a practical approach for estimating the escape ratio of near...
TRANSCRIPT

Contents lists available at ScienceDirect
Remote Sensing of Environment
journal homepage: www.elsevier.com/locate/rse
A practical approach for estimating the escape ratio of near-infrared solar-induced chlorophyll fluorescenceYelu Zenga,b,⁎,1, Grayson Badgleya,c,1, Benjamin Dechantd, Youngryel Ryud,e, Min Chenf,J.A. Berryaa Department of Global Ecology, Carnegie Institution for Science, Stanford, CA 94305, USAb State Key Laboratory of Remote Sensing Science, Institute of Remote Sensing and Digital Earth, Chinese Academy of Sciences, Beijing 100101, Chinac Department of Earth System Science, Stanford University, Stanford, CA 93405, USAd Research Institute of Agriculture and Life Sciences, Seoul National University, Seoul, Republic of Koreae Department of Landscape Architecture and Rural Systems Engineering, Seoul National University, Seoul, Republic of Koreaf Joint Global Change Research Institute, Pacific Northwest National Laboratory, College Park, MD, USA
A R T I C L E I N F O
Keywords:Solar-induced chlorophyll fluorescenceNear-infrared reflectanceCanopy structureEscape ratioSpectral invariant properties
A B S T R A C T
Solar-induced chlorophyll fluorescence (SIF) has emerged as a leading approach for remote sensing of grossprimary productivity (GPP). While SIF has an intrinsic, underlying relationship with canopy light capture andlight use efficiency, these physiological relationships are obscured by the fact that satellites observe a small andvariable fraction of total emitted canopy SIF. Upon emission, most SIF photons are reabsorbed or scatteredwithin the canopy, preventing their observation remotely. The complexities of the radiative transfer process,which vary across time and space, limit our ability to reliably infer physiological processes from SIF observa-tions. Here, we propose an approach for estimating the fraction of total emitted near-infrared SIF (760 nm)photons that escape the canopy by combining the near-infrared reflectance of vegetation (NIRV) and the fractionof absorbed photosynthetically active radiation (fPAR), two widely available remote sensing products. Ourapproach relies on the fact that NIRV is resilient against soil background contamination, allowing us to reliablycalculate the bidirectional reflectance factor of vegetation, which in turn conveys information about the escaperatio of SIF photons. Our NIRV-based approach explains variations in the escape ratio with an R2 of 0.91 and anRMSE of 1.48% across a series of simulations where canopy structure, soil brightness, and sun-sensor-canopygeometry are varied. The approach is applicable to conditions of low leaf area index and fractional vegetationcover. We show that correcting for the escape ratio of SIF using NIRV provides robust estimates of total emittedSIF, providing for the possibility of studying physiological variations of fluorescence yield at the global scale.
1. Introduction
The empirical relationship between solar-induced chlorophyllfluorescence (SIF) and gross primary productivity (GPP) is complicatedby the fact that we only observe a fraction of emitted SIF photons andthat this fraction depends on the direction of observation (Porcar-Castell et al., 2014). It is difficult, therefore, to distinguish physiologicalvariations in the raw SIF signal from variations in SIF caused by ra-diative transfer processes. Our goal here is to use multispectral remotesensing observations to estimate the total amount of SIF emitted by thechlorophyll of a canopy and to explore the use of this measurement forinterpreting canopy scale photosynthesis and light capture.
Many of the challenges of SIF radiative transfer have already been
addressed in models (e.g., van der Tol et al., 2009). Such models pro-vide a platform for simulation of chlorophyll to leaf scaling of SIF(Ramos and Lagorio, 2006; Vilfan et al., 2016), the reabsorption of SIFwithin the leaf and canopy (Gitelson et al., 1998; Romero et al., 2018),the directionality of SIF emissions from the canopy (Hernández-Clemente et al., 2017; Zhao et al., 2016), and the atmospheric scat-tering of SIF (Frankenberg et al., 2011a). Yet there does not exist anaccepted, broadly applicable way to measure the fraction of the totalleaf-level SIF emission that makes its way to the top of the canopy and,ultimately, to the SIF sensor.
The challenge posed by these leaf-to-canopy scale radiative pro-cesses is best thought of from a physical standpoint, where SIF, as ob-
https://doi.org/10.1016/j.rse.2019.05.028Received 28 July 2018; Received in revised form 11 April 2019; Accepted 17 May 2019
⁎ Corresponding author at: Department of Global Ecology, Carnegie Institution for Science, Stanford, CA 94305, USA.E-mail address: [email protected] (Y. Zeng).
1 These authors contributed equally.
Remote Sensing of Environment 232 (2019) 111209
Available online 26 July 20190034-4257/ © 2019 Elsevier Inc. All rights reserved.
T

served at the top of canopy (SIFObs), is defined in terms of total emittedSIF (SIFTotal):
=SIF PAR fPAR( ) · · ( ),Total f chl F f (1a)
=SIF SIF f( , ) · ( , ).Obs f Totalesc
f (1b)
This formulation explicitly decouples the leaf-level input variablesthat generate SIF (Eq. (1a)) from the canopy radiative transfer processesthat govern the fraction of SIFTotal that ultimately escapes the canopyfor detection (Eq. (1b)). More specifically, SIFTotal, defined as the sum ofall SIF photons at a given wavelength (λf) emitted by all leaves withinthe canopy in all directions, depends on: i) PAR, photosyntheticallyactive radiation, ii) fPARchl, the fraction of PAR absorbed by chlor-ophyll, and iii) ΦF, the quantum yield of fluorescence. Upon emissionfrom the leaf, the photons which comprise SIFTotal are scattered throughthe canopy and only a fraction, f esc(Ω), ultimately escape the canopyand are observed at the view angle Ω. Eq. (1) makes clear that detectingmeaningful variations in physiology (e.g., changes in ΦF) from mea-surements of SIFObs requires first accounting for changes in f esc(Ω).
Quantifying f esc is a complicated task that requires three types ofinformation: i) sun-sensor geometry, ii) canopy structural parameters(e.g., leaf area index, clumping index, and leaf angle distribution), andiii) leaf optical properties (e.g., leaf reflectance/transmittance). Each ofthese parameters must be known to accurately describe the radiativetransfer of SIF. If these data are available, there are numerous physi-cally based forward modeling techniques capable of precisely calcu-lating f esc and thereby quantifying both SIFObs and SIFTotal (van der Tolet al., 2009; Hernández-Clemente et al., 2017; Zhao et al., 2016; Heet al., 2017). Despite the success of these efforts, the critical parametersfor directly calculating f esc are rarely well-characterized at the sitelevel, let alone at scales relevant to remote sensing. As a result, f esc istreated as constant (e.g., Guanter et al., 2014) or calculated by makinga priori assumptions that in turn introduce errors into the calculation off esc. In fact, only a handful of studies have seriously considered theeffects of f esc on SIF (e.g., Fournier et al., 2012; Migliavacca et al.,2017; van der Tol et al., 2016), though the subject has recently gainedmore attention (Liu et al., 2018; Yang and van der Tol, 2018).
Alternatively, it is possible to use the shape of the bidirectional re-flectance factor (BRF) derived from reflectance-based measurements tocalculate the angular distribution of SIF radiance and thereby calculateSIFTotal. BRF-based approaches rely on the fact that solar and SIF photons ofthe same wavelength are confronted by roughly the same canopy radiativeenvironment, despite originating from distinct physical processes. As a re-sult, it should be possible to use directional information from optical mea-surements to correct the directional dependencies of SIF. Liu et al., 2016used this approach and found a strong linearity between bidirectional SIFradiance and the total scene near-infrared bidirectional reflectance factor(BRFT). Similarly, Yang and van der Tol, 2018 showed that for near-infraredSIF, f esc can be described as a function of NIR reflectance, canopy direc-tional interceptance (i0), and leaf albedo (ω).
The BRFT approach for estimating f esc is both computationally andtheoretically simple, requiring few inputs outside of multi-spectral BRFTmeasurements (Yang and van der Tol, 2018). Despite these advantages, theBRFT approaches, like those developed by Liu et al., 2018 and Yang and vander Tol, 2018, implicitly assume that the contribution of photons scatteredby the soil is negligible. This assumption holds for SIF, as the SIF signal isspecific to vegetation; soil simply cannot emit SIF photons. However, theassumption of minimal soil influence does not hold for reflected sunlight, assolar photons are strongly scattered by the soil. As a result, the BRFT ap-proach for estimating f esc can only be used i) over dense canopies, whereboth the leaf area index (LAI) and fractional vegetation cover (FVC) are highor ii) where soil reflectance is low.
Here, we propose a new BRF-based approach for calculating f esc usingthe NIR reflectance of vegetation (NIRV). From a theoretical standpoint,NIRV represents the fraction of reflected NIR light that originates from ve-getation. In this sense, it attempts to replicate the specificity SIF has to
vegetation. In practice, NIRV minimizes the influence of soil reflectance onthe retrieved reflectance value (Badgley et al., 2017). This enables accurateretrievals of the contribution of vegetation to observed NIR reflectanceunder a wide array of field conditions, including over sparse canopies andregardless of soil brightness. As a result, NIRV better satisfies the assumptionthat soil only negligibly contributes to total scene reflectance and therebyprovides the basis for a computationally simple and physically-groundedapproach for determining the f esc of near-infrared SIF and, as a result, totalemitted SIF.
We introduce the approach in four parts, beginning first with anexplicit theoretical derivation relating NIRV and fPAR to f esc. Second,we use both a one-dimensional and a three-dimensional radiativetransfer model to demonstrate the robustness of our proposed model off esc regardless of solar angle, view angle, soil brightness, and canopystructure. Third, using satellite observations of SIF and NIR reflectance,we directly compare the relationship between SIF-NIRV and SIF-NIRTacross a wide range of ecosystem types to test the empirical usefulnessof both measurements as the basis for estimating f esc. Finally, we de-monstrate how NIRV and fPAR can be used to translate virtual satelliteobservations of SIF (SIFObs) into robust measurements of total emittedSIF (SIFTotal).
2. Motivation and theoretical derivation
Our approach for estimating SIFTotal from measurements of SIFObs(Eq. (1)) takes its motivation from the recent empirical result byBadgley et al., 2017, which showed that the near-infrared reflectance ofvegetation (NIRV) is strongly correlated with satellite-measurements ofSIFObs in the near-infrared (740–760 nm) range, which we denote hereas SIFN. Such a strong empirical correspondence between reflected lightand SIF indicates that the scattering of reflected near-infrared solarphotons and emitted near-infrared SIFN are strongly related, a resultthat has been separately demonstrated from radiative transfer modeling(van der Tol et al., 2016; Yang and van der Tol, 2018). In terms of thetask at hand, the correlation between SIFN and NIRV presents the pos-sibility of using NIRV as an independent basis for estimating f esc of SIFN.Note, the approach discussed for the remainder of the manuscript onlypertains to near-infrared measurements of SIF.
The intuition underlying the link between NIRV and SIFN is bestillustrated by considering NIRV as a corresponding radiance, NIR′V:
=SIF S i f· · · ,N F Nesc
400 700 0 (2a)
=NIR S i f· · · .V N Nesc
760 0 (2b)
Eq. (2b) represents the total NIR radiance reflected by the vegetatedcomponent of the land surface, which is a real, measurable flux. Ex-pressing NIRV in this way emphasizes that SIFN and NIR radiance arestrongly related to each other, as both are jointly dependent on the fluxof incoming solar radiation, denoted Sλf (S400-700, or PAR, in the case ofSIF; S760 for NIR′V), the fractional interceptance of vegetation (i0, de-pendent on canopy gap fraction), and the fraction of photons that es-cape from the canopy ( fN
esc). Eq. (2a) differs slightly from the definitionof SIF presented in Eq. (1a), as i0 is not strictly equal to fPARchl.However, as discussed below, the two are strongly related and writingEq. (2a) in terms of i0 emphasizes the similarity of SIF to NIR′V.
While similar, Eq. (2) makes it clear that SIFN and NIRV are notidentical. In particular, SIF and NIRV originate from different physicalprocesses within the canopy. SIF photons, on the one hand, are con-trolled by a biochemically mediated conversion of absorbed light tofluorescence, termed fluorescence yield and denoted as ΦF. NIRV, on theother hand, results from the scattering of solar photons by interactionwith leaves, making NIRV dependent on the leaf single scattering albedoin the NIR band (ωN). The difference in the origin of the SIF and NIRVsignals means that the f esc of SIF does not strictly equal the f esc of NIRV.This is because the contribution of any one leaf to the top of canopy SIFor NIRV signal depends on the value of ΦF or ωN of that leaf. More
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
2

specifically, at a given location in the canopy, f esc of SIF and NIRV areindeed the same (when neglecting higher order terms for SIF excita-tion). However, when averaging over the whole canopy, each leaf isweighted by its respective value of either ΦF or ωN, causing slight dif-ferences in the two canopy-level f esc terms. There are similar, smalldifferences in the penetration of solar PAR and NIR photons within thecanopy that drive both SIF and NIRV. Yet given the strong empiricalrelationship between SIFN and NIRV, as well as the small relativevariability of ΦF at the leaf-level (van der Tol et al., 2014), it stands toreason that such differences are negligible (though not ignorable).Thus, it should be possible to use NIRV to independently estimate f esc
and thereby correct for view-angle and within-canopy radiative transferprocesses that mediate the relationship between SIFObs and SIFTotal. Theremainder of this manuscript explores that possibility.
2.1. Calculating BRFT in the NIR band
We begin our derivation with the definition of the total scene bi-directional reflectance factor at wavelength λf (BRFT(λf)), which can bebroken down as a linear combination of three parts:
= + +BRF BRF BRF BRF( ) ( ) ( ) ( ).T f V f S f M f (3)
The three components of BRFT are: i) BRFV, the contribution of photonsat wavelength λf reflected off only the scene's vegetative component, ii)BRFS, the contribution of photons at wavelength λf reflected off soilalone, and iii) BRFM which represents the contribution to BRFT frommultiple scattering between the vegetation and soil.2 In practice,however, satellites are only capable of measuring BRFT, as satellitescapture light reflected by vegetative and non-vegetative elements alike.For studying vegetation, we are primarily interested in BRFV, whichdescribes how viewing geometry, canopy structure, and leaf spectralproperties influence the scattering of both solar (reflected) and SIF(emitted) photons within the canopy.
More formally, BRFV can be described as:
=BRF i f( ) · · ,V fesc
0 f f (4)
where i0 is canopy interceptance, which represents the probability of asolar photon (of any wavelength) interacting with the canopy and isdefined as one minus the directional gap fraction (Smolander andStenberg, 2005), ωλf is the leaf single scattering albedo at wavelengthλf, and f esc
fis the fraction of photons at wavelength λf that escape the
canopy and are detected remotely (Knyazikhin et al., 2013). Yetwithout prior information on BRFS and BRFM, the task of disentanglingBRFV from BRFT becomes challenging (Eq. (3)).
For the problem at hand, we are interested in isolating the BRFV inthe NIR region, as this is the portion of BRFT that contains relevantinformation about f esc. For convenience, we rewrite Eq. (3) to explicitlyconsider the NIR case as:
= + +NIR NIR NIR NIR .T V S M (5)
In the most straightforward terms, we are interested in calculating thefraction of NIRT that originates from interactions with vegetation alone.We can call this fraction FV. By extension, it is possible to write:
+ + =F F F 1,V S M (6)
meaning that reflectance from vegetation, soil, and the multiple scat-tering between the two constitute the full NIRT signal. For the purposeof simplifying the remainder of the derivation, we treat FM as 0, thoughsubsequent radiative transfer experiments contained within thismanuscript make no such assumption.
With Eqs. (5) and (6) in hand, we can write:
==
NIR F NIRNIR F NIR
·· .
V V T
S S T (7)
The challenge now becomes finding an appropriate means of estimatingNIRV from measurements of NIRT. When framed in this way, the chal-lenge posed by Eq. (7) closely parallels the approach used to estimateSIF from remote sensing, which ultimately hinges on determining thefraction of SIF contained within measurements of NIRT.
One theoretical solution to isolating NIRV from NIRT was formulatedby Knyazikhin et al., 1998 in what they termed the “black soil pro-blem.” The black soil problem states that calculating NIRV (or BRFV atany wavelength, for that matter) would be trivial if the canopy wasilluminated from above and, instead of being backed by a soil with itsown reflectance properties, the soil was replaced by a perfectly absor-bent (black) background. In this scenario, both NIRS and NIRM becomezero and the retrieved NIRT signal can be safely attributed to vegeta-tion. While such a substitution is impossible in practice, it may bepossible to use radiative transfer simulations to identify optical-basedvegetation indices that behave similar to NIRT under black soil condi-tions.
2.2. An approximation of NIRV
Here, we explore an approach for approximating NIRV first de-scribed by Badgley et al., 2017:
NIR NDVI NIR· ,V T (8)
where NDVI is the normalized difference vegetation index, which de-pends on red (630–690 nm) and NIR reflectance (750–900 nm) (Tucker,1979). Eq. (8) makes it clear that the product of NDVI and NIRT is anapproximation of the proportion of reflected photons that directly in-teract with vegetation, which is a real, physical property of the scene. Itis also possible to calculate NIR′V (radiance) as NDVI ·NIR′T, however, weonly consider the reflectance case here. More specifically, NDVI ·NIRTserves as a reliable proxy of BRFV in the NIR band (NIRV), which can bemore succinctly expressed by combining Eq. (4) with Eq. (8):
NDVI NIR NIR i f· · · .T V N Nesc
0 (9)
There are two important messages to take from Eqs. (8) and (9).First, NDVI ·NIRT is one possible approach for approximating NIRV.Intuitively, the approach works because both NIRT and NDVI vary withsoil brightness, but in an opposite manner: darker soils have a higherNDVI but lower NIRT, while brighter soils have a lower NDVI but higherNIRT (Qi et al., 1994). As the fraction of vegetation cover approachesunity, NDVI also approaches unity and the product reduces to NIRT.Note that because NDVI does not necessarily span between 0 and 1, theapproach underestimates the true value of BRFV(NIR) for high fractionsof vegetation cover (Fig. 1; though these effects can be further miti-gated, see Discussion). Recognizing that there might exist alternativeapproaches for estimating NIRV, for purposes of both clarity andbrevity, from this point forward we will refer to the product of NDVIand NIRT as simply “NIRV.” We will contrast this approximation againstBRFV(NIR), which from this point will signify the true value of the BRFof vegetation.
Second, and more importantly for the derivation at hand, Eq. (9)suggests that NIRV (as approximated by NDVI ·NIRT) closely approx-imates the black soil condition imagined by Knyazikhin et al., 1998,providing a reliable proxy for estimating BRFV(NIR). The validity of thisclaim is immediately testable using SCOPE (van der Tol et al., 2009),which allows the simulation of canopy radiative transfer via an im-plementation of PROSAIL (Baret et al., 1994; Jacquemoud and Baret,1990; Verhoef, 1984). Using SCOPE, we allowed solar and view angles,soil brightness, and canopy structure to vary randomly across their fullvalid ranges (Table 1). The R2 between NIRV and BRFV exceeded 0.99across 5040 random combinations of input variables (Fig. 1). By con-trast, NIRT shows wide dispersion against BRFV at lower levels of2 Please see the Glossary for a full listing of terms and abbreviations.
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
3

vegetation cover, with gradual convergence of NIRT and BRFV as ve-getation comprises a larger fraction of the scene (Fig. S1).
2.3. Measuring fesc remotely
Eq. (9) can then be rearranged to calculate the common term, fNesc,shared between NIRV and SIFN:
f NIRi ·
.Nesc V
N0 (10)
Using Eq. (10) in practice requires two additional assumptions. First,ωN, the NIR leaf albedo, is relatively conservative both within andacross species (Gates et al., 1965; Asner, 1998) and, for purposes of thisderivation, we assume ωN takes a constant value of 1. Second, calcu-lating i0 requires additional knowledge of canopy architecture, in-cluding the leaf projection function (G(Ω); Ross, 1981) and theclumping index (CI; Chen, 1996). While both G(Ω) and CI are mea-surable quantities, both strongly vary in time and space, making themdifficult to quantify remotely (Raabe et al., 2015; Ryu et al., 2010).
Instead, we make the further simplifying assumption that i0 is ap-proximated by canopy fPAR, the fraction of absorbed photo-synthetically active radiation (PAR). PAR is strongly absorbed by plantcanopies; leaf albedo within the visible range (ωV) is small and, for ourpurposes, can be assumed to approach 0 (Gates et al., 1965). Following
Stenberg et al. (2016) and assuming ωV=0, we can explicitly relate i0to fPAR via canopy absorbance of visible light (ABSV):
=
=
fPAR ABS
ip
i
11
V
V
V0
0 (11)
Here, p represents the probability of a photon recolliding with the ca-nopy after its initial (non-absorbing) interaction with the canopy (seeAppendix). For clarity, Eq. (11) treats the soil as black, ignoring theminor contribution of solar photons reflected by the soil that are ulti-mately absorbed by vegetation. This assumption, however, is not in-voked at any point in our analysis, including the validation of Eq. (11)below. When evaluated using SCOPE, Eq. (11) holds across a wide arrayof conditions, where both LAI and FVC were varied (see Materials andmethods). For all combinations of LAI and FVC, fPAR is strongly cor-related with i0, having an the RMSE of 0.03, and an R2 exceeding 0.99,demonstrating that fPAR closely approximates i0 (Fig. 2). Furthermore,while the assumption that ωV=0 is useful for deriving the relationshipbetween f esc and NIRV, this assumption is not invoked in the radiativetransfer simulations used to validate our approach. It is also worthpointing out that while NDVI (used to approximate NIRV) and fPAR arerelated to each other, their relationship is not strictly linear due to thefact that canopy architecture (e.g., leaf inclination angle, CI) has un-equal effects on NDVI and fPAR (Fig. S2).
Together with our assumption that ωN=1, we can then rewrite Eq.(10) to read:
f NIRfPAR
,Nesc V
(12)
which is both computationally tractable and easily estimated from ex-isting in situ and remote sensing measurements. A more detailed deri-vation relating f esc of SIF to f esc of NIR reflectance is presented in theAppendix.
3. Materials and methods
3.1. Testing NIRV as the basis for estimating fesc
We evaluated Eq. (12) using both a one-dimensional (1-D) and athree-dimensional (3-D) radiative transfer model. Radiative transfermodels enabled us to exhaustively test numerous parameters that affect
Fig. 1. NIRV closely approximates BRFV, the vegetative component of pixelBRF, across a wide array of fractional vegetation cover, soil backgrounds, andleaf angle distributions. NIRV underestimates low values of BRFV and over-estimates higher values of BRFV. All values generated using SCOPE, withparameters drawn from ranges shown in Table 1.
Table 1Parameters varied in SCOPE simulations. Default values for SCOPE v1.70 wereused for all other parameters.
Variable Values
Canopy structure Leaf Area Index [0.5, 1, 3, 5]
Leaf Angle Distribution Spherical, Erectophile,Planophile
Sun-sensor geometry Solar Zenith Angle [20, 30, 40, 50, 60]
View Zenith Angle [0, 10, 20, 30, 40, 50, 60]
Soil Background Soil Spectra Four soil spectrum
Leaf Spectral Properties Leaf ChlorophyllContent (Cab)
[40, 60, 80]
Fig. 2. fPAR is a good approximation of canopy directional interceptance, i0,across canopies with varying leaf area and fraction of vegetation cover. Allvalues generated using SCOPE, with parameters drawn from ranges shown inTable 1.
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
4

f esc, allowing for a more comprehensive evaluation of Eq. (12) thanwould be possible using in situ measurements alone. For both models,our general approach was to randomly vary key parameters that affectf esc, including leaf-level spectral properties (e.g., chlorophyll content),leaf area index (LAI), leaf angle distribution (LAD), and soil brightness.
3.1.1. SCOPE: 1-D radiative transfer simulationsWe began our evaluation of Eq. (12) using the Soil Canopy Ob-
servation, Photochemistry and Energy (SCOPE) model, which is capableof simulating radiative transfer, energy balance, and photosynthesis, aswell as the SIF of individual leaves within the canopy and total emittedSIF across the full spectrum of chlorophyll fluorescence (van der Tolet al., 2009). Using SCOPE, we conducted a set of simulations where werandomly varied parameters affecting both f esc and scene NIR re-flectance. In particular, we randomly varied leaf chlorophyll content(Cab), LAD, LAI, the reflectance profile of soil, view zenith angle, andsolar zenith angle (Table 1). For each simulation, we compared simu-lated f esc against f esc as derived in Eq. (14), NIR
fPARV . For reference, we also
compared the NIRV-based approach against the NIRT approach pro-posed in Eq. (12) of Yang and van der Tol, 2018, which specifies f esc as:
=f NIRi·
.esc T
N 0 (13)
We further examined variations in FVC using a simple linear spectralmixture model, with the end members of i) vegetation, with valuesderived from SCOPE with an LAI of 3, and ii) bare soil. The spectralproperties of each scenario were calculated by the area proportional toeach end member, with FVC ranging between 0.1 and 1.
All SCOPE simulations were conducted using version 1.70 of themodel. For each simulation, directional and total emitted SIF wereextracted at 760 nm. NIRV was calculated using red reflectance at648 nm and NIR reflectance at 858 nm to be consistent with the red andNIR bands of the Moderate Resolution Imaging Spectroradiometer(MODIS) for future practical use, though it is worth noting that the NIRof vegetation has little variation from 760 to 900 nm (Gates et al.,1965). We used the SCOPE canopy gap fraction variable, gap.Ps, tocalculate i0 as 1− gap.Ps. Finally, we calculated BRFV from SCOPE foreach simulated condition by keeping all parameters equal, but insertinga black soil background.
3.1.2. DART: 3-D radiative transfer simulationsWe conducted a similar experiment testing the validity of Eq. (12)
using the Discrete Anisotropic Radiative Transfer (DART) model(Gastellu-Etchegorry et al., 2004). DART allows the simulation of lightwithin the optical domain and, due to recent developments described inGastellu-Etchegorry et al., 2017, DART is also capable of modeling SIF.Unlike SCOPE, which operates under the assumption of a horizontallyhomogeneous canopy, DART provides a more realistic representation ofpartial vegetation cover. As a result, DART provides a useful tool fortesting the relationship between NIRV and BRFV under conditions ofsparse vegetation cover.
Due to the computational demands of DART, we focused our at-tention on varying soil properties and viewing geometry across DARTruns. Leaf area, solar zenith angle, and chlorophyll content of the
canopy were held constant across all runs (Table 2). We instead con-centrated on varying soil brightness, using three separate soil spectraincluded in DART (Fig. S3). 3-D renderings of the simulated canopy areincluded in the supplementary materials (Fig. S4). DART directly cal-culates both red and NIR reflectance, from which we calculated NIRV.DART also simulates observed SIF (SIFObs), but does not provide readyaccess to leaf-level estimates of SIF, meaning we were unable to cal-culate SIFTotal and f esc of SIF.
Instead, we compared simulated NIRV under various soil brightnessconditions against BRFV, which we calculated by running a second setof DART simulations where the soil background was assumed to beblack. These simulations allowed us to evaluate the performance ofNIRV and NIRT for estimating BRFV, as outlined in Eq. (9). Recall thatBRFV comprises three parts: i0, ωN, and f esc.As a result, the utility ofNIRV as a predictor of f esc can be assessed by examining the linearity ofNIRV-BRFV relationship, when evaluating parameter combinations thatmainly affect f esc. Similarly, we directly evaluate the SIF-NIRT and SIF-NIRV relationship using data from the DART simulations. As with theSCOPE simulations, SIFObs was calculated at 760 nm, while NIRV wascalculated using reflectance red reflectance at 648 nm and NIR re-flectance at 858 nm. For continuity with Yang and van der Tol, 2018,NIRT was calculated at 760 nm.
3.2. Empirical evaluation of NIRV compared to NIRT
We performed an empirical evaluation of NIRV as the basis for es-timating f esc by directly comparing NIRV against measurements of SIF.Such a comparison allowed empirical validation of the SIF-NIRV re-lationship, with the goal of demonstrating that NIRV does indeed ap-proximate the “black soil” condition. Previously, Liu et al., 2018 andYang and van der Tol, 2018 proposed using NIRT for estimating f esc.However, both studies invoked the black soil assumption, assumingperfect absorption of solar photons by the soil background. Our deri-vation of f esc and the simulation results that follow make no such as-sumption. Combining our model-based results with an empirical com-parison of NIRV and NIRT offers a more comprehensive evaluation ofhow the differing approaches operate in practice.
We combined measurements of NIRV and NIRT from MODIS withnear-infrared SIF retrievals from the newly launched TROPOMI sensor(Köhler et al., 2018). Specifically, the proposed joint dependency of SIFand NIRV on i0 and fN
esc (as illustrated by Eq. (2)) can be more simplyexpressed and evaluated by exploring the proportionality of SIF to NIRVusing actual remote sensing measurements:
SIF NIR .V (14)
Black soil does not exist under real measurement conditions, meaningEq. (14) can be directly contrasted against the proportionality of SIF toNIRT:
SIF NIR .T (15)
If NIRT is sufficient for calculating f esc in non-idealized situations, thereshould be no significant difference between the SIF-NIRT and the SIF-NIRV relationship. Such a comparison provides an empirical test of thedifferences of NIRT and NIRV in approximating the “black soil condi-tion”, as opposed to specifically testing various techniques for esti-mating f esc that require additional inputs (e.g, the approach to esti-mating f esc proposed by Yang and van der Tol, 2018 requires i0 as aninput).
To perform the comparison, we selected five sites with variousfractions of vegetation cover from across North America. We then ex-tracted daily near-infrared SIF retrievals from the cloud filtered andglobally gridded TROPOMI SIF dataset described by Köhler et al., 2018,which has a spatial resolution of 0.2∘. We combined SIF data with dailyNIRV and NIRT observations from the nadir-corrected MODIS re-flectance product MCD43A4v6 regridded to 0.2∘ (Schaaf et al., 2002).MODIS data were used, rather than radiances from TROPOMI because
Table 2Parameters varied in DART simulations. Default values were used for all otherparameters.
Variable Values
Canopy structure Leaf Area Index 2Sun-sensor geometry Solar Zenith Angle 30∘
View Zenith Angle 0∘–70∘ with step of 5∘
Soil background Soil Spectra Three soil spectrum (Fig. S3)Leaf spectral properties Leaf Chlorophyll Content 58
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
5
atmospherically corrected TROPOMI radiance data at the necessarywavelengths are not presently available. This introduces angular dif-ferences between the TROPOMI and MODIS data, though such dis-crepancies do not influence the inference derived from comparing SIFagainst NIRT and NIRV. Furthermore, the majority of TROPOMI SIFretrievals have a phase angle that falls between 20∘ and 60∘ (Köhleret al., 2018), while the MODIS data has been adjusted to nadir. Thesetwo facts combine to mitigate the severity of the disagreement betweenSIF and reflectance measurements that are solely attributable toviewing geometry. To facilitate comparison between flux measurementsof SIF and unitless measurements of reflectance, we normalized SIFobservations by at-sensor solar radiance. This ensured that the observedSIF-NIR relationships were not driven by an underlying PAR-NIR re-lationship. Together, these data allow direct evaluation of Eqs. (14) and(15). MODIS data were processed using the Google Earth Engine PythonAPI. TROPOMI data were downloaded from ftp://fluo.gps.caltech.edu/data/tropomi/.
3.3. Correcting virtual satellite retrievals
Finally, we used SCOPE to construct a virtual experiment to de-monstrate the practical utility of using NIRV-derived estimates of f esc toestimate SIFTotal from directional-dependent simulations of SIFObs. Inrecent years, SIF has emerged as a useful remote measurement of APAR(Yang et al., 2018; Du et al., 2017) and GPP (Frankenberg et al., 2011b;Guanter et al., 2014; Smith et al., 2018; Yang et al., 2015; Zhang et al.,2016; Sun et al., 2017) at scales ranging from individual study sites tothe globe. Many of these studies, however, rely on using fixed scalingfactors between SIFObs and the variable of interest. These scaling fac-tors, however, often ignore variations in f esc that affect the value ofSIFObs. Failure to account for f esc makes it difficult to study variations inΦF at the global scale and potentially influences the interpretation ofcanopy- and global-scale relationships between SIF, GPP, and APAR.
To demonstrate this effect and explore the usefulness of Eq. (12) forcalculating SIFTotal from measurements of SIFObs, we extracted solar andviewing geometry data from the Global Ozone Monitoring Experi-ment–2 sensor (GOME-2; a widely used SIF sensor) for a single location.This allowed us to vary the solar and view angle parameters in SCOPEto match local conditions, as if the site were viewed by GOME-2(Table 3). We then calculated the relative error introduced in calcu-lating SIFTotal from SIFObs using NIRV and Eq. (12). We also evaluatedthe relative error of assuming that any one value of SIFObs has a con-stant scaling with SIFTotal and the relative error in estimating SIFTotalfrom NIRT-derived estimates of f esc, following the approach of Yang andvan der Tol, 2018 (Eq. (13)). Along with varying solar angle and viewangle, we also simulated various values of FVC ranging from 0.1 to 1.
4. Results
4.1. SCOPE Simulations of fesc
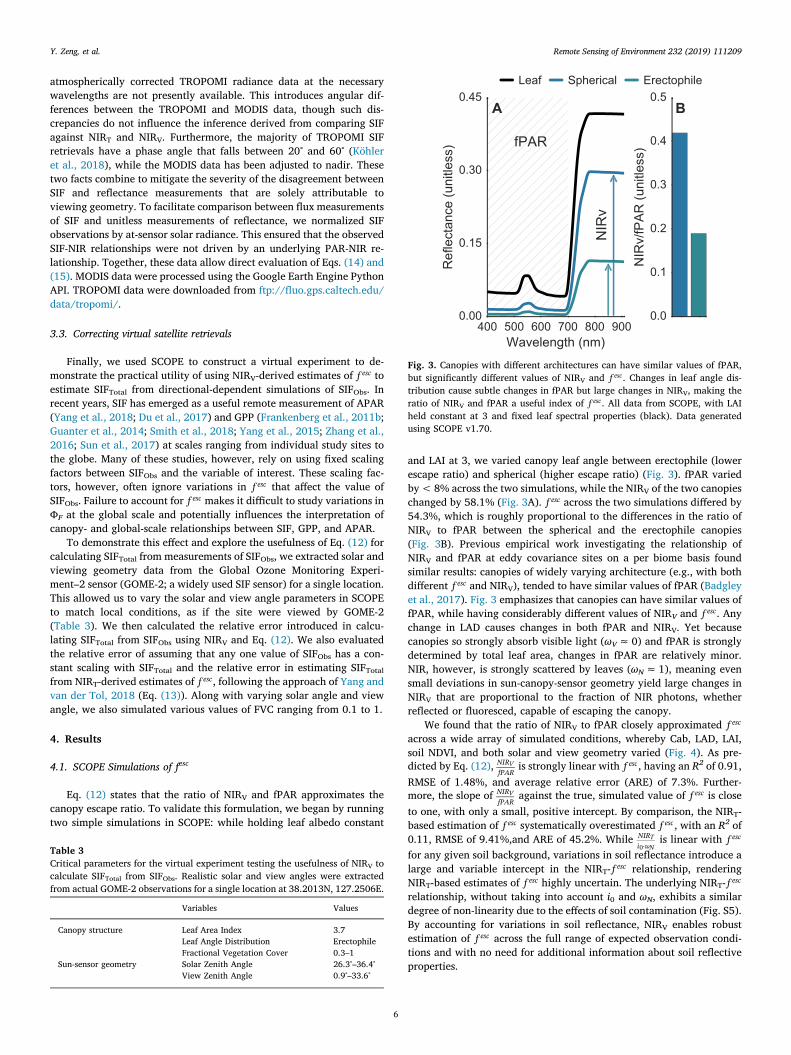
Eq. (12) states that the ratio of NIRV and fPAR approximates thecanopy escape ratio. To validate this formulation, we began by runningtwo simple simulations in SCOPE: while holding leaf albedo constant
and LAI at 3, we varied canopy leaf angle between erectophile (lowerescape ratio) and spherical (higher escape ratio) (Fig. 3). fPAR variedby< 8% across the two simulations, while the NIRV of the two canopieschanged by 58.1% (Fig. 3A). f esc across the two simulations differed by54.3%, which is roughly proportional to the differences in the ratio ofNIRV to fPAR between the spherical and the erectophile canopies(Fig. 3B). Previous empirical work investigating the relationship ofNIRV and fPAR at eddy covariance sites on a per biome basis foundsimilar results: canopies of widely varying architecture (e.g., with bothdifferent f esc and NIRV), tended to have similar values of fPAR (Badgleyet al., 2017). Fig. 3 emphasizes that canopies can have similar values offPAR, while having considerably different values of NIRV and f esc. Anychange in LAD causes changes in both fPAR and NIRV. Yet becausecanopies so strongly absorb visible light (ωV≈0) and fPAR is stronglydetermined by total leaf area, changes in fPAR are relatively minor.NIR, however, is strongly scattered by leaves (ωN≈1), meaning evensmall deviations in sun-canopy-sensor geometry yield large changes inNIRV that are proportional to the fraction of NIR photons, whetherreflected or fluoresced, capable of escaping the canopy.
We found that the ratio of NIRV to fPAR closely approximated f esc
across a wide array of simulated conditions, whereby Cab, LAD, LAI,soil NDVI, and both solar and view geometry varied (Fig. 4). As pre-dicted by Eq. (12), NIR
fPARV is strongly linear with f esc, having an R2 of 0.91,
RMSE of 1.48%, and average relative error (ARE) of 7.3%. Further-more, the slope of NIR
fPARV against the true, simulated value of f esc is close
to one, with only a small, positive intercept. By comparison, the NIRT-based estimation of f esc systematically overestimated f esc, with an R2 of0.11, RMSE of 9.41%,and ARE of 45.2%. While NIR
i ·TN0is linear with f esc
for any given soil background, variations in soil reflectance introduce alarge and variable intercept in the NIRT- f esc relationship, renderingNIRT-based estimates of f esc highly uncertain. The underlying NIRT- f esc
relationship, without taking into account i0 and ωN, exhibits a similardegree of non-linearity due to the effects of soil contamination (Fig. S5).By accounting for variations in soil reflectance, NIRV enables robustestimation of f esc across the full range of expected observation condi-tions and with no need for additional information about soil reflectiveproperties.
Table 3Critical parameters for the virtual experiment testing the usefulness of NIRV tocalculate SIFTotal from SIFObs. Realistic solar and view angles were extractedfrom actual GOME-2 observations for a single location at 38.2013N, 127.2506E.
Variables Values
Canopy structure Leaf Area Index 3.7Leaf Angle Distribution ErectophileFractional Vegetation Cover 0.3–1
Sun-sensor geometry Solar Zenith Angle 26.3∘–36.4∘
View Zenith Angle 0.9∘–33.6∘
Fig. 3. Canopies with different architectures can have similar values of fPAR,but significantly different values of NIRV and f esc. Changes in leaf angle dis-tribution cause subtle changes in fPAR but large changes in NIRV, making theratio of NIRV and fPAR a useful index of f esc. All data from SCOPE, with LAIheld constant at 3 and fixed leaf spectral properties (black). Data generatedusing SCOPE v1.70.
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
6

4.2. DART simulations of BRFV
A second set of simulations using the 3-D radiative transfer modelDART demonstrates the generality of the NIRV-BRFV relationship whenmore realistic radiative transfer processes are taken into account(Fig. 5). Because DART does not generate per-leaf estimates of SIF, weinstead evaluated the suitability of NIRV as the basis for estimating f esc
by comparing NIRV and NIRT against BRFV, following Eq. (9). While soilbrightness does introduce a bias into the estimation of BRFV by NIRV,the NIRV-BRFV relationship has a small RMSE of 0.026, a near-zerointercept, and a slope of 0.89 (Fig. 5A). Variations in soil backgroundresult in a weaker relationship between NIRT and BRFV (Fig. 5B). Asfound in the SCOPE simulations, NIRT is mostly linear with BRFV forany given soil brightness. However, variations in the soil backgrounddegrade the overall proportionality of NIRT to BRFV, resulting in aRMSE of 0.107, an intercept of 0.11, and a slope of 0.4. As a result,using NIRT to derive f esc would require a priori information about soilspectral properties (to account for the influence of NIRS, see Eq. (7)).NIRV has a more robust relationship BRFV and f esc that does not requireexplicit consideration of soil background reflectance.
NIRV also had a more consistent relationship with DART simulationsof SIFObs, when compared to NIRT (Fig. 6). The SIF-NIRV relationshiphas a small RMSE of 0.004 and a near-zerointercept of 0.01. Intrigu-ingly, the SIF-NIRT relationship is substantially stronger (R2=0.85)than the NIRT-BRFV (R2=0.42) relationship shown in Fig. 5. This
seemingly contradictory result has to do with the fact that for a dis-continuous canopy backed by a bright soil background, accurately es-timating f esc of SIF requires consideration of the interaction of SIFphotons with the soil background (BRFM from Eq. (3)). Under theseconditions, BRFM strongly contributes to the fraction of SIF photons thatultimately escape the canopy. In the case of a mostly uniform (e.g.,BRFS is small due to the spatial distribution of vegetation relative to thesoil background, Fig. S4), albeit discontinuous canopy, both NIRV andNIRT are capable of capturing the contribution of BRFM to f esc. How-ever, NIRV has still has a stronger relationship with SIF because it is lessaffected by BRFS. As a result, following Eqs. (14) and (15), NIRV offers amore robust starting point for estimating f esc of SIF. Finally, Fig. 6 canbe thought of as a best-case scenario for the performance of NIRT over adiscontinuous canopy. As the canopy becomes less uniform (e.g.,clumped in space) and BRFS increases, the insensitivity of NIRV to thesoil background results in the SIF-NIRV relationship remaining stronglylinear (R2=0.83), while the SIF-NIRT relationship rapidly weakens andbecomes non-linear (R2=0.62; Fig. S6).
4.3. Empirical differences between NIRV and NIRT
Satellite measurements of NIRV and SIF confirmed that NIRV mini-mizes the effects of variation in soil background, whereas the SIF-NIRTrelationship is highly non-linear for sparse vegetation types (Table 4;see Fig. S7 for individual site scatter plots). These data represent a
Fig. 4. A) The ratio of NIRV to fPAR closely approximates f esc, while B) f esc calculated as NIRTi N0·
, following Yang and van der Tol, 2018, is largely unrelated to modeled
f esc across the same range of conditions. All data produced using SCOPE, varying LAD, LAI, soil NDVI, view zenith angle, and solar zenith angle at random.
Fig. 5. 3-D radiative transfer simulations of the A) NIRV-BRFV and B) NIRT-BRFV relationships from DART. NIRV is strongly linear with BRFV across varying soilbackgrounds, demonstrating the usefulness of NIRV as the basis for estimating f esc, regardless of vegetation density.
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
7

direct test of Eqs. (14) and (15), as NIRV and NIRT must be proportionalto SIF if either measurement is to be useful as the basis for estimatingf esc (Eq. (2)). For each site, we calculated the difference in the coeffi-cient of determination (R2) between radiation normalized SIF, NIRV,and NIRT. While NIRT and radiation normalized SIF show strongagreement over dense, continuous canopies, like crops and broadleafforests (ΔR2=−0.01 and 0.11, respectively), the relationship is con-siderably weaker over sparse vegetation canopies. For biomes likesparse woodlands, NIRV has a substantially stronger and more linearrelationship with radiation normalized SIF than NIRT (ΔR2=0.53),though the SIF-NIRV relationship did break down at the grassland site.Interestingly, NIRV even outperformed NIRT at US-Ha1 (HarvardForest), a dense, mostly closed canopy deciduous broadleaf forestwhere soil contamination is expected to be minimal. However, early inthe season, before leaf flush, soil is readily visible from the satelliteperspective, which causes a divergence in NIRT from measured SIF (Fig.S7). Thus, even though NIRT is highly correlated with SIF for parts ofthe season, NIRV has a more consistent, inter-seasonal relationship withSIF. Taken together, these results nicely illustrate that NIRV, as definedin Eq. (9), does a better job of approximating the “black soil condition”,whereas soil contamination renders NIRT a less suitable candidate asthe basis for calculating f esc. NIRV also outperforms the SIF-NDVI re-lationship at these sites, further demonstrating the utility of combiningNDVI and NIRT together (Fig. S7).
4.4. Correcting virtual satellite retrievals
Accurate estimates of f esc allow the conversion of directional mea-surements of SIFObs into estimates of total emitted SIF, SIFTotal. Changesin both solar and viewing geometry cause variations in SIFObs that areunrelated to differences in the true value of total emitted SIF. Ideally,SIFObs should be converted to SIFTotal to serve as a common basis ofestimating canopy APAR and, potentially, gross primary production.When we ran SCOPE using view angles taken from actual GOME-2
satellite retrievals for a single site, variations in viewing geometry re-sulted in upwards of 70% variation in SIFObs (Fig. 7A). Naively relatingany one value of SIFObs to total emitted SIF yielded a± 30% relativeerror in estimated SIFTotal (Fig. 7B). Such variations have no direct linkto physiology and, if not accounted for, significantly bias attempts touse SIF to infer canopy-scale and leaf-level physiology (e.g., ΦF),especially across canopies with dissimilar architectures. By contrast,SIFTotal calculated from NIRV-derived estimates of f esc has a maximumrelative error of± 4.9% and ARE of 2.2% (Fig. 7C). Such minimal er-rors should allow SIF measurements to be more accurately comparedacross space and time to draw physiological inferences.
NIRV provides a robust estimate of f esc across all values of FVC,resulting in accurate estimation of SIFTotal even under sparsely vege-tated conditions (Fig. 8). NITT, on the on the other hand, is stronglyaffected by soil reflectance, resulting in errors exceeding 50% in esti-mated SIFTotal for low values of FVC. It is only under relatively highvalues of FVC (e.g., > 0.8) that NIRT-derived estimates of SIFTotal are onaverage more accurate than estimates of SIFObs alone. The average re-lative error of directly scaling any one directional value of SIFObs toSIFTotal remains constant at roughly 20% across all simulated values ofFVC because SIF is not influenced by soil background reflectance.Average relative error of NIRV-based estimates of SIFTotal falls wellbelow 10% for all values of FVC examined. Interestingly, the averagerelative error of NIRV-based SIFTotal was lower than NIRT-based esti-mates for all values of FVC, including when FVC reached unity. Thisresult demonstrates the usefulness of NIRV for both sparsely and denselyvegetated scenes.
5. Discussion
We demonstrated that the ratio of NIRV to fPAR is an accurate ap-proximation of the escape ratio of near-infrared SIF photons, providinga theoretically-grounded and computationally tractable approach fortranslating directional-based SIF measurements to comparable, whole-
Fig. 6. 3-D radiative transfer simulations of the A) SIF-NIRV and the B) SIF-NIRT relationship from DART. NIRV is less sensitive to variation in the soil background andhas a stronger linear relationship with simulated SIF. Reflectance profiles of each soil are provided in Fig. S3.
Table 4NIRV has a stronger, more consistent relationship with daily measurements of SIF than NIRT across multiple biomes. Daily MODIS reflectance values were comparedagainst daily, at-sensor radiance normalized SIF measurements from the TROPOMI sensor for the period spanning March to August 2018.
Site(Ameriflux Code)
Lat./Lon. Biome SIF-NIRVR2
SIF-NIRTR2
(NIRV−NIRT)ΔR2
US-Ha1 42.52, −72.17 Broadleaf Forest 0.72 0.61 0.11US-CZ1 37.11, −119.73 Oak/Pine Woodland 0.71 0.18 0.53US-SCg 33.74, −117.69 Grassland 0.24 0.02 0.22US-Ho1 45.20, −68.74 Evergreen Forest 0.70 0.16 0.54US-Ne1 41.17, −96.47 Cropland 0.87 0.88 −0.01
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
8

canopy estimates of total emitted SIF. Importantly, the approach re-quires minimal assumptions and can be calculated using widely avail-able optical remote sensing data. Our approach is immediately applic-able to in situ studies of SIF and a wide range of satellite-based SIFplatforms, including the newly launched TROPOMI sensor (Veefkindet al., 2012) and the upcoming FLEX mission (Drusch et al., 2017), bothof which have access to simultaneous or near simultaneous measure-ments of reflectance and SIF, which allows for per-sounding f esc ad-justments by NIRV. Our approach is also useful for in situ studies of SIF,where diurnal shifts in sun-sensor geometry can significantly influenceSIFObs and obscure underlying physiologyical changes in ΦF (Fig. 7).
To date, there have only been a handful of studies explicitly focusedon the escape ratio of SIF, including attempts to correct for it. Romeroet al., 2018 proposed an approach based on canopy reflectance, trans-mittance, and soil reflectance to estimate the spectral shape of SIFemitted at the leaf-level. While useful, the dependence on priorknowledge of soil reflectance and canopy transmittance limits thepractical application of the approach. Liu et al., 2018 took a purelystatistical approach, combining machine learning with reflectance-based inputs to infer f esc of both red and near-infrared SIF. While theirapproach performed well for even low LAI values, the approach wepropose here using NIR
fPARV is both mechanistically and computationally
simpler. In an independent study, Yang and van der Tol, 2018 showedthat NIR
i ·TN0was a good approximation f esc under the black soil condition.
Our work directly extends this concept by demonstrating that NIRVclosely approximates the black soil condition and is readily applicablein real-world remote sensing applications (Table 4).
5.1. Canopy spectral invariants
Our approach builds on recent advances in canopy radiative transfertheory, especially the development of spectral invariant properties (SIP;Huang et al., 2007, Knyazikhin et al., 2013, Stenberg et al., 2016). SIPdramatically reduce the complexity of characterizing whole-canopyradiative processes, breaking down the radiative transfer process intothree main components: i0, the probability a photon interacts with ve-getation; ρn, the probability a photon escapes the canopy on its n-thcollision; and pn, the probability a photon, after its n-th recollision,recollides with vegetation yet again. Together, i0, ρ, and p can becombined to characterize even the most complex radiative environ-ments, across all wavelengths of light. In many ways, the developmentof SIP is akin to the development of leaf-level spectral models, wherebydetailed knowledge of the internal complexity of the leaf (e.g., chlor-ophyll content, mesophyll density) and its effect on radiative transfer,can instead be characterized by only a few parameters (e.g.,Jacquemoud and Baret, 1990). SIP enables the same reduction incomplexity at the scale of the entire canopy (Stenberg et al., 2016).
Our results demonstrate that NIRV serves as a new tool for capturingSIP parameters from remote sensing. Indeed, the term f esc can be ex-plicitly expressed in terms of both ρ and p (see Appendix). Given thatNIRV also relates to i0 (Eq. (9)), NIRV can in principle serve as the basisfor individually retrieving i0, ρ, and p. Fully separating the influence ofeach parameter, however, likely requires use of multi-angular data andperhaps some ancillary data (e.g., fPAR, leaf spectral properties). In themeantime, the fact that NIRV captures the effects of i0, ρ, and p is suf-ficient for accurately characterizing the canopy radiative environmentand estimating SIFTotal from measurements of SIFObs. Our formulationof using NIRV to calculate f esc might also find use in process-basedecosystem models, by way of simplifying canopy radiative transferprocesses. Many of these models already simulate or take as inputs allthe variables needed to estimate f esc from NIRV. One potential use ofthese data suggested by our analysis would be explicitly scaling mod-eled estimates of SIFTotal to estimates of SIFObs for comparison withground-based and satellite observations of SIF. Finally, NIRV has thedistinct advantage of approximating black soil conditions, meaning it isreadily usable in remote sensing applications, where LAI and FVC areoften low and soil contamination is nearly universal (Fig. 4; Table 4).Previously proposed SIP-based indices, like the directional area scat-tering factor (DASF), require explicit consideration of soil backgroundeffects (Knyazikhin et al., 2013). In fact, NIRV and its relationship to SIFand BRFV can be expressed in terms of DASF (see Appendix, Eq. (A9)),
Fig. 7. Variation in solar angle and view angle cause large differences in SIFObs. A) View geometry alone causes upwards of a 70% difference in directional SIF, B)which if used as a proxy for total emitted SIF causes± 30% differences in estimated SIFTotal. C) Adjusting SIF by NIRV-derived f esc results in low relative errors ofestimated SIFTotal. All data calculated using SCOPE, with FVC equal to 0.5.
Fig. 8. SIFTotal, as estimated using f esc derived from NIRV, has a lower relativeerror than both SIFTotal estimated from assuming a constant scaling betweendirectional SIF and SIFTotal (green) and SIFTotal derived from NIRT-based esti-mates of f esc (orange). All data simulated using SCOPE, with critical parametersoutlined in Table 3. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
9

providing additional opportunities for the synthesis of NIRV, SIP, andglobal scale remote sensing of vegetation (Köhler et al., 2018). Suchsynthesis should help resolve outstanding questions about the relativecontrols of canopy-scale structure and physiology on plant productivity,a topic that has received growing attention in recent years (e.g.,Migliavacca et al., 2017).
5.2. The radiative transfer of fluorescence
Calculating total emitted near-infrared SIF, as opposed to direc-tional SIF, is an important step in using SIF to accurately estimate GPP.Several recent SIF studies, including Guanter et al., 2012, Zhang et al.,2016, and Sun et al., 2018 found that the slope of the SIF-GPP re-lationship varied by biome. However, these studies did not fully ac-count for directional effects and canopy escape ratio. As a result, the perbiome relationships previously described might be partly caused bydifferences in latitude and time of year, LAD, FVC, LAI and soilbrightness affecting f esc. Although the SIF-GPP relationship can be in-fluenced by many factors, including physiological (e.g., C3 and C4pathways of photosynthesis) and environmental factors, variations inthe radiative transfer of SIF (manifest in differing f esc) can resultin± 30% differences in observed SIF (Fig. 7), which likely plays animportant role in explaining spatial patterns of SIF at the global scale.
Accurate estimation of SIFTotal is especially critical to efforts to studyvariations in fluorescence yield (ΦF, see Eq. (1)) at the canopy scale andbeyond. ΦF is itself directly related to how energy is partitioned betweenphotochemical and non-photochemical processes within the leaf, makingthe measurement and prediction of ΦF an essential step in successfullyusing SIF to investigate plant physiology (Porcar-Castell et al., 2014).However, ΦF is both a small (approx. 1% ± 0.5% of APAR) and highlydynamic signal (Krause and Weis, 1991), varying on daily and seasonaltimescales (Yang et al., 2018; Miao et al., 2018; Porcar-Castell, 2011), aswell as across space (Malenovský et al., 2009; Atherton et al., 2017). Sucha small, complex signal, whose variations are small relative to its meanvalue, is easily obscured by f esc induced variations in SIF. NIRV offers oneapproach for removing the effects of f esc, which is a key step in thechallenge of mechanistically linking SIF to GPP. Previously, Badgley et al.,2017 used simulations to show that the ratio of SIF to NIRV can be used todiscern variations in ΦF. Our results relating NIRV to f esc provide a phy-sical explanation for why this approach works by linking NIRV to BRFV.
In evaluating the suitability of using NIRV to estimate f esc of SIF, it ishelpful to consider uncertainties in our approach. From a physicalstandpoint, SIF and NIRV are generated by different processes within thecanopy, which means that canopy-scale f esc of SIF can differ from the f esc
of NIRV. More specifically, vertical variations and differences in ΦF andωN cause SIF and NIR photons to originate from different places withinthe canopy. Thankfully, these differences are small, as demonstrated by aseries of supplementary SCOPE simulations we conducted to quantify theeffect (Table S1). We ran three simulations: i) a baseline for comparison,ii) a simulation where we varied meteorological and biochemical para-meters that affect ΦF but have no influence on ωN, and iii) a simulationwhere we varied leaf optical parameters affecting ωN but have no in-fluence on ΦF. In both instances, varying meteorological variables andleaf optical properties causes only the slightest change in the R2 andRMSE of the f esc of SIF and f esc of NIRV relationship, (Fig. S8).
One additional uncertainty in our approach concerns the spectralmismatch between near-infrared SIF (calculated between 740 and760 nm) and NIRV, measured at 858 nm to be consistent with MODIS.Differences in leaf albedo across these wavelengths mean that the f esc ofa solar photon at 858 nm is not strictly equal to the f esc of a SIF photonat a shorter wavelength. In practice, however, these differences aresmall. Using SCOPE, we performed two additional simulations: i) si-mulating both NIRV and SIF at 760 nm and ii) simulating SIF at 740 nmand NIRV at 858 nm. As expected, measuring NIRV and SIF at the samewavelength improves the linearity of f esc estimated by NIRV with si-mulated f esc of SIF (R2=0.93, up from 0.91; Fig. S9). In practical
terms, however, NIR reflectance is typically measured at much longerwavelengths to avoid several strong atmospheric absorption featuresthat complicate the accurate retrieval of surface reflectance from space-borne sensors. MODIS, VIIRS (the Visible Infrared Imaging RadiometerSuite), and Landsat all measure NIR at wavelengths extending beyond800 nm. Therefore, from a data availability standpoint, satellite-basedstudies that use NIRV from these sensors likely necessitate accom-modating some degree of spectral mismatch when compared againstSIF. However, studies should consider calculating f esc using the samewavelength as retrieved fluorescence if the NIR data is available. In thesame way, SIF measured at 740 nm, where chlorophyll more stronglyreabsorbs SIF photons, degrades the accuracy of NIRV-derived f esc (Fig.S10). While this effect is small, causing an increase of RMSE of< 0.2%and a reduction of R2 of 0.08, it is nonetheless worth mentioning.Future work to more accurately characterize the within-canopy re-ab-sorption of SIF (e.g., Romero et al., 2018) and incorporate such effectsinto SIF retrievals can help ameliorate the uncertainties that arise fromthe spectral mismatch of SIF and NIRV.
5.3. Approximating BRFV with NIRV
NIRV minimizes the effects of variations in soil brightness, evenunder conditions where the NDVI of the soil is high and LAI (or FVC) islow (Fig. 4). Yet in these cases, it is important to keep in mind that NIRVis still only an approximation of BRFV. In our simulations, NIRV wasslightly lower than BRFV when the LAI was low and, by contrast, washigher than BRFV when the LAI was high (Fig. 1). This is due to the factthat NDVI is not strictly equal to 0 when LAI is 0 and does not ne-cessarily equal 1 when LAI is high because non-vegetated surfaces (e.g.,soil) can have a non-zero NDVI and FVC rarely reaches 1.
In practical terms, this effect can be further reduced by normalizingNDVI on a per-pixel basis to account for spatial variation in the multi-year average value of NDVImin and NDVImax. More specifically, theNDVI in Eq. (8) could instead be replaced by NDVI′, which takes theform:
=NDVI NDVI NDVINDVI NDVI
.min
max min (16)
In initial simulations where Eq. (16) is substituted into Eq. (8), wefound a 35% reduction in RMSE of the NIRV-BRFV relationship, withespecially large improvements under extremely high and low values ofLAI and FVC (Fig. 9). Adjusting for the overall range of NDVI makes theNIRV-BRFV relationship nearly one-to-one, with a small intercept of0.02 and a slope of 1.01.
Fig. 9. NIRV can be further improved by accounting for per-pixel minimum(NDVImin) and maximum (NDVImax) values of NDVI. NDVImin and NDVImax canbe derived from multi-angular remote sensing data, allowing NIRV to even moreclosely approximate BRFV (see Eq. (9)).
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
10

Fully adopting Eq. (16) would introduce two additional unknownparameters, NDVImin and NDVImax, which might hinder application.However, there exist multi-angular remote sensing approaches forquantifying NDVImin and NDVImax that could lead to further improve-ments in NIRV, especially over sparse canopies (Song et al., 2017; Muet al., 2017). Our analysis of MODIS and TROPOMI data showed thatthe NIRV-SIF relationship was relatively non-linear at a sparsely vege-tated grassland site, which might arise from NIRV being an inadequateapproximation of the black soil condition when vegetation cover isextremely low (Table 4 and Fig. S7). Future work will need to establishthe exact lower bound of FVC where the NIRV approximation ofNDVI ·NIRT breaks down in practice. Further improvements could alsobe made by properly accounting for spatial and temporal variation inωN, the leaf single scattering albedo in the NIR band (see Eq. (4)). Sucha modification would introduce yet another per-pixel parameter,though accounting for variations in ωN might prove practical for site-level studies.
5.4. Applications without knowledge of fPAR
Perhaps the largest downside to our proposed measure of f esc is itsreliance on readily available and reliable fPAR data. In fact, using Eq.(12) to estimate total emitted SIF not only requires fPAR, it also re-quires that NIRV be calculated with near-identical sun-canopy-sensorgeometry to SIF. The forthcoming FLEX mission, for example, will fly intandem with another European satellite, Sentinel 3 (S-3), allowing theco-registration of a FLEX-based SIF product with the S-3 fPAR product(Gobron, 2010) and NIRV, as laid out in the FLEX mission concept plan(Drusch et al., 2017). Development of fPAR products based on datafrom existing SIF sensors that also make reflectance measurements inthe visible domain (such as GOME-2), could provide another avenue forusing our NIRV-based correction of f esc. Further complications are in-troduced by the need to measure fPARgreen, as opposed to simply theinteraction of photons with non-photosynthetic aspects of the canopy.This is a lingering challenge that continues to motivate advances in theremote sensing of fPAR (e.g., Zhang et al., 2005) that will ultimatelybenefit the more robust calculation of f esc. Gower et al., 1999 foundthat the difference between fPAR and fPARgreen diverged between 5%and 35%, though in practice they argued that the difference likely fallson the lower end of this range because leaves tend to preferentiallycluster around branches, as opposed to being randomly distributedthroughout the canopy (see also, Dufrêne and Bréda, 1995). Such rea-soning has empirical support as well. Both Kucharik et al., 1998 andRyu et al., 2012 found that woody elements of the canopy were largelymasked by vegetation, resulting in only small differences between plantarea index and leaf area index. While accurately calculating fPARcomplicates the global application of Eq. (12), the approach should bemore than suitable for in situ studies where SIF, NIRV, and fPAR can bemore robustly determined through careful site-level instrumentation. Ifcombined with in situ measurements of photosynthesis, these datamight help in further elucidating the nature of SIF-NIRV-photosynthesisrelationship.
Alternatively, it should still be possible to use Eq. (12) even withoutknowing fPAR to normalize SIF to a constant viewing geometry. Cross-sensor combinations of SIF and reflectance data should be possible solong as SIF and optical measurements are made in close succession,
such that the fPAR across the two satellite measurements can be as-sumed constant. While such an approach would enable the normal-ization of SIF and allow for comparisons of SIF measurements acrossspace, it would not allow the precise calculation of SIFTotal due to un-certainty in the true value of fPAR. In this mode, MODIS BRF productsmight be used to calculate NIRV and combined with off-nadir SIF andNIRV observations from TROPOMI or OCO-2 to generate a nadir-nor-malized SIF dataset. Even more promising is the newly launched EPICsensor, a high-resolution spectral camera capable of capturing sub-hourly, full-disc images of the Earth at roughly 10 km2 (Marshak andKnyazikhin, 2017). EPIC is situated such that it stares directly into thehot-spot, which offers two unique advantages. First, constant viewgeometry and high temporal resolution would allow normalizationusing near simultaneous observations to the EPIC view, which wouldeliminate seasonal changes in solar elevation that still complicate nadir-adjusted imagery. Second, looking into the hot-spot means that EPICmostly sees only fully-illuminated leaves. Sun-lit leaves, in turn, con-tribute the lion-share of total emitted fluorescence, meaning hot-spotobservations of NIRV may be expected to provide directional-correctedmeasurements of SIF that strongly correlate with total emitted SIF andin turn provide a strong basis for estimating APAR and GPP globally.
6. Conclusion
The escape ratio between directional SIF and total emitted SIF caninfluence the SIF-GPP relationship and is determined by the sun-ca-nopy-sensor geometry, canopy structure parameters and leaf/soil op-tical properties. The widely used radiative transfer forward modelsrequire knowledge of canopy structure parameters and leaf/soil opticalproperties, while the relatively easier BRFT approach can only be usedover dense canopies or against dark soil backgrounds. We developed asimple but accurate approach to estimate the escape ratio, NIR
fPARV , which
effectively removes the influence of soil reflectance and can be easilyapplied using existing in situ or remotely sensed NIRV and fPAR data-sets. The proposed escape ratio formula was evaluated for differentcanopy structure, soil brightness, and view geometry cases by SCOPEsimulations, and achieved higher accuracy than the BRFT approach.
Acknowledgements
We thank Kaiyu Guan, Ari Kornfeld, Guofang Miao, and Xi Yang fortheir helpful discussions. Y.Z. was in part supported by the National KeyResearch and Development Program (No. 2018YFA0605503) and theNational Natural Science Foundation of China (No. 41701401) at thebeginning of the research. Christian Frankenberg and Philipp Köhlerhelped in our interpretation of the TROPOMI results and provided theTROPOMI data, which was funded by the Earth Science U.S.Participating Investigator (Grant Number: NNX15AH95G). G.B. wassupported by a NASA Earth and Space Science fellowship. B.D. and Y.R.were supported by the National Research Foundation of Korea (NRF-2016M1A3A3A02018195). While writing this paper, Y.R. was sup-ported by the Carnegie Institution for Science during a sabbatical leave.M.C. was supported by a Laboratory Directed Research andDevelopment project sponsored by the Pacific Northwest NationalLaboratory of the U.S. Department of Energy.
Appendix A. Appendix
A.1. Canopy spectral invariants primer
This Appendix provides additional background on canopy spectral invariant theory and how spectral invariant properties (SIP) relate to BRF andf esc. For a complete introduction, see Knyazikhin et al., 2013 and Stenberg et al., 2016. For our purposes, we are more interested in the practicalapplication of SIP to better understand f esc. The three SIP parameters used in our derivation relating NIRV and fPAR to f esc are:
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
11

1. Escape probability (ρn(Ω)): the probability that a solar photon, on its n-th interaction with a vegetated element of the canopy, escapes the canopyin the direction Ω.
2. Recollision probability (pn): the probability that a solar photon, on its n-th interaction with a vegetated element of the canopy, recollides with thecanopy an n-th plus one time.
3. Canopy interceptance (i0): the probability that an incoming solar photon interacts with a vegetated element of the canopy.
Importantly, SIP hold for all wavelengths of incoming light. In this sense, SIP allows for canopy radiative transfer to be treated in the abstract, asopposed to requiring detailed knowledge of canopy geometry.
A.2. Spectral invariants and fesc
SIP are relevant to the manuscript at hand, as f esc can be described purely in terms of ρn, pn, and the near-infrared (NIR) leaf albedo, ωN, whichalthough not an invariant property, is relatively invariant across time and space (Gates et al., 1965; Asner, 1998). Using these terms, f esc can bedescribed as:
= + + + …f p p p( , ) ( ) · ( )· ( ) · ( )· · ( )· ( ) .BSesc
f f f f1 1 2 1 2 3 (A1)
Note that for purposes of this Text, we present all results in terms of the “black soil” condition, which we denote with the subscript “BS.” AllEquations can be expanded to account for the multiple scattering of a reflective soil background, but are excluded here both for brevity and becauseNIRV closely approximates the BS condition. Furthermore, for Eq. (A1), the SIF re-excited by multiple scattered SIF photons are regarded as neg-ligible (< 0.1%) (Zhao et al., 2016).
The geometric series in Eq. (A1) can be more succinctly written as:
=fp( , )( )
1 ( ),BS
escf
f (A2)
which more directly illustrates the relationship of ρ and p to f esc. As was the case in the Main Text, Eq. (A2) can again be simplified if ωN is assumedto equal 1. Of course, precise knowledge of ωN would improve estimates of f esc. Furthermore, the linearity of NIRV and f esc by Eq. (12) from the MainText underscores our claim that NIRV represents a new tool for exploring SIP at large scales.
A.3. The directional area scattering factor
SIP also allow use to describe the relationship between SIF, f esc, and NIRV in slightly more comprehensive terms. The following equations werehelpful in our own exploration of the NIRV- f esc relationship and, for this reason, we present them here.
The three SIP outlined above combine together to form a parameter known as the directional area scattering factor (DASF):
=DASF ip
( )1
,0
(A3)
which fully describes the canopy radiative transfer environment. Below, we use DASF as a bridging term between f esc, directional SIF, and NIRV. Wedo this by describing each of these variables in terms of DASF.
We start by relating f esc to DASF, which requires only a slight modification of Eq. (A2). By slightly rearranging how we reduce the Neumannseries presented in Eq. (A1) and simultaneously multiplying and dividing by i0, we get:
=
=
fpip
pp i
DASFi
( , )( )
1 ( )( )
1· 11 ( )
· 1
· 1 .
BSesc
ff
f
0
0
0 (A4)
Next, we can describe directional SIF as a function of total emitted SIF and the escape ratio. The total emitted SIF by all leaves within the canopycan be written as:
=SIF i Q M d( ) ( )· ( , )·Total f dir e e f F e0 400
750
(A5)
where Qdir(λe) is incoming photon flux density at wavelength λe (we ignore diffuse radiation for now), M(λe,λf) is the leaf fluorescence excitation-emission matrix where rows represent the excitation wavelength λe from 400 to 750 nm, and columns represent the emission wavelength λf from 640to 850 nm (van der Tol et al., 2009). M(λe,λf) is linearly scaled with the amplification factor F which is PAR-dependent and can vary throughoutthe canopy. SIFTotal becomes directional SIF (SIF(Ω)) by accounting for f esc:
=SIF SIF f( , ) ( )· ( , ),f Total f BSesc
f (A6)
which can be fully expanded to:
=
=
SIF ip
pp
Q M d
DASF CES
( , )( )
1· 1
1 ( )( )· ( , )·
· .
ff
dir e e f F e0
400
750
(A7)
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
12

Eq. (A7) neatly partitions SIF into two components: the purely structural/radiative transform component represented by DASF and what we callthe canopy emission-scattering (CES) coefficient, which relates to leaf biochemical properties by M(λe,λf), incident radiation Qdir(λe), and electrontransport via ΦF′.
The final step in relating NIRV to f esc via DASF is describing BRFV in terms of DASF. This step has previously been described by Knyazikhin et al.(2013) in the same paper in which DASF was first introduced:
=
=
BRF ip
pp
DASF CSC
( , )( )
1· 1
1 ( )( )
· .
V ff
f0
(A8)
As was the case in Eq. (A7), this rearrangement separates structural concerns (encapsulated by DASF) and the canopy scattering coefficient (CSC),which relates to leaf albedo (see also, Köhler et al., 2018).
With this, all the pieces are in place for relating NIRV, SIF, and f esc to each other via DASF. To fully complete the circle, from Eqs. (A4), (A8), and(10) from the Main Text, we can write:
=DASF NIRCSC
NIR
V
V
N (A9)
Appendix B. Supplementary information
Supplementary information to this article can be found online at https://doi.org/10.1016/j.rse.2019.05.028.
References
Asner, G.P., 1998. Biophysical and biochemical sources of variability in canopy re-flectance. Remote Sens. Environ. 640 (3), 0 234–253.
Atherton, J., Olascoaga, B., Alonso, L., Porcar-Castell, A., 2017. Spatial variation of leafoptical properties in a boreal forest is influenced by species and light environment.Front. Plant Sci. 8, 0 309.
Badgley, G., Field, C.B., Berry, J.A., 2017. Canopy near-infrared reflectance and terrestrialphotosynthesis. Sci. Adv. 30 (3), e1602244.
Baret, F., Vanderbilt, V.C., Steven, M.D., Jacquemoud, S., 1994. Use of spectral analogy toevaluate canopy reflectance sensitivity to leaf optical properties. Remote Sens.Environ. 480 (2), 253–260 0.
Chen, J.M., 1996. Optically-based methods for measuring seasonal variation of leaf areaindex in boreal conifer stands. Agric. For. Meteorol. 800 (2–4), 0 135–163.
Drusch, M., Moreno, J., Bello, U.D., Franco, R., Goulas, Y., Huth, A., Kraft, S., Middleton,E.M., Miglietta, F., Mohammed, G., et al., 2017. The fluorescence explorer missionconcept — ESA's earth explorer 8. IEEE Trans. Geosci. Remote Sens. 550 (3),1273–1284 0.
Du, S., Liu, L., Liu, X., Hu, J., 2017. Response of canopy solar-induced chlorophyllfluorescence to the absorbed photosynthetically active radiation absorbed by chlor-ophyll. Remote Sens. 90 (9), 911.
Dufrêne, E., Bréda, N., 1995. Estimation of deciduous forest leaf area index using directand indirect methods. Oecologia 1040 (2), 156–162.
Fournier, A., Daumard, F., Champagne, S., Ounis, A., Goulas, Y., Moya, I., 2012. Effect ofcanopy structure on sun-induced chlorophyll fluorescence. ISPRS J. Photogramm.Remote Sens. 68, 112–120.
Frankenberg, C., Butz, A., Toon, G.C., 2011a. Disentangling chlorophyll fluorescence fromatmospheric scattering effects in O2 a-band spectra of reflected sun-light. Geophys.Res. Lett. 38 (3), 0.
Frankenberg, C., Fisher, J.B., Worden, J., Badgley, G., Saatchi, S.S., Lee, J.-E., Toon, G.C.,Butz, A., Jung, M., Kuze, A., et al., 2011b. New global observations of the terrestrialcarbon cycle from GOSAT: patterns of plant fluorescence with gross primary pro-ductivity. Geophys. Res. Lett. 380 (17).
Gastellu-Etchegorry, J.P., Martin, E., Gascon, F., 2004. Dart: a 3d model for simulatingsatellite images and studying surface radiation budget. Int. J. Remote Sens. 250 (1), 073–96.
Gastellu-Etchegorry, J.-P., Lauret, N., Yin, T., Landier, L., K., A., Malenovskỳ, Z., Bitar,A.A., Aval, J., Benhmida, S., Qi, J., et al., 2017. Dart: recent advances in remotesensing data modeling with atmosphere, polarization, and chlorophyll fluorescence.IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 100 (6), 0 2640–2649.
Gates, D.M., Keegan, H.J., Schleter, J.C., Weidner, V.R., 1965. Spectral properties ofplants. Appl. Opt. 40 (1), 0 11–20.
Gitelson, A.A., Buschmann, C., Lichtenthaler, H.K., 1998. Leaf chlorophyll fluorescencecorrected for re-absorption by means of absorption and reflectance measurements. J.Plant Physiol. 1520 (2–3), 0 283–296.
Gobron, N., 2010. Ocean and land colour instrument (olci) fapar and rectified channelsover terrestrial surfaces algorithm theoretical basis document. Eur report no. xxxxxen. In: European Commission Joint Research Centre (JRC) Scientific and TechnicalReports.
Gower, S.T., Kucharik, C.J., Norman, J.M., 1999. Direct and indirect estimation of leafarea index, fapar, and net primary production of terrestrial ecosystems. Remote Sens.Environ. 700 (1), 0 29–51.
Guanter, L., Frankenberg, C., Dudhia, A., Lewis, P.E., Gómez-Dans, J., Kuze, A., Suto, H.,
Grainger, R.G., 2012. Retrieval and global assessment of terrestrial chlorophyllfluorescence from GOSAT space measurements. Remote Sens. Environ. 121 (0),236–251.
Guanter, L., Zhang, Y., Jung, M., Joiner, J., Voigt, M., Berry, J.A., Frankenberg, C., Huete,A.R., Zarco-Tejada, P., Lee, J.E., Moran, M.S., 2014. Global and time-resolved mon-itoring of crop photosynthesis with chlorophyll fluorescence. Proc. Natl. Acad. Sci.111 (14), E1327–E1333.
He, L., Chen, J.M., Liu, J., Mo, G., Joiner, J., 2017. Angular normalization of gome-2 sun-induced chlorophyll fluorescence observation as a better proxy of vegetation pro-ductivity. Geophys. Res. Lett. 440 (11), 0 5691–5699.
Hernández-Clemente, R., North, P.R.J., Hornero, A., Zarco-Tejada, P.J., 2017. Assessingthe effects of forest health on sun-induced chlorophyll fluorescence using the fluor-flight 3-d radiative transfer model to account for forest structure. Remote Sens.Environ. 193 (0), 165–179.
Huang, D., Knyazikhin, Y., Dickinson, R.E., Rautiainen, M., Stenberg, P., D., M., Lewis, P.,Cescatti, A., Tian, Y., Verhoef, W., et al., 2007. Canopy spectral invariants for remotesensing and model applications. Remote Sens. Environ. 1060 (1), 106–122 0.
Jacquemoud, S., Baret, F., 1990. Prospect: a model of leaf optical properties spectra.Remote Sens. Environ. 340 (2), 0 75–91.
Knyazikhin, Y., Martonchik, J.V., Myneni, R.B., Diner, D.J., Running, S.W., 1998.Synergistic algorithm for estimating vegetation canopy leaf area index and fraction ofabsorbed photosynthetically active radiation from modis and misr data. J. Geophys.Res. Atmos. 1030 (D24), 0 32257–32275.
Knyazikhin, Y., Schull, M.A., Stenberg, P., Mõttus, M., Rautiainen, M., Yang, Y., Marshak,A., Carmona, P.L., Kaufmann, R.K., Lewis, P., et al., 2013. Hyperspectral remotesensing of foliar nitrogen content. Proc. Natl. Acad. Sci. 1100 (3), 0 E185–E192.
Köhler, P., Frankenberg, C., Magney, T.S., Guanter, L., Joiner, J., Landgraf, J., 2018.Global retrievals of solar-induced chlorophyll fluorescence with tropomi: first resultsand intersensor comparison to OCO-2. Geophys. Res. Lett. 450 (19), 10–456.
Krause, G.H., Weis, E.L., 1991. Chlorophyll fluorescence and photosynthesis: the basics.Annu. Rev. Plant Biol. 420 (1), 313–349 0.
Kucharik, C.J., Norman, J.M., Gower, S.T., 1998. Measurements of branch area and ad-justing leaf area index indirect measurements. Agric. For. Meteorol. 910 (1–2),69–88 0.
Liu, L., Liu, X., Wang, Z., Zhang, B., 2016. Measurement and analysis of bidirectional sifemissions in wheat canopies. IEEE Trans. Geosci. Remote Sens. 540 (5), 2640–2651 0.
Liu, X., Guanter, L., Liu, L., Damm, A., Malenovsky, Z., Rascher, U., Peng, D., Du, S.,Gastellu-Etchegorry, J.P., 2018. Downscaling of solar-induced chlorophyll fluores-cence from canopy level to photosystem level using a random forest model. RemoteSens. Environ. 1–18.
Malenovský, Z., Bandhu Mishra, K., Zemek, F., Rascher, U., Nedbal, L., 2009. Scientificand technical challenges in remote sensing of plant canopy reflectance and fluores-cence. J. Exp. Bot. 600 (11), 0 2987–3004.
Marshak, A., Knyazikhin, Y., 2017. The spectral invariant approximation within canopyradiative transfer to support the use of the epic/dscovr oxygen b-band for monitoringvegetation. J. Quant. Spectrosc. Radiat. Transf. 191 (0), 7–12.
Miao, G., Guan, K., Xi, Y., Bernacchi, C.J., Berry, Joseph A., DeLucia, E.H., Jin, W., Moore,C.E., Meacham, K., Cai, Y., et al., 2018. Sun-induced chlorophyll fluorescence, pho-tosynthesis, and light use efficiency of a soybean field from seasonally continuousmeasurements. J. Geophys. Res. Biogeosci. 1230 (2), 610–623 0.
Migliavacca, M., Perez-Priego, O., Rossini, M., El-Madany, T.S., Moreno, G., van der Tol,C., Rascher, U., Berninger, A., Bessenbacher, V., Burkart, A., et al., 2017. Plantfunctional traits and canopy structure control the relationship between photo-synthetic CO2 uptake and far-red sun-induced fluorescence in a Mediterranean
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
13

grassland under different nutrient availability. New Phytol. 2140 (3), 0 1078–1091.Mu, X., Hu, R., Zeng, Y., McVicar, T.R., Ren, H., Song, W., Wang, Y., Casa, R., Qi, J., Xie,
D., et al., 2017. Estimating structural parameters of agricultural crops from ground-based multi-angular digital images with a fractional model of sun and shade com-ponents. Agric. For. Meteorol. 246, 162–177.
Porcar-Castell, A., 2011. A high-resolution portrait of the annual dynamics of photo-chemical and non-photochemical quenching in needles of Pinus sylvestris. Physiol.Plant. 1430 (2), 0 139–153.
Porcar-Castell, A., Tyystjärvi, E., Atherton, J., van der Tol, C., Flexas, J., Pfündel, E.E.,Moreno, J., Frankenberg, C., Berry, J.A., 2014. Linking chlorophyll a fluorescence tophotosynthesis for remote sensing applications: mechanisms and challenges. J. Exp.Bot. 650 (15), 0 4065–4095.
Qi, J., Chehbouni, A., Huete, A.R., Kerr, Y.H., Sorooshian, S., 1994. A modified soil ad-justed vegetation index. Remote Sens. Environ. 480 (2), 0 119–126.
Raabe, K., Pisek, J., Sonnentag, O., Annuk, K., 2015. Variations of leaf inclination angledistribution with height over the growing season and light exposure for eightbroadleaf tree species. Agric. For. Meteorol. 214 (0), 2–11.
Ramos, M.E., Lagorio, M.G., 2006. A model considering light reabsorption processes tocorrect in vivo chlorophyll fluorescence spectra in apples. Photochem. Photobiol. Sci.50 (5), 508–512 0.
Romero, J.M., Cordon, G.B., Lagorio, M.G., 2018. Modeling re-absorption of fluorescencefrom the leaf to the canopy level. Remote Sens. Environ. 204 (0), 138–146.
Ross, J., 1981. The Radiation Regime and Architecture of Plant Stands. Dr W. JunkPublishers.
Ryu, Y., Sonnentag, O., Nilson, T., Vargas, R., Kobayashi, H., Wenk, R., Baldocchi, D.D.,2010. How to quantify tree leaf area index in an open savanna ecosystem: a multi-instrument and multi-model approach. Agric. For. Meteorol. 1500 (1), 63–76 0.
Ryu, Y., Verfaillie, J., Macfarlane, C., Kobayashi, H., Sonnentag, O., Vargas, R., Ma, S.,Baldocchi, D.D., 2012. Continuous observation of tree leaf area index at ecosystemscale using upward-pointing digital cameras. Remote Sens. Environ. 126 (0),116–125.
Schaaf, C.B., Gao, F., Strahler, A.H., Lucht, W., Li, X., Tsang, T., Strugnell, N.C., Zhang, X.,Jin, Y., Muller, J.-P., et al., 2002. First operational brdf, albedo nadir reflectanceproducts from modis. Remote Sens. Environ. 830 (1–2), 135–148 0.
Smith, W.K., Biederman, J.A., Scott, R.L., Moore, D.J.P., He, M., Kimball, J.S., Yan, D.,Hudson, A., Barnes, M.L., MacBean, N., et al., 2018. Chlorophyll fluorescence bettercaptures seasonal and interannual gross primary productivity dynamics across dry-land ecosystems of southwestern North America. Geophys. Res. Lett. 450 (2),748–757 0.
Smolander, S., Stenberg, P., 2005. Simple parameterizations of the radiation budget ofuniform broadleaved and coniferous canopies. Remote Sens. Environ. 940 (3), 0355–363.
Song, W., Mu, X., Ruan, G., Gao, Z., Li, L., Yan, G., 2017. Estimating fractional vegetationcover and the vegetation index of bare soil and highly dense vegetation with aphysically based method. Int. J. Appl. Earth Obs. Geoinf. 58, 0 168–176.
Stenberg, P., Mõttus, M., Rautiainen, M., 2016. Photon recollision probability in model-ling the radiation regime of canopies — a review. Remote Sens. Environ. 183,98–108.
Sun, Y., Frankenberg, C., Wood, J.D., Schimel, D.S., Jung, M., Guanter, L., Drewry, D.T.,Verma, M., Porcar-Castell, A., Griffis, T.J., et al., 2017. Oco-2 advances photo-synthesis observation from space via solar-induced chlorophyll fluorescence. Science3580 (6360), 0 eaam5747.
Sun, Y., Frankenberg, C., Jung, M., Joiner, J., Guanter, L., Köhler, P., Magney, T., 2018.Overview of solar-induced chlorophyll fluorescence (sif) from the orbiting carbonobservatory-2: retrieval, cross-mission comparison, and global monitoring for gpp.Remote Sens. Environ. 209, 808–823.
van der Tol, C., Verhoef, W., Timmermans, J., Verhoef, A., Su, Zhongbo, 2009. An in-tegrated model of soil-canopy spectral radiances, photosynthesis, fluorescence,temperature and energy balance. Biogeosciences 60 (12), 0 3109–3129.
van der Tol, C., PKE Campbell, J.A.B., Rascher, U., 2014. Models of fluorescence andphotosynthesis for interpreting measurements of solar-induced chlorophyll fluores-cence. J. Geophys. Res. Biogeosci. 1190 (12), 0 2312–2327.
van der Tol, C., Rossini, M., Cogliati, S., Verhoef, W., Colombo, R., Rascher, U.,Mohammed, G., 2016. A model and measurement comparison of diurnal cycles ofsun-induced chlorophyll fluorescence of crops. Remote Sens. Environ. 186, 0663–677.
Tucker, C.J., 1979. Red and photographic infrared linear combinations for monitoringvegetation. Remote Sens. Environ. 80 (2), 0 127–150.
Veefkind, J.P., Aben, I., McMullan, K., Förster, H., De Vries, J., Otter, G., Claas, J., Eskes,H.J., De Haan, J.F., Kleipool, Q., et al., 2012. Tropomi on the ESA sentinel-5 pre-cursor: a GMES mission for global observations of the atmospheric composition forclimate, air quality and ozone layer applications. Remote Sens. Environ. 120, 70–83.
Verhoef, W., 1984. Light scattering by leaf layers with application to canopy reflectancemodeling: the sail model. Remote Sens. Environ. 160 (2), 125–141.
Vilfan, N., van der Tol, C., Muller, O., Rascher, U., Verhoef, W., 2016. Fluspect-b: a modelfor leaf fluorescence, reflectance and transmittance spectra. Remote Sens. Environ.186, 596–615.
Yang, K., Ryu, Y., Dechant, B., Berry, J.A., Hwang, Y., Jiang, C., Kang, M., Kim, J., Kim,
H., Kornfeld, A., Yang, X., 2018. Sun-induced chlorophyll fluorescence is morestrongly related to absorbed light than to photosynthesis at half-hourly resolution in arice paddy. Remote Sens. Environ. 216, 658–673.
Yang, P., van der Tol, C., 2018. Linking canopy scattering of far-red sun-induced chlor-ophyll fluorescence with reflectance. Remote Sens. Environ. 209, 456–467.
Yang, X., Tang, J., Mustard, J.F., Lee, J.-E., Rossini, M., Joiner, J., William Munger, J.,Kornfeld, A., Richardson, A.D., 2015. Solar-induced chlorophyll fluorescence thatcorrelates with canopy photosynthesis on diurnal and seasonal scales in a temperatedeciduous forest. Geophys. Res. Lett. 420 (8), 2977–2987.
Zhang, Q., Xiao, X., Braswell, B., Linder, E., Baret, F., Berrien Moore, I.I.I., 2005.Estimating light absorption by chlorophyll, leaf and canopy in a deciduous broadleafforest using modis data and a radiative transfer model. Remote Sens. Environ. 990(3), 0 357–371.
Zhang, Y., Guanter, L., Berry, J.A., van der Tol, C., Yang, X., Tang, J., Zhang, F., 2016.Model-based analysis of the relationship between sun-induced chlorophyll fluores-cence and gross primary production for remote sensing applications. Remote Sens.Environ. 187, 145–155.
Zhao, F., Dai, X., Verhoef, W., Guo, Y., van der Tol, C., Li, Y., Huang, Y., 2016. Fluorwps: aMonte Carlo ray-tracing model to compute sun-induced chlorophyll fluorescence ofthree-dimensional canopy. Remote Sens. Environ. 187, 385–399.
Glossary
ABSV: canopy absorbance of visible lightBRF: bidirectional reflectance factorBRFT: the total scene bidirectional reflectance factorBRFV: contribution to BRFT of photons reflected off only the scene's vegetative componentBRFS: contribution to BRFT of photons reflected off soil aloneBRFM: contribution to BRFT of photons from multiple scattering between the vegetation
and soilBS: black soil problemCES: Canopy Emission/Scattering coefficientCSC: Canopy Scattering CoefficientDASF: Directional Area Scattering FactorFVC: fractional vegetation coverFV: the fraction of NIRT that originates from interactions with vegetation aloneFS: the fraction of NIRT that originates from interactions with soil aloneFM: the fraction of NIRT that originates from the multiple scattering between the vege-
tation and soilfPAR: the fraction of absorbed photosynthetically active radiationfPARchl: the fraction of PAR absorbed by chlorophyllf esc(λf,Ω): escape ratio, which is the fraction of SIF photons at a given wavelength (λf)
that ultimately escape the canopy and are observed at the view angle of ΩΦF: the quantum yield of fluorescenceΦF
′: the amplification factor of the quantum yield of fluorescencei0: canopy interceptance, which is the probability that an incoming solar photon interacts
with a vegetated element of the canopyLAI: leaf area indexM(λe,λf): he leaf fluorescence excitation-emission matrix where rows represent the ex-
citation wavelength λe from 400 to 750 nm, and columns represent the emissionwavelength λf from 640 to 850 nm
NDVI: the normalized difference vegetation indexNIRT: shorthand for BRFT(NIR)NIR′T: the total NIR radiance reflected by the land surfaceNIRV: the NIR reflectance of vegetation, which can be calculated by NDVI •NIRT and is an
approximation of BRFV(NIR)NIR′V: the total NIR radiance reflected by the vegetated component of the land surfaceNIRS: contribution to NIRT of photons reflected off soil aloneNIRM: contribution to NIRT of photons from multiple scattering between the vegetation
and soilPAR: absorbed photosynthetically active radiationρn(Ω): Escape probability, which is the probability that a solar photon, on its n-th in-
teraction with a vegetated element of the canopy, escapes the canopy in the directionΩ
pn: Recollision probability, which is the probability that a solar photon, on its n-th in-teraction with a vegetated element of the canopy, recollides with the canopy an n-thplus one time.
Qdir(λe): the direct incoming photon flux density at wavelength λeSλ: the flux of incoming solar radiation at wavelength λSIF: solar-induced chlorophyll fluorescenceSIFTotal(λf): the sum of all SIF photons at a given wavelength (λf) emitted by all leaves
within the canopy in all directionsSIFObs(λf,Ω): SIF radiance observed at the top of canopy at a given wavelength (λf) and at
the view angle of ΩSIP: spectral invariant propertiesωλf: leaf single scattering albedo at wavelength λf
Y. Zeng, et al. Remote Sensing of Environment 232 (2019) 111209
14