adaptive dynamics studying the dynamic change of community dynamical parameters through mutation and...
TRANSCRIPT

Adaptive Dynamics
studying the dynamic change of community dynamical parameters
through mutation and selection
Hans (= J A J *) Metz
(formerly ADN) IIASA
QuickTime™ en eenTIFF (ongecomprimeerd)-decompressorzijn vereist om deze afbeelding weer te geven.
VEOLIA-Ecole Poly-technique
QuickTime™ en eenTIFF (ongecomprimeerd)-decompressor
zijn vereist om deze afbeelding weer te geven.
&Mathematical Institute, Leiden University
QuickTime™ and a decompressor
are needed to see this picture.

preamble: some terminology
micro-evolution: changes in gene frequencies on a population dynamical time scale,
meso-evolution: evolutionary changes in the values of traits of representative individuals and concomitant patterns of taxonomic diversification (through multiple mutant substitutions),
macro-evolution: changes where one cannot even speak in terms of a fixed set of traits, like anatomical innovations.
Goal: get a mathematical grip on meso-evolution.

functiontrajectories
formtrajectories
genome
development
selection
(darwinian)
(causal)
demography
physics
almost faithful reproduction
ecology
(causal)
fitness
environment
components of the evolutionary mechanism

fitness
functiontrajectories
formtrajectories
genome
development
selection
(darwinian)
(causal)
physics
almost faithful reproduction
ecology
(causal)environment
Adaptive Dynamics
demography

AD’s basis in Commmunity Dynamics
Populations are considered as measures over a space ot i(ndividual)-states (e.g. spanned by age and size).
Environments (E) are delimited such that given their environment individuals are independent,
and hence their mean numbers have linear dynamics.Resident populations are assumed to be so large that
we can approximate their dynamics deterministically.These resident populations influence the environment
so that they do not grow out of bounds.The resulting dynamical systems therefore have
attractors, which are assumed to produce ergodic environments.

AD’s basis in Commmunity Dynamics
Mutants enter the population singly.Therefore, initially their impact on the environment can
be neglected. The initial growth of a mutant population can be
approximated with a branching process. Invasion fitness is the (generalised) Malthusian parameter
(= averaged long term exponential growth rate of the mean) of this proces: (Existence guaranteed by the multiplicative ergodic theorem.)
Residents have fitness zero.

AD’s basis in Commmunity Dynamics
resident population size
population sizes of
other species
mutantpopulationsize
fitness as dominant transversal eigenvalue

resident population size
population sizes of
other species
mutantpopulationsize
or, more generally, dominant transversal Lyapunov exponent
AD’s basis in Commmunity Dynamics

Fitnesses are not given quantities, but depend on (1) the traits of the individuals, X, Y, (2) the environment in which they live:
(Y | E)The latter is set by the resident community:
E = Eattr(C), C={X1,...,Xk)
biological implications
Evolutionary progress is almost exclusively determined by the fitnesses of potential mutants.

AD: fitness landscapes change with evolution
Evolution proceeds through uphill movements in a fitness landscape that keeps changing so as to keep the fitness of the resident types at exactly zero.
Evolution proceeds through uphill movements in a fitness landscape
resident trait value(s) x
evol
utio
nary
tim
e
0
0
0
fitness landscape: (y,E(t))
mutant trait value y
0
0

type
morph
strategy
trait vector
point (in trait space)
type
morph
strategy
trait vector (trait value)
point (in trait space)
effective synonyms

The different spaces that play a role in adaptive dynamics:
the trait space in which their evolution takes place(= parameter space of their i- and therefore of their p-dynamics)
= the ‘state space’ of their adaptive dynamics
the physical space inhabited by the organisms
the state space of their i(ndividual)-dynamics
the space of the influences that they undergo(fluctuations in light, temperature, food, enemies, conspecifics):
their ‘environment’
the parameter spaces of families of adaptive dynamics
the state space of their p(opulation)-dynamics
scaling up from organisms to trait evolution

the simplifications underlying AD
essential formost conclusions
i.e., separated population dynamical and mutational time scales:the population dynamics relaxes before the next mutant comes
1. mutation limited evolution
2. clonal reproduction
3. good local mixing4. largish system sizes
5. “good” c(ommunity)-attractors6. interior c-attractors unique
7. fitness smooth in traits8. small mutational steps
essential conceptuallly
essential

from CD to AD: nature of the limits
x
adaptive dynamicslimit
individual-basedsimulation
classical largenumber limit
t
, rescale time, only consider traits
rescale numbers to densities
= system size, = mutations / birth
t

the mechanism of meso-evolution
C := {X1,..,Xk}: trait values of the residents
Environment: Eattr(C) Y: trait value of mutant
Fitness (rate of exponential growth in numbers) of mutant
sC(Y) := (Eattr(C),Y)
* Y has a positive probability to invade into a C community iff sC(Y) > 0.
* After invasion, Xi can be ousted by Y only if sX1,..,Y,.., Xk(Xi) ≤ 0.
* For small mutational steps Y takes over, except near so-called “ess”es.

time1
10
100
1000
10 200
# individuals
population dynamics: branching process results
or "grow exponentially” either go extinct, mutant populations starting from single individuals
In an a priori given ergodic environment:
(with a probability that to first order in | Y – X | is proportional
to their (fitness)+ , and with their fitness as rate parameter).

Invasion of a "good" c-attractor of X leads to a substitution such that this c-attractor is inherited by Y Y and up to O(2),
sY(X) = – sX(Y).
community dynamics: ousting the resident?
Proposition:
Let = | Y – X | be sufficiently small,
and let X not be close to an “evolutionarily singular strategy”, or to a c(ommunity)-dynamical bifurcation point.
“For small mutational steps invasion implies substitution.”

community dynamics: sketch of the proof
When an equilibrium point or a limit cycle is invaded, the relative frequency p of Y satisfies
= sX(Y) p(1-p) + O(2),
while the convergence of the dynamics of the total population densities occurs O(1).
dpdt
1
03 4 5 6 7
p
sX(Y) t
Singular strategies X* are defined by sX*(Y) = O(), instead of O().

Near where the mutant trait value y equals the resident trait value x there is a degenerate transcritical bifurcation:
community dynamics: the bifurcation structure
nx →
↑ny
n x→
↑ny
n x →
↑ny
y<xy=xy>x

community dynamics: the bifurcation structure
relativefrequency
of 2nd
'species'resident trait value x
0
1
mutant trait value y
resident trait value x0
1
mutant trait value y
evolution will be towards increasing x
evolution will be towards decreasing x
Near where the mutant trait value y equals the resident trait value x there is a degenerate transcritical bifurcation:
The effective speeds of evolutionary change are proportional to the probabilities that invading mutants survive the initial stochastic phase

population dynamics: the bifurcation structure
probability thatmutant
invades
evolution will be towards increasing x
evolution will be towards decreasing x
Near where the mutant equals the resident, the probability that the mutant invades changes as depicted below:
The effective speeds of evolutionary change are proportional to the probabilities that invading mutants survive the initial stochastic phase
resident trait value x0
1
mutant trait value y
resident trait value x0
1
mutant trait value y

+
+
-
y
x
-
fitness contour plotx: residenty: potential mutant
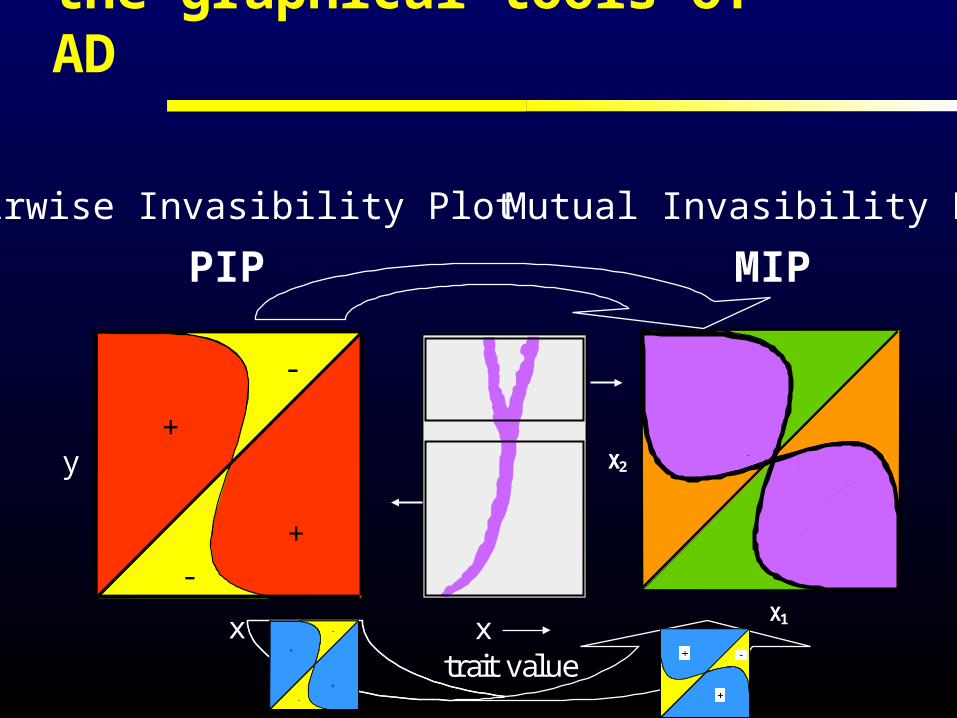
the graphical tools of AD
trait valuex
x0x1
x1
x2
x
X
X1
2
+
-
+
-
.
Mutual Invasibility Plot
MIP
y
xtrait value
X
x
the graphical tools of AD
+
+
-
-
Pairwise Invasibility Plot
PIP
y
x0
+
+
example 1 example 2
+
+
x0
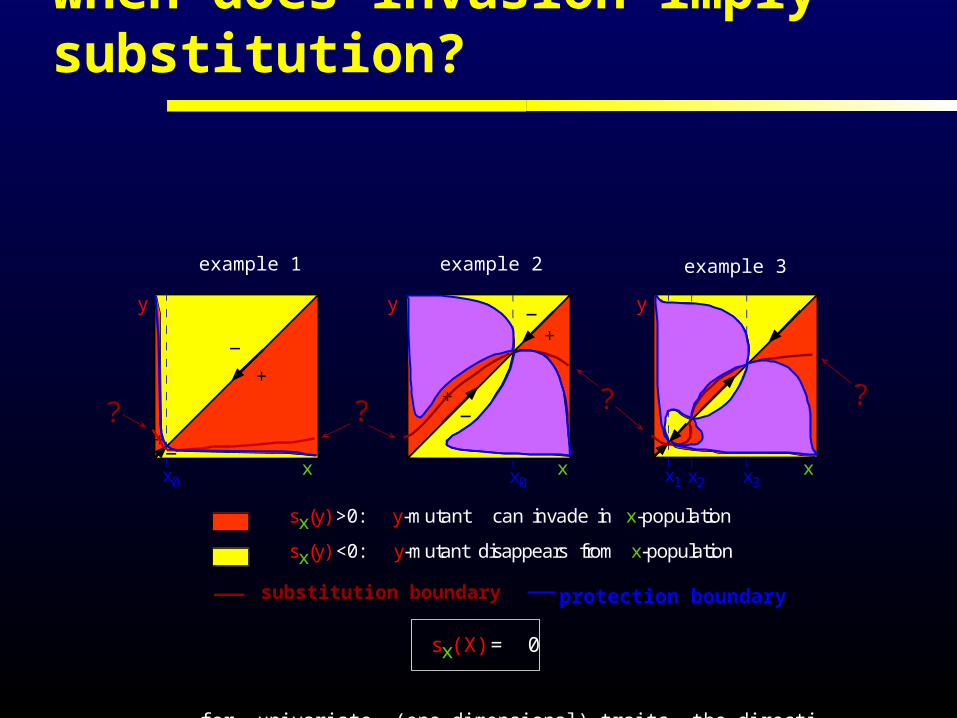
sx(y) >0: y-mutant can invade in x-population
sx(y) <0: y-mutant disappears from x-population
for univariate (one-dimensional) traits the direction of evolution is determined by sign sx(y)
sx(X) = 0
example 3
x x1 x2 x3
y y
x x
when does invasion imply substitution?
? ? ? ?
substitution boundary protection boundary

X2
X1
trait valuex
Trait Evolution Plot
TEP
x2
the graphical tools of AD
y
x
+
+
-
-
Pairwise Invasibility Plot
PIP

evolutionarily singular strategies
x*
x0
+
+
x*
x* is a singular point iff
dy
dsx(y) = 0y=x=x*
(x* is an extremum in the y-direction)
x*
x*
+
+

y
x* x
v=y-x*
u=x-x*
su(v) = a + b1u+b0v + c11u2+2c10uv+c00v2
+ h.o.t
b1=b0=0
a=0b1+b0=0c11+2c10+c00=0
neutrality of resident
x* is an extremum in y
s0(0)=0
su(u)=0su(v) = c11u2−(c11+c) +uv cv
+ . .h o t
(monomorphic) linearisation around y = x = x*
c11+2c10+c00=0
a=0b1+b0=0
neutrality of resident
su(u)= 0
b1=b0=0
x* is an extremum in y
s0(0)=0
su(v) = c11u2−(c11+c) +uv cv
+ . .h o t

monomorphicconvergenceto x0
yes
no
c00
noyes
noyes
c11c11
c00
x0 uninvadable
local PIP classification
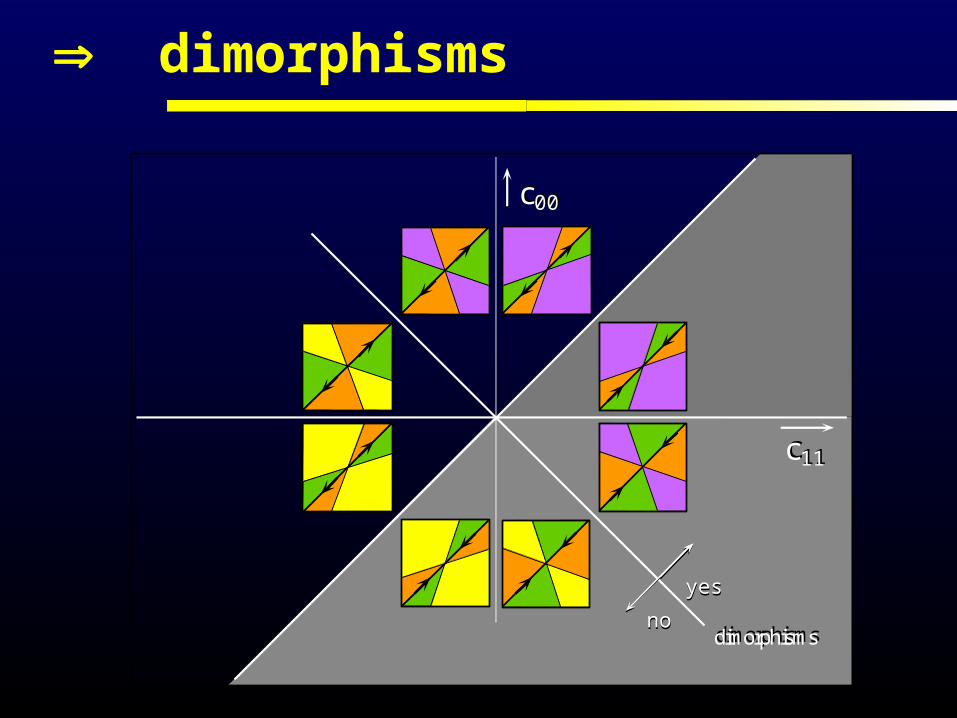
dimorphisms
c00
c11c11
c00
dimorphismsnono
yesyes
dimorphisms

dimorphic linearisation around y = x1 = x2 = x*
Local coordinates: v = y-x* mutant u1 = x1-x*, u2 = x2-x*
residents
Only directional derivatives (!):
u1 = uw1, u2 = uw2
Only directional derivatives (!)

n1 →
n1→^
↑
n2^
↑n2
A
B
B
A
community state space parameter space
parameter paths attractor paths
A
B
A
B
population dynamics: non-genericity strikes

dimorphic linearisation around y = x1 = x2 = x*
su ,u (v) = α+
β1(w1,w)u+β v +
γ11(w1,w)u+γ1(w1,w)uv+γV+
. . .h o t
1 (*)
Local coordinates: v = y-x* mutant u1 = x1-x*, u2 = x2-x*
residents
Only directional derivatives (!):
u1 = uw1, u2 = uw2
Only directional derivatives (!) :
u1=uw1, u2=uw2

dimorphic linearisation around y = x1 = x2 = x*
s00 (v) = s0(v)
su ,u (u1) = 0 =1 2
su ,u (u2)1 2
neutrality of residents
su ,u (v) =1 2
su ,u (v)2 1
symmetry
if u1=u2=0 we are back in themonomorphic resident case
su ,u (v) =1 2
expansion formula (*)
(v-u1) (v-u2) [c00+ h.o.t]

local types of dimorphic evolution
0
c00>0
V0
u1 u2
Su ,u (v)1 2
Su ,u (v) = (v-u1) (v-u2) [c00+ h.o.t]1 2
c00<0
Su ,u (v)1 2
V
u1 u2
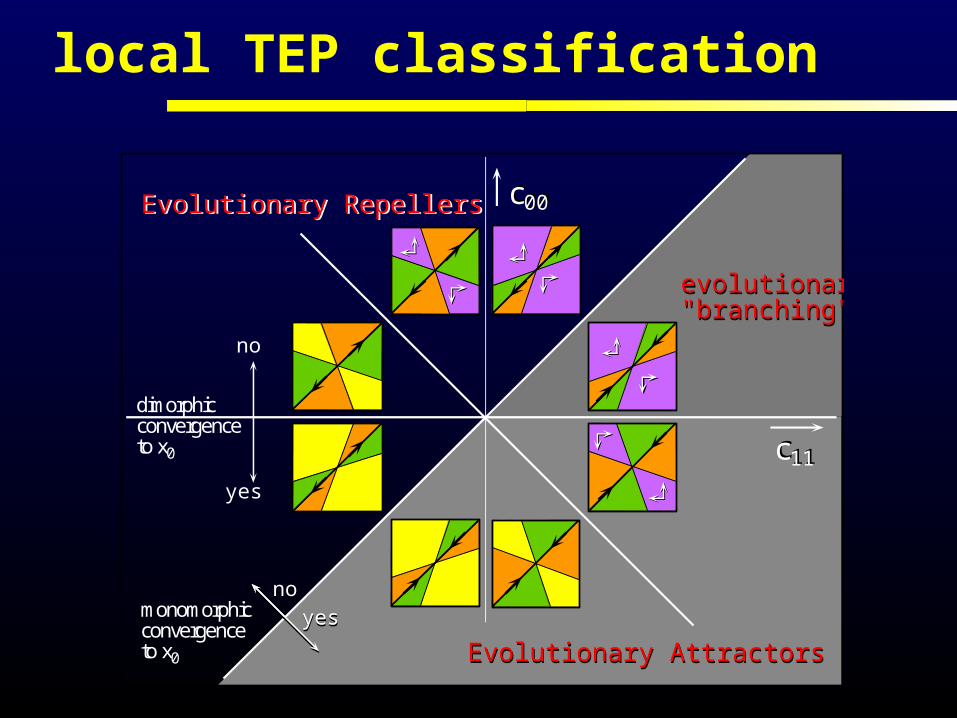
local TEP classification
monomorphicconvergenceto x0
dimorphicconvergenceto x0
yes
no
evolutionary"branching"evolutionary"branching"
Evolutionary AttractorsEvolutionary Attractors
Evolutionary RepellersEvolutionary Repellers c00
noyes
noyes
c11c11
c00

more about adaptive branching
t r a i t v a lu e
x
evol
utio
nary
tim
e
t i m e t r a
i t
fitne
ssfitness
minimum
population

. Summary
Ecological Character Simulation
beyond clonality: thwarting the Mendelian mixer
asso
rtativ
enes
s

a toy example
____ = 1 - Σa(xi,xj)nj/k(xi)dninidt j
k(x)=
a(xi,xj)=e-α(xi-xj) xi-xj→xi-xj→
↑a↑a
1/√α1/√α
Lotka-Volterra competition among individualsdifferentiated according to a one-dimensional trait x.
with
and
population equations:
1-x2 if -1<x<10 elsewhere{
↑k
-1 x → 1
↑k
-1 x → 1
Lotka-Volterra all per capita growth rates are linear functions of the population densities
Lotka-Volterra all per capita growth rates are linear functions of the population densities
LV models are unrealistic, but useful since they have explicit expressions for the invasion fitnesses.

a toy example
____ = 1 - Σa(xi,xj)nj/k(xi)dninidt j
k(x)=
a(xi,xj)=e-α(xi-xj) xi-xj→xi-xj→
↑a↑a
1/√α1/√α
Lotka-Volterra competition among individualsdifferentiated according to a one-dimensional trait x.
with
and
population equations:
1-x2 if -1<x<10 elsewhere{
↑k
-1 x → 1
↑k
-1 x → 1
viable range
competition kernelcompetition kernel
carrying capacity carrying capacity
widthwidth 1 ––––––––
√2α

matryoshka galore
x1
x
x2
Exploring parameter space
α=1/3: α=: α=3:
isoclines correspond to loci of monomorphic singular points.
interrupted: branching prone ( trimorphically repelling)

x1
x
x2
Exploring parameter space
α=1/3: α=: α=3:
of two lines about to merge one goes extinct

matryoshka galore polymorphisms are invariant under permutation of indices
X2
the six purple
volumesshould
be identified
!
adjacent purple volumes are mirror symmetric around a diagonal plane
X1
X3

matryoshka galore the sets of trimorphisms connect to the isoclines of the dimorphisms
isoclineof
species 1
isoclineof
species 3
X1
X2
X3
(x2 = x3) (x2 = x1)

more consistency conditions!
There also exist various global consistency relations!
The classification of the singular points was based on just a smoothness assumption and some ecologically reasonable consistency conditions.
x2
x1
y
x
+
-
+
-
is extinct. the coexistence set one type. Use that on the boundaries of

a more complicated example
.
Seed size 2
Seed size 1
seed size evolution: TEPs

a potential difficulty: heteroclinic loops
1 2
3
1 2
3
1 2
3
1 2
3
1 2 1 2
3 3
?

a potential difficulty: heteroclinic loops
4
3
1
2
4
23
1
?
The larger the number of types, the larger the fraction ofheteroclinic loops among the possible attractor structures !

much remains to be done!
(Many partial results are floating around.)
Classify the geometries of the fitness landscapes, and coexistence sets near singular points in higher dimensions.
Develop a fullfledged bifurcation theory for AD.
Develop (more) global geometrical results. Delineate to what extent, and in which manner, AD results stay intact
for Mendelian populations.Develop analogous theories for not fully smooth s-
functions.
Analyse how to deal with the heteroclinic loop problem.
(Some recent results by Odo and Barbara Boldin.)

The end