a north african-european transition fauna: water beetles...
TRANSCRIPT

AQUATIC CONSERVATION: MARINE A N D FRESHWATER ECOSYSTEMS, VOL. 6, 121-140 (1996)
A North A frican-European transition fauna: water beetles (Coleoptera) from the Ebro delta and other Mediterranean
coastal wetlands in the Iberian peninsula
IGNACIO RIBERA The Scottish Agricultural College, Environmental Sciences Department. Auchincruive, Ayr KA6 SH W, UK
DAVID T. BILTON University of Plymouth, Department of Biological Sciences, Drake Circus, Plymouth PL4 8AA
PEDRO AGUILERA Del Pou 21-23, atic 2, 08016 Barcelona, Spain
GARTH N. FOSTER The Scottish Agricultural College, Environmental Sciences Department, Auchincruive, Ayr KA6 SH W, UK
ABSTRACT
1. The aquatic Coleoptera of some major wetlands in the Mediterranean coast of Spain were surveyed, with the aim of assessing their conservation value and the status of some coastal species in the Iberian Peninsula.
2. Classification of 59 species from 13 sites between the deltas of the rivers Ebro and Llobregat using TWINSPAN suggested that water origin and amount and structure of vegetation were major factors influencing the composition of the coastal water beetle communities.
3. The Ebro delta is considered to be a key biogeographical area, being the limit of the distribution of a number of north African and European species. Coastal sites in the Iberian peninsula sustain the only known populations in continental Europe of at least three Ethiopian species, although two of them may have become extinct. Some European species known to have reached the north of Catalonia were also considered to be possibly extinct in the Iberian peninsula owing to the destruction of their habitats. Three circum-Mediterranean coastal species were recorded for the first time in the Iberian peninsula, one of them in a genus new to its fauna.
4. Despite the scarcity of true endemics-the result of the recent geological origin and the temporal dynamics of coastal systems-some species were exclusive to temporary habitats, and had very restricted distributions.
5 . Although most of the areas studied have some form of protection, this is mainly focused in large, permanent lagoons and on the vertebrate fauna. The need to include marginal areas and invertebrates in the protection plans for the area is stressed.
INTRODUCTION
The Mediterranean basin is recognized as an area of high biodiversity, both on a regional (Palaearctic) and on a global scale. For example, it harbours ca. 10% of the world’s vascular plant species (Greuter, 1995). It is also home to more than a thousand species of water beetles, about half of which occur in the Iberian
CCC 1052-761 3/96/030121-20 01996 by John Wiley & Sons, Ltd
Received 12 March 1996 Accepted 9 August 1996

122 1. RIBERA ET A L .
peninsula. This fauna is still incompletely known, and new species are being regularly discovered; e.g. 73 were added to the Iberian fauna list in the last 10 years, 46 of them new to science. Most of the Iberian endemics are restricted to mountain ranges in the south and the west, but many of the recently discovered non-endemic species were North African taxa found in coastal habitats. The coast of the Mediterranean countries is one of the most threatened areas in Europe, owing to the development of tourism and an increase in the affluence of the local population (see e.g. Ranwell, 1977; Folch, 1988; Ehrlich, 1995; Greuter, 1995).
In the Iberian peninsula the invertebrate fauna of coastal wetlands is largely ignored in conservation plans and in the designation of protected areas, which are based mainly-sometimes even exclusively-on the birds. The vertebrate bias in the protection of the coastal wetlands has led to a reduction in the number and extent of the marginal, temporary habitats, which are of less importance for sustaining larger species of vertebrates, but which often provide the only suitable habitats for specific invertebrate assemblages.
The aim of this work is to give the first detailed account of the aquatic Coleoptera of the Ebro delta and other coastal wetlands in Spain, and to discuss their conservation value in relation to the Iberian (and European) fauna and the biogeographical significance of these areas as a transition zone between Europe and North Africa.
METHODS
Study area
Areas included in the study ranged from the Albufera de Adra (province of Almeria) in the south, to the Aiguamolls de 1’Emporda (province of Girona) in the north (Table 1 and Figure 1). Most of the data were collected by the authors between 1985 and 1995. Previously published information has been collated but it is very scarce for the majority of the studied sites (see Appendix).
The Albufera Nueva de Adra (Almeria) is a coastal lagoon with a surface area of approximately 40 ha, with a mixture of fresh water from temporary streams and sea water infiltrating through the subsoil, surrounded by a belt of giant reed (Arundo donax) and other macrophytes.
The Santa Pola and Torrevieja salines (Alicante) are large, shallow enclosures in which sea water is progressively concentrated to obtain salt. The traditional management of these pools provides a wide range of different environmental conditions, although in most lagoons the water is too saline to sustain any but a few specialized species. The total surface of the salines in both of them is about 2500 ha, with individual lagoons up to 400 ha.
The Ebro delta (Tarragona) is the largest and most important wetland on the Spanish Mediterranean coast, being comparable only to the Camargue in France. It is well known mainly because of its importance in the migration of birds between Europe and Africa, but its invertebrate fauna is still poorly known, and there is virtually no information about the aquatic Coleoptera (Appendix). The Ebro river was a closed basin separated from the sea by the Catalonian littoral mountains until the end of the Miocene. During the Pliocene and Pleistocene, climatic changes and associated shifts in eustatic sea levels produced a series of progressions and regressions of the different consecutive deltas (Nelson and Maldonado, 1990; Marifio, 1992). During the Holocene, with a stable sea level, deposition rates reduced. What constitutes the emerged area of the present delta-about 300 km2, extending more than 20 km into the sea-evolved in the last 2000 years, when deforestation and overgrazing of the Ebro basin trebled sediment supply and doubled offshore deposition rates compared with natural Holocene amounts (Nelson and Maldonado, 1990). The construction of the main irrigation channels on both sides of the River Ebro in 1857 and 1912 respectively favoured the expansion of the rice fields, which now occupy the major part of the delta. Only about 20% of its surface is not occupied for agricultural, urban or tourist uses, mainly in its periphery (Folch, 1988). In the last 50 years the construction of dams had reduced the sediment supply to less than 5% of the former amount, leading to

Tab
le 1
. C
oast
al w
etla
nds
incl
uded
in th
e st
udy.
Wet
land
Si
te
Des
crip
tion
AD
: Adr
a A
lbuf
era
Nue
va
Fres
hwat
er, v
eget
ated
, eut
roph
ic,
SA: S
anta
Pol
a an
d To
rrev
ieja
Sa
linas
Sa
line,
not
veg
etat
ed, s
hallo
w,
ED:
Ebro
del
ta
AU
FA: L
'Auf
acad
a Sa
line,
not
veg
etat
ed, s
mal
l dun
e
larg
e la
goon
larg
e la
goon
s
pond
s Sa
line,
smal
l pon
ds, w
ith a
ve
geta
tion
belt
Silte
d po
nd w
ell-v
eget
ated
Wel
l-veg
etat
ed la
goon
and
ch
anne
ls o
ccas
iona
lly co
nnec
ted
with
the
sea
TAN
C:
La T
anca
da
Salin
e, sm
all d
une
pond
, with
a
vege
tatio
n be
lt V
EDI:
Esta
ny d
el C
anal
Vel
l: di
tch
Arti
ficia
l ditc
h be
side
the
fen,
w
ell-v
eget
ated
V
EFE:
Est
any
del C
anal
Vel
l: fe
n La
rge
fen
with
den
se re
ed, i
n th
e ed
ge o
f a
perm
anen
t la
goon
PI
DR
: Seq
uia
Maj
or:
drai
nage
Te
mpo
rary
, sha
llow
, wel
l-veg
etat
ed
chan
nels
ch
anne
ls, w
ith g
roun
dwat
er
sprin
gs
PIIR
: SZ
quia
Maj
or: i
rrig
atio
n Pe
rman
ent,
deep
, eut
roph
ic,
chan
nel
wel
l-veg
etat
ed c
hann
el
TO: T
orre
dem
barr
a G
AV
I: C
anal
Gav
ina
Shal
low
chan
nel c
onne
cted
with
ENC
A: L
'Enc
anyi
sada
ERM
S: E
ms
de C
asab
lanc
a EU
CA
: Els
Euca
liptu
s W
ell-v
eget
ated
cha
nnel
s M
AR
Q:
Plat
ja d
e la
Mar
ques
a
the
sea
Shal
low
, lar
ge d
une
lago
ons,
w
ith s
carc
e ve
geta
tion
Smal
l pon
ds w
ith a
veg
etat
ion
belt
Lago
ons
and
pond
s w
ith d
iffer
ent
salin
ity a
nd a
mou
nt o
f veg
etat
ion
MU
NT
: Els
Mun
tany
ans
NA
NI:
Cal
Nan
i
Fluv
ia d
elta
LD:
Llob
rega
t de
lta
AE:
Aig
uam
olls
de
1'Em
pord
a
PS: P
ined
a de
Sal
ou
Sam
plin
g da
tes
15.0
4.85
23.0
8.94
16.0
1.93
, 22.
01.9
4, 2
5.03
.95,
24.
06.9
5
17.0
5.94
17.0
5.94
21
.08.
84, 2
7.05
.94,
24.
06.9
5 26
.07.
92, 7
.0 1.
95, 2
4.06
.95
17.0
5.94
17.0
5.94
, 24.
06.9
5
17.0
5.94
, 24.
06.9
5
28.0
3.93
, 28.
05.9
4, 1
7.07
.94,
7.0
1.95
28.0
3.93
, 8.0
5.94
, 17.
07.9
4, 7
.01.
95
4.06
.94,
29.
04.9
5
4.06
.94,
29.
04.9
5
14.0
5.94
Not
sam
pled
s m 2 P
W m m 3 c: ?1
P s m W
P
0
W m 5 e
h)
W

Hyg
rolu
s par
aUel
ogra
mm
us
flyW
uS q
uadt
igut
talu
s H
ydat
icus
sem
nige
r G
raph
odor
us ci
nere
us
Aig
uam
olls
de
I'Em
pord
A
Del
ta d
el L
lobr
egat
G
ynnu
s mnu
lus
Hyd
ropo
rus n
orm
and s
la1
Hyg
IUlu
s par
eleb
gram
mus
H
Ydm
poN
s non
nand
s /a
t A
cWs
cana
kula
tus
Hyd
abcu
r lra
nsve
mah
s na
plU
s vane
gatu
s G
raph
oder
us ci
nere
us
Mel
hles
cnb
rate
Uus
c~
rale
lus
Cyh
sler
rnpu
ncta
fus
Hyd
q$yp
hus
s@rt
alel
us
Heb
phoN
s B
usln
s H
ydm
eLph
ut u
gra
leh
s *
de
I'E
bre
Hem
phyd
ms
mus
icus
H
ydm
@Ju
s Peu
s
Gra
phod
orus
une
reus
Och
lheb
lus w
nds
1 H
elop
horu
s Sus
lns
Ber
osus
lw
ws
B
eros
us ja
echr
C
hasm
ogen
us hv
orni
cus
Hyd
mph
#us
pkeu
s O
chlh
stnu
s de
nhfe
r O
chlh
etnu
s mac
ulal
us
Och
lheb
ius
subp
clus
Eb
Sal
inas
de
San
ta P
ola
Sal
inas
de
Torr
evie
ja 'I
Alb
ufer
a d
e A
dra C
anlh
ydru
s di
ophl
hslm
us
Cyb
isle
r vul
nera
lus
Mel
hles
crib
refe
lus
Och
lheb
iur c
omge
rus
Och
lheb
fus d
o@ga
do,
Och
Ute
bius
subp
iclu
s
Q;
Esl
any
del C
anal
Vel
l
Mar
ques
a
L'E
ncan
ylsa
da sQ
y
L'A
ulac
ada
Em
s de
Cas
abla
nca L
Els
Euc
alip
lus
- 2 m
m Tz h
Y b ?
Figu
re 1
. C
oast
al w
etla
nds
stud
ied,
with
som
e of
the
ir m
ost i
nter
estin
g sp
ecie
s (se
e A
ppen
dix
for
thei
r ge
nera
l dis
trib
utio
n an
d st
atus
).

WATER BEETLES FROM THE EBRO DELTA 125
an increase in erosion (Nelson and Maldonado, 1990). Sediment compacting in the Holocene delta, further reduction of the discharge due to planned water transfers, and possible increases in sea level due to global climate changes would add to the regression of the emerged surface, one of the major threats for the conservation of the delta ecosystems (Nelson and Maldonado, 1990; Mariiio, 1992; Prat and Ibaiiez, 1995).
The two sites in Pineda de Salou (Tarragona) are the only remnants of a much larger coastal wetland which has been progressively drained since the mid 17th century, mainly for agriculture and to eradicate malaria. Only a permanent irrigation channel (Skquia Major) and some temporary drainage channels remain, fed by groundwater and by rain. Their original invertebrate fauna is almost completely unknown.
Els Muntanyans, in Torredembarra (Tarragona) are a series of shallow coastal lagoons parallel to the coast, which fill with sea water during storms in winter and spring. Originally some of them also received fresh water from small temporary streams created by rain water. However, the construction of the railway between the lagoons and the land behind has disrupted this natural pattern of rain drainage, and now only some agricultural and urban effluents reach the lagoons. In some marginal areas, such as in Canal Gavina, there is still a mixture of sea and rain water.
The Delta del Llobregat (Barcelona) is one of the best known places in Spain entomologically (see Appendix), mainly because of its proximity to Barcelona. Unfortunately, most of the traditional sites have been completely destroyed or badly damaged-such as Can Tunis, now completely urbanized, or the Llobregat river itself, which is highly polluted. Some of the species previously recorded from the area are considered locally extinct (Ribera et al., 1995). Only about 5% of the 130 km2 of the delta retains natural vegetation (Folch, 1988).
The Aiguamolls de 1’Emporda are depressions lying behind coastal dunes, with occasional connection to the sea at high tides or during storms. Lagoons and ponds have different degrees of salinity, depending on the distance to the sea and on the existence of groundwater springs or superficial freshwater inlets from streams or drainage ditches. This site was not sampled, the only data being from published sources (Appendix).
Sampling
Water beetles were sampled with a D-framed pond net with a mesh size of 0.5 or 1 mm. Sites were worked across a representative range of microhabitats until no further taxa were collected. Small beetles such as Ochthebius spp. living in the edges of saline lagoons were sought for by splashing the banks with water and picking off the floating beetles using a strainer. All species were recorded on a presence/absence basis. Such a simple approach has proved valuable for similar studies in the past (e.g. Foster et al., 1990).
Multivariate analysis
A classification of the communities of aquatic Coleoptera was based on 13 sites and 59 species (Table 2) using TWINSPAN (Hill, 1979). Only sites for which detailed knowledge of the local distribution of the species was available were included in the analysis, to avoid the use of lists pooled from different habitats with vaned environmental conditions. The amount and type of vegetation, type of substratum, and salinity of the water of the sites included in the classification were broadly categorized in a qualitative scale (Table 3). Salinity in Mediterranean coastal habitats is highly variable, due to seasonality of the rainfall and changes in the water level, and in some of the studied sites individual measurements may be misleading. The origin of the water, the presence of Salicorniu spp in the surrounding area, and the habitual presence of marine fauna (usually Polychaeta and Decapoda), were used as a qualitative, indirect measure of saline regime.
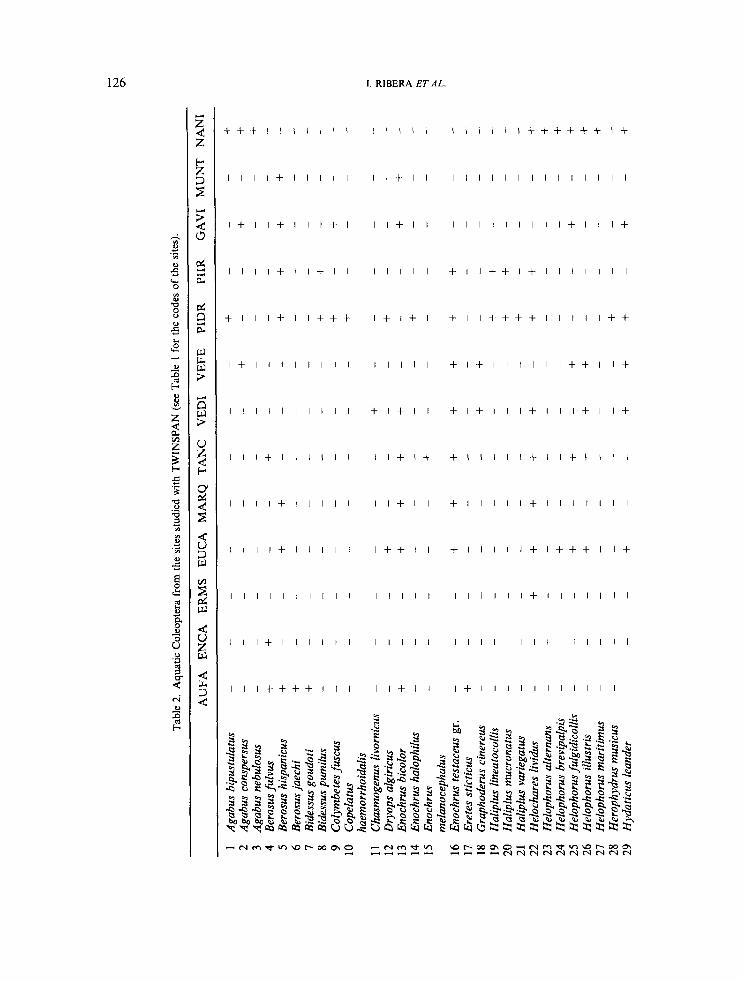
Tabl
e 2.
Aqu
atic
Col
eopt
era
from
the
sites
stu
died
with
TW
INSP
AN
(see
Tab
le 1
for
the
code
s of
the
sites
).
AU
FA
E
NC
A
ER
MS
EU
CA
M
AR
Q
TA
NC
V
ED
I V
EFE
P
IDR
PI
IR
GA
VI
MU
NT
N
AN
I
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
Agab
us b
ipus
tula
tus
Agab
us c
onsp
ersu
s Ag
abus
neb
ulos
us
Bero
sus f
ulvu
s Be
rosu
s his
pani
cus
Bero
sus j
aech
i Bi
dess
us g
oudo
ti Bi
dess
us pu
milu
s C
olym
bete
s fus
cus
Cop
elat
us
haem
orrh
oida
lis
Cha
smog
enus
livo
rnic
us
Dry
ops
algi
ricu
s En
ochr
us b
icol
or
Enoc
hrus
hal
ophi
lus
Enoc
hrus
rn
elan
ocep
halu
s En
ochr
us te
stac
eus g
r.
Eret
es s
tictic
us
Gra
phod
erus
cine
reus
H
alip
lus
linea
toco
llis
Hal
iplu
s muc
rona
tus
Hal
iplu
s va
rieg
atus
H
eloc
hare
s liv
idus
H
elop
horu
s alte
rnan
s H
elop
horu
s bre
vipa
lpis
H
elop
horu
s ful
gidi
colli
s H
elop
horu
s ill
ustr
is
Hel
opho
rus m
ariti
mus
H
erop
hydr
us m
usic
us
Hyd
atic
us le
ande
r
3
b
Y a.
P

30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
Hyd
robi
us fu
scip
es
Hyd
roch
us
smar
agdi
neus
H
ydro
glyp
hus p
usill
us
Hyd
rogl
yphu
s si
gnat
ellu
s H
ydro
philu
s pic
eus
Hyd
roph
ilus p
ista
ceus
H
ydro
poru
s lim
batu
s H
ydro
poru
s te
ssel
latu
s H
ydro
poru
s va
gepi
ctus
H
ydro
vatu
s cu
spid
atus
H
ygro
tus
conf
luen
s H
yphy
drus
au
bei
Ilybi
us m
erid
iona
lis
Lacc
ophi
lus h
yalin
us
Lacc
ophi
lus m
inut
us
Lacc
ophi
lus p
ontic
us
Lim
noxe
nus
nige
r M
ethl
es c
ribr
atel
lus
Neb
riop
orus
cer
isyi
N
oter
us c
lavi
corn
is
Not
erus
lae
vb
Och
theb
ius d
entif
er
Och
theb
ius d
ilata
tus
Och
theb
ius m
acul
atus
O
ch th
ebiu
s mar
inus
O
chth
ebiu
s sub
pict
us
Och
theb
ius v
irid
is 1
Pa
racy
mus
aen
eus
Pelto
dyte
s ro
tund
atus
Rh
antu
s su
tura
lis
Sp. N
o.
11
5 8
18
11
12
18
13
30
12
10
4 17

128 1. RIBERA ET AL.
Table 3. Some environmental characteristics of the sites classified with TWINSPAN.
Site Distance to sea (m) Salinity Vegetation cover Vegetation type Substratum
AUFA ENCA ERMS EUCA MARQ TANC VEDI VEFE PIDR PIIR GAVI MUNT NAN1
100 300 700 1000 300 300 1500 1500 600 600 300 100 1000
3 2 1 2 2 3 0 1 0 0 2 3 0
1 2 3 3 3 2 4 4 3 4 3 2 2
1 3 3 4 4 3 2 3 4 4 2 3 3
3 1 2 2 1 2 2 2 1 2 1 2 1
Salinity: 0 drainage or rain fresh water; 1 water of mixed origin, without Salicornia sp. or marine fauna; 2 water in areas with Salicornia sp.; 3 sea water with marine fauna. Vegetation cover: 1 no vegetation; 2 belt of emergent macrophytes; 3 less than 50% of the surface covered with macrophytes; 4 more than 50% of the surface covered with macrophytes. Dominant vegetation type: 1 none; 2 algae and macrophytes; 3 emergent macrophytes; 4 emergent and submerged macrophytes. Substratum: 1 silt or fine mud; 2 sand with mud; 3 sand.
Geographical distribution of the species
For the purpose of this study the species were classified as: (1) Trans-Iberian (present also in North Africa and France); (2) Iberian endemics sensu lato (Iberia plus southernmost France); (3) Species with a northern distribution (with the southern limit of their distribution within the Iberian peninsula); (4) Species with a southern distribution (with the northern limit of their distribution within the Iberian peninsula); and (5) Coastal circum-Mediterranean species (present only in coastal sites around the Mediterranean) (Appendix).
RESULTS
Classification of aquatic Coleoptera assemblages from coastal wetland habitats
In interpretation of the analysis it should be understood that some of the species have wider ecological tolerances than those reflected in this study, in which only a limited range of habitats was investigated. In consequence, some results cannot be considered representative of general autecology.
The first major split among the studied communities separated the habitats with water of mainly sea origin (groups 1 and 2) from the rest (groups 3 to 5) (Figure 2). Indicator species for groups 1 and 2 were Berosus fulvus and Hydroporus limbatus. Ochthebius subpictus also had a strong association with this group, occurring in the four sites, although it was also found in some of the sites in the other groups (see below). Group 1 included dune ponds without vegetation in L‘Aufacada, very close to the sea and filled with sea water from storms, and the lagoons in Els Muntanyans, also close to the sea, filled by storms and with no or little vegetation (Table 3) . Berosus hispanicus was found in both sites, although this species occurs also in other coastal sites as well as in habitats with bare substrata in central Spain (Ribera and Aguilera, 1995). Two of the circum-Mediterranean species with restricted distribution found in this study, Ochthebius dentifer and Berosus jaechi (see below), occur regularly in the small dune ponds in L’Aufacada. Group 2 included small ponds in the Ebro delta (marginal areas in La Tancada and L’Encanyisada), with sea or mixed water and a belt of vegetation outside the water (Scirpus spp. or Salicornia spp.). A species with strong preference for this group was Purucymus aeneus, which occurs regularly in coastal salt pans.

WATER BEETLES FROM THE EBRO DELTA
I E.
I
1 29
bicolor
3 [i] MUNT
1 f (E.bicolor)\ (E.~.esraceiis gr) (H.signatellirs) C O . siibpictiisy
4
M M Q VEDI VEFE
Figure 2. TWINSPAN dendrogram of the study sites. Species names are the indicators and the strongly associated species (in brackets) for each of the dichotomies. The five endgroups include the codified name of each site (see Table 1).
Some of the species had no defined preference, having a wide tolerance for salinity and other environmental conditions. These included Laccophilus hyalinus, Noterus clavicornir, Ochthebius dilatatus, Helophorus illustrir, and Helophorus fulgidicollis. Some were coastal species, e.g. H. illustris, but others have been found in central Spain, e.g. H. fulgidicollk in some arid areas (Angus, 1992; Ribera and Aguilera, 1995).
The indicator species for the sites with water from drainage, rain or with a mixed origin (groups 3 to 5) was Rhantus suturalis. Enochrus bicolor was the indicator species for groups 3 and 4. It is mostly associated with highly mineralized waters both on the coast and in inland (Ribera and Aguilera, 1995). Group 3 included only Cal Nani, a series of small ponds in a clay substratum with a belt of macrophytes. Group 4 included five sites with a mixture of sea water and rain water from drainage, such as Canal Gavina; lagoons with occasional connections with the sea, such as temporary marginal areas in Estany del Canal Vell; or lagoons and temporary inundated marshes in Platja de la Marquesa and Els Eucaliptus. Four of them had 0. subpictus and E. bicolor. All had a more developed vegetation, dominated by reed (Phragrnites spp.) (Canal Vell, Platja de la Marquesa and Els Eucaliptus), or submerged macrophytes (Canal Gavina and ditch in Canal Vell) (Table 3). Other species with a strong preference for this group were Enochrus testaceus gr., Hydroglyphus signatellus and Ochthebius subpictus. All of them are mainly coastal species, although widespread in other arid areas in central Spain (Ribera and Aguilera, 1995).

130 I. RIBERA ET AL.
Group 5 included three sites with fresh water and dense vegetation ( e g Typha angustijblia, Potamogeton nodosus, Chara spp., Iris pseudacorus, Carex hispida, Schoenus nigricans and others in Pineda de Salou). Indicator species for this group were Noterus laevis and Bidessus purnilus. Other species with preferences for this group were Methles cribratellus, together with all the species found exclusively in Pineda de Salou (see Table 2).
Habitats of the rarer species
The habitat of the dytiscid Methles cribratellus and the hydrophilid Chasrnogenus livornicus is of special interest, owing to the scarcity of published data and to their rarity. M . cribratellus was found both in Pineda de Salou and in the Ebro delta among detritus or at the base of aquatic vegetation, very close to the bottom or near the edges, a similar habitat to that recorded by Foster (1986) from Almeria. Hydroporus normandi s. lat. (a species complex, H. Fery, personal communication, 1995), is another species with a similar morphology (small, 3.2-3.7mm, narrow and flat, parallel-sided, and very shiny) and habitat that occurs in the Mediterranean coast. The taxon has been found in an area with common reed (Phragmites australis) in Torredembarra, with no open water and in a deep layer of dead plant fragments (near Els Muntanyans, 29.4.1995 IR), and in Delta del Llobregat, in a small pond apparently spring-fed and covered with Phragmites spp. and other macrophytes (near Ca L‘Arana, 14.5.1994 DTB). The latter two species are very poor swimmers (unpublished observations), probably adapted to crawl among detritus and rotten or dense vegetation (Ribera and Nilsson, 1995).
C. Zivornicus was found in a drainage channel in Estany del Canal Vell, among a dense layer of filamentous algae and other macrophytes, and with just a few centimetres of very cloudy water. The species has since been found in similar conditions in a natural habitat in the same delta, in Aigiies Mortes, an almost completely silted lagoon with only a very shallow layer of water during a short time of the year (J. Fresneda, personal communication, 1995).
Biogeographical remarks on the fauna of the Mediterranean coast of Spain
Species with a northern distribution The Ebro delta is the southernmost known western Mediterranean site in the distribution of the large Holarctic dytiscid beetle Graphoderus cinereus, which occurs in well preserved temporary fens in the north east of Spain. It has been found in Laguna de Pitillas (Navarra; Ribera and Aguilera, 1996), Estanys de Canadal (Girona; Ribera et al., 1994), Aiguamolls de 1’Emporda (Girona; Rosales and Lafuente, 1993), and Delta del Tordera (Barcelona; Sabater et al., 1986). There are some old records from the Delta del Llobregat and nearby localities in Barcelona (Cuni, 1888; Traizet, 1895), but the species has not been found in the area this century. Hydrophilus piceus, a Eurosiberian hydrophilid, is another large species with its southern limit in the Mediterranean coast. The species has been found further in the south than the Ebro delta in Oropesa (Catellbn) (E. Rico, personal communication, 1996). It occurs in some of the same habitats as Graphoderus (Valladares and Ribera, 1993; Ribera et al., 1995), in general coexisting with Hydrophilus pistaceus, a western Mediterranean species common in the Iberian peninsula.
Other European large species that reach the southern limit of their distribution in the western Mediterranean in coastal wetlands are Hydaticus serniniger, Ilybius quadriguttatus and Hydrochara caraboides. These were recently found in the Aiguamolls de 1’Emporda and in Estanys de Canadal (Girona) (Rosales and Lafuente, 1993; Valladares and Ribera, 1993; Ribera et al., 1995; see Appendix).
There are some old records of other European species from the Delta del Llobregat, such as the dytiscids Hydaticus transversalis (Cuni and Martorell, 1876) and Acilius canaliculatus (Traizet, 1895). For both of them this would represent the southern limit of their respective distributions in the western Mediterranean, but none of these species has been recorded in the area during this century. Hernando and Aguilera (1 994)

WATER BEETLES FROM THE EBRO DELTA 131
found one of the original specimens of H . transversalis collected by Martorell in a lagoon in Badalona, an area now completely urbanized, but other specimens have not been found. Most of the collections were destroyed or lost during and after the civil war. As discussed by Ribera et al. (1 995) some of these records may be trusted, although the species are probably now extinct in the Iberian peninsula.
Species with a southern distribution Methles cribratellus is an Ethiopian species that reaches isolated localities in North Africa, some Greek islands, Sicily and Sardinia (Franciscolo, 1979; Burmeister et al., 1987). This species was previously found in the Iberian peninsula in Baixo Alentejo (Franciscolo, 1966), Almeria (Foster, 1986) and Pineda de Salou (Hernando and Aguilera, 1994). The localities in Tarragona appear to be at the northern limit of the distribution of this species.
Herophydrus musicus is another north African species that has been recently found in the same habitat as M . cribratellus in Pineda de Salou (Hernando and Aguilera, 1994). It was previously found in the south-east corner of the Iberian peninsula (Millin and Soler, 1990). The localities in Tarragona are again the northernmost limit of its known distribution.
Among the large Dytiscidae, Cybister tripunctatus is a species widespread in the Ethiopian region and which has been recorded from different coastal habitats as far in the north as the Delta del Llobregat (Rico et al., 1990). Cybister vulneratus was last found in the Iberian peninsula in the Albufera de Adra by R. B. Angus in 1962 (personal communication), together with Canthydrus diophthalmus, recorded by Montes and Soler (1985) from the same site. Both are Ethiopian species reaching isolated localities in the southern Palacarctic region. They were found in small pools which have been destroyed by the development of huertas (vegetable growing areas) around the remaining lagoons. Despite recent visits to the Albufera and nearby areas by some of the authors, neither species has been found again, and their status in the Iberian peninsula is uncertain, but they are certainly endangered-if not extinct. Cunrhydrus has also disappeared from Sicily and the Balearics (Montes and Soler, 1985), and in its only other European site, Sardinia, records dated back to the last century (Franciscolo, 1979; Burmeister et al., 1987). C . vulneratus was otherwise known in Europe from Sicily (Franciscolo, 1979), at a site which is also apparently lost. Both species may well be extinct in Europe.
Coastal circum- Mediterranean species Chasmogenus Iivornicus was previously known only from isolated sites in the Eastern Mediterranean, in Italy, the former Yugoslavia, Greece and Israel (Hebauer, 1992). The record from the Ebro delta expands its distribution to the western Mediterranean, establishing it as a circum-Mediterranean species. This is the only European representative of this mostly Ethiopian genus.
Helophorus illustris. Berosus jaechi, Ochthebius dentifer and Ochthebius subpictus are circum- Mediterranean species which probably had a widespread distribution across the Mediterranean coast in historical times, but due to habitat fragmentation and loss they have been reduced to isolated populations with restricted distributions (Jach 1991a, 1991b; Schodl, 1991; Angus, 1992; Aguilera and Ribera, 1994). For the first three species, the only known populations in the Iberian peninsula are those reported here.
Iberian endemics sensu lato Four species among those found in coastal, stagnant habitats can be considered Iberian endemics in a broad sense (Appendix). Hydroporus vagepictus is a common species inhabiting a wide range of habitats from southern France to central Spain, with some isolated populations in the south (Rico et al., 1990; Ribera et al., 1995). As noted above, the taxonomic status of Hydroporus normandi requires review, and it is not possible to assert with certainty its distribution, but the form under discussion is most probably not exclusively coastal. Ochthebius delgadoi is a SE Iberian species mostly found in running waters (Jach, 1994). Thus, the only possible true coastal endemic is Dryops hispanus, described from Barcelona and known only

132 1. RIBERA E T A L .
from the holotype (Olmi, 1972), but also with an uncertain taxonomic status (E. Rico, personal communication, 1996).
DISCUSSION
Classification of coastal wetland beetles
In evaluating the importance of the salinity of the water in the classification of the studied assemblages of water beetles it has to be taken into account that salinity is related with other variables, such as distance to the sea or structure and development of the soil. It is possible that for some species the limiting factor is not salinity itself, but some other associated characteristic, such as suitable soil structure for pupation or lack of predators in saline waters.
The presence of typical coastal species in some arid areas in central Spain is a common pattern among several groups of aquatic Coleoptera (see Ribera and Aguilera, 1995 for detailed examples). This can be the result of a secondary colonization of their often highly mineralized waters, but also of the permanence of relict Miocenic populations originally inhabiting the shores of old continental seas (such is the case of some interstitial crustaceans: Coineau, 1994). The coastal water beetles found inland are in general good dispersers, and in consequence it is difficult to establish the origin of their populations.
The structure and amount of vegetation were other important factors in the classification of the communities. For phytophagous or detritivorous groups (e.g. Hydrophilidae or Hydraenidae) the type and amount of vegetation can have a direct influence. However, predators (Dytiscidae and Noteridae) are probably more related to the spatial structure that the vegetation provides. This is an important factor influencing the size and shape of the species that can be found, and probably also influences their swimming strategy (Ribera and Nilsson, 1995).
Biogeographical considerations
The absence of endemics among coastal species can be attributed to the temporal dynamics of the coastal lagoons, and their relatively recent geological origin. The species that populate these habitats are in general good colonizers-most of the sites are temporary over a scale of a few hundred years. Species presently restricted to these areas are not true endemics which evolved in situ, but remnant populations of former wide distributions.
The role of the Quaternary glaciations in the distribution of the invertebrate fauna of the Iberian peninsula, and in the Mediterranean basin in general, is not completely understood, and may be overestimated owing to the preponderance of north European studies. Some species formerly thought to have been locally extinct due to glaciations persisted in refugia (such as the walnut, Juglans regiu: Carrion and Shnchez-G6mez, 1992), and it is probable that at least some of the numerous stepparian species of plants and invertebrates found both in central Arag6n and the Asian steppes are relict populations that have persisted in the area through the Pleistocene (e.g. Braun-Blanquet and Bolos, 1957; Miracle, 1982; see Ribera and Aguilera, 1995 for a list of examples). The major patterns in the biogeography of the circum- Mediterranean fauna were determined during the Oligocene and the Miocene, when the formation of a continuous landmass allowed dispersal throughout the Mediterranean basin. The subsequent tectonic activity resulted in the isolation of different areas and the speciation of numerous taxa across the Mediterranean before the end of the Miocene (Oosterbroek and Arntzen, 1992).
During the frequent climatic shifts in the Quaternary, insects moved around Europe tracking the habitats that suited their autecological requirements (Coope, 1995). The Mediterranean coast of Spain could have provided a corridor, with continuous displacements of species that have resulted in a mixed series of assemblages. As a striking example, in the same site in the Ebro delta (Estany del Canal Vell), typical

WATER BEETLES FROM THE EBRO DELTA 133
Table 4. Composition of the communities of aquatic Coleoptera in the studied coastal wetands (see Table 1 for the codes of the sites).
Ds AD % SA % ED % PS % TO % LD % AE %
N 0 0.0 0 0.0 3 8.1 2 6.5 0 0.0 9 11.7 8 14.8 S 3 15.8 1 11.1 3 8.1 3 9.7 0 0.0 1 1.3 0 0.0 M 0 0.0 1 11.1 5 13.5 0 0.0 1 9.1 1 1.3 1 1.9 E 0 0.0 1 11.1 0 0.0 1 3.2 1 9.1 3 3.9 1 1.9 T 16 84.2 6 66.7 26 70.3 25 80.6 9 81.8 63 81.8 44 81.5
Total 19 9 37 31 11 77 54
Ds, distribution: N, Northern species; S, Southern species; M, Coastal circum-Mediterranean species; E, Iberian endemics semu lato; T, trans-Iberian species.
central and northern European species, such as Hydrophilus piceus or Graphoderus cinereus, coexist with African or circum-Mediterranean ones, such as Methles cribratellus or Chasmogenus livornicus, all of them at the limit of their respective distributions. The gradual change in the composition of the fauna from north to south is clear (Table 4). However, although the replacement of northern by southern species seems to be steady, the key role of the Ebro delta can still be appreciated: no northern species were found in the coastal sites studied south of it, and only few southern species had isolated coastal populations in the north.
The Ebro delta is also a key biogeographical area for several species of other organisms. Among plants, some southern species have their northern limit here (e.g. Limoniastr~m monopetalum, Lonicera biffora, Tamarix boveana, and Zygophyllum album, the last being the only known population in continental Europe), and some northern ones their southern limit (e.g. Fraxinus angustifolia ssp. oxycarpa, Alnus glutinosa) (Folch, 1988). The same pattern is found among many different groups, such as Mollusca (Sacchi, 1957) or ground beetles (Zaballos and Jeanne, 1994). In the case of the aquatic Coleoptera, the Ebro valley is a biogeographical barrier of much more importance than the Pyrenees, the Iberian fauna sensu strict0 being largely confined to the area south-west of the river. It is usually overlooked that the narrowest strip of land connecting Iberia with mainland Europe does not cross the Pyrenees, but the Ebro valley.
Conservation value of coastal habitats
The conservation value of the aquatic Coleoptera of the studied coastal wetlands is indisputedly high, but the reasons for this are not immediately apparent. The perceived value cannot depend solely on species richness, though this is high for many of the sites, with one-fifth of the Iberian fauna being found in the region as a whole. Species paucity also exists. The saline habitats separated out by TWINSPAN, and the salinas in Santa Pola and Torrevieja, were the sites with fewest species, but their fauna was highly characteristic, contributing an additional faunal component. The level of endemism cannot be a factor in dictating ‘high’ conservation quality as there is no evidence of true endemism in these coastal habitats. The primary criterion for high quality must therefore be associated with the occurrence of species with narrow ecological requirements and restricted distributions. Most of these species are at the limits of their distributions, being the only known populations in the Iberian Peninsula--or in Europe in the case of Ethiopian species. This presents a paradox as such species might reasonably be ignored when considering what constitutes the important, central elements within a region.
The natural conclusion is that the conservation value of the area depends on the mixture of species of different origin. The only alternative would be to reject the perception that this area has high value, and to

134 I. RIBERA ET AL.
claim that such a mixture of species indicates that it has no importance. This approach is unacceptable, nor is there any room for compromise; the area is of high value. At this stage, with an incomplete database for the circum-Mediterranean area, there are no easy means of providing a series of comparisons with other coastal complexes. The rate of loss of Mediterranean coastal complexes demands the drawing of attention to this region now.
The specificity of the fauna of temporary habitats has been documented for a wide range of taxonomic groups and environmental conditions (e.g. Wiggins et al., 1980; Collinson et al., 1995). Coleoptera are one of the dominant groups in temporary habitats (Larson, 1985; Jeffries, 1994; Collinson et al., 1995), and patterns in their biology and distribution have been used as an indication of the general characteristics of the system and of its biological and conservation quality (see e.g. Foster, 1991; Ribera and Foster, 1992).
The major effect of recent development in the Mediterranean coast of Spain has been habitat fragmentation and loss (Folch, 1988). This loss has accelerated in the last 50 years, following massive tourist developments on most of the coast. If we extend to water beetles the pattern of the Mediterranean flora, for which very few extinctions have been documented (Greuter, 1995), or that of the Quaternary insects in general (Coope, 1995), it may be assumed that in recent times this has had no major effect on the regional species richness, leading instead to local population extinctions and a possible loss of genetic biodiversity. The still rich and interesting fauna of the remaining relatively well preserved areas studied here would support this assumption, although the original fauna in most cases is unknown. The Ebro delta is particularly interesting, where all five exclusively coastal species can be found, some of them being the only known Iberian populations (Appendix). The lower Ebro also has one of the last known populations of Margaritifera auricularia in the world, a severely endangered species of freshwater bivalve (Altaba, 1990; Woodward, 1996), and other endangered or scarce species of macroinvertebrates typical of large European rivers (e.g. Raptobaeropus tenellus and Ephoron Virgo, both mayflies: Muiioz and Prat, 1994). Although the survey of the coastal wetlands is far from being comprehensive, and more species and populations are likely to be found in the near future, the suitable habitats are becoming increasingly scarce, and there is the real possibility that species may become extinct without being discovered.
Some of the Iberian Mediterranean endemics are not considered to be specially endangered, despite their very restricted distributions, because they are locally abundant (e.g. Ojeda et al., 1995, for plants in heathland communities, or Fernandez-Pedrosa et al., 1995 for Aphanius iberus, an endemic fish with only 15 known populations in the Iberian peninsula, several of them in the sites studied here). However, the effect of habitat destruction and fragmentation on the species richness of communities, and on the extinction of local populations, is far from understood (Lawton, 1995). Most of the coastal habitats have temporary waters, and the species have metapopulation dynamics at a local or a regional scale. Unless detailed knowledge of the distribution and the status of the individual populations is available, the fact that a species seems to be widespread and locally abundant does not mean that its persistence is guaranteed. Some of the species have good colonization abilities and reproduction rates, and a few populations can be the source for a large area, such as the African butterfly Colotis evagore in southern Spain: the species can survive the winter only in some coastal microclimate enclaves, from where it extends each year to other areas where it can pass up to six generations in spring and summer, colonizing up to 60 000 km2 in one breeding season (Jordan0 et al., 1991). Despite the apparent abundance of the species in some years, the whole Iberian metapopulation is totally dependent on a few source populations on the coast, which can be easily depleted by an unusual cold winter. The identification of source populations is not always so clear as in this example, and if the species can reproduce continuously in sink habitats, or if generations overlap in long-lived organisms, this may provide misleading information on the population dynamics and the autecology of the species (Watkinson and Sutherland, 1995). The destruction of small, misidentified source habitats could lead to the loss of large sink populations. The time lag in the extinction of reduced, non- viable populations has also to be considered in the evaluation of the status of species with restricted distributions (Brooks and Balmford, 1996).

WATER BEETLES FROM THE EBRO DELTA 135
If the natural dynamics of habitat creation and destruction are interrupted, there is no possibility of renewal, and the present habitats will disappear-and with them their special communities. The fact that most of the places studied are already protected (under regional, national and international laws-Santa Pola and Torrevieja salines, the Ebro delta and Aiguamolls de 1’Emporda are included in the Ramsar Convention) is not a solution for the long-term effect of habitat fragmentation, unless the dynamics of the temporary habitats are guaranteed through appropriate management practices, something that at present is not even considered necessary. Protection is usually focused on the large, permanent lagoons (e.g. in both Ebro and Llobregat deltas, or in the Albufera de Adra), and some of the most interesting temporary habitats studied here are not included in the protected areas, which include only the surface of the lagoons. The effect of the exclusive protection of the permanent lagoons is clear in the Albufera de Adra, in which the surrounding land is now completely devoted to intensive vegetable growing. This has resulted in the destruction of the surrounding habitats and the loss of their exclusive invertebrate assemblages.
Some of the wetlands studied have no effective protection: such is the case of Pineda de Salou, in which the most interesting sites, the temporary channels, are highly endangered because of tourist development and urban development plans for the area, while the persistence of the permanent channel, with its less interesting fauna, seems to be guaranteed because of its agricultural use. Both types of habitats are strongly affected by human activity and, as mentioned above, represent the only remnant of a former coastal lagoon. However, the fauna of the irrigation channel is much poorer, both in the number and the interest of the species: the species with restricted distributions, which are more interesting from a biogeographic point of view (Haliplus variegatus, Methles cribratellus and Herophydrus musicus) occur only in temporary drainage ditches (Table 2). This difference is not restricted to aquatic beetles: in the irrigation channel Gambusiu holbrooki, a small, non-native coastal and estuarine fish occurs, which is displacing the Mediterranean endemic species (Fernandez-Pedrosa et al., 1995). The drainage channels, however, harbour one of the few known natural populations of Aphanius iberus, one of these endemic fish species. Whether these differences are due to impermanence or to other factors is difficult to assess. They may result from the presence of agricultural effluents in the irrigation channel, or to the existence of groundwater springs in some of the drainage channels.
The conservation value of marginal, temporary habitats has only recently been recognized, and their preservation requires specific management practices (Foster, 1991; Biggs et al., 1994). It is important that future conservation and management plans for the wetlands of this region take account of the wider aquatic community, and maintain the dynamic system which ensures the continuity of a wide range of freshwater conditions. General recommendations in Ranwell (1977) and Collinson et al. (1995) are applicable to a wider range of temporary coastal habitats:
1. It is necessary to survey the invertebrate fauna of the coastal sites before major changes in the environment are undertaken. Invertebrates have to be considered in assessing the conservation value of coastal habitats.
2. The water regime and quality should be preserved. A wide range of salinity has to be available in the area, ranging from sea spills in dune ponds to fresh groundwater in sources and springs. The random, unpredictable consequences of the Mediterranean climate should be preserved whenever possible, avoiding dredging or deepening of temporary ponds, and regular, predictable water level changes.
3. It is necessary to provide new habitats if the natural dynamic of the system is interrupted, e.g. by producing new scrapes in silted old lagoons, or allowing the formation of dune ponds, letting them erode and making way for the regeneration of new dunes at the strandline.
ACKNOWLEDGEMENTS
We are grateful to Robert Angus, Natalio Rosales, Javier Fresneda and Eugenio Rico for unpublished records of coastal aquatic Coleoptera. Our thanks also go to David McCracken, Martin Drake, and an anonymous referee for

136 1. RIBERA ET A L .
helpful comments on the text. The Scottish Agricultural College receives financial support from the Scottish Office Agriculture, Environment and Fisheries Department.
REFERENCES
Aguilera, P. and Ribera, I. 1994. ‘Berosus jaechi Schodl in the Iberian peninsula’, Latissimus, 4, 3. Altaba, C. R. 1990. ‘The last known population of the freshwater mussel Margaritifera auricularia (Bivalvia,
Angus, R. B. 1992. Susswasserfauna von Mitteleuropa. Band 20110-2: Insecta Coleoptera Hydrophilidae Helophorinae,
Biggs, J., Corfield, A., Walker, D., Whitfield, M. and Williams, P. 1994. ‘New approaches to pond management’,
Braun-Blanquet, J. and Bolos, 0. 1957. ‘Les groupements vigttaux du basin de 1’Ebre et leur dynamisme’, Anales de la
Brooks, T. and Balmford, A. 1996. ‘Atlantic forest extinctions’, Nature, 380, 11 5. Burmeister, E. G., Dettner, K. and Holmen, M. 1987. ‘Die Hydtadephaga sardiniens (Insecta, Coleoptera: Gyrinidae,
Haliplidae, Noteridae, Hygrobiidae, Dytiscidae’, Spixiana, 10, 157-1 85. Carrion, J. S. and Sanchez-Gbmez, P. 1992. ‘Palynological data in support of the survival of walnut (Juglans regia L.) in
the western Mediterranean area during last glacial times’, Journal of Biogeography, 19, 623-630. Coineau, N. 1994. ‘Evolutionary biogeography of the Microparasellid isopod Microcharon (Crustacea) in the
Mediterranean basin’, Hydrobiologia, 287, 77-93. Collinson, N. H., Biggs, J., Corfield, A., Hodson, M. J., Walker, D., Whitfield, M. and Williams, P. J. 1995.
‘Temporary and permanent ponds: and assessment of the effects of drying out on the conservation value of aquatic macroinvertebrate communities’, Biological Conservation, 74, 125-1 33.
Coope, G. R. 1995. ‘Insect faunas in ice environments: why so little extinction?’, in Lawton, J. H. and May, R. M. (Eds), Extinction Rates, Oxford University Press, Oxford, 55-74.
Cuni, M. 1888. ‘Insectos observados en 10s alrededores de Barcelona’, Anales de la Sociedad Espanlola de Historia Natural, 17, 133-191.
Cuni, M. and Martorell, M. 1876. Catdogo Metodico y Razonado de 10s Coledpteros Observados en Catalunla. Imprenta Tomis Gorchs and cia, Barcelona.
Ehrlich, P. R. 1995. ‘The scale of human enterprise and biodiversity loss’, in Lawton, J . H. and May, R. M. (Eds), Extinction Rates, Oxford University Press, Oxford, 214-226.
Falkenstrom, G . 1939. ‘Halipliden und Dytisciden aus der Iberischen Halbinsel, gesamment in Sommer 1935 von Prof. Dr. 0. Lundblad’, Arkiv For Zoologi, 31A, 1-22.
Fernandez-Pedrosa, V., Gonzalez, A., Planelles, M., Moya, A. and Latorre, A. 1995. ‘Mitochondria1 DNA variability in three Mediterranean populations of Aphanius iberus’, Biological Conservation, 72, 251-256.
Folch, R. (Ed.) 1988. Natura, lis o abus? Llibre blanc de la gestid de la natura als Paiios Catalans. 2nd edn. Editorial Barcino, Barcelona.
Foster, G. N. 1986. ‘Los caballeros’, Balfour-Browne Club Newsletter, 36, 10-12. Foster, G. N. 1991. ‘Conserving insects of aquatic and wetland habitats, with special reference to beetles’, in Collins,
N. M. and Thomas, J. A. (Eds), The Conservation of Insects and their Environments, Academic Press, London, 213-236. Foster, G . N., Foster, A. P., Eyre, M. D. and Bilton, D. T. 1990. ‘Classification of water beetle assemblages in arable
fenland and ranking of sites in relation to conservation value’, Freshwater Biology, 22, 343-354. Franciscolo, M. E. 1966. ‘Notes on Iberian Dytiscoidea, I: Methlinae Guignot, 1936, a subfamily of Dytiscidae
(Coleoptera) apparently new to Europe’, Proceedings of the Royal Entomological Society, London, Ser. B, 35, 11-1 5. Franciscolo, M. E. 1979. Fauna d’ltalia. Vol. XZV. Coleoptera: Haliplidae, Hygrobiidae, Gyrinidae, Dytiscidae. Edizioni
Calderini, Bologna. Greuter, W. 1995. ‘Extinctions in Mediterranean areas’, in Lawton, J. H. and May, R. M. (Eds), Extinction Rates,
Oxford University Press, Oxford, 88-97. Hebauer, F. 1992. ‘The species of the genus Chasmogenus Sharp, 1882 (Coleoptera), Hydrophilidae’, Acta
Coleopterologica, 8, 61-92. Hernando, C. and Aguilera, P. 1994. ‘Nuevos datos sobre Dytiscidae (Coleoptera) para la peninsula Iberica’,
ZAPATERI Revista aragonesa de entomologia, 4, 45-48. Hernando, C. and Fresneda, J. 1986. ‘Hydradephaga nous o interesants per a la peninsula Iberica (Coleoptera,
Haliplidae i Dytiscidae)’, Buflleti de la Uni6 Excusionista de Catalunya, 130, 18-21. Hill, M. 0. 1979. TWINSPAN. a Fortran Program for Detrended Correspondence Analysis and Reciprocal Averaging.
Cornell University, Ithaca, New York.
Unionoida): a conservation priority’, Biological Conservation, 52, 271-286.
Gustav Fischer Verlag, Stuttgart.
British Wildlife, 5, 273-287.
Estacidn Experimental Aula Dei, 5 , 1-267.

WATER BEETLES FROM THE EBRO DELTA 137
Jach, M. A. 1990. ‘Revision of the Palearctic species of the genus Ochthebius Leach V. The subgenus Asiobates (Hydraenidae, Coleoptera)’, Koleopterologische Rundschau, 60, 37-105.
Jach, M. A. 1991a. ‘Revision of the Palearctic species of the genus Ochthebius Leach VI. The marinus group (Hydraenidae, Coleoptera)’, Entomologica Basiliensia, 14, 101-145.
Jach, M. A. 1991b. ‘Revision of the Palearctic species of the genus Ochthebius Leach. VII. Thefoveolatus-group (Coleoptera, Hydraenidae)’, Koleopterologische Rundschau, 61, 61-94.
Jach, M. A. 1994. ‘Revision der palaarktischen Arten der Gattung Ochthebius Leach. XIII. Beschreibung neuer Arten aus Spanien, der Tiikei, RuPland und China’, Annalen des Naturhistorischen Museums Wien, 96B, 199-208.
Jeffries, M. 1994. ‘Invertebrate communities and turnover in wetland ponds affected by drought’, Freshwater Biology, 32, 603-612.
Jordano, D., Retamosa, E. C. and Fernandez Haeger, J. 1991. ‘Factors affecting the continued presence of Colotis evagore (Klug, 1829) in southern Spain’, Journal of Biogeography, 18, 637-646.
Lagar, A. 1951. ‘Los Hidrocintaros del Delta del rio Llobregat’, Graellsia, 9, 75-80. Lagar, A. 1967. ‘Coleopteros del Delta del rio Llobregat. VI nota’, Graellsia, 23, 71-79. Lagar, A. 1968. ‘Los Haliplidae de Cataluiia’, Miscelunea Zooldgica, 2, 65-73. Lagar, A. 1970. ‘Coleopteros del Delta del no Llobregat’, VII nota. Graellsia, 26, 43-57. Larson, D. J. 1985. ‘Structure in temperate predaceous diving beetle communities (Coleoptera: Dytiscidae)’, Holarctic
Ecology, 8, 18-32. Lawton. J. H. 1995. ‘Population dynamic principles’, in Lawton, J. H. and May, R. M. (Eds), Extinction Rates, Oxford
University Press, Oxfbrd, 147-163.
(Eds), Climatic Change and the Mediterranean, Edward Arnold, London, 304327.
Ibirica’, Boletin de la Asociacidn esparSola de Entomologia, 14, 272.
9, 455467.
europeo (Col. Dytiscidae)’, Annuli &I Museo Civivo di Storia Naturale (di Genova) Giacomo Doria, 85, 187-199.
Marifio, M. G. 1992. ‘Implications of climatic change on the Ebro delta’, in Jeftic, L., Milliman, J. D. and Sestini, G.
Millan, A. and Soler, A. G. 1990. ‘Herophydrus musicus (Klug, 1833) (Col. Dytiscidae): primera cita para la Peninsula
Miracle, M. R. 1982. ‘Biogeography of the freshwater zooplanktonic communities of Spain’, Journal of Biogeography,
Montes, C. and Soler, A. G. 1985 .‘Canthydrus diophthalmus (Reiche and Saulcy, 1855), nuevo Noterinae para el continente
Muiioz, I. and Prat, N. 1994. ‘Macroinvertebrate community in the lower Ebro river (NE Spain)’, Hydrobiologia, 286,
Nelson, C. H. and Maldonado, A. 1990. ‘Factors controlling late Cenozoic continental margin growth from the Ebro
Ojeda, F., Arroyo, J. and Maraiibn, T. 1995. ‘Biodiversity components and conservation of Mediterranean heathlands
Olmi, M. 1972. ‘The Palearctic species of the genus Dryops Olivier (Coleoptera, Dryopidae)’, Bollettino del Museo di
Oosterbroek, P. and Arntzen, J. W. 1992. ‘Area-cladograms of Circum-Mediterranean taxa in relation to
Prat, N. and Ibaiiez, C. 1995. ‘Effects of water transfers projected in the Spanish national hydrological plan on the
Ranwell, D. S . 1977. ‘Strategies for the management of coastal systems’, in Jefferies, R. L. and Davy, A. J. (Eds),
Ribera, I. and Aguilera, P. 1995. ‘Colebpteros acuiticos de la provincia de Huesca (Aragbn, Espaiia)’, ZAPATERZ
Ribera, I . and Aguilera, P. 1996. ‘Els Estanys de Capmany: the missing Spanish pingo (or palsa) fens?’, Latissimus, 7,
Ribera, I., Aguilera, P., Bilton, D. T., Fery, H., Fresneda, J., Hernando, C. and Foster, G. N. 1995. ‘Towards a critical
Ribera, I . and Foster, G. N. 1992. ‘Us0 de colebpteros acuiticos como indicadores biologicos (Coleoptera)’, Elytron, 6,
Ribera, I., Isart, J. and Rigil, J. A. 1994. ‘Cole6pteros acuiticos de 10s estanques de Capmany (Gerona). Hydradephaga’, Scientia Gerundensis, 20, 17-34.
Ribera, I. and Nilsson, A. N. 1995. ‘Morphometric patterns among diving beetles (Coleoptera: Noteridae, Hygrobiidae, Dytiscidae’, Canadian Journal of Zoology, 73, 2343-2360.
Rico, E., Pirez, L. C. and Montes, C. 1990. Lista faunktica y bibliografica de 10s Hydradephaga (Coleoptera: Haliplidae. Hygrobiidae, Gyrinidae, Noteridae, Dytiscidae) de la Peninsula IbPrica e islas Baleares. Asociacion Espaiiola de Limnologia, Madrid.
65-78.
delta to the western Mediterranean deep sea’, Marine Geology, 95, 419440.
in Southern Spain’, Biological Conservation, 72, 61-72.
Zoologia dell’Universita di Torino, 5 , 69-1 32.
Mediterranean palaeogeography’, Journal of Biogeography, 19, 3-20.
ecology of the lower river Ebro (NE Spain) and its delta’, Water Science and Technology, 31, 79-86.
Ecological Processes in Coastal Environments, Blackwell, Oxford, 51 5-527.
Revista aragonesa de entomologia, 5 , 7-34.
2-6.
checklist of Iberian water beetles-Some old records reconsidered’, Latissimus, 6, 3-7.
61-75.

138 1. RIBERA ET AL.
Rosales, N. and Lafuente, M. C. 1993. ‘Hydaticus seminiger (DeGeer) in the Iberian peninsula’, Latissimus, 3, 4. Sabater, F., Murillo, J. and Campeny, R. 1986. ‘Macroinvertebrados de las lagunas temporales de Ca L‘Estany de
Tordera (Catalunya, NE Espafia)’, VII Jornadas de la Asociacidn espariola de Entomologia (Sevilla), 1241-1 251. Sacchi, C. F. 1957. ‘Lineamenti biogeografici della Spagna mediterranea su basi malacofaunistiche’, Publicaciones del
Instituto de Biologia Aplicada, 25, 5-48. Schodl, S. 1991. ‘Revision der Gattung Berosus Leach 1. Teil: Die palaarktischen Arten der Untergattung Enoplurus
(Coleoptera: Hydrophilidae)’, Koleopterologische Rundschau, 61, 11 1-1 35. Traizet, 1895. ‘Notas sobre Anticidos e Hidrocantaros de 10s alrededores de Barcelona’, Actas de la Sociedad Espariola
de Historia natural, 24, 182-1 83. Valladares, L. F. and Ribera, I. 1993. ‘Sobre la presencia de Hydrochara caraboides (Linnaeus, 1758) e Hydrophilus
piceus (Linnaeus, 1758) en la Peninsula IbCrica (Coleoptera: Hydrophilidae)’, Zoologica Baetica, 4, 7-12. Watkinson, A. R. and Sutherland, W. J. 1995. ‘Sources, sinks and pseudo-sinks’, Journal of Animal Ecology, 64, 126130. Wiggins, G. B., MacKay, R. J. and Smith, I. M. 1980. ‘Evolutionary and ecological strategies of animals in annual
temporary pools’, Archiv fur Hydrobiologie, Supplement, 58, 97-206. Woodward, F. R. 1996. ‘Margaritifera auricularis (Spengler, 1793)’, in van Helsdingen P. J., Willemse, L. and Speight,
M. C. D. (Eds), Background Information on Invertebrates of the Habitats Directive and the Bern Convention. Part 111, European Invertebrate Survey, Killarney, 495-500.
Zaballos, J. P. and Jeanne, C. 1994. ‘Nuevo catalog0 de 10s carabidos (Coleoptera) de la Peninsula Ibtrica’, Monografias de la Sociedad Entomologica Aragonesa, 1, 1-1 59.
Appendix. Aquatic Coleoptera from the Mediterranean coast of Spain. ~ ~~~
Ds St AD SA ED PS TO LD AE
GYRINIDAE 1 Gyrinus caspius Menetries, 1832 2 Gyrinus dejeani Brull6, 1832 3 Gyrinus minutus Fabricius, 1798 4 Gyrinus urinator Niger, 1807
5 Haliplus guttatus Aubi, 1836 6 Haliplus lineatocollis (Marsham, 1802) 7 Haliplus mucronatus Stephens, 1832 8 Haliplus rubidus Perris, 1857 9 Haliplus variegatus Sturm, 1834
HALIPLIDAE
10 Peltodytes caesus (Duftschmid, 1805) 11 Peltodytes rotundatus (AubC, 1836)
12 Canthydrus diophthalmus (Reiche & Saulcy, 1855) 13 Noterus clavicornis (De Geer, 1774) 14 Noterus laevis Sturm, 1834
DYTISCIDAE Copelatinae 15 Copelatus haemorrhoidalis (Fabricius, 1787)
H ydroporinae 16 Bidessus goudoti (Castelnau, 1834) 17 Bidessus pumilus (AubC, 1836) 18 Graptodytes flavipes (Olivier, 1795) 19 Graptodytes varius (AubC, 1836) 20 Herophydrus musicus (Klug, 1833) 21 Hydroglyphus pusillus (Fabricius, 178 1) 22 Hydroglyphus signatellus (Klug, 1834) 23 Hydroporus analis AubC, 1836 24 Hydroporus limbatus AubC, 1836 25 Hydroporus normandi s. lat. 26 Hydroporus planus (Fabricius, 1781)
NOTERIDAE
T + T N T +
S R,X 1 T + T + / I
T
T T + T T S T S T T E? ? T
+ +
+ 15
+ 16
+ + +/6 +
+
t
+ + 14 + + +/4 +/4
+ + + +
11 16 10 16 8 10
15 8 16 8 13
10 10 16
+/8 16 +/8 16
8 16
16 9 16 8 16
+ I8
+ 19 10 + 10

WATER BEETLES FROM THE EBRO DELTA 139
Appendix (continued).
Ds St AD SA PS TO LD AE ED
27 Hydroporus pubescens (Gyllenhal, 1808) 28 Hydroporus tessellatus Drapiez, 1819
30 Hydrovatus cuspidatus (Kunze, 1818) 31 Hygrotus confluens (Fabricius, 1787) 32 Hygrotus impressopunctatus (Schaller, 1783) 33 Hygrotus inaequalis (Fabricius, 1777) 34 Hygrotus parallelogrammus (Ahrens, 1812) 35 Hyphydrus aubei Ganglbauer, 1892 36 Metaporus meridionalis (Aubi, 1836) 37 Methles cribratellus (Fairmaire, 1880) 38 Nebrioporus canaliculatus (Lacordaire, 1835) 39 Nebrioporus cerisyi (Aube, 1836)
40 Agabus bipustulatus (Linnaeus, 1767) 41 Agabus conspersus (Marsham, 1802) 42 Agabus montanus (Stephens, 1828) 43 Agabus nebulosus (Forster, 1771) 44 Colymbetes fuscus (Linnaeus, 1758) 45 Ilybius fuliginosus (Fabricius, 1792) 46 Ilybius meridionalis AubQ 1836
48 Rhantus suturalis (McLeay, 1825)
49 Laccophilus hyalinus (De Geer, 1774) 50 Laccophilus minutus (Linnaeus, 1758) 5 1 Laccophilus ponticus Sharp, 1882
52 Acilius canaliculatus (Nicolai, 1822) 53 Acilius sulcatus (Linnaeus, 1758) 54 Cybister lateralimarginalis (De Geer, 1774) 55 Cybister tripunctatus Castelnau, 1834 56 Cybister vulneratus Klug, 1834 57 Dytiscus pisanus Castelnau, 1834 58 Dytiscus sernisulcatus Miiller, 1776 59 Eretes sticticus (Linnaeus, 1767) 60 Graphoderus cinereus (Linnaeus, 1758) 61 Hydaticus leander (Rossi, 1790) 62 Hydaticus seminiger (De Geer, 1774) 63 Hydaticus transversalis (Pontoppidan, 1763)
64 Ochthebius corrugatus Rosenhauer, 1856 65 Ochthebius delgadoi Jach, 1994 66 Ochthebius dentifer Rey, 1885 67 Ochthebius dilatatus Stephens, 1829 68 Ochthebius macutatus Reiche, 1872 69 Ochthebius marinus (Paykull, 1798) 70 Ochthebius meridionalis Rey, 1885 71 Ochthebius nanus Stephens, 1829 72 Ochthebius pilosus Waltl, 1835 73 Ochthebius subpictus Woilaston, 1857 74 Ochthebius viridis 1 sensu Jach, 1991a 75 Ochthebius viridis 2 sensu Jach, 1991a
T T E T T N T N T T S T T
T T T T T N T N T
T T T
N T T S S T T T N T N N
S E M T S T T T T M N T
29 Hydroporus vagepictus Fairmaire & Laboulbtne, 1854
Colymbetinae
47 Ilybius quariguttatus (Boisduval & Lacordaire, 1835)
Laccophilinae
Dytiscinae
HYDRAENIDAE
8 16 + +/I0 16 + 9 16 + 10 16
+/8 16 8 16
10 16 + + / I 0 16
+ / I0 16
16
+ /8
+ 14
+ +
+
2 + +
+ 8 16 + +/11 16 16
+ + 8 1 6 + /8
8/5 + 8 5 + + +/8 16
+
X?
+ + + 10 16 + + / l o 16
+/8 16 + 9 10 9 16 9
+ + +
C c,x +
8
8 16 8 16
+ + +/9 16 16
8
+ +
R.X
+ +
R + + + t7 + 17
+ 17 12
+
+? + 12 + 17
+ 12? 12? 17
C + +

140 I. RIBERA E T A L .
Appendix (continued).
Ds St A D SA ED PS TO L D AE
HELOPHORIDAE 76 Helophorus alternans Gent , 1836 77 Helophorus aquaticus (Linnaeus, 1758) 78 Helophorus brevipalpis Bedel, 1881 79 Helophorus fulgidicollis Motschulsky, 1860 80 Helophorus griseus Herbst, 1793 81 Helophorus illustris Sharp, 1916 82 Helophorus maritimus cplx Rey, 1885 83 Helophorus minutus Fabricius, 1775 84 Helophorus rufpes (Bosc, 179 1)
HY DROCHIDAE 85 Hydrochus smaragdineus Fairmaire, 1879
HYDROPHILIDAE Hydrophilinae 86 Anacaena lutescens (Stephens, 1829) 87 Berosus affnis Brullk, 1835 88 Berosus fulvus Kuwert, 1888 89 Berosus hispanicus Kiister, 1847 89 Berosus hispanicus Kiister, 1847 90 Berosus jaechi Schodl, 1991 91 Berosus signaticollis (Charpentier, 1825) 92 Chasmogenus livornicus (Kuwert, 1890) 93 Enochrus bicolor (Fabricius, 1792) 94 Enochrus halophilus (Bedel, 1878) 95 Enochrus isotae Hebauer, 1981 96 Enochrus melanocephalus (Olivier, 1792) 97 Enochrus quadripunctatus (Herbst, 1797) 98 Enochrus testaceus gr. sensu Ribera & Aguilera (1995) 99 Helochares lividus (Forster, 1771)
100 Hydrobius fuscipes (Linnaeus, 1758) 101 Hydrochara caraboides (Linnaeus, 1758) 102 Hydrochara fravipes (Steven, 1808) 103 Hydrophilus piceus (Linnaeus, 1758) 104 Hydrophilus pistaceus (Castelnau, 1840) 105 Limnoxenus niger (Zschach, 1788) 106 Paracymus aeneus (Germar, 1824)
107 Coelostoma hispanicum (Kiister, 1848)
108 Dryops algiricus (Lucas, 1849) 109 Dryops hispanus Olmi, 1972
Sphaeridiinae
DRYOPIDAE
Sp. No.
T N T T T M T T T
T
T T N T T M T M T T T T N T? T T N T N T T T
T
T E?
+ +
+
+ +
+ + 12
C + + 8
8 17
17
+ f 17
3 8 17 + + +/3 + + + +/3 + +
+ 17
+ + + 17
R + /3 + R +
+ 12 + + + + 12?
17
+ + + +/8 17
17 +/I2 17
8 8 12 17 12 17
+ / 5 + + + +
+
+ 8 17
+ + + + R,X? 14
19 9 37 31 11 77 54
Ds: General distribution of the species (see text): T, trans-Iberian; E, Iberian endemics sensu lato; N, Northern species; S, Southern species; M, Coastal circum-Mediterranean species. St: Conservation status of the species in the Iberian peninsula: X, Probably extinct; R, Only one known, coastal, population; C, Present only in the coast (some of them can be found inland in other geographical areas). Coastal wetlands surveyed (see Fig. 1): AD, Albufera de Adra (Almeria); SA, Torrevieja and Santa Pola salines (Alicante); ED, Ebro delta (Tarragona); PS, Pineda de Salou (Tarragona); TO, Torredembarra (Tarragona); LD, Llobregat delta (Barcelona); AE, Aiguamolls de I'Empordl (Girona). Records: +, new records; numbers refer to bibliographic records (only the first record of the species in each locality is provided). 1, Montes and Soler (1985); 2, Foster (1986); 3, Aguilera and Ribera (1994); 4, Hernando and Aguilera (1994); 5, Ribera er al. (1995); 6, Falkenstrom (1939); 7, Jach (1990); 8, Cuni (1888); 9, Traizet (1895); 10, Lagar (1951); 11, Lagar (1967); 12, Lagar (1970); 13, Lagar (1968); 14, Olmi (1972); 15, Hernando and Fresneda (1986); 16, Rosales and Lafuente (1993); 17, Natalio Rosales (personal communication, 1996).