adenovirus-mediated transfer of the thyroid sodium/iodide
TRANSCRIPT

[CANCER RESEARCH 60, 3484–3492, July 1, 2000]
Adenovirus-mediated Transfer of the Thyroid Sodium/Iodide Symporter Gene intoTumors for a Targeted Radiotherapy1
Anne Boland,2 Marcel Ricard, Paule Opolon, Jean-Michel Bidart, Patrice Yeh, Sebastiano Filetti,Martin Schlumberger, and Michel PerricaudetUMR1582 CNRS-IGR-Rhone-Poulenc [A. B., P. O., P. Y., M. P.] and Service de Physique [M. R.], Departement de Biologie Clinique [J-M. B.], and Service de Medecine Nucleaire[M. S.], Institut Gustave Roussy [M. S.], 94805 Villejuif, France, and Dipartimento di Medicina Sperimentale e Clinica, Universita di Catnzaro, 88100 Catanzaro, Italy [S. F.]
ABSTRACT
The Na1/I2 symporter (NIS) present in the membranes of thyroid cellsis responsible for the capacity of the thyroid to concentrate iodide. Thisallows treatment of thyroid cancers with 131I. We propose to enlarge thistherapeutic strategy to nonthyroid tumors by using an adenoviral vectorto deliver the NIS gene into the tumor cells. We constructed a recombinantadenovirus encoding the ratNIS gene under the control of the cytomeg-alovirus promoter (AdNIS). Infection of SiHa cells (human cervix tumorcells) with AdNIS resulted in perchlorate-sensitive125I uptake by thesecells to a level 125–225 times higher than that in noninfected cells. Similarresults were obtained for other human tumor cell lines, including MCF7and T-47D (mammary gland), DU 145 and PC-3 (prostate), A549 (lung),and HT-29 (colon), demonstrating that the AdNIS vector can function intumor cells of various origins. In addition, AdNIS-infected tumor cellswere selectively killed by exposure to131I, as revealed by clonogenicassays. To assess the efficiency of this cancer gene therapy strategyin vivo,we injected the AdNIS vector in human tumors (SiHa or MCF7 cells)established s.c. in nude mice. Immunohistological analysis confirmed theexpression of the NIS protein in the tumor. Three days after intratumoralinjection, AdNIS-treated tumors could specifically accumulate125I or 123I,as revealed by kinetics and imaging experiments. A quantitative analysisdemonstrated that the uptake in AdNIS-injected tumors was 4–25 timeshigher than that in nontreated tumors. On average, 11% of the totalamount of injected 125I could be recovered per gram of AdNIS-treatedtumor tissue. Altogether, these data indicate that AdNIS is very efficientin triggering significant iodide uptake by a tumor, outlining the potentialof this novel cancer gene therapy approach for a targeted radiotherapy.
INTRODUCTION
Iodide accumulation in the thyroid can reach concentrations 20–40-fold over the plasma levels (1). Iodide captured by the thyroid isorganified,i.e. oxidized into iodine and subsequently used to iodinatethyroglobulin, the precursor of thyroid hormones. This unique capac-ity to concentrate and organify iodide allows the use of radioactiveiodine isotopes (131I) for the treatment of differentiated thyroid can-cers and hyperthyroidism (2, 3). However, iodine metabolism abnor-malities such as defects in iodide uptake and/or organification arefrequently observed in thyroid cancer tissues, thereby seriously com-promising the efficiency of radioiodine treatment (4–6).
Iodide accumulation in the thyroid gland is ensured by the NIS,3 a
transmembrane glycoprotein present in the basolateral pole of thyroidfollicular cells. NIS-mediated iodide uptake is an active transportprocess that occurs against the electrochemical gradient in I2 anionsand is competitively inhibited by thiocyanate and perchlorate anions(7, 8). The cDNAs of the rat and humanNISgenes have been clonedrecently and code for proteins of 618 and 643 amino acids, respec-tively (9, 10). The two proteins are 84% identical and have beenpredicted to be integral membrane proteins displaying 12 (9–11) or 13membrane-spanning domains (12). NIS expression is not strictlylimited to the thyroid but also occurs in several extrathyroidal tissues,including the salivary glands, the gastric mucosa, and the mammarygland (13). In these tissues, however, iodide is not organified (13).
The cloning of theNIS gene constitutes an important step towardthe understanding of the molecular mechanisms underlying iodidetransport abnormalities in thyroid pathologies. Indeed, several casesof hypothyroidism with low iodide uptake were linked to inactivatingmutations in theNISgene (14, 15). Similarly, the low or absent iodidetransport observed in thyroid cancer tissues was correlated with a lowor absent NIS expression (6, 16, 17). Importantly, NIS-mediatediodide transport does not require the follicular organization of thethyroid, as demonstratedin vitro in bovine (18), porcine (19), human(20), and rat thyroid cells (7). Severalin vitro studies also showed thattransfer of theNISgene into nonthyroid cells, either by transfection ofNIScDNA (9, 10, 21, 22) or with a retroviral vector (23), led to iodideuptake by the transduced cells. Coupling delivery of theNISgene intotumor cells with131I administration may therefore open new avenuesto treat cancer.
Adenoviral vectors are particularly well suited for cancer genetherapy. They lead to a transient but robust expression of the trans-gene, and efficientin vivogene transfer has been reported in numeroustissues, including the thyroid (24–26). In addition, concentrated ade-novirus preparations can be obtained, which constitutes a clear ad-vantage over other viral vectors such as retroviruses for an optimalinvivo gene transfer. The commonly used adenoviral vectors are ex-tremely attenuated and lack at least the viral early transcriptionregions E1 (essential for replication) and E3. Such vectors still exhibitsome level of cytotoxicity, which can be viewed as an advantage fordestructive strategies such as cancer gene therapy (for review, seeRef. 27).
The aim of this work is to demonstrate the feasibility of using anadenoviral vector to deliver theNISgene into human tumors, with thegoal of enabling them to concentrate radioactive iodine. For thispurpose, we constructed and characterized a recombinant adenovirusexpressing the ratNISgene (AdNIS) as a first step toward a targetedradiotherapy of tumors.
MATERIALS AND METHODS
Construction of the Recombinant AdNIS Virus. AdNIS is aDE1–DE3recombinant adenovirus expressing the ratNIS gene under the control of theimmediate early promoter of the cytomegalovirus (pCMV). The ratNIS gene[nucleotides 74–2046 of the published sequence (9)] was cloned as aAatII(blunt-ended by Mung Bean nuclease treatment)-HindIII fragment in thePvuII-HindIII sites of the pCEP-4 vector (Invitrogen) to add a promoter
Received 12/13/99; accepted 4/27/00.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 A. B. is financed by a Marie Curie Research and Training Grant from the Biotech-nology research and technological development program from the European Community.This work was supported by grants from the Centre National de la Recherche Scientifique,the Institut Gustave Roussy, Electricite de France, and the Ligue Nationale Contre leCancer. We acknowledge the support of Associazione Italiana per la Ricerca sul Cancro(to S. F.).
2 To whom requests for reprints should be addressed, at Laboratoire de Vectorologieet Transfert de Genes UMR1582, Institut Gustave Roussy, 39 rue Camille DesmoulinsPR2, 94805 Villejuif, France. Phone: (33)-1-42-11-50-82; Fax: (33)-1-42-11-52-45; E-mail: [email protected].
3 The abbreviations used are: NIS, sodium iodide symporter; rNIS, rat NIS; FBS, fetalbovine serum; PFU, plaque-forming unit(s); MOI, multiplicity of infection; ATCC,American Type Culture Collection; bHBSS, buffered HBSS.
3484
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

(pCMV) and a polyA (SV40 polyA). The obtained plasmid was called pAB1.A 3711-bp SspI-EcoRV fragment of pAB1 was then cloned in the shuttlevector pXL3048 linearized byEcoRV. pXL3048 is a KmR-SacB-ColE1 de-rivative (28) containing the left end of the Ad5 genome (nucleotides 1–386),a polylinker with three unique cloning sites (EcoRV,BamHI, andSalI), andpart of the Ad5pIX gene (nucleotides 3446–4296). The shuttle vector con-taining the NIS expression cassette was called pAB2. The recombinant adeno-viral genome encoding NIS was obtained by homologous recombinationbetween plasmids pAB2 and pXL3215 inEscherichia coli, as describedpreviously (28). pXL3215 contains the Ad5 genome bordered by twoPacIsites and carrying deletions within E1 (nucleotides 386–3446) and E3 (nucle-otides 28592–30470). After recombinational cloning inE. coli, the adenoviralgenome was excised byPacI digestion, and the AdNIS virus was recovered bytransfecting 10mg of PacI-digested DNA into 293 cells by the Lipofectamine-based procedure (Life Technologies, Inc.).
Control Adenoviral Vectors. Recombinant adenoviruses expressing notransgene (AdCO1) or encodingb-galactosidase (Adbgal) were used as neg-ative controls in this study and have been described previously (29, 30).
Virus Amplification, Purification, and Titration. All viral stocks wereprepared from infected 293 cells (31) by standard procedures (30). After atwo-step purification on CsCl gradients, viral stocks were desalted by usingPharmacia G50 columns (Orsay, France) and frozen at280°C in PBS con-taining 7% glycerol. Viral titers were calculated by dilution plaquing onto 911cells (32) and expressed in PFU/ml.
Cell Lines. Unless stated otherwise, all cell culture media and reagentswere purchased from Life Technologies, Inc. Rat thyroid FRTL-5 cells (ATCCCRL-8305) were routinely grown in Coon’s modified Ham’s F12 medium(Sigma) supplemented with 5% donor calf serum (Life Technologies, Inc.16030) and 10mg/ml insulin (Sigma I 1882), 10 nM hydrocortisone (Sigma H0396), 5mg/ml transferrin (Sigma T 1147), 10 ng/ml somatostatin (Sigma S1763), 10 ng/ml glycyl-L-histidyl-L-lysine acetate (Sigma G 7387), and 10milliunits/ml thyrotropin (Sigma T 8931). SiHa (ATCC HTB-35), MCF7(ATCC HTB-22), T-47D (ATCC HTB-133), DU 145 (ATCC HTB-81), andHT-29 (ATCC HTB-38) cells were maintained in DMEM supplemented with10% heat-inactivated FBS. PC-3 cells (ATCC CRL-1435) were grown inF-12K Nutrient Mixture (Kaighn’s modification) supplemented with 10%heat-inactivated FBS. A549 cells (ATCC CLL-185) were maintained in min-imum Eagle’s medium supplemented with 10% heat-inactivated FBS and 1%nonessential amino acids. 293 and 911 cells were grown in minimum Eagle’smedium supplemented with 10% FBS and 1% nonessential amino acids andDMEM supplemented with 10% FBS, respectively.
Anti-NIS Antibodies. A peptide spanning the COOH-terminal (600–618)region of rNIS was synthesized by a conventional solid-phase method using anApplied Biosystems Model 431A peptide synthesizer. The identity and purityof the (600–618) peptide were verified by amino acid analysis and peptidemicrosequencing. The synthetic peptide was conjugated to keyhole limpethemocyanin using benzidine as the coupling agent on the Lys600 residue. Tworabbits were immunized by intradermal injection of the synthetic peptide-carrier conjugate. After two boosts at 3-week intervals, animals were bled, andtheir sera were tested in an ELISA. Antisera, at various dilutions, were verifiedfor their capacity to react with the rNIS synthetic peptide coated on microtiterplates. Antibody binding was then revealed by peroxidase-labeled goat anti-rabbit antibody (Nordic, Tilburg, the Netherlands).
Immunofluorescence.SiHa cells were seeded in 4-wells SonicSeal Slides(Nunc, Inc.) at a density of 43 105 cells/well. The next day, cells wereinfected with virus-containing culture supernatant from the first viral amplifi-cation step; a 5-fold dilution was used, and infection was carried out for 1 h in200 ml of medium before the addition of 800ml of medium. Twenty-four hafter infection, cells were fixed for 15 min in PBS-4% formaldehyde, washedtwice with PBS, and permeabilized with PBS-0.2% Triton X-100 for 10 min.After two other washes with PBS, cells were incubated overnight at 4°C inPBS-0.5% FBS. The NIS protein was detected using the rabbit polyclonalantibodies described above; cells were incubated for 90 min with the anti-NISantibody diluted 1:500 in PBS, washed three times with PBS-0.5% FBS, andthen incubated for 30 min with a fluorescein-conjugated secondary antibody(Vector Laboratories, Inc., Burlingame, CA). After further washing in PBS-0.5% FBS, slides were mounted and observed with a fluorescence microscope.
Infection Conditions for Iodide Uptake Experiments. Cells were seededin 24-well plates 3 days before the experiment to achieve between 53 105 and
106 cells/well at the day of infection. Cell numbers were determined immedi-ately before infection as the average cell content of two wells. Cells wereinfected at the indicated MOI in 200ml of medium for 1 h, and then 800ml ofmedium were added in each well. For each cell line, the medium used forinfection was the same as the culture medium.
In Vitro 125I Uptake Experiments. Iodide uptake experiments were per-formed 28–30 h after virus infection, using the method of Weisset al. (7).Briefly, cells were washed once with 1 ml of HBSS buffered to pH 7–7.5 with10 mM HEPES (bHBSS). Iodide uptake was then initiated by adding 0.5 ml ofbHBSS containing 0.1mCi of 125I per well. After the indicated time of contactwith iodide, cells were washed once with ice-cold bHBSS and incubated for 20min in 1 ml of ice-cold ethanol. The ethanol was then recovered, and radio-activity was quantified (cpm) with a well gamma-counter (Beckman gamma5500 B).
In Vitro Cell Killing with 131I and Clonogenic Assay.Cells were seededin 24-well plates and infected at a MOI of 10, as described above. Twenty-fiveh after infection, cells were washed once with 0.5 ml of bHBSS and incubatedwith 0.5 ml of bHBSS (control) or 0.5 ml of bHBSS containing 10mCi of 131I.After 5 h of contact with 131I, cells were washed twice with bHBSS,trypsinized, and counted. For each condition [noninfected cells, AdNIS-infected cells, and cells infected with a control adenovirus (AdCO1)], cellswere plated in triplicate in 6-well plates (1000 cells/well) and incubated for 1week at 37°C. Cells were then washed once with PBS and stained with a cristalviolet solution (for 250 ml, 0.5 g of cristal violet, 25 ml of 40% formaldehyde,50 ml of ethanol, and 175 ml of water). Colonies of more than 30 cells werecounted, and the means and SD were determined for each condition. Resultsare expressed as the percentage of surviving cells,i.e. the percentage ofcolonies obtained after treatment with131I compared to treatment with bHBSSalone, and are representative of two separate experiments.
Tumor Induction and in Vivo Iodide Uptake Experiments. Female nudemice (6 – 8 weeks of age) were irradiated (5 Gy) the day before injection ofthe tumor cells. Tumors were induced by s.c. injection of 200ml of sterilePBS containing 53 106 SiHa or MCF7 cells. In the case of MCF7 tumors,the cell suspension contained 50% Matrigel (Becton Dickinson). Whentumors had reached 5– 8 mm in diameter (approximatively 3 weeks aftercell injection), a 10-day thyroxine treatment was initiated to suppressthyroid iodine uptake; each day, animals were injected i.p. with 2mg ofL-thyroxine (Roche) diluted in 100ml of PBS. Seven days after the onsetof the L-thyroxine treatment, the AdNIS virus was injected into the tumors(2 3 109 PFU in 100ml of PBS), and iodide uptake was assessed 3 dayslater. For kinetics and quantitative uptake experiments, 6mCi of 125I wereinjected i.p. in 200ml of sterile PBS. The presence of radioactive iodide inthe tumors was recorded up to 400 min after the injection of radioactiveiodide, using a small radiation-sensitive probe (Europrobe-Eurorad, Stras-bourg, France). For quantitative analysis of the amount of125I present inthe tumors, mice were sacrificed 90 min after the injection of radioactiveiodide, the weight of the tumors was determined, and radioactivity wasquantified using a calibrated well gamma-counter (Compugamma 1282;LKB) for 1 min. For experiments with131I, mice were injected i.p. with 30,60, or 90mCi of 131I in 200 ml of sterile PBS (corresponding in terms ofradioactivity/weight to doses generally used in human therapeutics); fiveanimals were included in each group, and tumor sizes were followed for 2weeks. For imaging experiments, 50mCi of 123I were injected i.p. in 200ml of sterile PBS; 90 min later, animals were anesthetized, and an imagewas taken (gamma camera DHD, SMV, BUC France). In all cases, thetumors were removed and analyzed by immunohistology.
Immunohistology. Removed tumors were fixed in 5% acetic acid, 75%absolute ethyl alcohol, 2% of formalin (40%), and 18% water for paraffinblock preparations. To examine the histological aspect of the tumor, paraffinsections (5-mm thick) were stained with H&E-saffron. The presence of the NISprotein was revealed by incubating the sections for 1 h with anti-NIS poly-clonal antibodies (see above) at a 1:1300 dilution and then incubating sectionsfor 30 min with a goat antirabbit secondary antibody conjugated to peroxidase(Envision; Dako). 3-Amino-9-ethylcarbazole (Envision kit; Dako) was used toreveal the markers, and sections were counterstained with Mayer’s hematox-ylin (1:2).
3485
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

RESULTS
Construction and Characterization of the Recombinant Adeno-virus Encoding the NIS Gene. We constructed an E1/E3-defectiverecombinant adenovirus encoding theNIS gene from rat thyroidFRTL-5 cells under the control of the immediate early promoter fromthe cytomegalovirus (AdNIS). To confirm expression of the NISprotein from the engineered adenovirus, we infected SiHa cells (hu-man cervix tumor cells) with virus-containing culture supernatant.Twenty-four h after infection, cells were fixed and permeabilized. Thepresence of the NIS protein was then detected by immunofluorescenceusing a rabbit polyclonal antibody directed against the COOH-termi-nal part of the NIS protein and a FITC-labeled antirabbit antibody.Fig. 1Ashows that AdNIS-infected cells were clearly labeled and thatthe NIS protein was localized at the cell surface. On the contrary,noninfected SiHa cells were not labeled (Fig. 1B). In addition, cellsinfected with a control adenovirus (Adbgal) and AdNIS-infected cellstreated with PBS in place of the primary antibody did not display any
staining (data not shown), confirming that the observed signal waslinked to the expression of the NIS protein. The AdNIS vector thusleads to high-level expression and correct localization of the NISprotein within the infected cells.
Tumor Cells Infected with AdNIS Efficiently Accumulate Io-dide. To further characterize the AdNIS virus,in vitro iodide uptakeexperiments were carried out on AdNIS-infected SiHa cells. Cellswere infected with CsCl-purified AdNIS for 28–30 h at various MOIs.Iodide uptake was then measured by incubating the cells for 15 minwith bHBSS containing 0.1mCi of 125I. At a MOI of 10, cellsaccumulated 140 times more iodide than noninfected control cells(Fig. 2A). Surprisingly, at a higher MOI, a lower amount of iodide wastaken up (Fig. 2A); possible explanations include a direct cytotoxicityof the adenoviral vector or a toxic effect of NIS overexpression in thecell membranes. In addition, cells infected with a control adenovirus(Adbgal) did not accumulate more iodide than noninfected cells (datanot shown), demonstrating that iodide uptake was specifically linked
Fig. 1. The AdNIS vector is functionalin vitro. A andB, detection of the NIS protein by immunofluorescenceon AdNIS-infected SiHa cells (A) and noninfected, con-trol SiHa cells (B). Twenty-four h after infection, thepresence of the NIS protein was detected by immuno-fluorescence using an anti-NIS antibody directed againstthe COOH-terminal end of the protein. Note the mem-brane localization of the NIS protein in some AdNIS-infected cells (arrows).
3486
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
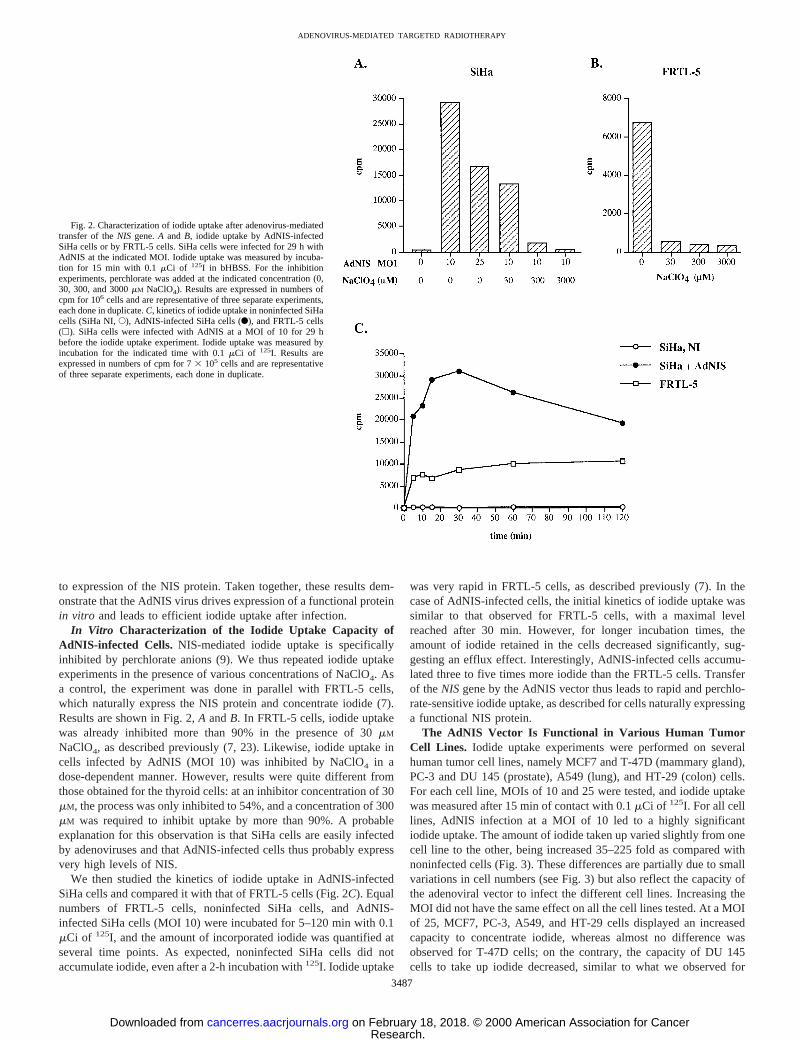
to expression of the NIS protein. Taken together, these results dem-onstrate that the AdNIS virus drives expression of a functional proteinin vitro and leads to efficient iodide uptake after infection.
In Vitro Characterization of the Iodide Uptake Capacity ofAdNIS-infected Cells. NIS-mediated iodide uptake is specificallyinhibited by perchlorate anions (9). We thus repeated iodide uptakeexperiments in the presence of various concentrations of NaClO4. Asa control, the experiment was done in parallel with FRTL-5 cells,which naturally express the NIS protein and concentrate iodide (7).Results are shown in Fig. 2,A andB. In FRTL-5 cells, iodide uptakewas already inhibited more than 90% in the presence of 30mM
NaClO4, as described previously (7, 23). Likewise, iodide uptake incells infected by AdNIS (MOI 10) was inhibited by NaClO4 in adose-dependent manner. However, results were quite different fromthose obtained for the thyroid cells: at an inhibitor concentration of 30mM, the process was only inhibited to 54%, and a concentration of 300mM was required to inhibit uptake by more than 90%. A probableexplanation for this observation is that SiHa cells are easily infectedby adenoviruses and that AdNIS-infected cells thus probably expressvery high levels of NIS.
We then studied the kinetics of iodide uptake in AdNIS-infectedSiHa cells and compared it with that of FRTL-5 cells (Fig. 2C). Equalnumbers of FRTL-5 cells, noninfected SiHa cells, and AdNIS-infected SiHa cells (MOI 10) were incubated for 5–120 min with 0.1mCi of 125I, and the amount of incorporated iodide was quantified atseveral time points. As expected, noninfected SiHa cells did notaccumulate iodide, even after a 2-h incubation with125I. Iodide uptake
was very rapid in FRTL-5 cells, as described previously (7). In thecase of AdNIS-infected cells, the initial kinetics of iodide uptake wassimilar to that observed for FRTL-5 cells, with a maximal levelreached after 30 min. However, for longer incubation times, theamount of iodide retained in the cells decreased significantly, sug-gesting an efflux effect. Interestingly, AdNIS-infected cells accumu-lated three to five times more iodide than the FRTL-5 cells. Transferof theNISgene by the AdNIS vector thus leads to rapid and perchlo-rate-sensitive iodide uptake, as described for cells naturally expressinga functional NIS protein.
The AdNIS Vector Is Functional in Various Human TumorCell Lines. Iodide uptake experiments were performed on severalhuman tumor cell lines, namely MCF7 and T-47D (mammary gland),PC-3 and DU 145 (prostate), A549 (lung), and HT-29 (colon) cells.For each cell line, MOIs of 10 and 25 were tested, and iodide uptakewas measured after 15 min of contact with 0.1mCi of 125I. For all celllines, AdNIS infection at a MOI of 10 led to a highly significantiodide uptake. The amount of iodide taken up varied slightly from onecell line to the other, being increased 35–225 fold as compared withnoninfected cells (Fig. 3). These differences are partially due to smallvariations in cell numbers (see Fig. 3) but also reflect the capacity ofthe adenoviral vector to infect the different cell lines. Increasing theMOI did not have the same effect on all the cell lines tested. At a MOIof 25, MCF7, PC-3, A549, and HT-29 cells displayed an increasedcapacity to concentrate iodide, whereas almost no difference wasobserved for T-47D cells; on the contrary, the capacity of DU 145cells to take up iodide decreased, similar to what we observed for
Fig. 2. Characterization of iodide uptake after adenovirus-mediatedtransfer of theNIS gene.A and B, iodide uptake by AdNIS-infectedSiHa cells or by FRTL-5 cells. SiHa cells were infected for 29 h withAdNIS at the indicated MOI. Iodide uptake was measured by incuba-tion for 15 min with 0.1mCi of 125I in bHBSS. For the inhibitionexperiments, perchlorate was added at the indicated concentration (0,30, 300, and 3000mM NaClO4). Results are expressed in numbers ofcpm for 106 cells and are representative of three separate experiments,each done in duplicate.C, kinetics of iodide uptake in noninfected SiHacells (SiHa NI,E), AdNIS-infected SiHa cells (F), and FRTL-5 cells(h). SiHa cells were infected with AdNIS at a MOI of 10 for 29 hbefore the iodide uptake experiment. Iodide uptake was measured byincubation for the indicated time with 0.1mCi of 125I. Results areexpressed in numbers of cpm for 73 105 cells and are representativeof three separate experiments, each done in duplicate.
3487
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

SiHa cells (see Figs. 2 and 3). These differences probably reflect themore or less pronounced cytotoxic effect of the adenoviral vector onthe various cell lines tested.
To assess the specificity of the process, we also measured iodideuptake in each cell line in the presence of NaClO4. As shown in Fig.3, iodide uptake was again inhibited in a dose-dependent manner byperchlorate anions. However, in contrast to what was observed withAdNIS-infected SiHa cells, a concentration of 30mM was sufficient toobtain an inhibition of more than 95%. This difference again reflectsthe capacity of the viral vector to infect the different cell lines, whichis at least 10–20 times higher for SiHa cells than for the other celllines.4 Taken together, these results demonstrate that AdNIS is func-tional in various tumor cell lines, whatever their origin.
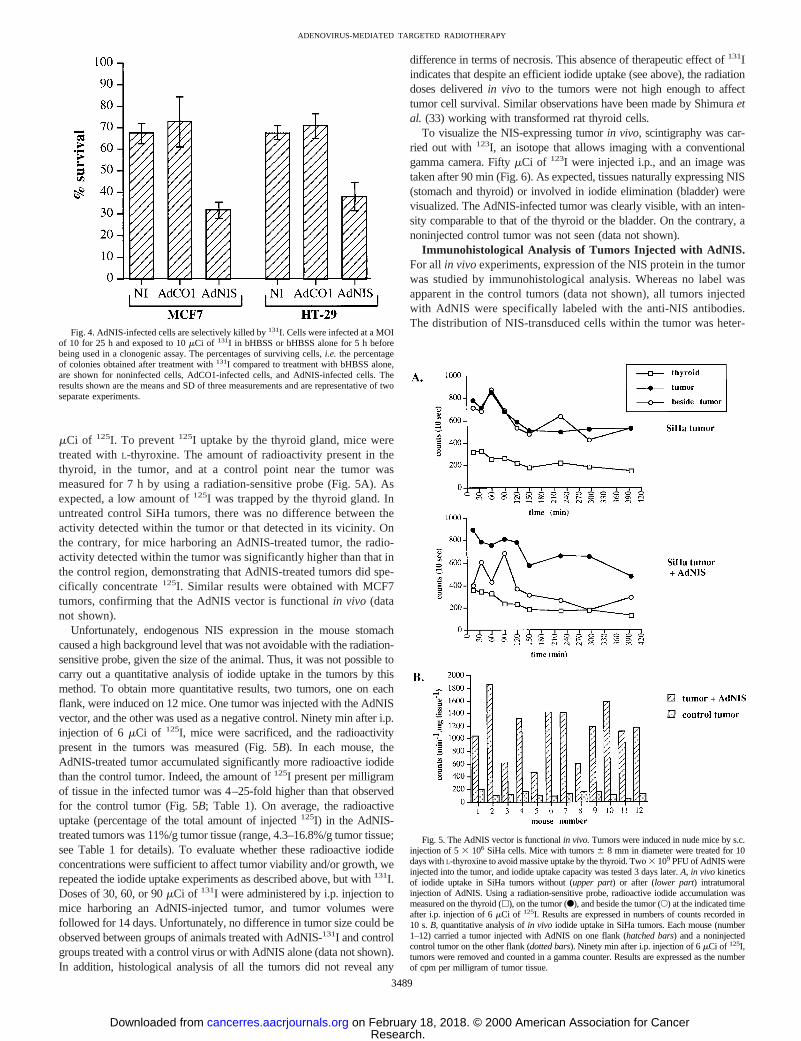
AdNIS-infected Human Tumor Cells Are Efficiently Killed by131I. In vitro 131I uptake experiments were performed on AdNIS-infected MCF7 and HT-29 cells to demonstrate that it was possible toobtain cell killing with the AdNIS-radioactive iodide system. Nonin-fected cells and cells infected with an adenoviral vector encoding no
transgene (AdCO1) were treated similarly, as controls. After131Itreatment, clonogenic assays were performed, and results are shown inFig. 4 and expressed as the percentage of surviving cells. In each case,for cells treated with bHBSS only, the numbers of colonies werecomparable, indicating that infection by an adenovirus (AdNIS orAdCO1) did not affect cell survival. On exposure to131I, around 30%of the cells were nonspecifically killed, as assessed by the resultsobtained for noninfected and AdCO1-infected cells. However, thenumber of colonies recovered from AdNIS-infected cells was signif-icantly lower than that from noninfected or AdCO1-infected cells,showing a selective killing effect of131I on NIS-expressing cells.These results thus demonstrate that coupling AdNIS and131I treat-mentsin vitro efficiently and specifically leads to cell killing, whichis the end goal of the system.
The AdNIS Vector Leads to Efficient Iodide Uptake in Vivo. Toevaluate the efficiency of the AdNIS vectorin vivo, SiHa tumors wereinduced on the right flank of nude mice by s.c. injection of 53 106
cells. The virus (23 109 PFU) was injected directly into the tumorswhen they had reached a diameter of 5–8 mm, and125I uptakeexperiments were carried out 3 days after by i.p. administration of 64 A. Boland, unpublished observations.
Fig. 3. The AdNIS vector leads to efficient iodide uptake in various cell lines. The iodide uptake capacity of MCF7 cells (mammary gland), DU 145 cells (prostate), A549 cells(lung), T-47D cells (mammary gland), PC-3 cells (prostate), and HT-29 cells (colon) was tested 29 h after infection with AdNIS at the indicated MOI. Iodide uptake was measuredby incubation for 15 min with 0.1mCi of 125I in bHBSS in the absence or presence of NaClO4 at the indicated concentration. Results are expressed in numbers of cpm and arerepresentative of at least two separate experiments, each done in duplicate. Cells numbers per well were as follows: MCF7 cells, 7.83 105; DU 145 cells, 6.53 105; A549 cells,5.5 3 105; T-47D cells, 6.43 105; PC-3 cells, 6.93 105; HT-29 cells, 13 106. NaClO4, sodium perchlorate concentration (mM); 2, no inhibitor.
3488
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
mCi of 125I. To prevent125I uptake by the thyroid gland, mice weretreated withL-thyroxine. The amount of radioactivity present in thethyroid, in the tumor, and at a control point near the tumor wasmeasured for 7 h by using a radiation-sensitive probe (Fig. 5A). Asexpected, a low amount of125I was trapped by the thyroid gland. Inuntreated control SiHa tumors, there was no difference between theactivity detected within the tumor or that detected in its vicinity. Onthe contrary, for mice harboring an AdNIS-treated tumor, the radio-activity detected within the tumor was significantly higher than that inthe control region, demonstrating that AdNIS-treated tumors did spe-cifically concentrate125I. Similar results were obtained with MCF7tumors, confirming that the AdNIS vector is functionalin vivo (datanot shown).
Unfortunately, endogenous NIS expression in the mouse stomachcaused a high background level that was not avoidable with the radiation-sensitive probe, given the size of the animal. Thus, it was not possible tocarry out a quantitative analysis of iodide uptake in the tumors by thismethod. To obtain more quantitative results, two tumors, one on eachflank, were induced on 12 mice. One tumor was injected with the AdNISvector, and the other was used as a negative control. Ninety min after i.p.injection of 6 mCi of 125I, mice were sacrificed, and the radioactivitypresent in the tumors was measured (Fig. 5B). In each mouse, theAdNIS-treated tumor accumulated significantly more radioactive iodidethan the control tumor. Indeed, the amount of125I present per milligramof tissue in the infected tumor was 4–25-fold higher than that observedfor the control tumor (Fig. 5B; Table 1). On average, the radioactiveuptake (percentage of the total amount of injected125I) in the AdNIS-treated tumors was 11%/g tumor tissue (range, 4.3–16.8%/g tumor tissue;see Table 1 for details). To evaluate whether these radioactive iodideconcentrations were sufficient to affect tumor viability and/or growth, werepeated the iodide uptake experiments as described above, but with131I.Doses of 30, 60, or 90mCi of 131I were administered by i.p. injection tomice harboring an AdNIS-injected tumor, and tumor volumes werefollowed for 14 days. Unfortunately, no difference in tumor size could beobserved between groups of animals treated with AdNIS-131I and controlgroups treated with a control virus or with AdNIS alone (data not shown).In addition, histological analysis of all the tumors did not reveal any
difference in terms of necrosis. This absence of therapeutic effect of131Iindicates that despite an efficient iodide uptake (see above), the radiationdoses deliveredin vivo to the tumors were not high enough to affecttumor cell survival. Similar observations have been made by Shimuraetal. (33) working with transformed rat thyroid cells.
To visualize the NIS-expressing tumorin vivo, scintigraphy was car-ried out with 123I, an isotope that allows imaging with a conventionalgamma camera. FiftymCi of 123I were injected i.p., and an image wastaken after 90 min (Fig. 6). As expected, tissues naturally expressing NIS(stomach and thyroid) or involved in iodide elimination (bladder) werevisualized. The AdNIS-infected tumor was clearly visible, with an inten-sity comparable to that of the thyroid or the bladder. On the contrary, anoninjected control tumor was not seen (data not shown).
Immunohistological Analysis of Tumors Injected with AdNIS.For all in vivoexperiments, expression of the NIS protein in the tumorwas studied by immunohistological analysis. Whereas no label wasapparent in the control tumors (data not shown), all tumors injectedwith AdNIS were specifically labeled with the anti-NIS antibodies.The distribution of NIS-transduced cells within the tumor was heter-
Fig. 4. AdNIS-infected cells are selectively killed by131I. Cells were infected at a MOIof 10 for 25 h and exposed to 10mCi of 131I in bHBSS or bHBSS alone for 5 h beforebeing used in a clonogenic assay. The percentages of surviving cells,i.e. the percentageof colonies obtained after treatment with131I compared to treatment with bHBSS alone,are shown for noninfected cells, AdCO1-infected cells, and AdNIS-infected cells. Theresults shown are the means and SD of three measurements and are representative of twoseparate experiments.
Fig. 5. The AdNIS vector is functionalin vivo.Tumors were induced in nude mice by s.c.injection of 53 106 SiHa cells. Mice with tumors6 8 mm in diameter were treated for 10days withL-thyroxine to avoid massive uptake by the thyroid. Two3 109 PFU of AdNIS wereinjected into the tumor, and iodide uptake capacity was tested 3 days later.A, in vivo kineticsof iodide uptake in SiHa tumors without (upper part) or after (lower part) intratumoralinjection of AdNIS. Using a radiation-sensitive probe, radioactive iodide accumulation wasmeasured on the thyroid (h), on the tumor (F), and beside the tumor (E) at the indicated timeafter i.p. injection of 6mCi of 125I. Results are expressed in numbers of counts recorded in10 s.B, quantitative analysis ofin vivo iodide uptake in SiHa tumors. Each mouse (number1–12) carried a tumor injected with AdNIS on one flank (hatched bars) and a noninjectedcontrol tumor on the other flank (dotted bars). Ninety min after i.p. injection of 6mCi of 125I,tumors were removed and counted in a gamma counter. Results are expressed as the numberof cpm per milligram of tumor tissue.
3489
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

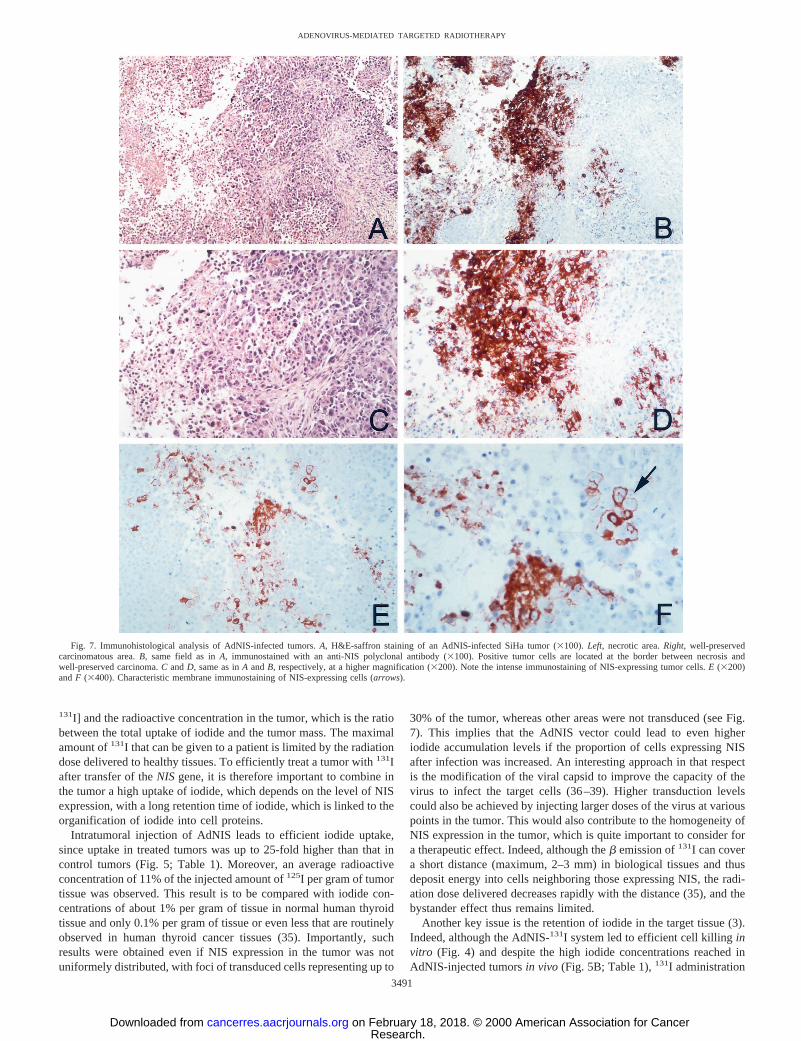
ogeneous, with foci of NIS-expressing cells representing up to 30% ofthe surface of the viable tumor tissue. Representative tumor sectionsare shown in Fig. 7. Interestingly, these foci were generally located atthe border between necrotic and viable regions of the tumor (see Fig.7, A–D). A possible explanation for this observation is that theadenoviral preparation injected could diffuse more easily at the con-tact of softer tissue (necrosis) and less so in viable areas. An alterna-tive hypothesis is that necrosis could be linked to a direct cytotoxiceffect of AdNIS infection. In some immunostained areas, it was clearthat labeling was confined to the cell membranes, demonstrating thatin vivo also, the AdNIS vector leads to correct localization of the NISprotein (Fig. 7,E andF).
DISCUSSION
The recent cloning of theNISgene is not only of major importancefor the understanding of the mechanisms underlying iodide transportin the thyroid but could also open new therapeutic perspectives.Indeed, achievement of efficient transfer of theNIS gene, coupled toradioactive iodide administration, may allow radioiodine treatment ofnonthyroid tumors as well as thyroid tumors presenting a defect intheir capacity to trap iodide.
In this study, we investigated the possibility of using an adenoviralvector to deliver theNIS gene into tumors, with the aim of treatingthem with radioiodine. We constructed a recombinant adenovirus,AdNIS, driving expression of the ratNISgene under the control of thecytomegalovirus promoter.In vitro infection of human tumor cellsfrom various origins, including mammary gland, prostate, lung, cer-vix, and colon, with AdNIS led to expression of a functional NISprotein, as revealed by efficient iodide uptake after infection. Iodideaccumulation in AdNIS-infected cells was rapid and perchlorate sen-sitive, two characteristics previously reported for NIS-mediated iodidetransport (9). Depending on the cell line tested, iodide accumulationwas 35–225-fold higher than that in noninfected control cells. Inaddition, AdNIS-infected cells were efficiently killed by131I, asrevealed by clonogenic assays. The AdNIS vector was also functionalin vivo, as revealed by the iodide accumulation observedin vivo intumors injected with AdNIS. Taken together, our results demonstratethat using the AdNIS vector is a valid approach to achieve efficientiodide uptake in tumors of nonthyroid origin.
While this study was in progress, transfer of theNIS gene with aretroviral vector was described in human [A375 (melanoma) andIGROV (ovarian carcinoma)] and mouse [CT26 (colon carcinoma)
BNL.1 ME (transformed liver)] tumor cells (23). In the cell linestested, iodide uptake reached to a maximum 21-fold that observedwith nontransduced cells and 35-fold that of cells transduced with acontrol retroviral vector (23). Delivery of theNIS gene by an adeno-viral vector is likely to be more efficient because the iodide uptakecapacity of AdNIS-infected cells was increased up to 225-fold ascompared with that of noninfected cells. Another major differencebetween both studies concerns the results obtainedin vivo. WhereasMandell et al. (23) induced tumors by injection of tumor cells firsttransducedin vitro with the NIS gene, we preferred to inject theAdNIS virus in established tumors, an approach required in a thera-peutic situation. In that respect, adenoviral vectors present a clearadvantage over retroviral vectors, namely, the ability to obtain con-centrated preparations, which facilitates efficientin vivo gene deliv-ery. Our results thus demonstrate for the first time that it is possibleto transfer theNISgene in a preformed tumor and to thereby confer asignificant and relevant iodide uptake capacityin vivo.
The therapeutic efficacy of radioiodine is dependent on the radia-tion dose delivered to the target tissue (34). The delivered radiationdose is proportional to both the effective half-life of131I in the tumor[combination of the physical (8.02 days) and biological half-lives of
Fig. 6. 123I imaging after intratumoral injection of AdNIS. Tumors were induced innude mice by s.c. injection of 53 106 SiHa cells. Mice with tumors6 8 mm in diameterwere treated for 10 days withL-thyroxine to avoid massive uptake by the thyroid.Two 3 109 PFU of AdNIS were injected in the tumor, and scintigraphy was performed3 days later using a gamma camera, 90 min after i.p. injection of 50mCi of 123I.
Table 1 Iodide concentrations in tumors after AdNIS treatment
Each mouse carried two tumors, one injected with 23 109 PFU of AdNIS and oneuntreated tumor used as a negative control. Ninety min after i.p. injection of 6mCi of 125I,tumors were removed, their weight was determined, and the radioactivity present in eachsample was counted. For each mouse, results were expressed as the percentage of the totalamount of 125I injected that was recovered per gram of tumor tissue (radioactiveconcentration). The last column represents the AdNIS-treated tumor:control tumor ratio.
Mouse no.
% of total 125I/g tissue
RatioAdNIS-treated tumor Control tumor
1 9.53 1.75 5.42 16.81 0.88 19.13 5.56 1.08 5.14 11.93 1.51 7.95 4.26 1.02 4.26 12.91 0.92 14.07 12.76 1.16 11.08 5.53 1.42 3.99 10.75 1.45 7.4
10 14.52 0.85 17.111 10.10 0.41 24.612 10.91 1.17 9.3
Mean6 SD 10.46 6 3.79 1.14 6 0.36 10.8 6 6.6
3490
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from
131I] and the radioactive concentration in the tumor, which is the ratiobetween the total uptake of iodide and the tumor mass. The maximalamount of131I that can be given to a patient is limited by the radiationdose delivered to healthy tissues. To efficiently treat a tumor with131Iafter transfer of theNIS gene, it is therefore important to combine inthe tumor a high uptake of iodide, which depends on the level of NISexpression, with a long retention time of iodide, which is linked to theorganification of iodide into cell proteins.
Intratumoral injection of AdNIS leads to efficient iodide uptake,since uptake in treated tumors was up to 25-fold higher than that incontrol tumors (Fig. 5; Table 1). Moreover, an average radioactiveconcentration of 11% of the injected amount of125I per gram of tumortissue was observed. This result is to be compared with iodide con-centrations of about 1% per gram of tissue in normal human thyroidtissue and only 0.1% per gram of tissue or even less that are routinelyobserved in human thyroid cancer tissues (35). Importantly, suchresults were obtained even if NIS expression in the tumor was notuniformely distributed, with foci of transduced cells representing up to
30% of the tumor, whereas other areas were not transduced (see Fig.7). This implies that the AdNIS vector could lead to even higheriodide accumulation levels if the proportion of cells expressing NISafter infection was increased. An interesting approach in that respectis the modification of the viral capsid to improve the capacity of thevirus to infect the target cells (36–39). Higher transduction levelscould also be achieved by injecting larger doses of the virus at variouspoints in the tumor. This would also contribute to the homogeneity ofNIS expression in the tumor, which is quite important to consider fora therapeutic effect. Indeed, although theb emission of131I can covera short distance (maximum, 2–3 mm) in biological tissues and thusdeposit energy into cells neighboring those expressing NIS, the radi-ation dose delivered decreases rapidly with the distance (35), and thebystander effect thus remains limited.
Another key issue is the retention of iodide in the target tissue (3).Indeed, although the AdNIS-131I system led to efficient cell killinginvitro (Fig. 4) and despite the high iodide concentrations reached inAdNIS-injected tumorsin vivo (Fig. 5B; Table 1),131I administration
Fig. 7. Immunohistological analysis of AdNIS-infected tumors.A, H&E-saffron staining of an AdNIS-infected SiHa tumor (3100).Left, necrotic area.Right, well-preservedcarcinomatous area.B, same field as inA, immunostained with an anti-NIS polyclonal antibody (3100). Positive tumor cells are located at the border between necrosis andwell-preserved carcinoma.C andD, same as inA andB, respectively, at a higher magnification (3200). Note the intense immunostaining of NIS-expressing tumor cells.E (3200)andF (3400). Characteristic membrane immunostaining of NIS-expressing cells (arrows).
3491
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

did not have any effect on tumor viability and growth in our study.This observation is most probably linked to the fact that the tumorsdid not retain iodide for a time period long enough to allow deliveryof a radiation dose affecting cell viability.In vitro data confirmed thatthe tumor cells used in this study do not have the capacity to organifyand thus retain the iodide taken up after AdNIS treatment (data notshown). This was similarly illustrated in a study of tumors consistingof malignantly transformed rat thyroid cells that had lost their capacityto concentrate iodide and that were transfectedin vitro with the ratNIS gene (33). Although the tumors efficiently concentrated iodide,no effect of 131I on tumor growth was observed because the rapidiodide efflux from the tumor did not allow the delivery of a radiationdose sufficient to inhibit cell growth (33). Several advances areconceivable to circumvent this lack of iodide retention in the tumors.First, the efficiency ofNIS gene transfer–and thus the iodide uptakecapacity of the target tissue–may be improved by the use of modifiedvectors and/or higher viral doses (see above). Second, the biologicalhalf-life of radioiodine in the tumor tissues could be increased bycoupling transfer of theNISgene with delivery of a gene involved inthe iodide organification process, such as the thyroperoxidase (1).
In view of the obtained results, coupling transfer of theNISgene byan adenoviral vector and radioiodine administration appears to be avery promising strategy for treating tumors of various origins. Al-though improvements to achieve higher radiation doses in the targettissue will be required, application of this “targeted radiotherapy”approach in patients will be facilitated by our long-standing experi-ence with radioiodine for thyroid cancer therapy.
ACKNOWLEDGMENTS
We thank Laure Franqueville and Stephanie Esselin for excellent technicalassistance and Elisabeth Connault for the immunohistological work. We arevery grateful to all of the staff of the animal facilities of the Institut GustaveRoussy for their help during thein vivo experiments.
REFERENCES
1. Carrasco, N. Iodide transport in the thyroid gland. Biochim. Biophys. Acta,1154:65–82, 1993.
2. Kaplan, M. M., Meier, D. A., and Dworkin, H. J. Treatment of hyperthyroidism withradioactive iodine. Endocrinol. Metab. Clin. N. Am.,27: 205–223, 1998.
3. Schlumberger, M. Papillary and follicular thyroid carcinoma. N. Engl. J. Med.,338:297–306, 1998.
4. Thomas-Morvan, C., Carayon, P., Schlumberger, M., Vignal, A., and Tubiana, M.Thyrotrophin stimulation of adenylate cyclase and iodine uptake in human differen-tiated thyroid cancer. Acta Endocrinol. (Copenh.),101: 25–31, 1982.
5. Smanik, P. A., Ryu, K-Y., Theil, K. S., Mazzaferri, E. L., and Jhiang, S. M.Expression, exon-intron organization, and chromosome mapping of the human so-dium iodide symporter. Endocrinology,138: 3555–3558, 1997.
6. Arturi, F., Russo, D., Schlumberger, M., du Villard, J-A., Caillou, B., Vigneri, P., Wicker,R., Chiefari, E., Suarez, H. G., and Filetti, S. Iodide symporter gene expression in humanthyroid tumors. J. Clin. Endocrinol. Metab.,83: 2493–2496, 1998.
7. Weiss, S. J., Philp, N. J., and Grollman, E. F. Iodide transport in a continuous line ofcultured cells from rat thyroid. Endocrinology,114: 1090–1098, 1984.
8. Eskandari, S., Loo, D. D. F., Dai, G., Levy, O., Wright, E. M., and Carrasco, N.Thyroid Na1/I2 symporter. Mechanism, stoichiometry, and specificity. J. Biol.Chem.,272: 27230–27238, 1997.
9. Dai, G., Levy, O., and Carrasco, N. Cloning and characterization of the thyroid iodidesymporter. Nature (Lond.),379: 458–460, 1996.
10. Smanik, P. A., Liu, Q., Furminger, T. L., Ryu, K., Xing, S., Mazzaferri, E. L., andJhiang, S. M. Cloning of the human sodium iodide symporter. Biochem. Biophys.Res. Commun.,226: 339–345, 1996.
11. Levy, O., Dai, G., Riedel, C., Ginter, C. S., Paul, E. M., Lebowitz, A. N., andCarrasco, N. Characterization of the thyroid Na1/I2 symporter with an anti-COOHterminus antibody. Proc. Natl. Acad. Sci. USA,94: 5568–5573, 1997.
12. Levy, O., De la Vieja, A., Ginter, C. S., Riedel, C., Dai, G., and Carrasco, N. N-linkedglycosylation of the thyroid Na1/I2 symporter (NIS). Implications for its secondarystructure model. J. Biol. Chem.,273: 22657–22663, 1998.
13. Spitzweg, C., Joba, W., Eisenmenger, W., and Heufelder, A. E. Analysis of humansodium iodide symporter gene expression in extrathyroidal tissues and cloning of itscomplementary deoxyribonucleic acids from salivary gland, mammary gland, andgastric mucosa. J. Clin. Endocrinol. Metab.,83: 1746–1751, 1998.
14. Fujiwara, H., Tatsumi, K., Miki, K., Harada, T., Miyai, K., Takai, S-I., and Amino,N. Congenital hypothyroidism caused by a mutation in the Na1/I2 symporter. Nat.Genet.,16: 124–125, 1997.
15. Pohlenz, J., Rosenthal, I. M., Weiss, R. E., Jhiang, S. M., Burant, C., and Refetoff, S.Congenital hypothyroidism due to mutations in the sodium/iodide symporter. Iden-tification of a nonsense mutation producing a downstream cryptic 39splice site.J. Clin. Invest.,101: 1028–1035, 1998.
16. Lazar, V., Bidart, J-M., Caillou, B., Mahe, C., Lacroix, L., Filetti, S., andSchlumberger, M. Expression of the Na1/I2 symporter gene in human thyroidtumors: a comparison study with other thyroid-specific genes. J. Clin. Endocrinol.Metab.,84: 3228–3234, 1999.
17. Caillou, B., Troalen, F., Baudin, E., Talbot, M., Filetti, S., Schlumberger, M., andBidart, J-M. Na1/I2 symporter distribution in human thyroid tissues: an immunohis-tochemical study. J. Clin. Endocrinol. Metab.,83: 4102–4106, 1998.
18. Knopp, J., Stolc, V., and Tong, W. Evidence for the induction of iodide transport inbovine thyroid cells treated with thyroid-stimulating hormone or dibutyryl cyclicadenosine 39,59-monophosphate. J. Biol. Chem.,245: 4403–4408, 1970.
19. Fayet, G., and Hovsepian, S. Active transport of iodide in isolated porcine thyroidcells. Application to anin vitro bioassay of thyrotropin. Mol. Cell. Endocrinol.,7:67–78, 1977.
20. Dickson, J. G., Hovsepian, S., Fayet, G., and Lissitzky, S. Follicle formation and iodidemetabolism in cultures of human thyroid cells. J. Endocrinol.,90: 113–124, 1981.
21. Kosugi, S., Sasaki, N., Hai, N., Sugawa, H., Aoki, N., Shigemasa, C., Mori, T., andYoshida, A. Establishment and characterization of a Chinese hamster ovary cell line,CHO-4J, stably expressing a number of Na1/I2 symporters. Biochem. Biophys. Res.Commun.,227: 94–101, 1996.
22. Spitzweg, C., Zhang, S., Bergert, E. R., Castro, M. R., McIver, B., Heufelder, A. E.,Tindall, D. J., Young, C. Y. F., and Morris, J. C. Prostate-specific antigen (PSA)promoter-driven androgen-inducible expression of sodium iodide symporter in pros-tate cancer cell lines. Cancer Res.,59: 2136–2141, 1999.
23. Mandell, R. B., Mandell, L. Z., and Link, C. J. J. Radioisotope concentrator genetherapy using the sodium/iodide symporter gene. Cancer Res.,59: 661–668, 1999.
24. Perricaudet, M., and Stratford-Perricaudet, L. D. Adenovirus-mediatedin vivo genetherapy.In: J-M. Vos (ed.), Human Viruses in Gene Therapy, pp. 1–32. Durham, NC:Carolina Academic Press, 1994.
25. Zhang, R., Straus, F. H., and DeGroot, L. J. Effective genetic therapy of establishedmedullary thyroid carcinomas with murine interleukin-2: dissemination and cytotox-icity studies in a rat tumor model. Endocrinology,140: 2152–2158, 1999.
26. Zhang, W-W. Development and application of adenoviral vectors for gene therapy ofcancer. Cancer Gene Ther.,6: 113–138, 1999.
27. Benihoud, K., Yeh, P., and Perricaudet, M. Adenovirus vectors for gene delivery.Curr. Opin. Biotechnol.,10: 440–447, 1999.
28. Crouzet, J., Naudin, L., Orsini, C., Vigne, E., Ferrero, L., Le Roux, A., Benoit, P.,Latta, M., Torrent, C., Branellec, D., Denefle, P., Mayaux, J. F., Perricaudet, M., andYeh, P. Recombinational construction inEscherichia coliof infectious adenoviralgenomes. Proc. Natl. Acad. Sci. USA,94: 1414–1419, 1997.
29. Griscelli, F., Li, H., Bennaceur-Griscelli, A., Soria, J., Opolon, P., Soria, C.,Perricaudet, M., Yeh, P., and Lu, H. Angiostatin gene transfer: inhibition of tumorgrowth in vivo by blockade of endothelial proliferation associated with a mitosisarrest. Proc. Natl. Acad. Sci. USA,95: 6367–6372, 1998.
30. Stratford-Perricaudet, L. D., Makeh, I., Perricaudet, M., and Briand, P. Widespreadlong-term gene transfer to mouse skeletal muscles and heart. J. Clin. Invest.,90:626–630, 1992.
31. Graham, F. L., Smiley, J., Russel, W. C., and Nairn, R. Characteristics of a human cellline transformed by DNA from human adenovirus type 5. J. Gen. Virol.,36:59–74, 1977.
32. Fallaux, F. J., Kranenburg, O., Cramer, S. J., Houweling, A., van Ormondt, H.,Hoeben, R. C., and Van Der Eb, A. J. Characterization of 911: a new helper cell linefor the titration and propagation of early region 1-deleted adenoviral vectors. Hum.Gene Ther.,7: 215–222, 1996.
33. Shimura, H., Haraguchi, K., Miyazaki, A., Endo, T., and Onaya, T. Iodide uptake andexperimental131I therapy in transplanted undifferentiated thyroid cancer cells ex-pressing the Na1/I2 symporter gene. Endocrinology,138: 4493–4496, 1997.
34. Maxon, H. R., Thomas, S. R., Hertzberg, V. S., Kereiakes, J. G., Chen, I. W., Sperling,M. I., and Saenger, E. L. Relation between effective radiation dose and outcome ofradioiodine therapy for thyroid cancer. N. Engl. J. Med.,309: 937–941, 1983.
35. Filetti, S., Bidart, J-M., Arturi, F., Caillou, B., Russo, D., and Schlumberger, M.Sodium/iodide symporter: a key transport system in thyroid cancer cell metabolism.Eur. J. Endocrinol.,141: 443–457, 1999.
36. Wickham, T. J., Roelvink, P. W., Brough, D. E., and Kovesdi, I. Adenovirus targetedto heparan-containing receptors increases its gene delivery efficiency to multiple celltypes. Nat. Biotechnol.,14: 1570–1573, 1996.
37. Wickham, T. J., Tzeng, E., Shears, L. L., Roelvink, P. W., Li, Y., Lee, G. M., Brough,D. E., Lizonova, A., and Kovesdi, I. Increasedin vitro and in vivo gene transfer byadenovirus vectors containing chimeric fiber proteins. J. Virol.,71: 8221–8229, 1997.
38. Haisma, H. J., Pinedo, H. M., van Rijswijk, A., van der Meulen-Muileman, I.,Sosnowski, B. A., Ying, W., van Beusechem, V. W., Tillman, B. W., Gerritsen,W. R., and Curiel, D. T. Tumor-specific gene transfer via an adenoviral vectortargeted to the pan-carcinoma antigen EpCAM. Gene Ther.,6: 1469–1474, 1999.
39. Vigne, E., Mahfouz, I., Dedieu, J-F., Brie, A., Perricaudet, M., and Yeh, P. RGDinclusion in the hexon monomer provides adenovirus type 5-based vectors with a fiberknob-independent pathway for infection. J. Virol.,73: 5156–5161, 1999.
3492
ADENOVIRUS-MEDIATED TARGETED RADIOTHERAPY
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from

2000;60:3484-3492. Cancer Res Anne Boland, Marcel Ricard, Paule Opolon, et al. Symporter Gene into Tumors for a Targeted RadiotherapyAdenovirus-mediated Transfer of the Thyroid Sodium/Iodide
Updated version
http://cancerres.aacrjournals.org/content/60/13/3484
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/60/13/3484.full#ref-list-1
This article cites 37 articles, 13 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/60/13/3484.full#related-urls
This article has been cited by 30 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/60/13/3484To request permission to re-use all or part of this article, use this link
Research. on February 18, 2018. © 2000 American Association for Cancercancerres.aacrjournals.org Downloaded from