a structural feature of human immunoglobulin light chains · a structural feature of human...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAI~ CHEMISTRY Vol. 247, No. 4, Issue of February 25, pp 1065-1073, 1972
Printed in U.S.A.
A Structural Feature of Human Immunoglobulin Light Chains
TWO COMPACT DOMAIKS CONNECTED BY A SMALL SWITCH REGIOK*
(Received for publication, October 8, 1971)
3’. ANDERS KARLSSON, PER A. PETERSON, AND INGEMAR BERGG~RD~
From the Institute of Medical Chemistry, University of Uppsala, Uppsala, Sweden
SUMMARY
A lambda immunoglobulin light chain from a myeloma patient, normal light chains, and fragments one-half the size of these chains have been examined by chemical and physical methods. Fragments produced from the X chain with trypsin, pepsin, and papain, and free urinary fragments of this chain were isolated. Characterization of the various fragments of the X chain by amino acid analyses, peptide mapping, NH2-terminal sequence determinations, gel chro- matography, and ultracentrifugations suggested that the fragments consist of whole variable or constant halves, and that they extend one to five residues into the adjacent half. Thus, the region at the mid-point of the chain which is partic- ularly susceptible to proteolysis apparently consists of a polypeptide stretch of less than ten residues. This region seems to be of similar size in normal light chains, since frag- ments of such chains had very similar Stokes’ radii and molecular weights as halves of the X chain. The urinary fragments of the X chain were identical or similar to frag- ments formed by proteolysis. These fragments probably are of catabolic origin.
The halves of the light chains had significantly lower fric- tional ratios than the intact chains, which suggests that whole light chains are more elongated than the halves or contain trapped water in the contact region between the halves (or both).
A characteristic structural feature of immunoglobulins is that their polypeptide chains are composed of an NHz-terminal, var- iable part, the sequence of which differs for each individual chain, and a COOH-terminal, constant part with a practically invariant amino acid sequence within each class of chain. The variable and constant parts are apparently encoded by separate germ- line genes (cf. Reference 1). In a light polypeptide chain (K or X chain’), the variable part (V, or Vi) and the constant part
* This investigation was supported by the Swedish Medical Re- search Council, Project 13x-512 and by grants from the Swedish Cancer Society.
1 Present address, Department of Physiological Chemistry, University of Lund, Sweden.
1 Nomenclature for human immunoglobulins according to Bull. World He&h Organ., 30, 447 (1964), and J. Bid. Chem., 246, 3033
(C, or CA) each constitutes approximately half of the chain. It has been suggested that successive segments of immunoglobulin chains, corresponding in size to half of a light chain, are folded into globular domains which are connected by more extended stretches of polypeptide (1). The domain hypothesis has gained support from studies which have shown that light chains can be cleaved into variable and constant halves by limited proteolysis (2, 3), that the resulting halves have compact shapes (3), and that they may exist as independently folded regions in the in- tact chain (4). This series of studies has emanated from the previous observations that fragments corresponding to variable or constant parts of light chains are excreted in urine by some patients with multiple mgeloma (512) and by normal individ- uals (10).
The main objectives of this investigation have been to compare the hydrodynamic properties of light chain halves with those of complete chains, and to derive an estimate of the size of the switch region2 between the variable and constant parts.
A monoclonal X chain and its isolated halves were examined in detail and additional results were obtained in similar investiga- tions on normal light chains and their halves. The physical studies suggested a more compact structure for the half-chains than for the intact molecule. The extension of the switch region between the halves was apparently less than ten amino acid residues.
EXPERIMEKTAL PROCEDURE
Urine was obtained from patients A. J. and N. D. with multiple myeloma. Urine collections were carried out as described pre- viously (13). A G myeloma protein was isolated from serum of patient A. J. by gel chromatography on Sephadex G-200 followed by zone electrophoresis. Normal human IgG was kindly sup- plied by AR Kabi, Stockholm, Sweden. Light chains from the myeloma protein and normal TgG were prepared by the method of Fleischman, Pain, and Porter (14) as previously outlined (15). Trypsin (TRTPCK twice crystallized) was purchased from Worthington. Mercuripapain, pepsin (twice crystallized) were
(1970). Other abbreviations used: dansyl, l-dimethylaminonaph- talene-5.sulfonyl; pap CA, pap VX, fragments obtained by papain digestion of A. J. x chain; try CA, try VX, fragments obtained by trypsin digestion of A. J. X chain: pep CA, pep VX, fragments ob- tained by pepsin digestion of .I. J. X chain.
2 This region is here defined ts the area which is particularly susceptible to proteolytic digestion.
1065
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1066 Two Compact Parts in Human InmunoglobuLin Light Chains Vol. 2-17, ATO. 4
obtained from Sigma. Sephadex G-100 was purchased from Pharmacia (Uppsala, Sweden), Pevikon C-870 from Kema Nord AU (Stockholm, Sweden). Cellulose (Whatman CC 41) was washed in ethanol, 0.5 RI KOII, 0.5 M HCl, and water before the preparation of thin layer sheets. Polyamide thin layer sheets were products of Cheng Ching Trading Co., Ltd., Hankow, Taiwan. Dansyl amino acids and dansyl chloride were obtained from BDH, England, and phenylisothiocyanate from Pierce Chemical Co. All other chemicals were of the highest grade available.
Methods
Concentration of Proteins-Urine and protein solutions were concentrated by ultrafiltration (16) with 236s inch dialysis tub- ing (Union Carbide Corp., Chicago, Ill.) as previously described (17).
Electrophoresis-Preparative zone electrophoresis was per- formed in blocks (18) of Pevikon C-870 (19), as described earlier (15). Sodium barbital buffer, pH 8.6, ionic strength 0.1, was used in these experiments.
Vertical polyacrylamide gel electrophoresis was carried out in a standard EC 470 Cell (E-C Apparatus Corp., Philadelphia). Electrophoresis was performed according to the instructions supplied by the manufacturer with 4% Cyanogum-41 (E-C Apparatus Corp.) in the spacer gel and 8% in the running gel. A discontinuous buffer system, described by Davies (20), with 0.4 M Tris-HCl buffer, pH 8.9, in the running gel, 0.1 M Tris- HCl buffer, pH 6.7, in the spacer gel, and 0.04 M Tris-glycine buffer, pH 8.3, in the electrode compart.ment was used.
Starch gel electrophoresis in 8 M urea and formate buffer, pH 2.9, (21) was carried out as described elsewhere (22).
Amino Acid Analyses-Quantitative amino acid analyses were carried out as described by Spackman, Stein, and Moore (23). The prot*ein samples (1 tjo 4 mg) were hydrolyzed in 6 N HCl at 110” for 24 and 72 hours (24). Chromatography was carried out on a Biochrom automatic amino acid analyzer (Biocal, Mu- nich). Total cysteine plus cystine was estimated as cysteic acid in samples hydrolyzed for 18 hours after oxidation with performic acid (2.5). Tryptophan was measured spectrophotometrically by the method of Edelhoch (26).
Peptide Jlapping-Protein samples (1 to 2 mg) were digested with trypsin according to Milstein (27). The digest was sub- mitted to electrophoresis on cellulose sheets (0.5 x 220 x 200 mm) in a pyridine-a&ate buffer, pH 5.5, followed by ascending chromatography in the solvent system described by Waley and Watson (28). The peptides were visualized by spraying with ninhydrin (0.2% in acetone).
NIIz Terminal Sequence Determinations-Approximately 0.10 ymole of the various polypeptides were used for determining the NH&erminal amino acid sequences, with sequential Edman deg- radation plus dansylation (29). Identification of the dansylated amino acids was performed by polyamide thin layer chromatog- raphy as described by Woods and Wang (30).
Profein Jleasuremet&-Protein concentrations in solutions of purified protein and in eluates from gel chromatography were determined by reading the absorbance at 280 nm. In other instances protein was measured by the Folin technique of Lowry et al. (31), with human IgG as the standard. The extinction coefficients at 280 nm (EE,) for tl ie various protein prepara- tions were obtained after corrections for content of moisture and ash. lLloisture was measured by drying to constant weight at
100” and 0.05 mm Hg. The following extinction coefficients were found: A. J. X chain, 14.6; normal light chains, 14.3; and halves of normal light chains, 14.5. The constant and variable halves of A. J. X chain were both given a value of 14.6 since the two halves contain the same amounts of tyrosine and tryptophan (see below).
UZtracentrilfiLggation-Sedimentation coefficients and molecular weights were estimated at 20” in a Spinco model E analytical ultracentrifuge, equipped with an RTIC temperature control unit and an electronic speed control device. All experiments were conducted in 0.02 M Tris-HCl buffer, pH 8.0, containing 0.15 M NaCl. The samples were diluted to the appropriate con- centrations and dialyzed in the cold against two changes of the solvent. Standard 12.mm double sector cells with sapphire windows were used throughout. The centrifuge was operated at 60,000 rpm for the sedimentation velocity determinations. The sedimenting boundary was recorded every 16 min with the phase plate schlieren optics at protein concentrations above 2 mg per ml and with the photoelectric scanning system set at 280 nm for concentrations of less than 2 mg per ml. Calcula- tions were carried out according to Schachman (32).
Sedimentation equilibrium experiments were performed with the meniscus depletion technique of Yphantis with speed settings according to his suggestions (33). Recordings were made either with Rayleigh interference optics or with the photoelectric scanner set at 280 nm. Equilibrium times were estimated according to the procedure of Teller et al. (34). In practice, equilibrium was assessed by comparing recordings taken several hours apart. The experiments were discontinued when no significant redistribution of material could be observed over a period of several hours.
Partial specific volumes for the calculations were estimated from the amino acid analyses reported below (35). Densities of the solutions were determined by pycnometry.
Analytical Gel Chromatography-Stokes’ molecular radii were determined by analytical gel chromatography on Sephadex G-100 columns (100 x 1 cm or 140 x 2 cm) equilibrated with 0.02 $1 Tris-HCl buffer, pH 8.0, containing 1.0 M NaCl. 1111 experiments were performed at +4” and analyses were carried out. at least in duplicate.
The void volume, Vo, of the columns was estimated by chro- matogrnphing a dye-marked destran (.lfw > 1.4 X 106). The void volume plus the inner volume was estimated from the elution position of 0.05 PCi 3H~0 in admixture with the chromatographed samples. Calculation of the volume of the gel matrix was based on a bed volume of 20 ml per g of dry Sephadex G-100 and a par- tial specific volume of 0.6 ml per g of Sephades. The total volume, Vt, was taken as the sum of the void volume, the inner volume, and the volume of the gel matrix. The elution volume, V,, of a given solute zone was taken as the concentration maxi- mum of the zone.
Stokes’ molecular radii were computed from the equ .tion given by Laurent and Killander (36), where r, is Stokes’ molecu- lar radius, and L and rr are constants characteristic of each gel. r, was assumed to be 6.5 A (36, 37). L was determined by chromatography of human albumin (r, = 35.5 A).
Calculations of Di$usion Coeficients, Molecular Weights, and Frictional Ratios-Apparent diffusion coefficient (D20,w) were computed from Stokes’ radii by use of the Stokes-Einstein equa- tion (38). Molecular weights were determined by sedimentation equilibrium ultracentrifugation as described above, or calculated
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Issue of February 25, 1972 F. A. I~a6wn, P. A. Peterson, and I. Berg&d 1067
from sedimeutation coefficients, diffusion coefficients, and partial specific volumes by the equation of Svedberg. Frictional ratios (f/j,,) were calculated from Stokes’ radii, sedimentation CO- efficients, molecular weights, and partial specific volumes. The formulas used are found in Reference 39.
Viscosity nfeasurements-Viscosity measurements were made at 25.5 f 0.01” with Cannon-Xanning semimicro viscometers. Flow times for 0.02 M Tris-HCl buffer, pH 8.0, containing 0.2 M NaCl were 225 to 270 see, and for protein solutions 15 to 65 set longer. Flow times were reproducible to at least f 0.20 sec. Measurements were carried out at protein concentrations rang- ing from 0.01 to 0.04 g per ml. Values for the intrinsic viscos- ities were obtained by treatment of the data according to No- elken et al. (40).
Other Xethods-Immunoglobulin components were reduced at room temperature by treatment with 0.1 M 2-mercaptoethanol (Calbiochem) in 0.2 M Tris-HCl buffer, pH 8.0. The resulting sulfhydryl groups were alkylated 1 hour later with a 10% molar excess of iodoacetamide (Mann). In some instances the reduc- tion was carried out in the presence of 8 til urea. The immuno- logical identification of light chain fragments was carried out with the Ouchterlony immunodiffusion technique (41). Specific anti- sera against the A. J. light chain and its free Vx and free CA fragments were used (3).
Isolation oj Bence-Jones Proteins and Urinary Light Chain Fragments
Concentrated urinary macromolecules from patient A. J. were chromatographcd at +4” on a Sephadex G-100 column equili- brated with 0.02 M Tris-HCl buffer, pH 8.0, containing 1.0 M NaCl. Several peaks were obtained (Fig. 1). The dimer, the monomer, and the fragments of light chain were located by their previously established relative elution volumes (15, 3). Each of these three materials were subjected to zone electrophoresis. The monomer and the dimer wcrc obtained as homogeneous fractions after this isolation step. The elcctrophoretic scpara-
ELUTION VOLUME (ml)
FIG. 1. Chromatography on Sephadex G-100 of concentrated urinary proteins (11,300 mg) from patient A. J. The colllmn (115 X 8 cm) was equilibrated with 0.02 M Tris-HCl buffer, pTI 8.0, con- taining 1 M NaCl. Fractions of 20 ml were collected at 20.min in- tervals. The el~lted protein was combined into three fractions as indicated by the arrows. The elution positions of the fractions (from the lej/) correspond to dimer, monomer, and fragments of light chains.
tion of constant and variable fragments is illustrated in Fig. 2. A second Sephadex G-100 chromatography was performed for final purification of the fragments. Fig. 3 shows a chromatogram of the constant fragment. The yield of dimer, monomer, con- stant, and variable fragment, from eight 24.hour urine specimens was 2800, 1430, 18, and 14 mg, respectively.
B dimeric X Bence-Jones protein was also isolated from corn centrated urinary macromolecules from patient K. D. as de- scribed above. No light chain fragments were obtained in this instance.
Preparation of Fragments from Normal Light Clzains
Light chains from normal IgG were digested with trypsin 30 min at 37” in 1 M Tris-HCl buffer, pH 8.0, with an enzyme to
r , I I I
FRACTION NUMBER FIG. 2. Zone electrophoresis in barbital buffer, pII 8.G, of light
chain fragments (294 mg) obtained by chromatography on Sepha- dex G-100. The potential gradient was 3.5 volts per cm and the time, 48 hours. Total protein was estimated by the Folin tech- nique. The slowly migrating constant fragment and the rapidly migrating variable fragment were identified by the Ouchterlony immunodiffusion technique with specific antisera against each fragment. Fractions containing these fragments were pooled as indicated by the WTOWS. The anode is to the righl.
F c 0.6
:
2
2 04
2
i
ELUTION VOLUME /ml)
FIG. 3. Chromatography on Sephadex G-100 of the fraction con- taining the free Cx fragment obtained by zone electrophoresis. The column (140 X 2 cm) was equilibrated with 0.02 M Tris-HCI buffer, pH 8.0, containing 1.0 1% NaCl. Fractions of 3.5 ml were collected at a flow rate of 10.5 ml per hour. Fractions containing free Cx fragments were pooled as indicated by the arrows.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1068 Two Compact Pads in Human Immunoglobulin Light Chains Vol. 247, No. 4
substrate ratio of 1: 250. The digest was separated on a Sepha- dex G-100 column (cj. Fig. 3, Reference 3). A typical experi- ment gave fragments corresponding to halves of light chains in a yield of 10%.
Fragments of normal light chains produced by trypsin were chosen in the present study since these fragments could be ob- tained in particularly good yields (3). As previously shown by immunodiffusion analyses (3), this material contained both C, and CA halves.
Preparation of Fragments from A. J. X Chain
X chains obtained by reduction and alkylation of the dimeric Bence-Jones protein A. J. was submitted to enzymatic digestion. Incubation with trypsin, papain, and pepsin produced different proportions of constant and variable fragments of the A. J. X chain. A polyacrylamide gel electrophoretic analysis, shown in Fig. 4, illustrates the difference between trypsin and papain di- gestion. Tryptic digestion gave rise to a variable fragment in high yield and smaller amounts of two constant fragments, whereas papain digestion produced a distinct constant fragment, but only traces of variable fragment. Pepsin digestion produced a constant fragment; no variable fragment could be detected.
Tryptiti fragments were prepared as follows. The protein (200 mg) was dissolved in 10 ml of 1 M Tris-HCl buffer, pH 8.0, and trypsin (4 mg in 0.001 M HCl) was added. After 1-min in- cubation at 37”, the digestion was stopped by the addition of soybean trypsin inhibitor (5 mg). The digest was separated by zone electrophoresis in sodium barbital buffer, pH 8.6. After electrophoresis, protein fractions in positions corresponding to the free constant and free variable fragments were pooled and concentrated. Each fragment was purified by gel chromatog- raphy on a Sephadex G-100 column (2 x 140 cm) equilibrated with 0.02 M Tris-HCl buffer, pH 8.0, containing 1 M NaCl. Constant fragment (try CA) was obtained in a yield of approxi-
mately 4 mg. A variable fragment (try VX) was obtained in a in a yield of 10 mg.
For the preparation of peptic fragments, light chains (150 mg) were dissolved in 15 ml of 0.1 M citrate-phosphate buffer, pH 3.2, and mixed with enzyme (6 mg in 0.6 ml buffer). The di- gestion at 37” was stopped after 5 min by adjusting the pH to 8 with 2 M Tris-HCI buffer. Peptic fragments were isolated by the same procedure as used for the tryptic fragments. No variable fragment could be isolated. A constant (pep Cx) was obtained in a yield of 11 mg.
Papain fragments were prepared by the following procedure. The protein (300 mg) was dissolved in 12 ml of 0.1 M sodium- phosphate buffer, pH 7.0, containing 4 mM cystein and 1 mM EDTA, and digested with papain (1.2 mg) at 37” for 3 min. The digestion was terminated by cooling to 4’ and by addition of iodoacetamide to 5 mM. After purification, as described above, 12.5 mg of a constant fragment (pap CA) was obtained. NO variable fragment could be isolated.
Homogeneity of Puri$ed Fragments
The various isolated VX and CX fragments appeared homo- geneous on Ouchterlony immunodiffusion analyses with anti- serum against the A. J. X chain. There was no indication of cross reactivity between the VA and CA fragments (cf. Fig. 2, Reference 3). All fragments emerged as symmetrical peaks from Sephadex G-100 columns, and were thus homogeneous with respect to size (see Fig. 3).
The purified free Vi, try VA, free CA, try CA, pep CA, and pap CA fragments were examined by electrophoresis in polyacrylamide gel at pH 8.3 and in urea starch gel at pH 2.9. On polyacryl- amide gel electrophoresis, the free CA fragment gave two zones, an intense, slowly migrating zone and a weak somewhat faster zone. The pap CA fragment showed the same pattern but most often with the proportions of the two zones reversed. The two
123456789
5
Al A2 A3 Bl 82 83
Cl C2 C3 Dl 02 D3
FIG. 4. Polyacrylamide gel electrophoresis of trypsin and papain digests of the A. J. x chain: Trypsin digests (1 to 4)) A. J. X chain (6), and papain digests (6 to 9). The enzyme to substrate ratios were, 1:50 for trypsin; and 1:250, for papain. The incubation times were: 0.5, 3. 10, and 30 min. Times increases in each instance from the left to the right.
FIG. 5. Starch gel electrophoresis in 8 M urea and formate buffer, pH 2.9, of: free CA (A), pap Ch (B), free VA (C) and try VA-fragments (D) ; 1, reduced and alkylated, 2, reduced and alkylated in the presence of 8 M urea, and 3, untreated.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from
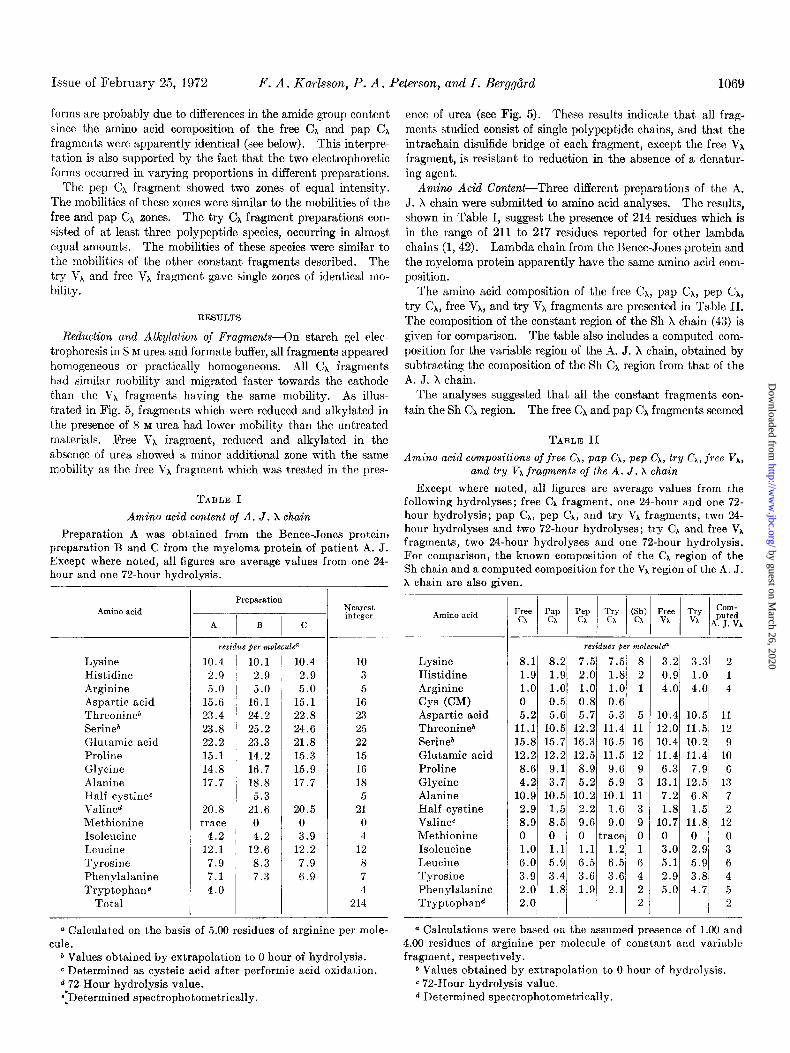
Issue of February 25, 1972 F. A. Karlsson, P. A. Peterson, and I. BerggiErd 1069
forms are probably due to differences in the amide group content ence of urea (see Fig. 5). These results indicate that all frag- since the amino acid composition of the free CA and pap CA ments studied consist of single polypeptide chains, and that the fragments were apparently identical (see below). This interpre- intrachain disulfide bridge of each fragment, except the free Vi tation is also supported by the fact that the two electrophoretic fragment, is resistant to reduction in the absence of a denatur- forms occurred in varying proportions in different preparations. ing agent.
The pep Cl fragment showed two zones of equal intensity. The mobilities of these zones were similar to the mobilities of the free and pap CA zones. The try CA fragment preparations con- sisted of at least three polypeptide species, occurring in almost equal amounts. The mobilities of these species were similar to the mobilities of the other constant fragments described. The try VA and free Vh fragment gave single zones of identical mo-
bility.
Amino Acid Content-Three different preparations of the A, J. X chain were submitted to amino acid analyses. The results, shown in Table I, suggest the presence of 214 residues which is in the range of 211 to 217 residues reported for other lambda chains (1,42). Lambda chain from the Bence-Jones protein and the myeloma protein apparently have the same amino acid com- position.
RESULTS
Reduction and Alkylation of Fragments-On starch gel elec- trophoresis in 8 M urea and formate buffer, all fragments appeared homogeneous or practically homogeneous. All CA fragments had similar mobility and migrated faster towards the cathode than the VX fragments having the same mobility. As illus- trated in Fig. 5, fragments which were reduced and alkylated in the presence of 8 M urea had lower mobility than the untreated materials. Free VA fragment, reduced and alkylated in the absence of urea showed a minor additional zone with the same mobility as the free Vx fragment which was treated in the pres-
The amino acid composition of the free CA, pap CA, pep CA, try CA, free Vi, and try Vi fragments are presented in Table II. The composition of the constant region of the Sh X chain (43) is given for comparison. The table also includes a computed com- position for the variable region of the A. J. X chain, obtained by subtracting the composition of the Sh CA region from that of the A. J. X chain.
The analyses suggested that all the constant fragments con- tain the Sh Cx region. The free CA and pap CA fragments seemed
TABLE II
Amino acid compositions of free CX, pap CX, pep CA, try CA, free VA, and try Vi fragments of the A. J. X chain
TABLE I
Amino acid content of A. J. X chain
Preparation A was obtained from the Bence-Jones protein, preparation B and C from the myeloma protein of patient A. J. Except where noted, all figures are average values from one 24- hour-and one 72-hour hydrolysis.
Except where noted, all figures are average values from the following hydrolyses; free Cx fragment, one 24.hour and one 72- hour hydrolysis; pap CX, pep CX, and try VX fragments, two 24- hour hydrolyses and two 72-hour hydrolyses; try CA and free Vh fragments, two 24-hour hydrolyses and one 72-hour hydrolysis. For comparison, the known composition of the CA region of the Sh chain and a computed composition for the VX region of the A. J. X chain are also given.
Preparation
Amino acid
A B C
residue pev molecule”
Lysine Histidine Arginine Aspartic acid Threonineb Serineb Glutamic acid Proline Glycine Alanine Half-cystinec Valined Methionine Isoleucine Leucine Tyrosine Phenylalanine Tryptophane
Total
10.4 2.9 5.0
15.6 23.4 23.8 22.2 15.1 14.8 17.7
10.1 2.9 5.0
16.1 24.2 25.2 23.3 14.2 16.7 18.8 5.3
21.6 0 4.2
12.6 8.3 7.3
10.4 2.9 5.0
15.1 22.8 24.6 21.8 15.3 15.9 17.7
20.8 trace
4.2 12.1 7.9 7.1 4.0
20.5 0 3.9
12.2 7.9 6.9
Amino acid
10 Lysine 8.1 8.2 7.5 7.5 8 3 Histidine 1.9 1.9 2.0 1.8 2 5 Arginine 1.0 1.0 1.0 1.0 1
16 CYS (CM) 0 0.5 0.8 0.6 23 Aspartic acid 5.2 5.6 5.7 5.3 5 25 Threonineb 11.1 10.5 12.2 11.4 11 22 Serineb 15.8 15.7 16.3 16.5 16 15 Glutamic acid 12.2 12.2 12.5 11.5 12 16 Proline 8.6 9.1 8.9 9.6 9 18 Glycine 4.2 3.7 5.2 5.9 3 5 Alanine 10.9 10.5 10.2 10.1 11
21 Half-cystine 2.9 1.5 2.2 1.6 3 0 Valinec 8.9 8.5 9.6 9.0 9 4 Methionine 0 0 0 trace 0
12 Isoleucine 1.0 1.1 1.1 1.2 1 8 Leucine 6.0 5.9 6.5 6.5 6 7 Tyrosine 3.9 3.4 3.6 3.6 4 4 Phenylalanine 2.0 1.8 1.9 2.1 2
214 Tryptophand 2.0 2
a Calculated on the basis of 5.00 residues of arginine per mole- cule.
b Values obtained by extrapolation to 0 hour of hydrolysis. c Determined as cysteic acid after performic acid oxidation. d 72-Hour hydrolysis value. e:Determined spectrophotometrically.
3.2 3.3 2 0.9 1.0 1 4.0 4.0 4
10.4 10.5 11 12.0 11.5 12 10.4 10.2 9 11.4 11.4 10 6.3 7.9 6
13.1 12.5 13 7.2 6.8 7 1.8 1.5 2
10.7 11.8 12 0 0 0 3.0 2.9 3 5.1 5.9 6 2.9 3.8 4 5.0 4.7 5
I 12 a Calculations were based on the assumed presence of 1.00 and
4.00 residues of arginine per molecule of constant and variable fragment, respectively.
b Values obtained by extrapolation to 0 hour of hydrolysis. c 72.Hour hydrolysis value. d Determined spectrophotometrically.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1070 Two Compact Parts in Human I~wwunoylobulin Light Chains Vol. 247, No. 4
~=C~NS~ANT BB= VARIABLE
“ORIGIN + J FIG. 6. Peptide maps of tryptic digests of (A) A. J. x chain; (B) free CL fragment; (C) try VA-fragment: and (D) A. J. X chain (draw-
ing). The tryptic peptides were separated in the first dimension by electrophoresis and in the second dimension by ascending chroma- tography as described under “Experimental Procedure.”
to be identical. These two fragments appear to consist of the Sh CA region plus glycine. Extra glycine was also present irr the pep CA and try CX fragments.
The amino acid contents of the free VA and try VA fragments suggest that the two fragments contain the entire or practically the entire variable region. The intact light chain apparently contains ten lysine residues, the constant region eight, and each of the variable fragments three lysine residues. These obscrvn- tions and the known specificity of trypsin suggest that the VA fragments extend to the first lysine residue in the constant region.
Peptide Maps-Tryptic peptides from various protein prepa- rations were separated by electrophoresis followed by chromu- tography. Fig. 6 shows the peptide maps of reduced, alkylatrd A. J. X chain, and its free CA and try VX fragments. The draw- ing in Fig. 6D illustrates the peptides, which are assumed to originate from the constant and the variable regions of the A. J. X chain. The interpretation is based on a comparison of the peptide maps of this chain and its CA and VA fragments with the peptide map of another reduced, alkylated X chain (N. I).).
The peptide maps of the complete A. J. X chain showed about fifteen distinct spots and a few faint spots. This number of peptides agrees well with the content of fifteen residues of lysine plus arginine per molecule (Table I). Some faint spots may have originated from incomplete cleavage or from chymotryptic:- like cleavage.
The maps of the CA fragments revealed nine to 10 distinct and two or three faint spots. All these spots, except one (in some instances two), were also found in the map of the complete A. J. X chain. The extra peptide most likely represents the NIL terminal sequence of the Ci-fragments.
Peptide mapping of the free Vi and try VA fragments gave very similar patterns. All the Vx peptides were contained in the pep tide map of the A. J. X chain.
NIIP-Terminal Amino Acid Sequences--In order to evaluate the results of the NH%-terminal sequence analyses, it is necessary to examine the sequences reported for the switch region between the variable and constant parts in lambda chains. Fig. 7 shows the amino acids most often found in this region. Nine human chains have been sequenced completely (I, 42). Variations have l)een established only for three residues in a sequence of seven residues adjacent to the constant half (Residues 102 to 108 in the numbering system for the Sh X chain, cf. Reference 43). The sequence in this part of the variable region is relatively conserva- tive. We have interpreted our findings on the basis of the se- quence indicated in the figure, assuming that the A. J. X chain has lysine, leucine, and glycine, in Positions 103, 104, and 108, respectively.
The free CL and pap CA fragments had identical NHz-terminal sequences. Three residues were determined and the sequence Gly-Gls-Pro- was found in both instances. These results, in conjunction with the results described above, seem to establish that these polypeptides consist of the constant region plus a glycine residue (Gly-108) from the variable region.
The NHz-terminal sequence found in the pep Cx fragment was Thr-Val-Leu-Gly-. This observation indicates that the pep CA fragment begins in the variable region at Position 105.
The analyses of the try Q, fragment preparation revealed more than one amino acid in each sequential step, indicating that this preparation was a mixture of fragments. The best interpreta- tion of possible NHt-terminal sequences is shown in Fig. 7. The results suggest that one fragment starts at Position 112, another at Position 104 and a third fragment at Position 105. This last mentioned polypeptide most likely derives from a chymotryptic cleavage, since it is not consistent with trypsin specificity. The three fragments seemed to be present in approximately equal amounts. No NHZ-terminal amino acid was found in the A. J.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Issue of February 25, I!)72 p’. A. Karlsson, I’. A . I’et~twm, and I. Beryydrd 1071
Variable region 1 Constant region
102 109
Gl.n( 1) Ar6(2)
Arg(2) Val(2) S-(3)
-T~LT(9)-Lys(6)-Leu(7)-'rhr(S))-Val(S))-Leu(S))-Gly(I~)-Gl n-Pro-Lys-Ala-Ala-Pro-Ser-
Free CA Gly- Glx-Pro-
Pap Cl Gly- Glx-Pro-
Pep Cl Thr- Val- Leu- Gly-
Try Cl Ala-Ala-Pro-
Leu- Tlir- Vnl-
Thr- Val- Leu-
FIG. 7. Comparison of the most, common amino acid residues in Position 102 to 115 in human lambda chains and NHp-terminal se- qrlences found in various constant fragments of the A. J. X chain. Numbs in parentheses refer to the frequency of amino acid residues in this part of the variable region found in nine human lambda. chailm (I, 42).
TABLE III
l’hyskal properties of light chains and thei? corresponding halves
A. J. X chain I)imer Monomer Free CL rap CA
I 1 I 1 1 I 1 I'cp cx 'lky CA
1.8 a Free Vx r3 'I'ry Vi
1.6- - Normal light chai Ijimers
I I I I I I I f Ialves 0.2 04 0.6
s
3.6 2.3 1.6 1.6e 1.6f 1.6 1.8 1.7f
ns 3.6
A m2sec
29.0 7.0 23.5 9.0 17.0 12.5 17.0 12.5 17.5 12.0 17.5 12.0 15.5 14.0 15.5 14.0
29.5 7.0 16.5 13.0
- D20.w” x 10’ i/fob
1.28 1.29 1.17 1.17 1.18 1.17 1.05 1.09
1.31 1.14g
I
L
Molecular weight
43,700c 22,500d 11,200d 11,200" 11,500d 11,300c 11,400c 11,100~
45,OOOd 10,500c
PROTEIN CONCENTRATION 1%)
FIG. ri. Sedimentation coefficients of A. J. h chain and its halves. The following symbols are used: disulfide linked dimer (O), monomer (o), free Vh half (A), and free CA half (A). The mono- mer was obtained by redllction and xlkylation of the disulfidf:. linked dimer.
X chain, in the free Vi fragment, or t,he try VA fragment. This
finding indicates that the nTH4-terminal residue of the three com-
ponents is blocked and suggests that the free VA and try VA poly- peptides contain the intact KHz-terminal end of the A. J. X
chain. Xo attempts Tvere made to identify pyrrolidone carhox-
ylic acid which is known to occur as a blocking residue in X chains. Physical Characteristics--Some, physical properties of the A.
J. X chain and its constant and variahle halves are given in Table III. Sedimentation velocity experiments at concentrations
ranging from 0.2 mg per ml to 8 mg per ml indicated homogeneity
for all preparations examined with exception for the A. J. X chain
monomer which apparently participated in a monomer-dime1
* Estimated from data obtained by gel chromatography. b Frictional ratios were calculated from the sedimentation co-
&Gents and the molecular weights. Frictional ratios calcu- lated from Stokes’ radii and molecular weights gave values in close correspondence to those listed.
c ljetermined by sedimentation equilibrium ultracentrifuga- l,ioll.
d Calculated from sedimentation and diffusion coefficients. c &%-value. f &Y-value. g Calculated from Stokes’ radius and molecular weight.
ccluilibrium ( cf. Fig. 8). As shown in Fig. 8, the sedimentat,ion
behavior of the covalently linked dimer and the constant and
variable halves were almost independent of protein concentra-
tion.
From the data given in Table III the frictional ratio (f/j%) was
calculutjed for each component. As can be seen in the Table, A.
.J. h chain dimer and monomer exhibit similar if not identical
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

1072 Two Compact Parts in Human Immunoglobulin Light Chains Vol. 247, No. 4
frictional ratios. The VA halves have considerably lower frictional ratios than the intact chain whereas the CA halves apparently have intermediate values, differing however signif- icantly from those of dimers and VA halves. All frictional ratios given, ranging from 1.05 to 1.31, are indicative of a globular structure for halves as well as for the intact chains (cf. Reference 44). The composite nature of the frictional ratio, being depend- ent on shape and hydration, makes an unambiguous interpreta- tion difficult. Assuming all components are equally hydrated the data would suggest that the halves are more spherical than the intact chain.
The intrinsic viscosities for the A. J. X chain dimer and for the constant half obtained by papain digestion were 3.9 ml per g and 3.6 ml per g, respectively. Globular proteins generally have intrinsic viscosities of 3.3 to 4.0 ml per g (44). Thus, these re- sults apparently are compatible with the conclusions drawn from the frictional ratios.
Physical characteristics of dimers and halves of normal light chains are presented in Table III. It is apparent that there is a significant difference between the frictional ratios of the intact chains and their halves and that this difference is of similar magnitude as that between frictional ratios of the A. J. X chain and its halves. Provided that cleavage is not accompanied by a large decrease in hydration, this observation lends further support to the suggestion that halves of human light clrsins are less elongated than the complete chains (see above).
DISCUSSIOX
The investigations which have shown that human immuno- globulin light chains can be cleaved into halves by limited pro- teolysis (2, 3) have originated from previous observations of the presence of fragments corresponding to variable or constant parts of light chains in the urine of patients with multiple mye- loma (5-12) and normal individuals (10). Metabolic studies with human Bence-Jones proteins in man (45) and rat (2) have established that such fragments can be formed by catabolism. However, recent work (2, 46) suggests that some fragments found in urine might arise from de novo synthesis. Thus, mouse myeloma cells have been reported to synthesize a protein one- half the size of a light chain (46). This protein was considered to be the variable half. Some biosynthetic studies with bone marrow cells from patients with multiple myeloma have sug- gested that both variable and constant halves may be synthe- sized de novo (2) although other such studies have proved in- conclusive (12, 47).
Urinary light chain components from myeloma patient A. J. included both free constant and free variable halves. The free CA fragment apparently was identical with the CA fragment pro- duced from the complete h chain by papain digestion. Both consist of the constant region plus a glycine residue from the variable region. The free VA fragment was very similar to the Vi fragment formed by trypsin digestion. Characterization of these two fragments suggested that they extend three residues into the constant region with lysine (Lys-111) as the COOH- terminal residue. It is of interest to note that in two of the three urinary X chain fragments which have been characterized pre- viously, lysine at Position 111 was designated as the COOH- terminal residue (12). It is most likely that the CA nrld Vx
fragments isolated from the urine of patient A. J. are catabolic products, since the same fragments seem to be produced by
proteolysis in vitro and since the fragments do not correspond exactly to variable and constant halves.
Proteolytic digestion of the A. J. X chain with papain, pepsin, and trypsin gave rise to one predominant type of fragment corresponding either to the constant or to the variable half. It has been suggested that the variable half of light chains is more resistant to proteolysis than the constant half (45, 2). This would be consistent with the fact that most urinary fragments described have corresponded to the variable part. As to the A. J. h chain, however, the proportions of constant and variable halves formed are apparently determined by the type of enzymic activity and not by different stabilities of the halves.
The properties of the various fragments of the A. J. X chain suggest that these fragments consist of whole variable or con- stant halves and that they extend one to five residues into the adjacent half. The switch region between the variable and con- stant parts, seemed to consist of a polypeptide stretch of less than ten amino acid residues. As demonstrated previously (3) tryp- tic fragments of normal light chains contain both C, and Ch halves. This material very likely also includes V, and Vi halves, since variable fragments often may be particularly resistant to proteolysis (see above) and since it was established in this work that trypsin digestion of the A. J. X chain gives predominantly Vi fragments. Fragments produced from normal light chains by trypsin, papain, and pepsin or isolated in free form from normal urine derived from both K chains and X chains and have approximately the same size (3). As shown in this study, such fragments had very similar Stokes’ radii and molecular weights as halves of the A. J. X chain. Accordingly, the switch regions of normal light chains should be of similar magnitude as the switch region of the A. J. X chain.
Previous work has shown that human light chains are pref- erentially split by proteolytic enzymes in the area connecting the variable and constant regions (2, 3), and that the resulting halves are compact structures (3) which seem to be folded rela- tively independent of each other (4). This supports the hypoth- esis that the polypeptide chains of immunoglobulins consist of linearly arranged globular domains connected by less tightly folded parts of the chains (1). The present observations that the frictional ratios for halves of light chains are significantly lower than for the complete chains further corroborate the do- main theory. It seems most likely that the differences in frictional ratios are due to the fact that the intact chains are more elongated than the halves, although at least part of these differ- ences could also be due to the presence in whole light chains of trapped water in the contact region between the domains.
Rcknozuledgments-We wish to thank Dr. L. Rask and Miss Y. Fernstedt for performing the NHI-terminal sequence deter- minations.
l(.IWlGiFNCP’S 2 1
1. I~~ELMAN, G. M., AND GALL, W. E. (1969) Annu. Rev. Biochem., 38, 415.
2. QOIOMON, A., AND MC LAUGHLIN,~. L. (1969) J.Biol. Chem., 244, 3393.
3. KAMLSSON, F. A., PISTMXSON, 1’. A., AND BEKGG~RD, I. (1969) hoc. Nat. Acad. Sci., Ii. S. A 1 64, 1257.
4. H.J~RK, I., KARLSSON, F. A.,ANDBISKGGBRD, I. (1971)Proc. Nat. A cad. Xci., U. S. A., 66, 1707.
5. I~I~:UITSCH, H. F. (19G3) Scierxe, 141, J35. 6. SIJLOMON, A., KILLANDER, J., GKI~Y, II. M., ASD KUNKEL, H.
G. (1966) Science, 161, 1237.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Issue of February 25, 1972 F. A. Karlsson, I’. A . I’rlerson, mad I. Bwggiird 1073
7. CIOLI, D., AND BAGLIONI, C. (1966) J. Mol. Biol., 16, 385. 8. WILLIBMS, R. C., JR., PINNELL, S. R., AND BRATT, G. T. (1!)66)
J. Lab. Clin. Med., 68, 81. 9.
10. 11.
TAN, M., AND EPSTEIN, W. (1967) J. Immunol., 98, 568. BERGG~~RD. I.. AND P~:TERSON. I?. (1967) in Nobel sumn.. 3. 71. I I VAN EYK, H. G., AND Myszeo&h. K. i1967) Clin. ?%A. i cla,
18, 101. 12. BAGLIONI, C., CIOLI, D., GORINI, G., RUFFILJ, A., AND ALEX-
CIO-ZONTA, L. (1967) Cold Spring Harbor Symposia on &an,- titative Biology, 32, 147.
13.
14.
15.
BERGG~RD, I., AND BEARN, A. G. (1968) J. Biol. Chem., 243, 4095.
FLEISCHMAN, J. B., PAIN, K. H., AND PORTER, R. It. (1902) Arch. Biochem. Biophys., 1, (suppl.), 174.
BIXRGG~RD, I., AND PETI<;RSON, P. A. (1969) J. Biol. Chem., 244, 4299.
16. EVERALL, P. H., AND WRI~XT, G. II. (1958) J. Med. Lab. ‘I’cch- nol., 16, 209.
17. BERGGHRD, I. (1962) Ark. Kemi, 18, 291. 18. KUNKEL, H. G. (1954) Methods Biochem. Anal., 1, 141. 19. MILLER-EBERHARD. H. J. (1960) Stand. J. Clin. Lab. Inwest., I
12, 33. 20. DAVIES, B. J. (1964) Ann. N. Y. Acad. Sci., 121, 404. 21. EDELMAN, G. M., AND Pourlrrc, M. D. (1961) J. Exp. Med., 113,
861. 22.
23.
PETERSON, P. A., AND B~:RGG~~RD, I. (1971) J. Biol. Chem., 246, 25.
SPACKMAN, P. H., STEIN, W. H., AND MOORE, S. (1958) Anal. Chem., 30, 1190.
24. VAN HOFSTEN, B., VAN KLEY, H., AND EAKER, D. (1965) l?io-
25. 26. 27.
chim. Biophys. Acta, 110, 585. MOORE, S. (1963) J. Biol. Chem., 236, 235. EDELHOCH, H. (1967) Biochemistry, 6, 1948. MILSTEIN, C. (1968) Biochem. J., 110, 652.
28. WALI~Y, S. G., AND WATSON, J. (1953) Biochem. J., 66, 328. 29. GRAY, W. R. (1967) Methods Enzymol., 11, 469. 30. WOODS, K. R., AND WANG, K.-T. (1967) Biochem. Biophys.
Acta, 133, 369. 31. TJOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RAND.~LL,
II. J. (1951) J. Biol. Chem., 193, 265. 3“. SCHACH~AN, H. K. (1957) Methods Enzymol., 4, 52. 33. YPHANTIS, P. A. (1964) Biochemistry, 3, 297. 34. TELLER, D. C., HORBETT, T. A., RICHARDS, E. G., AND SCHACH-
MAN, H. K. (1969) Ann. N. Y. Acad. Sci., 164, 66. 35. EDSaLL, J. T. (1943) in E. J. COHN AND J. T. EDSALL (Editors),
Proteins, amino acids and peptides, Reinhold Publishing Corporation, New York, p. 375.
36. LAURENT, T. C., AND KILLANDER, J. (1964) J. Chromatogr., 14, 317.
37. &RINK, B., LAURENT, T. C., AND RIGLER, R. (1967) J. Chro- matogr., 31, 48.
38. COSTING, L. J. (1956) Advan. Protein Chem., 11, 449. 39. SVF,DBF,RG, T., AND PEDERSEN, K. 0. (1940), The ultracentri-
fuse, Oxford University Press, Oxford, p. 34. 40. NOELKEN, M. E., NELSON. C. A., BUCKLEY, C. E., III, AND
TANFORD, C. (1965) J. Biol. Chem., 240, 218. 41. OUCHTERLONY, 0. (1958) Progr. AZZergy, 6, 1 42. GARVER, F. A., AND HILSCHMANN, N. (1971) FEBS (Fed. Eur.
Biochem. Sot.) Lett., 16, 128. 43. WIKLER, M., TITANI, K., SHINODA, T., AND PUTMAN, F. W.
(1967) J. Biol. Chem., 242, 1668. 44. TANFORD, C. (1963) Physical chemistry of macromolecules, John
Wiley and Sons, Inc., New York, p. 394. 45. CIOLI, D., AND BAGLIONI, C. (1968) J. Exp. Med., 128, 517. 46. SCHUBERT, D., AND COHN, M. (1970) J. Mol. Biol., 63, 305. 47. CIOLI, D., AND BAGLIONI, C. (1967) Nobel Symposium, Vol. 3,
p. 401.
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from

F. Anders Karlsson, Per A. Peterson and Ingemar BerggårdDOMAINS CONNECTED BY A SMALL SWITCH REGION
A Structural Feature of Human Immunoglobulin Light Chains: TWO COMPACT
1972, 247:1065-1073.J. Biol. Chem.
http://www.jbc.org/content/247/4/1065Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/4/1065.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 26, 2020
http://ww
w.jbc.org/
Dow
nloaded from