspects of idiopathic scoliosis. A Nicholas Andry Award essay, 1970. Clin.Orthop.Relat Res. Dev. 86, 121-131 (1972).
Czeizel,A., A.Bellyei, O.Barta, T.Magda and L.Molnar: Genetics of adolescent idiopathic scoliosis. J.Med.Genet. Dev. 15, 424-427 (1978).
Dalpe,G., M.Mathieu, A.Comtois, E.Zhu, S.Wasiak, Y.De Repentigny, N.Leclerc and R.Kothary: Dystonin-deficient mice exhibit an intrinsic muscle weakness and an instability of skeletal muscle cytoarchitecture. Dev.Biol. Dev. 210, 367-380 (1999a).
Dalpe,G., M.Mathieu, A.Comtois, E.Zhu, S.Wasiak, Y.De Repentigny, N.Leclerc and R.Kothary: Dystonin-deficient mice exhibit an intrinsic muscle weakness and an instability of skeletal muscle cytoarchitecture. Dev.Biol. Dev. 210, 367-380 (1999b).
De Laet,C., J.A.Kanis, A.Oden, H.Johanson, O.Johnell, P.Delmas, J.A.Eisman, H.Kroger, S.Fujiwara, P.Garnero, E.V.McCloskey, D.Mellstrom, L.J.Melton, III, P.J.Meunier, H.A.Pols, J.Reeve, A.Silman and A.Tenenhouse: Body mass index as a predictor of fracture risk: a meta-analysis. Osteoporos.Int. Dev. 16, 1330-1338 (2005).
DeRisi,J., L.Penland, P.O.Brown, M.L.Bittner, P.S.Meltzer, M.Ray, Y.Chen, Y.A.Su and J.M.Trent: Use of a cDNA microarray to analyse gene expression patterns in human cancer. Nat.Genet. Dev. 14, 457-460 (1996).
Dieudonne,M.N., F.Machinal-Quelin, V.Serazin-Leroy, M.C.Leneveu, R.Pecquery and Y.Giudicelli: Leptin mediates a proliferative response in human MCF7 breast cancer cells. Biochem.Biophys.Res.Commun. Dev. 293, 622-628 (2002).
Ducy,P., M.Amling, S.Takeda, M.Priemel, A.F.Schilling, F.T.Beil, J.Shen, C.Vinson, J.M.Rueger and G.Karsenty: Leptin inhibits bone formation through a hypothalamic relay: a central control of bone mass. Cell Dev. 100, 197-207 (2000).
Elefteriou,F., J.D.Ahn, S.Takeda, M.Starbuck, X.Yang, X.Liu, H.Kondo, W.G.Richards, T.W.Bannon, M.Noda, K.Clement, C.Vaisse and G.Karsenty: Leptin regulation of bone resorption by the sympathetic nervous system and CART. Nature Dev. 434, 514-520 (2005).
Fagan,A.B., D.J.Kennaway and A.D.Sutherland: Total 24-hour melatonin secretion in adolescent idiopathic scoliosis. A case-control study. Spine (Phila Pa 1976.) Dev. 23, 41-46 (1998).
Fishman,D., G.Faulds, R.Jeffery, V.Mohamed-Ali, J.S.Yudkin, S.Humphries and P.Woo: The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J.Clin.Invest Dev. 102, 1369-1376 (1998).
Gangwar,R., B.Mittal and R.D.Mittal: Association of interleukin-6 -174G>C promoter polymorphism with risk of cervical cancer. Int.J.Biol.Markers Dev. 24, 11-16 (2009).
Gao,X., D.Gordon, D.Zhang, R.Browne, C.Helms, J.Gillum, S.Weber, S.Devroy, S.Swaney, M.Dobbs, J.Morcuende, V.Sheffield, M.Lovett, A.Bowcock, J.Herring and C.Wise: CHD7 gene polymorphisms are associated with susceptibility to idiopathic scoliosis. Am.J.Hum.Genet. Dev. 80, 957-965 (2007).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
70
Gong,D.W., S.Bi, R.E.Pratley and B.D.Weintraub: Genomic structure and promoter analysis of the human obese gene. J.Biol.Chem. Dev. 271, 3971-3974 (1996).
Guo,X., W.W.Chau, Y.L.Chan and J.C.Cheng: Relative anterior spinal overgrowth in adolescent idiopathic scoliosis. Results of disproportionate endochondral-membranous bone growth. J.Bone Joint Surg.Br. Dev. 85, 1026-1031 (2003).
Hahn,L.W., M.D.Ritchie and J.H.Moore: Multifactor dimensionality reduction software for detecting gene-gene and gene-environment interactions. Bioinformatics. Dev. 19, 376-382 (2003b).
Hahn,L.W., M.D.Ritchie and J.H.Moore: Multifactor dimensionality reduction software for detecting gene-gene and gene-environment interactions. Bioinformatics. Dev. 19, 376-382 (2003a).
Higuchi,R., C.Fockler, G.Dollinger and R.Watson: Kinetic PCR analysis: real-time monitoring of DNA amplification reactions. Biotechnology (N.Y) Dev. 11, 1026-1030 (1993).
Hinuy,H.M., M.H.Hirata, N.Forti, J.Diament, M.F.Sampaio, D.Armaganijan, L.A.Salazar and R.D.Hirata: Leptin G-2548A promoter polymorphism is associated with increased plasma leptin and BMI in Brazilian women. Arq Bras.Endocrinol.Metabol. Dev. 52, 611-616 (2008).
Hirose,Y., K.Chiba, T.Karasugi, M.Nakajima, Y.Kawaguchi, Y.Mikami, T.Furuichi, F.Mio, A.Miyake, T.Miyamoto, K.Ozaki, A.Takahashi, H.Mizuta, T.Kubo, T.Kimura, T.Tanaka, Y.Toyama and S.Ikegawa: A functional polymorphism in THBS2 that affects alternative splicing and MMP binding is associated with lumbar-disc herniation. Am.J.Hum.Genet. Dev. 82, 1122-1129 (2008).
Hirschhorn,J.N., K.Lohmueller, E.Byrne and K.Hirschhorn: A comprehensive review of genetic association studies. Genet.Med. Dev. 4, 45-61 (2002).
Hoffstedt,J., P.Eriksson, S.Mottagui-Tabar and P.Arner: A polymorphism in the leptin promoter region (-2548 G/A) influences gene expression and adipose tissue secretion of leptin. Horm.Metab Res. Dev. 34, 355-359 (2002).
Holloway,W.R., F.M.Collier, C.J.Aitken, D.E.Myers, J.M.Hodge, M.Malakellis, T.J.Gough, G.R.Collier and G.C.Nicholson: Leptin inhibits osteoclast generation. J.Bone Miner.Res. Dev. 17, 200-209 (2002).
Holt,I., M.W.Davie and M.J.Marshall: Osteoclasts are not the major source of interleukin-6 in mouse parietal bones. Bone Dev. 18, 221-226 (1996).
Huang,d.W., B.T.Sherman and R.A.Lempicki: Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat.Protoc. Dev. 4, 44-57 (2009).
Ishii,A. and S.H.Lo: A role of tensin in skeletal-muscle regeneration. Biochem.J. Dev. 356, 737-745 (2001).
Jensen,L.J., M.Kuhn, M.Stark, S.Chaffron, C.Creevey, J.Muller, T.Doerks, P.Julien, A.Roth, M.Simonovic, P.Bork and C.von Mering: STRING 8--a global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. Dev. 37, D412-D416(2009).
Justice,C.M., N.H.Miller, B.Marosy, J.Zhang and A.F.Wilson: Familial idiopathic scoliosis: evidence of an X-linked susceptibility locus. Spine (Phila Pa 1976.) Dev. 28, 589-594 (2003).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
71
Kanehisa,M., S.Goto, M.Furumichi, M.Tanabe and M.Hirakawa: KEGG for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res. Dev. 38, D355-D360(2010).
Kesling,K.L. and K.A.Reinker: Scoliosis in twins. A meta-analysis of the literature and report of six cases. Spine (Phila Pa 1976.) Dev. 22, 2009-2014 (1997).
Kindsfater,K., T.Lowe, D.Lawellin, D.Weinstein and J.Akmakjian: Levels of platelet calmodulin for the prediction of progression and severity of adolescent idiopathic scoliosis. J.Bone Joint Surg.Am. Dev. 76, 1186-1192 (1994).
Kouwenhoven,J.W. and R.M.Castelein: The pathogenesis of adolescent idiopathic scoliosis: review of the literature. Spine (Phila Pa 1976.) Dev. 33, 2898-2908 (2008).
Kwan,T.S., M.Padrines, S.Theoleyre, D.Heymann and Y.Fortun: IL-6, RANKL, TNF-alpha/IL-1: interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. Dev. 15, 49-60 (2004).
Lao,M.L., D.H.Chow, X.Guo, J.C.Cheng and A.D.Holmes: Impaired dynamic balance control in adolescents with idiopathic scoliosis and abnormal somatosensory evoked potentials. J.Pediatr.Orthop. Dev. 28, 846-849 (2008).
Le Stunff,C., C.Le Bihan, N.J.Schork and P.Bougneres: A common promoter variant of the leptin gene is associated with changes in the relationship between serum leptin and fat mass in obese girls. Diabetes Dev. 49, 2196-2200 (2000).
Leon,L.R., A.A.White and M.J.Kluger: Role of IL-6 and TNF in thermoregulation and survival during sepsis in mice. Am.J.Physiol Dev. 275, R269-R277(1998).
Li,X.F., H.Li, Z.D.Liu and L.Y.Dai: Low bone mineral status in adolescent idiopathic scoliosis. Eur.Spine J. Dev. 17, 1431-1440 (2008).
Liu,T., W.C.Chu, G.Young, K.Li, B.H.Yeung, L.Guo, G.C.Man, W.W.Lam, S.T.Wong and J.C.Cheng: MR analysis of regional brain volume in adolescent idiopathic scoliosis: neurological manifestation of a systemic disease. J.Magn Reson.Imaging Dev. 27, 732-736 (2008).
Lo,H.S., Z.Wang, Y.Hu, H.H.Yang, S.Gere, K.H.Buetow and M.P.Lee: Allelic variation in gene expression is common in the human genome. Genome Res. Dev. 13, 1855-1862 (2003).
Lowe,T.G., M.Edgar, J.Y.Margulies, N.H.Miller, V.J.Raso, K.A.Reinker and C.H.Rivard: Etiology of idiopathic scoliosis: current trends in research. J.Bone Joint Surg.Am. Dev. 82-A, 1157-1168 (2000).
Machida,M., J.Dubousset, Y.Imamura, T.Iwaya, T.Yamada and J.Kimura: An experimental study in chickens for the pathogenesis of idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 18, 1609-1615 (1993).
Machida,M., J.Dubousset, Y.Imamura, Y.Miyashita, T.Yamada and J.Kimura: Melatonin. A possible role in pathogenesis of adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 21, 1147-1152 (1996).
Machida,M., I.Murai, Y.Miyashita, J.Dubousset, T.Yamada and J.Kimura: Pathogenesis of idiopathic scoliosis. Experimental study in rats. Spine (Phila Pa 1976.) Dev. 24, 1985-1989 (1999).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
72
Mammes,O., D.Betoulle, R.Aubert, B.Herbeth, G.Siest and F.Fumeron: Association of the G-2548A polymorphism in the 5' region of the LEP gene with overweight. Ann.Hum.Genet. Dev. 64, 391-394 (2000).
Mantzoros,C.S. and S.J.Moschos: Leptin: in search of role(s) in human physiology and pathophysiology. Clin.Endocrinol.(Oxf) Dev. 49, 551-567 (1998).
Marosy,B., C.M.Justice, N.Nzegwu, G.Kumar, A.F.Wilson and N.H.Miller: Lack of association between the aggrecan gene and familial idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 31, 1420-1425 (2006).
Martin,A., V.David, L.Malaval, M.H.Lafage-Proust, L.Vico and T.Thomas: Opposite effects of leptin on bone metabolism: a dose-dependent balance related to energy intake and insulin-like growth factor-I pathway. Endocrinology Dev. 148, 3419-3425 (2007).
Mason,M.M., Y.F.He, H.Chen, M.J.Quon and M.Reitman: Regulation of leptin promoter function by Sp1, C/EBP, and a novel factor. Endocrinology Dev. 139, 1013-1022 (1998).
Miller,N.H., C.M.Justice, B.Marosy, K.F.Doheny, E.Pugh, J.Zhang, H.C.Dietz, III and A.F.Wilson: Identification of candidate regions for familial idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 30, 1181-1187 (2005).
Miller,N.H., B.Marosy, C.M.Justice, S.M.Novak, E.Y.Tang, P.Boyce, J.Pettengil, K.F.Doheny, E.W.Pugh and A.F.Wilson: Linkage analysis of genetic loci for kyphoscoliosis on chromosomes 5p13, 13q13.3, and 13q32. Am.J.Med.Genet.A Dev. 140, 1059-1068 (2006).
Miller,N.H., B.Mims, A.Child, D.M.Milewicz, P.Sponseller and S.H.Blanton: Genetic analysis of structural elastic fiber and collagen genes in familial adolescent idiopathic scoliosis. J.Orthop.Res. Dev. 14, 994-999 (1996).
Misol,S., I.V.Ponseti, N.Samaan and J.T.Bradbury: Growth hormone blood levels in patients with idiopathic scoliosis. Clin.Orthop.Relat Res. Dev. 81, 122-125 (1971).
Montanaro,L., P.Parisini, T.Greggi, M.Di Silvestre, D.Campoccia, S.Rizzi and C.R.Arciola: Evidence of a linkage between matrilin-1 gene (MATN1) and idiopathic scoliosis. Scoliosis. Dev. 1, 21(2006).
Moreau,A., D.S.Wang, S.Forget, B.Azeddine, D.Angeloni, F.Fraschini, H.Labelle, B.Poitras, C.H.Rivard and G.Grimard: Melatonin signaling dysfunction in adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 29, 1772-1781 (2004).
Newton-Cheh,C. and J.N.Hirschhorn: Genetic association studies of complex traits: design and analysis issues. Mutat.Res. Dev. 573, 54-69 (2005).
Ocaka,L., C.Zhao, J.A.Reed, N.D.Ebenezer, G.Brice, T.Morley, M.Mehta, J.O'Dowd, J.L.Weber, A.J.Hardcastle and A.H.Child: Assignment of two loci for autosomal dominant adolescent idiopathic scoliosis to chromosomes 9q31.2-q34.2 and 17q25.3-qtel. J.Med.Genet. Dev. 45, 87-92 (2008).
Oegema,T.R., Jr., D.S.Bradford, K.M.Cooper and R.E.Hunter: Comparison of the biochemistry of proteoglycans isolated from normal, idiopathic scoliotic and cerebral palsy spines. Spine (Phila Pa 1976.) Dev. 8, 378-384 (1983).
Ogilvie,J.: Adolescent idiopathic scoliosis and genetic testing. Curr.Opin.Pediatr. Dev. (2009).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
73
Ostlund,R.E., Jr., J.W.Yang, S.Klein and R.Gingerich: Relation between plasma leptin concentration and body fat, gender, diet, age, and metabolic covariates. J.Clin.Endocrinol.Metab Dev. 81, 3909-3913 (1996).
Ostrowski,K., T.Rohde, S.Asp, P.Schjerling and B.K.Pedersen: Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. J.Physiol Dev. 515 ( Pt 1), 287-291 (1999).
Pedrini,V.A., I.V.Ponseti and S.C.Dohrman: Glycosaminoglycans of intervertebral disc in idiopathic scoliosis. J.Lab Clin.Med. Dev. 82, 938-950 (1973).
Peltonen,L. and V.A.McKusick: Genomics and medicine. Dissecting human disease in the postgenomic era. Science Dev. 291, 1224-1229 (2001).
Porter,R.W.: The pathogenesis of idiopathic scoliosis: uncoupled neuro-osseous growth? Eur.Spine J. Dev. 10, 473-481 (2001b).
Porter,R.W.: The pathogenesis of idiopathic scoliosis: uncoupled neuro-osseous growth? Eur.Spine J. Dev. 10, 473-481 (2001a).
Qi,L., R.M.van Dam, J.B.Meigs, J.E.Manson, D.Hunter and F.B.Hu: Genetic variation in IL6 gene and type 2 diabetes: tagging-SNP haplotype analysis in large-scale case-control study and meta-analysis. Hum.Mol.Genet. Dev. 15, 1914-1920 (2006).
Qi,L., C.Zhang, R.M.van Dam and F.B.Hu: Interleukin-6 genetic variability and adiposity: associations in two prospective cohorts and systematic review in 26,944 individuals. J.Clin.Endocrinol.Metab Dev. 92, 3618-3625 (2007).
Qiu,X.S., N.L.Tang, H.Y.Yeung, J.C.Cheng and Y.Qiu: Lack of association between the promoter polymorphism of the MTNR1A gene and adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 33, 2204-2207 (2008).
Qiu,X.S., N.L.Tang, H.Y.Yeung, K.M.Lee, V.W.Hung, B.K.Ng, S.L.Ma, R.H.Kwok, L.Qin, Y.Qiu and J.C.Cheng: Melatonin receptor 1B (MTNR1B) gene polymorphism is associated with the occurrence of adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 32, 1748-1753 (2007a).
Qiu,X.S., N.L.Tang, H.Y.Yeung, Y.Qiu and J.C.Cheng: Genetic association study of growth hormone receptor and idiopathic scoliosis. Clin.Orthop.Relat Res. Dev. 462, 53-58 (2007b).
Qiu,Y., X.Sun, X.S.Qiu, W.G.Li, Z.Z.Zhu, F.Zhu, B.Wang, Y.Yu and B.P.Qian: Decreased circulating leptin level and its association with body and bone mass in girls with adolescent idiopathic scoliosis. Spine Dev. 32, 2703-2710 (2007c).
Raggio,C.L., P.F.Giampietro, S.Dobrin, C.Zhao, D.Dorshorst, N.Ghebranious, J.L.Weber and R.D.Blank: A novel locus for adolescent idiopathic scoliosis on chromosome 12p. J.Orthop.Res. Dev. 27, 1366-1372 (2009).
Rasko,I.,Downes C.S.: Genes in Medicine Molecular biology and human genetic disorders (Chapman&Hall) (1994).
Ravaglia,G., P.Forti, F.Maioli, M.Chiappelli, P.Dolzani, M.Martelli, M.Bianchin, E.Mariani, L.Bolondi and F.Licastro: Associations of the -174 G/C interleukin-6 gene promoter polymorphism with serum interleukin 6 and mortality in the elderly. Biogerontology. Dev. 6, 415-423 (2005).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
74
Ribeiro,R., A.P.Araujo, A.Coelho, R.Catarino, D.Pinto, A.Araujo, C.Calcada, C.Lopes and R.Medeiros: A functional polymorphism in the promoter region of leptin gene increases susceptibility for non-small cell lung cancer. Eur.J.Cancer Dev. 42, 1188-1193 (2006).
Ribeiro,R., A.Vasconcelos, S.Costa, D.Pinto, A.Morais, J.Oliveira, F.Lobo, C.Lopes and R.Medeiros: Overexpressing leptin genetic polymorphism (-2548 G/A) is associated with susceptibility to prostate cancer and risk of advanced disease. Prostate Dev. 59, 268-274 (2004).
Riseborough,E.J. and R.Wynne-Davies: A genetic survey of idiopathic scoliosis in Boston, Massachusetts. J.Bone Joint Surg.Am. Dev. 55, 974-982 (1973).
Roach,J.W.: Adolescent idiopathic scoliosis. Orthop.Clin.North Am. Dev. 30, 353-viii(1999).
Roberts,S., J.Menage and S.M.Eisenstein: The cartilage end-plate and intervertebral disc in scoliosis: calcification and other sequelae. J.Orthop.Res. Dev. 11, 747-757 (1993).
Rodriguez,S., T.R.Gaunt and I.N.Day: Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am.J.Epidemiol. Dev. 169, 505-514 (2009).
Roth,J.A., B.G.Kim, W.L.Lin and M.I.Cho: Melatonin promotes osteoblast differentiation and bone formation. J.Biol.Chem. Dev. 274, 22041-22047 (1999).
Roth,M.: Idiopathic scoliosis caused by a short spinal cord. Acta Radiol.Diagn.(Stockh) Dev. 7, 257-271 (1968).
Sadat-Ali,M., A.Al Othman, D.Bubshait and D.Al Dakheel: Does scoliosis causes low bone mass? A comparative study between siblings. Eur.Spine J. Dev. 17, 944-947 (2008).
Saladin,R., P.De Vos, M.Guerre-Millo, A.Leturque, J.Girard, B.Staels and J.Auwerx: Transient increase in obese gene expression after food intake or insulin administration. Nature Dev. 377, 527-529 (1995).
Salehi,L.B., M.Mangino, S.De Serio, D.De Cicco, F.Capon, S.Semprini, A.Pizzuti, G.Novelli and B.Dallapiccola: Assignment of a locus for autosomal dominant idiopathic scoliosis (IS) to human chromosome 17p11. Hum.Genet. Dev. 111, 401-404 (2002).
Satomura,K., S.Tobiume, R.Tokuyama, Y.Yamasaki, K.Kudoh, E.Maeda and M.Nagayama: Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J.Pineal Res. Dev. 42, 231-239 (2007).
Sevastik,J., R.G.Burwell and P.H.Dangerfield: A new concept for the etiopathogenesis of the thoracospinal deformity of idiopathic scoliosis: summary of an electronic focus group debate of the IBSE. Eur.Spine J. Dev. 12, 440-450 (2003).
Shohat,M., T.Shohat, M.Nitzan, M.Mimouni, R.Kedem and Y.L.Danon: Growth and ethnicity in scoliosis. Acta Orthop.Scand. Dev. 59, 310-313 (1988).
Skibola,C.F., E.A.Holly, M.S.Forrest, A.Hubbard, P.M.Bracci, D.R.Skibola, C.Hegedus and M.T.Smith: Body mass index, leptin and leptin receptor polymorphisms, and non-hodgkin lymphoma. Cancer Epidemiol.Biomarkers Prev. Dev. 13, 779-786 (2004).
Smith,A.J., F.D'Aiuto, J.Palmen, J.A.Cooper, J.Samuel, S.Thompson, J.Sanders, N.Donos, L.Nibali, D.Brull, P.Woo and S.E.Humphries: Association of serum interleukin-6 concentration with a functional IL6 -6331T>C polymorphism. Clin.Chem. Dev. 54, 841-850 (2008).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
75
Snoussi,K., A.D.Strosberg, N.Bouaouina, S.Ben Ahmed, A.N.Helal and L.Chouchane: Leptin and leptin receptor polymorphisms are associated with increased risk and poor prognosis of breast carcinoma. BMC Cancer Dev. 6, 38(2006).
Soyka,L.A., S.Grinspoon, L.L.Levitsky, D.B.Herzog and A.Klibanski: The effects of anorexia nervosa on bone metabolism in female adolescents. J.Clin.Endocrinol.Metab Dev. 84, 4489-4496 (1999).
Spencer,G.S. and M.J.Eccles: Spinal muscle in scoliosis. Part 2. The proportion and size of type 1 and type 2 skeletal muscle fibres measured using a computer-controlled microscope. J.Neurol.Sci. Dev. 30, 143-154 (1976).
Stokes,I.A. and D.D.Aronsson: Disc and vertebral wedging in patients with progressive scoliosis. J.Spinal Disord. Dev. 14, 317-322 (2001).
Stokes,I.A. and J.P.Laible: Three-dimensional osseo-ligamentous model of the thorax representing initiation of scoliosis by asymmetric growth. J.Biomech. Dev. 23, 589-595 (1990).
Strobel,A., T.Issad, L.Camoin, M.Ozata and A.D.Strosberg: A leptin missense mutation associated with hypogonadism and morbid obesity. Nat.Genet. Dev. 18, 213-215 (1998).
Taguchi,Y., M.Yamamoto, T.Yamate, S.C.Lin, H.Mocharla, P.DeTogni, N.Nakayama, B.F.Boyce, E.Abe and S.C.Manolagas: Interleukin-6-type cytokines stimulate mesenchymal progenitor differentiation toward the osteoblastic lineage. Proc.Assoc.Am.Physicians Dev. 110, 559-574 (1998).
Taylor,T.K., P.Ghosh and G.R.Bushell: The contribution of the intervertebral disk to the scoliotic deformity. Clin.Orthop.Relat Res. Dev. 79-90 (1981).
Thalmann,S. and C.A.Meier: Local adipose tissue depots as cardiovascular risk factors. Cardiovasc.Res. Dev. 75, 690-701 (2007).
THILLARD,M.J.: [Vertebral column deformities following epiphysectomy in the chick.]. C R.Hebd.Seances Acad.Sci. Dev. 248, 1238-1240 (1959).
Thomas,K.A., S.D.Cook, T.C.Skalley, S.V.Renshaw, R.S.Makuch, M.Gross, T.S.Whitecloud, III and J.T.Bennett: Lumbar spine and femoral neck bone mineral density in idiopathic scoliosis: a follow-up study. J.Pediatr.Orthop. Dev. 12, 235-240 (1992).
Thomas,T., F.Gori, S.Khosla, M.D.Jensen, B.Burguera and B.L.Riggs: Leptin acts on human marrow stromal cells to enhance differentiation to osteoblasts and to inhibit differentiation to adipocytes. Endocrinology Dev. 140, 1630-1638 (1999).
Trayhurn,P. and I.S.Wood: Adipokines: inflammation and the pleiotropic role of white adipose tissue. Br.J.Nutr. Dev. 92, 347-355 (2004).
Upadhyay,R., M.Jain, S.Kumar, U.C.Ghoshal and B.Mittal: Association of interleukin-6 (-174G>C) promoter polymorphism with risk of squamous cell esophageal cancer and tumor location: an exploratory study. Clin.Immunol. Dev. 128, 199-204 (2008).
Veldhuizen,A.G., D.J.Wever and P.J.Webb: The aetiology of idiopathic scoliosis: biomechanical and neuromuscular factors. Eur.Spine J. Dev. 9, 178-184 (2000).
Walsh,P.S., D.A.Metzger and R.Higuchi: Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques Dev. 10, 506-513 (1991).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
76
Wang,J.L., R.Liu, M.Hawkins, N.Barzilai and L.Rossetti: A nutrient-sensing pathway regulates leptin gene expression in muscle and fat. Nature Dev. 393, 684-688 (1998).
Wang,T.N., M.C.Huang, W.T.Chang, A.M.Ko, E.M.Tsai, C.S.Liu, C.H.Lee and Y.C.Ko: G-2548A polymorphism of the leptin gene is correlated with extreme obesity in Taiwanese aborigines. Obesity (Silver.Spring) Dev. 14, 183-187 (2006).
Weimann,E., W.F.Blum, C.Witzel, S.Schwidergall and H.J.Bohles: Hypoleptinemia in female and male elite gymnasts. Eur.J.Clin.Invest Dev. 29, 853-860 (1999).
Wise,C.A., R.Barnes, J.Gillum, J.A.Herring, A.M.Bowcock and M.Lovett: Localization of susceptibility to familial idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 25, 2372-2380 (2000).
Wynne-Davies,R.: Familial (idiopathic) scoliosis. A family survey. J.Bone Joint Surg.Br. Dev. 50, 24-30 (1968).
Xiao,P., P.Y.Liu, Y.Lu, Y.F.Guo, D.H.Xiong, L.H.Li, R.R.Recker and H.W.Deng: Association tests of interleukin-6 (IL-6) and type II tumor necrosis factor receptor (TNFR2) genes with bone mineral density in Caucasians using a re-sampling approach. Hum.Genet. Dev. 117, 340-348 (2005).
Yamada,R., T.Tanaka, M.Unoki, T.Nagai, T.Sawada, Y.Ohnishi, T.Tsunoda, M.Yukioka, A.Maeda, K.Suzuki, H.Tateishi, T.Ochi, Y.Nakamura and K.Yamamoto: Association between a single-nucleotide polymorphism in the promoter of the human interleukin-3 gene and rheumatoid arthritis in Japanese patients, and maximum-likelihood estimation of combinatorial effect that two genetic loci have on susceptibility to the disease. Am.J.Hum.Genet. Dev. 68, 674-685 (2001).
Yapijakis,C., M.Kechagiadakis, E.Nkenke, Z.Serefoglou, D.Avgoustidis, A.Vylliotis, D.Perrea, F.W.Neukam, E.Patsouris and E.Vairaktaris: Association of leptin -2548G/A and leptin receptor Q223R polymorphisms with increased risk for oral cancer. J.Cancer Res.Clin.Oncol. Dev. 135, 603-612 (2009).
Yeung,H.Y., N.L.Tang, K.M.Lee, B.K.Ng, V.W.Hung, R.Kwok, X.Guo, L.Qin and J.C.Cheng: Genetic association study of insulin-like growth factor-I (IGF-I) gene with curve severity and osteopenia in adolescent idiopathic scoliosis. Stud.Health Technol.Inform. Dev. 123, 18-24 (2006).
Yiannakouris,N., L.Melistas, M.Yannakoulia, K.Mungal and C.S.Mantzoros: The-2548G/A polymorphism in the human leptin gene promoter region is associated with plasma free leptin levels; interaction with adiposity and gender in healthy subjects. Hormones.(Athens.) Dev. 2, 229-236 (2003).
Zhang,F., M.B.Basinski, J.M.Beals, S.L.Briggs, L.M.Churgay, D.K.Clawson, R.D.DiMarchi, T.C.Furman, J.E.Hale, H.M.Hsiung, B.E.Schoner, D.P.Smith, X.Y.Zhang, J.P.Wery and R.W.Schevitz: Crystal structure of the obese protein leptin-E100. Nature Dev. 387, 206-209 (1997).
Zhang,H.Q., S.J.Lu, M.X.Tang, L.Q.Chen, S.H.Liu, C.F.Guo, X.Y.Wang, J.Chen and L.Xie: Association of estrogen receptor beta gene polymorphisms with susceptibility to adolescent idiopathic scoliosis. Spine (Phila Pa 1976.) Dev. 34, 760-764 (2009).
Zhang,Y., R.Proenca, M.Maffei, M.Barone, L.Leopold and J.M.Friedman: Positional cloning of the mouse obese gene and its human homologue. Nature Dev. 372, 425-432 (1994).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
77
Zhao,L.J., H.Jiang, C.J.Papasian, D.Maulik, B.Drees, J.Hamilton and H.W.Deng: Correlation of obesity and osteoporosis: effect of fat mass on the determination of osteoporosis. J.Bone Miner.Res. Dev. 23, 17-29 (2008).
Zhao,Y. and G.X.Qiu: [Expression of calmodulin and nNOS in the paraspinal muscles in idiopathic scoliosis]. Zhonghua Yi.Xue.Za Zhi. Dev. 84, 1358-1361 (2004).

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
78
KÖSZÖNETNYILVÁNÍTÁS
Ezúton szeretnék köszönetet mondani mindazoknak, akik a munkámban és a dolgozatom
elkészítésében segítettek.
Elıször is hálásan köszönöm Dr. Raskó Istvánnak, hogy lehetıvé tette a kutatómunkám
végezését a csoportjában, köszönöm a tudományos irányítást és az anyagi támogatás
biztosítását.
Nagyon köszönöm a Humán Molekuláris Munkacsoport minden volt és jelenlegi tagjának a
jó munkahelyi légkört, a segítıkészséget és azt a támogatást, amit a hosszú évek alatt
nyújtottak. Külön köszönöm Dr. Mórocz Mónikának, Szécsényi Anitának, Dr. Álmos
Péternek, Dragon Annamáriának és Grózer Zsuzsannának, hogy a dolgozat megírásához
szakmai segítséget nyújtottak. Nem tudok elég hálás lenni Lehıcz Istvánnénak és Radóné
Dudás Máriának az asszisztensi munkájukért és barátságukért.
Köszönetemet szeretném kifejezni Dr. Endreffy Emıkének és Dr. Illés Tamásnak a
munkához szükséges mintákért; Dr. Zvara Ágnesnek a microarray kísérletekben nyújtott
baráti segítségért és végül, de nem utolsó sorban Dr. Monostori Évának az irántam való
türelméért és a dolgozat megírásához szükséges baráti támogatásért.
Köszönöm szüleimnek, férjemnek és gyerekeimnek, hogy a munkámmal járó nehézségeket
elviselték, a dolgozat megírását türelemmel fogadták és mindenben támogattak.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
79
ÖSSZEFOGLALÁS
A XXI. században a társadalom nagyobb részét érintı, komplex betegségek etiológiai
vizsgálatai kerültek a humángenetikai kutatások középpontjába. Ezekre a betegségekre
jellemzı, hogy több gén hibás mőködése a környezeti hatásokkal kölcsönhatásban alakítja ki
ıket és genetikailag is heterogének lehetnek, például a szív- és érrendszeri betegségek. Az
általunk vizsgált, a felnıttkor elején valamely ismert ok nélkül kialakuló gerincferdülés (AIS)
olyan komplex betegség, amelynek genetikai hátterérıl a kiterjedt kutatások ellenére nem
tudunk sokat. Az AIS klinikai kórképe változatos, eltérı lehet a kialakulás ideje, a görbület
progressziójának mértéke, sebessége és nagysága. Az esetek döntı többségében a
gerincferdülés kezdete a serdülıkorra tehetı, ezért valószínősítik, hogy a növekedéssel
kapcsolatos hormonális, metabolikus és biomechanikai változások szerepet játszhatnak az
AIS pathomechanizmusában. A lányoknál a rövidebb idı alatt és gyorsabban lezajló
serdülıkori fejlıdés lehet az oka annak, hogy 3.4-szer gyakoribb a nagyobb mértékő
gerincferdülés a körükben, mint a fiúknál. Az elmúlt 40 év alatt számos területen keresték a
deformitás kialakulásának okát, vizsgálták: a csont- és a kötıszövet, valamint a harántcsíkolt
izmok strukturális eltéréseit; a neurológiai rendellenességeket; a hormonháztartás
egyensúlyát; a biomechanikai hatásokat; a genetikai tényezıket. Az etiológiai kutatások
kezdetén kevés figyelem irányult a genetikai háttér felé, mivel a betegség többnyire
szórványosan lépett fel, és gyakran a családon belül halmozódó esetek sem mutattak
egyértelmő öröklıdést. Az AIS ma elfogadott, genetikai modellje szerint a betegség
kialakulása két szakaszra osztható, egy iniciációs és egy progressziós fázisra és feltételezések
szerint más-más gének hibái felelısek a hajlamosításra és a súlyosbodásra.
Magyarországon a Pécsi Tudományegyetem Általános Orvosi Kar Ortopédiai Klinikáján
végzik az évi 450 új eset döntı többségének mőtéti korrekcióját. A dolgozatomban leírt
munkát szoros együttmőködésben végeztük a klinika tanszékvezetı professzorával Dr. Illés
Tamással, akitıl az AIS-es betegek vérmintáit és a korrekciós mőtét során a gerincoszlop két
oldaláról származó izombiopsziákat kaptuk.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
80
A munkánk során célul tőztük ki, hogy molekuláris szintő vizsgálatokban az AIS-re
jellemzı további eltéréseket tárjunk fel a harántcsíkolt izmok transzkriptoma analízisével és
az eltérések hátterében álló genetikai tényezıket vizsgáljuk.
Elıször CHIP microarray módszerrel kívántuk összehasonlítani külön-külön az:
1. Egészséges és scoliotikus paravertebrális izmok génkifejezıdési mintázatát,
valamint a:
2. Scoliotikus izmokban a görbület konvex és konkáv oldali génkifejezıdését.
Másodszor a génexpressziós vizsgálatok által jelölt gének SNP polimorfizmusát terveztük
vizsgálni asszociációs analízissel eset-kontroll tanulmányban.
A két microarray analízis eredményeként egy 187, illetve egy 63 tagból álló génlistát
kaptunk, melyek tagjait négy szempont alapján elemeztük annak eldöntésére, melyek azok a
gének, amelyek számításba jöhetnek a további vizsgálatokban.
1. Elıször biológiai folyamatokat jelölı génontológiai (GO) kategóriákhoz rendeltük a
génlista tagjait és olyan GO kategóriákat kerestünk, amelyekben a vizsgált gének a
humán genom átlagos GO eloszlásához képest magasabb arányban vannak jelen és
szerepük lehet az AIS kialakításában. A gerincferdüléses betegek gerincoszlopának
konvex és konkáv oldalán génkifejezıdésbeli különbséget mutató gének két GO
kategóriában, a lipid anyagcserében (GO:0006629) és a morfogenezisben
(GO:0007275) nagyobb arányban jelentek meg, ebbe a két kategóriába a 63 génbıl
összesen 18-at tudtunk besorolni.
2. A génlisták tagjainak közös biológiai, metabolikus útvonalban való szereplése és
együttes génexpressziós változásuk jelzi az adott folyamat fontosságát, ilyen útvonal
keresésekor több gén is expressziós változást mutatott az adipociták jelátviteli
mechanizmusában (KEGG_Pathway: hsa04920), ami a lipid anyagcsere érintettségét
az AIS-ben alátámasztja.
3. Meghatároztuk a génlistáinkon lévı gének kromoszóma lokalizációját és két
szempont szerint is átnéztük a gének elhelyezkedését. Egyrészrıl kerestünk olyan
kromoszóma szakaszokat, ahová több gén is térképezıdik közel egymáshoz, mert egy
helyen lévı és egyforma génkifejezıdésbeli eltérést mutató gének esetén
gondolhatunk valamiféle genomszintő átrendezıdésre az adott régióban (delécióra
vagy inszercióra). Ilyet nem tudtunk kimutatni. Másrészrıl kiválogattuk azokat a
géneket, amelyek olyan kromoszóma szakaszra esnek, amit családokon kapcsoltsági
analízissel a szakirodalom az AIS-hez kötöttnek tekint. Nyolc ennek megfelelı gént

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
81
találtunk, amelyeknek további vizsgálata szükséges.
4. Irodalmi kutatásokat végeztünk és a vonatkozó GeneCards leírások és számos
publikáció alapján összefüggést próbáltunk találni a gének jelenleg ismert funkciója
és az AIS pathomechanizmusára vonatkozó jelenleg ismert modellek között. Az
érdekesnek talált gének közül ötöt már tovább vizsgáltunk, de két ígéretes gén, az
izomerı meghatározásában szereplı a disztonin és a sérült izmok regenerálódásában
fontos tenzin célzott vizsgálata még várat magára.
A microarray kísérletek kiváló kiindulási alapként szolgálnak, mivel látótérbe hoznak a
vizsgált jelenséggel kapcsolatban álló új géneket. Következı lépésként azonban mindenképp
szükséges ellenırizni egy független módszerrel, hogy valóban fennáll-e a megfigyelt
génkifejezıdésbeli különbség. Ez a független módszer napjainkban általában a kvantitatív
valós idejő polimeráz láncreakció (QRT-PCR). A vizsgálatra kiválasztott 13 gén közül
egyedül a leptin (LEP) esetén tudtunk kimutatni eltérést a QRT-PCR-rel is. A scoliosisos
betegekben a görbült gerincoszlop konvex oldalán a leptin relatív mRNS mennyisége 2,5-
szerese a konkáv oldalénak és ennek az eredménynek kapcsán kezdtük el vizsgálni a LEP
gént.
A leptin szerepe az AIS kialakításában napjainkban merült fel a szakirodalomban és
eddig egyetlen közlemény jelent meg, amelyben az AIS-es lányokban alacsonyabb leptin
szérumszintet találtak az egészséges kontrollokhoz viszonyítva. Egy eset-kontroll
tanulmányban asszociációs analízissel a leptin gén promóter régiójában vizsgáltuk annak az
SNP-ének az elıfordulási gyakoriságát, amelynek szerepe lehet a leptin gén szabályozásában.
Az asszociációs vizsgálatba bevontuk az interleukin 6 (IL6) szabályozó régiójában lévı
polimorfizmust is, mivel egy közlemény szerint a gerincferdüléssel asszociációt mutatott. A
két SNP, a LEP G-2548A és a IL6 G-174C kimutatására a PCR-RFLP módszert használtuk
és 126 idiopathiás scoliosisos beteg és 197 egészséges, egymással nem rokon személy
esetében határoztuk meg a jelenlévı allélokat. Mindkét populáció Hardy-Weinberg
egyensúlyban volt valamennyi vizsgált lókuszra. Egyik SNP esetén sem találtunk
szignifikáns különbséget sem az allélgyakoriságban, sem a genotípus gyakoriságban az AIS-
es és a kontroll csoport között de a különbözı genotípusok esélyhányadosainak
meghatározása után úgy tőnik a LEP-2548A allél jelenléte lehet hajlamosító faktor az AIS-
ben. Vizsgálatainkban az AA vs. GG: OR=1.513(CI:0.894-2.562), ami a komplex betegségek
esetén a hajlamosító allélokra jellemzı értéknek tekinthetı. Az esélyhányados értéke megnıtt
akkor, ha külön vizsgáltuk a lányokat (AA vs. GG: OR=2.02(CI:0.824-4.961)), ami azt jelzi,
hogy a lányokban nagyobb szerepe lehet az AIS kialakításában a LEP-2548A allélnak, mint a

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
82
fiúkban. Ugyanakkor nem találtunk összefüggést a görbület mértéke (Cobb˚) és az SNP által
meghatározott allélok között. Az IL6 G-174C polimorfizmusát vizsgálva nem tudtuk
megerısíteni a korábban, mások által leírt asszociációt. A reprodukálhatóság hiánya
egyáltalán nem egyedi, sıt a komplex betegségekre jellemzınek tartja a szakirodalom, ezért
szükséges több, megfelelı statisztikai erıvel rendelkezı, független populáción elvégezni az
asszociációs vizsgálatokat.
Mivel multifaktoriális öröklıdéső betegségekben a gének nem önmagukban hatnak,
hanem egymással kölcsönhatásban, ezért megvizsgáltuk létezik-e episztatikus kapcsolat az
általunk vizsgált két gén alléljei között. A gén-gén kölcsönhatás feltárásához két eljárást is
használtunk, az elsı az MDR analízis, amely kisszámú mintaelem esetén is kimutatja az
episztázist és alkalmas a gének közötti szinergizmus kimutatására akkor is ha az allél
eloszlásban nem mutatkozik eltérés a beteg és a kontroll csoport között. Az analízis
eredményeként megkaptuk azokat a genotípus kombinációkat , amelyek elıfordulása esetén
nagyobb a kockázata az AIS kialakulásának. A másik statisztikai módszer, ami a
génkombinációk hatásának maximum likelihood becslését alkalmazva megadja, hogy egy
adott génkombináció milyen mértékben emeli meg a betegség kialakulásának esélyét egy
másik génkombinációhoz viszonyítva. Az elemzés után a leginkább hajlamosító
allélkombináció (IL6 CC-LEP AA) hordozójának 4.667-szor nagyobb az esélye az AIS
kialakulására, mint az (IL6 GG-LEP GG) kombináció hordozójának. Ez az esélyhányados
még magasabb a lányok esetén, tehát a hajlamosító allélkombináció (IL6 CC-LEP AA)
hordozása még nagyobb kockázatot jelent náluk (OR= 6.667 (CI:0.597-74.506). Statisztikai
módszerrel megvizsgálva a genotípus kombinációk és a Cobb˚ közötti kapcsolatot, nem
találtunk semmiféle összefüggést köztük. A vizsgálati eredményeink alapján úgy gondoljuk,
hogy a LEP AA-IL6 CC genotípus kombináció az AIS kialakulására hajlamosít, nem pedig a
progressziójában játszhat szerepet.
Jelen dolgozatban az AIS kutatásában újszerő megközelítést használva a paravertebrális
izmok génexpressziós mintázatát vizsgáltuk és sikerült transzkriptoma szinten megjelenı
eltérést feltárni és független módszerrel visszaigazolni a leptin gén esetén. A további
genetikai és statisztikai elemzések az AIS kialakításában szerepet játszható gén-gén
kölcsönhatás felderítéséhez vezetett és elsıként sikerült kimutatnunk, hogy két adipokint
(leptint, interleukin-6-ot) termelı gén szabályozását befolyásoló polimorfizmus együttes
jelenléte szóba jöhet, mint hajlamosító faktor az AIS etiológiájában.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
83
SUMMARY
In the 21st Century the human genetic studies focus on revealing etiology of common
complex diseases. Complex diseases are believed to be caused by a several number of genes,
usually interacting with various environmental factors and to be genetically heterogenic like
coronary heart diseases. Adolescence idiopathic scoliosis, a multifactorial disease represents
a three-dimensional deformity of spine with lateral curvature in which the causes are
unknown. Many studies are conducted to uncover contributing genetic factors of AIS,
however, genetics underlying AIS have not been identified yet. Adolescent idiopathic
scoliosis occurs at the peripubertal period (between 10 and 16 years of age) supposing the
role in AIS pathomechamism the changes in endocrine, metabolic and biomechanical
alterations during growth spurt. It is widely accepted that scoliosis has a sex-conditioned
manifestation, girls have a higher risk for development of curve progression the ratio of girls
to boys is 3.4/1 with curves greater than 30 degrees. In the last 40 years many different
research area were involved in hunting for etiology factors of spine deformity like connective
tissue abnormalities, asymmetries in the central nervous system, hormonal variation and
genetics.
According to the most accepted model for inheritance the genetic factors can be divided into
predisposing factors, initiating factors and contributing factors in accordance with their role
in the pathogenesis of AIS. Moreover, the current view is that there are no major genes
causing AIS, but combinations of sequence variants of possible predisposition genes may
have synergic effects on determining the deformity. Therefore, it is important to identify such
sequence variants and combined genotype pattern of these susceptibility genes to get a better
understanding of AIS pathogenesis.
The major objectives of the work presented in my dissertation were the investigation of the
genetic background of AIS and the identification of novel molecular alterations that are
associated with this disease. The study was performed in close collaboration with Prof.
Tamás Illés at Department of Orthopaedics, Medical School University of Pécs, who
provided us the clinical samples of AIS patients.
For molecular biology studies I used two different molecular biological approaches.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
84
In the first approach I have investigated the gene expression pattern of the paravertebral
muscles responsible for the stability of the spinal column. By using the DNA microarray
technology in transcriptome profiling experiments I have compared:
I. The gene expression pattern of the paravertebral muscles in healthy control
individuals and AIS patients.
II. The gene expression pattern of the paravertebral muscles in the concave and the
convex side of the spinal column deformity.
I have collected those genes that could be candidates for further studies according to the
following criteria:
1. They are significantly accumulated in a functionally relevant Gene Ontology
group.
2. They are accumulated in a functionally relevant metabolic or signal transduction
pathway.
3. They are located in a chromosome region has been previously reported to be
involved in AIS.
4. According to the scientific publications reliable hypothesis could be built regarding
its involvement in AIS pathomechanism.
In the second approach the identified potential candidate genes were investigated in case-
control SNP polymorphism association studies. The data resulted by these experiments was
subjected to different statistical evaluation processes suitable for the identification of the
genetic interactions and the determination whether a particular gene is involved in the onset
or in the later progression phase of the AIS.
For revealing the gene expression differences in AIS 95 total RNA samples have been
prepared from muscle biopsies of patients subjected to correction surgery interventions. In
collaboration with the Functional Genomics Group of the BRC two microarray transcription
profiling experiments have been performed. In the first experimental layout I have compared
the gene expression levels in healthy and AIS individuals. In the second I have revealed the
differences in the transcriptional landscape of muscle samples derived from the concave and
the convex side of the spinal column deformity. To select genes for further genetic
investigations, the elements of the gene lists have been evaluated regarding to the following
four criteria:
1. The members of the gene lists has been classified according to their Gene Ontology
annotation, and the significantly overrepresented GO categories that functionally

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
85
might be involved in the AIS etiology have been identified. Genes from the lipid
metabolism (GO:0006629) and morphogenesis (GO:0007275) categories were
enriched in gene set showing expression difference in the concave and convex side of
the spinal cord deformity. These GO categories contained 18 out of the total 63 genes
of this list.
2. The simultaneous accumulation of genes with significantly altered expression in
particular metabolic or signal transduction networks might help to reveal molecular or
cell physiological processes involved in the appearance of a disease phenotype. In our
microarray investigations an adipocyte signalling cascade (KEGG_Pathway:
hsa04920) proved to be overrepresented in the significant gene list, indicating the
potential role of lipid metabolism in AIS disorder.
3. I have identified the chromosomal localization of genes showing significant
expression difference in microarray experiments and selected 8 of them to be
localized to chromosomal region have previously been reported in family based
linkage analysis as could be involved in AIS.
4. Performing very exhaustive data mining in different biomedical literature and gene
annotation databases like Pubmed, GenCards or OMIM, I tried to find connections
between the known functions of the selected genes and the different AIS pathogenesis
models suggested up till now by the scientific community working in this field. Two
of the identified genes might be very interesting in this respect. Distrophin (DTS) has
well characterized role in affecting the muscle strength, while tensin has an important
function in the regeneration process of the wounded muscles.
It is always necessary to validate the microarray results by an independent method. In bulk
majority of the currently applied experimental strategies this independent method is the
quantitative real time polymerase chain reaction (QRT-PCR). In QRT-PCR assay the
abundance of leptin gene relative mRNA was 2.5 times higher in the convex side of the spine
in AIS patients. These findings inspired me to make further studies concerning the
involvement of leptin gene in AIS pathogenesis.
In a case-control association study I have investigated the association of a promoter SNP in
the leptin gene with the manifestation of the AIS phenotype. In the same assay I have also
analyzed another SNP from IL6 gene promoter region that has been reported to be associated
with AIS. By using the PCR-RFLP method for the concurrent detection of LEP G-2548A and
IL6 G-174C polymorphisms I have determined the genotypes in 126 AIS and 197 healthy
individuals. Both investigated populations were in Hardy-Weinberg equilibrium at these loci.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
86
No significant differences were found for allele and genotype frequencies of the
polymorphisms of LEP G-2548A and IL6 G-174C between cases or controls. However the
determination of the odds ratio of the different genotypes indicates that the presence of the
LEP-2548A allele might be a susceptibility factor for AIS. In my studies the odds ratio of
LEP-2548 AA vs. LEP-2548 GG was OR=1.513 (CI:0.894-2.562) which is generally
accepted as an indicator for susceptibility alleles in the case of complex diseases. The odds
ratio value was further increased (LEP-2548 AA vs. LEP-2548 GG: OR=2.02(CI:0.824-
4.961)) when the female subpopulations was subjected, suggesting that in females LEP-
2548A allele might have a greater impact on the manifestation of AIS than in males. In the
other hand I didn’t find any association between the alleles determined by these SNPs and the
extent (Cobbo) of the spinal curve.
I have investigated whether there is any epistatic interaction between the alleles of leptin and
IL6 in respect of the manifestation of idiopathic scoliosis. I have used two different
approaches for revealing the gene-gene interactions. MDR analysis is suitable for detection of
epistatic interactions in such situations when the sample size is relatively small for the
investigation. As a result of this study the genotype combinations had increased risk of the
appearance of AIS could be emerged. The other statistical process based on the maximum
likelihood estimation can give a quantitative answer how a particular allelic combination
increases the chance of AIS manifestation compared to the other allelic patterns. By
performing these studies I have demonstrated that the IL6 CC-LEP AA allele combination
has the strongest susceptibility effect, which is 4.667 times higher to that of appear in the
presence of IL6 GG-LEP GG alleles. This relative odds ratio is even higher in the case of
female target population (OR= 6.667 (CI:0.597-74.506) indicating a higher genetic risk for
idiopathic scoliosis.
In the studies described in my Ph.D. dissertation I used a novel approaches, namely
microarray analyses in AIS research for the investigation of the gene expression pattern of the
paravertebral muscles. I could identify several genes where the altered gene expression
indicates their potential role in AIS pathogenesis. An independent method (QRT-PCR) was
successfully used to collect supporting evidence regarding the detected gene expression
difference in the case of leptin. Further genotyping studies and statistical investigations lead
to the identification of a gene-gene interaction resulting in increased susceptibility for AIS. I
could demonstrate that the simultaneous occurrence of allelic variants determined by single
nucleotide polymorphisms in the regulatory promoter regions of two adipokine genes (IL6
and leptin) might be a susceptibility factor in AIS etiology.

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
87
KÖZLEMÉNYEK LISTÁJA
1. Czibula, A; Leiker, G; Rasko, I
Changes in alkylation damage removal during in vitro neuronal differentiation
ACTA BIOLOGICA HUNGARICA Volume: 48 Issue: 1 Pages: 113-120 (1997)
2. Czibula, A; Morocz, M; Bachrati, CZ, et al.
Hunt for genetic susceptibility in a complex disease
JOURNAL OF MOLECULAR STRUCTURE-THEOCHEM Volume: 666 Pages: 681-686
(2003)
3. Gervain J, Czibula A, Simon J, Kalmár T.
Structural analysis of the PKR-binding region of HCV 1b samples from patients with chronic
hepatitis C and the correlation with IFN-sensitivity
ORV HETIL. Volume:144 Issue: 24 Pages: 1179-84( 2003) Hungarian.
4. Bogácsi-Szabó E, Kalmár T, Csányi B, Tömöry G, Czibula A, Priskin K, Horváth F,
Downes CS, Raskó I.
Mitochondrial DNA of ancient Cumanians: Culturally Asian steppe nomadic immigrants with
substantially more western Eurasian mitochondrial DNA lineages
HUMAN BIOLOGY Volume: 77 Issue: 5 Pages: 639-662 ( 2005)
5. Ion G, Fajka-Boja R, Kovács F, Szebeni G, Gombos I, Czibula A, Matkó J, Monostori E.
Acid sphingomyelinase mediated release of ceramide is essential to trigger the mitochondrial
pathway of apoptosis by galectin-1
CELLULAR SIGNALLING Volume: 18 Issue: 11 Pages: 1887-1896 (2006)
6. Hegedus Z, Czibula A, Kiss-Toth, E
Tribbles: novel regulators of cell function; evolutionary aspects
CELLULAR AND MOLECULAR LIFE SCIENCES Volume: 63 Issue: 14 Pages: 1632-
1641 (2006)

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
88
7. Zana M, Szécsényi A, Czibula A, Bjelik A, Juhász A, Rimanóczy A, Szabó K, Vetró A,
Szucs P, Várkonyi A, Pákáski M, Boda K, Raskó I, Janka Z, Kálmán J.
Age-dependent oxidative stress-induced DNA damage in Down's lymphocytes
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Volume:345
Issue:2 Pages: 726-733 (2006)
8. Hegedus Z, Czibula A, Kiss-Toth, E
Tribbles: A family of kinase-like proteins with potent signalling regulatory function
CELLULAR SIGNALLING Volume: 19 Issue: 2 Pages: 238-250 (2007)
9. Tömöry G, Csányi B, Bogácsi-Szabó E, Kalmár T, Czibula A, Csosz A, Priskin K, Mende
B, Langó P, Downes CS, Raskó I
Comparison of maternal lineage and biogeographic analyses of ancient and modern
Hungarian populations
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY Volume: 134 Pages: 354-368
(2007)
10. Sung HY, Guan H, Czibula A, King AR, Eder K, Heath E, Suvarna SK, Dower SK,
Wilson AG, Francis SE, Crossman DC, Kiss-Toth E.
Human tribbles-1 controls proliferation and chemotaxis of smooth muscle cells via MAPK
signaling pathways
JOURNAL OF BIOLOGICAL CHEMISTRY Volume: 282 Issue: 25 Pages: 18379-18387
(2007)
11. Almos PZ, Horváth S, Czibula A, Raskó I, Sipos B, Bihari P, Béres J, Juhász A, Janka Z,
Kálmán J.
H1 tau haplotype-related genomic variation at 17q21.3 as an Asian heritage of the European
Gypsy population.
HEREDITY Volume: 101 Issue: 5 Pages: 416-419 (2008)
12. Csányi B, Bogácsi-Szabó E, Tömöry G, Czibula A, Priskin K, Csõsz A, Mende B, Langó
P, Csete K, Zsolnai A, Conant EK, Downes CS, Raskó I
Y-chromosome analysis of ancient Hungarian and two modern Hungarian-speaking
populations from the Carpathian Basin

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
89
ANNALS OF HUMAN GENETICS Volume: 72 Pages: 519-534 (2008)
13. Matusek,T.; Gombos,R.; Szecsenyi,A.; Sanchez-Soriano,N.; Czibula,A.; Pataki,C.;
Gedai,A.; Prokop,A.; Rasko,I.; Mihaly,J.
Formin proteins of the DAAM subfamily play a role during axon growth
JOURNAL OF NEUROSCIENCES Volume: 28 Issue: 49 Pages: 13310-13319 (2008)
14. Nagy,D.; Bogacsi-Szabo,E.; Varkonyi,A.; Csanyi,B.; Czibula,A.; Bede,O.; Tari,B.;
Rasko,I.
Prevalence of adult-type hypolactasia as diagnosed with genetic and lactose hydrogen breath
tests in Hungarians
EUROPEAN JOURNAL OF CLINICAL NUTRITION Volume: 63 Issue: 7 Pages: 909-912
(2009)
15. Mórocz M, Czibula A *, Grózer Zs, Szécsényi A, Álmos P, Dragon A, Raskó I, Illés T
Association study of MMP-3, IL-6, Leptin, BMP4, MTNR1B Gene Promoter Polymorphisms
and Adolescent Idiopathic Scoliosis
Közlésre elküldve a SPINE-ba, revízió alatt
*megosztott elsı szerzıség
Konferencia kiadványok
1. Regulation of the bovine leukemia-virus basal transcription
Kiss-Toth E, Unk I, Bachrati C, Czibula A
JOURNAL OF CELLULAR BIOCHEMISTRY Pages: 48-48 Supplement: 18C (1994)
2. Repair of alkylation damage by differentiating mouse teratocarcinoma cells
Czibula A, Leiker G, Margison GP, T, Rasko I
JOURNAL OF CELLULAR BIOCHEMISTRY Pages: 278-278 Supplement: (1995)
3. Changes of DNA repair during in vitro cellular differentiation
Czibula A, Gerencser A, Bachrati C, Borda S, Johnson RT, Rasko I
CELL BIOLOGY INTERNATIONAL Volume: 20 Pages: 220-220
(1996)

Czibula Ágnes: Az idiopathiás scoliosis molekuláris biológiai vizsgálta Ph.D. értekezés, 2010
90
4. A putative molecular genetic susceptibility allele for idiopathic scoliosis
Czibula A, Morocz M, Csiszar A, Bachrati C, Olah E, Szeszak F, Morava E, Szappanos L,
Rasko I
EUROPEAN JOURNAL OF HUMAN GENETICS Volume: 10 Pages: 76-77 (2002)
5. The correlation of genom mutations in HCV-1b with IFN-sensitivity in Hungarian patients
Gervain J, Simon J, Czibula A, Kalmar T
JOURNAL OF GASTROENTEROLOGY AND HEPATOLOGY Volume: 21 Pages: A139-
A139 Supplement: 2 (2006)