aac accepts, published online ahead of print on 30 ... · department of immunology and...
TRANSCRIPT

Chloroquine Modulates HIV-1 Induced Plasmacytoid Dendritic Cell IFNα:
Implication for T cell Activation
Running Title: Chloroquine Inhibits pDC Activation
Jeffrey A. Martinson1, Carlos J. Montoya
2, Xiomara Usuga
1, Rollie Ronquillo
1, Alan L. Landay
1,
Seema N. Desai1*
1. Department of Immunology and Microbiology, Rush University Medical Center, Chicago, IL-
USA
2. Group of Immunovirology-Biogenesis, University of Antioquia, Medellin-Colombia
Please address correspondence to:
Seema N. Desai, Ph.D.
Department of Immunology/Microbiology
Rush University Medical Center
1735 West Harrison St (Cohn Research Building Lab 641)
Chicago, IL 60612
Phone: 312 942 9292; FAX: 312 942-5206
Key Words: type 1 interferon, innate immunity, immune activation, endosomal inhibitors
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Antimicrob. Agents Chemother. doi:10.1128/AAC.01246-09 AAC Accepts, published online ahead of print on 30 November 2009
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Abstract
Plasmacytoid dendritic cell (pDC) mainly contribute to antiviral immunity through recognition
of microbial products and viruses via intracellular toll-like receptors (TLR) 7 or 9 that results in
the production of type I interferons. Though interferons reduce viral burden in acute phase of
infection their role in chronic phase is unclear. Presence of elevated plasma IFNα levels in
advanced HIV disease and its association with microbial translocation in chronic HIV infection
lead us to hypothesize that IFNα could contribute to immune activation. Blocking IFNα
production using chloroquine an endosomal inhibitor was tested in a novel in vitro model system
with the aim to characterize the effects of chloroquine on HIV-1 mediated TLR signaling, IFNα
production and T cell activation. Our results indicate that chloroquine blocks TLR mediated
activation of pDC and MyD88 signaling as shown by decrease in the downstream signaling
molecules IRAK-4, IRF-7 and inhibition of IFNα synthesis. Chloroquine decreased CD8 T cell
activation induced by AT-2 HIV-1 in PBMC cultures. In addition to blocking pDC activation,
chloroquine also blocked negative modulators of T cell response like IDO (indoleamine 2, 3-
dioxygenase) and PDL-1 (programmed death ligand-1). Our results indicate that TLR
stimulation and production of IFN-α by pDC contribute to immune activation, and blocking
these pathways using chloroquine may interfere with events contributing to HIV pathogenesis.
Our results suggests that a safe well tolerated drug such as chloroquine can be proposed as an
adjuvant therapeutic candidate along with HAART to control immune activation in HIV-1
infection.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Introduction
Plasmacytoid dendritic cells (pDC) are an integral part of the innate and adaptive immune
systems that recognize pathogens through toll-like receptor (TLR) 7 and 9 (2, 34, 42, 43, 45, 73).
Since these TLRs are intracellular, their ligands require cellular uptake and endosomal
maturation to trigger NF-κB and MAP kinase-mediated signals through the MyD88-dependent
pathway. These TLR signals result in pDC activation/maturation as well as the production of
pro-inflammatory cytokines and a large amount of type 1 interferon (IFN-α/β) (17, 43, 46). PDC
also provide negative regulatory signals that modulate and establish immune tolerance (62).
Several candidate molecules expressed by pDC are implicated in negative modulation of T cell
responses including the production of indoleamine 2, 3 dioxygenase (IDO) and the expression of
PDL-1. IDO, the rate limiting enzyme involved in tryptophan catabolism, inhibits CD4 T-cell
proliferation (8, 10, 53) and enhances T regulatory cell generation and suppressor cell function
(16). The PD-1/PDL-1 interaction results in decreased T cell proliferation, cytokine production
and cell mediated cytotoxicity (72). TLR mediated induction of PDL-1 expression on pDC may
contribute to T cell exhaustion and dysfunction through the PD-1/PDL-1 pathway. Despite a
decrease in the number of circulating pDC in HIV-1 infected individuals (3, 24, 26, 73, 74) their
stimulation by microbial products via TLR7, TLR9 (48) or by HIV itself (5) results in activation
of PDC and interferon-alpha (IFNα) production that contributes to pathogenesis in chronic HIV-
1 disease(14). IFNα is known to be associated with elevated LPS levels in chronic HIV infection
that may contribute to persistent immune activation (14).
Chloroquine, an anti-malarial drug, has immunomodulatory properties and is used in the
treatment of autoimmune disorders (28, 69, 81). It is also commonly used in vitro to study the
role of endosomal acidification in cellular processes, such as the TLR activation pathways in
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

pDC induced by HIV-1 (5, 9, 25, 35, 71). It is a weak base that accumulates within the
endosome of cells leading to an elevated vacuolar pH thereby inhibiting endosomal maturation
and nucleic acid binding to TLR7 and TLR9 (68). HIV-1 activates pDC through TLR7 (5) which
makes chloroquine a candidate for preventing HIV-1 induced activation and subsequent down-
stream effects on T cell activation and function.
The goal of the present study was to characterize the effects of chloroquine on HIV-1
mediated stimulation of pDC and its potential effect on modulating regulators of T cell function
including IFNα induced T cell activation. Our results show that chloroquine inhibits pDC
activation/maturation, up-regulation of MyD88 pathway signaling molecules IRF-7 and IRAK-4,
IFN-α production, IDO synthesis, and PDL-1 expression. We also observed that CD8+ T cell
activation induced by IFN-α can be modulated by chloroquine through its ability to decrease
IFNα production by pDC. The central role that TLRs play in innate and adaptive immunity
makes them an ideal target for therapeutic intervention using chloroquine. We propose that in
HIV infection, chronic TLR stimulation and production of IFNα by pDC contributes to immune
activation and immune cell dysfunction, and that blocking or modulation of these pathways with
chloroquine may interfere with the events of HIV pathogenesis.
Materials and Methods
Monoclonal antibodies and reagents
The following fluorochrome conjugated mouse anti-human monoclonal antibodies were used in
these studies: BD Biosciences (BD; San Jose, CA): Lin1-FITC cocktail (CD3, CD14, CD16,
CD20, CD56), CD123 PerCP-Cy5.5, HLA DR APC, biotin-HLA DR, HLA DR FITC, CD86
APC, CD83 PE, CD38 PE, CD3 Pacific Blue, CD8 APC-H7; eBiosciences (San Diego, CA):
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

PDL-1 APC; and CalTag (Carlsbad, CA): CD4 PE-Texas Red. Unconjugated rabbit anti-human
polyclonal antibodies included IRF-7 (H-246; Santa Cruz Biotechnology; Santa Cruz, CA),
IRAK-4 (Invitrogen; Carlsbad, CA) and the anti-human IDO polyclonal rabbit antibody directed
against the DLIESGLRERVEKLNMLC peptide (60) from Dr. David Munn (Medical College of
Georgia; Augusta, GA). Fluorochrome conjugated secondary reagents included streptavidin-
Pacific Blue (Invitrogen) and donkey anti-rabbit PE (Jackson ImmunoResearch Laboratories;
West Grove, PA). The BD Cytofix/Cytoperm™ kit was used for intracellular staining of IRF-7,
IRAK-4, and IDO. Cells were treated with the FcγR blocking reagent (Miltenyi Biotec; Auburn,
CA) and/or normal donkey serum (Jackson ImmunoResearch) to prevent non-specific antibody
binding to the cells.
The synthetic small molecule imidiazoquinoline, 3M-011 (TLR7/8) was provided by 3M
Pharmaceuticals (St. Paul, MN). Endotoxin-free A-class CpG ODN 2336 or 2216 were provided
by Coley Pharmaceutical Group (Pfizer Wellesley, MA). Non-infectious aldithriol-2 (AT-2)
treated HIV-1Ada (R5-tropic) and HIV-1MN (X4-tropic) viral preparations along with matched
control microvesicles, SUPT1-CCR5 and CEMX174 (T1) isolated from uninfected cell cultures
(6) were kindly provided by Dr. Jeff Lifson from the AIDS Vaccine Program, National Cancer
Institute (Frederick, MD). Recombinant IFNα2a (rIFNα2a) was obtained from PBL
(Piscacaway, NJ) and the 64G12 monoclonal antibody against the human interferon-alpha
receptor (anti-IFNαR) was provided by Michael Tovey from the Laboratory of Viral Oncology
(Villejuif, France).
TLR agonists and AT-2 HIV-1 for stimulation of PBMC
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

The TLR agonists were used at the following concentrations; TLR7/8 agonist (3M-011; 3.0 µM)
and the TLR 9 agonist A-class CpG ODN (2336 or 2216; 4 µg/ml) for in vitro stimulation of
PBMC. It is now known that endocytosis of HIV-1 activates plasmacytoid dendritic cells via
Toll-like receptor-viral RNA interactions (5), we stimulated PBMC using non-infectious
aldithriol-2 (AT-2) treated HIV-1Ada (R5-tropic) and HIV-1MN (X4-tropic) as more than 99% of
HIV-1 particles detected in the circulation are non infectious (35, 64). AT-2 HIV-1Ada (AT-2
Ada) and HIV-1MN (AT-2 MN) were used at a concentration of 500 ng/ml of p24 capsid
equivalent. The concentration of HIV-1 (500ng/mL) used in this study is within the range found
in the plasma of infected individuals, children and adults which is known to vary depending on
stage of HIV disease (44, 66). Optimal concentrations of the TLR agonists (32, 54, 55) and AT-
2 HIV-1 (54) were determined previously. Media and matched microvesicle controls, MV-
CCR5 and MV-CEMX served as the negative controls in these experiments.
Chloroquine for inhibition of endosomal acidification and blocking pDC TLR signaling
To study the effect of chloroquine on regulating the endosomal TLR7 and TLR9 pathways in
response to inactivated HIV-1 or TLR agonists we used a concentration of 100 µM chloroquine
based on a previous report evaluating HSV viral activation of pDC (56). For the T cell activation
studies we used a concentration of chloroquine (0.5-5 µM) which is the achievable physiological
concentration in blood as described in the Medsafe datasafe database [www.medsafe.govt.nz]
(13). PBMC were pre-incubated for 1 hour in media containing chloroquine (diphosphate salt)
(InvivoGen, San Diego, CA) which was present throughout the cell culture.
Cell culture for pDC activation ± chloroquine
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Heparinized peripheral blood was collected via venipuncture from healthy HIV-seronegative
volunteers recruited from Rush University Medical Center (Chicago, IL) after informed consent.
Peripheral blood mononuclear cells (PBMC) were isolated by density gradient centrifugation
using lymphocyte separation medium (Cambrex, Gaitherburg, MD). PBMC (1x106/ml) were
cultured in RPMI 1640 (BioWhittaker, Walkersville, MD) supplemented with 10% heat-
inactivated fetal bovine serum (Invitrogen-Gibco), 100 U/ml penicillin, 100 µg/ml streptomycin
(Sigma, St. Louis, MO) and 2 mM L-glutamine (Sigma) in 6-well polystyrene tissue culture
plates in a humidified 370C 5% CO2 incubator. PBMC were pre-incubated with media
containing 100 µM of chloroquine or media alone for 1 hour prior to stimulation with TLR
agonists or AT-2 inactivated HIV-1. Cells were cultured for 20 hours, harvested and evaluated
for pDC activation/maturation and intracellular IDO, IRF-7 and IRAK-4 protein expression
using flow cytometry. Supernatants were stored at -200C for IFNα cytokine analysis by ELISA.
Cell culture for T cell activation ± chloroquine
PBMC or enriched CD8+ T cell subsets (indirect isolation, Miltenyi (Auburn, CA) Microbead
Isolation Kit) were pre-incubated for 1 hour with media containing chloroquine or 30 minutes
with the 64G12 anti-IFNαR blocking antibody (10 µg/ml). AT-2 HIV-1 or 1000 U/ml rIFNα2a
was added and cells were cultured for 20 hours, harvested and evaluated for T cell activation by
measuring CD38 surface expression on CD4+ and CD8
+ cells by flow cytometry. Logical gating
was used to identify the CD4 (CD3+/CD4
+) and CD8 (CD3
+/CD8
+) T cell subsets. T cell
activation is expressed as CD38 mean fluorescence intensity (CD38-MFI) for each parent T cell
subset.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Cell culture for pDC IDO activity ± Chloroquine
PBMC pre-incubated (1 hour) with media containing 100 µM chloroquine or media alone were
cultured with TLR agonists or AT-2 inactivated HIV-1 in 96-well Tissue Culture plates (1x106
cell/well; 0.250 ml/well) in a humidified 370C 5% CO2 incubator. Cells were cultured for 20
hours, harvested and evaluated for pDC PDL-1 expression by flow cytometry. Supernatants
were stored at -200C for evaluation of IDO activity in fluorometric assays to detect tryptophan
and kynurenine in the pathway of L-tryptophan catabolism.
Flow cytometric analysis
Chloroquine treated or untreated PBMC from control or TLR agonist and AT-2 HIV-1
stimulated cultures were harvested, washed and resuspended in PBS containing 0.5% bovine
serum albumin and 0.1% sodium azide (FACS buffer). Non-specific antibody binding to Fc
receptors was blocked by pre-incubation of the cells with the Fcγ-receptor blocking reagent
(Miltenyi Biotec). PBMC for pDC activation/maturation and PDL-1 along with PBMC and
purified CD4+ and CD8
+ T cells for T cell activation were surface stained with the appropriate
antibodies, washed with FACS buffer, fixed with 2% formaldehyde and stored at 40C until
analysis. Cells for intracellular IRF-7, IRAK-4 and IDO staining were surface stained to identify
pDC (Lin1-/CD123
+/HLA DR
+), washed with FACS buffer, blocked with normal donkey serum
(Jackson ImmunoResearch) and processed with BD Cytofix/Cytoperm™ solution. Cells were
then washed in Perm/Wash™ buffer and stained with the appropriate unconjugated polyclonal
rabbit anti-human IRF-7, IRAK-4, or IDO antibodies. Cells were washed in Perm/Wash™
buffer and stained with donkey anti-rabbit PE secondary antibody (Jackson ImmunoResearch).
Finally, the cells were washed twice in Perm/Wash™ buffer, fixed with 1% formaldehyde, and
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

stored at 40C until analysis. All samples were evaluated within 24 hours of staining using a
FACSCalibur™ or LSRII flow cytometer.
Logical gating was used to identify the pDC (Lin1-/CD123
+/HLA DR
+) population. PDC
activation (pDC-CD86) and maturation (pDC-CD83) marker expression are expressed as a
percentage of the pDC parent population. Logical gating was used to identify the CD8
(CD3+/CD8
+) T cell population. Mean fluorescence intensity (MFI) was used to evaluate IRF-7,
IRAK-4, IDO, and PDL-1 levels within the parent pDC population and CD38 level within the
parent T cell subsets.
Measurement of IFNα by ELISA
Commercial ELISA kits were used to measure the concentration of IFNα (PBL Biomedical
Laboratories; Piscataway, NJ) in cell culture supernatants. ELISA assays were performed
according to the manufacturer’s guidelines.
IDO activity: kynurenine and tryptophan fluorescence assays
Depletion of tryptophan and accumulation of the immunosuppressive catabolite kynurenine is a
measure of IDO activity. Detection of tryptophan and kynurenine in the culture supernatants
was performed using fluorometric assays (7, 59). In brief, duplicate eppendorf tubes containing
cell culture supernatants or stock kynurenine solution (Sigma; 25–0.39 µg/ml) were incubated
with 30% trichloroacetic acid (TCA, Sigma) for 30 minutes in 500C water bath. The tubes were
allowed to cool to room temperature and then centrifuged at 2400 rpm for 10 minutes. An
aliquot from each tube was transferred into individual wells of a flat bottom polystyrene 96-well
plate. Freshly made 2% Erlich’s reagent (Sigma) in glacial acetic acid was added to each well
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

and the plate was incubated at room temperature for 10 minutes in the dark. Finally the plate
was then read on a BioTek Synergy HT plate reader (BioTek; Winooski, VT) at 490 nm
wavelength. A kynurenine (0 –25 µg/ml) standard curve was used to obtain the kynurenine
concentration in the culture supernatants.
The tryptophan fluorescence assay was performed as follows: Stock L- tryptophan
(Sigma; 1.25 – 20 µg/ml) or cell culture supernatants were incubated in 20% TCA in a 96 well
plate (duplicate wells/sample) for 1 hour at 40C. The plates were centrifuged at 2400 rpm for 15
minutes and supernatants were transferred into polypropylene tubes. The supernatants were then
incubated with a mixture of 10% TCA, 1.8% in PBS formaldehyde and 6 mM FeCL3 at 900C for
2 hours. The tubes were allowed to cool to room temperature and aliquots from each tube were
transferred into individual wells of a flat bottom polystyrene 96-well plate. The plates were read
on a BioTek Synergy HT fluorescence plate reader (Ex=360/40; Em = 460/40; sensitivity =70).
The tryptophan (1.25–20 µg/ml) standard curve was used to obtain the tryptophan concentration
in the culture supernatants. IDO activity is expressed as the kynurenine: tryptophan ratio.
Statistical analysis
Results are expressed as the mean ± standard error of the mean (SEM). Differences between
groups were performed using an unpaired Student’s t-test (confidence level of 95%) with
GraphPad PRISM software v4.03. p values of <0.05 were considered statistically significant.
Results
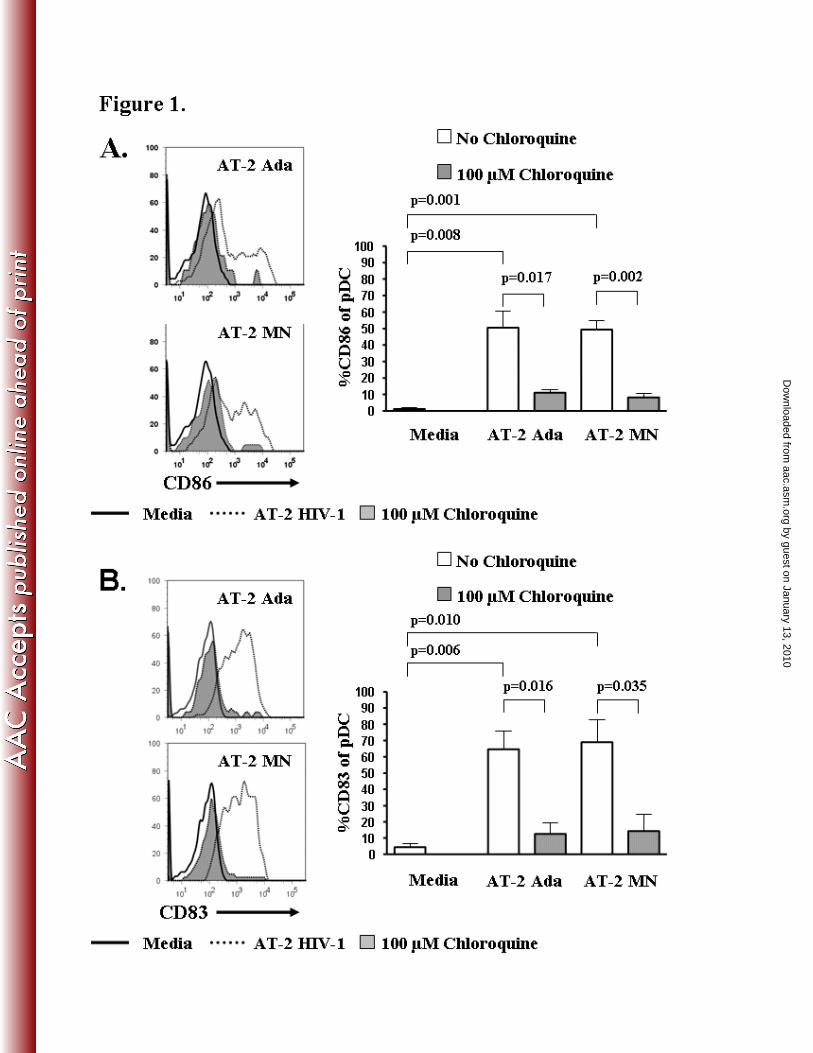
AT-2 HIV-1 induced pDC activation/maturation is down-regulated by chloroquine
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

We investigated the ability of chloroquine, an inhibitor of endosomal acidification, to abrogate
HIV-1 induced pDC activation (pDC-CD86) and maturation (pDC-CD83). PBMC, treated in the
presence or absence of chloroquine (100 µM), were stimulated with non-infectious HIV AT-2-
Ada, AT-2-MN, or control microvesicles overnight. Both CD86 and CD83 were significantly
up-regulated on pDC (Lin1-/CD123
+/HLA DR
+) exposed to AT-2 Ada (CD86 p=0.008; CD83
p=0.006) or AT-2 MN (CD86 p=0.001; CD83 p=0.010) compared to the media control (Fig 1
A,B). AT-2 Ada induced pDC activation markers (% pDC expressing CD86) decreased from
50.5 ± 9.8% to 11.1 ± 19.8% (p=0.017, Fig 1A) and pDC maturation markers (% pDC
expressing CD83) decreased from 64.6 ± 11.1% to 12.6 ± 6.8% (p=0.016, Fig 1B) in the
chloroquine treated cultures. Similarly, AT-2 MN induced pDC activation markers decreased
from 49.3 ± 5.4% to 8.2 ± 2.5% (p=0.002, Fig 1A) and maturation markers decreased from 69.0
± 13.9% to 14.3 ± 10.6% (p=0.035, Fig 1B) in chloroquine treated cultures. Control
microvesicles, CCR5-MV and CEMX-MV, did not induce significant pDC activation/maturation
(CD86: Media 1.23 ± 0.54%, CEMX-MV 1.65 ± 0.38%, CCR5-MV 2.89 ± 1.29% or CD83:
Media 4.42 ± 2.94%, CEMX-MV 5.24 ± 4.31%, CCR5-MV 3.03 ± 1.97%) compared to media
controls.
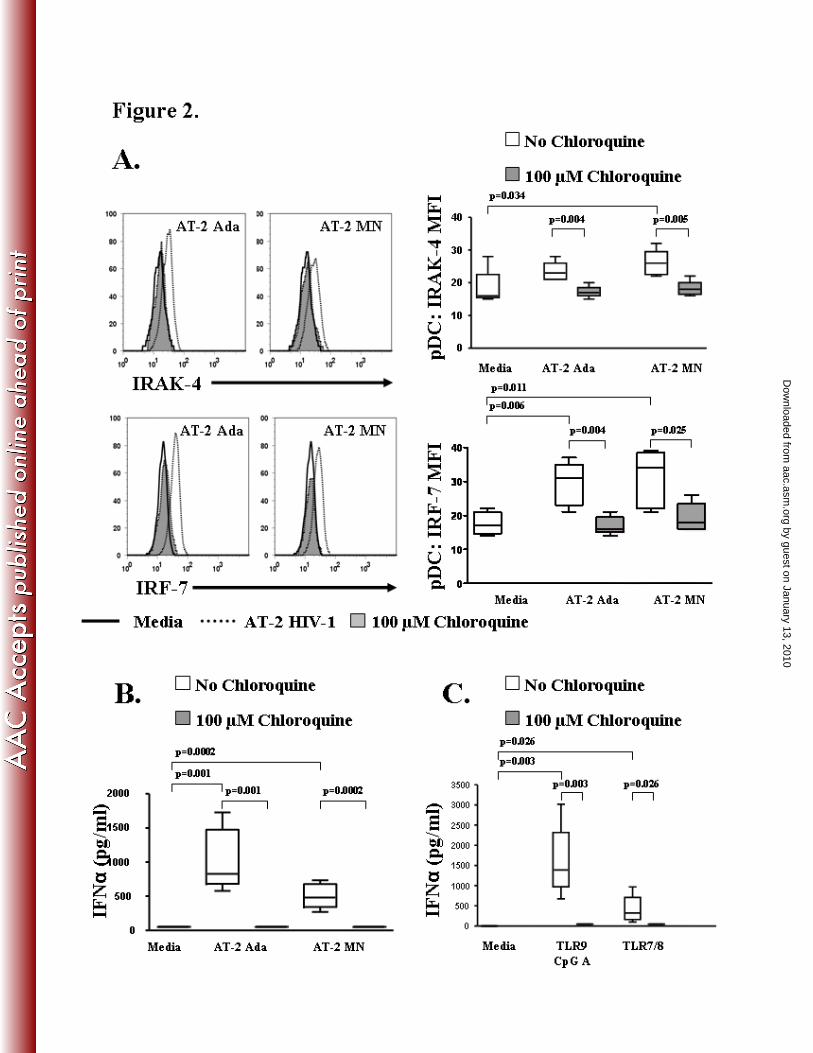
TLR signaling and IFNα production by pDC is inhibited by chloroquine
HIV-1 induces the secretion of type-1 interferons (IFNα/β) by pDC partly through TLR 7 (5).
TLR7 recognition of ssRNA (HIV-1 virus) depends on the kinase activity of interleukin-1
receptor-associated kinase 4 (IRAK-4), the interferon regulatory factor 7 (IRF-7) transcription
factor and the adaptor molecule MyD88 (2, 38, 39, 47). In this study, we tested the effects of
chloroquine on AT-2 HIV-1 induced TLR signaling by measuring downstream transcription
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

molecules IRAK-4 and IRF-7 and subsequent interferon production in PBMC from five donors.
AT-2 MN significantly up-regulated IRAK-4 MFI (p=0.034) while IRF-7 MFI was significantly
increased by both AT-2 MN (p=0.011) and AT-2 Ada (p= 0.006) (Fig 2A). Chloroquine
treatment prevented AT-2 Ada (p=0.004) and AT-2 MN (p=0.005) induced up-regulation of
IRAK-4 in pDC compared with AT-2 HIV-1 stimulated cultures in the absence of chloroquine
(Fig 2A). Further down-stream in the TLR7 induced MyD88 signaling pathway, the
transcription factor, IRF-7, was also decreased in AT-2 Ada (p=0.004) and AT-2 MN (p=0.025)
stimulated cultures treated with chloroquine (Fig 2A) compared to AT-2 HIV-1 cultures in the
absence of chloroquine. Control microvesicles, CCR5-MV and CEMX-MV, did not induce
significant pDC IRAK-4 (Media 18.40 ± 2.42, CEMX-MV 16.25 ± 0.95, CCR5-MV 16.50 ±
1.19) or IRF-7 (Media 17.60 ± 1.50, CEMX-MV 17.80 ± 1.69, CCR5-MV 16.40 ± 0.60)
expression compared to media controls.
Modulation of the TLR7 signaling pathway via chloroquine should be reflected in the
production of IFNα. Therefore, we evaluated AT-2 HIV-1 stimulated culture supernatants for
IFNα using a commercially available ELISA assay. Supernatants from either the AT-2 Ada
(p=0.001) or AT-2 MN (p=0.0002) stimulated cultures contained significantly higher
concentrations of IFNα compared to the media control (Fig 2B). Chloroquine completely
abrogated the production of IFNα in PBMC cultures stimulated with AT-2 Ada (p=0.001) or AT-
2 MN (p= 0.0002) (Fig 2B). To confirm the ability of chloroquine to prevent IFNα secretion via
TLR7 or TLR9 signaling of pDC we used synthetic TLR7/8 (3M-011) and TLR 9 (CpG A)
agonists. Supernatants from the TLR9 CpG A (p=0.003) or TLR7/8 (p=0.026) agonist
stimulated cultures contained significantly higher concentrations of IFNα compared to the media
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

control (Fig 2C). As observed in the AT-2 HIV-1 stimulated cultures, neither TLR9 CpG A
agonist (p=0.003) nor the TLR7/8 agonist (p=0.026) induced IFNα production in the presence of
chloroquine (Fig 2C). Taken together, these results demonstrate that chloroquine inhibits TLR
signaling as evidenced by a decrease in the IRAK-4 and IRF-7 signaling molecules along the
MyD88 specific pathway and the subsequent production of IFNα by preventing endocytosis and
the subsequent endosomal degradation of HIV-1 required for TLR signaling by pDC.
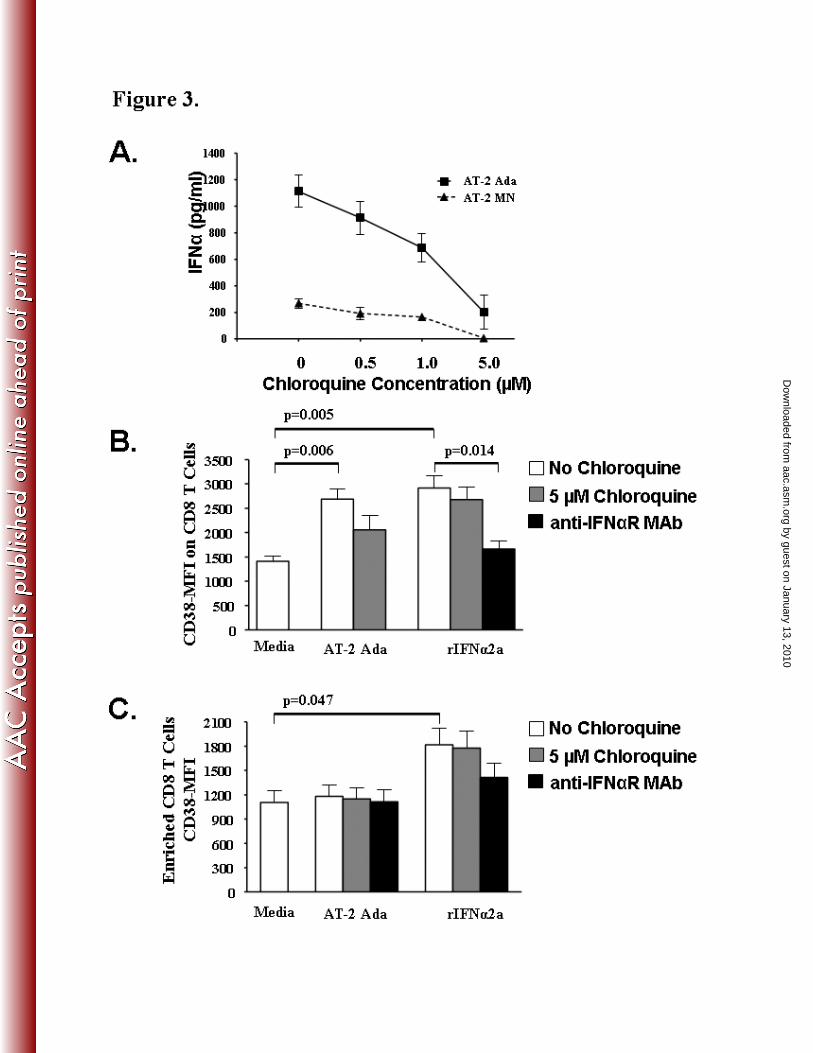
Choroquine down-regulates IFN-α induced T cell activation
T cell activation, a predictor of HIV disease progression, can be measured by the elevated
expression of the CD38 cell surface marker (31, 52, 75) on CD8 T cells. We tested the ability of
physiologic concentrations of chloroquine (0.5–5 µM) to modulate IFNα production and its
subsequent effect on T cell expression of CD38 during exposure to non-infectious AT-2 HIV-1
viruses or exogenous rIFNα2a. Using a commercially available ELISA assay we observed a
dose-dependent decrease in IFNα concentration in the supernatants of AT-2 HIV-1 stimulated
PBMC cultures (n=3) treated with 0.5, 1, or 5 µM concentrations of chloroquine (Fig 3A). At a
concentration of 5 µM Chloroquine, AT-2 Ada (p=0.007) and AT-2 MN (p=0.002) HIV-1
stimulated PBMC supernatants contained significantly reduced levels of IFNα compared to AT-
2-HIV-1 stimulated supernatants without chloroquine. Since IFNα production in response to
AT-2 MN (268.4 ± 35.1 pg/mL) was lower compared to AT-2 Ada (1113 ± 120.6 pg/mL), we
studied the effect of 5 µM chloroquine on AT-2 Ada stimulated PBMC and purified T cells.
Flow cytometric analyses revealed a significant increase in CD38-MFI on CD8+
T- cells (Fig 3B)
after exposure of PBMC to AT-2 Ada (p=0.006), and exogenous rIFNα2a (p=0.005) as
compared to the media control. Chloroquine (5 µM) decreased CD38-MFI on CD8 T cells
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

23.4% in the AT-2 Ada stimulated PBMC (Fig 3B). In contrast, blocking of the IFNα-receptor
with the 64G12 monoclonal antibody significantly down-regulated the CD38-MFI on CD8+
T-
cells (p=0.014; 43% decrease CD38-MFI) in PBMC stimulated with exogenous rIFNα.
Next, we utilized Miltenyi bead enriched CD8+ T cells (n=3) to evaluate the role of IFNα
or HIV-1 to induce CD8+ T cell activation. CD38 expression (CD38-MFI) was significantly
increased on CD8+ cells exposed to exogenous rIFNα2a (p=0.047) but not on CD8
+ cells
exposed to AT-2 Ada HIV-1 as compared to media (Fig 3C). Blocking of the IFNα-receptor by
pre-treatment of enriched CD8+ cells with the 64G12 monoclonal antibody partially decreased
CD38-MFI by 22.3% in the rIFNα2a stimulated cultures (Fig 3C). As expected, chloroquine did
not inhibit rIFNα2a induced CD8+ T cell activation. These data suggest that using chloroquine
to block IFNα produced by pDC during HIV-1 infection may play a critical role in reducing
chronic T cell activation.
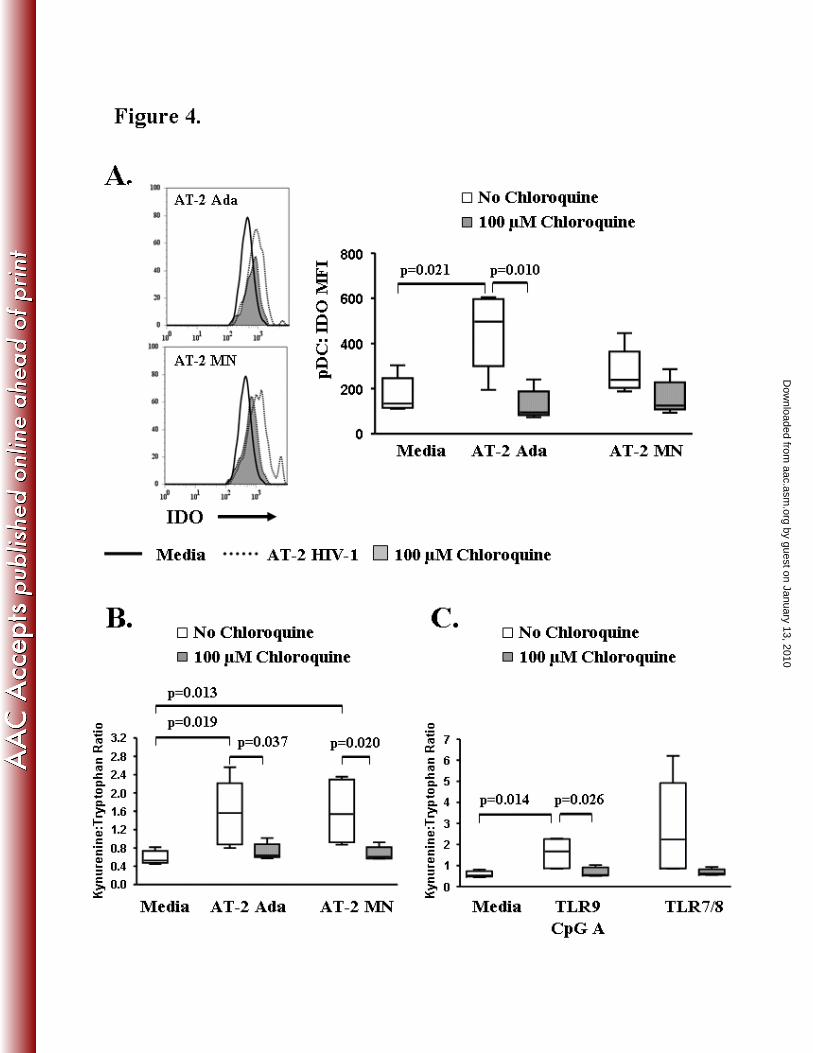
AT-2 HIV-1 induced pDC-IDO expression and IDO tryptophan catabolism is down-regulated by
chloroquine
PDC produce indoleamine 2, 3 dioxygenase (IDO), a key enzyme in tryptophan degradation
along the kynurenine pathway that has been implicated in decreased CD4 T cell proliferative
capacity in HIV-1 infection (10). In this study, PBMC from five individuals were tested for the
effect of chloroquine on AT-2 HIV-1 induced intracellular expression of IDO in the pDC
population. Intracellular levels of IDO in pDC (pDC-IDO mean fluorescence intensity; MFI)
were up-regulated following overnight stimulation with AT-2 Ada or AT-2 MN (Fig 4A,
representative examples shown). PDC-IDO expression was significantly up-regulated in the
cultures stimulated with AT-2 Ada (MFI = 448.0 ± 96.0; p=0.021) compared to the media (MFI
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

= 170.8 ± 35.8) (Fig 4A). Chloroquine treatment abrogated AT-2 HIV-1induced pDC-IDO
expression to baseline MFI values similar to the media control cultures (Ada+CHL MFI = 126.4
± 30.1, p=0.010; MN+CHL MFI = 159.0 ± 34.0; Media MFI = 170.8 ± 35.8) (Fig 4A). These
results demonstrate that chloroquine can suppress TLR mediated IDO expression by pDC which
may prove beneficial in blocking tryptophan degradation.
Next, we evaluated IDO functional activity by measuring kynurenine (KYN) and
tryptophan (TRYP) levels in cell culture supernatants. Results are expressed as the KYN:TRYP
ratio, whereby, an increase in the KYN:TRYP ratio indicates active IDO catabolism of
tryptophan along the kynurenine pathway. The KYN:TRYP ratios (Fig 4B) were significantly
increased in the supernatants of cultures stimulated with AT-2 Ada (1.548 ± 0.320; p=0.019) or
AT-2 MN (1.594 ± 0.308; p=0.013) as compared to the media control (0.589 ± 0.066).
Chloroquine treatment resulted in decreased tryptophan catabolism in the AT-2 Ada (Ada vs.
Ada-CHL p=0.037) and AT-2 MN (MN vs. MN-CHL p=0.020) stimulated culture supernatants.
We also evaluated IDO activity in cultures stimulated with synthetic TLR9 (CpG A) and
TLR7/8 agonists (Fig 4C). The KYN:TRYP ratio was significantly increased in the supernatants
of cultures stimulated with TLR9 (1.590 ± 0.314; p=0.014) as compared to the media control
(0.589 ± 0.066). KYN:TRYP ratios were significantly lower in TLR9 CpG A (p=0.026)
stimulated cultures containing chloroquine compared to cultures without chloroquine. The
KYN:TRYP ratio was elevated in the supernatants stimulated with TLR7/8 agonist compared to
media and were reduced in chloroquine treated cultures though no statistical differences were
observed. These results demonstrate that increased IDO expression in pDC can be directly
mediated by TLR7 and/or TLR9 stimulation and the abrogation of the TLR signal with
chloroquine may offer a means to regulate pDC-IDO expression in HIV-1 infection.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

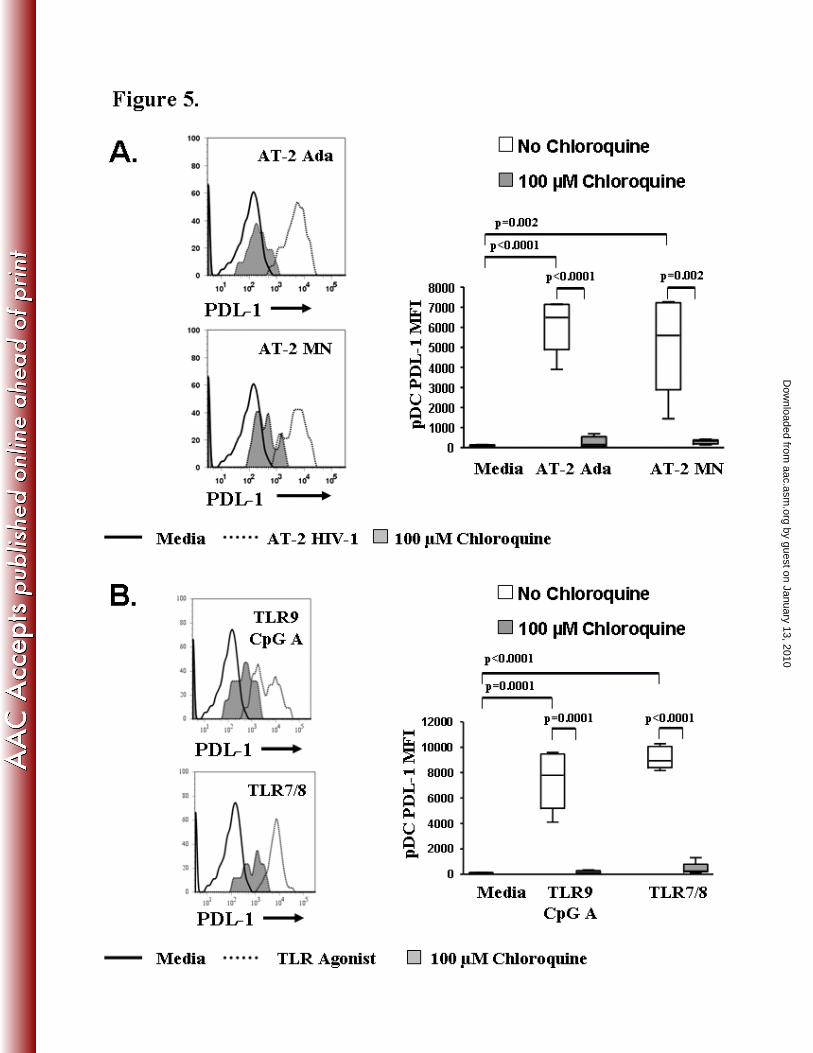
AT-2 HIV-1 and TLR agonist induced PDL-1 expression on pDC is down-regulated by
chloroquine
Microbial persistence may be related to the exploitation of the PD-1 and PDL-1 pathway by
certain pathogens (72). PDL-1 is expressed constitutively on pDC. AT-2 Ada (p<0.0001), AT-2
MN (p=0.002), TLR9 CpG A (p=0.0001), and TLR7/8 (p<0.0001) significantly increased the
PDL-1 expression on pDC compared to the media control (Fig 5A,B). We tested the ability of
chloroquine to regulate AT-2 HIV-1 or synthetic TLR7/8 and TLR9 agonist induced PDL-1
expression on pDC. PDL-1 mean fluorescence intensity on pDC (pDC-PDL-1 MFI) was
significantly reduced in the presence of chloroquine (Fig 5A,B). Chloroquine completely
abrogated the up-regulation of PDL-1 on pDC in cultures stimulated with either AT-2 HIV-1 or
TLR agonists [AT-2 Ada (p<0.0001), AT-2 MN (p=0.002), TLR9 CpG A (p=0.0001), TLR7/8
(p<0.0001)]. These results demonstrate that chloroquine down-modulates the potential
contribution of pDC to T cell exhaustion/dysfunction through the PD-1/PDL-1 pathway by
suppressing PDL-1 expression on pDC.
Discussion
HIV infection is associated with the impairment of immune function with deficiencies observed
in nearly every type of immune cell. A better understanding of the process by which HIV
hijacks innate immune cell function could be explored for alternative therapeutic approaches.
Among potential mechanisms, chronic activation of pDC by direct interaction with infectious
and non-infectious HIV-1 particles contribute to the maintenance of immune cell dysfunction
and HIV-associated pathogenesis (8, 11, 21, 36). In this study, we used an in vitro model to
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

demonstrate the potential immunoregulatory function of chloroquine to modulate HIV-1 induced
pDC activation/maturation, IFNα production, IDO expression/activity, and PDL-1 expression.
We also show that IFNα, an outcome of pDC activation, directly contributes to CD8+ T cell
activation. Using chloroquine, an inhibitor of endosomal fusion and acidification, we were able
to suppress HIV-1 mediated TLR signaling in pDC and inhibit the potential deleterious effects of
chronic pDC activation. Our data shows that interfering with TLR signaling reduces HIV-1
induced pDC activation products that are known to contribute to immune dysregulation in HIV-1
infection.
More than 99% of HIV-1 particles detected in the circulation are not productively
infectious (35, 64). These non-infectious particles contribute to HIV-induced
immunopathogenesis (21, 36). We and others have shown that AT-2 HIV-1 at concentrations
between 300-1000ng/mL induces pDC activation/maturation and production of IFNα (27, 35, 36,
54, 82). The concentration of HIV (500ng/mL) used in this study is within the range found in the
plasma of infected individuals, children and adults which is known to vary depending on stage of
HIV disease (44, 66). Using 500ng/mL of noninfectious AT-2 inactivated HIV-1 Ada and MN
virus we could activate pDC IFNα pathway. Activation of pDC by HIV-1 requires endocytosis
of the virion leading to endosomal TLR7 recognition of viral RNA (5, 58) and initiation of the
MyD88 pathway for the production of IFNα by pDC. Chloroquine, an inhibitor of endocytosis
and endosomal acidification/maturation, prevented HIV-1 mediated TLR7 signaling in pDC as
shown by the failure of pDC to up-regulate the down-stream IRAK-4 and IRF-7 signaling
molecules within the MyD88 pathway. By showing that chloroquine decreases IFNα production
it can be inferred that AT-2 HIV-1 utilizes TLR7 signaling in pDC and not the cytosolic anti-
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

viral pathway mediated by RIG-1 recognition of ssRNA (41, 65). These data highlight the
importance of the endolysosomal pathway in HIV-1 mediated signaling of pDC.
The production of IFNα by pDC in HIV-1 infection may also have important
consequences for T cell activation and survival (67). During acute infection HIV-1 high plasma
titers of IFNα provide a protective antiviral effect and help trigger the adaptive immune response
(73). However, in late-stage disease elevated levels of serum IFNα is an indicator of poor
clinical prognosis (30, 80). We demonstrate that 100µM chloroquine has an inhibitory effect on
HIV-1 mediated production of IFNα. It has been shown that 100µM chloroquine used in
treatment of leucocytes in vitro results in intracellular concentrations comparable to those
obtained in vivo during chloroquine therapy (29). Our findings are similar to those of other
investigators using chloroquine to modulate IFNα production in HSV infected systems (25, 50).
However, in our study at the declining concentrations of chloroquine (0.5-5 µM) we observed a
dose-dependent decrease in HIV-1 induced IFNα. At our lowest tested concentration of
chloroquine( 0.5 µM), we observed a partial blockage of IFNα which is in contrast to a published
report (71) of complete blockage of IFNα with similar concentration of chloroquine (0.5 µM)
used to induce pDC activation through TLR9. These differences in concentration of chloroquine
required for blocking IFNα could be attributed to their activation pathways. As previously
reported, a higher concentration of chloroquine is necessary to block TLR7 endosomal
maturation and acidification compared to TLR9 (51). Therefore, our results provide evidence
that HIV-1 induced IFNα can be modulated with chloroquine at concentrations that are
obtainable for therapeutic application.
Chronic immune activation is a hallmark of HIV-1 pathogenesis. Previous reports of
elevated IFNα levels in the plasma and lymphoid tissues along with increased expression of
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

interferon-inducible genes in HIV-infected patients has been shown to correlate with elevated T
cell activation characteristic of HIV progression (14, 36, 37). In this study, CD38 activation
marker, commonly used to measure the T cell activation state in HIV-infected patients (8, 20, 31,
75), was down-modulated in PBMC cultures when utilizing 5µM chloroquine . As cell separation
procedures affect the uptake of chloroquine (29) we used lower dose of chloroquine (5µM) to
study its effect on T cell activation in enriched T cells and compared with a similar dose of
chloroquine in PBMC cultures. Chloroquine downmodulates T cell activation in AT-2 HIV-1
stimulated PBMC cultures and not in AT-2 HIV-1 stimulated enriched T cells suggesting that
inhibition of pDC IFNα decreases T cell activation. Using enriched CD8+ cells we demonstrate
that increased CD38 expression is induced by exogenous rIFNα2a and not by direct interaction
with HIV-1 suggesting that pDC IFNα certainly contributes to CD8+ T cell activation. As
previously reported(67), we did not find IFNα induced activation of CD4+ T cell (data not
shown). The ability to block IFNα mediated CD8+ activation with the anti-IFNαR monoclonal
antibody (64G12) and not chloroquine highlights the importance of modulating IFNα induced
CD8+ T cell activation. This is the first direct evidence of chloroquine blocking pDC IFN-α
mediated activation and is supported by a previous study demonstrating the ability of
chloroquine to control TLR9 (CpG) or LPS induced pro-inflammatory cytokine production in
mice (40).
HIV-1 also modulates immune function through the induction of negative regulatory
signals which directly affect the development of the adaptive immune response. One such
mechanism evaluated in our studies was the induction of the immunosuppressive enzyme, IDO
in pDC following HIV-1 stimulation. We also observed an increase in kynurenine metabolites in
the culture supernatants indicative of IDO regulated tryptophan catabolism. As shown by other
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

in vitro studies HIV-1 induces IDO in pDC (10, 22, 57) which directly inhibits T cell
proliferation and function through tryptophan depletion and through the activity of kynurenine
metabolites (4, 12, 61). Another key regulatory function of IDO is the induction and stimulation
of T regulatory cells during HIV infection (18, 23, 33) which dampen HIV specific T cell
responses. Thus, development of new treatment modalities (i.e. chloroquine or
hydroxychloroquine) to reverse the induction of IDO levels found in both blood cells and
lymphoid tissues (4, 10, 61) may prove beneficial for the improvement of T cell proliferation and
function in addition to controlling aberrant immune activation in HIV infected individuals.
A second mechanism induced by HIV-1 infection to dampen anti-viral responses is the
PD-1/PDL-1 pathway (9, 19, 63, 79). Our data suggests that “non-infectious” HIV-1 (AT-2
HIV-1) exposure up-regulates PDL-1 ligand expression on pDC and is consistent with reports of
other investigators showing TLR induced PDL-1 on pDC and IFNα induced PDL-1 on mDC (1,
15, 78). Manipulation of the PD-1/PDL-1 pathway in HIV-1 infection may prove beneficial in
the restoration of virus specific CD8+ T cell responses by decreasing programmed death pathway
induced apoptosis.
In addition to the immunomodulatory effects observed in these studies chloroquine has
been shown in vitro to have direct inhibitory activity on newly produced HIV virions by altering
the glycosylation of the 2G12 epitope which is located on the gp120 envelope surface protein
required for virus infectivity (69, 70). Several clinical trials have been performed where
chloroquine or hydroxychloroquine was given to HIV infected individuals along with anti-
retroviral therapy. In one study a decrease in HIV viral load measurements was reported (77).
Whilst in another a decrease in plasma p24 capsid antigen levels was observed in chloroquine
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

treated individuals while no alterations in CD4+ T lymphocyte counts were identified compared
to control group (76).
HIV-1 infection hijacks the innate immune response which directly contributes to disease
pathogenesis. Chronic stimulation of pDC with non-infectious and infectious virions leads to
their enhanced activation, state, production of the cytokine IFNα and an up-regulation of IDO
and PDL-1, all of which are known to directly contribute to decreased T cell survival,
proliferation, and function. In this study, we have shown that chloroquine blocks TLR signaling
in pDC, a critical step in its activation pathway. By blocking pDC activation we observed a
decrease in the immunoregulatory cytokine IFNα which in turn could reduce IFNα mediated
immune activation and improve T cell survival by blocking negative modulators like IDO and
PDL-1. Chloroquine is widely accessible, inexpensive and is well tolerated when administered
over periods of several years making it a good candidate for adjuvant therapy along with
HAART to control immune activation in HIV-1 infection. The relevance of the findings
presented in this study are particularly important as it is now known that non AIDS defining
illnesses such as atherosclerosis, liver disease, renal diseases that occur despite effective HAART
contribute to mortality. Immune activation and inflammation are the key contributing factors
linked to these co-morbidities (49). The use of chloroquine as an adjuvant with HAART could
be an effective and inexpensive approach to control immune activation and reduce the risk of co-
morbidities in HAART treated HIV infected individuals. The use of chloroquine in HIV-1
infected individuals in resource rich and resource poor countries need to be further investigated.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

References
1. Abe, M., Z. Wang, A. de Creus, and A. W. Thomson. 2005. Plasmacytoid dendritic
cell precursors induce allogeneic T-cell hyporesponsiveness and prolong heart graft
survival. Am J Transplant 5:1808-19.
2. Akira, S., K. Takeda, and T. Kaisho. 2001. Toll-like receptors: critical proteins linking
innate and acquired immunity. Nat Immunol 2:675-80.
3. Almeida, M., M. Cordero, J. Almeida, and A. Orfao. 2005. Different subsets of
peripheral blood dendritic cells show distinct phenotypic and functional abnormalities in
HIV-1 infection. Aids 19:261-71.
4. Andersson, J., A. Boasso, J. Nilsson, R. Zhang, N. J. Shire, S. Lindback, G. M.
Shearer, and C. A. Chougnet. 2005. The prevalence of regulatory T cells in lymphoid
tissue is correlated with viral load in HIV-infected patients. J Immunol 174:3143-7.
5. Beignon, A. S., K. McKenna, M. Skoberne, O. Manches, I. DaSilva, D. G. Kavanagh,
M. Larsson, R. J. Gorelick, J. D. Lifson, and N. Bhardwaj. 2005. Endocytosis of HIV-
1 activates plasmacytoid dendritic cells via Toll-like receptor-viral RNA interactions. J
Clin Invest 115:3265-75.
6. Bess, J. W., Jr., R. J. Gorelick, W. J. Bosche, L. E. Henderson, and L. O. Arthur.
1997. Microvesicles are a source of contaminating cellular proteins found in purified
HIV-1 preparations. Virology 230:134-44.
7. Bloxam, D. L., and W. H. Warren. 1974. Error in the determination of tryptophan by
the method of Denkla and Dewey. A revised procedure. Anal Biochem 60:621-5.
8. Boasso, A., A. W. Hardy, S. A. Anderson, M. J. Dolan, and G. M. Shearer. 2008.
HIV-induced type I interferon and tryptophan catabolism drive T cell dysfunction despite
phenotypic activation. PLoS ONE 3:e2961.
9. Boasso, A., A. W. Hardy, A. L. Landay, J. L. Martinson, S. A. Anderson, M. J.
Dolan, M. Clerici, and G. M. Shearer. 2008. PDL-1 upregulation on monocytes and T
cells by HIV via type I interferon: restricted expression of type I interferon receptor by
CCR5-expressing leukocytes. Clin Immunol 129:132-44.
10. Boasso, A., J. P. Herbeuval, A. W. Hardy, S. A. Anderson, M. J. Dolan, D. Fuchs,
and G. M. Shearer. 2007. HIV inhibits CD4+ T-cell proliferation by inducing
indoleamine 2,3-dioxygenase in plasmacytoid dendritic cells. Blood 109:3351-9.
11. Boasso, A., and G. M. Shearer. 2008. Chronic innate immune activation as a cause of
HIV-1 immunopathogenesis. Clin Immunol 126:235-42.
12. Boasso, A., M. Vaccari, J. Nilsson, G. M. Shearer, J. Andersson, V. Cecchinato, C.
Chougnet, and G. Franchini. 2006. Do regulatory T-cells play a role in AIDS
pathogenesis? AIDS Rev 8:141-7.
13. Boelaert, J. R., J. Piette, and K. Sperber. 2001. The potential place of chloroquine in
the treatment of HIV-1-infected patients. J Clin Virol 20:137-40.
14. Brenchley, J. M., D. A. Price, T. W. Schacker, T. E. Asher, G. Silvestri, S. Rao, Z.
Kazzaz, E. Bornstein, O. Lambotte, D. Altmann, B. R. Blazar, B. Rodriguez, L.
Teixeira-Johnson, A. Landay, J. N. Martin, F. M. Hecht, L. J. Picker, M. M.
Lederman, S. G. Deeks, and D. C. Douek. 2006. Microbial translocation is a cause of
systemic immune activation in chronic HIV infection. Nat Med 12:1365-71.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

15. Chen, L., Z. Zhang, W. Chen, Z. Zhang, Y. Li, M. Shi, J. Zhang, L. Chen, S. Wang,
and F. S. Wang. 2007. B7-H1 up-regulation on myeloid dendritic cells significantly
suppresses T cell immune function in patients with chronic hepatitis B. J Immunol
178:6634-41.
16. Chen, W., X. Liang, A. J. Peterson, D. H. Munn, and B. R. Blazar. 2008. The
indoleamine 2,3-dioxygenase pathway is essential for human plasmacytoid dendritic cell-
induced adaptive T regulatory cell generation. J Immunol 181:5396-404.
17. Colonna, M., G. Trinchieri, and Y. J. Liu. 2004. Plasmacytoid dendritic cells in
immunity. Nat Immunol 5:1219-26.
18. Curti, A., S. Pandolfi, B. Valzasina, M. Aluigi, A. Isidori, E. Ferri, V. Salvestrini, G.
Bonanno, S. Rutella, I. Durelli, A. L. Horenstein, F. Fiore, M. Massaia, M. P.
Colombo, M. Baccarani, and R. M. Lemoli. 2007. Modulation of tryptophan
catabolism by human leukemic cells results in the conversion of CD25- into CD25+ T
regulatory cells. Blood 109:2871-7.
19. Day, C. L., D. E. Kaufmann, P. Kiepiela, J. A. Brown, E. S. Moodley, S. Reddy, E.
W. Mackey, J. D. Miller, A. J. Leslie, C. DePierres, Z. Mncube, J. Duraiswamy, B.
Zhu, Q. Eichbaum, M. Altfeld, E. J. Wherry, H. M. Coovadia, P. J. Goulder, P.
Klenerman, R. Ahmed, G. J. Freeman, and B. D. Walker. 2006. PD-1 expression on
HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature
443:350-4.
20. Dyrhol-Riise, A. M., P. Voltersvik, J. Olofsson, and B. Asjo. 1999. Activation of CD8
T cells normalizes and correlates with the level of infectious provirus in tonsils during
highly active antiretroviral therapy in early HIV-1 infection. Aids 13:2365-76.
21. Esser, M. T., J. W. Bess, Jr., K. Suryanarayana, E. Chertova, D. Marti, M.
Carrington, L. O. Arthur, and J. D. Lifson. 2001. Partial activation and induction of
apoptosis in CD4(+) and CD8(+) T lymphocytes by conformationally authentic
noninfectious human immunodeficiency virus type 1. J Virol 75:1152-64.
22. Fallarino, F., S. Gizzi, P. Mosci, U. Grohmann, and P. Puccetti. 2007. Tryptophan
catabolism in IDO+ plasmacytoid dendritic cells. Curr Drug Metab 8:209-16.
23. Fallarino, F., U. Grohmann, K. W. Hwang, C. Orabona, C. Vacca, R. Bianchi, M. L.
Belladonna, M. C. Fioretti, M. L. Alegre, and P. Puccetti. 2003. Modulation of
tryptophan catabolism by regulatory T cells. Nat Immunol 4:1206-12.
24. Feldman, S., D. Stein, S. Amrute, T. Denny, Z. Garcia, P. Kloser, Y. Sun, N.
Megjugorac, and P. Fitzgerald-Bocarsly. 2001. Decreased interferon-alpha production
in HIV-infected patients correlates with numerical and functional deficiencies in
circulating type 2 dendritic cell precursors. Clin Immunol 101:201-10.
25. Feldman, S. B., M. Ferraro, H. M. Zheng, N. Patel, S. Gould-Fogerite, and P.
Fitzgerald-Bocarsly. 1994. Viral induction of low frequency interferon-alpha producing
cells. Virology 204:1-7.
26. Finke, J. S., M. Shodell, K. Shah, F. P. Siegal, and R. M. Steinman. 2004. Dendritic
cell numbers in the blood of HIV-1 infected patients before and after changes in
antiretroviral therapy. J Clin Immunol 24:647-52.
27. Fonteneau, J. F., M. Larsson, A. S. Beignon, K. McKenna, I. Dasilva, A. Amara, Y.
J. Liu, J. D. Lifson, D. R. Littman, and N. Bhardwaj. 2004. Human immunodeficiency
virus type 1 activates plasmacytoid dendritic cells and concomitantly induces the
bystander maturation of myeloid dendritic cells. J Virol 78:5223-32.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

28. Fox, R. I. 1993. Mechanism of action of hydroxychloroquine as an antirheumatic drug.
Semin Arthritis Rheum 23:82-91.
29. French, J. K., N. P. Hurst, M. L. O'Donnell, and W. H. Betts. 1987. Uptake of
chloroquine and hydroxychloroquine by human blood leucocytes in vitro: relation to
cellular concentrations during antirheumatic therapy. Ann Rheum Dis 46:42-5.
30. Fuchs, D., G. M. Shearer, R. N. Boswell, D. R. Lucey, M. Clerici, G. Reibnegger, E.
R. Werner, R. A. Zajac, and H. Wachter. 1991. Negative correlation between blood
cell counts and serum neopterin concentration in patients with HIV-1 infection. Aids
5:209-12.
31. Giorgi, J. V., L. E. Hultin, J. A. McKeating, T. D. Johnson, B. Owens, L. P.
Jacobson, R. Shih, J. Lewis, D. J. Wiley, J. P. Phair, S. M. Wolinsky, and R. Detels. 1999. Shorter survival in advanced human immunodeficiency virus type 1 infection is
more closely associated with T lymphocyte activation than with plasma virus burden or
virus chemokine coreceptor usage. J Infect Dis 179:859-70.
32. Gorden, K. B., K. S. Gorski, S. J. Gibson, R. M. Kedl, W. C. Kieper, X. Qiu, M. A.
Tomai, S. S. Alkan, and J. P. Vasilakos. 2005. Synthetic TLR agonists reveal
functional differences between human TLR7 and TLR8. J Immunol 174:1259-68.
33. Grohmann, U., C. Orabona, F. Fallarino, C. Vacca, F. Calcinaro, A. Falorni, P.
Candeloro, M. L. Belladonna, R. Bianchi, M. C. Fioretti, and P. Puccetti. 2002.
CTLA-4-Ig regulates tryptophan catabolism in vivo. Nat Immunol 3:1097-101.
34. Grouard, G., M. C. Rissoan, L. Filgueira, I. Durand, J. Banchereau, and Y. J. Liu.
1997. The enigmatic plasmacytoid T cells develop into dendritic cells with interleukin
(IL)-3 and CD40-ligand. J Exp Med 185:1101-11.
35. Hardy, A. W., D. R. Graham, G. M. Shearer, and J. P. Herbeuval. 2007. HIV turns
plasmacytoid dendritic cells (pDC) into TRAIL-expressing killer pDC and down-
regulates HIV coreceptors by Toll-like receptor 7-induced IFN-alpha. Proc Natl Acad Sci
U S A 104:17453-8.
36. Herbeuval, J. P., J. C. Grivel, A. Boasso, A. W. Hardy, C. Chougnet, M. J. Dolan, H.
Yagita, J. D. Lifson, and G. M. Shearer. 2005. CD4+ T-cell death induced by
infectious and noninfectious HIV-1: role of type 1 interferon-dependent, TRAIL/DR5-
mediated apoptosis. Blood 106:3524-31.
37. Herbeuval, J. P., J. Nilsson, A. Boasso, A. W. Hardy, M. J. Kruhlak, S. A. Anderson,
M. J. Dolan, M. Dy, J. Andersson, and G. M. Shearer. 2006. Differential expression of
IFN-alpha and TRAIL/DR5 in lymphoid tissue of progressor versus nonprogressor HIV-
1-infected patients. Proc Natl Acad Sci U S A 103:7000-5.
38. Honda, K., Y. Ohba, H. Yanai, H. Negishi, T. Mizutani, A. Takaoka, C. Taya, and T.
Taniguchi. 2005. Spatiotemporal regulation of MyD88-IRF-7 signalling for robust type-I
interferon induction. Nature 434:1035-40.
39. Honda, K., H. Yanai, H. Negishi, M. Asagiri, M. Sato, T. Mizutani, N. Shimada, Y.
Ohba, A. Takaoka, N. Yoshida, and T. Taniguchi. 2005. IRF-7 is the master regulator
of type-I interferon-dependent immune responses. Nature 434:772-7.
40. Hong, Z., Z. Jiang, W. Liangxi, D. Guofu, L. Ping, L. Yongling, P. Wendong, and W.
Minghai. 2004. Chloroquine protects mice from challenge with CpG ODN and LPS by
decreasing proinflammatory cytokine release. Int Immunopharmacol 4:223-34.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

41. Hornung, V., J. Ellegast, S. Kim, K. Brzozka, A. Jung, H. Kato, H. Poeck, S. Akira,
K. K. Conzelmann, M. Schlee, S. Endres, and G. Hartmann. 2006. 5'-Triphosphate
RNA is the ligand for RIG-I. Science 314:994-7.
42. Hornung, V., S. Rothenfusser, S. Britsch, A. Krug, B. Jahrsdorfer, T. Giese, S.
Endres, and G. Hartmann. 2002. Quantitative expression of toll-like receptor 1-10
mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to
CpG oligodeoxynucleotides. J Immunol 168:4531-7.
43. Ito, T., R. Amakawa, T. Kaisho, H. Hemmi, K. Tajima, K. Uehira, Y. Ozaki, H.
Tomizawa, S. Akira, and S. Fukuhara. 2002. Interferon-alpha and interleukin-12 are
induced differentially by Toll-like receptor 7 ligands in human blood dendritic cell
subsets. J Exp Med 195:1507-12.
44. Jackson, G. G., D. A. Paul, L. A. Falk, M. Rubenis, J. C. Despotes, D. Mack, M.
Knigge, and E. E. Emeson. 1988. Human immunodeficiency virus (HIV) antigenemia
(p24) in the acquired immunodeficiency syndrome (AIDS) and the effect of treatment
with zidovudine (AZT). Ann Intern Med 108:175-80.
45. Janeway, C. A., Jr., and R. Medzhitov. 2002. Innate immune recognition. Annu Rev
Immunol 20:197-216.
46. Kadowaki, N., S. Antonenko, J. Y. Lau, and Y. J. Liu. 2000. Natural interferon
alpha/beta-producing cells link innate and adaptive immunity. J Exp Med 192:219-26.
47. Kawagoe, T., S. Sato, A. Jung, M. Yamamoto, K. Matsui, H. Kato, S. Uematsu, O.
Takeuchi, and S. Akira. 2007. Essential role of IRAK-4 protein and its kinase activity in
Toll-like receptor-mediated immune responses but not in TCR signaling. J Exp Med
204:1013-24.
48. Kawai, T., S. Sato, K. J. Ishii, C. Coban, H. Hemmi, M. Yamamoto, K. Terai, M.
Matsuda, J. Inoue, S. Uematsu, O. Takeuchi, and S. Akira. 2004. Interferon-alpha
induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88
and TRAF6. Nat Immunol 5:1061-8.
49. Kuller, L. H., R. Tracy, W. Belloso, S. De Wit, F. Drummond, H. C. Lane, B.
Ledergerber, J. Lundgren, J. Neuhaus, D. Nixon, N. I. Paton, and J. D. Neaton. 2008.
Inflammatory and coagulation biomarkers and mortality in patients with HIV infection.
PLoS Med 5:e203.
50. Lebon, P. 1985. Inhibition of herpes simplex virus type 1-induced interferon synthesis by
monoclonal antibodies against viral glycoprotein D and by lysosomotropic drugs. J Gen
Virol 66 ( Pt 12):2781-6.
51. Lee, J., T. H. Chuang, V. Redecke, L. She, P. M. Pitha, D. A. Carson, E. Raz, and H.
B. Cottam. 2003. Molecular basis for the immunostimulatory activity of guanine
nucleoside analogs: activation of Toll-like receptor 7. Proc Natl Acad Sci U S A
100:6646-51.
52. Liu, Z., W. G. Cumberland, L. E. Hultin, H. E. Prince, R. Detels, and J. V. Giorgi.
1997. Elevated CD38 antigen expression on CD8+ T cells is a stronger marker for the
risk of chronic HIV disease progression to AIDS and death in the Multicenter AIDS
Cohort Study than CD4+ cell count, soluble immune activation markers, or combinations
of HLA-DR and CD38 expression. J Acquir Immune Defic Syndr Hum Retrovirol 16:83-
92.
53. Malleret, B., B. Maneglier, I. Karlsson, P. Lebon, M. Nascimbeni, L. Perie, P.
Brochard, B. Delache, J. Calvo, T. Andrieu, O. Spreux-Varoquaux, A. Hosmalin, R.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Le Grand, and B. Vaslin. 2008. Primary infection with simian immunodeficiency virus:
plasmacytoid dendritic cell homing to lymph nodes, type I interferon, and immune
suppression. Blood 112:4598-608.
54. Martinson, J. A., A. Roman-Gonzalez, A. R. Tenorio, C. J. Montoya, C. N. Gichinga,
M. T. Rugeles, M. Tomai, A. M. Krieg, S. Ghanekar, L. L. Baum, and A. L. Landay. 2007. Dendritic cells from HIV-1 infected individuals are less responsive to toll-like
receptor (TLR) ligands. Cell Immunol 250:75-84.
55. Martinson, J. A., A. R. Tenorio, C. J. Montoya, L. Al-Harthi, C. N. Gichinga, A. M.
Krieg, L. L. Baum, and A. L. Landay. 2007. Impact of class A, B and C CpG-
oligodeoxynucleotides on in vitro activation of innate immune cells in human
immunodeficiency virus-1 infected individuals. Immunology 120:526-35.
56. Megjugorac, N. J., H. A. Young, S. B. Amrute, S. L. Olshalsky, and P. Fitzgerald-
Bocarsly. 2004. Virally stimulated plasmacytoid dendritic cells produce chemokines and
induce migration of T and NK cells. J Leukoc Biol 75:504-14.
57. Mellor, A. L., and D. H. Munn. 2004. IDO expression by dendritic cells: tolerance and
tryptophan catabolism. Nat Rev Immunol 4:762-74.
58. Miyauchi, K., Y. Kim, O. Latinovic, V. Morozov, and G. B. Melikyan. 2009. HIV
enters cells via endocytosis and dynamin-dependent fusion with endosomes. Cell
137:433-44.
59. Muller, A. J., J. B. DuHadaway, P. S. Donover, E. Sutanto-Ward, and G. C.
Prendergast. 2005. Inhibition of indoleamine 2,3-dioxygenase, an immunoregulatory
target of the cancer suppression gene Bin1, potentiates cancer chemotherapy. Nat Med
11:312-9.
60. Munn, D. H., M. D. Sharma, J. R. Lee, K. G. Jhaver, T. S. Johnson, D. B. Keskin, B.
Marshall, P. Chandler, S. J. Antonia, R. Burgess, C. L. Slingluff, Jr., and A. L.
Mellor. 2002. Potential regulatory function of human dendritic cells expressing
indoleamine 2,3-dioxygenase. Science 297:1867-70.
61. Nilsson, J., A. Boasso, P. A. Velilla, R. Zhang, M. Vaccari, G. Franchini, G. M.
Shearer, J. Andersson, and C. Chougnet. 2006. HIV-1-driven regulatory T-cell
accumulation in lymphoid tissues is associated with disease progression in HIV/AIDS.
Blood 108:3808-17.
62. Ochando, J. C., C. Homma, Y. Yang, A. Hidalgo, A. Garin, F. Tacke, V. Angeli, Y.
Li, P. Boros, Y. Ding, R. Jessberger, G. Trinchieri, S. A. Lira, G. J. Randolph, and J.
S. Bromberg. 2006. Alloantigen-presenting plasmacytoid dendritic cells mediate
tolerance to vascularized grafts. Nat Immunol 7:652-62.
63. Petrovas, C., J. P. Casazza, J. M. Brenchley, D. A. Price, E. Gostick, W. C. Adams,
M. L. Precopio, T. Schacker, M. Roederer, D. C. Douek, and R. A. Koup. 2006. PD-1
is a regulator of virus-specific CD8+ T cell survival in HIV infection. J Exp Med
203:2281-92.
64. Piatak, M., Jr., M. S. Saag, L. C. Yang, S. J. Clark, J. C. Kappes, K. C. Luk, B. H.
Hahn, G. M. Shaw, and J. D. Lifson. 1993. High levels of HIV-1 in plasma during all
stages of infection determined by competitive PCR. Science 259:1749-54.
65. Pichlmair, A., O. Schulz, C. P. Tan, T. I. Naslund, P. Liljestrom, F. Weber, and C.
Reis e Sousa. 2006. RIG-I-mediated antiviral responses to single-stranded RNA bearing
5'-phosphates. Science 314:997-1001.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

66. Read, J. S., K. C. Rich, J. J. Korelitz, L. M. Mofenson, R. Harris, J. H. Moye, Jr., W.
A. Meyer, 3rd, S. G. Pahwa, J. W. Bethel, and R. P. Nugent. 2000. Quantification of
human immunodeficiency virus type 1 p24 antigen and antibody rivals human
immunodeficiency virus type 1 RNA and CD4+ enumeration for prognosis. National
Institute of Child Health and Human Development Intravenous Immunoglobulin Clinical
Trial Study Group. Pediatr Infect Dis J 19:544-51.
67. Rodriguez, B., M. M. Lederman, W. Jiang, D. A. Bazdar, K. Garate, C. V. Harding,
and S. F. Sieg. 2006. Interferon-alpha differentially rescues CD4 and CD8 T cells from
apoptosis in HIV infection. Aids 20:1379-89.
68. Rutz, M., J. Metzger, T. Gellert, P. Luppa, G. B. Lipford, H. Wagner, and S. Bauer.
2004. Toll-like receptor 9 binds single-stranded CpG-DNA in a sequence- and pH-
dependent manner. Eur J Immunol 34:2541-50.
69. Savarino, A., J. R. Boelaert, A. Cassone, G. Majori, and R. Cauda. 2003. Effects of
chloroquine on viral infections: an old drug against today's diseases? Lancet Infect Dis
3:722-7.
70. Savarino, A., M. B. Lucia, E. Rastrelli, S. Rutella, C. Golotta, E. Morra, E.
Tamburrini, C. F. Perno, J. R. Boelaert, K. Sperber, and R. Cauda. 2004. Anti-HIV
effects of chloroquine: inhibition of viral particle glycosylation and synergism with
protease inhibitors. J Acquir Immune Defic Syndr 35:223-32.
71. Schmidt, B., B. M. Ashlock, H. Foster, S. H. Fujimura, and J. A. Levy. 2005. HIV-
infected cells are major inducers of plasmacytoid dendritic cell interferon production,
maturation, and migration. Virology 343:256-66.
72. Sharpe, A. H., E. J. Wherry, R. Ahmed, and G. J. Freeman. 2007. The function of
programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat
Immunol 8:239-45.
73. Siegal, F. P., N. Kadowaki, M. Shodell, P. A. Fitzgerald-Bocarsly, K. Shah, S. Ho, S.
Antonenko, and Y. J. Liu. 1999. The nature of the principal type 1 interferon-producing
cells in human blood. Science 284:1835-7.
74. Soumelis, V., I. Scott, F. Gheyas, D. Bouhour, G. Cozon, L. Cotte, L. Huang, J. A.
Levy, and Y. J. Liu. 2001. Depletion of circulating natural type 1 interferon-producing
cells in HIV-infected AIDS patients. Blood 98:906-12.
75. Sousa, A. E., J. Carneiro, M. Meier-Schellersheim, Z. Grossman, and R. M.
Victorino. 2002. CD4 T cell depletion is linked directly to immune activation in the
pathogenesis of HIV-1 and HIV-2 but only indirectly to the viral load. J Immunol
169:3400-6.
76. Sperber, K., G. Chiang, H. Chen, W. Ross, E. Chusid, M. Gonchar, R. Chow, and O.
Liriano. 1997. Comparison of hydroxychloroquine with zidovudine in asymptomatic
patients infected with human immunodeficiency virus type 1. Clin Ther 19:913-23.
77. Sperber, K., M. Louie, T. Kraus, J. Proner, E. Sapira, S. Lin, V. Stecher, and L.
Mayer. 1995. Hydroxychloroquine treatment of patients with human immunodeficiency
virus type 1. Clin Ther 17:622-36.
78. Stylianou, E., A. Yndestad, L. I. Sikkeland, V. Bjerkeli, J. K. Damas, T. Haug, H. G.
Eiken, P. Aukrust, and S. S. Froland. 2002. Effects of interferon-alpha on gene
expression of chemokines and members of the tumour necrosis factor superfamily in
HIV-infected patients. Clin Exp Immunol 130:279-85.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

79. Trautmann, L., L. Janbazian, N. Chomont, E. A. Said, S. Gimmig, B. Bessette, M. R.
Boulassel, E. Delwart, H. Sepulveda, R. S. Balderas, J. P. Routy, E. K. Haddad, and
R. P. Sekaly. 2006. Upregulation of PD-1 expression on HIV-specific CD8+ T cells
leads to reversible immune dysfunction. Nat Med 12:1198-202.
80. von Sydow, M., A. Sonnerborg, H. Gaines, and O. Strannegard. 1991. Interferon-
alpha and tumor necrosis factor-alpha in serum of patients in various stages of HIV-1
infection. AIDS Res Hum Retroviruses 7:375-80.
81. Wallace, D. J., M. Linker-Israeli, S. Hyun, J. R. Klinenberg, and V. Stecher. 1994.
The effect of hydroxychloroquine therapy on serum levels of immunoregulatory
molecules in patients with systemic lupus erythematosus. J Rheumatol 21:375-6.
82. Yonezawa, A., R. Morita, A. Takaori-Kondo, N. Kadowaki, T. Kitawaki, T. Hori,
and T. Uchiyama. 2003. Natural alpha interferon-producing cells respond to human
immunodeficiency virus type 1 with alpha interferon production and maturation into
dendritic cells. J Virol 77:3777-84.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

Figure Legends
Figure 1. Chloroquine abrogates AT-2 HIV-1 induced DC activation and maturation.
PBMC were stimulated with AT-2 HIV-1 for 20 hours in the presence or absence of 100 µM
chloroquine and analyzed by flow cytometry. Logical gating was used to identify the pDC
population (Lin1-/HLA DR
+/CD123
+) for analyses. A). Representative flow cytometric analysis
and summary of pDC activation marker CD86 and B) Representative flow cytometric analysis
and summary of pDC maturation marker CD83. Results are expressed as the mean % expression
± SEM, n=3
Figure 2. Chloroquine abrogates TLR activated MyD88-dependent signaling pathway and
IFNα production in pDC.
PBMC were stimulated with AT-2 HIV-1 or the TLR9 (CpG A) and TLR7/8 (3M-011) agonists
for 20 hours in the presence or absence of 100 µM chloroquine. Logical gating was used to
identify the pDC population (Lin1-/HLA DR
+/CD123
+) for analyses. A). Representative flow
cytometric analysis and summary of IRAK-4 and IRF-7 signaling molecules expressed in pDC
stimulated with AT-2 HIV-1 ± 100 µM chloroquine. B) Summary of IFNα (pg/ml)
concentration in the culture supernatants from PBMC cultures stimulated with AT-2 HIV-1 and
C) synthetic TLR9 (CpG A) and TLR7/8 (3M-011) agonists ± 100 µM chloroquine. Results are
expressed as the mean ± SEM, n=5.
Figure 3. Chloroquine decreases IFNαααα induced CD8+ T cell activation.
PBMC were stimulated with AT-2 HIV-1 (Ada and MN) for 20 hours in the presence or absence
of physiologicically achievable concentrations of chloroquine (0.5 – 5.0 µM). A) IFNα (pg/ml)
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

concentration in supernatants of AT-2 Ada and MN stimulated PBMC ± chloroquine. B). PBMC
cultures stimulated with AT-2 Ada HIV-1 or 1000 U/ml rIFNα2a were analyzed by flow
cytometry for expression of the CD38 activation marker in the CD8+ T cell subset (CD3
+/CD8
+)
± 5 µM chloroquine or the anti-IFNαR blocking monoclonal antibody, 64G12. C) Miltenyi bead
purified CD8+ T cells stimulated with AT-2 HIV-1 or 1000 U/ml rIFNα2a were analyzed by flow
cytometry for expression of the CD38 activation marker in the CD8+ T cell subset (CD3
+/CD8
+)
± 5 µM chloroquine or the anti-IFNαR blocking monoclonal antibody, 64G12. Results are
expressed as mean ± SEM, n=3.
Figure 4. Chloroquine abrogates AT-2 HIV-1 induced IDO (indoleamine 2-3-dioxygenase)
expression and activity in pDC.
PBMC were stimulated with AT-2 HIV-1 for 20 hours in the presence or absence of 100 µM
chloroquine and analyzed by flow cytometry. A). Representative flow cytometric analysis and
summary of AT-2 HIV-1 induced IDO expression (IDO mean fluorescence intensity; IDO-MFI)
in pDC (Lin1-/HLA DR+/CD123+) ± 100 µM chloroquine. Supernatants from PBMC cultures
stimulated with B) AT-2 HIV-1 or C) TLR agonists for 20 hours ±100 µM chloroquine were
analyzed for IDO activity using fluorometric assays to detect kynurenine and tryptophan. Results
are expressed as the kynurenine: tryptophan ratio (KYN: TRYP). Results are expressed as mean
± SEM, n=5.
Figure 5. Chloroquine abrogates PDL-1 expression in pDC.
PBMC were stimulated with AT-2 HIV-1 for 20 hours in the presence or absence of 100 µM
chloroquine and analyzed by flow cytometry. A). Representative flow cytometric analysis and
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from

summary of AT-2 HIV-1 induced PDL-1 expression on pDC (Lin1-/HLA DR+/CD123+) ± 100
µM chloroquine. B). Representative flow cytometric analysis and summary of TLR agonist
induced PDL-1 expression on pDC (Lin1-/HLA DR+/CD123+) ± 100 µM chloroquine. Results
are expressed as mean ± SEM, n=5.
by guest on January 13, 2010 aac.asm
.orgD
ownloaded from