adalto lourenÇo - a diversidade das algas
TRANSCRIPT

CAPÍTULO 2
A DIVERSIDADE DAS ALGAS
As algas são organismos ubíquos em sistemas aquáticos sob incidência de luz solar.Sua importância na natureza refere-se principalmente à elevada participação no balanço globalda fotossíntese, contribuindo com grande parcela da produção primária do planeta. No mar,mais de 90% da fotossíntese é realizada pelas diversas microalgas que constituem o fito-plâncton. As microalgas são os principais produtores primários marinhos, portanto, sãofundamentais para a estruturação de quase todos os ecossistemas costeiros e oceânicos.Embora muitas funções sejam genericamente atribuídas às algas, elas constituem um conjuntoartificial e heterogêneo de organismos, os quais eventualmente exibem mais diferenças doque semelhanças entre si. Neste capítulo, discute-se brevemente a importância das algas nanatureza, sendo apresentadas as características gerais dos grandes grupos de algas.
2.1 CONCEITOS ELEMENTARES
Algas são seres muito diversos presentes em sistemas aquáticos (ou zonas úmidas), sãofotossintetizantes em sua grande maioria (neste caso, dotados de clorofila a) e apresentamestrutura vegetativa conhecida como talo, cuja diferenciação celular é caracteristicamentepequena ou nula. Assim, o termo “alga” é completamente desprovido de valor taxonômico,pois designa organismos muito distintos entre si quanto à origem, composição química emorfologia. Envolve seres unicelulares e multicelulares, com hábitos planctônicos e bentô-nicos. A partir dessas características derivam os termos de uso consagrado: microalgas (algascom dimensões microscópicas) e macroalgas (algas com dimensões macroscópicas). Enquantoas macroalgas são quase sempre bentônicas (exceto algumas formas que podem ser flutuantes),a maioria das microalgas tem hábito planctônico, embora haja também muitas espéciesbentônicas e terrestres (habitando ambientes úmidos). As microalgas planctônicas de dadolocal constituem o fitoplâncton, ou seja, são componentes fotoautotróficos do plâncton.Os diversos organismos fitoplanctônicos podem se apresentar como células isoladas, agru-padas formando colônias ou encadeadas sob a forma de segmentos lineares de células. Emtodos os casos, porém, ocorre pouca ou nenhuma diferenciação das funções ou especializaçãodas células, ou seja, cada célula realiza todas as funções vitais.
O fitoplâncton é uma das categorias do plâncton, que envolve também o zooplâncton(animais planctônicos, incluindo também o ictioplâncton, constituído por ovos e larvas de peixes),o bacterioplâncton (bactérias planctônicas), o micoplâncton (fungos planctônicos), o proto-zooplâncton (protistas heterotróficos planctônicos) e o virioplâncton (vírus presentes na colunad’água). Assim, o termo fitoplâncton se refere ao conjunto de algas microscópicas que vivemem suspensão na coluna d’água. Microalgas bentônicas fazem parte de uma categoria ecoló-gica chamada de microfitobentos. Os termos “fitoplâncton” e “microalgas”, portanto, nãosão sinônimos.
A evolução das algas e o estabelecimento de relações filogenéticas entre os gruposmodernos desses organismos são temas extremamente controvertidos e sujeitos a diferentesinterpretações, resultantes da acumulação de novas informações. O estudo dos grandes gruposde algas está numa das fronteiras da Biologia e da própria compreensão da evolução dos seres
33

34
vivos, pois em muito contribui para o entendimento dos organismos conhecidos comoprotistas. Há várias propostas de organização, e muitas vezes diferem pelos caracteres escolhi-dos como mais importantes para fundamentar o sistema de classificação.
Nesta obra, os grandes grupos de algas são apresentados sucintamente, pois não há ointuito de aprofundar a discussão sobre esse assunto, o qual vem sendo tratado em livrossobre Ficologia (por exemplo: Hoek et al., 1995; Sze, 1998; Lee, 1999; Graham & Wilcox,2000; Reviers, 2006). Busca-se aqui apenas oferecer elementos para contextualizar ade-quadamente a posição sistemática das microalgas utilizadas em atividades de cultivo. Assim,grupos com representantes macroscópicos e dulciaqüícolas também são abordados, eviden-ciando as diferenças entre as muitas espécies cultivadas com finalidades diversas.
2.1.1 AS ALGAS PLANCTÔNICAS COMO PRODUTORES PRIMÁRIOS
No mar, a atividade fotossintética é realizada por vários tipos de organismos, desta-cando-se, dentre eles, o fitoplâncton, o microfitobentos, as algas macroscópicas, as plantasvasculares e as associações simbióticas envolvendo algas, como as zooxantelas, zooclorelas ecianelas. Esse vasto conjunto de organismos fotossintetizantes apresenta papel ecológicoequivalente ao das plantas terrestres em ecossistemas continentais. Os organismos fotoauto-tróficos marinhos apresentam grandes diferenças morfológicas, fisiológicas, bioquímicas eecológicas entre si, diferentemente do que ocorre no ambiente terrestre, em que as diversasespécies vegetais não algáceas apresentam características relativamente similares (Lourenço& Marques Júnior, 2002). Tal contraste resulta do processo evolutivo que envolve os diversosprodutores primários existentes. Assume-se que vários grupos de algas não apresentamrelações filogenéticas entre si, tendo origens distintas e polifiléticas. Na terra, as plantas comflores, as briófitas (musgos, hepáticas e antóceros) e as plantas vasculares sem sementes (psilo-tófitas, licófitas, esfenófitas e pterófitas) derivam de um mesmo ancestral, presumivelmente algascarofíceas, por isso compartilham muitas semelhanças químicas e estruturais.
Em função de sua natureza procarionte, para muitos autores, as cianobactérias não devemser consideradas parte do fitoplâncton, que então seria constituído apenas por organismoseucariontes (neste caso, todas as algas seriam protistas). Entretanto, vários outros autoresargumentam que na natureza os produtores primários planctônicos eucarióticos e procarióticosdesempenham o mesmo papel ecológico, o que justificaria a inclusão das cianobactériasplanctônicas no fitoplâncton. Essa é a abordagem adotada nesta obra, que valoriza mais o papelecológico dos componentes do que as características de sua organização celular ou posiçãosistemática.
O fitoplâncton, as algas macroscópicas, as gramas marinhas e o microfitobentos são osprincipais grupos de produtores primários fotossintetizantes que ocorrem no mar. Plantas demangue e plantas de marisma são vegetais importantes nos locais onde ocorrem, gerando grandeprodução primária em áreas costeiras. Porém, são plantas terrestres tolerantes à salinidade daágua do mar, não se tratando de organismos marinhos, ao contrário dos outros produtoresfotoautotróficos mencionados acima. De qualquer forma, as plantas de mangue e as de marismaocorrem em áreas de transição entre o ambiente marinho e o terrestre; como grande parte desuas contribuições à produção primária dirige-se para o mar, torna-se plausível sua avaliaçãojuntamente com os produtores primários marinhos verdadeiros.
Bactérias e árqueas quimioautotróficas1 também são produtores primários importantesno mar, especialmente em determinados ambientes profundos ricos em minerais dissolvidos
1. Ver também bactérias e árqueas quimiolitoautotróficas.

35
que são envolvidos em processos de oxirredução ou emanações de hidrocarbonetos simples: asaberturas hidrotermais e as nascentes frias. Bactérias e árqueas quimioautotróficas sãoprodutores primários não-fotossintetizantes que utilizam CO
2 como fonte de carbono, mas
obtêm energia para sua redução pela oxidação aeróbica de substratos inorgânicos. Osprocariontes quimioautotróficos exibem grande especificidade de substrato para obter ener-gia, de forma que determinada espécie tende a apresentar capacidade de metabolizar apenasum substrato, como o metano e derivados inorgânicos de enxofre, por exemplo. Há tambéma ocorrência de bactérias quimioautotróficas em águas mais rasas, geralmente associadas asedimentos ricos em enxofre, como aqueles encontrados em muitos manguezais. Embora sejalocalmente essencial, a produção primária resultante da quimiossíntese representa parcela muitopequena para o balanço global da produção primária marinha.
Estima-se que a fotossíntese e a respiração envolvam 85 bilhões de toneladas métricas(85 x 1015 g) de carbono nos ecossistemas da Terra (1015 g = 1,0 gigatonelada, Gt – o mesmo que1,0 petagrama, Pg). Esse valor impressionante representa pequena parcela do carbono incorporadoà matéria orgânica dos seres vivos que constituem a biosfera, que, segundo Ricklefs (2003), atinge2.650 Gt, com tempo médio de residência na biota de 31 anos (calculado como 2.650 Gtexistentes na biota divididos pelas 85 Gt que representam a atividade fotossintética anual nomundo). Tais números podem ser diferentes, se outros autores forem considerados. Behrenfeldet al. (2001) estimaram que a fotossíntese anual da biosfera seria de 111-117 Pg de carbono, oque estabeleceria menor tempo médio de residência do carbono na biosfera, de 23,2 anos. Estima-se que os produtores primários marinhos fixem anualmente nos oceanos cerca de 45-50 Gt decarbono. Tais valores corresponderiam a 52,9%-58,8% da produção primária global, segundoRicklefs (2003), mas apenas a 38,5%-45,0% da produtividade global da biosfera, de acordo comBehrenfeld et al. (2001).
Calcula-se que pelo menos 90% da produção primária marinha global seja derivada daatividade do fitoplâncton. Assim, todos os demais grupos de produtores primários marinhosreunidos contribuem com fração menor da produção primária dos oceanos mundiais. Essainformação eventualmente causa estranheza a alguns leitores pelo reconhecimento da existênciade ecossistemas dominados por produtores primários, como as algas macroscópicas e gramasmarinhas, por exemplo, em áreas costeiras. Nesses casos ocorre inversão em relação aos argumentosjá apresentados, e a contribuição do fitoplâncton para a produção primária em tais ecossistemaspode ser relativamente pequena, bem menor do que a de outros produtores primários locais.Porém, ecossistemas dominados por gramas marinhas e algas macroscópicas correspondem aparcela pequena dos oceanos mundiais: eles ocorrem apenas em áreas rasas e, quando reunidas,correspondem a menos de 1% da área dos oceanos do mundo. Na enorme vastidão dos oceanospredomina a presença e a contribuição do fitoplâncton, que se distribui por todo o espaço onde hápenetração de luz solar, que pode corresponder a mais de 200 m de profundidade. Uma vez que aprodução primária se refere à incorporação de carbono no ambiente, as taxas de produção primáriageralmente são expressas como gramas de carbono fixado por área ou volume de água, por intervalode tempo, constituindo a chamada produtividade primária. Por exemplo, a produtividade primáriade determinado local pode ser registrada em gramas de carbono por metro quadrado por ano(g C.m–2.ano–1), representando a produção integrada em todas as profundidades da superfície até abase da zona eufótica. A Tabela 2.1 exibe medidas de produtividade primária em diferentes ecos-sistemas marinhos e estuarinos, os quais são dominados por diversos produtores primários.
Mesmo sendo o fitoplâncton a principal categoria de produtores primários marinhos, aprodutividade primária por ele exercida no ambiente pelágico é extremamente variável nasdiferentes regiões dos oceanos. Tais diferenças resultam principalmente da combinação da

36
disponibilidade de nutrientes dissolvidos e de luz nas diversas regiões dos oceanos do mundo.Nutrientes e luz são os principais fatores que afetam a produção primária fotossintetizante. Naszonas costeiras, os valores da produção primária fitoplanctônica anual tipicamente oscilam entre120 e 250 g C m–2.ano–1, nas áreas de ressurgência, a produtividade primária pode atingir 370 gC m–2.ano–1; e nas áreas oceânicas tropicais, observam-se valores médios baixos, em torno de 30g C m–2.ano–1 (Tabela 2.2).
Tabela 2.1 Medidas de produtividade primária em diferentes ecossistemas costeiros do mundo. As medidasestão expressas em g C m–2 ano–1.
Ecossistema e respectiva localização
Produtividade primária
Fonte
Pradarias de gramas marinhas Cymodocea nodosa (Mediterrâneo) 2.000-6.750 Zieman & Wetzel (1980) Halodule wrightii (EUA: Carolina do Norte) 182-730 Zieman & Wetzel (1980) Halophila (Austrália) 16-736 Mann (2000) Poisidonia oceanica (Malta) 730-2.190 Zieman & Wetzel (1980) Phyllospadix scouleri (México: Baixa Califórnia), valor máximo 8.000 Ramirez-Garcia et al. (1998) Thalassia (Caribe), valor médio 800 Mann (2000) Thalassia testudium (lagunas das Bahamas) 602-836 Koch & Madden (2001) Thalassia hemprichii (Indonésia) 912-1314 Erftemeijer et al. (1993) Thalassodendron ciliatum (Moçambique) 3.468-9.322 Bandeira (2002) Zostera (Dinamarca), valor médio 800 Mann (2000) Zostera marina (EUA: Carolina do Norte e Alasca) 73-2.920 Zieman & Wetzel (1980) Marismas Bancos de Spartina (Canadá: Nova Escócia) 803-1.051 Livingstone & Patriquin (1981) Bancos de Spartina (EUA: Nova Jersey) 500-2.300 Smith et al. (1979) Bancos de Spartina (EUA: Pacífico Noroeste) 100-1.000 Seliskar & Gallagher (1983) Scirpus maritimus (estuário da lagoa dos Patos, RS) 1.013-1.984 Costa et al. (1997) Spartina alterniflora (estuário da lagoa dos Patos, RS) 1.693-4.873 Marinucci (1982) Manguezais Manguezais (América tropical) 1.022-8.760 Brown & Lugo (1982) Kandelia candel (Vietnã) 176-1.339 Coulter et al. (2001) Manguezais de Cananéia (SP), valor médio 610 Adaime (1987) Manguezais da baixada Santista (SP), valor médio 460 Ponte et al. (1990) Bancos de macroalgas Laminaria (Atlântico Norte), valor médio 2.000 Castro & Huber (2005) Macrocystis (EUA: Califórnia), valor médio 1.500 Castro & Huber (2005) Ecklonia (Austrália), valor médio 1.000 Castro & Huber (2005) Recifes de coral Atol Enewetak (ilhas Marshall, Pacífico Central) 1.500-5.000 Atkinson (1992) Recifes de coral (diversos) 1.500-3.700 Castro & Huber (2005) Recifes de coral (diversos) 2.000-5.000 Sze (1998) Recifes de coral (diversos) 1.500-5.000 Lalli & Parsons (1997)

37
Tabela 2.2 Medidas de produtividade primária em diferentes áreas geográficas do mundo. Os númerosreferem-se à produção primária atribuída ao fitoplâncton.
Ambientes Produtividade(g C m–2 ano–1)
Fonte
Áreas de ressurgência costeira 110-370 Castro & Huber (2005)Áreas de ressurgência equatorial 70-180 Castro & Huber (2005)Estuários (média) 256 Day Jr. et al. (1989)Giros oceânicos centrais 4-40 Castro & Huber (2005)Mares subpolares 50-110 Castro & Huber (2005)Oceano Antártico 40-260 Castro & Huber (2005)Oceano Ártico < 1-100 Castro & Huber (2005)Oceanos temperados (costeiros) 110-220 Castro & Huber (2005)Oceanos temperados (oceânicos) 70-120 Nybakken & Bertness (2005)Oceanos tropicais 18-50 Nybakken & Bertness (2005)Plataforma continental 100-160 Nybakken & Bertness (2005)Zonas costeiras (média) 100 Ryther (1963)
2.1.2 CLASSES DE TAMANHO DO PLÂNCTON MARINHO
Uma característica inerente ao plâncton é o alto grau de mistura de seus componentes,já que eles apresentam movimentos limitados e são transportados pela água. Este fato temcomo conseqüência o estabelecimento de estratégias de amostragem que contemplem aobtenção dos organismos de interesse no plâncton. Uma forma simples de dividir os com-ponentes do plâncton a fim de estudá-los é estabelecendo classes de tamanho. Os diversosconstituintes do plâncton são convencionalmente separados em duas categorias fundamentaisde tamanho, em função de sua retenção ou não em redes com malha de 20 μm. Todas as espéciesque ficam retidas na rede e que, por conseguinte, apresentam tamanho igual ou superior a20 μm são designadas como plâncton de rede (as frações que atravessam a rede não recebemdesignação especial). Alguns autores consideram plâncton de rede as frações retidas em malhasmaiores, como 50 μm, 63 μm ou 80 μm, mas o valor de 20 μm é universalmente mais aceitopara designar o plâncton de rede. Essa é a abordagem adotada nesta obra.
A classificação do plâncton em apenas duas classes de tamanho é uma simplificaçãoaceita e utilizável apenas com caráter abrangente e didático. Nos estudos desenvolvidos emplanctonologia (ou planctologia)2 normalmente é necessário derivar mais a distribuição dasclasses de tamanho de componentes do plâncton para melhor entender a dinâmica e osprocessos que envolvem tais organismos. Constata-se, por exemplo, que, embora os organis-mos planctônicos sejam microscópicos ou submicroscópicos em sua quase totalidade, algunscomponentes, como as medusas, podem alcançar grandes dimensões (até 1 metro de diâ-metro). O simples agrupamento dos organismos planctônicos em uma única categoria,abrangendo seres com mais de 20 μm de diâmetro, constitui grande distorção em muitoscasos.
2. Estes termos criados para designar o estudo científico do plâncton ainda são relativamente pouco usadosno Brasil e no exterior (seus equivalentes em outras línguas), embora sejam formalmente reconhecidos.

38
Assim, maior detalhamento na distribuição das classes de tamanho é necessário paracontemplar os organismos de maiores dimensões. Similarmente, o desenvolvimento daspesquisas tem revelado a existência de componentes extremamente pequenos no plâncton,com dimensões de poucos micrômetros ou mesmo décimos de micrômetro. Ainda poucoconhecidos pela Ciência, esses componentes minúsculos do plâncton constituem uma dasfrentes mais promissoras de estudos dentro da abrangência da Biologia Marinha e Oceano-grafia; o avanço nas pesquisas sobre esses organismos poderá revelar informações fundamentaispara a compreensão de processos que envolvem componentes pelágicos e bentônicos.
Tendo por base esse conjunto de informações, autores modernos têm proposto outrascategorias de tamanho para os organismos planctônicos. Uma dessas propostas foi feita porSieburth (1979). Segundo esse autor, sete categorias de tamanho devem ser consideradas.O megaplâncton seria constituído por organismos que alcançam mais de 20 cm (por exemplo,muitas medusas, caravelas e sargaços flutuantes). O macroplâncton envolveria indivíduosque medem entre 2 e 20 cm (por exemplo, ctenóforos e diversos crustáceos, como o krill).O mesoplâncton compreenderia componentes cujas dimensões variam entre 200 mm e 20mm (por exemplo, algumas algas unicelulares grandes ou em cadeias de células, protozoáriosgrandes, formas larvares de metazoários e parte dos ovos de peixes). O microplâncton contem-plaria espécies que apresentam tamanho entre 20 e 200 μm (por exemplo, grande fraçãodas algas unicelulares e dos protozoários marinhos, diversas formas larvares de metazoários).O nanoplâncton envolveria organismos com dimensões entre 2 e 20 μm (por exemplo, partedas algas unicelulares, leveduras, protozoários e ovos de invertebrados). O picoplâncton éconstituído por organismos que apresentam dimensões entre 0,2 e 2 μm (por exemplo,bactérias heterotróficas, cianobactérias e algumas algas eucarióticas). E, finalmente, o autorpropõe o fentoplâncton, envolvendo seres de 0,02 a 0,2 μm (por exemplo, vírus marinhos,seus únicos componentes).
Finalmente, nota-se que as classes de tamanho segundo as quais o plâncton é divididonão apresentam identidade taxonômica quanto a seus componentes. Verifica-se a existênciade espécies de determinado filo ou divisão com tamanhos variados e distribuídas por diversasclasses de tamanho. Por exemplo, a maioria das diatomáceas é encontrada no microplâncton(por exemplo, gêneros Cyclotella e Ditylum), mas diatomáceas pequenas podem ser encon-tradas no nanoplâncton (por exemplo, Minutocellus) e cadeias de células (por exemplo,Melosira) ou indivíduos unicelulares grandes (por exemplo, Coscinodiscus) podem ser coletadosno mesoplâncton. Indivíduos adultos de crustáceos do gênero Calanus (classe Copepoda)pertencem ao mesoplâncton, mas seus ovos e estádios larvares iniciais pertencem ao micro-plâncton. Não é possível, tampouco, usar determinado artefato de coleta no campo (porexemplo, rede de plâncton, garrafa oceanográfica) e obter apenas os organismos de um grupode interesse.
Coletas de organismos planctônicos são definitivamente marcadas pelo elevado graude mistura de espécies que fazem parte de categorias taxonômicas distintas. Assim, não háuma “rede de fitoplâncton” nem uma “rede de zooplâncton”, mas, sim, redes cujos tamanhosdos poros são adequados para coletar mais organismos do fitoplâncton (por exemplo, malhascom poros de 20 µm) ou do zooplâncton (por exemplo, malhas com poros de 150 µm). Oesquema de classificação de Sieburth (1979) para categorias de tamanho de organismosplanctônicos é apresentado na Tabela 2.3, que permite fazer relações breves com grupostaxonômicos e métodos de coleta.

39
Tabe
la 2
.3 C
lass
ific
ação
dos
org
anis
mos
pla
nctô
nico
s po
r cl
asse
s de
tam
anho
, dim
ensõ
es t
ípic
as, e
xem
plos
e m
étod
os d
e co
leta
dos
org
anis
mos
.
Fe
ntop
lânc
ton
(0,0
2-0,
2 μm
)
Pic
oplâ
ncto
n
(0,2
-2,0
μm
)
Nan
oplâ
ncto
n
(2,0
-20 μm
)
Mic
ropl
ânct
on
(20-
200 μm
)
Mes
oplâ
ncto
n (0
,2-2
0 m
m)
Mac
ropl
ânct
on
(2-2
0 cm
) M
egap
lânc
ton
(20-
200
cm)
Vir
iopl
ânct
on
----
----
----
----
-
B
acte
riop
lânc
ton
--
----
----
----
-
Mic
oplâ
ncto
n
--
----
----
----
--
Fito
plân
cton
----
----
----
----
--
----
----
----
--
----
----
----
----
- --
----
----
----
---
Pro
tozo
oplâ
ncto
n
--
----
----
----
--
----
----
----
----
- --
----
----
----
---
Met
azoo
plân
cton
----
----
----
----
- --
----
----
----
---
----
----
----
----
--
----
----
----
----
- T
aman
ho (m
) 10
–7
10–6
10
–5
10–3
10
–2
10–1
10
0 E
xem
plos
de
orga
nism
os
víru
s ba
ctér
ias
e eu
cari
onte
s m
inús
culo
s
coco
litof
oríd
eos,
diat
omác
eas
pequ
enas
, fla
gela
dos
auto
tróf
icos
e
hete
rotr
ófic
os
diat
omác
eas,
dino
flage
lado
s,
cilia
dos,
fla
gela
dos
auto
tróf
icos
e
hete
rotr
ófic
os
copé
podo
s,
larv
as d
e in
vert
ebra
dos
e
peix
es,
quet
ogna
tos,
m
icro
alga
s em
ca
deia
s
salp
as, m
edus
as,
cten
ófor
os
sifo
nófo
ros
colo
niai
s,
sarg
aço
flutu
ante
, m
edus
as
gran
des
Mét
odos
de
cole
ta
mai
s co
mun
s ga
rraf
a oc
eano
gráf
ica,
co
leta
com
bo
mba
de
sucç
ão
garr
afa,
fil
traç
ão,
sedi
men
taçã
o,
bom
ba d
e su
cção
garr
afa,
fil
traç
ão,
sedi
men
taçã
o,
cent
rifu
gaçã
o bo
mba
de
sucç
ão
rede
s de
pl
ânct
on,
garr
afa,
bom
ba
de s
ucçã
o
rede
s de
pl
ânct
on,
bom
ba d
e su
cção
rede
s de
pl
ânct
on, c
olet
a m
anua
l
rede
s de
pl
ânct
on,
cole
ta m
anua
l

40
2.1.3 PLASTOS E PIGMENTOS FOTOSSINTÉTICOS
Com exceção das cianobactérias (incluindo as proclorofíceas), as quais são bactériasespecializadas que possuem clorofila a, as chamadas algas consistem em espécies eucarióticasdotadas das diversas organelas celulares, dentre as quais os plastos. Estes são estruturas verdes,pardas, alaranjadas ou vermelhas (de acordo com os pigmentos predominantes que apre-sentam), delimitados por duas, três ou quatro membranas. Internamente, os plastos sãoconstituídos por camadas de membranas internas, os tilacóides, em que ocorrem as principaisreações bioquímicas da fotossíntese, e por uma matriz amorfa, o estroma (Figura 2.1).Embora os plastos presentes nas algas nem sempre sejam verdes, na literatura eles freqüen-temente são chamados de forma genérica de cloroplastos, o mesmo termo atribuído aosplastos verdes e fotossintetizantes de plantas terrestres. Atualmente, a maioria dos autoresacredita que a origem dos plastos seja resultado de relação simbiótica estável entre umorganismo eucarionte heterotrófico (o hospedeiro) e um organismo procarionte fotossin-tetizante (o endossimbionte). Relações endossimbióticas desse tipo são comuns na natureza,e acredita-se que mitocôndrias tenham sido originadas de forma similar. A presença dealgumas membranas envolvendo os plastos é um indício do processo de endossimbiose, quepode ter sido primária, secundária ou mesmo terciária, entre os diferentes grupos de algas(Figura 2.2). A existência de ribossomas 70S, típico de organismos procariontes e de materialgenético (ADN) no interior dos plastos, é encarada como evidência adicional da ocorrênciado processo de endossimbiose.
a) Membranasexternas einternas
Lamelas doestroma
Estroma
Grana
Tilacóide
Estroma
b)
Lamelas doestroma
Granum
Figura 2.1 Micrografia eletrônica de um cloroplasto da folha da poácea Phleum pratense (18.000x), comindicação dos componentes estruturais da organela (a). A mesma preparação em uma magnificação maior(52.000x) (b). Modificado de Taiz & Zeiger (1998).
Grande variedade de pigmentos fotossintéticos é encontrada em algas. Todas as espéciesfotossintetizantes são dotadas de clorofila a, pigmento mais importante para a fotossíntesee que apresenta papel central no arranjo dos fotossistemas para a captação de energia luminosa.

41
A maioria das algas dispõe de uma segunda clorofila, que pode ser a clorofila b, c ou d.Diferenças estruturais na molécula de clorofila determinam os diversos tipos conhecidos(Figura 2.3). As clorofilas b, c e d são consideradas pigmentos acessórios da fotossíntese, poiso papel principal nos sistemas de absorção de luz é desempenhado pela clorofila a, cabendoàs demais clorofilas aumentar o total de luz absorvido pelo organismo. A clorofila a apresentapicos de absorção de radiação luminosa em 430 e 664 nm; a clorofila b, em 435 e 645 nm;a clorofila c1, em 440, 583 e 634 nm; a clorofila c2, em 452, 586 e 635 nm; e a clorofila d,em 400, 456 e 700 nm. Entretanto, nem todas as algas possuem uma segunda clorofila, deforma que diversas espécies realizam fotossíntese apenas com a participação de clorofila a ede outros pigmentos não clorofilianos.
Dinoflagelado complasto derivado deuma criptomônada
Redução doendossimbionte
Dinoflagelado comcriptomônadaendossimbionte
Endossimbioseterciária
Criptomônada
Endossimbiosesecundária
Alga vermelha
Cianobactéria
Endossimbioseprimária
DN
CN
RN
DN
CN
RN
RNFigura 2.2 Representação de processos de en-dossimbiose primária, secundária e terciária,envolvendo diferentes grupos de algas, comredução das estruturas celulares dos endossim-biontes. RN = núcleo da alga vermelha; CN =núcleo da criptomônada e DN = núcleo dodinoflagelado. Modificado de Wilcox &Wedemayer (1985).

42
Mg
N
O
N
N
N H
HH
H
H
HHH
H
H
H
HH
H
H HHH
H
H
H
O
O
H
HHH
HH
OO
H
H
H
H
HH
H
HHHH
H H H
HH
H
HHHH
H
H
H
HH
H
HHHH
H H H
HH
H
H H
Mg
N
O
N
N
N H
HH
H
H HH
H H
H
HH
H
HH
H
H
O
O
H
HHH
H
OO
H
H
HH
H
HH
H
O HHH
H
OH
HH
H
a, b
d
a, d b
HH
H
c1
c2
Uma segunda classe de pigmentos fotossintetizantes é representada pelos carotenóides(Figura 2.4), que são pigmentos solúveis em solventes orgânicos e dotados de coloraçãoamarela, laranja, vermelha, marrom ou variações dessas cores. Carotenóides são divididosem dois grupos: os carotenos, moléculas dotadas apenas de carbono e hidrogênio; e as
Figura 2.3 Estruturas das moléculas de clorofilaa, b, c1, c2 e d. Obtido de Wikipedia (2006).

43
xantofilas, moléculas que, além de carbono e hidrogênio, apresentam oxigênio em suaestrutura. Há dezenas de carotenóides na natureza, mas cada espécie fotossintetizante dispõede apenas alguns deles, geralmente não mais do que 10 diferentes carotenóides. É importantesalientar que os conjuntos de carotenóides variam entre os grupos de algas, os quais assumemimportante papel para a organização sistemática das algas, havendo substâncias quefuncionam como marcadores taxonômicos. Um bom exemplo é a aloxantina, uma xantofilaexclusiva das algas conhecidas como criptófitas (membros da divisão Cryptophyta). Os carote-nóides têm duas funções principais: aumentar a captação de luz dos fotossistemas (a exemplodas clorofilas b, c e d) e proteger os fotossistemas quando há luz em excesso, funcionando comouma espécie de filtro. Esta segunda função é especialmente interessante, pois revela que, emboraa luz seja essencial para o funcionamento da vida fotossintetizante, sua presença excessiva podeser deletéria, desencadeando um processo conhecido como fotoinibição.
a)
HO
OH
b)
HO
OH
c)
O
d)
O
e)
O O
f)
O
OH
HO
g)
OH
HO
O
O
h)O
OCOCH3
HOO
HO
i)
H C O3C O
OHO
OHO
O
Figura 2.4 Exemplos de alguns caro-tenóides encontrados em algas: (a) ß-caroteno; (b) luteína; (c) zeaxantina;(d) equinenona; (e) cantaxantina; (f )astaxantina; (g) violaxantina; (h) fuco-xantina; (i) peridinina.

44
Uma terceira classe de pigmentos fotossintetizantes é composta pelas ficobiliproteínas,que são pigmentos azuis ou vermelhos, solúveis em água, presentes em apenas quatros gruposde algas (cianobactérias, rodófitas, glaucófitas e criptófitas). A presença de ficobiliproteínasconfere às algas coloração azulada ou avermelhada, dependendo das substâncias (e concen-trações) existentes. As ficobiliproteínas são compostas de duas partes: a proteína em si e aficobilina, fortemente unidas por ligações covalentes. A ficobilina (Figura 2.5) é efetivamenteo pigmento fotossintético, mas ele não apresenta papel funcional se não houver uma proteínaacoplada. Genericamente, as ficobilinas azuis são chamadas de ficocianinas e as vermelhas,de ficoeritrinas. Há alguns tipos diferentes de ficocianina e de ficoeritrina entre os gruposde algas e, a exemplo dos carotenóides, os conjuntos de pigmentos têm importância taxonô-mica. Com exceção das criptófitas, nas demais algas as ficobiliproteínas se dispõem sobre asmembranas dos tilacóides, constituindo estrutura visível por microscopia eletrônica conhe-cida como ficobilissomo. Estes se dispõem exatamente sobre os centros de reação da fotossíntesee são capazes de transferir energia para os fotossistemas. Assim, as ficobiliproteínas aumentamo espectro de captação de luz pela fotossíntese, mas também apresentam uma segunda função:atuar como reserva de nitrogênio. Em situações de carência de nitrogênio, as proteínas quecompõem as ficobiliproteínas são degradadas e os ficobilissomos são eliminados, liberandonitrogênio para os processos metabólicos essenciais. A alga realiza menos fotossíntese, assumecoloração esverdeada (em vez da coloração original típica, avermelhada ou azulada), mas podesobreviver a um período de carência de nitrogênio. Se o nitrogênio tornar-se disponível emconcentração suficiente, as ficobiliproteínas podem ser sintetizadas e os ficobilissomos podemser novamente constituídos.
CH3
NO
CH CH3 CH2 CH2 CH3 CH3 CH2
N N N
CH2 CH2
COOH COOH
CH3 CH3
H H HO
a)
CH3
NO
CH CH3 CH2 CH2 CH3 CH3 CH
N N N
CH2 CH2
COOH COOH
CH3 CH2
H H HO
b)
Figura 2.5 Estruturas de ficobilinas: (a) ficocianina; (b) ficoeritrina. Extraído de Sze (1998).
2.1.4 REPRODUÇÃO E CICLOS DE VIDA EM ALGAS
Um dos aspectos mais importantes no estudo das algas é a existência de padrões reprodutivosmarcantes e bastante variados entre os grupos de algas. A reprodução pode ser assexuada ousexuada, e esses padrões básicos apresentam elevado grau de diversificação dos processos.

45
Figura 2.6 Representação dos tipos básicosde reprodução quanto à morfologia dos ga-metas formados: (a) isogamia; (b) aniso-gamia; (c) e (d) oogamia.
A reprodução assexuada envolve a produção de esporos. Estes são produzidos a partirde células vegetativas ou em regiões especiais do talo de formas multicelulares, denominadasesporângios ou esporocistos. Os esporos podem ser móveis, chamados de zoósporos, ouimóveis, conhecidos como aplanósporos (estes têm a capacidade de se converterem emzoósporos). Zoósporos são dotados de um ou mais flagelos, ao passo que os aplanósporossão desprovidos de flagelos. Zoóporos podem apresentar uma mancha ocelar, que é umapequena estrutura rica em pigmentos, localizada numa das extremidades da célula, cujafunção está relacionada à fotorrecepção e orientação dos movimentos. Outras formas deesporos não-móveis são os autósporos ou monósporos, que diferem dos aplanósporos porapresentarem capacidade de se converterem em zoósporos. Algumas espécies podem sereproduzir por meio do processo conhecido como produção de autocolônia. A fragmentaçãodo talo pode ser entendida como uma forma de reprodução vegetativa, que pode ser realizadapor meio da mera ruptura do talo (em espécies multicelulares), e cada fração resultante crescee forma um novo talo. A reprodução vegetativa pode se manifestar por meio de divisão bináriada célula, no caso de espécies unicelulares.
A reprodução sexuada envolve grande variedade de possibilidades entre os diversosgrupos de algas. A principal maneira de classificar a reprodução sexuada diz respeito aos tipose formas de gametas produzidos (Figura 2.6). As formas apresentadas pelos gametas que sefundem podem ser altamente variadas. Na isogamia, dois gametas móveis são idênticos ouquase idênticos; na anisogamia, os dois gametas são diferentes em tamanho ou compor-tamento; e na oogamia, um gameta pequeno, móvel ou não, se funde com um gameta bemmaior e imóvel. Na anisogamia e na oogamia, os dois tipos de gametas podem ser produzidospor um mesmo indivíduo, que é designado como monóico. Se os gametas forem produzidospor indivíduos diferentes, estes são ditos dióicos. Se os gametas produzidos por um mesmoorganismo puderem se fundir, gerando autofecundação, o organismo é designado comohomotálico. Se a autofecundação não for viável em razão de fatores genéticos, então, diz-seque o organismo formado é heterotálico. Os gametas são produzidos a partir de célulasvegetativas ou são formados em regiões especiais do talo de formas multicelulares, deno-minadas gametângios ou gametocistos.
(a) (b)
(c) (d)

46
Algas podem apresentar talos que produzem esporos ou gametas, podendo ter cargacromossomial haplóide (N) ou diplóide (2N). Indivíduos que formam gametas são designadoscomo gametófitos, ao passo que aqueles que produzem esporos são os esporófitos. Há trêspadrões básicos, de acordo com o momento em que acontece a meiose: (a) a fase vegetativa(maior porção do ciclo) é haplóide e a meiose ocorre após a fecundação do zigoto, que édiplóide (Figura 2.7a); (b) a fase vegetativa é diplóide e a meiose dá origem a gametashaplóides (Figura 2.7b); (c) ocorrem duas ou três fases, com gametófitos haplóides (produçãode gametas por mitose) e esporófitos diplóides (produção de esporos por meiose), caracte-rizando alternância de gerações (Figura 2.7c).
Os ciclos de vida representados pelas Figuras 2.7a e 2.7b são chamados haplobiônticos,pois neles apenas uma fase predomina na natureza. Se a fase predominante for constituídapor organismos haplóides, o ciclo de vida é chamado de haplobiôntico haplonte ou haplôn-tico; se a fase predominante for constituída por organismos diplóides, o ciclo de vida échamado de haplobiôntico diplonte ou diplôntico. Por outro lado, se não há predomíniode fases na natureza, existindo formas diplóides e haplóides de vida livre, o ciclo de vida échamado de diplobiôntico (Figura 2.7c); se apresentar duas fases com cargas cromossomiaisdistintas ele é chamado de diploaplôntico. Se as duas fases do ciclo diploaplôntico sãomorfologicamente idênticas, caracteriza-se um ciclo isomórfico; se elas forem diferentes, diz-se que o ciclo é heteromórfico.
Em algumas algas é possível também que gametas não produzam fecundação e zigotos,dando origem diretamente a indivíduos haplóides. Uma vez que não há fecundação, a formade reprodução é assexuada, apesar do envolvimento de gametas. Esse processo especial dereprodução é conhecido como partenogênese.
Singamia
Singamia
Meiose
Meiose
Gametas
Gametas
Mitose
Mitose
Talo (n)
Talo (n)
Células (n)
Células (n)
(a)
(c)
Zigoto (2n)
Mitose
Mitose
Singamia
Meiose
Gametas
Mitose
Talo (2n)
(b)Zigoto (2n)
Mitose
Zigoto (2n)Mitose
Talo (2n)
Figura 2.7 Representação simplificada dos trêstipos básicos de ciclos de vida de algas: (a) ha-plobionte haplôntico; (b) haplobionte diplôn-tico; (c) diploaplôntico. Os retângulos repre-sentam as formas vegetativas e exibem as res-pectivas cargas cromossomiais existentes. Notaras diferenças quanto à posição da meiose nosciclos de vida representados, para formar ga-metas ou esporos.

47
2.1.5 NOMENCLATURA DOS GRUPOS DE ALGAS
Há fundamentalmente três códigos internacionais de nomenclatura biológica: zoo-lógica, botânica e bacteriana. Trata-se de formas organizadas de estabelecer normas para aescolha de nomes para os grupos de organismos representados, sobretudo táxons elevados(ordem, classe, etc.). Tradicionalmente, as algas têm sido classificadas utilizando-se o CódigoInternacional de Nomenclatura Botânica (CINB). As regras para nomenclatura biológicapodem mudar com o tempo, um resultado de avanços no conhecimento científico. Para asalgas são seguidos os princípios estabelecidos na última versão do CINB, conhecido comocódigo de Saint Louis, cidade norte-americana onde ocorreu o último conclave internacionalque estabeleceu atualização das regras. O Código de Saint Louis foi traduzido para o por-tuguês por Bicudo & Prado (2003).
O CINB é muito extenso e contempla todos os procedimentos para atribuição denomes a plantas e algas. Aqui o interesse restringe-se apenas a apresentar os sufixos empregadospara táxons elevados.
Divisão (= filo): -phyta. Ex.: divisão Ochrophyta.Classe: -phyceae. Ex.: classe Dinophyceae.Subclasse: -phycidae. Ex.: subclasse Florideophycidae.Ordem: -ales. Ex.: ordem Pavlovales.Subordem: -ineae. Ex.: subordem Chromulineae.Família: -aceae. Ex.: família Zygnemataceae.Subfamília: -oideae. Ex.: subfamília Corallinoideae.Para mais detalhes da nomenclatura botânica é essencial consultar o CINB.
2.2 OS GRANDES GRUPOS DE ALGAS
Algas que fazem parte do fitoplâncton são as mais utilizadas em cultivos. Microalgasbentônicas são também cultiváveis, mas geralmente crescem mais lentamente e por isso têmsido menos utilizadas em pesquisas e aplicações específicas. Por essa razão, maior ênfase seráconferida a espécies fitoplânctônicas neste texto.
Todos os grandes grupos de algas (divisões ou filos) têm representantes no fitoplâncton(Tabela 2.4). As espécies dotadas de clorofila a e outros pigmentos acessórios (Figuras 2.3-2.5) são fotoautotróficas e constituem-se em fonte primária de alimento aos animais eprotistas heterotróficos da coluna d’água e dos sedimentos. Diversos grupos de algas perten-centes ao fitoplâncton não apresentam relações filogenéticas entre si, exibindo diferençasexpressivas que se manifestam em seus produtos de reserva (Figura 2.8), pigmentos fotossin-tetizantes acessórios e também substâncias constituintes das paredes celulares (quandopresentes).
Há também espécies desprovidas de capacidade fotossintética, mas que pertencem aosmesmos grupos taxonômicos de algas fotossintetizantes. Interpretam-se esses casos comoresultantes da perda de pigmentos fotossintetizantes durante o processo evolutivo ou, aparen-temente em alguns casos, pela não aquisição dos pigmentos ao longo da evolução das espécies.Tais algas heterotróficas, apesar do parentesco próximo com componentes do fitoplâncton,não são tratadas como pertencentes ao fitoplâncton, que, pelo conceito adotado nesta obra,é composto apenas por espécies fotossintetizantes.

48
Tabe
la 2
.4 C
arac
terí
stic
as m
orfo
lógi
cas,
quí
mic
as e
rep
rodu
tiva
s fu
ndam
enta
is d
os p
rinc
ipai
s gr
upos
mod
erno
s de
alg
as. M
odif
icad
o e
expa
ndid
o a
part
ir d
eG
raha
m &
Wilc
ox (
2000
).
Div
isão
/gru
po
Prin
cipa
is p
igm
ento
s fo
toss
inté
tico
s P
rodu
to d
e re
serv
a Fl
agel
o R
epro
duçã
o se
xuad
a C
ober
tura
cel
ular
D
imen
sões
Chl
orox
ybac
teri
a
(= C
yano
phyt
a)
clor
ofila
a*,
fico
cian
ina,
al
ofic
ocia
nina
, ß-c
arot
eno,
xan
tofi
las
grân
ulos
de
cian
ofi-
cina
, am
ido
das
cia-
nofí
ceas
(gl
icog
ênio
)
ause
nte
ause
nte
pept
idog
lican
o to
das
são
mic
rosc
ópic
as
P
rocl
orof
ícea
s cl
orof
ila a
e b
, ß-c
arot
eno,
xan
tofi
las
amid
o da
s ci
anof
í-ce
as (
glic
ogên
io)
ause
nte
ause
nte
pept
idog
lican
o to
das
são
mic
rosc
ópic
as
C
hlor
arac
hnio
phyt
a cl
orof
ila a
, clo
rofil
a b,
ß-c
arot
eno,
xa
ntof
ilas
glic
anos
ß1-
3 hi
dros
solú
veis
pr
esen
te
pres
ente
pr
esen
te**
* to
das
são
mic
rosc
ópic
as
G
lauc
ophy
ta
clor
ofila
a, f
icoc
iani
na,
alof
icoc
iani
na, f
icoe
ritr
ina,
ß-
caro
teno
, xan
tofi
las
amid
o pr
esen
te
ause
nte
celu
lose
(pa
rte
das
espé
cies
) to
das
são
mic
rosc
ópic
as
Eug
leno
phyt
a cl
orof
ila a
e b
, ß-c
arot
eno,
out
ros
caro
teno
s, x
anto
filas
pa
ram
ilo
pres
ente
au
sent
e pe
lícul
a fo
rmad
a po
r ba
n-da
s pr
otei
náce
as e
spir
alad
as
situ
ada
sob
a M
P#
toda
s sã
o m
icro
scóp
icas
Cry
ptop
hya
clor
ofila
a e
c, fi
coci
anin
a,
ficoe
ritr
ina,
alo
ficoc
iani
na, α
- e ß
-ca
rote
no, x
anto
fila
s
amid
o pr
esen
te
reco
nhec
ida
em a
lgum
as
espé
cies
peri
plas
to c
ompo
sto
de
plac
as p
rote
inác
eas
e/ou
es
cam
as a
ssoc
iado
à M
P
toda
s sã
o m
icro
scóp
icas
Pry
mne
siop
hyta
(=
Hap
toph
yta)
cl
orof
ila a
e c,
ß-c
arot
eno,
xan
tofil
as
cris
olam
inar
ina
pres
ente
pr
esen
te
célu
las
nuas
ou
reco
bert
as
com
esc
amas
de
CaC
O3
(coc
olit
ofor
ídeo
s)
toda
s sã
o m
icro
scóp
icas
Din
ophy
ta
(= P
irrh
ophy
ta)
clor
ofila
a e
c, ß
-car
oten
o, x
anto
filas
am
ido
pres
ente
pr
esen
te
plac
as c
elul
ósic
as e
m
vesí
cula
s ab
aixo
da
MP
ou
célu
las
nuas
toda
s sã
o m
icro
scóp
icas

49
Tabe
la 2
.4 C
arac
terí
stic
as m
orfo
lógi
cas,
quí
mic
as e
rep
rodu
tiva
s fu
ndam
enta
is d
os p
rinc
ipai
s gr
upos
mod
erno
s de
alg
as. M
odif
icad
o e
expa
ndid
o a
part
ir d
eG
raha
m &
Wilc
ox (
2000
) (c
onti
nuaç
ão).
Div
isão
/gru
po
Pri
ncip
ais
pigm
ento
s fo
toss
inté
tico
s Pr
odut
o de
res
erva
Fl
agel
o R
epro
duçã
o se
xuad
a C
ober
tura
cel
ular
D
imen
sões
Och
roph
yta
(dia
tom
ácea
s,
cris
ofíc
eas,
feof
ícea
s,
silic
ofla
gela
dos,
pi
ngui
ofíc
eas,
ra
fido
fíce
as, e
tc.)
clor
ofila
a e
c, ß
-car
oten
o, x
anto
fila
s cr
isol
amin
arin
a,
lipíd
eos
pres
ente
ao
men
os e
m
célu
las
repr
odut
i-va
s
pres
ente
al
gum
as s
ão n
uas,
out
ras
poss
uem
síli
ca o
u es
cam
as
orgâ
nica
s; ce
lulo
se e
al
gina
tos
estã
o pr
esen
tes
em
algu
ns g
rupo
s
mai
oria
das
es
péci
es é
m
icro
scóp
ica,
mas
nu
m g
rupo
(f
eofí
ceas
) tod
as a
s es
péci
es s
ão
mac
rosc
ópic
as
Eus
tigm
atof
ícea
s cl
orof
ila a
, ß-c
arot
eno,
xan
tofi
las
cris
olam
inar
ina
pres
ente
**
ause
nte
pres
ente
, mas
de
com
posi
ção
quím
ica
aind
a de
scon
heci
da
toda
s as
esp
écie
s sã
o m
icro
scóp
icas
R
hodo
phyt
a cl
orof
ila a
, fic
ocia
nina
, fic
oeri
trin
a,
alof
icoc
iani
na, ?
- e
ß-ca
rote
no,
xant
ofila
s
amid
o da
s flo
ríde
as
ause
nte
pres
ente
ce
lulo
se, p
olis
saca
ríde
os
sulfa
tado
s; a
lgum
as s
ão
calc
ifica
das
mai
oria
das
es
péci
es é
m
acro
scóp
ica
Chl
orop
hyta
cl
orof
ila a
e b
, ß-c
arot
eno,
out
ros
caro
teno
s, x
anto
fila
s am
ido
pres
ente
pr
esen
te
celu
lose
e o
utro
s po
límer
os,
esca
mas
em
alg
umas
; al
gum
as e
spéc
ies
são
nuas
, ou
tras
são
cal
cific
adas
mai
oria
das
es
péci
es é
m
icro
scóp
ica,
mas
em
alg
uns g
rupo
s to
das
as e
spéc
ies
são
mac
rosc
ópic
as
*Há
pelo
men
os u
ma
cian
obac
téri
a do
tada
de
clor
ofila
d: A
cary
ochl
oris
mar
ina
(Miy
ashi
ta e
t al.,
200
3).
**O
s fla
gelo
s es
tão
pres
ente
s ap
enas
nas
for
mas
veg
etat
ivas
de
algu
mas
esp
écie
s e
em z
oósp
oros
.**
*A m
aior
ia d
as e
spéc
ies
alte
rna
fase
s am
ebói
des
(sem
par
ede
celu
lar)
com
out
ras
cocó
ides
, cuj
a co
mpo
siçã
o qu
ímic
a da
par
ede
celu
lar
é de
scon
heci
da.
# M
embr
ana
plas
mát
ica.

50
CH OH2
a)O
O
CH OH2
O
O
CH OH2
O
O
CH OH2
O
O
CH OH2
b)O
O
CH OH2
O
O
CH OH2
O
O
CH OH2
O
O
Figura 2.8 Polissacarídeos de reserva apresentados por algas. (a) Amido, formado por moléculas de glicoseunidas por ligações glicosídicas do tipo α-1,4. (b) Estrutura básica de laminarina, crisolaminarina eparamilo, polissacarídeos formados por unidades de glicose unidas por ligações glicosídicas do tipo ß-1,3.O paramilo apresenta mais ramificações laterais que as outras duas substâncias; a laminarina apresentamanitol nas extremidades de parte das cadeias de glicose, característica que não ocorre na crisolaminarina.Reproduzido de Sze (1998).
Parte das espécies dos dinoflagelados, das criptomônadas e das euglenófitas apresentanutrição exclusivamente heterotrófica e, neste caso, não são consideradas fitoplanctônicas.Somente as espécies fotoautotróficas ou mixotróficas desses grupos são membros do fito-plâncton. Assim, por exemplo, os dinoflagelados fotoautotróficos Pyrocystis lunula eProrocentrum minimum são membros do fitoplâncton, mas os dinoflagelados heterotróficosPfiesteria piscicida e Noctiluca scintillans são membros do protozooplâncton. Esse exemploreforça a idéia de grande diversidade de formas e de exploração dos recursos disponíveis noambiente pelas algas microscópicas, demonstrando também de forma inequívoca que ostermos “alga” e “fitoplâncton” não têm valor taxonômico.
O fitoplâncton marinho é formado por um número de espécies ainda não determinadocom exatidão, pelo menos em parte em decorrência de limitações relativas à identificaçãode seus constituintes. Uma das poucas estimativas existentes é a de Sournia et al. (1991),que sugerem a existência de 5.000 espécies no fitoplâncton marinho. Apesar de respeitada,essa referência é também bastante conservadora. Stoermer & Smol (1999) estimam a existên-cia de pelo menos 10.000 espécies de diatomáceas, mas consideram também a possibilidadede haver mais de 100.000 espécies. Considerando-se que a maior parte das espécies conhe-cidas de diatomáceas tem hábitos planctônicos (principalmente em águas rasas) e ocorre nomar, mesmo sendo assumida a estimativa mais conservadora de Stoermer & Smol (1999)de 10.000 espécies, é natural admitir que haja mais de 5.000 espécies no fitoplânctonmarinho, distribuídas por diversos grupos taxonômicos. Táxons pequenos e de difícil iden-tificação, como muitas cianobactérias e flagelados fotossintetizantes, devem envolver grandenúmero de espécies a serem descritas.
Sendo o fitoplâncton constituído por algas e cianobactérias, pode-se assumir que suascaracterísticas estruturais básicas correspondem àquelas dos principais grupos de algas exis-tentes. Nesta obra são considerados dez grupos fundamentais de algas, cujas característicasbásicas são sumariadas na Tabela 2.4.
Uma descrição mais detalhada das características das algas que formam o fitoplânctoné apresentada a seguir.

51
2.2.1 DIVISÃO CHLOROXYBACTERIA (= CYANOPHYTA)As cianobactérias constituem um grupo bem definido de eubactérias, sendo as únicas
bactérias capazes de produzir oxigênio como produto colateral da fotossíntese. As ciano-bactérias são também chamadas de cloroxibactérias, cianoprocariontes, cianoclorontes, algasverde-azuladas ou cianofíceas. Clorofila a e diversos pigmentos acessórios de proteção eampliação da captação de luz (ficobilinas e carotenóides) estão presentes, associados a tilacói-des membranosos. As cianobactérias são encaradas como o grupo mais antigo de produtoresprimários fotossintetizantes. Há evidências fósseis atribuídas a organismos semelhantes a cia-nobactérias encontradas em rochas datadas de 3,46 bilhões de anos no oeste da Austrália(Schopf, 1993). Cianobactérias são também bastante abundantes em registros fósseis menosantigos, de 1 até 3 bilhões de anos. Acredita-se também que as cianobactérias tiveram papeldecisivo na transformação das características da atmosfera primitiva da Terra, pois, por meiode sua atividade fotossintética, o oxigênio (escasso nos primórdios do planeta) pôde ser acumu-lado lenta e paulatinamente na atmosfera. Esse processo de longo prazo proporcionou condiçõespara o surgimento de algas eucarióticas mais complexas, bem como de protistas heterotróficose animais, todos com maior demanda por oxigênio que organismos procariontes.
Dentre os organismos autotróficos, as cianobactérias são singulares por apresentaremorganização celular procariótica e pela ausência marcante de flagelos e da maioria das organelascelulares (mitocôndrias, plastos, núcleo, retículo endoplasmático, etc.). Os ribossomos estãopresentes e são do tipo 70S, característicos de bactérias, sendo menores do que os apresentadospor algas eucarióticas. Na região central da célula de cianobactérias (conhecida como centro-plasma) o material genético é organizado num único cromossomo circular (às vezes com cópias),sem histonas associadas, mas com íntrons presentes. Alças de ADN e plasmídios também podemestar presentes. Cianobactérias planctônicas podem formar vesículas de gás, com membranasdelimitando os espaços preenchidos por gases. Sistemas de túbulos dispersos pelo citoplasmaestão presentes e assemelham-se aos elementos de citoesqueleto típicos de células eucarióticas.
Juntamente com os ribossomos, os tilacóides são as únicas estruturas subcelularespresentes em cianobactérias que podem ser tratadas como organelas (Figura 2.9). Cadatilacóide compreende um espaço distinto do citoplasma, e os fotossistemas estão organizadosem torno de centros de reação contendo clorofila a. Quando presentes, ficobiliproteínascompõem estruturas granulares chamadas de ficobilissomos sobre as membranas externasde tilacóides. Cada ficobiliproteína consiste em várias ficobilinas firmemente ligadas a umaproteína. Quatro tipos de ficobiliproteínas ocorrem em cianobactérias: ficoeritrina, fico-cianina, aloficocianina e ficoeritrocianina. Todas as cianobactérias que contêm ficobilinaspossuem aloficocianina e ficocianina, sendo esta última a responsável pela coloração verde-azulada típica da maioria das espécies. Recentemente foi descoberta uma espécie de ciano-bactéria dotada de clorofila d, Acaryochloris marina (Miyashita et al., 2003), a qual possuihábitos bentônicos e comumente é encontrada associada a algas vermelhas (Murakami etal., 2004). Atualmente, tem se formado consenso de que a clorofila d eventualmente medidaem algumas algas vermelhas deva ser na realidade produzida por A. marina (Larkum & Kühl,2005) ou por outras possíveis espécies de cianobactérias ainda não conhecidas (ver o quadro“A intrigante presença de clorofila d em algas vermelhas”). A. marina apresenta altas con-centrações de clorofila d, seu principal pigmento fotossintético, mais importante que a própriaclorofila a, característica singular desse organismo (Kühl et al., 2005).

52
Bainha depolissacarídeo
Parede celularcom 4 camadas
Plasmalema
Tilacóide
Ficobilissoma
Ficobilissoma
Tilacóide
b) c)
0,02 µm
Vesículas de gás
Membranasdo tilacóide
Corpo poliédrico(carboxissomo)
Fibrilas de ADNno nucleoplasma
Ficoeritrina
Ficocianina
Aloficocianina
Grânulos decianoficina
Pilhas detilacóides
Fibrilas de ADN
Ficoeritrina
Ficocianina
Aloficocianina
Corpo poliédrico(carboxissomo)
d)
Membranado tilacóide
e)
0,5 µm
f)
Vacúolo(= tilacóide distendido)
0,5 µm
Parede celularcom 4 camadas
Plasmalema
Ribossomos
a)
Ribossomos
Figura 2.9 Características morfológicas de uma célula da cianobactéria Synechocystis (a) e da proclorofíceaProchloron (f ). Detalhes dos tilacóides, exibindo fileiras de ficobilissomos hemidiscoidais em corte trans-versal (b). Disposição dos ficobilissomos em corte longitudinal (c). Representação do arranjo tridimensionaldos constituintes moleculares de um ficobilissomo hemidiscoidal (d). Representação do arranjo tridimensionaldos constituintes moleculares de um ficobilissomo hemisférico (e). Modificado de Hoek et al. (1995).
O produto de reserva das cianobactérias é um polissacarídeo formado por monômerosde glicose unidos por ligações glicosídicas do tipo α-1,4, conhecido como amido das ciano-fíceas. Essa substância difere do amido por apresentar ramificações mais abundantes emrelação à cadeia principal de polissacarídeo. Em última análise, o amido das cianofíceas ébastante semelhante ao glicogênio encontrado em animais.
No citoplasma de cianobactérias são encontradas estruturas granulares associadas àacumulação de reservas: carboxissomos e grânulos, que podem ser constituídos de cianoficina,de amido das cianofíceas e de polifosfato. Os carboxissomos são também chamados de corpospoliédricos em razão da aparência geométrica que exibem. Eles apresentam diâmetro de 200-

53
300 nm e reservas da enzima ribulose-1,5-bisfosfatase-carboxilase-oxigenase (RuBisCO), quecatalisa a primeira etapa das reações no escuro da fotossíntese (ciclo de Calvin), conhecidacomo fixação fotossintética de CO
2. Os grânulos de cianoficina são estruturas levemente
angulares de até 500 nm de diâmetro, que podem ser vistos em microscopia óptica semcorantes específicos. Tais estruturas tendem a se acumular próximo à parede de células deespécies coloniais e filamentosas ou entre o centroplasma e o cromatoplasma. São constituídasfundamentalmente por proteínas decompostas rapidamente por ação de pepsinas. Trata-sede proteínas formadas por apenas dois ácidos aminados, arginina e asparagina, cuja funçãoé armazenar nitrogênio. Em situações de carência de nitrogênio, os grânulos de cianoficinapodem ser decompostos e o conteúdo de nitrogênio presente pode ser aproveitado para asdiversas funções metabólicas da célula. Grânulos de polifosfato têm papel equivalente aosgrânulos de cianoficina, porém servem para armazenar fósforo. Bactérias, fungos e algaseucarióticas também podem apresentar grânulos de polifosfato, mas não ocorrem em briófitase plantas vasculares. Os grânulos de amido das cianofíceas constituem a principal reservaenergética da célula. Eles possuem 30 a 65 nm de diâmetro, são bastante numerosos e ocorrementre tilacóides. As vesículas de ácido poli-â-hidroxibutírico, também de estrutura granular,ocorrem apenas em algumas cianobactérias e possuem cerca de 200 nm de diâmetro. Taisvesículas e produto de reserva são comuns em muitas bactérias heterotróficas.
A parede celular de cianobactérias é formada principalmente por peptidoglicano e ésemelhante àquela tipicamente apresentada por bactérias Gram-negativas. Há duas camadasna parede celular: uma rígida de peptidoglicano adjacente à membrana da célula e umaexterna de lipopolissacarídeos. A camada de peptidoglicano consiste em polímeros de N-acetilglucosamina e ácido N-acetilmurâmico com ácidos aminados inseridos (intercalados)em sua estrutura. Uma bainha de mucilagem freqüentemente se estende além da paredecelular, constituída essencialmente de carboidratos. A bainha de mucilagem geralmente éincolor, mas em algumas espécies pode assumir coloração amarelada ou marrom, nesses casos,provavelmente, exercem função de fotoproteção. Em diversas cianobactérias, a mucilagemproduzida causa deslizamento ou escorregamento quando as células estão em contato comum substrato rígido ou quando estão emaranhadas entre si (no caso de espécies que formamcadeias de células) na água, formando agregação macroscópica. Quando há grande númerode indivíduos nas agregações, estas podem assumir a forma de um biofilme colorido, conhe-cido popularmente como limo (diversas algas podem formar o chamado “limo”, que não éexclusividade de cianobactérias).
O número real de espécies de cianobactérias é desconhecido, especialmente porque aidentificação de espécies com base em características morfológicas é bastante limitada. Aprópria aplicação do conceito morfológico de espécie é limitada, dadas as grandes semelhan-ças morfológicas entre as espécies. Similarmente, a aplicação do conceito biológico de espéciea cianobactérias é inviável à luz dos conhecimentos disponíveis atualmente, pois não háregistro de reprodução sexuada no grupo. Na realidade, a aplicação do conceito biológicode espécie a microrganismos é em geral limitada, sobretudo para procariontes. As cia-nobactérias consistem num grupo em que o conceito filogenético de espécie tem sidoaplicado com mais freqüência. Apesar dessas limitações, reconhece-se a existência de 150gêneros e 2.000 espécies de cianobactérias na natureza (Hoek et al., 1995). Especula-se queessas estimativas sejam conservadoras, havendo maior número de gêneros e espécies. Ascianobactérias podem ocorrer como células solitárias (freqüentemente cocóides), em fila-mentos, agregações e colônias, e normalmente são envolvidas por bainha de mucilagem.Nas espécies filamentosas utiliza-se o termo tricoma para designar a série de células em si e

54
o termo filamento para fazer referência às células juntamente com a bainha de mucilagemque as envolve. Na reprodução, a divisão binária é o processo mais comum, sendo que ascélulas simplesmente se alongam e se dividem em duas células filhas. Algumas espécies podemse reproduzir por meio de esporos de dois tipos: exósporos e endósporos. Os primeirosresultam da realização de sucessivas divisões restritas na extremidade de uma célula, ao passoque os endósporos são formados por meio de múltiplas divisões dentro de uma célula, sendoliberados quando a parede celular se rompe. Espécies filamentosas podem se reproduzir pormeio de ruptura do filamento. Normalmente esse processo ocorre em partes terminais deum filamento, de maneira que a porção menor de filamento formada é chamada de hormo-gônio. A formação de hormogônios geralmente se dá quando uma célula do filamento morreou a parede entre células se separa. Os hormogônios comumente são diferenciáveis detricomas vegetativos pela análise da forma das células, por sua maior mobilidade e pelapresença de vesículas de gás.
As células de cianobactérias geralmente são pequenas, quase sempre menores que 2µm, o que determina a inclusão das cianobactérias no picoplâncton. Raríssimas são as espéciescom células livres grandes, como Chroococcus giganteus, que atingem 50-60 µm de diâmetro.Espécies que formam filamentos, entretanto, assumem dimensões maiores e são componentesdo nanoplâncton e do microplâncton. Embora as células dessas espécies também sejampequenas, o filamento (dotado de poucas unidades a dezenas de células) funciona como umaunidade, independentemente do tamanho de cada um de seus componentes.
Algumas cianobactérias filamentosas possuem células especializadas, chamadas deheterocitos e acinetos. Heterocitos (Figura 2.10) são especializados na fixação de nitrogêniomolecular (N2), sendo dotados de nitrogenases em grandes concentrações. N2 é a forma maisabundante de nitrogênio na natureza, porém, algas eucarióticas e plantas vasculares (alémda maioria das cianobactérias) não possuem aparato bioquímico capaz de romper a ligaçãoquímica tripla entre os átomos de nitrogênio. As espécies de cianobactérias com essa capa-cidade possuem vantagem competitiva relevante, podendo ocorrer em ambientes pobres emnitrogênio combinado. Mais do que o benefício próprio gerado, a incorporação de N
2 por
cianobactérias promove enriquecimento significativo de nitrogênio no ambiente, pois coma morte, consumo por protozoários e animais ou perda de matéria pelas espécies ocorre aentrada de mais nitrogênio na trama trófica aquática. Os heterocitos geralmente são maioresque as células vegetativas do filamento e possuem coloração diferente, freqüentementeamarelada. Nos heterocitos, o nitrogênio gasoso é convertido em amônia, que é incorporadaem ácidos aminados. Um deles, a glutamina, pode ser transportado para células vegetativasvizinhas, suprindo-as de nitrogênio. A nitrogenase, entretanto, é uma enzima inibida pelapresença de oxigênio, de forma que a atividade de fixação de nitrogênio, em princípio, éincompatível com a fotossíntese. Apesar disso, a fixação de N
2 por células de cianobactérias
não é feita com exclusividade pelos heterocitos, já tendo sido demonstrado que várias espéciesque não formam essas células especiais também fixam nitrogênio gasoso. Algumas espéciescocóides de cianobactérias, como as do gênero Gloeocapsa, que apresentam células solitáriasou agregadas em pequeno número, também são capazes de fixar nitrogênio (Wyatt & Silvey,1969; Matsui et al., 1986). Em casos como este, especula-se que deve haver algum mecanismopara proteger a nitrogenase do oxigênio gerado por meio da fotossíntese, criando um mi-croambiente de anaerobiose dentro da célula. Espécies atuais de cianobactérias que fixamN2 e não possuem heterocitos são interpretadas como “fósseis vivos” ou sobreviventes daera anterior à acumulação de oxigênio na atmosfera (Hoek et al., 1995). Alguns estudosdemonstraram que em condições de baixa disponibilidade de nitrogênio combinado noambiente, células vegetativas podem se diferenciar em heterocitos e desencadear o processo

55
de fixação de nitrogênio gasoso (Sze, 1998). Os heterocitos são conectados de forma tênuea células vegetativas vizinhas, sendo comum a ruptura do filamento junto a eles. Acinetos(Figura 2.11) são células de resistência formadas quando células vegetativas espessamfortemente suas paredes celulares e tornam-se dormentes. Acinetos geralmente são formadosquando, após período efêmero de crescimento intenso, predominam condições adversas aocrescimento. Eles geralmente são maiores que as células vegetativas e possuem um citoplasmagranular, com abundância de grânulos de cianoficina. Os acinetos podem permanecer inativospor muitos anos, havendo registros de acinetos de Anabaena viáveis após 64 anos de formação.Ao sofrerem germinação, os acinetos têm seu protoplasto liberado pela ruptura da paredeou através de um poro, deflagrando um processo de geração de célula e crescimento.
a) b) c)5 µm 10 µm10 µm 5 µm
Figura 2.11 Um acineto, indicado pela seta, em Cylindrospermum. Extraído de University of Wisconsin –Madison (2002).
As cianobactérias são comuns em virtualmente todos os ambientes aquáticos (dulcia-qüícolas, estuarinos e marinhos), bem como em ambientes terrestres úmidos; são ubíquas,mas tendem a ocorrer em maior biomassa em ambientes de água doce, como lagos, e mesmoem ambientes artificiais, como reservatórios. Diversas espécies, sobretudo as filamentosas,têm hábitos bentônicos e são capazes de formar biofilmes sobre sedimento e rochas, sendocomponentes importantes do chamado microfitobentos. Biofilmes formados por filamentosintrincados de cianobactérias podem agregar outros microrganismos (bactérias, protozoários,microalgas eucarióticas, fungos), sedimento e partículas minerais diversas, com conseqüenteaumento gradativo de volume ao longo do tempo. Formações desse tipo, conhecidas comoestromatólitos, assumem formato de esfera ou de coluna em águas costeiras rasas, decorrentesdo crescimento orientado pela luz. Vários estromatólitos sofreram fossilização e muitos delesforam datados como possuindo mais de três bilhões de anos. Estromatólitos eram muitoabundantes no passado remoto da Terra, presumivelmente porque inicialmente não havia
Figura 2.10 Diferenças morfológicas dos hetoro-citos (indicados pelas setas) das cianobactériasAnabaena (a), Scytonema (b) e Aulosira (c). Repro-duzido de Graham & Wilcox (2000).

56
organismos para consumi-los. O processo evolutivo global das espécies, influenciado pelaprópria atividade das cianobactérias, levou ao surgimento de novas formas de vida quepassaram a consumir os estromatólitos. Atualmente, estromatólitos em desenvolvimento sãoencontrados somente em locais quentes e inóspitos a organismos pastadores e perfurantes,como Shark Bay (oeste da Austrália), ambiente hipersalino; há também registros dessasformações especiais nas proximidades da ilha Exuma (Bahamas), onde correntes de maréfortes inibem os consumidores de estromatólitos.
Em outros casos, os filamentos intrincados de cianobactérias ocorrem de forma flu-tuante em razão da presença de grande quantidade de mucilagem e vesículas de gás, podendoser facilmente coletados na superfície da água. No mar, as cianobactérias planctônicas sãomais abundantes em áreas tropicais e temperadas afastadas da costa, as quais são tipicamenteoligotróficas. Em áreas costeiras de ambientes tropicais e temperados e em regiões de altaslatitudes as cianobactérias estão presentes em densidades mais baixas, ocorrendo abundânciade outros grupos de produtores primários, como as diatomáceas.
Alguns membros do grupo possuem também clorofila b e são conhecidos como proclo-rofíceas. A descoberta das proclorofíceas (Lewin & Withers, 1975) causou enorme furorcientífico, pois inicialmente tais bactérias foram encaradas como derivadas de um possívelancestral primitivo de algas verdes (clorófitas), originado a partir de cianobactérias. A presençade clorofila b foi considerada característica marcante o suficiente para a criação de uma divisãoà parte para acomodar as proclorofíceas, a chamada divisão Prochlorophyta. Entretanto,análises moleculares, bioquímicas, morfológicas e filogenéticas posteriores indicaram queas proclorofíceas não estão relacionadas às algas verdes, conforme se pensava; mais do queisto, as pesquisas indicam claramente que as proclorofíceas constituem um grupo especialde cianobactérias. As proclorofíceas são desprovidas de ficobilinas como pigmentos acessóriosda fotossíntese. Uma das implicações dessas pesquisas foi o reconhecimento de que a clorofilab pode ter surgido na natureza mais de uma vez, em linhagens evolutivas distintas e inde-pendentes. Tendo por base principalmente os pigmentos fotossintéticos, as cianobactériaspodem ser divididas em dois grandes grupos, conforme apresentado na Tabela 2.5.
Tabela 2.5 Comparação de cianobactérias típicas e proclorofíceas. Modificado de Sze (1998).
Cianobactérias típicas Proclorofíceas
Pigmentos fotossintéticos principais Clorofila a, ficobiliproteínas Clorofila a, bReserva de carboidrato Amido das cianofíceas Amido das cianofíceasOrganização de tilacóides Separados PareadosCobertura celular Parede de peptidoglicano Parede de peptidoglicano
Há apenas três gêneros conhecidos de proclorofíceas. Prochloron e Prochlorococcus são célulasesféricas, não formam filamentos e não apresentam bainha de mucilagem. Ambos ocorrem apenasno mar, sendo que Prochloron ocorre associado simbioticamente a tunicados e Prochlorococcusé amplamente distribuído no plâncton, especialmente em águas oceânicas oligotróficas.Prochlorotrix é formado por filamentos sem heterocitos, com vesículas de gás nas células, eocorre apenas em água doce.
As cianobactérias são dotadas de extraordinária capacidade de se adaptar com sucessoa alterações ambientais. Em geral, elas toleram grandes flutuações de temperatura, salinidade,pH e disponibilidade de nutrientes, por exemplo, bem maiores do que a maioria das espéciesde algas microscópicas eucarióticas. Possivelmente, sua enorme tolerância a variações am-

57
bientais está relacionada à condição de procarionte e à simplicidade de suas células. Essacapacidade é fundamental para a compreensão de seu papel como organismos pioneiros emambientes recém-formados, bem como sua presença ubíqua na natureza. Como exemplosda tolerância de cianobactérias a ambientes extremos, espécies do gênero Synechococcus (célulascocóides) são capazes de tolerar temperaturas de até 74°C, ao passo que espécies dos gênerosPhormidium e Lyngbya (filamentosos) formam biofilmes no fundo de lagos antárticos perma-nentemente cobertos por gelo, com reduzidíssima penetração de luz mesmo no verão. NoParque Nacional de Yellowstone (EUA), famoso pela ocorrência de gêiseres, diversas ciano-bactérias ocorrem na água empoçada quente, ácida e plena de derivados de enxofre em altasconcentrações (tóxicas para a grande maioria dos organismos), formando mosaicos coloridos.Populações naturais da cianobactéria Arthrospira (Figura 8.1) ocorrem abundantemente emsistemas lacustres fortemente alcalinos (pH > 10,0) da África e do México, nos quais poucasespécies conseguem sobreviver.
Algumas cianobactérias planctônicas são capazes de formar florações intensas em corposde água, alterando drástica e subitamente o aspecto e as características do ambiente. Taismudanças são geralmente efêmeras, sendo possível o retorno às características originais dolocal antes do evento de floração. No mar, florações de cianobactérias são comuns emambientes oligotróficos, ao passo que em águas continentais o fenômeno geralmente ocorreem áreas eutróficas ou em locais associados a enriquecimento recente por nutrientes. Alémdisso, várias espécies de cianobactérias são capazes de produzir toxinas de diversas classes eações, com efeitos nocivos relevantes para o homem. Toxinas classificadas como hepa-totoxinas, neurotoxinas e irritantes por contato estão entre aquelas produzidas por ciano-bactérias. Já foram registrados acidentes fatais com humanos como resultado da ação detoxinas de cianobactérias. O caso mais famoso e trágico ocorreu no Brasil, em 1996, quando56 pacientes de uma clínica de hemodiálise de Caruaru, PE, morreram em função da expo-sição à água não tratada e com altas concentrações de microcistina (Figura 2.12), umahepatotoxina produzida por Microcystis (Jochimsen et al., 1998).
MeO
MeMe
Me
HN
O
NHN
Me O
OO
CO H2
CO H2
Me
Me
O
O
Me
HN
NH
HN NH2
O
HN
HN
Me
N
Figura 2.12 Molécula da microcistina, uma hepatotoxina produzida por cianobactérias.
Assume-se genericamente em evolução que longos períodos de tempo seriam favoráveisao processo de especiação, culminando na formação de maior número de espécies numdeterminado grupo. Entretanto, a enorme importância das cianobactérias ao longo de erasgeológicas e a ampla presença das mesmas em inúmeros ambientes contrastam com umreduzido número de espécies reconhecidas atualmente. Esse fato apresenta três vertentes

58
possíveis de interpretação: i) o número real de espécies é maior do que o reconhecido, emfunção de limitações no processo de identificação das espécies, conforme já mencionado nestetexto; ii) a ausência de reprodução sexuada no grupo tornaria a velocidade de aquisição decaracterísticas novas (e daí a especiação) bem lenta, acarretando o baixo número de espéciesexistente; iii) a extraordinária plasticidade adaptativa das cianobactérias as levaria a sofrerpoucas mudanças genéticas ao longo do tempo, contribuindo para o surgimento de pouquíssi-mas espécies novas durante o processo evolutivo do grupo. Embora os três argumentos sejamplausíveis, não há dados conclusivos para determinar a veracidade de cada um deles. Alémdisso, torna-se natural assumir que os três argumentos não sejam excludentes e possam, aomenos em parte, contribuir para a compreensão da diversidade conhecida de espécies decianobactérias.
Muitas cianobactérias são extremamente bem-sucedidas no estabelecimento de relaçõessimbióticas estáveis com outros organismos, fornecendo material orgânico, oxigênio e/ousubstâncias nitrogenadas a seus hospedeiros. A Tabela 2.6 indica uma lista de organismosque apresentam cianobactérias como simbiontes. Uma das associações mais bem estudadasenvolve a planta vascular sem semente Azolla e a cianobactéria Anabaena azollae. Azolla crescesem se fixar a um substrato, flutuando na superfície da água de lagos e pântanos. Cavidadesespeciais nas folhas da planta contêm filamentos de A. azollae com elevada proporção deheterocitos em relação a células vegetativas, comparadas com a proporção típica apresentadapor formas livres da cianobactéria. A planta utiliza a maior parte da amônia produzida porAnabaena. Azolla estimula a fixação contínua de nitrogênio molecular, inibindo a enzimaglutamina sintetase, de forma a impedir que a cianobactéria utilize a amônia gerada comoresultado da redução do nitrogênio molecular. A planta torna-se bastante enriquecida comnitrogênio e após sua morte, ou perda de partes (como folhas), promove significativo enrique-cimento de nitrogênio combinado no meio. Na Ásia, Azolla é amplamente utilizada emcampos de produção de arroz como fertilizante natural (Sze, 1998). Outro exemplo muitoimportante é o de algas glaucófitas, existentes apenas em ambientes de água doce. Tais algasdispõem de uma a várias cianobactérias endossimbióticas chamadas cianelas, envoltas poruma membrana. As cianelas atuam como plastos e fornecem material orgânico a seus hospe-deiros. Elas apresentam parede de peptidoglicano e outras estruturas encontradas em ciano-bactérias de vida livre, como carboxissomos e tilacóides. Contudo, seu material genético ébastante reduzido (semelhante em tamanho ao material genético de plastos) se comparadoao de cianobactérias de vida livre, e as cianelas não sobrevivem se removidas de seus hospe-deiros. Acredita-se que as cianelas das glaucófitas representam estádio intermediário naevolução de cloroplastos e constituem-se em evidência viva para os defensores da teoriaendossimbiótica de origem e evolução de plastos.
Conforme mencionado anteriormente, muitos autores acreditam que os plastos de algaseucarióticas sejam derivados direta ou indiretamente de cianobactérias. A ocorrência de duasmembranas associadas, envolvendo os plastos de algas verdes e vermelhas, sugere que os plastosoriginaram-se diretamente de cianobactérias simbióticas. Nesse campo, inclusive, evidênciasmoleculares mostram que os plastos de algas verdes são mais semelhantes a cianobactériastípicas do que a proclorofíceas, enfraquecendo a hipótese de origem de algas verdes a partirde proclorofíceas. As membranas adicionais que envolvem plastos de outros grupos de algassugerem a origem a partir de um evento de endossimbiose secundária ou terciária, no qualo endossimbionte teria sido uma alga eucariótica (Sze, 1998).

59
Tabela 2.6 Exemplos de associações simbióticas envolvendo cianobactérias. Modificado e expandido apartir de Sze (1998).
Hospedeiro Cianobactéria simbionte
Diatomáceas planctônicas (Rhizosolenia, Hemiaulus) RicheliaAlgas glaucófitas CianelasEsponjas Aphanocapsa, PhormidiumTunicados ProchloronPêlos de urso-polar (Ursus maritimus) Aphanocapsa ou GloeocapsaFungos (líquens) Nostoc, Calothrix e outrosGeosiphon (fungo de solo) NostocBriófitas Anabaena e NostocAzolla (planta vascular sem semente) AnabaenaRaízes de cicadáceas (gimnospermas) NostocRaízes de plantas de mangue SynechococcusGlândulas de Gunnera (angiosperma) Nostoc
A sistemática de cianobactérias é bastante controvertida. Historicamente podem seridentificadas duas vertentes principais de classificação e organização dos grupos. Uma vertenteé mais antiga, tendo por base a escola botânica e os estudos de Geitler (1932); a segundavertente é mais recente, valoriza informações sobre fisiologia, cultivo e bioquímica dasespécies, sendo ligada a práticas típicas de microbiologia e mais relacionada à sistemática debactérias proposta por Stanier et al. (1978). A convergência de abordagem entre escolasdistintas nem sempre ocorre, gerando sistemas de classificação eventualmente muito dife-rentes. A classificação apresentada na Tabela 2.7 é baseada em Sze (1998), que reconhece aexistência de apenas uma classe e cinco ordens no grupo. Essa proposta não corresponde deforma alguma a um consenso entre os autores, mas seguramente é válida para mostrar relaçõesentre os principais grupos de cianobactérias.
Tabela 2.7 Classificação de cianobactérias de acordo com Sze (1998).
Divisão Cyanophyta, classe Cyanophyceae
Ordem Chroococales
Células solitárias, agregações e colônias, não realizando reprodução por esporos
Ordem Chamaesiphonales
Células solitárias, agregações e filamentos curtos, realizando reprodução por esporos
Ordem Oscillatoriales
Filamentos não ramificados e sem células especializadas
Ordem Nostocales
Filamentos não ramificados, com heterocitos e acinetos
Ordem Stigonematales
Filamentos ramificados (às vezes multisseriados), com heterocitos e acinetos
Os gêneros Aphanothece, Chroococcus, Cyanocystis, Gloeocapsa, Gloeothrichia,Merismopedia, Microcystis, Synechococcus, Synechocystis (cocóides), Anabaena, Aphanizomenon(Figura 8.25), Arthrospira (Figura 8.1), Cylindrospermopsis, Cylindrospermum, Lyngbia,Nodularia, Nostoc, Oscillatoria, Phormidium, Spirulina e Trichodesmium (filamentosos) são

60
alguns dos mais comuns dentre as cianobactérias. Prochloron, Prochlorococcus e Prochlorotrixsão os únicos gêneros conhecidos de proclorofíceas.
2.2.2 DIVISÃO CHLORARACHNIOPHYTA
As cloraracniófitas formam uma divisão pequena, com apenas cinco gêneros (quatrodeles são monoespecíficos) e oito espécies, todas marinhas. A divisão foi criada para acomodaruma espécie amebóide verde, Chlorarachnion reptans (Figura 2.13), cujo posicionamentonão se mostrava adequado em outros grupos de algas. Originalmente, C. reptans, descobertanas ilhas Canárias, Espanha, foi classificada como uma xantofícea (Geitler, 1930). A redes-coberta da espécie no México (Norris, 1967), bem como a descrição de outras espéciessemelhantes, tornou a criação da divisão ainda mais necessária. A seguinte combinação decaracterísticas torna o grupo singular: (i) presença de clorofila a e b; (ii) quatro membranascobrindo o plasto; (iii) presença de um nucleomorfo em cada compartimento periplastidial;(iv) zoósporos com um flagelo simples; (v) produtos de reserva não amiláceos; (vi) ausênciade mancha ocelar (Ota et al., 2005). O nome do gênero tipo do grupo (Chlorarachnion,significa “teia verde de aranha”, numa alusão à típica formação de colônias reticuladas sobreo substrato.
10 µma) b)10 µm
Figura 2.13 A cloraracniófita Chlorarachnion vista através de microscopia de campo claro (a) e de contrastede fase (b). Reproduzido de Graham & Wilcox (2000).
O interesse pelas cloraracniófitas em grande parte resulta das evidências que indicamque o grupo é derivado de um evento de endossimbiose secundária envolvendo um organismoamebóide (uma tecameba com pseudópodes) e uma alga verde. Análises moleculares confir-mam o caráter monofilético do grupo (Reviers, 2006). Os representantes modernos do grupoapresentam células vegetativas tipicamente amebóides, como supõe-se que seriam as célulasdo hospedeiro ancestral que estabeleceu relação endossimbiótica. Porém, todas as espéciesdo grupo alternam uma fase cocóide ou monadal com a fase amebóide. Aparentemente, aúnica exceção é Lotharella globosa, da qual se conhece apenas a fase cocóide. A duração dosestádios amebóide e cocóide/monadal varia entre as espécies, mas para a maioria delas a faseamebóide é mais duradoura. Apenas em Bigelowiella natans a fase monadal é mais duradouraque a fase amebóide. Este é o único grupo de organismos fotossintetizantes que contêm umplasto dotado conjuntamente de clorofila b e nucleomorfo, reforçando a hipótese da endossim-biose secundária. Os plastos são numerosos e envolvidos por quatro membranas, sendo a maisexterna desprovida de ribossomos. Muitos plastos ocorrem em células de cloraracniófitas e cadaum deles contém um pirenóide grande. Os tilacóides são empilhados em números variáveis,mas freqüentemente em três, formando lamelas. Além das clorofilas a e b, os plastos são dotadosde ß-caroteno, neoxantina, violaxantina, luteína e dodecanato de loroxantina.

61
O produto de reserva é um polissacarídeo derivado de moléculas de glicose unidas porligações glicosídicas do tipo ß1-3. São produtos hidrossolúveis, semelhantes àqueles produ-zidos por euglenófitas e ocrófitas, e que não coram em presença de iodo. Os produtos dereserva são acumulados em vesículas no citoplasma, formadas pelo aparato de Golgi.
Nas células flageladas ocorre apenas um único flagelo, inserido lateralmente. Há umúnico núcleo, cuja posição pode ser central ou periférica. Dictiossomos e mitocôndrias sãonumerosos nas espécies do grupo. Organelas semelhantes a tricocistos já foram encontradasem Chlorarachnion reptans, mas sua natureza e ação ainda não foram suficientemente estu-dadas (Reviers, 2006). Paredes celulares podem ser formadas em alguns estádios transitóriose em cistos de repouso, mas sua natureza química ainda não foi elucidada.
Não são conhecidos fósseis do grupo, pois a conservação das células certamente é muitodifícil. A reprodução do grupo dá-se por divisão binária das células e pela formação dezoósporos dotados de apenas um flagelo. Os zoósporos são formados em tétrades, grupos dequatro células imóveis dotadas de parede celular e flagelo. Reprodução sexuada já foi documen-tada em Chlorarachnion reptans e Cryptochlora perforans. Em C. reptans o gameta masculino éamebóide e o feminino é cocóide; em C. perforans os dois gametas são amebóides (Reviers,2006). Não há detalhes sobre a ploidia das fases que se alternam nem sobre a meiose praticadapelas espécies do grupo. Não são conhecidos efeitos nocivos de cloraracniófitas à biota ou aohomem.
As espécies conhecidas da divisão Chlorarachniophyta são: Bigelowiella natans,Chlorarachnion reptans, Cryptochlora perforans, Gymnochlora stellata, Lotharella amoeboformis,L. globosa, L. polymorpha e L. vacuolata. As espécies são amebóides ou cocóides encontradasno bentos, exceto B. natans, que apresenta hábito planctônico e suas células vegetativasapresentam um flagelo. Detalhes da ultra-estrutura do pirenóide e a posição do nucleomorfono compartimento periplastidial são importantes para a identificação das espécies bentônicasdo grupo. Ota et al. (2005) sugerem a existência de diversas espécies não descritas cultivadasem coleções de culturas de microalgas (vide Capítulo 7), a exemplo da cepa CCMP240(mantida pelo Provasoli-Guillard Center for Culture of Marine Phytoplankton, EUA), cujoestudo resultou na descrição da oitava espécie do grupo, L. vacuolata.
2.2.3 DIVISÃO GLAUCOPHYTA
Embora não haja representantes desta divisão no ambiente marinho, uma breve des-crição de suas características é importante para situar esse grupo em meio aos demais, queenvolvem algas marinhas e dulciaqüícolas.
As glaucófitas são algas raras, planctônicas ou bentônicas, podendo ocorrer comoepífitas de outras algas, musgos e angiospermas. Os membros da divisão Glaucophyta (Gr.glaukos, azulado-esverdeado + Gr. phyton, planta) são eucariotos que possuem plastos dotadosde pigmentos azul-esverdeados, conhecidos como cianelas (Figura 2.14). Esses plastos asse-melham-se a cianobactérias cocóides de várias formas, incluindo a ocorrência de paredesformadas por peptidoglucano. As cianelas/plastos das glaucófitas ocorrem no interior devacúolos e possuem clorofila a, ficobilinas e carotenóides. Sua coloração é verde-azulada,como efeito da presença de ficocianina e aloficocianina, mascarando a clorofila existente.Os tilacóides são separados uns dos outros, como nas cianobactérias típicas, e não ocorremempilhados, como em proclorofíceas e na maioria das algas eucarióticas (Figura 2.14). Assimcomo nas cianobactérias típicas e nas algas vermelhas, os pigmentos acessórios da fotossínteseocorrem organizados em ficobilissomos dispostos sobre a membrana dos tilacóides. Oscarotenóides presentes incluem o â-caroteno, a zeaxantina e a â-criptoxantina. Há material

62
genético no plasto/cianela, o qual se dispõe no centro da organela. Além disso, o plasto/cianelatambém dispõe de carboxissomos na sua porção central, dotados da enzima RuBisCO, quecatalisa a fixação fotossintética de CO
2 em ribulose 1,5-bisfosfato.
Vacúolopulsátil
Filopódio
Envelope emforma de urna
Cianela
Núcleo
a) b)
10
µm
Membrana do vacúoloque contém a cianela
Parede de peptidoglicano
Carboxissomos
Tilacóides
1µ
m
0,1 µm
Cianela
10 µm
Tilacóides
c)
Figura 2.14 As glaucófitas Cyanophora paradoxa (a) e Paulinella chromatophora (b). Detalhes da morfologiada cianela de P. chromatophora (c) vistos em corte transversal. Modificado de Hoek et al. (1995).
As características especiais das cianelas tornam as glaucófitas extremamente importantesno estudo da evolução das algas e da própria evolução dos seres fotossintetizantes, uma vezque são consideradas por muitos autores exemplos modernos do processo de aquisição deendossimbiose estável. Parcela importante dos estudos sobre endossimbiose em algas foirealizada com o gênero Cyanophora.
O polissacarídeo de reserva produzido pelas glaucófitas é o amido, que é acumuladoem grânulos fora do plasto/cianela, a exemplo do que ocorre nas algas vermelhas. Nas algasverdes, briófitas e plantas vasculares, o amido é formado e acumulado dentro do cloroplasto.
As células flageladas apresentam dois flagelos desiguais em tamanho, que se inserem numadepressão próxima ao ápice da célula. Os flagelos apresentam o arranjo típico de ultra-estrutura“9 + 2” (nove pares de microtúbulos periféricos circundando um par central), encontrado emquase todas as células eucarióticas, e duas filas de pêlos delicados; os flagelos das glaucófitas,entretanto, são inteiramente diferentes dos pêlos tubulares rígidos de outras algas, conhecidos

63
como mastigonemas, como os apresentados por algas criptófitas e ocrófitas. As formas vege-tativas não flageladas (células isoladas ou colônias) produzem células reprodutivas assexuadasflageladas (zoósporos). Não se conhece reprodução sexuada neste grupo de algas.
A divisão é formada por apenas três gêneros e 14 espécies: Cyanophora (com três espécies),Glaucocystis (com dez espécies) e Gloeochaete (com uma única espécie) (Reviers, 2006). Algunsautores reconhecem a existência de um quarto gênero na divisão (Glaucosphaera), constituídopor células não flageladas. Estudos recentes sugerem que esse gênero, na realidade, deve seralocado na divisão Rhodophyta, ordem Porphyridiales.
2.2.4 DIVISÃO EUGLENOPHYTA
A divisão Euglenophyta (Gr. eu, bom, verdadeiro ou primitivo + Gr. glene, olho + Gr.phyton, planta) inclui os chamados flagelados euglenóides, os quais ocorrem como célulaslivres ou em formas coloniais. A imensa maioria ocorre como mônadas (Figura 2.15), masalgumas espécies apresentam estádios durante os quais são aglomeradas dentro de uma cápsulade mucilagem. Há cerca de 40 gêneros e 900 espécies (650 a 1.050 espécies, dependendodos autores consultados), 2/3 das quais são heterotróficas (praticando saprotrofia ou fago-trofia), dotadas de plastos incolores, e há também espécies desprovidas de plastos. Dentreos gêneros e espécies restantes (1/3 do total), ocorrem as euglenofíceas dotadas de plastospigmentados, com clorofila a, clorofila b e carotenóides, muitos dos quais idênticos aos dealgas verdes. Os carotenóides mais importantes presentes são â-caroteno, neoxantina, dia-dinoxantina, equinenona, diatoxantina e zeaxantina. Os pigmentos acessórios não mascarama cor da clorofila, o que confere coloração verde às espécies fotossintetizantes. As espécies capazesde realizar fotossíntese são em sua imensa maioria mixotróficas. Os tilacóides se agrupam emtrês, formando lamelas, um arranjo também encontrado em dinoflagelados e em algas hetero-contófitas. Os plastos de euglenófitas apresentam envelope constituído por três membranas enunca são conectados ao núcleo através do retículo endoplasmático. O ADN do plasto ocorrecomo uma mistura de grânulos pequenos, espalhados por toda a organela.
Reservatório
Vacúolocontrátil
Plasto
Paramilo
Estigma
Núcleo
E
Py
P
a)
b)
Figura 2.15 Ilustrações das feições mor-fológicas típicas de euglenófitas e suasestruturas celulares. E = estigma; Py =pirenóide; P = plasto; N = núcleo. (a)Segundo Gojdics (1953); (b) segundoWalne & Arnott (1967).

64
Em vez de parede celular, há uma película associada a microtúbulos constituída defaixas de proteínas abaixo da membrana plasmática, a qual confere flexibilidade e resistênciaà célula. As faixas de natureza protéica formam espirais em torno da célula e têm capacidadede contração e distensão. Muitas euglenófitas têm forma plástica que se altera com seusmovimentos. À medida que a célula se movimenta, as faixas que compõem a película escorre-gam umas sobre as outras, contraem-se e distendem-se, promovendo as mencionadas mu-danças na forma das células que configuram os chamados movimentos euglenóides oumetabolia. Algumas euglenófitas (por exemplo, Menodium, Rhobdomonas) têm películasrígidas e outras (por exemplo, Ascoglena, Strombomonas, Trachelomonas) têm uma lórica queenvolve a célula. Quase todas as espécies apresentam uma fina camada de mucilagem, a qualé secretada por corpos de mucilagem ou mucocistos, derivados do complexo de Golgi,situados abaixo da película. A produção intensa de mucilagem pode resultar na formaçãode estádios palmelóides.
Os flagelos emergem de uma invaginação especial denominada ampulla, ampola oucitofaringe, localizada na extremidade anterior da célula, consistindo em um canal e umreservatório. Quase sempre há dois flagelos, mas um deles é curto e não emerge da ampulla,aparentando, portanto, tratar-se de um único flagelo. Assim, além dos movimentos euglenói-des, as euglenófitas podem também se movimentar pela ação de seus flagelos. O flagelo queemerge mostra-se espesso em observações por microscopia óptica, em razão da ocorrênciade uma haste paraflagelar, que se situa num dos lados do axonema. Os flagelos são dotadosde um arranjo unilateral de pêlos delicados e curtos (2-3 µm de comprimento), tendo dooutro lado um segundo arranjo de cerdas ainda mais curtas e finas. Junto ao reservatórioocorre um estigma alaranjado ou avermelhado, pigmentado pela presença de gotículas decarotenóides. Estigmas semelhantes aos das euglenófitas (livres no citoplasma) são encontradassomente em eustigmatofíceas e dinoflagelados, pois nas demais algas elas ocorrem nos plastos.Em espécies de água doce ocorre também um vacúolo contrátil, na porção anterior da célula,que lança seus conteúdos dentro do reservatório e que provavelmente tem papel importantena osmorregulação da célula. O vacúolo contrátil é rodeado por vacúolos menores, acessóriosque também se intumescem de líquidos e lançam seus conteúdos no reservatório.
O produto de reserva é um polissacarídeo derivado de glicose, formado por ligaçõesglicosídicas do tipo ß-1,3, o paramilo, que ocorre em grânulos no citoplasma de espéciespigmentadas e mesmo em espécies não pigmentadas dotadas de plastos. O paramilo não coraem presença de lugol e forma estruturas em formato de anel. O paramilo é um polissacarídeosemelhante à crisolaminarina, porém, a molécula apresenta mais ramificações laterais dacadeia linear principal e é insolúvel. Se pirenóides estão presentes, eles se projetam para forado plasto e o paramilo pode ser visto associado a eles. Gotículas de gordura também podemser encontradas nas espécies do grupo, constituindo reserva energética secundária. Inicial-mente acreditava-se que o paramilo era uma substância exclusiva de euglenófitas, até queKreger & van der Veer (1970) demonstraram a presença de paramilo em grânulos na algaprimnesiófita Pavlova mesolychnon.
A mitocôndria das euglenófitas apresenta algumas peculiaridades: são grandes (0,5-10 µm de comprimento) e às vezes são conectadas umas às outras formando redes. As mito-côndrias são maiores e mais interconectadas em espécies heterotróficas do que naquelas quepraticam fotossíntese. As membranas externas das mitocôndrias são particularmente bastanteonduladas e as internas apresentam cristas bem desenvolvidas em forma de discos.
O núcleo interfásico apresenta cromossomos condensados, os quais podem ser vistosem células vivas. A mitose é fechada (o envelope nuclear se mantém intacto) e o nucléolo

65
persiste durante a divisão celular. Não se conhece reprodução sexuada em euglenófitas,embora em Scytomonas já tenha sido verificada a ocorrência de fusão de células que, assim,poderia representar uma forma de reprodução sexuada por isogamia (Reviers, 2006). Naausência de mais informações e detalhes sobre a sexualidade das euglenófitas, assume-se queas células se reproduzem meramente por divisão binária, que se inicia com a duplicação doreservatório e dos flagelos.
A maioria das espécies conhecidas ocorre em ambientes dulciaqüícolas, embora diversasespécies marinhas e de ambientes salobros também sejam conhecidas. Euglenófitas tambémsão encontradas em solos úmidos e formas parasitas de copépodes e de batráquios são reconhe-cidas. Algumas espécies de euglenófitas pigmentadas têm hábito bentônico e são capazes dealterar visivelmente o sedimento sobre o qual ocorrem, quando geram aglomerações densas –nesses casos, o sedimento geralmente torna-se esverdeado. Tais espécies são constituintes dochamado microfitobentos, juntamente com outros grupos de algas, como as diatomáceas e ascianobactérias, tendo importância relevante na ecologia de praias, estuários, pântanos e lagos.
Especula-se que esse grupo seja antigo, pois há registros fósseis atribuídos a ele emdepósitos de 410 a 460 milhões de anos. As análises filogenéticas modernas indicam que aseuglenófitas estão relacionadas evolutivamente aos kinetoplastídeos, formando um cladobem definido (Brusca & Brusca, 2003). Os kinetoplastídeos constituem um grupo deprotistas heterotróficos que envolvem diversas formas de parasitas que afetam o homem, comoo gênero Trypanosoma (causador da doença de Chagas e da doença do sono) e Leishmannia(causador da leishmaniose). O grupo envolve também formas livres que consomem algasmicroscópicas, como o gênero Bodo, comum em sistemas de cultivo de microalgas de largaescala (ver Capítulo 8). As relações das euglenófitas com os protozoários kinetoplastídeosconstituem mais uma evidência da inexistência de valor taxonômico do termo “alga”, confor-me discutido no início deste capítulo.
A divisão Euglenophyta é constituída por apenas uma classe, Euglenophyceae. Esta édividida em seis ordens, fundamentalmente com base na morfologia das células: Euglenales,Eutreptiales, Euglenamorphales, Heteronematales, Rhabdomonadales e Sphenomonadales.As três primeiras ordens apresentam representantes fotossintetizantes e não fotossintetizantes;as três últimas ordens são constituídas apenas por representantes heterotróficos. Diversosestudos independentes de sistemática molecular, com base na análise das seqüências do ARNr18S, sugerem que as euglenófitas fotossintetizantes foram originadas por endossimbiosesecundária entre uma alga verde e formas incolores heterotróficas desprovidas de plastos,como Petalomonas e Pseudoperanema (Reviers, 2006). Essa interpretação é conveniente econtribui para o entendimento da constatação de que a maioria das euglenófitas não éfotossintetizante, uma vez que o processo citado de endossimbiose secundária envolveu apenasparte das espécies primitivas do grupo.
Os gêneros Euglena, Eutreptia, Phacus, Trachelomonas (formas pigmentadas), Astasia,Colacium, Peranema e Petalomonas (formas incolores) são alguns dos representantes maiscomuns da divisão Euglenophyta. Espécies do gênero Eutreptia ocorrem em água doce e nomar e são capazes de formar florações ocasionais. Colacium é um gênero bentônico e colonialque cresce principalmente como epizoonte do cladócero Daphnia em ambientes de água doce.
2.2.5 DIVISÃO CRYPTOPHYTA
Esta divisão compreende número inexato de flagelados unicelulares, conhecidos comocriptomônadas ou criptófitas, que significam literalmente “células flageladas livres escondidas”ou “planta escondida”, respectivamente. O grupo vem recebendo essas designações por sua

66
presença discreta (muitas vezes imperceptível) em amostras naturais. São reconhecidos de 12a 23 gêneros, de acordo com a fonte consultada. Algumas espécies não são fotossintetizantes.
As criptófitas, em vez de parede celular, possuem revestimento especial, o periplasto(Figura 2.16), constituído por placas proteináceas de diferentes tamanhos e formas (geral-mente poliédricas) sob a membrana plasmática e também por uma camada de placas e/ouescamas situada sobre a mesma. Forma-se uma estrutura complexa, em que as camadas externae interna do periplasto estão diretamente associadas à membrana plasmática. As célulasgeralmente têm forma elipsóide, são achatadas dorsiventralmente, sendo assimétricas edotadas de dois flagelos originados na extremidade subapical ou lateral da célula, na base deuma depressão conhecida como vestíbulo. Um vacúolo contrátil proeminente lança seuconteúdo no vestíbulo; este forma a extremidade do sulco ou citofaringe da célula. Forma-se um sulco quando a depressão é aberta ao longo de seu comprimento e uma citofaringequando a abertura ocorre apenas na extremidade anterior, sendo formado um tubo. Ovestíbulo pode ser aberto ou fechado supostamente por meio da ação de fibrilas contráteisda proteína centrina, a qual é controlada pelos níveis da Ca2+ e ATP (Graham & Wilcox,2000). Detalhes sobre a posição e forma do vestíbulo têm importância taxonômica. Juntoao vestíbulo há duas estruturas, conhecidas como corpos de Maupas, cuja função é desconhe-cida. Acredita-se que elas possam estar envolvidas no processo de remoção e digestão demembranas supérfluas (Hoek et al., 1995).
Vestíbulo
Ejectissômios
Nucleomorfo
Mitocôndria
Núcleo
Periplasto
Plasto
Núcleo
Nucleomorfo
Plasto
a)
b)
Figura 2.16 Representação esquemática deCryptomonas ovata em corte longitudianal (a)e transversal (b). Notar a existência de doisplastos grandes, nucleomorfo e ejectissômios.Extraído de Santore (1985).

67
Os flagelos são levemente diferentes entre si em forma e tamanho: o flagelo maiorapresenta duas fileiras de mastigonemas (de 1,5 µm de comprimento), ao passo que o menorapresenta apenas uma fileira de mastigonemas mais curtos (1,0 µm de comprimento). Osmastigonemas são formados entre o envelope nuclear e o retículo endoplasmático e supos-tamente aumentam a eficiência dos flagelos. Os flagelos são cobertos por escamas orgânicasminúsculas (150 nm de diâmetro), as quais apresentam padrão de roseta heptagonal, cujafunção é desconhecida. Vistas em movimento, percebe-se que as criptomônadas realizamrotação durante o deslocamento. Esse padrão de movimento decorre da assimetria da célulae da ação desigual dos dois flagelos, pois um deles é duro e quase imóvel, enquanto o outroefetivamente impulsiona a célula.
Algumas poucas espécies são desprovidas de plastos e pigmentos, apresentando nutriçãoheterotrófica, mas a grande maioria das espécies é fotoautotrófica ou heterotrófica facultativa.A maioria das criptófitas possui um único plasto bilobado (algumas espécies apresentam doisplastos), de grandes dimensões, no qual se observa freqüentemente a proeminência de umlóbulo em relação ao outro (Figura 2.16). Geralmente, a forma da célula determina a formado plasto, o qual é dotado de um único pirenóide, que freqüentemente se projeta no espaçointerlobular. O plasto está envolvido por dois pares de membranas: um deles é mais interno,o envelope do plasto; o outro, mais externo, é oriundo do chamado retículo endoplasmáticoperiplastidial. Além dos plastos, o núcleo e os grânulos de amido também estão envolvidosentre os dois pares de membranas mencionados (Lee, 1999). O espaço situado entre os doispares de membranas do plasto, que compreende um conjunto de organelas e estruturascelulares, é conhecido como compartimento periplastidial. Assim, o amido, principalproduto de reserva das criptófitas, deposita-se caracteristicamente no compartimentoperiplastidial, sendo, portanto, envolvido pelas membranas dos plastos, uma feição únicaentre organismos fotossintetizantes. Gotículas de lipídeos também são formadas comomaterial de reserva, mas estas ocorrem de forma dispersa pela célula. As criptófitasfotossintetizantes possuem um conjunto singular de pigmentos: clorofila a, clorofila c2 (aclorofila c2 transfere energia das ficobilinas para a clorofila a), carotenóides (carotenos: á-caroteno, â-caroteno; xantofilas: aloxantina, crocoxantina, monadoxantina, zeaxantina) eficobilinas (ficoeritrina e ficocianina). A xantofila aloxantina é um pigmento acessório queocorre exclusivamente nesse grupo, funcionando como marcador taxonômico que indica apresença de criptomônadas em amostras naturais de fitoplâncton, que tipicamenteapresentam misturas de muitas espécies de grupos taxonômicos distintos. As criptomônadaspossuem ficocianina ou ficoeritrina, sendo que determinada espécie não produz as duassubstâncias. Como resultado da diversidade pigmentar do grupo e das diferentes proporçõesde pigmentos nas espécies, as diferentes criptófitas podem apresentar coloração altamentediversificada, como azul, avermelhada, verde-azulada, verde oliva, marrom-amarelada emarrom. As ficobiliproteínas das criptomônadas não se localizam em ficobilissomos, comoem cianobactérias e algas vermelhas, mas, sim, no lúmen dos tilacóides, conferindo a essaregião celular aparência opaca em imagens geradas por microscopia eletrônica. A ausênciade ficobilissomos permite que os tilacóides de criptomônadas formem lamelas (quase sempreconstituídas por apenas dois tilacóides pareados), diferentemente das algas vermelhas ecianobactérias, que têm tilacóides dispersos (Graham & Wilcox, 2000). Poucas espéciespodem não ter tilacóides organizados em lamelas.
Normalmente, as criptomônadas não possuem mancha ocelar, mas quando presentesconsistem em um conjunto de glóbulos esféricos, como nas algas ocrófitas. A mancha ocelarsitua-se no centro da célula, entre os lóbulos do plasto, e não está associada aos flagelos, aocontrário do que acontece em outros grupos de algas (Hoek et al., 1995).

68
As células possuem um único núcleo, que é grande e localizado na porção posteriorda célula. Há elevado número de cromossomos, variando entre 40 e 210 nas diversas espéciesexaminadas. A mitose é aberta (com desintegração total do envelope nuclear), e na metáfaseos cromossomos se congregam de forma a gerar uma placa massiva. A divisão celular éfinalizada com a formação de um sulco de clivagem, que consiste em uma invaginação doplasmalema em forma de anel (Hoek et al., 1995).
Uma organela especial, o nucleomorfo, ocorre em criptomônadas no compartimentoperiplastidial, muitas vezes numa depressão na superfície do pirenóide. O nucleomorfoapresenta estrutura semelhante a um nucléolo: ele é envolvido por uma membrana dupla, aqual apresenta poros e contém ADN distribuído em três cromossomos; o material genéticodo nucleomorfo tem genes ativos para transcrição de ARN ribossômico, os quais controlama própria manutenção da organela, capaz de realizar autoduplicação. O nucleomorfo éinterpretado como o núcleo vestigial de um endossimbionte eucariótico incorporado porum organismo heterotrófico ancestral das criptófitas. O compartimento da célula envolvidopelo retículo endoplasmático do plasto é encarado como um vestígio do organismo endossim-bionte original (Hoek et al., 1995). Tais interpretações recebem respaldo de análises filo-genéticas moleculares que envolvem material extraído do núcleo e do nucleomorfo. Resul-tados obtidos por diversos autores independentes mostram semelhança entre o ADN donucleomorfo e o do núcleo de algas vermelhas, sugerindo que estes poderiam ter sido osorganismos eucarióticos endossimbiontes incorporados de forma estável para a formação dascriptófitas. Assim, sugere-se a ocorrência de endossimbiose secundária para a formação dogrupo (Graham & Wilcox, 2000).
Outra organela especial presente em criptomonadas é o ejectissômio (Figura 2.16).Trata-se de estruturas em forma de fita enrolada em espiral, que são explosivamente descarre-gadas de suas vesículas mediante estimulação. A fita se desenrola e estica rápida e dras-ticamente durante a descarga, projetando-se à frente. Os ejectissômios são formados por umaporção pequena e uma grande (duas fitas enroladas), dispostas num ângulo superior a 100°entre si. Cada célula tem vários ejectissômios grandes localizados junto ao vestíbulo e outrosmenores distribuídos pela periferia da célula. Os ejectissômios pequenos podem atingirobjetos distantes até 4 µm da célula, ao passo que os ejectissômios grandes podem se estenderpor distâncias de até 20 µm (Lee, 1999). Presume-se que a ação dos ejectissômios seja ummecanismo de defesa contra a perturbação causada por herbívoros ou mesmo um recursopara fugir da predação. Quando uma criptófita é seriamente irritada ou submetida a choqueambiental repentino, como mudança de pH, condições osmóticas, temperatura ou excessode luz (e calor), numa lâmina de microscópio óptico, descargas massivas de vários ejectissô-mios podem levar a uma rápida desintegração da célula. Esta é uma das razões pelas quais ascriptomônadas são mais difíceis de preservar com soluções químicas do que outras algas. Ouso de baixas concentrações de glutaraldeído tamponado (1%-3%) é recomendado, pois essasubstância penetra rapidamente e atinge as proteínas que desencadeiam a reação dos ejectissô-mios antes que ocorra o processo massivo de destruição das células (Graham & Wilcox, 2000).
A reprodução ocorre fundamentalmente por mitose assexual e citocinese, durante aqual as células continuam a se movimentar. Células que estão prestes a se dividir apresentamaspecto arredondado e quatro flagelos, em dois pares. A reprodução sexuada já foi docu-mentada em algumas espécies, mas não se sabe se é um aspecto amplamente presente nogrupo. Isogamia seguida de cariogamia, por exemplo, foi registrada em Cryptomonas sp.(Kugrens & Lee, 1988). Na espécie Proteomonas sulcata o ciclo de vida envolve um estádioflagelado haplóide e um estádio morfologicamente distinto e diplóide, mas também flagelado.

69
Cada estádio pode se reproduzir vegetativamente, mas ocasionalmente ocorre a formaçãode estádios alternados, caracterizando um ciclo de vida heteromórfico diplohaplôntico emflagelados (Hill & Wetherbee, 1986).
Há cerca de 100 espécies marinhas e 100 espécies dulciaqüícolas. As criptófitas sãopequenas e a maioria das espécies pertence ao nanoplâncton. Em algumas espécies, estádiossésseis palmelóides podem ser formados. O gênero Bjornbergiella apresenta um talo filamen-toso extremamente simples. As criptófitas são particularmente abundantes em águas oligo-tróficas, temperadas e de altas latitudes, em lagos e oceanos. Poucas espécies toleram altastemperaturas oceânicas (23°C ou mais), sendo o grupo raro ou ausente em águas hipersalinas(salinidade > 40). No ambiente, as criptófitas tendem a ocorrer mais abundantemente emáguas subsuperficiais (10-15 m de profundidade) e muitas espécies são adaptadas a níveisbaixos de irradiância. Certas espécies de crescimento rápido (como algumas dos gênerosCryptomonas e Rhodomonas) vêm sendo utilizadas de forma bem-sucedida em aqüiculturacomo alimento para outros organismos. O sucesso de sua utilização decorre, além do cres-cimento rápido, da composição química favorável a vários animais e da ausência de paredecelular, fato que facilita a digestão (ver Capítulo 8). Algumas espécies são capazes de realizarflorações, como espécies dulciaqüícolas do gênero Cyanomonas ou diversas espécies no mardo Norte. Florações de Chroomonas lacustris e Cryptomonas spp. são comuns em lagos antár-ticos perenemente cobertos por gelo, contribuindo com mais de 70% da biomassa dofitoplâncton naqueles locais (Graham & Wilcox, 2000).
Um aspecto extremamente interessante quanto às criptófitas é sua incorporação nointerior de células de organismos mixotróficos. As criptófitas, ou apenas algumas de suas partes(como os plastos), ocorrem endossimbioticamente (temporária ou permanentemente) emalgumas espécies de protozoários (por exemplo, os ciliados Myrionecta rubra e Cyclotytrichiummeunieri) e dinoflagelados (por exemplo, Pfiesteria piscicida e Amphidinium wigrense), contri-buindo de forma fundamental para a sobrevivência dessas outras espécies. Algumas dessasespécies de ciliados e dinoflagelados podem formar proliferações intensas, e especula-se queao menos em parte essa capacidade decorra da aquisição de energia através da endossimbiosede criptófitas, como no caso do ciliado Mesodinium rubrum.
Lee (1999) classifica as criptófitas em apenas uma classe, Cryptophyceae, constituídapor apenas três ordens: Goniomonadales, constituída por espécies incolores e/ou sem plastos;Cryptomonadales, formada por células geralmente avermelhadas com plastos contendo aficobiliproteína ficoeritrina-cr; e Chroomonadales, formada por células geralmente azul-esverdeadas com plastos contendo a ficobiliproteína ficocianina-cr.
Cryptomonas é o gênero mais estudado da divisão; muitas das informações assumidascomo sendo comuns à divisão inteira foram levantadas em estudos realizados apenas comespécies do gênero Cryptomonas. Outros gêneros comuns de criptófitas são Chilomonas (possuiplastos incolores), Chroomonas, Goniomonas (= Cyathomonas) (desprovido de plastos),Hemiselmis, Hillea, Rhodomonas e Storeatula.
2.2.6 DIVISÃO PRYMNESIOPHYTA (= HAPTOPHYTA)Esta divisão compreende organismos flagelados unicelulares e formas vegetativas solitárias
ou coloniais não flageladas, que possuem estádios flagelados em alguma parte de seu ciclo devida. Os representantes dessa divisão são conhecidos como haptófitas ou primnesiófitas. A feiçãomais marcante do grupo é a presença de uma estrutura em forma de fio, conhecida comohaptonema, normalmente situada entre os dois flagelos típicos (Figura 2.17). A função dohaptonema não é devidamente compreendida até o momento. Segundo alguns autores, o

70
haptonema é uma estrutura vestigial que tenderia a desaparecer nas células; segundo outros,o haptonema poderia ter a função de orientar o movimento das células ou auxiliar na buscapor alimento. O nome Haptophyta foi atribuído pela presença do haptonema, não tendonenhuma relação com algum dos gêneros do grupo. Posteriormente, o nome da divisão foialterado para Prymnesiophyta (Gr. prymnesium, cabo rígido + Gr. phykos, alga ou Gr. phyton,planta), utilizando-se o gênero Prymnesium como tipo. Como o nome Haptophyta e adesignação de seus representantes como haptófitas já estavam consagrados, atualmente sãoreconhecidas formalmente as duas designações para o grupo e para seus representantes.
5 µm
1 µm
a)
b)
c
ss
h
l
sf
lf
c
h
Figura 2.17 Esquemas de duas primnesiófitas: Chrysochromulina ephippum (a) e Pavlova mesolychnon (b).c = plasto; l = vesícula de leucosina; SS = escamas; sf = flagelo curto; lf = flagelo longo; h = haptonema.Reproduzido de Lee (1999).
Há cerca de 280 espécies conhecidas, sendo a maioria marinha e exclusivamentefotoautotrófica, mas há também espécies mixotróficas (por osmotrofia ou fagotrofia). Asprimnesiófitas são pequenas e em sua maioria pertencem ao nanoplâncton, podendo serdominantes naquela fração do plâncton marinho. As células das primnesiófitas são comu-mente cobertas por escamas e, em muitas delas, as escamas são mineralizadas, formandoornamentação muito marcante. Do total, 40 gêneros (envolvendo cerca de 200 espécies)constituem os cocolitoforídeos, grupo extremamente abundante e importante no mar,caracterizado pela belíssima ornamentação das células com escamas, os cocólitos, que namaioria dos casos são impregnadas com carbonato de cálcio (Figura 2.18). Outros 11 gênerosapresentam células não ornamentadas com cocólitos e englobam cerca de 80 espécies (asespécies mixotróficas de primnesiófitas, mencionadas acima, não apresentam cocólitos). Apartir de registros fósseis, estima-se que os cocolitoforídeos tenham surgido há 250 milhõesde anos, logo após o maior evento de extinção em massa ocorrido na Terra, no final doPermiano. Estima-se que 85% das espécies marinhas e mais de 90% das espécies terrestrestenham sido extintas naquele evento (Benton, 2003), possivelmente em decorrência de

71
atividades vulcânicas muito intensas.3 Entretanto, a produção de cocólitos é considerada umacaracterística derivada, que teria surgido posteriormente entre as primnesiófitas, o que sugereque a divisão provavelmente seja mais antiga que os cocolitoforídeos (Graham & Wilcox,2000). Não há registros antigos de células sem ornamentação, cuja conservação e identificaçãosão muito mais difíceis.
a)
2 µm 2 µm
b)
3 µm
3 µmc)
d)2 µm
e) 2 µmf)
Figura 2.18 Micrografias eletrônicas de varredura de alguns cocolitoforídeos: (a) Coccolithus pelagicus;(b) Emiliania huxleyi; (c) Dicosphaera tubifera; (d) Pontosphaera syacusana; (e) Syracosphaera nodosa;(f ) Braarudosphaera bigelowii. Reproduzido de Faber & Preisig (1994).
3. A famosa extinção em massa ocorrida no final do Cretáceo (há 65 milhões de anos) provocou odesaparecimento de cerca de 50% das espécies existentes, incluindo os dinossauros, uma percentagemmenor do que na grande extinção em massa do Permiano.

72
Os plastos são envolvidos dentro de uma dobra da membrana do retículo endo-plasmático, que se constitui num retículo endoplasmático plastidial. O plasto se dispõe juntoao núcleo e o retículo endoplasmático plastidial forma um contínuo que envolve também onúcleo, constituindo seu envelope nuclear. Os pigmentos fotossintéticos exibidos por essegrupo incluem clorofila a, clorofila c1, clorofila c2 e carotenóides como fucoxantina (geral-mente o mais abundante deles), ß-caroteno, diadinoxantina, diatoxantina e derivados dafucoxantina, como 19’-hexanoiloxifucoxantina e 19’-butanoiloxifucoxantina. A presença decarotenóides em grande concentração confere a seus representantes uma coloração amarelada,dourada ou diversos tons de marrom, mascarando a cor da clorofila. Entretanto, pode haverdiferenças expressivas na composição de carotenóides, sendo que algumas sequer apresentamfucoxantina. De acordo com Falkowski & Raven (1997), a clorofila c apresenta uma viabiossintética distinta daquela das clorofilas a e b; a clorofila c é considerada como umintermediário na transferência de energia entre carotenóides e clorofila a dentro do complexode antena carotenóide/clorofila.
O produto de reserva é um polissacarídero derivado de glicose formado por ligaçõesglicosídicas do tipo ß -1,3, a crisolaminarina, que é levemente diferente do paramilo existentenos euglenóides, e ocorre no interior de vacúolos citoplasmáticos. Lipídeos também sãoacumulados pelo citoplasma como produto de reserva.
As primnesiófitas possuem dois flagelos lisos e geralmente iguais, desprovidos demastigonemas e inseridos na porção anterior ou lateral da célula. Em algumas espécies osdois flagelos apresentam diferenças discretas de comprimento. Embora o haptonema situe-se entre os flagelos, sua ultra-estrutura é inteiramente diferente daqueles. Enquanto os flagelosapresentam o típico arranjo 9 + 2 de microtúbulos (9 pares periféricos + 2 microtúbuloscentrais), os haptonemas têm seis ou sete microtúbulos isolados (não ocorrem em pares) emum arranjo de meia-lua; além disso, entre o corpo do haptonema e a membrana plasmática,ocorre uma dobra do retículo endoplasmático.
A reprodução vegetativa por divisão binária é a forma mais comum de reprodução nogrupo. Porém, reprodução sexuada existe e já foi documentada em várias espécies. Um ciclode vida heteromórfico diplohaplôntico foi descoberto em algumas espécies, no qual umestádio flagelado diplóide e planctônico se alterna com um estádio filamentoso haplóide ebentônico. Em outras espécies existe uma alternância entre estádios flagelados e não flagelados,mas detalhes sobre esse padrão de ciclo de vida permanecem obscuros.
Os cocolitoforídeos são pequenos e pertencentes ao nanoplâncton; eles ocorrem emtodo o mundo, sendo especialmente abundantes em águas oligotróficas e quentes, como astípicas de oceano aberto tropical. Os cocolitoforídeos são particularmente interessantes porsuas relações com os ciclos biogeoquímicos do enxofre e do carbono. Os cocólitos sãoformados pela deposição de seus materiais de origem via secreção do complexo de Golgi.Essa organela secreta substâncias orgânicas como fibrilas celulósicas que constituem a matrizpara posterior acumulação de carbonato de cálcio. Em algumas espécies, entretanto, não háacumulação de carbonato de cálcio e os cocólitos formados são, portanto, de naturezaorgânica. De qualquer forma, o desenvolvimento de cocólitos envolve a secreção de substân-cias orgânicas no interior de cisternas de um único e extremamente grande aparato de Golgisituado próximo dos corpos basais dos flagelos, com sua face de formação voltada para aparte anterior da célula. Esse aparato de Golgi especial é típico desse grupo de algas. Ashaptófitas que produzem cocólitos podem ser separadas em dois grupos fundamentais, deacordo com sua estrutura e onde são produzidos: há aquelas formadoras de “heterococólitos”,sendo estes formados internamente (pelo aparato de Golgi), e aquelas possuindo os chamados

73
“holococólitos”, que são formados extracelularmente. Os holococólitos são formados regular-mente por cristais hexagonais ou romboédricos, enquanto os heterococólitos são formadospor cristais romboédricos, cujas superfícies e ângulos podem ser total ou parcialmentesuprimidos (Graham & Wilcox, 2000). Os heterococólitos são mais robustos que os pequenose mais delicados holococólitos. Alguns ciclos de vida incluem formas que se alternam naprodução de holococólitos e heterococólitos. No gênero Coccolithus, as formas que produzemheterococólitos são imóveis e se alternam com formas flageladas dotadas de holococólitos.No segundo caso, as escamas orgânicas são produzidas pelo aparato de Golgi dentro da célula,são secretadas externamente e, então, calcita é depositada sobre a escama (cocólito) dentrodo espaço coberto por um envelope orgânico (Graham & Wilcox, 2000).
A calcificação dos cocólitos é dependente da fotossíntese, pois é exatamente por meiodesse processo que a energia necessária para o transporte de substâncias no aparato de Golgie através do citoplasma torna-se disponível. Além disso, a fotossíntese também funciona comoum mecanismo de captação do dióxido de carbono gerado por intermédio da equação dacalcificação (Equação 2.1), que tem como produto principal carbonato de cálcio:
2HCO3– + Ca2+ → CaCO
3 + CO
2 + H
2O 2.1
A produção de CO2, prontamente utilizado na fotossíntese, é considerada como umavantagem adaptativa extremamente relevante dos cocolitoforídeos. Dados experimentaisderivados de estudos com cepas que realizam processos intensos de calcificação com outrasque realizam calcificação em menor intensidade apóiam essa interpretação (Graham &Wilcox, 2000). Existe uma grande variedade de cocólitos entre os cocolitoforídeos, queparecem exibir relações com o nicho ecológico ocupado pelas espécies, ou pelo habitat. Otipo mais simples de cocólito consiste em discos basais sobre os quais paredes de calcita sãogeradas ao longo das laterais para formar um cilindro curto (ex.: Syracosphaera). Outrasespécies produzem cocólitos que consistem em dois escudos unidos por uma coluna curta(ex.: Emiliania). Esse padrão permite que cocólitos vizinhos ocorram de forma imbricada,aumentando a coesão da cobertura celular.
Alguns cocolitoforídeos possuem um haptonema bem desenvolvido, mas em outrostal estrutura é vestigial ou ausente. A presença de cocólitos de carbonato de cálcio é umafeição única dentre algas microscópicas e é encarada por certos autores como uma respostaadaptativa para a necessidade de uma camada de proteção a células que habitam ambientessupersaturados de carbonato de cálcio. Os cocolitoforídeos têm taxas de afundamento maisrápidas que os demais membros do fitoplâncton de mesmo tamanho, presumivelmentedevido ao aumento de densidade gerado pelos cocólitos. Essa característica capacita oscocolitoforídeos a absorver nutrientes disponíveis em águas mais profundas, diminuindo acompetição por nutrientes escassos de águas superficiais. A liberação de cocólitos em águasprofundas, aliado aos movimentos de seus flagelos, permitiria o retorno dos indivíduos aáguas mais rasas, onde a fotossíntese é favorecida. Esse mecanismo é possivelmente responsávelpelo sucesso de ocorrência de cocolitoforídeos em águas pobres em nutrientes, como o oceanoaberto tropical. Estudos sobre dinâmica de nutrientes indicam que os cocolitoforídeos sãocapazes de consumir nitrogênio nas formas de nitrato, uréia e nitrito, embora amônia sejaa forma preferida, conforme ocorre com a maioria das espécies do fitoplâncton. Fosfato podeser obtido via ruptura de moléculas orgânicas por fosfatases extracelulares. Sob condiçõesótimas de crescimento, algumas espécies de cocolitoforídeos podem se dividir até 2,5 vezespor dia, mas outras têm seu máximo em apenas uma divisão por dia. Muitas espécies sãosensíveis ao excesso de luz, principalmente aquelas oriundas de oceano aberto tropical, sendo

74
levadas à morte se submetidas à luz contínua. Esse fato sugere que essas espécies são depen-dentes da ocorrência de ciclos diários de claro-escuro. Espécies oriundas de ambientescosteiros, polares e temperados parecem ser menos sensíveis à ausência de fotoperíodo.
Muitas das informações relativas às funções dos cocólitos para os cocolitoforídeos foramobtidas a partir de estudos com Emiliania huxleyi, espécie cosmopolita presente em todosos ambientes oceânicos e neríticos do mundo, exceto nos ambientes polares. Uma excelenterevisão sobre o conhecimento disponível sobre essa espécie foi feita por Paasche (2001). Oscocólitos possuem provavelmente várias funções para as células, como: i) restrição do contatodas células com patógenos como bactérias e vírus; ii) proteção contra predação exercida porprotozoários (proteção virtualmente não efetiva contra muitos membros do zooplâncton, comocopépodos); iii) regulação da flutuabilidade das células (produção e/ou liberação de cocólitosmais pesados para regular a posição da célula na coluna d’água). Alguns autores sugerem tambémque cocólitos poderiam servir para concentrar o foco de luz no interior da célula e atividadena absorção de nutrientes. Estudos recentes indicam também que os cocólitos podem ter funçãoimportante na dispersão de luz na coluna d’água, pois dependendo do ângulo no qual os raiossolares atingem as células eles poderiam ser refletidos de volta para a atmosfera ou seremprojetados para outras profundidades (Voss et al., 1998; Gordon & Du, 2001).
Algumas espécies de primnesiófitas são muito abundantes no mar, como Phaeocystis antarctica,possivelmente a espécie do fitoplâncton individualmente presente em maior biomassa no oceanoAntártico. Outras espécies estão envolvidas em fenômenos intensos de floração algácea, as quais sãonocivas à biota marinha; dentre elas estão a espécie colonial Phaeocystis globosa e alguns representantesdo gênero Prymnesium, como P. parvum e P. patelliferum, além de Chrysochromulina, Emiliania huxleyie Gephryocapsa oceanica. Crescimentos densos de Chrysochromulina podem causar danos a guelrasde peixes, com conseqüente mortalidade dos animais; representantes do gênero Prymnesium produ-zem ictiotoxinas que são responsáveis pela morte de peixes. Florações de Phaeocystis são comunsnos mares do Norte e Báltico, e nos eventos mais intensos provocam a formação espetacularde uma espuma esbranquiçada que flutua sobre a água. E. huxleyi freqüentemente realizaflorações de imensas dimensões no Atlântico Norte, eventualmente atingindo milhares dequilômetros quadrados.
Embora os cocolitoforídeos constituam uma fração menor de vasas calcárias recentesdos oceanos, no Cretáceo eles dominaram o nanoplâncton, numa indicação de que o grupojá foi mais abundante e diversificado que atualmente (Lee, 1999). Os cocolitoforídeosconstituíram-se nos principais componentes de depósitos calcários de sedimentos oceânicosdo Mesozóico (Jurássico e Cretáceo) e Terciário. A abundância de cocólitos fósseis atingiuum pico no Cretáceo Final (95-63 milhões de anos atrás), quando enormes depósitos de gizse formaram no norte da Europa e em muitas outras regiões pelo mundo. O impacto de umasteróide na costa da península de Yucatán (México), o famoso “evento K/T”, presumi-velmente teria provocado a extinção dos dinossauros e amonitas; estima-se que no mesmoevento cerca de 80% das espécies de cocolitoforídeos foram também extintas. Acredita-seque a extinção em massa dos cocolitoforídeos pode ter sido relacionada à diminuição datemperatura da Terra após o “evento K/T”, pois a maioria das espécies modernas de cocolito-forídeos tem afinidade por águas quentes. As espécies sobreviventes irradiaram-se e originaramvárias outras, as quais sofreram segunda extinção em massa entre 50 e 30 milhões de anosatrás (Graham & Wilcox, 2000).
Como os cocólitos são comuns, pequenos e exibem baixo endemismo, eles são usadoscomo indicadores estratigráficos para identificar rochas de diferentes idades de lugaresdistintos (Graham & Wilcox, 2000). A presença de cocólitos em rochas sedimentares pode

75
ser usada como marcador na descoberta e modo de deposição de jazidas de petróleo. Porexemplo, os depósitos de xisto de Kimmeridge Clays (Inglaterra, Reino Unido) são dispostosentre faixas de pedra calcária que são compostas principalmente de cocólitos de uma espécie,Ellipsagelosphaera britannica (Lee, 1999). Outras rochas associadas a petróleo têm tambémcocólitos característicos indicadores. Assim, em termos práticos, ao examinar o sedimentoobtido por meio de uma perfuração, a constatação da presença de certos cocólitos pode indicaralta probabilidade de existência de jazidas de petróleo. Cerca de 1.000 espécies fósseis decocolitoforídeos são amplamente utilizadas como indicadores de presença de petróleo (Younget al., 1994). Esse fato também mostra contundentemente que as primnesiófitas já forammuito mais abundantes no passado, tendo em vista a existência de menos de 300 espéciesmodernas.
As primnesiófitas também desempenham papel importante no ciclo do enxofre no mar.Devido à sua abundância, Phaeocystis e vários cocolitoforídeos como Emiliania são fontesimportantes de dimetilsulfito (DMS). Primnesiófitas utilizam dimetilsulfonio propionato(DMSP) como osmorregulador. Ao serem consumidas e digeridas pelo zooplâncton e pro-tozooplâncton, ou mesmo quando as células algáceas se rompem, os conteúdos celulares sãoliberados no meio circundante, incluindo DMSP, que na água do mar é convertido a DMSe ácido acrílico. DMS é uma substância volátil que se difunde para a atmosfera, onde éoxidado. Tais substâncias contêm enxofre oxidado e geram duas conseqüências distintas: a)proporcionam a formação de chuva ácida; b) seus aerossóis contribuem para a nucleação denuvens sobre os oceanos. Embora significativa, a ocorrência de chuva ácida por contribuiçãode primnesiófitas é bem menor do que aquelas decorrentes de ação antrópica sobre os conti-nentes e áreas costeiras. A formação de nuvens sobre os oceanos aumenta o albedo das áreascobertas pelas nuvens, sendo este um efeito importante no contexto do aquecimento global,pois menos calor tenderia a atingir a superfície da Terra. O ácido acrílico, também formadocom a quebra do DMSP, possui propriedades antibióticas e dados experimentais indicam queele pode inibir o crescimento de bactérias junto a células de primnesiófitas (Sze, 1998).
Algumas espécies, sobretudo as dos gêneros Isochrysis e Pavlova, são largamente utili-zadas em maricultura em todo o mundo por suas propriedades nutritivas adequadas, cresci-mento rápido, tamanho pequeno e digestão fácil por animais.
Segundo Lee (1999), a divisão Prymnesiophyta apresenta apenas uma classe,Prymnesiophyceae, a qual é dividida em duas ordens: Prymnesiales, cujos representantesapresentam dois flagelos lisos iguais, sem mancha ocelar e com escamas comumente cobrindoa célula; e Pavlovales, que apresenta espécies dotadas de dois flagelos desiguais, freqüentementecobertos com cerdas e depósitos, cujas células podem apresentar mancha ocelar.
Os gêneros mais comuns da divisão são Chrysochromulina, Isochrysis, Pavlova e Prymnesium(primnesiófitas nuas) e os cocolitoforídeos Calyptrolithophora, Coccolithus, Emiliania, Gephyrocapsa,Phaeocystis (gênero colonial, dotado de pouquíssimos cocólitos, quando presentes) e Syracosphaera.Alguns gêneros de cocolitoforídeos (ex.: Hymenomonas, Ochrosphaera e Pleurochrysis) alternamfases dotadas de cocólitos calcificados e outras com cocólitos de natureza orgânica.
2.2.7 DIVISÃO DINOPHYTA
Esta divisão compreende os dinoflagelados, organismos unicelulares dotados de doisflagelos distintos entre si. Existem cerca de 550 gêneros, e especula-se a existência de 2.100a 4.000 espécies, um número incerto pela dificuldade de identificação dos dinoflagelados.A grande maioria das espécies ocorre no ambiente marinho, havendo apenas cerca de 220espécies dulciaqüícolas. Cerca de metade das espécies de dinoflagelados consiste em heteró-

76
trofos não pigmentados e não fotossintetizantes que realizam nutrição heterotrófica. Asespécies portadoras de plastos fotossintetizantes variam significantemente na composiçãode pigmentos e tipo de RuBisCO, a enzima responsável pela fixação fotossintética de carbono.Muitas espécies fotossintetizantes são capazes de realizar também nutrição heterotrófica,configurando um quadro de mixotrofia. Os dinoflagelados ocorrem em registro fóssil emdepósitos de 400 milhões de anos. Os achados mais antigos são atribuídos à espécie iden-tificada como Arpylorus antiquus, a qual se assemelha morfologicamente a dinoflageladosmodernos. Embora a grande maioria dos dinoflagelados seja constituída por flageladosunicelulares, existem colônias de células flageladas, células (ou estádios do ciclo de vida) nãoflageladas e também filamentos.
As células dos dinoflagelados podem ser classificadas em duas categorias básicas emfunção das posições dos flagelos. Os dinoflagelados desmocontos (Figura 2.19a) têm os doisflagelos inseridos na posição anterior da célula, sendo que um deles, tipicamente espiralado,pode circundar a célula ao longo de um sulco discreto e difícil de visualizar em microscopiaóptica. O segundo tipo básico de dinoflagelados consiste nos dinocontos (Figura 2.19b),os quais dispõem de flagelos inseridos lateralmente. Um dos flagelos (flagelo transversal) sealoja num grande sulco que circunda completamente a célula, o cíngulo (Figura 2.19b). Oflagelo transversal dispõe-se inteiramente dentro do cíngulo e tende a ser maior que o segundoflagelo, o longitudinal; este se projeta para trás e se encontra num sulco menor, que se estendepor apenas uma parte da célula. O cíngulo, disposto na porção equatorial, divide a célulatípica dos dinoflagelados dinocontos em epicone e hipocone. Os dois flagelos emergem naface ventral da célula e dispõem-se entre si num ângulo aproximadamente reto. O arranjodos flagelos faz com que seus movimentos provoquem giros ou rodopios nos dinoflageladosem deslocamento, de forma que o nome do grupo (Dinophyta – Gr. dineo, girar + Gr. phyton,planta) deriva dessa característica. Entende-se que o giro é provocado pela ação do flagelotransversal, que é espiralado, pois espécies que não dispõem desse flagelo (como aquelas dogênero Noctiluca) não apresentam movimentos giratórios ao se deslocar. O flagelo longitudinalpode se estender por 100 µm além do limite da célula, apresenta duas fileiras de cerdasdelicadas e supostamente confere direção ao movimento do dinoflagelado. Apesar dos movi-mentos giratórios resultantes da ação conjunta dos flagelos, várias espécies são capazes derealizar deslocamentos rápidos, atingindo 200-500 µm s–1; ou movimentos mais lentos, masde longa duração, atingindo algumas dezenas de metros por dia, como ocorre nas espéciesque realizam migrações na coluna d’água.
Flagelotransversal
Cíngulo
Sulco
Flagelolongitudinal
a) b)
Figura 2.19 Representações dos doistipos básicos de células de dinoflagelados:(a) dinoflagelado desmoconto; (b)dinoflagelado dinoconto. Modificado deSteidinger & Tangen (1997).

77
Espécies heterotróficas freqüentemente apresentam uma estrutura tubular ou pedún-culo que se projeta para fora da célula e que é usada para sugar os conteúdos de suas presas.O pedúnculo normalmente não se apresenta permanentemente exposto, sendo projetadopor ocasião da captura da presa e recolhido após o consumo do alimento. O pedúnculo seinsere próximo à base dos flagelos. Certas espécies de dinoflagelados produzem estruturassemelhantes a pseudópodes, capazes de envolver e digerir as células das presas (Figura 2.20).Outra organela especial dos dinoflagelados é a púsula, também localizada próximo à basedos flagelos. Trata-se de uma série de vesículas que supostamente participam da osmorre-gulação, absorção de macromoléculas orgânicas e secreção celular. As púsulas são formadaspor invaginações da membrana plasmática e não funcionam como vacúolos contráteis: elasapenas se abrem nas imediações da base dos flagelos, não realizando movimentos de contraçãoe relaxamento. As púsulas são mais desenvolvidas em espécies heterotróficas de dinoflagelados.
50 µm
50 µm
a) b) c) d)
Figura 2.20 Alimentação de alguns dinoflagelados não-fotossintetizantes. (a) Protoperidinium spinulosumalimentando-se de uma cadeia de células da diatomácea Chaetoceros curvatus, por meio da ação de umpseudópodo com estrutura fibrilar e com forma de tela. (b-d) Protoperidinium conicum consumindo adiatomácea Corethron hystrix. Captura da presa por um filamento adesivo (b); desenvolvimento de umpseudópodo a partir do poro sulcal (c); retração do pseudópodo contendo a presa. O processo completodura vários minutos. Reproduzido de Hoek et al. (1995).
Além dos flagelos e estruturas associadas, três outras características das células dosdinoflagelados são particularmente especiais: a teca, o núcleo e os plastos. As células sãorecobertas por um sistema de camadas periféricas de vesículas membranáceas conhecido comoanfiesma. Em muitas espécies o anfiesma contém placas de celulose, conferindo uma estruturarígida à célula. Essas espécies que apresentam anfiesma enrigecido são ditas tecadas ou dotadasde armadura. As demais espécies, desprovidas de anfiesma com placas rígidas, são chamadasde não-tecadas ou sem armadura. Na realidade, todos os dinoflagelados são tecados, masvaria entre as espécies o grau de estruturação e rigidez da teca. O tamanho, o número e oarranjo das placas tecais são características valiosas para a taxonomia do grupo (Figura 2.21).
Nas espécies dotadas de plastos, estes são envoltos por três membranas, nenhuma dasquais é conectada ao retículo endoplasmático (Hoek et al., 1995). A maior parte dos autoresinterpreta esse padrão como sendo resultante da incorporação de uma célula eucariótica comoendossimbionte, a qual teria sofrido redução intensa de suas organelas (Sze, 1998). As espécies

78
desprovidas de plastos são geralmente incolores. A coloração dos dinoflagelados com plastosgeralmente varia de marrom a dourado (daí o nome alternativo da divisão, Pyrrhophyta, emdesuso, que significa “plantas de cor de fogo”), refletindo a ocorrência, em concentraçõesdistintas, de diferentes carotenóides, dentre os quais o â-caroteno e a xantofila peridinina,4
exclusiva do grupo. A clorofila c2 está presente nas espécies fotossintetizantes. Os tilacóidesestão geralmente agrupados de três em três, formando lamelas periféricas dentro do plasto.Pirenóides estão sempre presentes, mas podem variar bastante em sua forma; há pirenóidesgrandes e pedunculados, outros são pequenos e inseridos entre os tilacóides, e existemtambém aqueles discretamente protuberantes na superfície do plasto. O ADN do cloroplastoé encontrado em pequenos nódulos dispersos no interior da organela.
Pré-cingulares
Pós-cingulares
Cingulares
Antapicais
Pontas antapicais
Apicais
Poroapical
Antero-intercalares
Bordasda cintura
Poro flagelar
Sulcais
Suturas
Pré-apical
Figura 2.21 Nomenclatura de placas tecais aplicada a dinoflagelados com morfologia semelhante aPeridinium. Modificado de Taylor (1990).
Grânulos de amido ocorrem no citoplasma como produto de reserva. O amido ésintetizado fora dos plastos e é acumulado em forma de grânulos pequenos e numerosos. Lipídeostambém se constituem em material de reserva, dispersos em gotículas pelo citoplasma, mas têmmenor importância que o amido.
Diferentes tipos de estigmas podem ser encontrados em dinoflagelados (Hoek et al., 1995):glóbulos esféricos dispersos pelo citoplasma; glóbulos arranjados em linha no interior do plasto; camadasde glóbulos envolvidas por uma membrana; ocelos complexos compostos por lentes; e um retinóide(encontrado apenas na família Warnowiaceae). Seja qual for a complexidade estrutural das manchasocelares, elas são compostas por gotículas de lipídeos contendo carotenóides. A presença de manchasocelares determina a orientação dos dinoflagelados à luz (fototaxia positiva), embora o funcionamentodesse mecanismo ainda não seja suficientemente compreendido.
Os dinoflagelados podem apresentar também algumas estruturas que se projetam da célulapara o ambiente. Os tricocistos são as mais comuns delas e ocorrem na maioria das espécies naregião periférica das células. Trata-se de estruturas de natureza protéica em forma de haste que se
4. Embora seja um carotenóide exclusivo do grupo, algumas espécies não apresentam peridinina.

79
desenvolvem no interior de sáculos gerados pelo aparato de Golgi, que migram para a periferiada célula e orientam-se perpendicularmente em relação a ela e junto a poros, no caso de espéciestecadas. Quando o dinoflagelado é perturbado por fatores externos como temperatura ou tur-bulência, por exemplo, ocorre ruptura dos sáculos, com conseqüente entrada de água e alteraçãoda conformação física da proteína que constitui os tricocistos; como resultado, a estrutura alonga-se o equivalente a oito vezes seu tamanho em repouso e provoca liberação explosiva dos tricocistospara fora da célula. A função dos tricocistos está supostamente relacionada à defesa da célula contrapredadores microscópicos. Nematocistos constituem-se em outro tipo de estrutura de defesaapresentada por algumas espécies de dinoflagelados (sobretudo dos gêneros Polykrikos eNematodinium). Eles são maiores (até 20 µm) e mais elaborados que os tricocistos, originados porprocesso semelhante e ejetados das células também em situações de perturbação. São estruturascompletamente diferentes dos nematocistos apresentados por cnidários, embora recebam o mesmonome. Existem também os mucocistos, que são meros sacos que liberam mucilagem para o exteriore que são produzidos pelos corpos de mucilagem; ocorrem também em euglenófitas.
A reprodução vegetativa por divisão binária das células é a forma predominante de re-produção dos dinoflagelados. A produção de zoósporos é um tipo comum de reproduçãoassexuada em dinoflagelados. Diversas espécies são capazes de formar cistos de resistência, osquais podem ser conservados na natureza por períodos extremamente longos. Cistos de váriasespécies primitivas estão conservados em depósitos de centenas de milhões de anos e têm papelimportantíssimo na reconstrução da história evolutiva do grupo. Reprodução sexuada é re-conhecida no grupo para parte das espécies, mas se especula que essa característica seja comumna maioria das espécies. Com base nas espécies mais estudadas, assume-se que os dinoflageladostenham um ciclo de vida haplôntico, sendo que apenas o zigoto possui um núcleo diplóide(Hoek et al., 1995). Células vegetativas diplóides já foram detectadas em espécies heterotróficasde dinoflagelados (Sze, 1998).
O EXTRAORDINÁRIO CICLO DE VIDA DE DINOFLAGELADOSDO COMPLEXO PFIESTERIA
Pfiesteria piscicida Steidinger & Burkholder é um dinoflagelado de vida livre,formador de cistos, encontrado em águas rasas e sedimentos de estuários eutróficos. Éuma espécie predominantemente heterotrófica, mas é capaz também de realizar fo-tossíntese na ausência de alimento orgânico, sendo, portanto, mixotrófica (Burkholder& Glasgow, 1997). A descrição recente desse dinoflagelado (Burkholder et al., 1992)representou uma nova família, gênero e espécie dentro da ordem Dinamoebales, tendosido descoberta devido a patologias observadas em peixes, que implicaram a repentinamorte de tilápias em cultivo. Os peixes morreram quando expostos à água recentementecoletada do rio Pamlico, Carolina do Norte (EUA). Após o exame minucioso da água,foi detectada a presença desse novo dinoflagelado. Ele tem sido responsável por diversoseventos de mortandade em massa de peixes em estuários da Carolina do Norte. P.piscicida é uma espécie diferente de todos os dinoflagelados conhecidos, dotada dasseguintes características especiais:
demonstra forte atração por peixe vivo e outros tecidos frescos, secreções e excretas,detectando-os quimicamente;é estimulada a produzir toxinas pela presença de peixe vivo;

80
apresenta complexo ciclo de vida, com estádios flagelados, amebóides e de resis-tência (cistos);possui nutrição extremamente diversificada, sendo capaz de assimilar nutrientesorgânicos e inorgânicos e de consumir organismos que fazem parte de todos osníveis tróficos de estuários, desde bactérias a tecidos de mamíferos;está relacionada à produção de aerossóis que causam danos neurocognitivos emhumanos.
Os dinoflagelados que se assemelham morfologicamente a P. piscicida fazemparte do complexo tóxico Pfiesteria, representado por “predadores de emboscada”,que detectam quimicamente suas presas. A capacidade que essas algas têm de detectarquimicamente suas vítimas é altamente desenvolvida; quando isso ocorre, grandenúmero de células faz um ataque direto à presa, consumindo seus tecidos mais exter-nos. P. shumwayae faz parte desse complexo e é distinguida de P. piscicida apenas emsuas formas de zoósporo (Burkholder et al., 2001). Especialistas admitem a existênciade outras espécies do gênero Pfisteria, mas especula-se que elas ainda não foramencontradas fundamentalmente por causa da dificuldade na identificação.
P. piscicida apresenta o ciclo de vida mais complexo de todos os dinoflagelados conhe-cidos, com pelo menos 24 estádios diferentes, segundo Burkholder & Glasglow (1997),incluindo várias formas flageladas amebóides e encistadas, que variam de 5 a 450 µm dediâmetro (Litaker et al., 2002). Algumas dessas formas são tóxicas e ativas na colunad’água e no sedimento. Outra distinção ocorre porque P. piscicida apresenta hábitoalimentar generalista. Essa espécie ataca grande variedade de peixes, porém, quandoestes não estão disponíveis, ela consome bactérias, algas (principalmente criptomônadas)e microfauna (Burkholder & Glasglow, 1997). Essa espécie também apresenta alimen-tação autotrófica, marcada por sua capacidade de cleptoplastia, processo em que cloro-plastos funcionais são retidos pela predação de componentes do fitoplâncton (Lewituset al., 1999).
O ciclo de vida de P. piscicida envolve transformações rápidas entre formasflageladas, amebóides e cistos. Um estádio biflagelado haplóide móvel é a forma maiscomum e duradoura no ciclo de vida da espécie. Essa forma é comumente referidacomo zoósporo, apresentando em média 10,2 ± 1,1 µm de comprimento (Litaker etal., 2002). Eles apresentam também elasticidade da área sulcal, o que possibilita aoszoósporos quase dobrarem de tamanho durante a alimentação. Eles se alimentam dealgas e peixes através de um pedúnculo. Os zoósporos podem ser tóxicos ou não-tóxicos, podendo representar a mesma célula na presença ou ausência do peixe vivo,respectivamente. Os zoósporos variam consideravelmente em forma e tamanho,dependendo do estádio anterior às suas origens e da atividade alimentar, mas sempreapresentam um flagelo helicoidal transversal e outro longitudinal espesso. Além dezoósporos, os estádios flagelados compreendem também gametas e planozigotos.Especula-se que as transformações de um estádio para outro estejam associadas amudanças de temperatura, salinidade, teores de fósforo na água, presença de matériaorgânica e disponibilidade de peixe e outras presas, mas ainda não são plenamentecompreendidas (Burkholder & Glasgow, 1997). Formas presentes na coluna d’águapodem se converter em amebas e repousar no sedimento; por outro lado, amebaspodem deixar o sedimento e se converter em formas flageladas que exploram a coluna
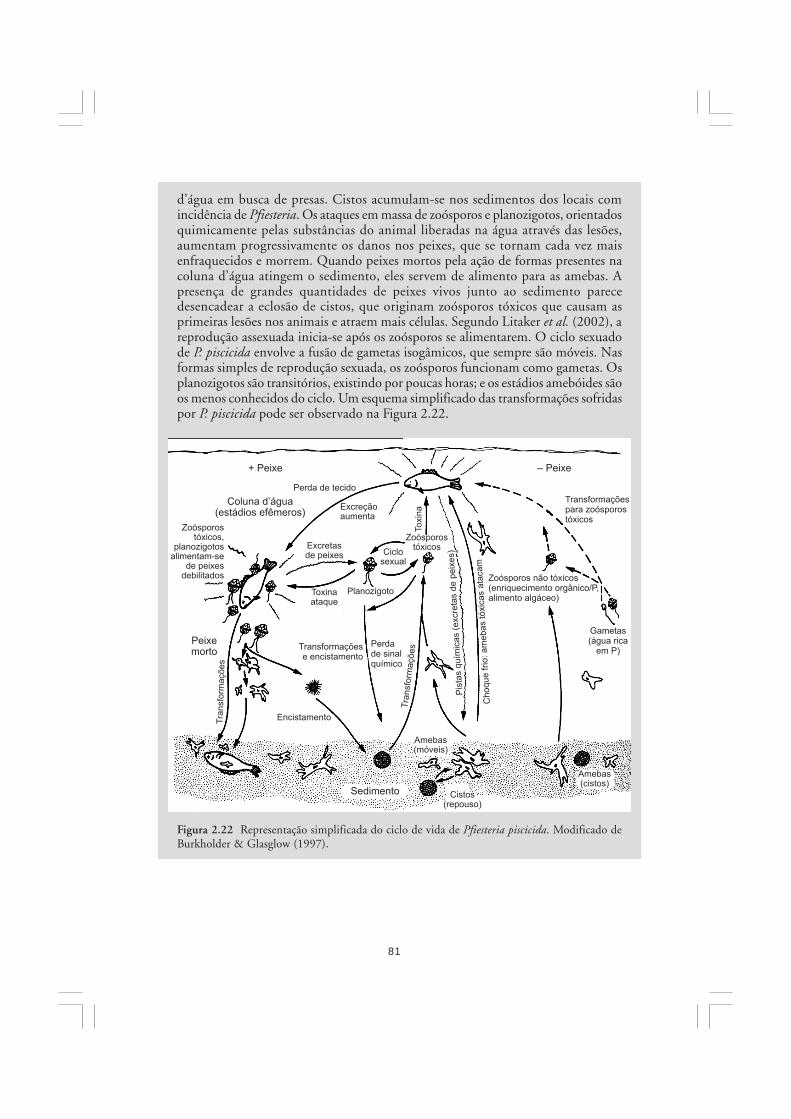
81
d’água em busca de presas. Cistos acumulam-se nos sedimentos dos locais comincidência de Pfiesteria. Os ataques em massa de zoósporos e planozigotos, orientadosquimicamente pelas substâncias do animal liberadas na água através das lesões,aumentam progressivamente os danos nos peixes, que se tornam cada vez maisenfraquecidos e morrem. Quando peixes mortos pela ação de formas presentes nacoluna d’água atingem o sedimento, eles servem de alimento para as amebas. Apresença de grandes quantidades de peixes vivos junto ao sedimento parecedesencadear a eclosão de cistos, que originam zoósporos tóxicos que causam asprimeiras lesões nos animais e atraem mais células. Segundo Litaker et al. (2002), areprodução assexuada inicia-se após os zoósporos se alimentarem. O ciclo sexuadode P. piscicida envolve a fusão de gametas isogâmicos, que sempre são móveis. Nasformas simples de reprodução sexuada, os zoósporos funcionam como gametas. Osplanozigotos são transitórios, existindo por poucas horas; e os estádios amebóides sãoos menos conhecidos do ciclo. Um esquema simplificado das transformações sofridaspor P. piscicida pode ser observado na Figura 2.22.
+ Peixe
Coluna d’água(estádios efêmeros)
Peixemorto
– Peixe
Sedimento
Perda de tecido
Excreçãoaumenta
Zoósporostóxicos,
planozigotosalimentam-se
de peixesdebilitados
Amebas(móveis)
PlanozigotoToxinaataque
Excretasde peixes Ciclo
sexual
To
xin
a
Zoósporostóxicos
Tra
nsfo
rmações
Perdade sinalquímico
Encistamento
Transformaçõese encistamento
Tra
nsfo
rmações
Cistos(repouso)
Amebas(cistos)
Pis
tas
quím
icas
(excre
tas
de
peix
es)
Choque
frio
:am
ebas
tóxic
as
ata
cam
Zoósporos não tóxicos(enriquecimento orgânico/P,alimento algáceo)
Gametas(água rica
em P)
Transformaçõespara zoósporostóxicos
Figura 2.22 Representação simplificada do ciclo de vida de Pfiesteria piscicida. Modificado deBurkholder & Glasglow (1997).

82
Os dinoflagelados são notáveis por suas associações simbióticas, sobretudo com inver-tebrados marinhos. Alguns desses dinoflagelados, conhecidos como zooxantelas, se associama corais formadores de recifes e são essenciais para o desenvolvimento de todo o ecossistemarecifal. Espécies do gênero Symbiodinium são particularmente comuns em associações simbió-ticas, sendo encontradas em corais, esponjas e bivalves gigantes.
Muitas espécies de dinoflagelados são capazes de realizar florações nocivas e várias sãoprodutoras de ficotoxinas potentes. As ficotoxinas apresentadas por dinoflagelados incluemneurotoxinas (por exemplo, brevetoxina), toxinas diarréicas (por exemplo, ácido okadaiko)e toxinas paralisantes (por exemplo, saxitoxina, Figura 2.23). A formação de floraçõesintensas, conhecidas como marés vermelhas, e a produção de ficotoxinas não são feiçõesexclusivas de dinoflagelados; há muitas espécies de outros grupos taxonômicos com taiscapacidades. Entretanto, o número de espécies de dinoflagelados capazes de gerar floraçõese produzir ficotoxinas é bastante elevado, se comparado com outros grupos. A última listade espécies tóxicas divulgada pela Comissão Oceanográfica Intergovernamental da UNESCOindica a existência de 89 espécies tóxicas no fitoplâncton marinho, das quais 63 são dino-flagelados (IOC, 2003). Fatos como esse tendem a levar o público leigo a fazer associaçõesindevidas entre marés vermelhas, toxidez e dinoflagelados. Mais do que isso, o público leigotende a associar esses fenômenos como sendo exclusivamente causados por dinoflagelados,o que não procede.
OHN
OH2N
NH2
NHNOH
OH
HN
N
Cerca de 30 espécies fotossintetizantes de dinoflagelados são capazes de realizar biolu-minescência. Trata-se das únicas espécies conhecidas de organismos fotossintetizantes (oumixotróficos, em alguns casos) que também são bioluminescentes, sendo pertencentes aosgêneros Lingulodinium (= Gonyaulax), Protogonyaulax, Pyrodinium, Pyrocystis e Ceratium.Há também espécies bioluminescentes em dois gêneros não fotossintetizantes: Noctiluca eProtoperidinium. Quimicamente, a bioluminescência envolve reação exergônica de oxigêniomolecular com diferentes substratos e enzimas, resultando em fótons de luz visível (≅ 50 kcal).Uma substância (luciferina) é oxidada por uma enzima (luciferase), resultando em umproduto eletricamente excitado que na decomposição irá emitir um fóton (Lee, 1999). Aimportância da bioluminescência nos dinoflagelados ainda não foi bem compreendida. Duashipóteses postulam que a bioluminescência seria um mecanismo selecionado como resposta àpredação. Na chamada “hipótese do susto”, a estimulação mecânica de um dinoflagelado porum predador desencadearia um lampejo que assustaria o predador e o faria fugir com osapêndices retraídos. Outra hipótese, a chamada “hipótese do alarme de roubo”, pondera quepossíveis predadores movimentando-se por entre os dinoflagelados os estimulariam mecani-camente (principalmente através da movimentação da água) a emitirem lampejos, o quechamaria a atenção dos predadores de seus predadores. Entretanto, no mundo microscópicodos dinoflagelados, as hipóteses não foram adequadamente testadas, exceto por meio de algunspoucos experimentos com copépodos. Por fim, pode-se considerar que a produção de luz talvez
Figura 2.23 Molécula de saxitoxina, uma toxinaparalisante produzida por dinoflagelados.

83
não apresente (nem tenha nunca apresentado) uma função, sendo um subproduto acidentalde intercâmbios energéticos, como acredita-se que seja o caso do fungo basidiomiceto Mycenalux-coeli, o qual espontaneamente exibe uma bela luz esverdeada à noite.
Devido à grande importância ecológica dos dinoflagelados, bem como seus possíveis efeitossobre o homem pela formação de florações e produção de ficotoxinas, o grupo vem recebendoatenção especial quanto aos esquemas de classificação. De acordo com Hoek et al. (1995), umaúnica classe é reconhecida na divisão: classe Dinophyceae. Esta é dividida em 12 ordens:Gymnodiniales, Gloeodiniales, Thoracosphaerales, Phytodiniales, Dinotrichales, Diamoebiales,Noctilucales, Blastiodinales, Synediales, Peridiniales, Dinophysiales e Prorocentrales.
É difícil apontar alguns dos principais gêneros de dinoflagelados dentre as centenasexistentes, pois muitos estão envolvidos com processos extremamente importantes na natureza,afetando inclusive o homem. Os seguintes gêneros têm importâncias distintas: Alexandrium,Amphidinium, Ceratium, Cystodinium, Dinophysis, Diplopsalis, Gonyaulax, Gymnodinium,Noctiluca, Ornithocercus, Oxyrrhis, Peridinium, Prorocentrum, Protoperidinium e Symbiodinium.
2.2.8 DIVISÃO OCHROPHYTA
Esta divisão envolve grande número de grupos de algas que já constituíram, no passado,unidades taxonômicas elevadas, tratadas de forma independente ou não relacionada. A divisãoinclui diferentes grupos de algas, como as diatomáceas, as eustigmatofíceas, as rafidofíceas, aspelagofíceas, os silicoflagelados, os pedinelídeos, as tribofíceas (ou xantofíceas) e as feofíceas(algas pardas), dentre outros grupos, reunindo linhagens “castanho-douradas” de algas. Emboraatualmente haja consenso sobre a adequação de reunir os grupos de algas mencionados numaúnica divisão, há discordâncias entre os autores sobre o nome da divisão Ochrophyta (= “plantasde cor ocre”). Assim, encontra-se na literatura um nome alternativo para a divisão, Heterokontophyta(= “plantas com flagelos diferentes”), que valoriza as características dos flagelos dos membrosda divisão para defini-la. Entretanto, outros protistas não-algáceos, como os oomicetos, oslabirintulomicetos e os opalinídeos (dentre outros), também compartilham característicasem comum com os membros desta divisão, sobretudo os flagelos heterocontes. Evidênciasmoleculares mostram que os protistas heterocontes podem ser reunidos num táxon maiselevado, o sub-reino Stramenopiles (= Heterokonta). Assim, os termos estramenópilas e hete-rocontes referem-se a um conjunto maior do que o das algas. Nesse sentido, as designaçõesocrófita e heterocontófita são mais restritas, sendo mais convenientes para fazer referência aalgas heterocontes. O termo ocrófita também exclui, de forma discreta, os demais protistasnão-algáceos do sub-reino Stramenopiles, os quais estão mais diretamente relacionados àsocrófitas do que qualquer outro grupo de protistas heterotróficos. Por fim, há tambémdiscordâncias entre alguns autores sobre os grupos de organismos que devem constituir adivisão Ochrophyta (ou Heterokontophyta) e, dependendo da fonte consultada, os compo-nentes indicados podem ser diferentes. Nesta obra será adotada a classificação proposta porReviers (2006).
Os membros da divisão Ochrophyta variam desde células microscópicas até algasgigantes de dezenas de metros de comprimento, com diferenciação de tecidos. A clorofila aestá presente na maior parte das ocrófitas, mas há também algumas formas heterotróficasnão pigmentadas. Nas espécies pigmentadas (maioria das espécies da divisão), os pigmentosacessórios e protetores dos fotossistemas incluem a clorofila c e os carotenóides, como afucoxantina e a vaucheriaxantina. Os produtos de reserva consistem em gotículas de gordurano citoplasma e/ou um polissacarídero solúvel e derivado de glicose formado por ligaçõesglicosídicas do tipo ß-1,3: a crisolaminarina ou laminarina. Esta é virtualmente a mesma

84
substância apresentada pelas primnesiofíceas, ocorrendo nos vacúolos citoplasmáticos. Háusualmente dois flagelos heteromórficos (daí o nome alternativo da divisão, Heterokontophyta),um deles dotado de três ramificações na extremidade, com aspecto plumoso, conhecido comomastigonema. Mais do que dotados de flagelos meramente diferentes, na realidade adesignação de um organismo como heteroconte significa que um de seus flagelos apresentamastigonemas tubulares tripartidos, o que confere aspecto plumoso ao flagelo, em contrastecom o segundo flagelo; esta característica pode ter sido perdida ou os mastigonemastripartidos podem ter se tornado secundariamente bipartidos em algumas espécies (Reviers,2006).
A cobertura das células varia consideravelmente entre as espécies, ocorrendo desdeestruturas de sílica (exemplo: diatomáceas) até paredes de celulose e ácido algínico (algasfeofíceas). A reprodução sexuada é comum na divisão Ochrophyta e vários tipos de ciclo devida ocorrem nos diferentes grupos que a compõem.
Segundo Medlin et al. (1997), as ocrófitas surgiram no final Permiano, há 260 milhõesde anos. O número de espécies da divisão é de difícil determinação. Há mais de 250 gênerose 10.000 espécies somente de diatomáceas, e estimativas que variam entre 900 e 2.000espécies de feofíceas. Diversos grupos que constituem a divisão Ochrophyta contêm poucasespécies, mas presume-se haver muitas espécies ainda não descritas. Dentre os grupos queconstituem a divisão, alguns são primariamente dulciaqüícolas (exemplo: synurofíceas),outros são tipicamente marinhos (exemplo: feofíceas) e outros, como as diatomáceas, sãocomuns em ambientes marinhos e dulciaqüícolas. As algas pardas (feofíceas) incluem asmaiores algas conhecidas. A diversidade morfológica e química das ocrófitas é a maior dentretodas as divisões de algas.
Há enorme diversidade de características e grande volume de informações sobre cadaclasse que constitui a divisão Ochrophyta. Assim, as informações sobre as 13 classes reconhe-cidas são apresentadas a seguir, com ênfase apenas em diatomáceas (por sua grande impor-tância no plâncton marinho). As demais classes são tratadas resumidamente por apresentaremimportância menor no plâncton, por envolverem poucas espécies marinhas ou, ainda, porhaver pequena disponibilidade de informações sobre elas.
2.2.8.1 CLASSE BACILLARIOPHYCEAE
A principal característica morfológica distintiva das diatomáceas diz respeito a suasfrústulas silicosas, as quais representam caráter importantíssimo para sua classificação devidoa seus ricos detalhes de forma e ornamentação (Figura 2.24). A palavra diatomácea é tambémuma alusão às frústulas, pois vem do grego dia, “de um extremo ao outro”, e tomos, “corte”;frústulas são estruturas bivalves complexas, que se encaixam formando uma espécie de tecaque protege a célula. Diatoma é também o nome de um gênero cosmopolita de diatomáceasbentônicas. As valvas têm tamanhos desiguais, sendo a maior chamada de epivalva e a menor,de hipovalva. A região de contato entre as valvas é o cíngulo (Figura 2.25).
Round et al. (1990) reconhecem a existência de 285 gêneros, envolvendo 10.000-12.000 espécies conhecidas. O número exato de espécies é impreciso devido à existência demuitos casos de sinonímia. Essa estimativa faz das diatomáceas o segundo maior grupo emnúmero de espécies, menor apenas que as algas verdes. Admite-se que ainda há muitas espéciesde diatomáceas a serem descritas, e Round & Crawford (1990) sugerem que pode haver100.000 espécies de diatomáceas na natureza.

85
Figura 2.24 Prancha de diatomáceas preparada pelo naturalista Ernst Haeckel em 1904. 1. Triceratiumdigitale; 2. Navicula lyra; 3. Navicula excavata; 4. Triceratium mirificum; 5. Triceratium pentacrinus; 6.Actinoptychus constellatus; 7. Aulacodiscus mammosus; 8. Navicula wrightii; 9. Auliscus crucifer; 10.Biddulphia pulchella; 11. Auliscus craterifer; 12. Auliscus mirabilis; 13. Aulacodiscus grevilleanus; 14. Surirellamacraeana; 15. Denticella regia; 16. Asterolampra eximia; 17. Actinoptychus heliopelta; 18. Plagiogrammabarbadense; 19. Pinnularia mulleri; 20. Biddulphia granulata; 21. Triceratium pentacrinus; 22. Triceratiummoronense. Obtido de Wikipedia (2006).

86
Frústula
Pseudosepto
EpitecaSepto
Hipoteca
Epivalva
Epicíngulo
Hipocíngulo
Hipovalva
Figura 2.25 Nomenclatura para as diferentes partes de frústulas de diatomáceas. Modificado de Hasle &Syvertsen (1997).
Tradicionalmente, as diatomáceas são classificadas em dois grandes grupos, em funçãoda morfologia da frústula: diatomáceas cêntricas, com frústulas de simetria radial; e diato-máceas penadas, com frústulas de simetria bilateral (Figura 2.26). Algumas diatomáceaspenadas são dotadas de uma ranhura ao longo do eixo longitudinal da frústula, conhecidacomo rafe (Figura 2.26a). Essa estrutura relaciona-se à produção de movimentos de desliza-mento sobre o substrato. Diatomáceas penadas desprovidas de rafe e diatomáceas cêntricasnão são capazes de realizar tais movimentos. Estudos moleculares indicam que as diatomáceascêntricas e as penadas desprovidas de rafe são parafiléticos. Assim, os esquemas de classificaçãoadotados atualmente (fundamentalmente derivados de informações sobre morfologia) deve-rão ser revistos em breve, pois não são consistentes com os dados moleculares já disponíveis.
b) 20 µm20 µma)
Figura 2.26 Tipos morfológicos básicos de diatomáceas: (a) diatomácea penada, com simetria bilateral;(b) diatomácea cêntrica, com simetria radial. A diatomácea penada apresenta rafe. Reproduzido de Graham& Wilcox (2000).
As frústulas de diatomáceas são caracteristicamente muito ricas em detalhes e orna-mentação, tendo grande valor taxonômico. Poros, projeções, depressões, processos labiais,etc. são abundantes nas células (Figura 2.27), e a observação cuidadosa dessas feições morfo-
Pseudo-septo

87
lógicas permite identificação segura das espécies. A visualização dos detalhes das frústulasnormalmente exige o uso de microscopia eletrônica ou tratamento químico das frústulasantes da observação por microscopia óptica.
1 2
3 4
5a 5b
Figura 2.27 Imagens de microscopia eletrônica de varredura de algumas diatomáceas penadas do gêneroThalassionema, ressaltando as diferenças entre as frústulas das espécies. Extremidades das frústulas de (1)T. javanicum, (2) T. nitzschioides, (3) T. bacillare, (4) T. pseudonitzschioides e (5a) T. frauenfeldii. (5b)Porção intermediária da valva de T. frauenfeldii. Reproduzido de Hasle & Syvertsen (1997).
Assim como as algas verdes e as cianobactérias, as diatomáceas são ubíquas: ocorremno mar, em água salobra, em água doce e em ambientes terrestres úmidos. Elas podemapresentar hábitos planctônicos ou bentônicos. As diatomáceas bentônicas podem ser epilí-ticas, epizoóicas, epífitas, cavernícolas; podem viver sobre detritos flutuantes, sedimentos,matéria orgânica e neve. Diatomáceas podem participar da formação de biofilmes micro-bianos coloridos (limo), fazendo parte do chamado microfitobentos, e podem ser encontradasem associações simbióticas com foraminíferos e esponjas. Em ambientes polares, muitasdiatomáceas (sobretudo formas penadas) ocorrem aderidas ao gelo flutuante e constituem-se numa fonte fundamental de alimento para animais e protistas heterotróficos naquelesambientes.
As células de diatomáceas variam desde 2,0 µm (picoplâncton) até 2,0 mm de diâmetro(mesoplâncton). Diatomáceas apresentam um núcleo mais ou menos centralizado. Os plastosapresentam clorofilas a e c, â-caroteno, fucoxantina, diatoxantina e diadinoxantina comoseus principais pigmentos fotossintéticos. As lamelas apresentam três tilacóides e os plastossão envolvidos por quatro membranas, a mais externa é conectada ao retículo endoplasmático.Os plastos geralmente são pequenos, discóides e numerosos em diatomáceas cêntricas, aopasso que as diatomáceas penadas tendem a apresentar apenas dois plastos, grandes e

88
achatados. Pirenóides são comuns em plastos de diatomáceas. Os produtos de reserva são acrisolaminarina e gotículas de óleo. Estes apresentam também a função de contribuir parao controle do posicionamento de diatomáceas planctônicas na coluna d’água; a acumulaçãode mais óleos diminui a densidade da célula e aumenta sua flutuabilidade.
Diatomáceas são desprovidas de movimentos, exceto gametas uniflagelados de partedas espécies e diatomáceas penadas dotadas de rafe. Essas espécies podem lançar substânciasmucilaginosas com capacidade adesiva; microfibrilas protéicas associadas ao material secretadoorganizam-se rapidamente, proporcionando uma forma de contração, que produz o deslo-camento da célula. Diatomáceas que se deslocam sobre sedimento deixam rastros de muci-lagem que contribuem para estabilizar o substrato, que pode eventualmente formar pequenasplacas compactadas de sedimento. O papel das diatomáceas na ecologia de sedimentos é muitorelevante, pois elas podem ser as principais espécies constituintes do microfitobentos em várioslocais. Diversas diatomáceas praticam migração vertical diária em diferentes estratos desedimento. Não se conhecem sistemas de fotorrecepção, como manchas ocelares, nas espéciesmóveis de diatomáceas.
As frústulas de diatomáceas são muito resistentes à dissociação em meio aquático,conferindo-lhes boa conservação na natureza. Há registros fósseis de diatomáceas de 150milhões de anos, ou seja, desde o início do Cretáceo. Isso sugere que as diatomáceas devemter se originado milhões de anos antes. Medlin et al. (1997) sugerem que as diatomáceaspodem ter derivado de seus ancestrais há cerca de 250 milhões de anos. Todos os registrosfósseis mais antigos são de diatomáceas cêntricas, ao passo que os primeiros registros dediatomáceas penadas são mais recentes, de 70 milhões de anos, no final do Cretáceo. As formasmais antigas de diatomáceas fósseis de ambientes dulciaqüícolas são penadas, registradas emdepósitos de 60 milhões de anos. Somente no Mioceno (24 milhões de anos) as diatomáceaspassaram a ser abundantes em registros fósseis, sugerindo possível dominância em parte dossistemas aquáticos (Graham & Wilcox, 2000).
Células vegetativas de diatomáceas são diplóides, reproduzindo-se predominantementede forma assexuada, por meio da divisão celular vegetativa. Nesse processo, cada uma das clelulasfilhas recebe uma das valvas da célula mãe e sintetiza a segunda valva. As clelulas filhas fazemda valva recebida a epivalva, de forma a produzir uma segunda valva menor, a hipovalva(Figura 2.28). Há, portanto, tendência à redução do tamanho médio da população da espéciecom o avanço do processo de reprodução vegetativa ao longo das gerações. Ao atingir tamanhocelular crítico, haveria a deflagração do processo de reprodução sexuada, por intermédio doqual as células vegetativas sofrem meiose e formam gametas. No caso de diatomáceas cêntricas,ocorre oogamia. Algumas células comportam-se como femininas e geram apenas uma ouduas oosferas (duas ou três oosferas morrem); cada célula masculina produz numerososanterozóides (4-128) dotados de apenas um flagelo, como resultado da ocorrência de mitoseapós a meiose. Os anterozóides são lançados na água e fundem-se à oosfera, gerando umzigoto que a seguir se desenvolve numa célula grande conhecida como auxósporo. Este nãoherda as frústulas da célula parental feminina e sintetiza novas frústulas de maior tamanho,restabelecendo uma média de tamanho típica da espécie (Figura 2.29). Diatomáceas penadastambém realizam reprodução sexuada ao atingirem tamanho celular criticamente pequeno,mas não produzem gametas flagelados, nem oosferas. Ocorre meiose e quatro núcleos ha-plóides são formados em cada célula (dos quais dois se desintegram posteriormente). Diato-máceas penadas pareiam-se e mantêm-se unidas pela ação de mucilagem extracelular; forma-se espaço entre as valvas que compõem as frústulas das duas células e ocorre fusão dos pro-toplastos. Isso permite que os núcleos se movimentem entre as células; núcleos haplóides

89
das células diferentes sofrem fusão e dois auxósporos com carga cromossomial 2N sãoformados. Os auxósporos dão origem a células vegetativas de maior tamanho (Figura 2.30).O processo de reprodução sexual de diatomáceas penadas é especial e difere dos três tiposbásicos quanto à formação de gametas (isogamia, anisogamia e oogamia), sendo chamado poralguns autores de gametangiogamia, que é uma forma de conjugação, semelhante àquelapraticada por outras algas. É importante ressaltar que diatomáceas também podem sofrerautogamia (fusão de dois núcleos haplóides irmãos) e apogamia (ausência de fusão de gametas).
A formação de auxósporos é um meio de recuperar o tamanho médio das populaçõesde diatomáceas, sendo muitas vezes assumido como processo universal entre as espécies.Entretanto, esse processo ainda não foi documentado em várias espécies. Além disso, aexperiência com cultivos em laboratório, obtida por muitos pesquisadores independentes,tem demonstrado que diversas espécies de diatomáceas não sofrem redução progressiva detamanho em função da reprodução vegetativa no decorrer de gerações consecutivas. Issosignifica que os mecanismos de síntese de valvas podem não conduzir à redução das célulasou, se isso ocorrer, pode haver aumento vegetativo das células vivas independente da açãode auxósporos, ainda não conhecido em detalhes (Nagai et al., 1995; Mann & Chepurnov,2004). Diatomáceas também podem formar cistos de repouso ou resistência, os quais sãomorfologicamente semelhantes às células vegetativas, mas com frústulas mais grossas egeralmente com formas mais arredondadas e menos elaboradas. Os cistos de diatomáceassão densos e tendem a se acumular nos sedimentos, podendo germinar quando as condiçõestornarem-se novamente adequadas para as células vegetativas.
A acumulação milenar de frústulas (de células vegetativas mortas e cistos) em certosambientes rasos deu origem a depósitos silicosos do mineral diatomito (também conhecidopopularmente como farinha-fóssil e terra de diatomáceas) e também a vasas de diatomáceasem áreas profundas dos oceanos. O diatomito é utilizado comercialmente na fabricação deelementos filtrantes, tintas, abrasivos, creme dental, isolantes, etc. Além disso, depósitos dediatomito têm grande importância como indicadores estratigráficos para a exploração petro-lífera e na reconstituição de paleoambientes. A inalação de diatomito pode acarretar sériosproblemas respiratórios, devendo-se tomar cuidados extremos nas áreas de mineração.
a)
b)
c)
Figura 2.28 Representação esquemática do processo de redução progressiva do tamanho médio de célulasde diatomáceas, após gerações sucessivas originadas por reprodução vegetativa.

90
Divisão celularvegetativa
Célula vegetativa“normal”
2N
N
Célulainicial
Auxósporo
Fusão
Meiose
Tamanhocelularcrítico
Figura 2.29 Representação do processo de reprodução sexuada por oogamia, realizado por diatomáceascêntricas. Modificado de Hasle & Syvertsen (1997).
Muitas diatomáceas são cultivadas para uso na alimentação de animais marinhos deinteresse comercial em aqüicultura (vide Capítulo 8). Várias espécies apresentam alto valornutricional e facilidade de cultivo, como Phaeodactylum tricornutum e Thalassiosira fluviatilis,dentre outras. O alto valor nutricional de diatomáceas deriva, pelo menos em parte, dapresença de ácidos graxos poliinsaturados (como o ácido eicosapentaenóico) em altas con-centrações.
Poucas espécies de diatomáceas estão relacionadas a efeitos nocivos. Algumas espéciesde Chaetoceros podem causar danos no aparato respiratório de peixes em caso de floraçõesintensas. Algumas espécies do gênero Pseudo-nitzschia são produtoras de ácido domóico(Figura 2.31), substância envolvida no envenenamento amnésico por moluscos e que podecausar a morte de humanos se ingerida em grandes concentrações.
É virtualmente impossível listar os principais gêneros de diatomáceas sem omitir algunsdeles, exceto se um rol exaustivamente grande for apresentado. Uma lista de gêneros impor-tantes, com inúmeras omissões, pode ser constituída por Asteriomphalus, Asterionella,Asterionellopsis, Biddulphia, Chaetoceros, Cocconeis, Coscinodiscus, Cyclotella, Cymatopleura,Cymbella, Epithemia, Fragillaria, Gomphonema, Gyrosigma, Licmorpha, Melosira, Navicula,Nitzschia, Odontella, Pinnularia, Pseudo-nitzschia, Rhizosolenia, Rhopalodia, Skeletonema,Stauroneis, Stephanodiscus, Synedra, Tabellaria, Thalassiosira e Triceratium.

91
Divisão celularvegetativa
Célula vegetativa“normal”
2N
NCélulainicial
Auxósporo
Fusão
Meiose
Tamanhocelularcrítico
Figura 2.30 Representação do processo de reprodução sexuada por gametangiogamia, realizado pordiatomáceas penadas. Modificado de Hasle & Syvertsen (1997).
HN
H3CHOOC
COOH
HOOC
CH3
Figura 2.31 Molécula de ácido domóico, uma toxina amnésica produzida por diatomáceas do gêneroPseudo-nitzschia.
2.2.8.2 CLASSE BOLIDOPHYCEAESabe-se pouco a respeito desta classe, constituída por um único gênero conhecido,
Bolidomonas, e apenas duas espécies. Trata-se de flagelados marinhos pertencentes aopicoplâncton, com diâmetro celular de apenas 1,2 µm, dotados de notável capacidade dedeslocamento proporcionada por um flagelo anterior longo, com mastigonemas bipartidos,e por um flagelo posterior pequeno e liso (Figura 2.32). Os movimentos rápidos das célulasconstituem o caráter utilizado para nomear o gênero e a classe (“bólide”, que significa rápido).

92
As duas espécies conhecidas de bolidofíceas são autotróficas e aparentemente não apresentammixotrofia.
FiT
HT
PlL
Gen
Fl1
Di
Fl2MB 1-2
MB 3-4N
M
Figura 2.32 Características morfológicas de Bolidomonas. Di = dictiossomo; FiT = filamentos terminaisdos mastigonemas; Fl1 = flagelo anterior; Fl2 = flagelo posterior; Gen = genoma plastidial em forma deanel, nas duas extremidades do plasto; HT = haste tubular dos mastigonemas bipartidos; L = lamela formadade três tilacóides; M = mitocôndria; MB 1-2 = membranas mais internas do envoltório plastidial; MB 3-4 = membranas mais externas do envoltório plastidial (a quarta membrana tem continuidade com oenvelope nuclear); N = núcleo; Pl = plasto. Notar que a inserção dos flagelos é ventral. Reproduzido deReviers (2006).
Não há informações sobre a reprodução do grupo nem se conhecem fósseis. Análisesmoleculares indicam que as bolidofíceas são o grupo filogeneticamente mais próximo dasdiatomáceas. Características da inserção dos flagelos na célula e a distribuição das organelasconstituem-se em particularidades que separam o grupo das demais ocrófitas.
Bolidomonas pacifica tem distribuição ao longo do Pacífico equatorial e B. mediterranicaé restrita ao mar Mediterrâneo.
2.2.8.3 CLASSE PARMOPHYCEAE
A classe é constituída apenas por espécies marinhas, as quais compreendem indivíduoscocóides de 2,0 a 5,5 µm de diâmetro, portanto, pertencentes ao nanoplâncton. As células sãodotadas de cinco a oito placas de natureza silicosa, característica peculiar do grupo (Figura 2.33).
Não se conhece a reprodução das parmofíceas e, apesar da cobertura silicosa das células,não há informações sobre fósseis no grupo. A falta de informações básicas sobre o grupotorna sua acomodação na divisão Ochrophyta sujeita a questionamentos. A presença de placasde silício nas células e a ausência de células vegetativas flageladas sugerem parentesco com asdiatomáceas, ainda não confirmado por análises moleculares.
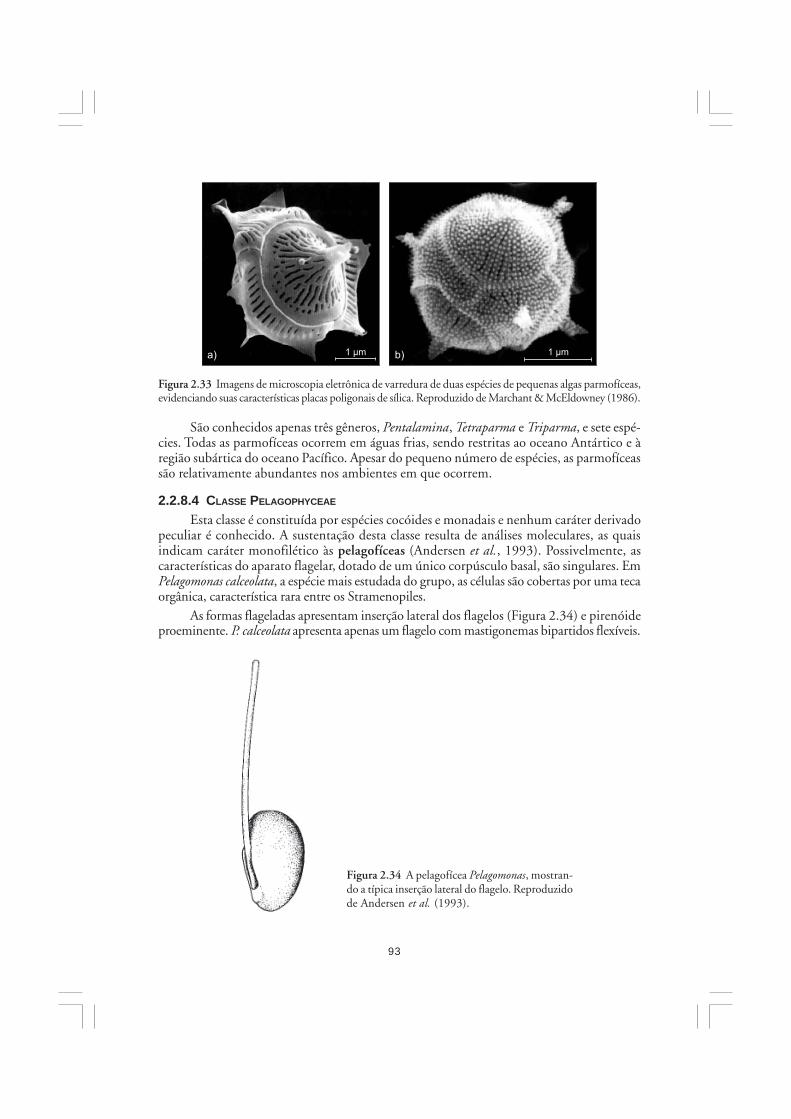
93
a) b)1 µm 1 µm
Figura 2.33 Imagens de microscopia eletrônica de varredura de duas espécies de pequenas algas parmofíceas,evidenciando suas características placas poligonais de sílica. Reproduzido de Marchant & McEldowney (1986).
São conhecidos apenas três gêneros, Pentalamina, Tetraparma e Triparma, e sete espé-cies. Todas as parmofíceas ocorrem em águas frias, sendo restritas ao oceano Antártico e àregião subártica do oceano Pacífico. Apesar do pequeno número de espécies, as parmofíceassão relativamente abundantes nos ambientes em que ocorrem.
2.2.8.4 CLASSE PELAGOPHYCEAE
Esta classe é constituída por espécies cocóides e monadais e nenhum caráter derivadopeculiar é conhecido. A sustentação desta classe resulta de análises moleculares, as quaisindicam caráter monofilético às pelagofíceas (Andersen et al., 1993). Possivelmente, ascaracterísticas do aparato flagelar, dotado de um único corpúsculo basal, são singulares. EmPelagomonas calceolata, a espécie mais estudada do grupo, as células são cobertas por uma tecaorgânica, característica rara entre os Stramenopiles.
As formas flageladas apresentam inserção lateral dos flagelos (Figura 2.34) e pirenóideproeminente. P. calceolata apresenta apenas um flagelo com mastigonemas bipartidos flexíveis.
Figura 2.34 A pelagofícea Pelagomonas, mostran-do a típica inserção lateral do flagelo. Reproduzidode Andersen et al. (1993).

94
Todas as espécies são marinhas e algumas podem ser encontradas em águas salobrasde sistemas estuarinos. A maioria ocorre no plâncton, mas há também pelagofíceas ben-tônicas. Como as espécies são pequenas, variando entre 1,0 e 3,0 µm de diâmetro, as formasplanctônicas são posicionadas entre o nanoplâncton (minoria) e o picoplâncton (maioria).Há também formas filamentosas e coloniais.
As pelagofíceas produzem zoósporos, mas a reprodução sexuada não é conhecida nogrupo. Ignora-se também a possível existência de fósseis.
São conhecidos 10 gêneros (Ankylochrysis, Aureococcus, Aureoumbra, Chrysocystis,Chrysonephos, Chrysoreinhardia, Nematochrysopsis, Pelagococcus, Pelagomonas e Sarcinochrysis)e 15 espécies. Aureoumbra é capaz de formar florações, e a espécie bentônica Nematochrysopsismarina eventualmente produz proliferações intensas no mar Mediterrâneo.
2.2.8.5 CLASSE DICTYOCHOPHYCEAE
As dictiocofíceas não apresentam caráter derivado marcante, porém, as análises mole-culares já realizadas confirmam o caráter monofilético do grupo e sua separação das demaisclasses de Ochrophyta. O único caráter peculiar do grupo é a posição do complexo de Golgina célula, que se encontra disperso ou na parte posterior da célula; nas demais ocrófitas ocomplexo de Golgi encontra-se na parte anterior da célula, entre o núcleo e a membranaplasmática.
A grande maioria das espécies é monadal e ocorre no mar. Um dos flagelos é pleurone-mático e longo e o segundo flagelo é curto e liso. As espécies são planctônicas e fotossin-tetizantes, algumas das quais são mixotróficas. Dois gêneros (Ciliophrys e Pteridomonas, antesclassificados como heliozoários) são heterotróficos e fagotróficos.
A classe é formada por três ordens (Dictyochales, Pedinellales e Rhizochromulinales).Os silicoflagelados fazem parte da ordem Dictyochales e a ocorrência de um esqueleto externode sílica é sua característica mais marcante (Figura 2.35). Atualmente, os silicoflagelados sãorepresentados por apenas um gênero, Dictyocha, com poucas espécies, embora com ampladistribuição nos oceanos mundiais, sobretudo em águas mais frias, onde podem ser bastanteabundantes e até formar florações. Representantes do gênero Dictyocha são caracteristicamentedotados de um citoplasma altamente reticulado. Porém, os silicoflagelados são extremamenteabundantes em registros fósseis desde 120 milhões de anos (meio do Cretáceo), tendo sidodescritos 20 gêneros que atingiram seu ápice no Mioceno. Estima-se a existência de 100.000espécies fósseis de silicoflagelados.
A ordem Pedinellales compreende 12 gêneros e 27 espécies distribuídas principalmenteno mar, embora algumas espécies ocorram em água doce e em água salobra. Os pedinelídeossão caracterizados por simetria radial perfeita ao longo de seu eixo longitudinal. A maioriatem hábitos planctônicos, mas há também espécies bentônicas. Fotoautotrofia, mixotrofiae fagotrofia são reconhecidas no grupo. Os pedinelídeos apresentam um flagelo anterior,que emerge de uma pequena depressão, e uma pequena estrutura em forma de bastão,conhecida como pedicelo, localizada numa extensão que se assemelha a uma asa minúscula.O pedicelo é uma estrutura comparável ao haptonema das primnesiofíceas em termos decomplexidade e funções. As espécies fotossintetizantes apresentam plastos numerosos (cadaum deles com um pirenóide), dispostos na periferia da célula, formando uma espécie de anel.A terceira ordem, Rhizochromulinales, é representada por apenas uma espécie, Rhizochromulinamarina, organismo amebóide cuja distribuição conhecida restringe-se ao mar Mediterrâneo.

95
a)a) b)b)
c)c) d)d) 20 µm
20 µm
20 µm
10 µm
Figura 2.35 Esqueletos de sílica altamente perfurados de dictiocofíceas fósseis do final do Cretáceo. (a)Dictyocha; (b) Vallacerta; (c) Lyramula; (d) Corbisema. Reproduzido de Graham & Wilcox (2000).
2.2.8.6 CLASSE PINGUIOPHYCEAE
Esta classe é formada por espécies cocóides, palmelóides, monadais ou loricadas, todasmarinhas (Figura 2.36). As pinguiofíceas são extremamente ricas em ácidos graxos poliinsa-turados da série ômega-3, característica que dá nome ao grupo (do latim pingue, “graxo”,“gordo”). A presença de altas concentrações de ácidos graxos (sobretudo o ácido eicosapen-taenóico – EPA) tem motivado estudos sobre o cultivo das espécies, com vistas a seu usocomo alimento em aqüicultura. A classe é pequena, formada por apenas cinco gênerosmonoespecíficos: Glossomastix chrysoplasta, Phaeomonas parva, Pinguiochrysis pyriformis,Pinguiococcus pyrenoidosus e Polypodochrysis teissieri. Três espécies (dos gêneros Phaeomonas,Pinguiochrysis e Pinguiococcus) são planctônicas e as demais são bentônicas. A classe foi descritarecentemente (Kawachi et al., 2002), acomodando uma espécie cuja posição era incertadentro das Chrysophyceae (Polypodochrysis teissieri) e quatro espécies novas.
As pinguiofíceas são fotoautotróficas e apresentam um plasto por célula (excetoGlossomatix, que possui dois plastos por célula), dotado de um pirenóide, que pode ser saliente,pedunculado ou estar imerso no estroma. Além das clorofilas a e c, as pinguiofíceas possuemfucoxantina (carotenóide principal), violaxantina, zeaxantina e â-caroteno como pigmentosfotossintetizantes. Carotenóides presentes em outros grupos de ocrófitas (ex.: neoxantina,diadinoxantina, heteroxantina e vaucheriaxantina) não estão presentes nas pinguiofíceas. Osplastos das cinco espécies são circundados pela membrana do retículo endoplasmático plastidial,a qual é contínua ao envelope nuclear, exceto para Glossomatix (Kawachi et al., 2002).
As espécies apresentam diferenças marcantes quanto ao envoltório das células. Em doisgêneros (Glossomatix e Pinguiococcus) ocorre uma camada fina de natureza polissacarídica;as células de Phaeomonas e Pinguiochrysis são nuas; e há uma lórica orgânica rica em alguns

96
metais (Zn e Mg) e enxofre em Polypodochrysis. Vesículas periféricas derivadas de membranastubulares são observadas na periferia de células de Glossomatix e Pinguiococcus. Curiosamente,uma mitocôndria circunda o núcleo de Phaeomonas e Pinguiochrysis, um caráter singularem Stramenopiles.
1
23
4
5
6
7
8
9
Figura 2.36 Diversidade de pinguiofíceas. 1. Pinguiochrysis pyriformis. 2,3. Phaeomonas parva; célulanatante (2); célula cocóide (3). 4. Pinguiococcus pyrenoidosus. 5-7. Glossomastix chrysoplasta; célula vegetativa(5); colônia (6); zoósporo (7), com pseudópode paralelo ao único flagelo. 8,9. Polypodochrysis teissieri;célula vegetativa com lórica (8); zoósporo (9), com pseudópode paralelo ao único flagelo. Barra da escala =10 µm (1-5, 7-9) ou = 80 µm (6). Reproduzido de Kawachi et al. (2002).
Não são conhecidos fósseis desta classe, nem eventuais efeitos prejudiciais em sistemasnaturais. A reprodução sexuada é desconhecida no grupo. A reprodução assexuada ocorrepor divisão binária ou pela produção de zoósporos. Estes, curiosamente, não nadam, mesmodispondo de flagelo. Os zoósporos movem-se deslizando sobre um substrato ou por ação deum pseudópode paralelo ao flagelo.
2.2.8.7 CLASSE RAPHIDOPHYCEAE
Esta classe é constituída por algas monadais relativamente grandes (30-80 µm) e nuas,sendo desprovidas de parede celular ou escamas (Figura 2.37). Estigmas não ocorrem nestaclasse e as rafidofíceas formam o único grupo de ocrófitas dotadas de tricocistos, uma desuas características mais marcantes. São conhecidas espécies marinhas (em oceano aberto eem águas estuarinas) e dulciaqüícolas nos oito gêneros descritos.
Células de rafidofíceas podem ser ovóides, piriformes, assimétricas ou achatadas, emdiferentes graus. São dotadas de um flagelo longo (geralmente voltado para frente), commastigonemas tripartidos, e de um flagelo mais curto (geralmente voltado para trás) e liso.

97
As rafidofíceas possuem núcleo grande e conspícuo mesmo durante a mitose. A divisão celulardá-se pela formação de um sulco de clivagem. O retículo endoplasmático periplastidial nãose conecta com o envelope nuclear. Aparatos de Golgi são também grandes e numerosos esua atividade envolve a secreção de mucilagem extracelular eventualmente abundante. Asrafidofíceas são fotoautotróficas e dotadas de plastos numerosos e discóides. Além de clorofilaa, algumas espécies possuem clorofilas c1 e c2 simultaneamente. Há diferenças na composiçãode carotenóides entre espécies marinhas e dulciaqüícolas; as espécies marinhas apresentamfucoxantina e violaxantina em grandes quantidades, ao passo que as espécies de água docesão ricas em diadinoxantina, heteroxantina e vaucheriaxantina. Pirenóides são encontradosapenas em espécies marinhas, e gotículas de óleo constituem-se no principal produto dereserva das rafidofíceas (Graham & Wilcox, 2000).
Fósseis não são conhecidos no grupo. Reprodução sexuada nunca foi observada, masreconhece-se a formação de cistos e esporos supostamente haplóides (Lee, 1999).
Algumas rafidofíceas podem formar florações nocivas à biota marinha. Florações derafidofíceas já foram registradas no Brasil, sendo notável um evento de grandes proporçõesna baía de Sepetiba (RJ) em fevereiro de 1998 (Odebrecht et al., 2002). Algumas espécies,como Heterosigma akashiwo, são produtoras de neurotoxinas sem efeito ao homem, mas quepodem afetar animais marinhos.
Os gêneros marinhos Chattonella, Fibrocapsa, Haramonas, Heterosigma e Olisthodiscusformam a classe juntamente com os gêneros dulciaqüícolas Gonyostomum, Merotricha eVacuolaria.
2.2.8.8 CLASSE PHAEOTHAMNIOPHYCEAE
Esta classe possui apenas representantes dulciaqüícolas e um gênero presente em águasalobra (Stichogloea), sendo tratada brevemente nesta obra. O grupo é reconhecido pelaidentidade gerada por meio de análises moleculares (Andersen et al., 1998), mas não seconhece caráter derivado próprio facilmente identificável.
Figura 2.37 Representação de duasrafidofíceas: (a) Heterosigma akashiwoe (b) Fibrocapsa japonica. C = plasto;M = mucocisto. Reproduzido de Lee(1999).

98
As espécies conhecidas apresentam talo filamentoso, palmelóide ou cocóide (Figura2.38). Pouco se sabe sobre o aparato fotossintético dos representantes desta classe, mas acombinação das xantofilas fucoxantina e heteroxantina é singular na natureza. Fósseis sãodesconhecidos. A reprodução sexuada nunca foi constatada nas feotamniofíceas e a multi-plicação ocorre pela produção de zoósporos (desprovidos de estigma e com inserção lateraldos dois flagelos heterocontes) ou autósporos.
a)
b)
c) d) 10 µm10 µm
10 µm
20 µm
Figura 2.38 Diversidade morfológica de feotamniofíceas. (a) Phaeothamnion confervicola. (b) Chrysocloniumramosum. (c) Phaeobotrys solitaria. (d) Phaeoschizochlamys mucosa. Reproduzido de Reviers (2006).
Os gêneros Chrysapion, Chrysoclonium, Phaeogloea, Phaeoschizochlamys, Phaeothamnion(o gênero mais estudado da classe), Selenophaea, Stichogloea e Tetrapion são os mais comuns,dentre os 15 gêneros existentes na classe.
2.2.8.9 CLASSE CHRYSOMEROPHYCEAE
Esta classe é constituída exclusivamente por algas bentônicas marinhas, cujos talos sãofilamentosos ou palmelóides (Figura 2.39). A organização das crisomerofíceas como umaclasse não é sustentada por nenhum caracter derivado evidente nem há, até o momento,respaldo de análises moleculares. A classe pode ser artificial e talvez no futuro o grupo sejareorganizado sistematicamente.
a)
b)
c)
d)
10 µm
10 µm
5 µm
20 µm
e)
f)
Figura 2.39 Diversidade morfológica de crisomerofíceas. (a,b) Chrysomeris ramosa (b, zoósporos). (c, d)Chrysowaernella hieroglyphica (d, zoósporos). (e, f ) Giraudyopsis stellifera (f, filamento plurisseriado).Reproduzido de Reviers (2006).,

99
Os organismos que constituem esta classe apresentam algumas afinidades com as algasfeofíceas, manifestadas pela inserção lateral dos zoóides e pela ultra-estrutura flagelar.Giraudyoides, um dos gêneros mais estudados do grupo e que apresenta talo heterótrico, foioriginalmente tratado como pertencente à classe Phaeophyceae. Não são conhecidos fósseisde espécies desta classe e a reprodução é realizada por zoósporos, não havendo registro dereprodução sexuada.
Sete gêneros são conhecidos na classe, Antarctosaccion, Chrysoderma, Chrysomeris,Chrysowaernella, Giraudyopsis, Phaeosaccion e Rhamnochrysis, com o total de 12 espécies.
2.2.8.10 CLASSE TRIBOPHYCEAE (= XANTHOPHYCEAE)Esta classe compreende 90 gêneros e cerca de 600 espécies conhecidas, exibindo enorme
diversidade morfológica, cujos talos podem ser cocóides, monadais, amebóides, palmelóides,sifonadas ou filamentosos (Figura 2.40). As tribofíceas ou xantofíceas são algas amarelo-esverdeadas ou verde-claro e são facilmente confundidas com algas clorófitas; porém, astribofíceas nunca apresentam amido nas células, o que permite distingui-las das algas verdesem observação de preparações para microscopia. A maioria das espécies ocorre em água docee no solo úmido, mas algumas tribofíceas ocorrem em água salobra e marinha, como algumasdas espécies de Vaucheria que ocorrem em marismas. Uma característica peculiar do grupoé a formação de cistos endógenos de natureza silicosa, os estomatocistos ou estatósporos.Entretanto, esses cistos especiais não ocorrem em todas as espécies do grupo e, além disso,formas semelhantes de estomatocistos também são encontradas em Chrysophyceae.
a)
b)
c)
d)
10 µm
10 µm
10 µm1
0µ
m
e)
f) g) j)
h)
k)
l)
i)
1 µm
10
µm
Figura 2.40 Diversidade morfológica de tribofíceas. (a) Botrydium granulatum. (b) Characiopsis dubia.(c) Chloramoeba heteromorpha. (d) Rhizochloris congregata. (e) Heterogloea smithiana. (f-i) Vaucheria repens(f-h, formação de um filamento sinzoósporo). (f ) Numerosos núcleos dispõem-se perifericamente na célula,mas cada núcleo tem relação com um par de flagelos ligeiramente desiguais (i). (g) Esporocisto vazio. (h)Sinzoósporo. (j,k) Vaucheria thuretii (j, porção de um talo sifonado, mostrando um oogônio com umaoosfera e cinco espermatocistos, três vazios e dois cheios. Cada espermatocisto contém vários anterozóides.Vaucheria thuretii é monóica, mas outras espécies do gênero são dióicas; k, anterozóide heteroconte). (i)Tribonema vulgare. Reproduzido de Reviers (2006).

100
A parede celular das células de tribofíceas é constituída primariamente por microfibrilasde celulose (em algumas espécies há impregnação por sílica), e em algumas espécies a paredecelular dispõe-se num arranjo tal que ocorrem valvas superpostas. Os plastos são numerosos,discóides e distribuem-se pela periferia das células e em muitas espécies são dotados de umpirenóide (inserido no plasto). O ADN plastidial é arranjado em forma de anel, gerandoum nucleóide do plasto. A clorofila c está presente em baixas concentrações. Os principaiscarotenóides acessórios são o â-caroteno, a diatoxantina, a heteroxantina e a diadinoxantina;a fucoxantina (que é uma xantofila marrom) não existe no grupo, o que faz seus plastosassumirem coloração verde ou verde-amarelada. Gotículas de lipídeos constituem o principalproduto de reserva das tribofíceas, mas supõe-se que a crisolaminarina também ocorra, aexemplo de outras ocrófitas, estando encerrada em vacúolos citoplasmáticos. Nos zoóidesos flagelos são inseridos junto ao ápice da célula (não estão dispostos lateralmente) e umaparato fotorreceptor típico de organismos heterocontes está presente, consistindo em umaintumescência junto ao flagelo curto e liso e uma mancha ocelar disposta dentro em umplasto (Hoek et al., 1995).
Fósseis do grupo são escassos, havendo mais registros relativamente recentes, desde oMioceno, atribuídos a Vaucheria. Entretanto, Xiao et al. (1998) descobriram fósseis na Sibéria(Rússia), em depósitos de um bilhão de anos, interpretados como de um organismo se-melhante a Vaucheria. Knoll (1996) também obteve achado na Rússia de um organismocomparável a Vaucheria, em depósitos de 900 milhões de anos; o fóssil encontrado foi descritocomo Paleovaucheria. Não há comprovação de que tais fósseis sejam mesmo representantesdo grupo.
A reprodução assexuada ocorre pela formação de células não-flageladas, como autós-poros, aplanósporos e cistos, e também pela formação de zoósporos grandes, dotados de váriospares de flagelos heterocontes, conhecidos como sinzoósporos. A reprodução sexuada somentefoi registrada nos gêneros Botrydium (por isogamia ou anisogamia) e Vaucheria (por oogamia),dois gêneros formados por espécies macroscópicas pequenas (mas visíveis a olho nu) comorganização cenocítica de talo.
Os principais gêneros de tribofíceas são Botrydium, Characiopsis, Chloridella, Ophiocytium,Stipitococcus, Tribonema e Vaucheria. São reconhecidos casos de proliferações bentônicas deVaucheria, as quais podem trazer efeitos nocivos à biota.
2.2.8.11 CLASSE PHAEOPHYCEAE
As feofíceas constituem um grupo muito bem definido de ocrófitas que engloba maisde 250 gêneros e 1.500 espécies. São também chamadas de algas pardas ou marrons. Sãoalgas de grande complexidade morfológica: no grupo ocorrem espécies que consistem emformas filamentosas microscópicas até algas gigantes – as maiores algas conhecidas, comdezenas de metros de comprimento e elevado grau de especialização de células, tecidos eórgãos. As feofíceas são as únicas ocrófitas que possuem plasmodesmas. Elas são dotadastambém de estruturas reprodutivas bem diferenciadas. Quase todas as feofíceas ocorrem nomar, apenas cinco gêneros (Bodanella, Heribaudiella, Lithoderma, Pleurocladia e Sphacelaria)com cerca de 30 espécies são de água doce. Não há espécies terrestres. As algas pardas ocorremno mundo inteiro, mas são particularmente mais abundantes em ambientes temperados eem águas frias, onde podem apresentar grande porte e ser amplamente dominantes emambientes rasos, desde o supralitoral até o infralitoral. A zona entremarés de ambientestemperados tende a apresentar enorme biomassa de feofíceas (Figura 2.41), muitas vezescorrespondendo a mais de 90% da biomassa total de algas do local. As feofíceas também

101
ocorrem em ambientes tropicais, mas apresentam porte menor e raramente exercem domi-nância nas comunidades algáceas. Todas as algas pardas são bentônicas, exceto por formasflutuantes de Sargassum (com hábito planctônico), constituindo o chamado mar dos Sargaçosao largo da costa sudeste dos EUA.
Todas as feofíceas são multicelulares. A diversidade morfológica do grupo é muitogrande, envolvendo filamentos microscópicos, filamentos em forma de mechas, formascrostosas, leques, lâminas pequenas, lâminas gigantes, vesículas ocas, estruturas reticuladas,fios longos, fitas sem aerocistos, fitas com aerocistos, arbúsculos, etc. O talo pode serfilamentoso (presente nas formas mais simples), pseudoparenquimatoso ou parenquimatoso,análogo àquele das plantas com flores. Espécies que apresentam tecidos parenquimatososverdadeiros alcançam alto grau de especialização de funções. Algumas dessas espécies apresentamcélulas especializadas dotadas de poros nas paredes celulares que permitem o transporte dosprodutos da fotossíntese, assemelhando-se aos tubos crivados das plantas vasculares.
A parede celular é constituída por uma rede de microfibrilas de celulose, a qual épreenchida e reforçada por alginas, principalmente na forma de alginato de cálcio. As alginassão formadas por dois ácidos urônicos, o ácido D-manurônico e o ácido L-gulurônico (Figura2.42). A porção estrutural da parede celular (celulose + alginas) geralmente é ainda cobertapor uma matriz mucilaginosa amorfa, composta de fucoidana e alginatos mucilaginosos(Hoek et al., 1995). A presença das alginas confere mais flexibilidade ao talo e supõe-setambém que possa aumentar a proteção das células à ação microbiana. As diferentes pro-porções de ácidos urônicos presentes, bem como a ocorrência natural de diferentes sais comcátions (Ca2+, Mg2+ ou Na+), determinam as diferentes propriedades das alginas de espéciesdistintas, com implicações importantes quanto ao uso de tais substâncias com fins comerciais.Os alginatos, produzidos principalmente a partir dos gêneros Macrocystis, Laminaria eAscophyllum, são usados como agentes geleificantes, estabilizantes e emulsificantes em váriasaplicações industriais, como na fabricação de tintas, sorvetes, cerveja, etc. As aplicaçõesindustriais de alginatos movimentam cerca de US$ 250 milhões ao ano, em todo o mundo(McHugh, 2003).
Figura 2.41 Aspectoda zona entremarés emPortaferry, Irlanda doNorte, durante umamaré baixa, onde seconstata a enormebiomassa de algasfeofíceas, sobretudoFucus vesiculosus eAscophyllum nodosum.Foto do autor.

102
OHO
C
OHO
OO
–
OH
O
C
OH
OO
–
O
O OH
C
OHO
O
OH
O
C
OH
OO–
O
O
O–
G G G G
OCO
OH
HO
HO
O
O
O
HOO
HO
OHO
M M M
OCO
OH
HO
HO
O
O
HOO
HO
OHO
OCO
O–
HO
HO
O
M
OOH
O
C
OH
OO
–
O
G
OCO
O–
HO
HO
M
O
Figura 2.42 Estrutura química de alginas, constituídas por dois ácidos urônicos: o ácido gulurônico (G)e o ácido manurônico (M). As proporções e as seqüências de moléculas de ácidos urônicos determinamalginas com propriedades diferentes. Obtido de Genialab (2006).
Dependendo do gênero considerado, pode haver de um a vários plastos por célula, osquais apresentam formas variadas, servindo de critério de classificação taxonômica para várioscomponentes do grupo. Porém, a forma mais comum é discóide. Além da clorofila a, asfeofíceas possuem clorofila c1, clorofila c2, â-caroteno, violaxantina e fucoxantina, sendoeste último pigmento (marrom) presente em grandes concentrações. Há um retículo endo-plasmático periplastidial contínuo com o envelope nuclear. Os pirenóides estão presentesapenas em parte das espécies, as quais correspondem àquelas pertencentes às ordens maisprimitivas.
O ADN plastidial está organizado num nucleóide em forma de anel. O produto dereserva gerado pela atividade fotossintética é a laminarina, um polissacarídeo constituído pormonômeros de glicose unidos por ligações glicosídicas â-1,3, semelhante à crisolaminarina.Além da laminarina, o manitol (um álcool derivado de uma hexose) pode ser acumulado comoproduto de reserva em grandes quantidades. Especula-se que a presença acentuada de manitolpelo citoplasma represente também uma segunda função como agente de crioproteção, jáque essas algas são abundantes em ambientes frios. Substâncias de baixo peso molecular comoo manitol, o glicerol e a sacarose são reconhecidos como crioprotetoras e como estabilizadorasdo balanço osmótico das células (Graham & Wilcox, 2000).
Algas pardas apresentam vesículas especiais espalhadas pelo citoplasma conhecidascomo fisóides. São corpos altamente refringentes e que eventualmente podem ser encontradosdentro do vacúolo celular. Os fisóides contêm grandes quantidades de taninos, consistindo

103
em polímeros de floroglucinol (também conhecidos como polifenólicos) e terpenos, cujafunção está supostamente relacionada à defesa contra herbívoros, pelo fortalecimento daparede celular por sua interação com alginas, à proteção contra danos provocados pelaradiação UVB (Graham & Wilcox, 2000) ou ao aumento da resistência ao ataque de mi-crorganismos.
A maioria das espécies apresenta ciclo de vida diplohaplôntico, com alternância entrereprodução sexuada e assexuada. Neste caso, as fases que se alternam podem ser isomórficasou heteromórficas. Dentre as algas da ordem Laminariales, as diferenças entre o esporófito(gigante) e o gametófico (microscópico) são extremas. Esporos e gametas são produzidospela maioria das espécies. Representantes da ordem Fucales apresentam ciclo de vida comapenas uma fase de vida livre diplóide, e os gametas são produzidos em porções diferenciadasdo talo, especializadas em reprodução.
Algumas espécies apresentam estruturas reprodutivas especiais: os órgãos pluriloculares,que ocorrem em gametófitos e esporófitos, originando células móveis (gametas ou esporos)por mitose; e os órgãos uniloculares, presentes apenas em esporófitos, originando esporosmóveis por meiose. Em parte das algas pardas o ciclo de vida consiste numa alternânciaisomórfica de gerações (Figura 2.43). Todas as algas pardas produzem células reprodutivasmóveis, dotadas de dois flagelos heterocontes típicos, um com mastigonemas tripartidos (oflagelo longo) e outro curto e simples. Os flagelos inserem-se lateralmente no zoósporo ougameta e há um aparato fotorreceptor típico de heterocontes, consistindo em intumescimentojunto ao flagelo posterior liso e de mancha ocelar encerrada dentro do plasto, numa porçãopróxima ao flagelo posterior.
As características da organização do talo, o modo de crescimento, o tipo de reproduçãosexual e o ciclo de vida são caracteres considerados na distribuição de algas pardas em diversasordens, conforme sumarizado na Tabela 2.8.
Zoósporo
Esporófito2N
Esporângioplurilocular
Meiose
GametófitoN
Gametâniopluricelular
FertilizaçãoZigoto
Esporângiounilocular Zoósporo
Gametas
Figura 2.43 Representação esquemática do ciclo de vida de Ectocarpus, que apresenta alternância de gera-ções isomórficas. Modificado de Sze (1998).
Gametângio

104
Ordem Tipo de talo Modo de crescimento
Reprodução sexual Tipo de ciclo de vida
Exemplos de gêneros
Ectocarpales Filamentos unisseriados, ramificados ou não ramificados
Geralmente difuso Isogamia morfológica com anisogamia ou oogamia funcional
Alternância isomórfica de gerações
Bachelotia, Ectocarpus, Ralfsia
Sphacelariales Filamentos multisseriados
ramificados Célula apical Isogamia, anisogamia
ou oogamia Geralmente isomórfico
Sphacelaria
Dictyotales Parenquimatoso Apical/marginal Oogamia Isomórfico Dictyota,
Dictyopteris, Padina, Stypopodium
Scytosiphonales Esporófito filamentoso pequeno; gametófito parenquimatoso grande
Apical no espo-rófito; intercalar no gametófito
Anisogamia Heteromórfico Scytosiphon
Cutleriales Gametófito
parenquimatoso maior que o esporófito
Apical no espo-rófito; tricotálico no gametófito
Anisogamia Heteromórfico Cutleria
Dictyosiphonales Gametófito filamentoso;
esporófito parenquimatoso Difuso no espo-rófito; apical no gametófito
Isogamia ou anisogamia
Heteromórfico Asperococcus, Dictyosiphon
Chordales Filamentos unisseriados
que podem formar discos ou cordas pseudoparen-quimatosos
Tricotálico no esporófito; apical no gametófito
Isogamia com anisogamia funcional
Heteromórfico Chorda, Elachista
Sporochnales Pseudoparenquimatosos;
esporófito maior que gametófito
Tricotálico Oogamia Heteromórfico Sporochnus
Desmarestiales Gametófitos filamentosos
pequenos; esporófito pseudoparenquimatoso maior
Tricotálico Oogamia Heteromórfico Desmarestia
Laminariales Gametófitos filamentosos
pequenos; esporófito parenquimatoso grande
Intercalar no esporófito; apical no gametófito
Oogamia; ovos eventualmente flagelados
Heteromórfico Alaria, Ecklonia, Laminaria, Macrocystis, Nereocystis
Fucales Parenquimatoso Célula apical Predominantemente
oogamia Aparentemente apenas uma geração diplóide simples, meiose gamética
Ascophyllum, Cystoseira, Fucus, Hormoseira, Sargassum
Durvillaeales Parenquimatoso Difuso em por-
ções apicais do talo; sem célula apical solitária
Oogamia Uma geração diplóide; meiose gamética
Durvillaea
Syringodermatales Esporófito parênquima-
toso; gametófito com apenas 2-4 células
Marginal/apical Isogamia Heteromórfico; gametófitos não são de vida livre
Syringoderma
Ascoseirales Esporófito
parenquimatoso Intercalar no esporófito
Isogamia Heteromórfico; gametófitos não
Ascoseira
Tabela 2.8 Sumário das características das principais ordens de Phaeophyceae. Modificado e expandidoa partir de Graham & Wilcox (2000).

105
Algumas ordens apresentadas na Tabela 2.8 são novas (por exemplo, Syringodermatales,Ascoseirales) e parcialmente aceitas pela comunidade científica. Outras (por exemplo,Laminariales, grupo provavelmente polifilético) possivelmente poderão ser divididas embreve, sobretudo em função da aquisição de dados novos de sistemática molecular.
Algumas algas pardas são amplamente utilizadas na alimentação humana, sobretudoem países do Extremo Oriente, onde recebem nomes populares. Laminaria (“kombu”),Undaria (“wakame”) e Hizikia (“hijiki”) são muito apreciadas por povos orientais; Durvillaeae Lessonia são consumidas no Chile. Algumas algas pardas (por exemplo, Laminaria) sãoutilizadas no tratamento de problemas de saúde segundo procedimentos da tradicionalmedicina chinesa.
Algumas algas pardas estão envolvidas com efeitos adversos, sobretudo aqueles con-seqüentes da introdução de espécies, intencional ou não. Undaria pinnatifida foi introduzidana França para fins de cultivo e alastrou-se pelo ambiente natural de vários países europeus.Também na Europa, Sargassum muticum foi introduzida acidentalmente e atualmente vemeliminando populações naturais de Laminaria, com a qual compete diretamente, na Grã-Bratanha, Irlanda, Espanha e França.
Além dos gêneros citados na Tabela 2.8, Colpomenia, Costaria, Cystoseira, Egregia,Hedophyllum, Lessonia, Phaeostroma, Pelagophycus, Pelvetia, Postelsia, Spatoglossum, Streblonemae Zonaria são alguns dos principais gêneros de algas pardas. Sargassum é o maior gênero, comcerca de 400 espécies.
2.2.8.12 CLASSE EUSTIGMATOPHYCEAE
O nome do grupo deriva da ocorrência de uma mancha ocelar alaranjada ou aver-melhada fora do(s) plasto(s), na porção anterior de células flageladas. Trata-se de um estigmaverdadeiro (Figura 2.44), o qual não é encerrado por uma membrana; a ocorrência de umestigma é singular entre as ocrófitas. Outra característica peculiar do grupo entre as ocrófitasé a ausência de clorofila c. As eustigmatofíceas compreendem 8 gêneros e 15 espécies, todasunicelulares cocóides ou coloniais, com distribuição na água doce, no solo úmido e no mar,sendo predominantemente planctônicas.
As eustigmatofíceas são células pequenas (2-32 µm) com parede celular rígida, cujanatureza química ainda não foi elucidada, ou cobertas apenas por uma camada de mucilagem.Acredita-se que as eustigmatofíceas sejam organismos fotoautotróficos obrigatórios, nãohavendo registros de nutrição heterotrófica entre as espécies. Ocorre número pequeno evariável de plastos entre as espécies, sendo mais comum um ou dois plastos por célula. Ostilacóides ocorrem agrupados em três. O retículo endoplasmático periplastidial está presente,mas não se conecta ao envelope nuclear. Em algumas espécies pode ser formado um pirenóidepedunculado; pirenóides jamais são encontrados em zoósporos e somente voltam a ser geradosapós a formação da célula vegetativa. A violaxantina é o principal pigmento acessório nacaptação de luz; o â-caroteno e a vaucheriaxantina são também carotenóides importantesdo grupo, havendo ainda outros pigmentos pouco abundantes; a fucoxantina não ocorreno grupo.
A estrutura química do produto de reserva das eustigmatofíceas não é conhecida, massabe-se que não se trata de amido, pois não há reação com lugol (Graham & Wilcox, 2000).Apenas algumas das espécies produzem células flageladas, as quais apresentam um flageloanterior longo, dotado de mastigonemas tripartidos, e um flagelo posterior muito curto ou

106
ausente, sendo neste caso representado apenas pelo corpo basal (Santos, 1996). A coloração,a forma da célula e a forma dos plastos tornam as eustigmatofíceas muito semelhantes a algasverdes cocóides, sendo necessário o uso de microscopia eletrônica ou análise de pigmentosfotossintetizantes para distingui-las.
E*
MM
NN
P
1 µm
Não são conhecidos fósseis desta classe. A reprodução sexuada também é desconhecidaentre as eustigmatofíceas, e as formas de reprodução assexuada registradas no grupo (alémda divisão binária) envolvem a produção de zoósporos nus (com um ou dois flagelos), deautósporos e a fragmentação de colônias, dependendo da espécie considerada.
Botryochloropsis, Chlorobotrys, Ellipsoidion, Eustigmatos, Monodopsis, Nannochloropsis,Polyedriella e Pseudocharaciopsis são os gêneros que constituem esta classe. Não são conhecidosefeitos nocivos de eustigmatofíceas na natureza. Espécies do gênero Nannochloropsis sãobastante utilizadas em aqüicultura em decorrência da facilidade de cultivo, do tamanhopequeno, da velocidade de crescimento acentuada e do alto teor de ácidos graxos poliinsa-turados (Lourenço et al., 2002b).
2.2.8.13 CLASSE CHRYSOPHYCEAE
As algas crisofíceas constituem um grupo relativamente grande de ocrófitas, com cercade 200 gêneros e de 1.000 espécies. A maioria das espécies é dulciaqüícola, mas há tambémalgumas espécies marinhas, sobretudo formas pequenas, pertencentes ao nanoplânctoncosteiro e oceânico. O grupo é mais comum em águas temperadas e frias. O nome do grupoderiva da palavra grega chrysos, que significa “ouro”, referência à coloração dos plastos dasespécies, que varia do dourado ao marrom. A característica mais peculiar do grupo é a
Figura 2.44 Micrografia eletrônica de umaeustigmatofícea típica, apresentando um es-tigma (E) na porção anterior da célula e forado plasto (P). Notar também o núcleo (N),a mitocôndria (M) e uma região expandidado flagelo (*). Modificado de Santos (1996).

107
formação de uma célula especial com paredes celulares silicosas, num estádio de repousodenominado estomatocisto, semelhante ao que ocorre com xantofíceas. Os estomatocistosapresentam uma abertura através da qual a célula é liberada após o período de repouso. Acombinação do estomatocisto com a composição de pigmentos peculiar do grupo o indivi-dualiza das demais ocrófitas. As crisofíceas são microscópicas, ocorrendo como célulasmonadais ou coloniais; formas filamentosas também existem. As crisofíceas são predominan-temente planctônicas quando ocorrem como células monadais, mas espécies coloniais ben-tônicas são comuns. A crisofícea Ochromonas (Figura 2.45) é um modelo perfeito de organis-mo não somente para o grupo, mas para a própria divisão Ochrophyta.
A mixotrofia é uma feição muito comum no grupo, que conta também com repre-sentantes exclusivamente heterotróficos (por exemplo, Oikomonas). Várias espécies sãocapazes de fagocitar células bacterianas ou mesmo outras algas, como diatomáceas e clorófitascocóides. Representantes dos gêneros Dinobryon e Uroglena são particularmente importantesno consumo de bactérias de lagos, contribuindo para controlar proliferações bacterianas commais eficiência do que protozoários ciliados, rotíferos e crustáceos juntos (Graham & Wilcox,2000). Os plastos contêm clorofilas a, c1 e c2, além das xantofilas fucoxantina e violaxantina,presentes em altas concentrações, mascarando a coloração das clorofilas. Pirenóides sãocomuns no grupo e consistem numa área granular que difere em aparência do estroma. Acrisolaminarina é o principal produto de reserva do grupo, sendo armazenada em vacúolos(principalmente no grande vacúolo posterior da célula). Os flagelos heterocontes inserem-se na porção anterior da célula, formando um ângulo aproximadamente reto entre si (Figura2.45). Escamas flagelares foram descritas para alguns gêneros. Na porção anterior do plasto,bem perto da inserção dos flagelos, forma-se uma mancha ocelar discreta, constituída porgotículas de lipídeos; estas dispõem-se entre o envelope do plasto e a primeira banda detilacóides (Lee, 1999).
MM
FM
LF
B
Mt
FT
E
PN
L
2µ
m
Figura 2.45 A crisofícea Ochromonas danica. B = inserçãobasal; LF = filamento lateral; Mt = haste microfibrilar; TF =filamento terminal; E = estigma; FM = pêlo fibrilar; MM =pêlo microtubular; P = plasto; N = núcleo; L = vesícula con-tendo crisolaminarina. Reproduzido de Lee (1999).

108
São conhecidos cistos fósseis do grupo em depósitos continentais desde o Cretáceo(145 milhões de anos), mas supõe-se que o grupo seja mais antigo. Acredita-se que o grupopossa ter sido dominante em muitos ambientes no passado, pois em certos depósitos aquantidade de cistos silicosos de crisofíceras supera amplamente as frústulas de diatomáceasem abundância.
Na classificação das crisofíceas, merece atenção especial o posicionamento de um pequenogrupo de algas de água doce, as Synurales, formado por quatro gêneros: Chrysodidymus, Synura,Tessellaria e Mallomonas. Este grupo apresenta caráter monofilético e bem individualizadodentro das crisofíceas, assim como caracteres derivados próprios bastante importantes, comoa ausência de clorofila c2, um sistema flagelar de raízes flagelares, corpúsculos basais paralelose escamas silicosas de simetria bilateral (Figura 2.46) (Reviers, 2006). Alguns autores acre-ditam que o grupo deveria ser elevado a uma subclasse das crisofíceas. Nesta obra o grupoé tratado como uma ordem, mas não há dúvidas de que essa questão é alvo de intenso debateda comunidade científica e continua em aberto.
A reprodução sexuada é conhecida para parte das espécies do grupo, mas é relativamenterara e envolve a formação de isogametas. Em Synura, um dos gêneros mais estudados, forma-se um estomatocisto zigótico reconhecível pela ocorrência de quatro plastos e dois núcleos.Estomatocistos também podem ser formados sem envolvimento de gametas, configurandouma forma de reprodução assexuada. A reprodução vegetativa por divisão binária das célulasé predominante no grupo e cada célula filha recebe metade das escamas da célula mãe. Mesmoque o suprimento externo de silício seja pequeno, as células podem se dividir, passando aapresentar ausência de cobertura silicosa (Sandgren et al., 1996).
1 µm
Diversas crisofíceas são capazes de formar florações intensas em lagos, as quais podemser prejudiciais à biota pelo consumo do oxigênio do sistema aquático ou pelo entupimentodas vias respiratórias de animais. Não há conhecimento de toxinas com ação específica sobreorganismos aquáticos. Porém, alguns representantes do grupo são conhecidos por sua capaci-
Figura 2.46 Escamas silicosas de Chrysodidymus synuroideus,membro da ordem Sinurales. Reproduzido de Graham et al.(1993).

109
dade de excretar substâncias orgânicas (como cetonas e aldeídos), as quais alteram o odor e osabor da água, podendo constituir graves problemas quando envolvem águas de abastecimento.
Crisofíceas são mais abundantes em lagos oligotróficos e mesotróficos; a presença dogrupo diminui muito em ambientes eutróficos com alta produtividade fitoplanctônica.Supõe-se que nessas condições as crisofíceas apresentam desvantagens em relação a outrasespécies do fitoplâncton na competição por fósforo, ou pelo consumo grande por cladócerosno verão (Hoek et al., 1995). Além disso, as crisofíceas fotossintetizantes ocorrem em pequenabiomassa em lagos com pH maior que 7,5, supostamente por necessitarem de CO
2 livre
para fotossíntese; em pH alcalino, o carbono está presente predominantemente como bi-carbonato (Hoek et al., 1995).
Os principais gêneros de crisofíceas são: Chromulina, Chrysamoeba, Chrysocapsa,Chrysodidymus, Chrysonephele, Chrysostephanosphaera, Cyclonexis, Dinobryon, Hydrurus,Lagynoin, Mallomonas, Ochromonas, Paraphysomonas, Synura, Uroglena e Uroglenopsis.
2.2.9 DIVISÃO RHODOPHYTA
Esta divisão corresponde às algas vermelhas, cujo número exato de espécies é des-conhecido, pois, embora haja cerca de 10.000 espécies descritas, assume-se que há grandequantidade de sinônimos e o número real deve situar-se entre 4.000 e 6.000 espécies. Amaioria das rodófitas ocorre no mar, especialmente em águas tropicais e temperadas quentes;são reconhecidos cerca de 20 gêneros e 150 espécies que ocorrem em ambientes de águadoce. Formas macroscópicas constituem a grande maioria das espécies, ao passo que espéciesunicelulares são poucas. Há grande diversidade morfológica e estrutural no grupo, queenvolve espécies unicelulares, calcificadas, filamentosas e até mesmo talos com forma delâminas com organização parenquimatosa. A maior parte das formas macroscópicas não atingegrandes portes, geralmente variando de alguns milímetros a poucas dezenas de centímetros.Uma das maiores espécies de algas vermelhas é Schizymenia borealis, encontrada no Alasca(EUA) e no Canadá, cujo talo (de forma laminar) pode atingir dois metros de comprimento.
Os registros fósseis de algas vermelhas são de difícil conservação, exceto formas queapresentam carbonato de cálcio em suas paredes celulares. Há registros fósseis segurosatribuídos a algas vermelhas de 570 milhões de anos encontrados no sul da China, os quaisenvolvem inclusive estruturas reprodutivas bem conservadas (Xiao et al., 1998). Entretanto,acredita-se que o grupo seja bem mais antigo, havendo registros atribuídos a algas vermelhasde 1,2 bilhão de anos encontrados na ilha Somerset, no Ártico canadense. Os vestígios fósseisencontrados foram descritos como Bangiomorpha pubescens (Butterfield, 2000), consistindoem formas filamentosas semelhantes a gêneros modernos, como Bangia.
Formas filamentosas constituem-se no tipo mais comum de organização de talo emalgas vermelhas. Neste caso, o crescimento geralmente ocorre através de uma ou mais célulasapicais. As algas filamentosas com crescimento apical muitas vezes apresentam-se comformato tipicamente cilíndrico, eventualmente com ramificações laterais hierarquizadas. Naextremidade do ramo há uma célula apical conspícua, dotada de divisões nos segmentosdistais, caracterizando um sistema de crescimento uniaxial (Figura 2.47), típico de algasvermelhas menores e de talos mais delicados. Entretanto, em muitas espécies o crescimentoocorre por meio da atividade de várias células apicais. Neste caso, o talo torna-se mais robusto,sendo constituído por vários filamentos, cada um deles apresentando uma célula apical inicial,o que caracteriza crescimento multiaxial (Figura 2.48). Como conseqüência dessa disposição,a maioria das espécies apresenta talos em forma de feixes de filamentos fortemente com-pactados; as células do talo apresentam-se tão justapostas que em um corte transversal podem

110
dar a impressão de constituírem parênquimas verdadeiros, sobretudo porque é difícil distin-guir os filamentos nas ramificações laterais. Esse tipo de organização de talo é comum a muitasalgas rodófitas, sendo chamada de pseudoparenquimatosa. Apesar do alto grau de compactaçãodos filamentos do talo, é possível distinguir células corticais e medulares. Além da disposiçãoperiférica, as células corticais apresentam pigmentação intensa e são pequenas, enquanto ascélulas medulares são pouco pigmentadas (ou mesmo sem coloração) e têm maiores dimen-sões. Embora o crescimento apical seja predominante no grupo, há também espécies cujocrescimento é intercalar, difuso, localizado, periférico, tricotálico ou terminal. Apesar dagrande diversidade morfológica, todas as evidências disponíveis atualmente apontam parao caráter monofilético das algas vermelhas, que seriam, portanto, um grupo natural.
As células de algas rodófitas podem apresentar número variável de núcleos. Nummesmo indivíduo é possível encontrar células multinucleadas em um ramo e células uninu-cleadas em outro. A presença de vários núcleos é interpretada como resultado da divisão donúcleo (sem divisão subseqüente da célula inteira), de fusões celulares ou da formação desinapses secundárias (Reviers, 2006). Um vacúolo grande pode preencher toda a região centralda célula. As mitocôndrias e o aparato de Golgi geralmente encontram-se associados nocitoplasma. Inclusões celulares são comuns em algas vermelhas, apresentando formas deesferas, discos, agulhas, espirais, poliedros, etc. As inclusões podem apresentar capacidadede refletir luz e se estiverem presentes em grande quantidade podem acarretar uma irides-cência característica ao talo da alga.
40 µm
Figura 2.47 Meristema apical da alga vermelhauniaxial Platysiphonia. Reproduzido de Bold &Wynne, (1978).

111
As algas desta divisão não possuem flagelos em quaisquer fases de seus ciclos de vida,fato que difere definitivamente as rodófitas das demais algas eucarióticas. Os corpúsculosbasais dos flagelos de algas de outras divisões são centríolos, porém essas organelas estãoausentes em algas vermelhas. Os centríolos têm função na organização do fuso mitóticodurante a divisão celular, mas no caso das algas vermelhas eles são substituídos por dois anéispolares ou organela associada ao núcleo (OAN). Esta última designação é preferida porquenem sempre as estruturas têm forma de anel, sendo mais comum a forma de um par decilindros ocos (Graham & Wilcox, 2000).
Um ou mais plastos estão presentes na periferia (mais comum) ou no centro da célula.Os plastos de rodófitas têm um envelope constituído por apenas duas membranas, o quesugere origem derivada de uma endossimbiose com cianobactérias. O ADN dos plastos dealgas rodófitas ocorre como numerosos nucleóides, de 1-2 µm de diâmetro, distribuídos portoda a organela. Pirenóides ocorrem em parte das espécies de algas vermelhas, mas sua funçãoé desconhecida. Diferentemente de outras algas eucarióticas, os tilacóides estão separadosuns dos outros e têm estruturas granulares sobre suas superfícies externas, os ficobilissomos(Figura 2.9c, d), que são as principais estruturas de captação de luz das rodófitas. Os pig-mentos fotossintéticos, ausentes em algumas formas parasitas,5 incluem a clorofila a, fico-bilinas (ficoeritrina-b, -r e -c, aloficocianina-c e ficocianina-r e -c) e carotenóides, na formade xantofilas (zeaxantina, neoxantina, luteína, anteraxantina, etc.) e carotenos (princi-palmente â-caroteno). A ficoeritrina-b é o pigmento dominante em muitas rodófitas, o quedetermina a coloração vermelha que dá nome ao grupo. Entretanto, muitas espécies têmoutros pigmentos abundantes, de forma que a coloração final resultante pode variar ampla-mente entre as espécies, que podem apresentar coloração amarelo-esverdeada, esverdeada,
5. Há muitas espécies parasitas (e parcialmente parasitas) dentre as rodófitas, as quais podem seradelfoparasitas ou aloparasitas.
Figura 2.48 Agardhiella, um exemplo de algarodófita multiaxial. Reproduzido de Gabrielson &Hommersand (1982).

112
rosa, vermelho-escuro e até marrom. Em decorrência da composição de pigmentos quepossuem, muitas algas vermelhas são capazes de viver sob baixa intensidade de luz, já tendosido registrada a ocorrência de indivíduos de uma espécie não identificada e coralinácea emprofundidade de 268 m, numa montanha submarina na região das Bahamas (Littler et al.,1985). Este é o registro mais profundo de ocorrência de um organismo multicelular fotos-sintetizante já realizado, que recebe apenas 0,0005% da radiação luminosa incidente nasuperfície (= 0,015 a 0,025 µE.m–2.s–1 da luz solar em seu zênite máximo). Durante algumasdécadas vários autores relataram a presença de clorofila d como uma feição característica dogrupo, ao passo que outros consideravam a presença de clorofila d como mero artefatoresultante do isolamento da clorofila a. Essa matéria vem recebendo novos elementos recen-temente, e evidências atuais sugerem que a clorofila d é, na realidade, produzida por cia-nobactérias epifíticas associadas a algas vermelhas (Murakami et al., 2004).
A INTRIGANTE PRESENÇA DE CLOROFILA d EM ALGAS VERMELHAS
A clorofila d foi descoberta na alga vermelha Erythrophyllum delesserioides (eposteriormente em outras espécies) há mais de seis décadas (Manning & Strain, 1943).Trata-se de uma substância singular, a qual apresenta picos de absorção de luz in vivona região do infravermelho próximo, entre 700-730 nm, com pico de absorção em720 nm; apenas a bacterioclorofila e, outro pigmento fotossintético, tem comporta-mento espectral comparável à clorofila d, apresentando absorção máxima de luz nafaixa entre 715 e 725 nm. Apesar das características espectrais peculiares da clorofilad, seu papel na fotossíntese de rodófitas sempre foi motivo de controvérsia. A clorofilad foi encontrada apenas em algumas espécies de algas vermelhas e, mesmo assim, váriasvezes a substância não era detectada regularmente: em muitas situações, o pigmentonão era encontrado em amostras da mesma espécie oriundas do mesmo sítio. Mesmoquando a clorofila d era medida em algas vermelhas, os valores nunca atingiamconcentrações altas, dando margem para questionamentos sobre a participação efetivadesse pigmento na fotossíntese praticada por rodófitas. Alguns autores especulavamque a clorofila d poderia ser meramente um produto de degradação da clorofila a,dada a grande semelhança estrutural das duas moléculas, sendo, assim, um artefatoanalítico gerado durante o processo de isolamento da clorofila a. A clorofila d é idênticaa um dos produtos de degradação da clorofila a por oxidação com permanganato(Murakami et al., 2004). Outro aspecto importante é o fato de que a clorofila d nuncafoi detectada em algas vermelhas de água doce.
As interpretações sobre a presença de clorofila d em algas vermelhas ganharamum novo elemento em 1996, quando uma descoberta começou a mudar o cenário.Miyashita et al. (1996) relataram a descoberta de uma cianobactéria, entãodesconhecida, associada a ascídias de recifes de coral da República de Palau, OceanoPacífico, no oeste equatorial. O grupo tentava, na realidade, isolar a proclorofíceaProchloron didemni, espécie comumente encontrada como simbionte de ascídias deambientes recifais e primeiro organismo procarionte possuidor de clorofila bdescoberto no mundo (Lewin & Withers, 1975). Surpreendentemente, o grupodescobriu um isolado com concentrações de clorofila d superiores a 90% do total declorofila (o restante era constituído de clorofila a), valores jamais encontrados emalgas vermelhas. A cepa isolada (MBIC-11017) continuou a ser pesquisada (Miyashitaet al., 1997), mas somente mais tarde a espécie foi formalmente descrita comoAcaryochloris marina (Miyashita et al., 2003). A distância filogenética entre a

113
cianobactéria e as algas vermelhas constituiu-se em mais um elemento intrigante paraa ocorrência do pigmento na natureza. Estudos morfológicos, químicos e molecularesrecentes convergem para a interpretação de que a cianobactéria marinha A. marina, deampla distribuição geográfica no mundo, é na realidade responsável pela detecção declorofila d em algas vermelhas (Murakami et al., 2004; Larkum & Kühl, 2005). Váriospesquisadores identificaram a presença de A. marina como epífita de diversas espéciesde rodófitas, formando pequenas manchas sobre os talos. Esse caráter de distribuiçãodescontínua sobre o substrato (a macroalga) contribui também para a ocorrência deelevada variabilidade nas medidas de clorofila d feitas em macroalgas. Assim, tem-seformado consenso de que não há algas vermelhas produtoras de clorofila d e que suaocorrência seria meramente derivada da atividade de cianobactérias com hábito epífito.
As mudanças de concepções sobre a produção de clorofila d na natureza parecemainda estar em curso. Agregando mais informações sobre o assunto, Miller et al. (2005)descobriram cianobactérias planctônicas semelhantes a A. marina em Salton Sea, lagoahipersalina (salinidade de 41-45 ups) e eutrófica da Califórnia (EUA). Trata-se doprimeiro registro de um organismo produtor de clorofila d fora do ambiente marinho,além de envolver um componente do plâncton. Provavelmente muitas informaçõesnovas nesse campo serão levantadas em breve.
O produto de reserva é um polissacarídero derivado de glicose formado por ligaçõesglicosídicas do tipo á-1,4, o amido das florídeas ou rodamilo, que ocorre como grânulosno citoplasma, os quais assumem formato aproximadamente esférico ou de escamas arredon-dadas. As cadeias lineares de amido das florídeas ramificam-se lateralmente no sexto carbono(ligações glicosídicas do tipo á-1,6), assim como acontece no glicogênio e na amilopectina,que é uma das formas de amido. Ao ser exposto a lugol, o amido das florídeas torna-secastanho-avermelhado. O produto de reserva das algas vermelhas é formado junto à mem-brana plasmática, mesmo quando um pirenóide está presente.
As células de rodófitas sempre apresentam paredes celulares, as quais são constituídasfundamentalmente por celulose. Entretanto, outros polímeros podem ocorrer associados àcelulose em algumas espécies, como mananas (polímeros de manose) e xilanas (polímerosde xilose). Há também polissacarídeos sulfatados derivados de galactose, as agaranas e ascarragenanas (Figura 2.49). A celulose constitui arranjo fibrilar na parede celular e outrassubstâncias depositam-se sobre a celulose, formando freqüentemente uma porção mucila-ginosa da parede celular. Agaranas e carragenanas diferem apenas pelo grau de sulfatação(carragenanas têm mais átomos de enxofre em sua estrutura); ambas têm grande importânciaeconômica, com vastíssimas aplicações, como, por exemplo, aditivos de laticínios, gelatinas,tintas, meios de cultura sólidos, etc. As mucilagens e os polissacarídeos sulfatados caracte-rísticos são produzidos por meio de atividade intensa do tipicamente grande e complexoaparato de Golgi de rodófitas. Algumas espécies apresentam parede celular impregnada porcarbonato de cálcio, exibindo aspecto pétreo. As espécies dotadas de paredes celulares calci-ficadas são conhecidas como algas calcárias, amplamente distribuídas no mar, sendo espe-cialmente importantes na construção de recifes biológicos. Plasmodesmas não ocorrem entrecélulas de algas vermelhas, mas na maioria das espécies as células vizinhas mantêm conexãoentre si através da abertura de pequenos trechos da parede celular, designada sinapse ouligação citoplasmática. Durante a mitose, a parede transversal formada entre as clelulas filhas

114
fecha-se como um diafragma, mas não por completo. O espaço onde ocorre a sinapse éparcialmente ocupado por uma estrutura complexa conhecida como rolha sináptica, com-posta, por sua vez, por proteínas e polissacarídeos de natureza ácida. A rolha sináptica podeser recoberta de cada lado por uma ou várias camadas contendo polissacarídeos ou mesmopela própria membrana plasmática (Reviers, 2006). Além das sinapses formadas durante adivisão celular, designadas sinapses primárias (Figura 2.50), há também um segundo tipode sinapse, designada sinapse secundária, que consiste numa conexão entre células defilamentos vizinhos. As sinapses são características com elevada importância para a taxonomiadas algas vermelhas.
CH OH2
HOO
R = H (Kappa)R = SO (Iota)3
OH
O
O
O
O
HOCH2
a)
CH OH2
O SO3
O
OH
O
O
CH2
O
b)
O
O
OR
� - 1 - 3 � - 1 - 4
CH OH2
HOO
OSO3
O
O
CH OSO2 3c)
O
O
OSO3
� - 1 - 3 � - 1 - 4
OH
Figura 2.49 Representações moleculares de biopolímeros presentes em algas vermelhas. (a) Agar.(b) Carragenana kappa ou iota, dependendo do radical (R). (c). Carragenana lambda. As setas indicam arepetição da estrutura da molécula básica para formar a macromolécula. Reproduzido de Oliveira et al.(1992).
Nas algas vermelhas ocorre reprodução sexuada e assexuada, as quais são manifestadaspor meio de reprodução vegetativa, espórica e gamética. Para muitas espécies multicelulares,a reprodução vegetativa pode ocorrer pela fragmentação do talo, cujas porções menoresdesprendidas funcionam como propágulos. Por outro lado, para as poucas espécies unice-lulares, a divisão binária consiste na forma predominante de reprodução. A forma maiscomum de reprodução assexuada no grupo baseia-se na formação e liberação de monósporos.A produção de monósporos é mais comum nos grupos mais primitivos de algas vermelhas,
α
α β
β

115
sendo mais raro ou mesmo ausente em grupos mais derivados, que podem praticar reproduçãosexuada quase com exclusividade. A reprodução sexuada envolve alternância de gerações,embora a reprodução gamética não seja conhecida para todos os gêneros (presume-se queela exista). O ciclo de vida trifásico caracteriza a maioria das espécies de algas vermelhas e éexclusivo dessa divisão. A reprodução gamética do grupo ocorre através de oogamia, seja ociclo de vida trifásico ou bifásico. O gameta feminino é denominado carpogônio e apresentauma porção prolongada, a tricogine, relacionada à captação do gameta masculino. Este édenominado espermácio, que é um aplanogameta, ou seja, um gameta desprovido de flagelos.
Membrana plasmática
Parede
a) b)
c) d)
R.E.
Sinapse
Figura 2.50 Representação da formação de sinapses em algas vermelhas. (a) A parede transversal começaa ser formada entre as células filhas, a partir da acumulação de substâncias oriundas do citoplasma. (b) Osepto é completo, exceto pela permanência de um orifício no centro. (c) O retículo endoplasmáticoposiciona-se através do orifício na parede celular e condensa-se no local. (d) A sinapse é formada, consistindoem uma rolha com a membrana plasmática contínua de célula para célula. Modificado a partir de Lee(1999).
Gametas masculinos aflagelados constituem uma feição bastante peculiar das algasvermelhas. Características do ciclo de vida de algas vermelhas, são importantes para a taxo-nomia do grupo, contribuindo fortemente para a determinação de classes, ordens e famílias.Tradicionalmente, a classificação das algas vermelhas sempre teve como seus principaissustentáculos as características morfológicas das estruturas reprodutivas de indivíduos femi-ninos e os eventos que ocorrem após a fecundação. Atualmente, outras características impor-tantes são também levadas em conta (por exemplo, seqüências gênicas, ultra-estrutura dassinapses, etc.), mas aspectos reprodutivos continuam tendo importância acentuada. Os ciclosde vida das duas principais subclasses da divisão, Florideophycidae e Bangiophycidae, sãobastante distintos e serão exemplificados por dois gêneros: Polysiphonia (Florideophycidae)e Porphyra (Bangiophycidae). As descrições dos ciclos de vida de Polysiphonia e Porphyra sãobaseadas em Hoek et al. (1995).

116
Espermácios (n)
Espermatângios
Gametófitofeminino
(n)
Ramocarpogonial
Célulaauxiliar (n)
Espermácio (n)
Tricogine
Oosfera(porção basal docarpogônio com
núcleo (n)
Núcleozigótico (2n)
Célulaaxial
Célulasuporte
Fertilização
Gametófitomasculino
(n)
Núcleozigótico (2n)
Germinaçãodos tetrásporos
Tetrásporos(n)
Meiose
Tetrasporófito(2n)
Germinaçãodo carpósporo
Carpósporos (2n)
Pericarpo(n)
Carposporófito(dentro do pericarpo)
(2n)
Carposporângios
Tetrasporângios
Figura 2.51 Representação diagramática do ciclo de vida de Polysiphonia, uma alga vermelha da classeFlorideophycidae. A descrição do ciclo encontra-se no texto. Adaptado de Raven et al. (1996).
O ciclo de vida de espécies do gênero Polysiphonia é apresentado na Figura 2.51.Algumas células axiais podem se projetar além das células pericentrais e dar origem a filamen-tos laterais especiais chamados de tricoblastos. Indivíduos masculinos formam esperma-tângios nos tricoblastos, projetando-se inteiramente para fora do filamento. No caso dosindivíduos femininos, o tricoblasto forma um ramo especial de sete células, o ramo car-pogonial, em que a segunda célula possui uma porção basal, que contém o núcleo e as demais

117
organelas, e uma porção alongada que se projeta para fora do filamento: é o carpogônio.Após a fecundação do carpogônio, o zigoto é transferido para a célula auxiliar, que dá inícioà formação do gonimoblasto. Durante o desenvolvimento subseqüente do gonimocarpo,células do ramo carpogonial e células dos filamentos estéreis fundem-se e geram uma grandecélula de fusão. O gonimocarpo maduro é envolto num pericarpo em forma de urna; ogonimocarpo e o pericarpo juntos formam o chamado cistocarpo.
O gonimocarpo constitui-se numa fase do ciclo de vida das Florideophycidae, a fasecarposporofítica, sendo protegido e nutrido pelo indivíduo materno, numa forma de para-sitismo.
A fase carposporofítica tem número diplóide de cromossomos e produz carpósporos,que são esporos formados por mitose. Ao atingirem a maturidade, os carpósporos são libe-rados para o meio através de um poro na extremidade do cistocarpo, e ao germinarem originama terceira fase do ciclo de vida, o esporófito. Este é morfologicamente idêntico ao gametófito, nocaso de Polysiphonia (e na maioria das Florideophycidae), constituindo ciclo de vida isomórfico;em algumas algas vermelhas as duas fases de vida livre podem ser bastante diferentes entre si,determinando ciclos de vida heteromórficos. Os esporófitos apresentam esporângios em ramosvegetativos. Por meio de meiose nos esporângios, formam-se quatro esporos haplóides. Poressa razão, os esporófitos das algas vermelhas são chamados de tetrasporófitos, os esporângiossão chamados de tetrasporângios e os esporos haplóides são designados tetrásporos. Os tetrás-poros são lançados no meio e dão origem a gametófitos haplóides, fechando o ciclo de vidatrifásico das Florideophycidae: fase gametofítica (n), fase carposporofítica (2n) e fase tetras-porofítica (2n).
O ciclo de vida de Porphyra (Bangiophycidae) é heteromórfico, com formação de duasfases alternantes extremamente distintas entre si (Figura 2.52). As diferenças extremas namorfologia das duas fases do ciclo de vida fizeram com que elas fossem tratadas como gênerosdistintos por muito tempo. Somente após os estudos detalhados do desenvolvimento dePorphyra, realizados pela britânica Kathleen M. Drew, pôde-se elucidar o ciclo de vida dasespécies do gênero (Drew, 1949). Tais descobertas abriram caminho para o desenvolvimentointenso do cultivo de Porphyra, uma alga comestível muito apreciada, principalmente empaíses do Extremo Oriente, cujo cultivo e comercialização movimentam atualmente recursosda ordem de 2,5 bilhões de dólares ao ano (McHugh, 2003). A fase “conchocelis” é oesporófito, com dimensões microscópicas e organização filamentosa; a fase é diplóide (2n)e ocorre em camadas internas superficiais de conchas de moluscos e mesmo carapaças decracas, predominantemente nos meses do verão e outono. A fase “conchocelis” alterna-secom a fase foliácea, que é o gametófito das espécies de Porphyra; o gametófito é macroscópico,desenvolve-se no médio e supralitoral de costões rochosos, ocorrendo no inverno e naprimavera. Os gametófitos são geralmente hermafroditas, mas também há espécies queapresentam gametófitos com sexos separados. Nos indivíduos masculinos (ou em certasporções dos talos de indivíduos hermafroditas), células vegetativas transformam-se emespermatângios. Após sucessivas divisões mitóticas, os espermatângios produzem muitas (64ou 128) espermácias, as quais são transportadas passivamente no plâncton. Os gametasfemininos, os carpogônios, são quase indistinguíveis das células vizinhas, exceto pelas peque-nas projeções que emergem na superfície do talo. As espermácias atingem o talo aleato-riamente e algumas se encontram com as projeções dos carpogônios. A espermácia entãoinjeta seu núcleo na projeção do carpogônio, ocorrendo cariogamia e formação de um úniconúcleo diplóide. Após a fertilização, o carpogônio (que se torna diplóide) sofre várias mitosessucessivas e forma células especiais, os carpósporos, que são liberados através da desintegração

118
da parede celular do carpogônio. Geralmente, nesta fase ocorre a desintegração de todo otalo da alga feminina. Os carpósporos atingem conchas calcárias, onde se fixam e originamfilamentos curtos, dotados de poucas células, a fase conchocelis, o esporófito diplóide dePorphyra. Quando atingem a maturidade, algumas células do esporófito diferenciam-se emesporângios, conhecidos como conchosporângios. Estes entram em mitose, originam esporosdiplóides conhecidos como conchósporos6 que são liberados na água, sofrem um processode meiose e formam quatro células haplóides. Ao atingirem um substrato adequado, essascélulas haplóides dão origem à fase foliácea de Porphyra, fechando seu ciclo de vida.
Margens da lâmina (corte)
Carpósporos
Concha de ostra
Primavera
Verão
Outono
Inverno
Conchocelis
Conchósporos
Lâmina
Fio de rede
Conchocelisfértil
Ramosconchosporangiais
Figura 2.52 Representação diagramática do ciclo de vida de Porphyra, uma alga vermelha da classeBangiophycidae. A descrição do ciclo encontra-se no texto. Nesta representação, a fixação dos gametófitosocorre em redes (em vez de rochas), pois o esquema refere-se ao cultivo comercial da alga. Modificado deMumford Jr. & Miura (1988).
Numerosos autores reconhecem a existência de apenas uma classe de algas vermelhas:Rhodophyceae. Três subclasses são reconhecidas atualmente: Cyanidiophycidae,Bangiophycidae e Florideophycidae. A subclasse Bangiophycidae é parafilética e deverá sersubdividida em breve. A própria subclasse Cyanidiophycidae é constituída por três gêneros(Cyanidium, Cyanidioschyzon e Galdeiria, todos de água doce) que antes eram classificadosdentro da subclasse Bangiophycidae. Como ainda não há proposta formal de subdivisão dasBangiophycidae, por conveniência nesta obra elas são apresentadas como um grupo único. Asordens de Rhodophyceae reconhecidas por Reviers (2006) são apresentadas na Tabela 2.9. Emfunção do grande número de táxons, as características diagnósticas de cada um deles não sãoapresentadas.
6. Antes imaginava-se que a meiose ocorria na formação dos conchósporos, mas atualmente sabe-se que aredução da ploidia ocorre posteriormente, após a liberação dos carpósporos (Graham & Wilcox, 2000).

119
É extremamente difícil listar os principais gêneros da divisão Rhodophyta, dada aimensa diversidade, importância ecológica e mesmo pelo aproveitamento econômico dealguns de seus representantes. Entretanto, alguns gêneros não podem deixar de ser citados,como Cyanidium, Glaucosphaera e Porphyridium (unicelulares), Bangia, Batrachospermum,Boldia, Bostrychia, Botryocladia, Caloglossa, Ceramium, Chondrus, Corallina, Dasya, Palmaria,Eucheuma, Gelidium, Gracilaria, Gymnogongrus, Hildenbrandia, Halosaccion, Hypnea,Kappaphycus, Laurencia, Lithothamnion, Lithotrix, Mastocarpus, Mazzaella, Polysiphonia,Porphyra, Pterocladia, Pterocladiella, Rhodymenia e Spirydia (multicelulares).
Tabela 2.9 Sumário da classificação das algas vermelhas em grandes grupos, com indicação do habitat deocorrência das espécies: A = subaéreo; D = dulciaqüícola; M = marinho; S = água salobra; T = termófilo.Segundo Reviers (2006).
Divisão Rhodophyta, classe Rhodophyceae
Subclasse Cyanidiophyceae Ordem Cyanidiales (D, T)
Subclasse Bangiophyceae Ordens Erythropeltiales (M, D),Rhodochaetales (M), Porphyridiales (M, D,A), Compsopogonales (D, S), Bangiales (M,D)
Subclasse Florideophyceae Ordens Hildenbrandiales (M, D), Ahnfeltiales(M), Gracilariales (M), Rhodymeniales (M),Ceramiales (M, D, S), Plocamiales (M),Halymeniales (M), Gelidiales (M),Bonnemaisoniales (M), Gigartinales (M),Corallinales (M), Rhodogorgonales (M),Balliales (M), Thoreales (D), Balbianales (D),Batrachospermales (D), Acrochaetiales (M, D),Palmariales (M), Nemaliales (M, D),Colanematales (M)
2.2.10 DIVISÃO CHLOROPHYTA
Compreende as algas verdes ou clorófitas e envolve formas unicelulares (maioria dasespécies) e espécies dotadas de talos multicelulares. Há cerca de 17.000 espécies, com repre-sentantes em ambientes dulciaqüícolas, salobros e marinhos; além desses habitats básicoshá outros especiais, como neve, troncos de árvores, folhas de plantas terrestres, pêlos deanimais, salinas, desertos secos, cinzas vulcânicas, etc. Juntamente com as cianobactérias, asclorófitas levam ao extremo os habitats possíveis para a existência de algas. Apesar dessa grandevariabilidade de habitats, nota-se que cerca de 90% do total de espécies (sobretudo as formasmicroscópicas) ocorrem em água doce. Há também enorme variabilidade morfológica nadivisão, que compreende formas unicelulares cocóides, unicelulares monadais, colôniaspequenas, colônias de tamanho intermediário, colônias grandes e filamentos, além de formasmacroscópicas que incluem espécies que podem atingir alguns metros de comprimento (porexemplo, algumas espécies de Codium) e mais de 1,0 m2 de área (por exemplo, formasflutuantes de Ulva). Dentre as espécies macroscópicas de clorófitas, há elevado número de

120
formas cenocíticas, como alguns representantes da classe Ulvophyceae. A complexidademorfológica apresentada por carofíceas multicelulares dos gêneros Chara e Nitella atingemaior grau na divisão Chlorophyta, com formação de estruturas reprodutivas bemdiferenciadas e protegidas.
As clorófitas constituem o grupo mais semelhante às plantas com flores. Vários estudosmoleculares (além de estudos químicos, morfológicos e citológicos) indicam que os doistáxons, Chlorophyta e Embryophyta, podem ser reunidos num mesmo táxon monofilético,as Viridiplantae. As algas verdes e as embriófitas têm em comum: plasto envolto por duasmembranas, dotadas de clorofilas a e b; tilacóides empilhados de dois a seis nos grana;ocorrência de neoxantina, xantofila que não está presente em nenhum outro grupo de alga,exceto nas cloraracniófitas; amido como produto de reserva, produzido dentro dos plastos;nas formas flageladas, presença de uma estrutura estrelada na zona de transição dos flagelos,conhecida como “peça em H” (Reviers, 2006). Porém, as clorófitas formam um grupoheterogêneo, havendo, efetivamente, táxons polifiléticos dentro da divisão Chlorophyta. Umgrupo de flagelados (as prasinofíceas) divergiu primitivamente da linhagem principal dogrupo e outra linhagem (as ulvofíceas) é formada apenas por organismos multicelulares. Asprasinofíceas e as ulvofíceas são predominantemente marinhas. As trebouxiofíceas e asclorofíceas são predominantemente dulciaqüícolas ou terrestres. Uma linhagem compostaprincipalmente de formas multicelulares, freqüentemente calcificadas, constitui as carofíceas.Estas deram origem às plantas terrestres (as embriófitas). As principais características dascinco classes de Chlorophyta são apresentadas na Tabela 2.10.
As clorófitas são dotadas de pelo menos um plasto fotossintetizante. Há, entretanto,grande variação no número de plastos por célula dentre as espécies, que pode atingir algumasdezenas. Duas membranas circundam os plastos, mas não há envelope adicional associadoao retículo endoplasmático. Essa característica assemelha as clorófitas às rodófitas, glaucófitas,briófitas e traqueófitas. As formas dos plastos também variam amplamente entre as espécies,conforme pode ser observado na Figura 2.53. O número e a forma dos plastos, entretanto,são características relativamente regulares dentro dos gêneros e têm grande importânciataxonômica. Estigmas podem estar presentes em clorófitas e, quando isso ocorre, estão semprelocalizados dentro dos plastos. Além da clorofila a, a clorofila b, o ß-caroteno e váriasxantofilas, como a luteína, a violaxantina, a zeaxantina, a anteraxantina, a neoxantina, aastaxantina e a cantaxantina, dente outras, ocorrem em plastos.
Apesar da existência de muitos carotenóides no grupo, os plastos de clorófitas sãoverdes, pois na grande maioria das espécies as concentrações dos carotenóides não suplantamas das clorofilas. Há várias décadas o aparato fotossintético de clorófitas vem sendo estudadoem função de sua grande semelhança em relação ao de plantas com flores, sendo o maisestudado dentre todas as algas. De forma exclusiva dentre os organismos conhecidos comoalgas, o produto de reserva, amido, é produzido dentro dos plastos das algas verdes, comonas plantas com flores. Pirenóides ocorrem em parte das espécies de clorófitas; quandopresentes, eles ocorrem dentro dos cloroplastos, são muitas vezes penetrados por tilacóidese cada um deles é circundado por uma espécie de anel de grãos de amido. As moléculascirculares de ADN plastidial ocorrem concentradas em numerosas manchas pequenas (quelembram gotículas) de 1-2 µm de diâmetro; o ADN plastidial de clorófitas nunca ocorreorganizado como um nucleóide único em forma de anel (Hoek et al., 1995). Há formasdesprovidas de aparato fotossintético, como Polytoma e Prototheca, sendo heterótrofosobrigatórios, cuja inclusão dentre as clorófitas decorre de suas características ultra-estruturaise moleculares.

121
Tab
ela
2.10
Car
acte
ríst
icas
das
cin
co p
rinc
ipai
s cl
asse
s de
alg
as v
erde
s. M
odif
icad
o e
expa
ndid
o a
part
ir d
e G
raha
m &
Wilc
ox (
2000
).
Cla
sse
Apa
rato
fla
gela
r/
cito
esqu
elet
o
Enz
imas
fo
torr
espi
-ra
tóri
as
Mit
ose
Cit
ocin
ese
Hab
itat
pr
imár
io
Cic
lo d
e vi
da
Exe
mpl
os d
e gê
nero
s
Pras
inop
hyce
ae
Raí
zes
cruc
iada
s,
rizop
last
os,
algu
ns c
om
MLS
, esc
amas
ce
lula
res
e fla
gela
res
com
uns
Var
iáve
l V
ariá
vel
Sulc
ada
Mar
inho
M
eios
e zi
gótic
a N
ephr
osel
mis,
Pt
eros
perm
a,
Tet
rase
lmis
Ulv
ophy
ceae
R
aíze
s X
-2-X
-2
cruc
iada
s,
orie
ntaç
ão
CC
W, +
/-
esca
mas
fla
gela
res
e ce
lula
res,
riz
opla
sto
pres
ente
Glic
olat
o de
sidr
ogen
ase
Fech
ada,
fuso
pe
rsist
ente
Su
lcad
a M
arin
ho
Mei
ose
zigó
tica
ou
alte
rnân
cia
de
gera
ções
ou
mei
ose
gam
étic
a
Ana
dyom
ene,
D
erbe
sia,
Rhi
zocl
oniu
m,
Siph
onoc
ladu
s, U
dote
a, U
lva,
V
alon
ia
Tre
boux
ioph
ycea
e R
aíze
s X
-2-X
-2
cruc
iada
s,
orie
ntaç
ão
CC
W, s
em
esca
mas
, riz
opla
sto
pres
ente
Glic
olat
o de
sidr
ogen
ase
Sem
i-fe
chad
a, fu
so
não-
pers
isten
te
Sulc
ada
Dul
ciaq
üíco
la
ou te
rres
tre
Mei
ose
zigó
tica
Chl
orel
la,
Gol
enki
nia,
T
rebo
uxia
Chl
orop
hyce
ae
Raí
zes
X-2
-X-2
cr
ucia
das,
or
ient
ação
CW
ou
DO
, esc
amas
ra
ram
ente
oc
orre
m,
rizop
last
os
Glic
olat
o de
sidr
ogen
ase
Fech
ada,
fuso
nã
o-pe
rsist
ente
Sulc
ada,
fico
plas
to,
algu
ns c
om p
laca
ce
lula
r e
plas
mod
esm
ata
Dul
ciaq
üíco
la
Mei
ose
zigó
tica
Ank
istro
desm
us,
Chl
amyd
omon
as,
Coe
lastr
um, G
oniu
m,
Hyd
rodi
ctyo
n,
Scen
edes
mus
, T
etra
edro
n, V
olvo
x
Cha
roph
ycea
e R
aíze
s as
simét
rica
s, M
LS, e
scam
as
flage
lare
s e
celu
lare
s ge
ralm
ente
pr
esen
tes,
riz
opla
sto
raro
Glic
olat
o ox
idas
e e
cata
lase
em
pe
roxi
ssom
as
Abe
rta,
fuso
pe
rsist
ente
Su
lcad
a, a
lgun
s co
m fr
agm
opla
sto,
pl
aca
celu
lar
e pl
asm
odes
mat
a
Dul
ciaq
üíco
la
Mei
ose
zigó
tica
Cha
ra, C
hlor
okyb
us,
Chl
oste
rium
, Nit
ella
, O
nyco
nem
a ,
Spir
ogyr
a, Z
ygne
ma

122
a)
b)
c)
d)
e)
f)
g)
Figura 2.53 Exemplos de plastos encontrados em algas verdes: (a) parietal em forma de taça; (b) parietalem forma de anel; (c) estrelado; (d) axial em forma de placa; (e) em forma de fita; (f ) reticulado; (g) discóide(numerosas unidades). Reproduzido de Graham & Wilcox (2000).
Numerosas clorófitas são capazes de suplementar a aquisição de carbono por meio dafotossíntese, pela utilização de matéria orgânica dissolvida na forma de açúcares, ácidosaminados e outras moléculas orgânicas pequenas, caracterizando um quadro de osmotrofiae mixotrofia (Graham & Wilcox, 2000).
As paredes celulares são constituídas por estrutura fibrilar de celulose (como nas plantasterrestres) inserida numa matriz não fibrilar (geralmente hemicelulose). Essa estrutura estáausente em formas flageladas. Certas espécies apresentam diferente composição química daparede celular, sendo constituídas por polímeros de xilose (por exemplo, Bryopsis e Caulerpa)ou manose (por exemplo, Acetabularia). Algumas são calcificadas, apresentando revestimentode CaCO
3 (por exemplo, Halimeda), e outras são nuas (por exemplo, Dunaliella). Clorófitas
dotadas de paredes celulares recobertas com CaCO3 são comuns em ambientes tropicais,
onde podem participar da constituição de recifes biológicos.Algumas espécies são células vegetativas flageladas, outras apresentam formas flageladas
apenas nas células reprodutivas; a maioria das espécies apresenta dois flagelos isocontes emalguma fase do ciclo de vida, embora haja clorófitas com quatro ou mais flagelos. Os flagelossão idênticos ou muito semelhantes em estrutura, mas podem diferir em comprimento.Características do aparato flagelar constituem marcadores filogenéticos importantes para

123
classificar grupos que compõem as Chlorophyta; é particularmente importante a disposiçãodos corpúsculos basais. Células flageladas de clorófitas das classes Ulvophyceae,Trebouxiophyceae e Chlorophyceae apresentam as raízes do sistema flagelar aproximadamentecruciadas, em que um número variável (X) de radículas de microtúbulos alterna-se comradículas compostas de dois microtúbulos para formar um arranjo conhecido como “X-2-X-2” (Graham & Wilcox, 2000). Dentre as algas com sistemas flagelares de raízes cruciadas,ocorrem três variações fundamentais na orientação das bases dos flagelos quando as célulassão observadas em vista apical: horário, no qual os corpos basais são deslocados em sentidohorário a partir de uma linha imaginária paralela entre eles (Figura 2.54a); anti-horário, noqual o aparato flagelar tem orientação contrária à descrição anterior e os corpos basaisgeralmente ocorrem de forma sobreposta (Figura 2.54b); e diretamente oposto, quando oscorpúsculos basais se dispõem como se fossem prolongamentos um do outro (Figura 2.54c).A orientação horária dos corpos basais ocorre na classe Chlorophyceae, ordens Chlorococcalese Chlamydomonadales. A orientação anti-horária é típica de Ulvophyceae e Trebouxiophyceae.A orientação oposta dos corpos basais ocorre na classe Chlorophyceae, ordens Sphaeropleales,Chaetophorales e Chaetopeltidales. As algas carofíceas não apresentam aparato flagelar cruciado,sendo dotadas de raízes flagelares altamente assimétricas e de estrutura especial e singularentre as algas verdes, a estrutura em camadas múltiplas (MLS) (Figura 2.54d). A MLS decarofíceas está associada aos corpos basais flagelares e é extremamente semelhante a MLSsgeradas por gametas flagelados produzidos por plantas terrestres. Supõe-se que as MLSs atuamcomo centros organizadores de microtúbulos, mas ainda não se sabe ao certo se este érealmente seu papel. As prasinofíceas apresentam sistemas flagelares com raízes cruciadas,mas há grande variação nas características de seu aparato flagelar, não se enquadrando bemem nenhum dos tipos básicos descritos anteriormente. Ocorrem espécies com número ímparde raízes microtubulares e espécies com estrutura semelhante a MLS das carofíceas (Hoek etal., 1995). Além dos componentes já citados, o aparato flagelar de clorófitas pode incluirtambém rizoplastos, organelas especiais que ocorrem na maioria das clorófitas.
Características da mitose e da citocinese são também de importância acentuada no estudodas clorófitas. Em clorofíceas e ulvofíceas, a mitose ocorre sem que haja ruptura do envelopenuclear, configurando mitose fechada. Nas clorofíceas ocorre duplicação do núcleo e os doisnúcleos filho permanecem próximos entre si; os microtúbulos do fuso organizam-se na regiãoequatorial da célula em divisão, entre os núcleos e de forma paralela ao plano de divisão celular,formando um ficoplasto (Figura 2.55a).
Nas ulvofíceas não há formação de ficoplasto e o fuso mitótico é persistente na telófase;a citocinese não se processa imediatamente após a mitose e consiste em invaginação centrípetada membrana plasmática. Nas carofíceas há desintegração completa do núcleo e o fuso é abertodurante o processo de divisão celular. Os núcleos filho são formados e transportados paraextremos opostos e o fuso persiste formando um fragmoplasto (Figura 2.55b). Na maioriadas algas a divisão celular se completa com a formação de um sulco, que consiste no crescimentoda periferia para o interior da célula da membrana plasmática na região equatorial, seguido daformação da parede celular (Figura 2.56a). Em poucas algas e nas briófitas e traqueófitas a divisãocelular se completa com a formação de placas celulares na região mediana entre as duas clelulasfilhas. A deposição de materiais que compõem a parede celular decorre de atividade intensa doaparato de Golgi e inicia-se na região central da parede em formação, progredindo para a periferiadas células, até que se processa a união com as paredes laterais (Figura 2.56b). Esse padrão ocorreem parte das clorofíceas e das carofíceas.

124
sd
PF
DF
a) d
s
d
s
s
d
d
s
d
s
s
dd
s
b)
tc
s
dd
s
c)
s
d
d
s
s
MLS
d
s
d)
d
Figura 2.54 Quatro tipos de aparato flagelar encontrados em algas verdes, vistos de lado (esquerda) oude cima (direita). Estes caracterizam as principais linhagens evolutivas da divisão. Os aparatos geralmenteincluem dois ou quatro corpos basais (mostrados aqui como cilindros ou retângulos), raízes microtubulares(s ou d) e fibras conectivas distais (DF) e/ou proximais (PF). (a) Aparato flagelar com raízes em formacruciada, mostrando deslocamento em sentido horário dos corpos basais flagelares a partir de uma linhaimaginária paralela e entre eles. (b) Aparato flagelar com raízes cruciadas e corpos basais deslocados emsentido anti-horário. (c) Aparato flagelar com corpos basais diretamente em oposição. (d) Aparato flagelarcom distribuição assimétrica das raízes flagelares, mostrando a característica estrutura em camadas múltiplas(MLS). Reproduzido de Graham & Wilcox (2000).
A reprodução vegetativa em clorófitas ocorre através de divisão celular simples e defragmentação de talos filamentosos, multicelulares macroscópicos, por exemplo. Formasunicelulares também podem produzir esporos, como já é bem documentado paraChlamydomonas. Formas coloniais podem gerar colônias filha a partir da produção de zoósporos,

125
que se agregam, se diferenciam morfologicamente e originam novas colônias da espécie.Formas multicelulares complexas, como Chara, não formam esporos, mas são capazes derealizar propagação vegetativa do talo.
nu
nu
nu
nu
a) b)
a) b)
A reprodução sexuada é comum nas clorófitas, envolvendo isogamia, anisogamia eoogamia. Os gametas de clorófitas são células especializadas e não células vegetativas, excetono caso de espécies de Volvocales, nas quais células vegetativas podem agir como gametas(Lee, 1999). Gametângios bem marcantes são formados nas espécies que praticam oogamia,ao passo que os gametângios são discretos ou ausentes no caso das espécies que realizamanisogamia e isogamia. Na grande maioria das espécies os gametas são flagelados (zooga-metas), embora aplanogametas e gametas amebóides sejam reconhecidos em alguns grupos,como as Zygnematales. A maioria das prasinofíceas, clorofíceas, trebouxiofíceas e carofíceasapresenta uma fase vegetativa haplóide, sendo o zigoto a única fase diplóide do ciclo de vida,a qual sofre meiose. As ulvofíceas apresentam alternância de gerações, com meiose espórica.Apesar dessas considerações básicas, há enorme riqueza de padrões e detalhes nos ciclos sexuaisde algas verdes, alguns dos quais são apresentados a seguir. Em formas unicelulares comoChlamydomonas ocorrem duas mitoses sucessivas que culminam na formação de quatrogametas haplóides menores que as células vegetativas. O pareamento, a fusão e a cariogamiados gametas resultam na formação de um zigoto diplóide, que sofre meiose e gera célulasvegetativas haplóides. Em formas filamentosas como Spirogyra ocorre conjugação. Primei-ramente, há o pareamento entre filamentos, que se mantêm próximos pela ação adesiva dabainha de mucilagem. Algumas células de um dos filamentos, tratado como masculino,desenvolvem projeções em direção ao segundo filamento, dito feminino. Forma-se um tuboconjugativo entre os filamentos e os conteúdos das células masculinas migram para dentrodas células femininas, funcionando como gametas especiais. A fusão acontece nos filamentosfemininos e formam-se zigotos diplóides, os zigósporos, que têm conteúdo denso e são
Figura 2.55 Padrões de arranjos de microtúbulose de separação dos núcleos encontrados em algasverdes. (a) Formação de ficoplasto. (b) Formaçãode fragmoplasto. Reproduzido de Sze (1998).
Figura 2.56 Padrões de citocinese em algas verdes.(a) Formação de sulco. (b) Deposição de placascelulares. Reproduzido de Sze (1998).

126
liberados no meio. Os zigósporos sofrem meiose e três dos quatro núcleos formados sofremdesintegração. A célula resultante, dotada de apenas um núcleo, origina os filamentos vege-tativos da espécie (Figura 2.57).
Filamentomasculino
Tuboconjugativo
Gametamasculino
Gametafeminino
Fusão
Mucilagem
PapilaN
2N
Núcleo
Cloroplasto
Pirenóides
Germinação
Zigósporos
GerminaçãoMeiose
Desintegraçãode três núcleos
Figura 2.57 Ciclo de vida de Spirogyra. Modificado de Lee (1999).
Outra forma peculiar de ciclo de vida é apresentada por carofíceas multicelulares, comoChara. A reprodução sexuada ocorre através de oogamia, sendo que as estruturas reprodutivassão formadas nos nós. Trata-se de estruturas multicelulares complexas, conhecidas comoglóbulos e núculas. Núculas apresentam ovócitos protegidos por células tubulares e umaabertura apical pela qual penetram os anterozóides. Glóbulos também são protegidos porcélulas especiais, as células-escudo, que circundam os filamentos anteridiais. Os anterídiossão liberados na água, atingem e penetram as núculas e fertilizam os ovócitos. Os zigotossão liberados na água, sofrem meiose e as células resultantes são capazes de originar as formasmacroscópicas típicas da espécie (Figura 2.58).
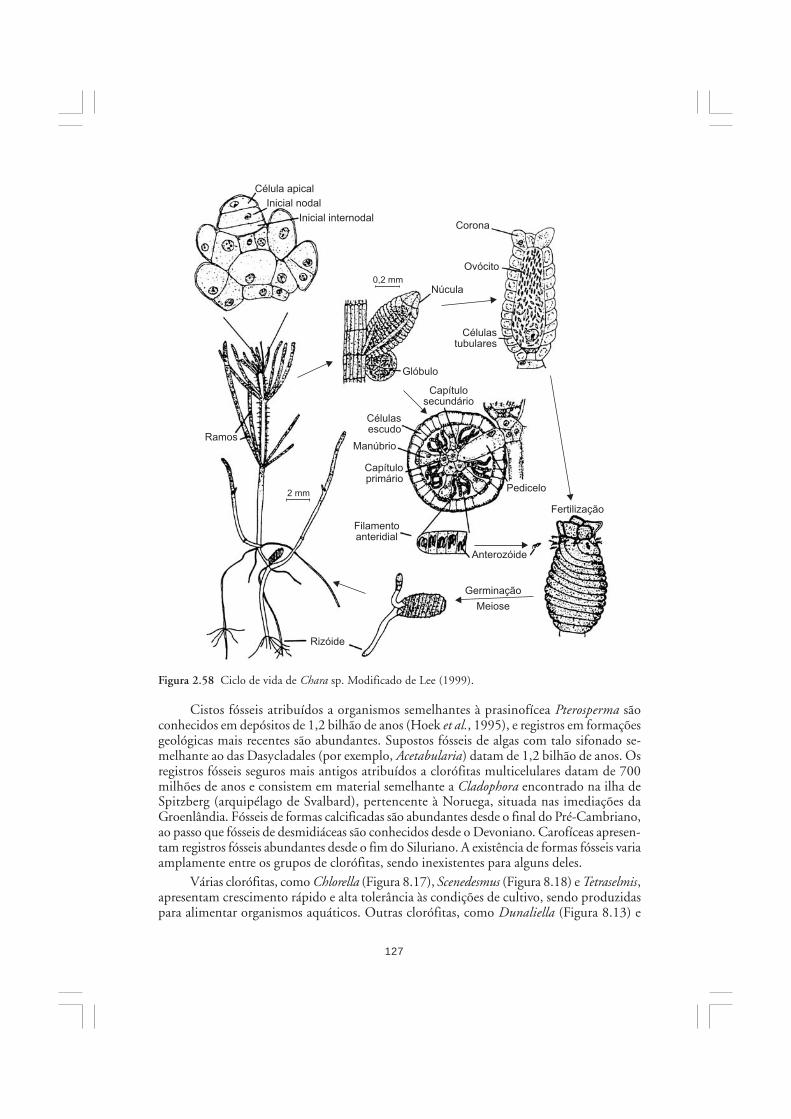
127
0,2 mm
Célula apical
Inicial nodal
Inicial internodal
Núcula
Glóbulo
Corona
Ovócito
Célulastubulares
Pedicelo
Capítulosecundário
Célulasescudo
Manúbrio
Capítuloprimário
2 mm
Fertilização
Anterozóide
Filamentoanteridial
Germinação
Meiose
Rizóide
Ramos
Figura 2.58 Ciclo de vida de Chara sp. Modificado de Lee (1999).
Cistos fósseis atribuídos a organismos semelhantes à prasinofícea Pterosperma sãoconhecidos em depósitos de 1,2 bilhão de anos (Hoek et al., 1995), e registros em formaçõesgeológicas mais recentes são abundantes. Supostos fósseis de algas com talo sifonado se-melhante ao das Dasycladales (por exemplo, Acetabularia) datam de 1,2 bilhão de anos. Osregistros fósseis seguros mais antigos atribuídos a clorófitas multicelulares datam de 700milhões de anos e consistem em material semelhante a Cladophora encontrado na ilha deSpitzberg (arquipélago de Svalbard), pertencente à Noruega, situada nas imediações daGroenlândia. Fósseis de formas calcificadas são abundantes desde o final do Pré-Cambriano,ao passo que fósseis de desmidiáceas são conhecidos desde o Devoniano. Carofíceas apresen-tam registros fósseis abundantes desde o fim do Siluriano. A existência de formas fósseis variaamplamente entre os grupos de clorófitas, sendo inexistentes para alguns deles.
Várias clorófitas, como Chlorella (Figura 8.17), Scenedesmus (Figura 8.18) e Tetraselmis,apresentam crescimento rápido e alta tolerância às condições de cultivo, sendo produzidaspara alimentar organismos aquáticos. Outras clorófitas, como Dunaliella (Figura 8.13) e

128
Haematococcus (Figura 8.27), têm sido cultivadas para extração de carotenóides utilizadoscomo corantes naturais. Espécies multicelulares dos gêneros Enteromorpha e Monostromasão cultivadas no Extremo Oriente para uso na alimentação humana, atividade que movi-menta milhões de dólares ao ano. Algumas algas clorófitas unicelulares podem formarflorações intensas, mas não há espécies produtoras de ficotoxinas. Algumas espécies multice-lulares têm capacidade de se tornarem invasoras, como Codium fragile e, em especial, Caulerpataxifolia, pelo enorme impacto já causado no Mar Mediterrâneo após sua introdução acidental(Meinesz, 1999). Algumas clorófitas multicelulares, como Ulva, Cladophora e Enteromorpha,são bastante resistentes aos efeitos de poluição doméstica e industrial e tendem a se tornardominantes em locais rasos sob forte impacto antrópico.
Numa divisão composta por milhares de espécies é bastante difícil listar gêneros maisimportantes ou representativos. Porém, em razão de sua importância ambiental ou usos pelohomem, além dos gêneros citados na Tabela 2.10, certamente alguns outros não podem deixarde ser citados, como Acetabularia, Acrosiphonia, Botryococcus, Bryopsis, Caulerpa,Chaetomorpha, Codium, Cosmarium, Desmidium, Dictyosphaera, Dunaliella, Enteromorpha,Halimeda, Micrasterias, Pandorina, Pediastrum, Pyramimonas, Staurastrum, Stichococcus,Trentepohlia e Ulothrix.
2.3 ALGUMAS REFLEXÕES SOBRE A DIVERSIDADE DAS ALGAS
A criação e a aplicação de sistemas de classificação de seres vivos é uma necessidade daBiologia: permite melhor avaliação das relações filogenéticas entre organismos e ofereceperspectiva mais sistemática de estudo dos componentes dos grupos. Os sistemas de classi-ficação são aplicados em vários níveis, desde os táxons menores (por exemplo, família) atéos táxons elevados (por exemplo, divisão ou filo). As mudanças na abrangência e na com-posição dos grupos são normais e até esperadas, dada a aquisição de mais conhecimentossobre os organismos e a própria descrição de novas espécies. Grupos grandes, que encerramcomponentes com elevado grau de diversidade estrutural e fisiológica (por exemplo, crus-táceos), são naturalmente sujeitos a mais mudanças. Entretanto, estabelecer relações filo-genéticas entre algas é possivelmente um dos desafios supremos da Biologia.
Na realidade, os protistas são, desde sua conceituação, problemáticos do ponto de vistasistemático, pois não constituem um grupo natural. Trata-se, de fato, de um conjunto muitogrande de grupos de organismos eucarióticos de organização celular simples. Numa simpli-ficação, os protistas consistem em todos os organismos que não se enquadram em outrosreinos. O termo “alga” envolve predominantemente organismos classificados como protistas,mas pode ser estendido a bactérias e até a uma linhagem que envolve as plantas (carofíceas).É uma abrangência gigantesca, a qual cria alguns problemas sistemáticos: enquanto algunsgrupos de algas não exibem qualquer evidência de parentesco, outros são semelhantes entresi e outros apresentam grau intermediário de semelhança.
Por muitas décadas, os grupos de algas foram organizados de forma meramente con-veniente, e nem sempre as características diagnósticas mais importantes eram consideradas.Um bom exemplo dessa tendência era a classificação das algas pardas como uma divisão,Phaeophyta. Possivelmente, por tradição e para manter as algas pardas na mesma hierarquiaem relação às formas multicelulares de rodófitas e clorófitas. Hoje sabe-se que as algas pardasestão muito mais relacionadas a diatomáceas e rafidofíceas do que às rodófitas e clorófitas.As dimensões dos organismos têm pouco valor para sua classificação. Informações sobrecomposição química, ultra-estrutura, reprodução e composição molecular têm sido decisivaspara o estabelecimento de relações filogenéticas entre algas.

129
A elucidação de caminhos evolutivos após o estabelecimento de possíveis processosestáveis de endossimbiose também é fundamental para a compreensão das algas. Eventos deendossimbiose primária, secundária e terciária devem ter determinado grupos modernos dealgas, e mudanças ocorridas desde então (por exemplo, perda de certos pigmentos por algumasespécies ou grupos) são peças de um quebra-cabeça bastante complexo. A ocorrência denutrição exclusivamente heterotrófica em alguns dinoflagelados e euglenóides, por exemplo,mostra que a própria fotossíntese não é característica universal para as algas. O termo “alga”certamente continuará a ser uma palavra meramente conveniente, pois decididamente nãorepresenta uma unidade taxonômica.
Apesar dessas considerações, a busca pelo estabelecimento de relações filogenéticas entregrupos de algas continuará ocorrendo, e espera-se que a agregação de muitas informaçõesnovas no futuro conduza a modelos mais consistentes. A Figura 2.59 é uma representaçãodas possíveis relações entre os grandes grupos de organismos eucarióticos, enfatizando asdiversas posições ocupadas pelas algas.
ClorófitasEmbriófitas
Rodófitas
Metazoários
Fungosbasidiomicetos,ascomicetos,
quitridiomicetos
Estramenópilascrisofíceas, diatomáceas,
feofícias, oomicetos,opalinídeos
Alveolatosciliados,
dinoflagelados,apicomplexos
Euglenozoárioseuglenóides,
kinetoplastídeos
Priminesiófitas Criptomônadas
Glaucófitas
Cloraracniófitas
Figura 2.59 Relações entre algas eucarióticas e outros grupos de organismos eucarióticos com base eminformações codificadas pela subunidade menor do ARN ribossômico. Modificado de Graham & Wilcox(2000).

Correções do Capítulo 2 Cultivo de Microalgas Marinhas – Princípios e Aplicações
1 ok Página 33 Linha 12. Retirar o negrito de “fitoplâncton”. 2 ok Página 33 Penúltima linha do primeiro parágrafo. Substituir “não ocorre” por “ocorre pouca ou nenhuma”. 3 Não foi feito Página 34 Segunda linha. Retirar a vírgula após “organização”. 4 ok Página 34 Terceira linha do quarto parágrafo. Retirar o “s” de “marismas”. 5 ok Página 34 Sexta linha do quarto parágrafo. Retirar o “s” de “marismas”. 6 ok Página 35 Primeira linha do segundo parágrafo. Passar “seja derivada” para o plural: “sejam derivadas”. 7 Não foi feito Página 35 Sétima linha do segundo parágrafo. Retirar a vírgula após “apresentados”. 8OK Página 36 Tabela 2.1, terceira coluna. Retirar o “n” de “Gallangher”: o correto é “Gallagher”. 9 OK Página 36 Tabela 2.1, terceira coluna. Em “Ponte et al. (1984)”, substituir o ano por “1990”. 10 ok Página 37 Este é muito grave. Sétima linha do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”. A letra grega “μ” não entrou aqui e em várias outras partes do texto. 11ok Página 37 Igual ao anterior. Nona linha do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”. A letra grega “μ” não entrou aqui e em várias outras partes do texto. 12ok Página 37 Igual ao anterior. Linha 11 do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”, quatro vezes na mesma linha: “50 μm”, “63 μm”, “80 μm” e “20 μm”. 13 ok Página 37 Igual ao anterior. Penúltima linha do segundo parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”: “20 μm”. 14 ok Página 38 Igual ao anterior. Décima linha do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”: “200 μm”.

15 ok Página 38 Igual ao anterior. Linha 12 do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”: “20 μm”. 16 ok Página 38 Igual ao anterior. Linha 14 do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”: “2 μm”. 17 ok Página 38 Igual ao anterior. Linha 16 do primeiro parágrafo. Símbolo distorcido: em vez de “mm”, o correto é “μ”: “0,2 μm”. 18 ok Página 39 Tabela 2.3, interseção da terceira coluna (picoplâncton) com a quinta linha (fitoplâncton). Colocar um pontilhado no espaço (como está na célula vizinha, à direita), faltou preencher este espaço. 19 ok Página 39 Tabela 2.3, interseção da oitava coluna (megaplâncton) com a oitava linha (tamanho). Retirar o “-“ de “10-0”: deve ficar como “100”. 20 ok Página 41 Legenda da Figura 2.2. Após a palavra “criptomônada”, acrescentar um “;” e “DN = núcleo do dinoflagelado”. 21 ok Página 41 Legenda da Figura 2.2. Substituir “1993” por “1985”. 22 ok Página 43 Quinta linha. Substituir “dessas algas” por “das algas”. 23 ok Página 43 Sétima linha. Retirar o negrito de “criptófitas”. 24 ok Página 43 Figura 2.4. Problema grave: a figura está cortada e não aparecem as letras “a”, “b”, ... “i”, citadas na legenda. A parte de baixo da última fórmula está também cortada. 25 ok Página 44 Sexta linha. Retirar o negrito de “ficobilina”. 26 ok Página 44 Oitava linha. Retirar o negrito de “ficocianinas”. 27 ok Página 44 Nona linha. Retirar o negrito de “ficoeritrinas”. 28 ok Página 44 Figura 2.5. Problema grave: a figura está cortada e a parte de cima da fórmula “a” está sem as duas carboxilas (COOH). 29 ok

Página 45 Figura 2.6. Eliminar uma linha vertical que está aparecendo no lado direito da ilustração. 30 ok Página 46 Terceira linha. Retirar o negrito de duas palavras: “gametófitos” e “esporófitos”. 31ok Página 48 Tabela 2.4, primeira coluna. Retirar o negrito de “Proclorofíceas”, pois este é um grupo de “Chloroxybacteria” (célula acima). 32 ok Página 48 Tabela 2.4, sexta coluna (“Cobertura celular”). Alinhar o texto à esquerda – ele está centralizado. 33 ok Página 49 Tabela 2.4, primeira coluna. Retirar a divisão silábica de “feo-fíceas”. Grafar “feofíceas”. 34 ok Página 49 Tabela 2.4, primeira coluna. Retirar o negrito de “Eustigmatofíceas”, pois este é um grupo de “Ochrophyta” (célula acima). 35 ok Página 49 Tabela 2.4, quarta coluna (“Flagelo”). Alinhar o texto à esquerda – ele está centralizado. 36 ok Página 49 Tabela 2.4, sexta coluna (“Cobertura celular”). Alinhar o texto à esquerda – ele está centralizado. 37 ok Página 50 Este é muito grave. Legenda da Figura 2.8. Símbolo distorcido: em vez de “a-1,4”, o correto é “α-1,4”. A letra grega “α” não entrou aqui e em várias outras partes do texto. 38 ok Página 51 Terceiro parágrafo, quarta linha. Retirar o negrito de “ficobiliproteínas”. 39 ok Página 51 Terceiro parágrafo, quinta linha. Substituir “ficobilissomas” por “ficobilissomos” (trocar o “a” por “o”). 40 ok Página 52 Primeiro parágrafo, segunda linha. Símbolo distorcido: em vez de “á-1,4”, o correto é “α-1,4”. A letra grega “α” não entrou aqui e em várias outras partes do texto. 41 ok Página 52 Segundo parágrafo, segunda linha. Substituir “carboxissomas” por “carboxissomos” (trocar o “a” por “o”). 42 ok Página 52 Segundo parágrafo, terceira linha. Mesma correção anterior. Substituir “carboxissomas” por “carboxissomos” (trocar o “a” por “o”). 43 ok

Página 53 Linha 16. Símbolo distorcido: em vez de “poli-â-hidroxibutírico”, o correto é “poli-β-hidroxibutírico”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 44 ok Página 53 Segunda linha do primeiro parágrafo. Usar inicial maiúscula em “Gram”, que é um nome próprio: “Gram-negativas”. 45 Não foi feito Página 54 Terceira linha. Substituir “células-filhas” por “células filhas”, sem hífen. O texto será todo padronizado assim. 46 ok Página 55 Legenda da Figura 2.11. Substituir “Universidade de” por “University of”. Assim está nas referências bibliográficas. 47 ok Página 56 Quinta linha. Acrescentar o artigo “um” antes da palavra “ambiente”. 48 ok Página 57 Linha 11. Substituir “Figura 8.29” por “Figura 8.1”. 49 ok Página 57 Primeira linha do segundo parágrafo. Retirar o negrito de “florações”. 50 ok Página 58 Linha 20 do primeiro parágrafo. Substituir “carboxissomas” por “carboxissomos” (trocar o “a” por “o”). 51ok Página 59 Sexta linha do primeiro parágrafo. Substituir “1971” por “1978”. 52ok Página 59 Tabela 2.7. Há uma linha vazia “sobrando” antes da que se inicia com “Ordem Nostocales”. 53ok Página 59 Último parágrafo, terceira linha. Substituir “Figura 8.30” por “Figura 8.25”. 54 ok Página 59 Último parágrafo, terceira linha. Substituir “Figura 8.29” por “Figura 8.1”. 55 ok Página 60 Primeiro parágrafo, quinta linha. Substituir “Geiter” por “Geitler”. Falta um “l”. 56 ok Página 60 Primeiro parágrafo, antepenúltima linha. Está tudo correto, pode apagar seu alerta (ESTÁ CERTO?). 57 ok Página 60

Segundo parágrafo, sétima linha. Colocar a palavra “monadal” em negrito, pois é a primeira citação deste verbete do glossário. 58 ok Página 61 Terceira linha. Retirar o negrito da palavra “euglenófitas”. 59 não foi feito Página 61 Terceiro parágrafo, sétima linha. Acrescentar uma vírgula depois da palavra “alternam”. 60ok Página 61 Quarto parágrafo, antepenúltima linha. Substituir a preposição “por” por “em”. 61 ok Página 61 Última linha. Substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 62 Página 61 Última linha. Substituir “â-criproxantina” por “β-criproxantina”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 63 Não foi feito Página 62 Terceira e última linha do segundo parágrafo. Retirar a vírgula após a palavra “vasculares”. 64 ok Página 63 Terceira a quinta linhas do segundo parágrafo. Antecipar a citação da “Figura 2.15” (quinta linha) para logo após a palavra “mônadas” (terceira linha). 65 ok Página 63 Décima linha do segundo parágrafo. Substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 66 ok Página 63 Linha 14 do segundo parágrafo. Colocar a palavra “heterocontófitas” em negrito, pois é a primeira citação deste verbete do glossário. 67 Não foi feito Página 64 Sexta linha do primeiro parágrafo. Substituir “se contraem e se distendem” por “contraem-se e distendem-se”. A ênclise é mais indicada neste caso. 68 ok Página 67 Nona linha do segundo parágrafo. Substituir “plastidial” por “periplastidial”, em negrito. É assim que o verbete está apresentado no glossário. 69 ok Página 67 ok Linha 18 do segundo parágrafo. Dois símbolos distorcidos. Substituir “á-caroteno, â-caroteno” por “α-caroteno, β-caroteno”. 70 ok Página 67 Linha 28 do segundo parágrafo. Substituir “ficobilissomas” por “ficobilissomos” (trocar o “a” por “o”). 71 ok Página 67 Linha 30 do segundo parágrafo. Mesma correção anterior. Substituir “ficobilissomas” por “ficobilissomos” (trocar o “a” por “o”). 72 ok Página 70 Legenda da Figura 2.17. Antes de “sf = flagelo curto”, acrescentar “SS = escamas”. Isto ficou faltando.

73 ok Página 70 Legenda da Figura 2.18. Substituir “Syacosphaera” por “Syracosphaera”. Faltou um “r”. 74 ok Página 72 Sétima linha do primeiro parágrafo. Grafar “19’-hexanoiloxifucoxantina” em vez de “19’-hexanoyloxyfucoxantina” (trocar os dois “y” por dois “i”). 75 ok Página 72 Sétima linha do primeiro parágrafo. Grafar “19’-butanoiloxifucoxantina” em vez de “19’-butanoyloxyfucoxantina” (trocar os dois “y” por dois “i”). 76 ok Página 72 Primeira linha do quarto parágrafo. Substituir “assexuada” por “vegetativa”. 77 não foi feito Página 74 Segunda linha do segundo parágrafo. Grafar “oceano” com “o” minúsculo: “oceano Atlântico”. 78 não foi feito Página 75 Oitava linha do primeiro parágrafo. Substituir “oxidado” por “oxidada” (trocar o segundo “o” por “a”). 79 ok Página 75 IMPORTANTE. Última frase do segundo parágrafo: eliminar toda a frase (“A reprodução sexuada é bastante difundida no grupo e envolve uma alternância de gerações heteromórfica”), pois ela já foi escrita páginas atrás. 80 ok Página 75 Segunda linha do quarto parágrafo. Substituir Gephryocapsa por Gephyrocapsa. As letras “r” e “y” estão invertidas. 81 ok Página 76 Legenda da Figura 2.19. Substituir “Jangen” por “Tangen”. 82 ok Página 78 Quarta linha. Símbolo distorcido. Substituir “â-caroteno” por “β-caroteno”. 83 ok Página 79 Linha 13. Tirar o negrito de “mucocisto”. 84 ok Página 79 Nona linha do primeiro parágrafo. Tirar o negrito de “haplôntico”. 85 ok Página 79 Décima linha do quadro “O extraordinário ciclo...”. Apagar “(Burkholder et al., 1992)”. É desnecessário citar o trabalho mais uma vez. 86 ok Página 80 Décima linha do terceiro parágrafo do quadro “O extraordinário ciclo...”. Apagar “(Lee, 1999)”. É desnecessário citar o trabalho mais uma vez. 87 ok

Página 80 Linha 11 do terceiro parágrafo do quadro “O extraordinário ciclo...”. Apagar “(Burkholder & Glasgow, 1997)”. É desnecessário citar o trabalho mais uma vez. 88 ok Página 81 ok Linha 13 do quadro “O extraordinário ciclo...”. Apagar “(Litaker et al., 2002)”. É desnecessário citar o trabalho mais uma vez. 89 ok Página 82 Primeira linha do terceiro parágrafo. Retirar o negrito de “bioluminescência”. 90ok Página 82 Sétima linha do terceiro parágrafo. Símbolo distorcido. Acrescentar “≈” antes de “50 kcal”. 91 ok Página 83 Linha 16 do terceiro parágrafo. Retirar o negrito das palavras “ocrófita” e “heterocontófita”. 92 ok Página 84 Terceira linha. Eliminar a frase “cujo significado foi apresentado anteriormente”, a qual está dentro dos parênteses. 93ok Página 86 Sétima linha. Acrescentar a palavra “tais” antes de “movimentos”. 94 ok Página 86 Figura 2.26. MUITO IMPORTANTE. A figura está cortada no topo e na base. As barras que indicam os tamanhos das células não estão aparecendo na base da figura. 95 ok Não foi feito Página 86 Terceira linha do último parágrafo. Acrescentar uma vírgula após “etc.”. 96 ok Página 85 Figura 2.27. MUITO IMPORTANTE. A figura está cortada à esquerda. 97 ok Página 87 Legenda da Figura 2.27. Substituir “5A” e “5B” por “5a” e “5b” (usar letras minúsculas). 98 ok Página 87 Legenda da Figura 2.27. Eliminar o espaço antes de “T. frauenfeldii”. 99 ok Página 87 Segundo parágrafo, terceira linha. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 100 não foi feito Página 88 Terceira linha do terceiro parágrafo. Substituir “células-filha” por “células filhas”, sem hífen.

101 Não foi feito Página 92 Primeira linha do primeiro parágrafo. Acrescentar uma vírgula após a palavra “grupo”. 102 ok Página 92 Retirar o espaço em branco entre o terceiro e o quarto parágrafos. 103 ok Página 93 Legenda da Figura 2.33. Falta o nome de um dos autores do trabalho: o correto é “Marchant & McEldowney (1986)”. 104 ok Página 93 Terceira linha. Grafar “oceano” com “o” minúsculo: “oceano Antártico”. 105 Não foi feito Página 93 Quarta linha. Grafar “oceano” com “o” minúsculo: “oceano Pacífico”. 106 ok Página 95 Primeira linha. Substituir “loricados” por “loricadas” (trocar o segundo “o” por “a”). 107 ok Página 95 Quarta linha do segundo parágrafo. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 108 ok Página 96 Legenda da Figura 2.36, segunda linha. Acrescentar “(3)” após a palavra “cocóide”. 109 ok Página 97 Décima linha. Retirar a vírgula após a conjunção “e”. A vírgula está “sobrando”. 110 ok Página 97 Legenda da Figura 2.37. Substituir “carterae” por “akashiwo” (a espécie mudou de nome recentemente). 111 ok Página 99 Terceira linha do terceiro parágrafo. Retirar o negrito de “xantofíceas”. 112 ok Página 99 Legenda da Figura 2.40, antepenúltima linha. Substituir “espermatozóides” por “anterozóides”. 113 ok Página 99 Legenda da Figura 2.40, penúltima linha. Substituir “espermatozóide” por “anterozóide”. 114 ok Página 100 Sétima linha. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 115 ok Página 101 Quinta linha após a figura. Após “filamentoso”, colocar a frase “, (presente nas formas mais simples),” entre parênteses (colocar os parênteses onde agora estão as vírgulas).

116 ok Página 102 Quarta linha. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 117 ok Página 102 Segunda linha do segundo parágrafo. Retirar o negrito de “laminarina”. 118 ok Página 102 terceira linha do segundo parágrafo. Símbolo distorcido: substituir “â-1,3” por “β-1,3”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 119ok Página 103 Sexta linha do segundo parágrafo. Substituir “heterocontos” por “heterocontes” (trocar o terceiro “o” por “e”). 120 ok Página 103 Oitava linha do segundo parágrafo. Substituir “heterocontos” por “heterocontes” (trocar o terceiro “o” por “e”). 121 ok verificar É desmarestiales? Página 104 Tabela 2.8. Primeira coluna. Substituir “Desmerestiales” por “Desmerestiales” (trocar o segundo “e” por “a”). 122 ok Página 104 Tabela 2.8. Segunda coluna. Retirar o sublinhado de “multisseriados” (na terceira linha). 123 ok Página 104 Tabela 2.8. Quarta coluna. Retirar a separação silábica de “anisoga-mia” (na terceira linha). 124 ok Página 104 Tabela 2.8. Sexta coluna. Retirar a separação silábica de “Ecklo-nia” (na linha 11). 125 ok Página 105 Terceira linha do segundo parágrafo. Substituir “hiziki” por “hijiki” (trocar o “z” por “j”). 126ok Página 105 Retirar o espaço em branco entre os parágrafos 5 e 6. 127 ok Página 105 Sexto parágrafo, oitava linha. Substituir “, que” (após “zoósporos”) pela conjunção “e”. 128 ok Página 105 Quarta linha. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 129ok Página 106

Segundo parágrafo, última linha. Substituir “2002” por “2002b”. 130ok Página 107 Legenda da Figura 2.45. Substituir “C” por “P”. 131 ok Página 108 Segunda linha do segundo parágrafo. Acrescentar um gênero após Synura: Tessellaria. A frase refere-se a quatro gêneros, mas só três estão escritos. 132 ok Página 108 Quarta linha do segundo parágrafo. Está tudo correto, pode apagar seu alerta (ESTÁ CERTO?). 133 ok Página 109 Segunda e terceira linhas linha do último parágrafo. Colocar “células apicais” em negrito. 134ok Página 110 Legenda da Figura 2.47. Eliminar “1985” e “1995” e acrescentar “(1978)”. 135ok Página 111 Sétima linha do segundo parágrafo, quinta linha. Substituir “ficobilissomas” por “ficobilissomos” (trocar o “a” por “o”). 136ok Página 112 Linha 12 do segundo parágrafo. Símbolo distorcido: substituir “â-caroteno” por “β-caroteno”. A letra grega “β” não entrou aqui e em várias outras partes do texto. 137não foi feito Página 112 Terceira linha do segundo parágrafo do quadro. Grafar “oceano” com inicial minúscula: “oceano Pacífico”. 138ok Página 113 MUITO IMPORTANTE. O quadro termina após o segundo parágrafo. Todo o restante dever ser apresentado como texto normal. 139 ok Página 113 Terceiro parágrafo, segunda linha. Símbolo distorcido: substituir “á-1,4” por “α-1,4”. A letra grega “α” não entrou aqui e em várias outras partes do texto. 140 ok Página 113 Terceiro parágrafo, quinta linha. Símbolo distorcido: substituir “á-1,6” por “α-1,6”. A letra grega “α” não entrou aqui e em várias outras partes do texto. 141ok Página 113 Última linha. Substituir “parece” por “parede”. 142ok Página 114

Legenda da Figura 2.49. Escrever “Ágar” em negrito. 143 ok Página 114 Legenda da Figura 2.49. Escrever “kappa” sem itálico. 144 ok Página 115 Segunda linha. Escrever “monósporos” sem negrito. 145 ok Página 119 Legenda da Figura 2.52. Os autores estão errados: substituir “Graham & Wilcox (2000)” por “Mumford Jr. & Miura (1988)”. 146ok Página 120 Terceira linha. Eliminar as informações entre parênteses: “(cerca de 90% das espécies)”. Esta informação aparece depois com mais clareza. 147ok Página 120 Terceira linha. Eliminar as informações entre parênteses: “(cerca de 90% das espécies)”. Esta informação aparece depois com mais clareza. 148 ok Página 121 Legenda da Tabela 2.10. Substituir “grupos” por “classes”. 149 ok Página 121 Tabela 2.10, primeira coluna. Acrescentar a palavra “Classe” no topo da coluna, que está em branco. 150 ok Página 121 Tabela 2.10, primeira coluna, terceira linha. Substituir “Tribouxiophyceae” por “Trebouxiophyceae” (trocar o primeiro “i” por “e”). 151ok Página 121 Tabela 2.10, terceira coluna, terceira linha. Retirar a divisão silábica de “desidro-genase”. 152ok Página 121 Tabela 2.10, terceira coluna, quarta linha. Retirar a divisão silábica de “desidro-genase”. 153ok Página 121 Tabela 2.10, terceira coluna, quinta linha. Retirar a divisão silábica de “desidro-genase”. 154ok Página 121 Tabela 2.10, oitava coluna, quarta linha. Substituir “Tribouxia” por “Trebouxia” (trocar o primeiro “i” por “e”). 155ok Página 122 Sexta linha. Retirar a vírgula após os parênteses. 156 ok Página 123 Juntar o segundo parágrafo ao primeiro – é o mesmo assunto.

157 não foi feito Página 123 |(124) Quarta linha do último parágrafo. Substituir “núcleos-filho” por “núcleos filhos”, sem hífen. 158 Não foi feito ok Página 124 Última linha. Substituir “núcleos-filho” por “núcleos filhos”, sem hífen. 159 Não foi feito ok Página 125 Quarta linha do primeiro parágrafo. Substituir “colônias-filha” por “colônias filhas”, sem hífen. 160 não foi feito Página 125 Quinta linha do primeiro parágrafo. Substituir “se diferenciam” por “diferenciam-se”. 161 ok Página 125 Antepenúltima linha do último parágrafo. Tirar o negrito de “aplanogametas”. 162 ok Página 126 Sétima linha. Tirar o negrito de “conjugação”. 163 ok Página 128 Primeira linha do primeiro parágrafo. Substituir “Figura 8.31” por “Figura 8.17”. 164 ok Página 128 Primeira linha do primeiro parágrafo. Após Scenedesmus, acrescentar “(Figura 8.18)”. 165 ok Página 128 Terceira linha do primeiro parágrafo. Após Dunaliela, acrescentar “(Figura 8.13)”. 166 ok Página 128 Terceira linha do primeiro parágrafo. Substituir “Figura 8.32” por “Figura 8.27”. 167 Não foi feito ok Página 128 Décima linha do primeiro parágrafo. Grafar “mar” com inicial minúscula: “mar Mediterrâneo”. 168 Não foi feito Página 129 Quinta linha do primeiro parágrafo. Substituir “sabe-se” por “se sabe”. A próclise é preferível após um advérbio. 169 ok Página 129 Sétima linha do primeiro parágrafo. Substituir “não têm nenhum” por “têm pouco”. 170 Não foi feito Página 129 Segunda linha do último parágrafo. Retirar a vírgula após “ocorrendo”. 171 ok Página 130 Legenda da Figura 2.59. Substituir “ribossomal” por “ribossômico”.