a krüppel-associated box–zinc finger protein, nt2, represses cell
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, June 2002, p. 4256–4267 Vol. 22, No. 120270-7306/02/$04.00�0 DOI: 10.1128/MCB.22.12.4256–4267.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
A Krüppel-Associated Box–Zinc Finger Protein, NT2, RepressesCell-Type-Specific Promoter Activity of the �2(XI) Collagen Gene
Kazuhiro Tanaka,1,2 Noriyuki Tsumaki,1 Christine A. Kozak,3 Yoshihiro Matsumoto,2Fumihiko Nakatani,2 Yukihide Iwamoto,2 and Yoshihiko Yamada1*
Craniofacial Developmental Biology and Regeneration Branch, National Institute of Dental and Craniofacial Research,1 andLaboratory of Molecular Microbiology, National Institute of Allergy and Infectious Diseases,3 National Institutes of
Health, Bethesda, Maryland 20892, and Department of Orthopaedic Surgery, Graduate School ofMedical Sciences, Kyushu University, Fukuoka 812-8582, Japan2
Received 4 January 2002/Returned for modification 8 February 2002/Accepted 13 March 2002
Type XI collagen is composed of three chains, �1(XI), �2(XI), and �3(XI), and plays a critical role in theformation of cartilage collagen fibrils and in skeletal morphogenesis. It was previously reported that the�530-bp promoter segment of the �2(XI) collagen gene (Col11a2) was sufficient for cartilage-specific expres-sion and that a 24-bp sequence from this segment was able to switch promoter activity from neural tissues tocartilage in transgenic mice when this sequence was placed in the heterologous neurofilament light gene (NFL)promoter. To identify a protein factor that bound to the 24-bp sequence of the Col11a2 promoter, we screeneda mouse limb bud cDNA expression library in the yeast one-hybrid screening system and obtained the cDNAclone NT2. Sequence analysis revealed that NT2 is a zinc finger protein consisting of a Krüppel-associated box(KRAB) and is a homologue of human FPM315, which was previously isolated by random cloning andsequencing. The KRAB domain has been found in a number of zinc finger proteins and implicated as atranscriptional repression domain, although few target genes for KRAB-containing zinc finger proteins hasbeen identified. Here, we demonstrate that NT2 functions as a negative regulator of Col11a2. In situ hybrid-ization analysis of developing mouse cartilage showed that NT2 mRNA is highly expressed by hypertrophicchondrocytes but is minimally expressed by resting and proliferating chondrocytes, in an inverse correlationwith the expression patterns of Col11a2. Gel shift assays showed that NT2 bound a specific sequence within the24-bp site of the Col11a2 promoter. We found that Col11a2 promoter activity was inhibited by transfection ofthe NT2 expression vector in RSC cells, a chondrosarcoma cell line. The expression vector for mutant NT2lacking the KRAB domain failed to inhibit Col11a2 promoter activity. These results demonstrate that KRAB-zinc finger protein NT2 inhibits transcription of its physiological target gene, suggesting a novel regulatorymechanism of cartilage-specific expression of Col11a2.
Cartilage is a highly specialized tissue and serves as thetemplate for the growth and development of most bones. Car-tilage development is initiated by mesenchymal cell condensa-tion, followed by a series of chondrocyte maturation processesincluding resting, proliferative, and hypertrophic stages. Car-tilage contains an extensive extracellular matrix which includestype II, IX, and XI collagens and provides mechanical strengthto resist compression in joints. Type II collagen, a major col-lagen in cartilage, forms collagen fibrils and provides a struc-tural framework for cartilage matrix. Type XI collagen coas-sembles stoichiometrically with type II collagen in the fibrils,whereas the type IX collagen is associated with the surface ofthe fibrils (30, 46).
The type XI collagen molecule is composed of three sub-units: �1(XI), �2(XI), and �3(XI) (32). The �3(XI) chain is aposttranslational variant of the �1(II) chain (14), whereas the�1(XI) and �2(XI) chains are distinct gene products (5, 22).Type XI collagen plays a critical role in regulating the forma-tion of the collagen fibrils (30, 38). It has been reported that anull mutation in the gene for the �1(XI) chain results in chon-drodysplasia in cho/cho mice (27). Mutations in the �2(XI)
chain cause chondrodysplasias in humans, such as Stickler syn-drome and otospondylomegaepiphyseal dysplasia, indicatingthat type XI collagen is intimately involved in skeletal mor-phogenesis (47). These observations indicate that the fidelity oftype XI collagen expression is essential for maintaining normalcartilage structure and function.
Expression of Col11a2 appears to be predominantly re-stricted to cartilage (43). Transcriptional regulation of Col11a2is mediated by tissue-specific regulatory elements within the�742-bp promoter of Col11a2 (44). It was shown that the�530-bp promoter sequence is sufficient for cartilage-specificexpression of Col11a2 (45). It has been suggested that SOX9,a member of the transcription factor family with an high-mobility-group (HMG)-type DNA binding domain homolo-gous to that of SRY (17, 54), plays an important role in theregulation of Col11a2 expression. Mutations in the gene forSOX9 cause campomelic dysplasia, a severe dwarfism syn-drome, which affects all cartilage-derived structures (12, 49,52). SOX9 binds to HMG-box-like sequences in the Col11a2promoter and increases the promoter activity (6). It has beenshown that a 24-bp sequence from �530 to �507 in theCol11a2 promoter is able to switch the activity of the heterol-ogous neurofilament light gene (NFL) promoter from neuraltissues to cartilage (45). It was also found that the deletion ofa sequence between �530 and �500 abolished reporter gene
* Corresponding author. Mailing address: Building 30, Room 405,NIDCR, NIH, Bethesda, MD 20892. Phone: (301) 496-2111. Fax:(301) 402-0897. E-mail: [email protected].
4256
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
on M
arch 17, 2018 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
on M
arch 17, 2018 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
on M
arch 17, 2018 by guesthttp://m
cb.asm.org/
Dow
nloaded from

expression in cartilage in transgenic mice and induced neuraltissue-specific expression (45). These observations suggest thatCol11a2 expression is regulated by both positive and negativeregulators.
A number of genes encoding the C2H2-type zinc finger do-main have been identified (4, 23). The Krüppel-associated box(KRAB) is a highly conserved motif of 75 amino acids that isfound in approximately one-third of the C2H2-type zinc fingerproteins (3). It has been suggested that the KRAB domain actsas a potent transcriptional repression domain (9, 29, 34, 37, 48,53, 58); however, these studies were done using artificial DNAbinding motifs fused to the KRAB domains and target DNAsequences such as the GAL4 binding domain and GAL4 up-stream activation sequence to demonstrate repressor activity ofthe KRAB domains. Therefore, little is known about physio-logical target genes for KRAB domain-containing proteins andtheir functional interactions.
Previous observation using reporter gene constructs in trans-genic mice suggested that a 24-bp sequence in the Col11a2promoter inhibits Col11a2 expression in neural tissues but isnecessary for cartilage-specific expression of the gene (45). Tounderstand the cartilage-specific regulatory mechanism in-volved in the 24-bp sequence, we screened a mouse limb budcDNA library using the yeast one-hybrid system (26, 50) andidentified KRAB-zinc finger protein factor NT2, which boundto the 24-bp sequence. We found that NT2 expression wasinversely correlated with expression of Col11a2 and that itinhibited Col11a2 promoter activity via binding to the 24-bpsite through the KRAB domain. Our results suggest a novelmechanism by which cartilage-specific expression of Col11a2 isnegatively regulated during embryonic development and chon-drocyte differentiation.
MATERIALS AND METHODS
Yeast strains and gene constructs. Saccharomyces cerevisiae YM4271 (MATaura3-52 his3-200 leu2-3,112 trp1-903) and reporter vectors pHISi and pLacZiwere obtained from Clontech (Palo Alto, Calif.). The reporter construct wasgenerated by inserting three head-to-tail copies of the 24-bp sequence (5�-CAGGGAGGAGGGAGAGCGGCTGCT-3�), which corresponds to the mouseCol11a2 promoter sequence (�530 to �507) (44), into the EcoRI and XbaI sitesof pHISi or the EcoRI and XhoI sites of pLacZi. These plasmids were linearizedand integrated into yeast YM4271 genomes. The yeast host strain was main-tained by selection on synthetic dextrose medium lacking histidine and uracil.
For a GAL4 activation domain-tagged cDNA library, poly(A)� RNA wasextracted from the limb buds of 13.5-day-old mouse embryos using the Micro-FastTrack kit (Invitrogen, Carlsbad, Calif.). An oligo(dT)-primed cDNA librarywas constructed with the HybriZap phage vector (Stratagene, La Jolla, Calif.).The plasmid (pAD-GAL4) library was obtained by in vivo excision according tothe manufacturer’s instructions (Stratagene). The library had a complexity of 2.2� 106 PFU and an average insert size of about 1.7 kb.
Screening of the GAL4 activation domain cDNA library. Screening of thecDNA library was performed with a yeast strain carrying the HIS3 and lacZreporter genes containing three copies of the 24-bp sequence of the Col2a1promoter (described above) by a lithium acetate method (40). The transformedyeast cells were plated under selective conditions with synthetic dextrose mediumlacking histidine and leucine. The cells grown on the selective plates weretransferred onto nitrocellulose filters. The membranes were frozen in liquidnitrogen and assayed for �-galactosidase activity. An estimated 2.7 � 106 trans-formants were selected, and 44 positive clones were obtained from the firstscreening. Twenty positive clones were recovered in the secondary screening.
Cell lines. Chondrogenic cells (ATDC5) (41) and rat chondrosarcoma cells(RCS) (33) were from Yuji Hiraki and James H. Kimura. Mouse cells (BALB/3T3 and 10T1/2), rat myoblast cells (L6), mouse myoblast cells (C2C12), mouseosteoblastic cells (MC3T3), and rat osteosarcoma cells (ROS17/2.7) were ob-tained from the American Type Culture Collection (Manassas, Va.).
Northern hybridization. Total RNA was extracted from various cell lines andnewborn mouse tissues using the RNeasy Mini kit (Qiagen). Northern blotanalysis was performed by electrophoresis of 20 �g of total RNA or 2 �g ofpoly(A)� RNA, and RNA was transferred onto Nytran membranes (Schleicher& Schuell) as described (39). cDNAs were labeled with [�-32P]dCTP using thePrime-it II kit (Stratagene). The membranes were hybridized with the labeledprobes at 42°C in 50% formamide, washed first at room temperature in 1� SSC(1� SSC is 0.15 M NaCl plus 0.015 M sodium citrate) and 0.1% sodium dodecylsulfate (SDS) and then at 60°C in 0.1� SSC and 0.1% SDS and exposed toautoradiography film.
Western blotting. Cell lysates and nuclear extracts were prepared as describedpreviously (25). Two micrograms of protein samples was fractionated by SDS-polyacrylamide gel electrophoresis (PAGE) and transferred onto a nylon mem-brane. The blots were incubated with anti-NT2 or anti-Flag antibody, and signalswere detected with an ECL kit (Amersham). The anti-Flag antibodies were obtainedfrom Sigma (St. Louis, Mo.). The anti-NT2 polyclonal antibodies were raised againstthe C-terminal portion of NT2 (amino acid residues 323 to 345) (Fig. 1) by immu-nizing rabbits. The antibodies were purified using an ImmunoPure IgG Purificationkit (Pierce, Rockford, Ill.). The immunoglobulin G fractions were also passedthrough an NT2-Sepharose affinity column to deplete reactivity to NT2 and wereused to demonstrate specific reactivity of the anti-NT2 antibodies.
In situ hybridization. Digoxigenin-11-UTP-labeled single-strand antisenseRNA probes for Col11a2, Col10a1, and NT2 were prepared using the DIG RNAlabeling kit (Boehringer Mannheim, Indianapolis, Ind.) according to the manu-facturer’s instructions. The sense riboprobes for Col11a2 and NT2 were alsoprepared and used as negative controls. In situ hybridization was performed asdescribed previously (18). After deparaffinization, the sections were treated with10 �g of proteinase K/ml for 15 min at room temperature and subjected to 0.2N HCl to inactivate endogenous alkaline phosphatase. Hybridization was per-formed at 50°C with 50% formamide, and washes were carried out at a stringencyof 2� SSC containing 50% formamide at 55°C. Then, the slides were subjectedto 10 mg of RNase A/ml in TNE (10 mM Tris-HCl [pH 8.0], 500 mM NaCl, 1mM EDTA) at 37°C for 30 min for the digestion of nonhybridized transcripts andwashed. A Genius Detection system (Boehringer Mannheim) was used to detectsignals according to the manufacturer’s instructions.
Electrophoretic mobility shift assays (EMSAs). The expression vectorspCA1F (42) and pcDNA3.1 (Invitrogen) were used to express Flag-tagged ornontagged NT2 proteins. A PCR product for full-length NT2 was cloned into theEcoRI and XhoI sites of pCA1F or pcDNA3.1. In vitro-translated NT2 proteinwas prepared by the TNT Coupled Reticulocyte Lysate system (Promega, Mad-ison, Wis/). WT, a 24-bp, double-stranded, wild-type probe (5�-GGCAGGGAGGAGGGAGAGCGGCTGCT) corresponding to the Col11a2 promoter se-quence, was used for the yeast screening described above. G residues were addedfor labeling with [�-32P]dCTP by Klenow fragment (Life Technologies, Gaith-ersburg, Md.). Substitution mutation probes, used as competitors, were preparedand their plus strand sequences are as follows (mutated nucleotides are under-lined): M1, 5�-CATTTGTTAGGGAGAGCGGCTGCT-3�; M2, 5�-CAGGGAGGAGTTGTGTCGGCTGCT3�; M3, 5�-CAGGGAGGAGGGAGAGCTTACTAT3�; M4, 5�-AGGGGAGGAGGGAGAGCGGCTGCT-3�; M5, 5�-CATTGAGGAGGGAGAGCGGCTGCT-3�; M6, 5�-CAGGTGGGAGGGAGAGCGGCTGCT-3�; M7, 5�-CAGGGATTAGGGAGAGCGGCTGCT-3�; M8, 5�-CAGGGAGGGTGGAGAGCGGCTGCT-3�; M9, 5�-CAGGGAGGAGTTAGAGCGGCTGCT-3�; M10, 5�-CAGGGAGGAGGGGTAGCGGCTGCT-3�; M11, 5�-CAGGGAGGAGGGAGGTCGGCTGCT-3�; and M12, 5�-CAGGGAGGAGGGAGAGATGCTGCT-3�.
EMSA was performed using the GelShift assay kit (Stratagene) according tothe manufacturer’s instructions.
DNA transfection assays. DNA was transfected into various cells using Fugene6 (Boehringer Mannheim) according to the manufacturer’s instructions. Theexpression vector pCA1F-NT2 was transiently transfected into RCS cells toexpress Flag-tagged NT2 protein. To express deletion mutants of pNT2, pNT2�1(amino acids 130 to 680), pNT2�2 (amino acids 210 to 680), and pNT2�3 (aminoacids 300 to 680) were generated by subcloning EcoRI/XhoI PCR fragments fromNT2 into the corresponding sites of the pCA1F vector. pNT2�4 was generatedby subcloning EcoRI/SalI (amino acids 1 to 216) and SalI/XhoI (amino acids 258to 680) PCR fragments of NT2 into the corresponding sites of the pCA1F vector.The anti-Flag antibodies were used to detect the expression of Flag-tagged NT2proteins. The reporter constructs p742luc, p530luc, p518luc, and p500luc con-tained the 742, 530, 518, and 500 bp of the Col11a2 promoter, respectively, andthese were linked to the luciferase reporter gene (44, 45). The reporter constructp530Mluc is the same as p530luc except for a substitution mutation at the NT2binding site. The reporter constructs containing the promoter of human NFL(hNF-L), pNFluc, and three tandem copies of the 24-bp sequence (�530 to
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4257
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

�507) of Col11a2 fused to the hNF-L, p24 � 3-NFluc, were linked to theluciferase reporter gene. pGL3-Control and pRL-SV40 (Promega) were used asa positive control and an internal control for normalization of transfectionefficiency, respectively. The transfected cells were harvested 48 h after transfec-tion and assayed for luciferase activity using the Dual-Luciferase Reporter assaysystem (Promega).
Genetic mapping. NT2 was mapped by Southern blot analysis of two geneticcrosses: (NFS/N or C58/J � M. musculus musculus) � M. m. musculus (23a) and(NFS/N � Mus spretus) � M. spretus or C58/J (1a). Digestion of parental mouseDNAs with EcoRI produced NT2 fragments of 9.4 kb in NFS/N and 8.2 kb in M.spretus and M. m. musculus.
RESULTS
Isolation of cDNA clones for proteins interacting with theCol11a2 promoter. We previously identified a 24-bp sequence(�530 to �507) as a regulatory site for Col11a2 transcription.To identify a nuclear factor that bound to the sequence, wescreened a 13.5-day-old mouse embryo limb bud cDNA libraryby the yeast one-hybrid screening method using a yeast strainharboring three tandem copies of the 24-bp sequence as atarget. After secondary screening, 20 positive clones were iso-lated and sequenced. Seventeen of them were found to encode
the NT2 protein and were further characterized, whereas theremaining 3 clones were nonspecific clones. DNA sequenceanalysis revealed that NT2 encodes a 690-amino acid polypep-tide with the KRAB-A subdomain, a leucine-rich region (LeR)at the N terminus, and nine C2H2-type zinc finger motifs at itscarboxyl terminus. The LeR domain is rich in leucine andglutamic acid residues and contains alpha helices. The LeR isoften found in KRAB-containing zinc finger proteins (2, 7, 10,24, 35, 36, 51, 55). The NT2 clone appears to be a homologueof a human cDNA clone, FPM315, previously identified byrandom cloning and sequencing (56). Chromosomal mappingsupported this notion. FPM315 is mapped on chromosome16p13.3 (56) (GenBank number AC004232). We have mappedthe NT2 gene at the centromeric end of mouse chromosome 16with the following gene order and recombinational distances:Ftl-rs8 � 3.4 1.1 � Znf263 � 5.1 1.4 � Igl, Hcf2. The genemaps to regions of conserved synteny in the human and mousegenomes. Sequence comparison between NT2 and humanFPM315 revealed that overall nucleotide and amino acid iden-tities were 85.0 and 84.1%, respectively (Fig. 1). The LeR and
FIG. 1. Alignment of mouse NT2 and human FPM315 amino acid sequences. The sequence of the mouse clone NT2 (upper sequence, M;GenBank no. AF499776) is compared with that of human FPM315 (lower sequence, H; GenBank no. AC004232) (56). Numbers refer to aminoacid positions in the mouse protein. Vertical lines denote amino acid identities. Boxes outline nine conserved zinc finger motifs. The boldfaceunderlining indicates the KRAB-A subdomain, and the thin underlining indicates the LeR domain.
4258 TANAKA ET AL. MOL. CELL. BIOL.
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

KRAB domains are highly conserved between human andmouse species: 97.4% amino acid identity in zinc finger motifs,97.0% in the LeR domain, and 90.2% in the KRAB-A subdo-main, respectively.
Inverse correlation of the expression pattern of NT2 andCol11a2 in chondrocyte differentiation and developing carti-lage. The expression of mouse NT2 mRNA was analyzed intissues from newborns and various cell types by Northern blot-ting and in mouse embryos by in situ hybridization. Northernanalysis of total RNA from newborn mice revealed that NT2mRNA was strongly expressed in the brain, thymus, and spleenand in rib cartilage, whereas little expression was found inskeletal muscle or in the heart, liver, or kidney (Fig. 2A). NT2mRNA was highly expressed in chondrogenic cell linesATDC5, BALB/3T3, and 10T1/2 and an osteoblastic cell line,MC3T3 (Fig. 2B), which did not synthesize Col11a2 mRNA. Incontrast, the level of NT2 mRNA was weak in chondrocyticRCS cells, a rat chondrosarcoma cell line which produces alarge amount of Col11a2 mRNA. A low level of NT2 mRNAwas observed in muscle cell lines L6 and C2C12 and osteosar-coma cell line ROS17/2.7; Col11a2 expression was also weak.We also examined the protein expression of NT2 in RCS, NIH3T3, and undifferentiated ATDC5 cells by Western blottingwith anti-NT2 antibodies (Fig. 2C). In RCS cells, althoughsome expression of NT2 mRNA was observed, the level of NT2protein was very low (Fig. 2C, lane 4). On the other hand, highlevels of NT2 protein, approximately 75 kDa in size, werefound in NIH 3T3 and undifferentiated ATDC5 cells (Fig. 2C,lanes 2 and 3), corresponding to strong expression of NT2mRNA. In vitro-translated NT2 protein also showed a singleband of 75 kDa (Fig. 2C, lane 1). This protein band was notdetected when the antibodies had depleted reactivity againstpurified NT2 synthesized in vitro (Fig. 2C, lanes 5 and 6),indicating specificity of the NT2 antibodies.
We next examined nuclear factor binding to the 24 bp of theCol11a2 promoter by EMSA (Fig. 2D). The nuclear extractsfrom NIH 3T3 and undifferentiated ATDC5 cells formed aprotein complex with the 24-bp probe (Fig. 2D). The complexwas supershifted by the addition of anti-NT2 antibodies, sug-gesting that it contains NT2 protein (Fig. 2, lanes 3 to 6). RCScell nuclear extracts formed two protein-DNA complexeswhose gel mobilities were different from those obtained withextracts from NIH 3T3 and undifferentiated ATDC5 cells(lanes 1 to 2) and were not supershifted by anti-NT2 antibod-ies, indicating that the complexes did not contain NT2.
In situ hybridization in the forearm of 16.5-day-old mouseembryos revealed that NT2 mRNA was expressed in the hy-pertrophic zone, whereas its expression was very low in theresting and proliferative zones (Fig. 3A and D). As expected,strong signals for Col11a2 mRNA were detected in the prolif-erating and prehypertrophic zones (Fig. 3B), while Col10a1mRNA was expressed predominantly in the prehypertrophicand hypertrophic zones (Fig. 3C). The expression of NT2mRNA was strong in the prehypertrophic and hypertrophiczones but weak in the proliferating zone (Fig. 3D), similar tothat of Col10a1, a marker of hypertrophic chondrocytes. Sincethe 24-bp sequence has the ability to switch promoter activityof the neurofilament gene from neural tissues to cartilage, wealso examined the expression pattern of NT2 in brain. In thefrontal cortex of the brain of 16.5-day-old mouse embryos, the
FIG. 2. mRNA and protein expressions of NT2 in various tissuesand cell types. (A) Analysis of NT2 expression in newborn mousetissues by Northern blotting. For each lane, 20 �g of total RNA fromvarious tissues was loaded, transferred to the nylon membrane, andhybridized with labeled NT2 cDNA. NT2 mRNA was strongly ex-pressed in the brain, thymus, spleen, and rib cartilage. Lane 1, brain;lane 2, thymus; lane 3, heart; lane 4, liver; lane 5, spleen; lane 6, kidney;lane 7, skeletal muscle; lane 8, rib cartilage. (B) Total RNA (20 �g/lane) extracted from various cells was analyzed by Northern blottingusing the NT2 cDNA probe. NT2 mRNA was highly expressed inBALB/3T3, 10T1/2, undifferentiated ATDC5, and MC3T3 cells(ATDC5 is a chondrocytic cell line and MC3T3 is an osteoblastic cellline), whereas low expression was seen with RCS cells (RCS is a ratchondrosarcoma cell line). Lane 1, BALB/3T3; lane 2, 10T1/2; lane 3,undifferentiated ATDC5; lane 4, RCS; lane 5, MC3T3; lane 6, ROS17/2.7; lane 7, L6; lane 8, C2C12. The lower panels show ethidium bro-mide-stained gels. (C) Western blot analysis of NT2 protein expressionin cell lines. Nuclear extracts (2 �g) from cells were fractionated bySDS-PAGE, blotted, and incubated with NT2 antibodies. In vitro-translated NT2 (2 �g) was also subjected to Western blot analysis. Toshow the specificity of NT2 antibodies, the antibodies with NT2 reac-tivity depleted by NT2-Sepharose affinity column chromatographywere also used as a control (�NT2 antibody). Lane 1, in vitro-trans-lated NT2; lane 2, NIH 3T3; lane 3, undifferentiated ATDC5; lane 4,RCS; lane 5, NIH 3T3 with �NT2 antibody; lane 6, undifferentiatedATDC5 with �NT2 antibody. Protein standards are indicated on theleft. The arrowhead indicates the NTZ protein band. (D) DNA bind-ing of NT2 in cells to the labeled wild-type Col11a2 promoter. The24-bp DNA sequence CAGGGAGGAGGGAGAGCGGCTGCT wasused to prepare a double-stranded probe. EMSAs were performedwith nuclear extracts from RCS (lanes 1 and 2), NIH 3T3 (lanes 3 and4), and undifferentiated ATDC5 (lanes 5 and 6) cells. The presence(�) or absence (�) of antibodies against NT2 is indicated. The largearrow indicates NT2-promoter complexes (lanes 3 and 5). The smallarrow marks supershifted NT2-promoter complexes by the anti-NT2antibodies (lanes 4 and 6). The arrowheads indicate RCS cell-specificprotein-promoter complexes (lanes 1 and 2). These complexes arespecific to RCS cells and are not found in either NIH 3T3 or undif-ferentiated ATDC5 cells and are not supershifted by anti-NT2 anti-bodies.
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4259
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

expression of NT2 mRNA was observed in the marginal zone,cortical plate, and ventricular zone where NFL mRNA wasfound (Fig. 3F and G). Sense NT2 probes showed no stainingin the limb and brain sections (Fig. 3E and H). NT2 mRNAexpression during chondrogenesis was next examined by in situhybridization using sections of mouse embryos at variousstages. The signals for NT2 mRNA were ubiquitous except inthe neural tube and somites in 8.5- and 9.5-day-old mouseembryos (Fig. 4A and B). The mesenchymal condensation wasobserved in 12.5-day-old embryos with Col11a2 mRNA expres-sion in the ribs and vertebral cartilage primordia (Fig. 4D),whereas NT2 mRNA expression was almost absent in theselocations (Fig. 4F). Sense NT2 and Col11a2 probes showed nostaining in these tissues (Fig. 4C, E, and G). These results alsoindicate an inverse correlation between the expression patternsof NT2 and those of Col11a2.
Next, expression patterns of NT2 mRNA were examined
during differentiation of ATDC5 cells (Fig. 5). In the presenceof insulin, ATDC5 cells differentiate into a proliferating chon-drocyte phenotype and subsequently into a hypertrophic chon-drocyte phenotype concomitant with the formation of cellularnodules (41). After 1 week in cultures in a confluent conditionwith insulin, the cells began to form nodules and to expressCol11a2, at which time NT2 expression was downregulated.We separated the nodule- and non-nodule-forming ATDC5cell populations, because these cells are mixed populations,with nodule-forming cells representing the chondrocytic phe-notype. In 2-week cultures, NT2 expression was not detectablein the nodule-forming cells where Col11a2 mRNA was ex-pressed at high levels (Fig. 5, lane 2). In the non-nodule-forming cells, NT2 mRNA was expressed at high levels,whereas Col11a2 mRNA was not present (Fig. 5, lane 1).Further culturing for 3 to 5 weeks switched the expression ofCol11a2 to Col10a1, indicative of terminal differentiation of
FIG. 3. In situ hybridization of longitudinal sections of the radius or forebrain of 16.5-day-old mouse embryos with antisense Col11a2, Col10a1,and NT2 or with sense NT2 riboprobes labeled with digoxigenin-11-UTP. (A) Staining with hematoxylin and eosin of the radius in the forelimb.h, hypertrophic chondrocytes; r, resting chondrocytes; p, proliferating chondrocytes. (B) Expression of Col11a2 in a semiserial section. Strongsignals of Col11a2 were detected in the resting and proliferating chondrocytes. (C) Expression of Col10a1 was observed in hypertrophicchondrocytes. (D) NT2 mRNA was highly expressed in the hypertrophic zone; however, its expression was very weak in the resting and proliferativezones. The localization of NT2 mRNA was similar to that of Col10a1, a marker gene of hypertrophic chondrocytes. (E) Sense NT2 riboprobesshowed no signals in the serial section. (F) Expression of NT2 mRNA in the forebrain of 16.5-day-old mouse embryos. Expression of NT2 wasobserved in the frontal cortex (fc) of the forebrain. (G) Higher magnification of the frontal cortex demonstrates that the signal for NT2 mRNAwas detected in the marginal zone (mz), cortical plate (cp), and ventricular zone (vz) but was weak in the intermediate zone (im). (H) Sense NT2riboprobes showed no signals in the brain section.
4260 TANAKA ET AL. MOL. CELL. BIOL.
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

ATDC5 cells. As the expression of the collagen types wasswitched from Col11a2 to Col10a1 in differentiating ATDC5cells, NT2 expression was induced again. In 5-week cultures,the nodule-forming cells expressed Col10a1 and NT2, whereasCol11a2 mRNA was not detectable in these cells (Fig. 5, lane3). These results indicate an inverse correlation in the expres-sion pattern between NT2 and Col11a2.
NT2 protein specifically binds to the Col11a2 promoter se-quence. The Flag-tagged, full-length NT2 protein was synthe-sized by an in vitro transcription-translation system and exam-ined for its binding activity to the Col11a2 promoter (Fig. 6).NT2 was able to bind to the 24-bp sequence from the Col11a2promoter (WT) (Fig. 6B, lane 2). We performed competitionexperiments with unlabeled oligonucleotides with substitutionmutations to determine a target sequence within the 24-bpsequence for NT2 binding. An excess of the unlabeled WTabolished binding of NT2 to the 24-bp site (Fig. 6B, lane 3).
Mutated oligonucleotide M3 inhibited the binding similar toWT, whereas M1 and M2 failed to block NT2 binding to the24-bp site (Fig. 6B, lanes 4 to 6). These results suggest that thesequences used to create the M1 and M2 substitution muta-tions are critical for the NT2 binding site. Binding of NT2 tothe labeled WT was also inhibited by excess unlabeled M4, M5,M11, and M12 (Fig. 6B, lanes 9, 10, 16, and 17) but not by M6to M10 (Fig. 6B, lanes 11 to 15). These results indicate that theNT2 protein binds specifically to the Col11a2 promoter andthat the core binding sequence is GAGGAGGGAG.
NT2 represses chondrocyte-specific Co1l1a2 promoter activ-ity via KRAB-A subdomain. An expression vector of NT2(pCA1F-NT2) was cotransfected with Col11a2 promoter-lucif-erase reporter gene constructs into RCS cells to examinewhether NT2 exerts its inhibitory activity on the Col11a2 pro-moter. The reporter constructs p742luc and p530luc, whichcontain different sizes of the functional promoter, were both
FIG. 4. In situ hybridization of axial sections of 8.5- (A), 9.5- (B), and 12.5- (C and D)day-old mouse embryos with antisense or sense NT2 andCol11a2 riboprobes labeled with digoxigenin-11-UTP. The signals for NT2 mRNA were ubiquitous but were almost absent in the neural tube andsomites in the 8.5- and 9.5-day-old mouse embryos (A and B). The sense NT2 probes showed no signals in the serial section of 9.5-day-old embryos(C). In the 12.5-day-old embryo, Col11a2 mRNA was strongly expressed at the mesenchymal condensations for rib and vertebral cartilageprimordia (D), whereas NT2 mRNA was not expressed in these locations (F). Negative controls using sense Col11a2 (E) and NT2 (G) showed nosignals in the semiserial sections. lb, limb bud; nt, neural tube; r, rib cartilage primordia; s, somite; v, vertebral cartilage primordia.
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4261
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

active in RCS cells but inactive in BALB/3T3, 10T1/2, or NIH3T3 cells, indicating cell type specificity of promoter activity.p500luc was inactive in all cell types (data not shown). WhenpCA1F-NT2 was cotransfected with p530luc into RCS cells,Col11a2 promoter activity was reduced in a dose-dependentmanner to approximately 20% of that of the control (Fig. 7).The repression by NT2 was not due to a general suppressiveeffect on transcriptional regulation, since NT2 showed no sig-nificant effects on the luciferase activity of the pGL3-Controlplasmid driven by the simian virus 40 (SV40) promoter (Fig.7). NT2 also failed to repress the luciferase activity ofp530Mluc, which contained substitution mutations in the NT2binding site (Fig. 7). These results suggest that NT2 specificallyinhibits Col11a2 promoter activity.
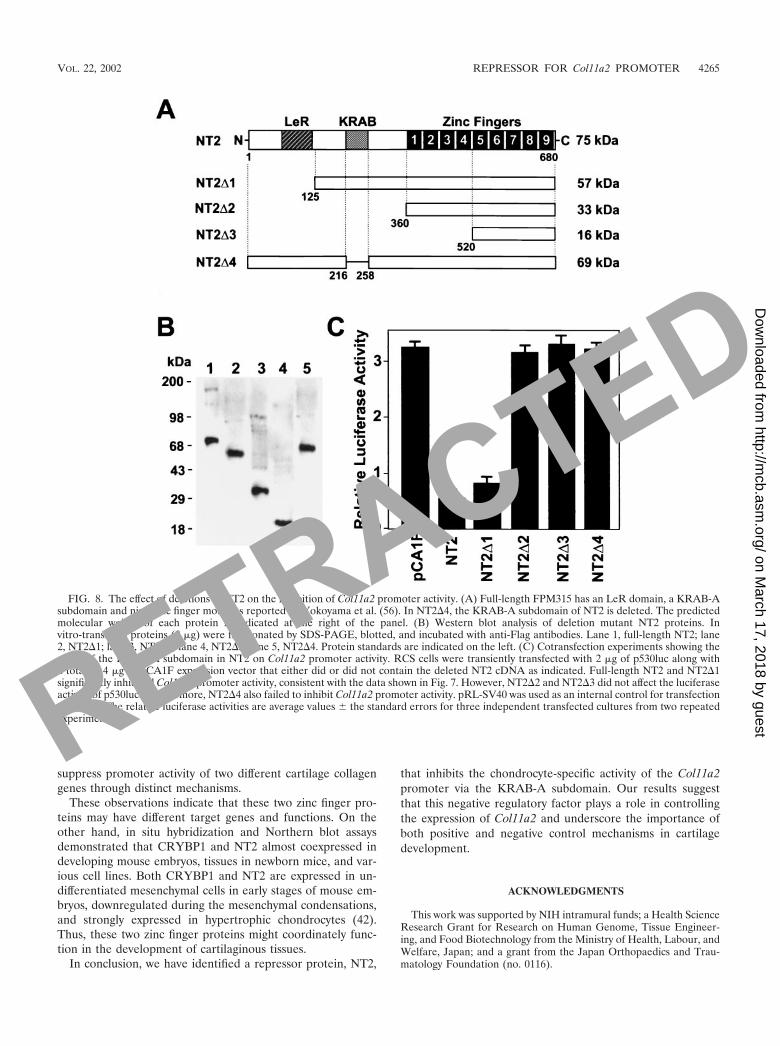
To examine which region of the NT2 protein is important forthe suppression of the promoter activity, we prepared NT2expression constructs with deletion mutations (Fig. 8A). Theprotein expression from each plasmid was confirmed by West-ern blotting with anti-Flag antibody (Fig. 8B). The mutantproteins containing zinc finger motifs were active for DNAbinding (data not shown). When pCA1F-NT2 or NT2�1 wascotransfected with p530luc in RCS cells, inhibition of promoteractivity was observed. However, NT2�2 and NT2�3 did not
exhibit any inhibitory effect on Col11a2 promoter activity.Moreover, NT2�4, in which only the KRAB-A subdomain wasmissing, also lost the inhibitory effect on Col11a2 promoteractivity. These data indicate that the suppressive effect of NT2on Col11a2 promoter activity is mediated, at least in part,through the KRAB-A subdomain.
DISCUSSION
Transcriptional factors play central roles in gene regulationand cellular differentiation. Cumulative evidence has demon-strated that transcriptional repression plays crucial roles inregulating cell-type-specific gene expression and that a numberof genes are controlled by a balance between activation andrepression factors (15, 20). While much attention has beenfocused on understanding transcriptional activation, the pre-cise mechanisms of transcriptional repression are not well un-derstood. Compared to the number of activation elements,only a few DNA motifs that mediate transcriptional repressionhave been identified (3, 8, 11, 16). The KRAB domain wasoriginally identified as a conserved motif consisting of 75amino acids that is present in about one-third of the Krüppel-type C2H2 zinc finger proteins. The KRAB domain is subdi-vided into two subdomains, KRAB-A and -B. The KRAB-Asubdomain is always present in the KRAB-containing proteins,whereas the KRAB-B subdomain is not always there (3). It hasbeen shown that several KRAB-A domains act as potent tran-scriptional repressors when heterologously tethered to pro-moters (29, 37, 53) and that KRAB-B potentiates the repres-sion exerted by the KRAB-A domain (48). Recently, theKRAB domain has been shown to associate with a KRAB-binding protein, KAP-1/TIF1�/KRIP-1, that acts as a universalcorepressor for the KRAB domain and plays an important rolein KRAB-mediated transcriptional repression (1, 13, 21, 31,34). Although it has been well demonstrated that the KRABdomain is a potent DNA binding-dependent repressor domain,these studies were performed by using heterologous DNAbinding motifs fused to the KRAB domain (9, 58). Few phys-iological target genes of the KRAB-containing zinc finger pro-teins have been identified. It has been shown that a KRAB-zincfinger protein, AJ18, bound to a CCACA motif that is also thebinding site for Runx2/Cbfa-1/Osf2, a master gene for osteo-genic differentiation, and that AJ18 suppressed Runx2-medi-ated transactivation of osteocalcin promoter construct (19).However, these studies used the promoter with six copies ofthe consensus binding sequence utilized by Runx2, and thedirect interaction of AJ18 with the osteocalcin promoter hasnot been demonstrated (19). Zheng et al. recently reportedthat a novel KRAB protein, ZBRK1, repressed transcriptionalactivity of the GADD45 gene by binding to a specific sequencewithin intron 3 of GADD45 (57). In the present study, weidentified a KRAB-containing zinc finger protein, NT2, thatbound to the 24-bp sequence in the Col11a2 promoter that iscritical for tissue-specific expression of Col11a2. NT2 showedan inverse correlation of expression patterns with Col11a2 andinhibited Col11a2 promoter activity via the KRAB domain.Our finding is the first demonstration that the KRAB-zincfinger protein is involved in the control of cartilage-specificgene expression.
We previously reported that Col11a2 is regulated by modu-
FIG. 5. Expression of NT2 in differentiating ATDC5 cells in vitro.ATDC5 cells were cultured with 10-�g/ml bovine insulin for differen-tiation. After 2 weeks in culture under confluent conditions, the cellsdifferentiated into the proliferative chondrocyte phenotype andformed nodules. The nodule- and nonnodule-forming cell populationswere separated using sharp forceps under a stereomicroscope, andtotal RNA (20 �g/lane) from the two cell populations was electropho-resed in each lane, transferred to a nylon membrane, and hybridizedwith NT2, Col11a2, and Col10a1 cDNA probes as indicated at the leftof the panels (lanes 1 and 2). The nodule-forming cells then differen-tiated into the hypertrophic chondrocyte phenotype after the cellswere incubated for 5 weeks. The total RNA was extracted and sub-jected to Northern blot analysis (lane 3). NT2 mRNA was stronglydetected in undifferentiated cells (lane 1), but the expression level wasdecreased when the cells differentiated into the proliferative chondro-cyte and began to express Col11a2 (lane 2). As the expression of thecollagen types was switched from Col11a2 to Col10a1 in differentiatingATDC5 cells, NT2 expression was induced again (lane 3). The lowerpanels show the ethidium bromide staining of the gel.
4262 TANAKA ET AL. MOL. CELL. BIOL.
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

lar arrangement of several cis-acting elements, the cartilage-specific element (�530 to �501), a neural tissue-specific ele-ment (�501 to �454), and a basal promoter element withoutany tissue specificity (distal to �453) (45). The deletion of a
sequence between �530 and �500 abolished reporter geneexpression in cartilage and induced neural tissue-specific ex-pression in transgenic mice. We have also shown that the 24-bpsequence (�530 to �507) is able to switch promoter activity of
FIG. 6. Specific DNA binding of NT2 to Col11a2 promoter analyzed by EMSA. (A) The gene structure of the promoter (�742 to �1) and exon1 of Col11a2. The 3�-portion of the retinoid X receptor � gene (Rxrb) was indicated. The coding strand sequences of the WT oligonucleotide probescorresponding to the target sequence (�530 to �507) for the yeast one-hybrid screening and the competitors with substitution mutations (M1through M12) used in EMSA were also indicated. Only mutated nucleotides are shown. (B) In the left panel, lane 2 shows that NT2 bound to theWT Col11a2 promoter sequence. Lanes 3 to 6 show competition between the labeled WT probe and the 50-fold molar excess of cold probes. Lane1, no competitor. DNA binding of NT2 was inhibited by the addition of WT or M3, whereas M1 and M2 probes showed minimal inhibition. Theright panel shows that the DNA binding of NT2 to labeled WT oligonucleotides (lane 7) was inhibited by addition of cold WT (lane 8), M4 (lane9), M5 (lane 10), M11 (lane 16), and M12 (lane 17) probes. The M6, M7, M8, M9, and M10 probes did not affect the binding of NT2 to thepromoter (lanes 11 to 15), indicating that the core binding sequence of NT2 in the promoter is GAGGAGGGAG. Lane 7, no competitor.
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4263
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

the NFL gene from neural tissues to cartilage (45). In thepresent study, the reporter construct pNFluc containing thefunctional neurofilament promoter showed no activity in RCScells; however, p24 � 3-Nfluc, containing three copies of the24-bp Col11a2 sequence in the neurofilament promoter, hadactivity in RCS cells (data not shown), consistent with theresults with transgenic mice (45). These results suggest that the24-bp cartilage-specific cis element has dual activities, one forinactivation of transcription in certain Col11a2-nonexpressingtissues, such as neural tissues, and the other for activation oftranscription in cartilage. In this study, we showed that NT2functions as a suppressor to inactivate cartilage-specific activityof the Col11a2 promoter through binding to the 24-bp site.Since the expression pattern of the reporter gene bearing the�500-bp promoter of Col11a2 in neural tissues in transgenicmice was very similar to that of NT2 (45) (Fig. 3F and G), NT2may inhibit promoter activity in these neural tissues.
The most prominent cell types where NT2 is expressed aremesenchymal cells and hypertrophic chondrocytes. NT2 isstrongly expressed in undifferentiated ATDC5 cells, whereas itis downregulated during differentiation of ATDC5 cells intothe chondrocytic phenotype where Col11a2 is expressed. Littleexpression of NT2 was also observed in RCS cells, which ex-press Col11a2 (Fig. 2). When ATDC5 cells further differenti-ate into hypertrophic chondrocytes, in which Col11a2 expres-sion is downregulated, the expression level of NT2 is increasedagain (Fig. 5). These observations are consistent with the insitu hybridization data, in which expression of NT2 is weak inthe resting and proliferating chondrocytes in developing carti-lage but is expressed strongly in the hypertrophic zones whereCol11a2 is downregulated (Fig. 3). In 8.5- and 9.5-day-oldmouse embryos, NT2 mRNA showed very low levels of expres-
sion in the somites and sclerotomes (Fig. 4A and B), suggestingthat downregulation of NT2 occurs during early mesodermdifferentiation. The level of NT2 expression is very reduced inmesenchymal condensations in 12.5-day-old embryos, indicat-ing an inverse relationship of the expression patterns in vivobetween NT2 and Col11a2 (Fig. 4D and F). These findingssupport the notion that NT2 is involved in both the early andlate stages of chondrocyte differentiation.
Although SOX9 has been shown to interact with both thepromoter and enhancer elements of Col11a2 (6, 28), the 24-bpCol11a2 sequence used in the present study does not containHMG consensus sites for SOX9 binding. EMSA with nuclearextracts from RCS cells demonstrated cell-type-specific proteinbinding to the 24-bp sequence (Fig. 2D). The supershift assayusing anti-NT2 antibodies revealed that the DNA-proteincomplexes did not contain NT2, consistent with the data fromNorthern blotting that NT2 expression was weak in RCS cells.Interestingly, anti-SOX9 antibodies also failed to supershiftthe cell-type-specific complex (data not shown). Therefore, inRCS cells, positive chondrocyte-specific factors other thanSOX9 are likely to interact with the 24-bp site.
NT2 is likely a homologue of the human protein FPM315,based on DNA and protein homology and chromosomal maplocation. The overall nucleotide and amino acid identities be-tween NT2 and FPM315 were 85.0 and 84.1%, respectively.The LeR, KRAB-A, and nine zinc-finger motifs are highlyconserved between humans and mice: 97.0% amino acid iden-tity in the LeR domain, 90.2% in the KRAB-A domain, and97.4% in zinc finger motifs (Fig. 1). The LeR domain is rich inleucine and highly conserved in a number of other zinc fingerproteins (2, 7, 10, 24, 35, 36, 51, 55). The LeR domain issometimes found in zinc finger proteins containing KRAB;however, our deletion experiments suggest that this domain isnot involved in the KRAB-mediated suppression of Col11a2promoter by NT2 (Fig. 8). The LeR domain is also enriched inglutamic acid residues and contains alpha helices. Althoughthe function of the LeR domain is not known, the conservationof the domain and its alpha-helical structures suggest that itmediates interactions with other proteins containing similarmotifs. The highly conserved, nine zinc finger motifs are foundin the C-terminal portion of NT2. The DNA binding of NT2 ismediated by five zinc finger motifs at the C terminus, since thisportion still retains binding activity to the 24-bp sequence (datanot shown). The four remaining zinc finger motifs may not playa central role in DNA binding; however, they may stabilize theprotein-DNA interaction.
Recently, a zinc finger transcription factor, CRYBP1, wasidentified as a repressor for Col2a1 enhancer activity throughbinding to the enhancer with a yeast one-hybrid screeningsystem (42). The CRYBP1-binding sequence on the Col2a1enhancer is located just 1 bp upstream from the SOX9 bindingsequence (25). Although CRYBP1 does not contain theKRAB domain, CRYBP1 competes for binding to the Col2a1enhancer with SOX9 and inhibits transcriptional activationby SOX9 (42). Cotransfection experiments revealed thatCRYBP1 did not inhibit promoter activity of Col11a2 and thatNT2 could not reduce enhancer activity of Col2a1 (data notshown). Since the 24-bp sequence of Col11a2 does not containthe SOX9 binding site, it is likely that NT2 and CRYBP1
FIG. 7. Cotransfection experiments showing the suppressive effectof NT2 on Col11a2 promoter activity. RCS cells were transiently trans-fected with 2 �g of reporter plasmid (pGL3-Control, p530luc, orp530Mluc) along with a total of 4 �g of pCA1F expression vector thateither did or did not contain the NT2 cDNA as indicated. NT2 reducedCol11a2 promoter activity of p530luc to 21% of the control in adose-dependent manner, whereas it did not affect the luciferase activ-ity of the pGL3-Control plasmid driven by the SV40 promoter. NT2did not affect the luciferase activity of p530Mluc, which has substitu-tion mutations at the NT2 binding site. A Renilla luciferase expressionvector, pRL-SV40, was used as an internal control for transfectionefficiency. The relative luciferase activities are average values thestandard errors for three independent transfected cultures from tworepeated experiments.
4264 TANAKA ET AL. MOL. CELL. BIOL.
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
suppress promoter activity of two different cartilage collagengenes through distinct mechanisms.
These observations indicate that these two zinc finger pro-teins may have different target genes and functions. On theother hand, in situ hybridization and Northern blot assaysdemonstrated that CRYBP1 and NT2 almost coexpressed indeveloping mouse embryos, tissues in newborn mice, and var-ious cell lines. Both CRYBP1 and NT2 are expressed in un-differentiated mesenchymal cells in early stages of mouse em-bryos, downregulated during the mesenchymal condensations,and strongly expressed in hypertrophic chondrocytes (42).Thus, these two zinc finger proteins might coordinately func-tion in the development of cartilaginous tissues.
In conclusion, we have identified a repressor protein, NT2,
that inhibits the chondrocyte-specific activity of the Col11a2promoter via the KRAB-A subdomain. Our results suggestthat this negative regulatory factor plays a role in controllingthe expression of Col11a2 and underscore the importance ofboth positive and negative control mechanisms in cartilagedevelopment.
ACKNOWLEDGMENTS
This work was supported by NIH intramural funds; a Health ScienceResearch Grant for Research on Human Genome, Tissue Engineer-ing, and Food Biotechnology from the Ministry of Health, Labour, andWelfare, Japan; and a grant from the Japan Orthopaedics and Trau-matology Foundation (no. 0116).
FIG. 8. The effect of deletions in NT2 on the inhibition of Col11a2 promoter activity. (A) Full-length FPM315 has an LeR domain, a KRAB-Asubdomain and nine zinc finger motifs as reported by Yokoyama et al. (56). In NT2�4, the KRAB-A subdomain of NT2 is deleted. The predictedmolecular weight of each protein is indicated at the right of the panel. (B) Western blot analysis of deletion mutant NT2 proteins. Invitro-translated proteins (2 �g) were fractionated by SDS-PAGE, blotted, and incubated with anti-Flag antibodies. Lane 1, full-length NT2; lane2, NT2�1; lane 3, NT2�2; lane 4, NT2�3; lane 5, NT2�4. Protein standards are indicated on the left. (C) Cotransfection experiments showing theeffect of the KRAB-A subdomain in NT2 on Col11a2 promoter activity. RCS cells were transiently transfected with 2 �g of p530luc along witha total of 4 �g of pCA1F expression vector that either did or did not contain the deleted NT2 cDNA as indicated. Full-length NT2 and NT2�1significantly inhibited Col11a2 promoter activity, consistent with the data shown in Fig. 7. However, NT2�2 and NT2�3 did not affect the luciferaseactivity of p530luc. Furthermore, NT2�4 also failed to inhibit Col11a2 promoter activity. pRL-SV40 was used as an internal control for transfectionefficiency. The relative luciferase activities are average values the standard errors for three independent transfected cultures from two repeatedexperiments.
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4265
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

REFERENCES
1. Abrink, M., J. A. Ortiz, C. Mark, C. Sanchez, C. Looman, L. Hellman, P.Chambon, and R. Losson. 2001. Conserved interaction between distinctKruppel-associated box domains and the transcriptional intermediary factor1 beta. Proc. Natl. Acad. Sci. USA 98:1422–1426.
1a.Adamson, M. C., J. Silver, and C. A. Kozak. 1991. The mouse homolog of theGibbon ape leukemia virus receptor: genetic mapping and a possible recep-tor function in rodents. Virology 183:778–781.
2. Attar, R. M., and M. Z. Gilman. 1992. Expression cloning of a novel zincfinger protein that binds to the c-fos serum response element. Mol. Cell. Biol.12:2432–2443.
3. Bellefroid, E. J., D. A. Poncelet, P. J. Lecocq, O. Revelant, and J. A. Martial.1991. The evolutionarily conserved Kruppel-associated box domain defines asubfamily of eukaryotic multifingered proteins. Proc. Natl. Acad. Sci. USA88:3608–3612.
4. Berg, J. M., and Y. Shi. 1996. The galvanization of biology: a growingappreciation for the roles of zinc. Science 271:1081–1085.
5. Bernard, M., H. Yoshioka, E. Rodriguez, M. Van der Rest, T. Kimura, Y.Ninomiya, B. R. Olsen, and F. Ramirez. 1988. Cloning and sequencing ofpro-alpha 1 (XI) collagen cDNA demonstrates that type XI belongs to thefibrillar class of collagens and reveals that the expression of the gene is notrestricted to cartilaginous tissue. J. Biol. Chem. 263:17159–17166.
6. Bridgewater, L. C., V. Lefebvre, and B. de Crombrugghe. 1998. Chondrocyte-specific enhancer elements in the Col11a2 gene resemble the Col2a1 tissue-specific enhancer. J. Biol. Chem. 273:14998–15006.
7. Chowdhury, K., M. Goulding, C. Walther, K. Imai, and H. Fickenscher.1992. The ubiquitous transactivator Zfp-38 is upregulated during spermat-ogenesis with differential transcription. Mech. Dev. 39:129–142.
8. Dawson, S. R., D. L. Turner, H. Weintraub, and S. M. Parkhurst. 1995.Specificity for the hairy/enhancer of split basic helix-loop-helix (bHLH)proteins maps outside the bHLH domain and suggests two separable modesof transcriptional repression. Mol. Cell. Biol. 15:6923–6931.
9. de Haan, G., S. Chusacultanachai, C. Mao, B. S. Katzenellenbogen, andD. J. Shapiro. 2000. Estrogen receptor-KRAB chimeras are potent ligand-dependent repressors of estrogen-regulated gene expression. J. Biol. Chem.275:13493–13501.
10. Denny, P., and A. Ashworth. 1991. A zinc finger protein-encoding geneexpressed in the post-meiotic phase of spermatogenesis. Gene 106:221–227.
11. Deweindt, C., O. Albagli, F. Bernardin, P. Dhordain, S. Quief, D. Lantoine,J. P. Kerckaert, and D. Leprince. 1995. The LAZ3/BCL6 oncogene encodesa sequence-specific transcriptional inhibitor: a novel function for the BTB/POZ domain as an autonomous repressing domain. Cell Growth Differ.6:1495–1503.
12. Foster, J. W., M. A. Dominguez-Steglich, S. Guioli, G. Kowk, P. A. Weller, M.Stevanovic, J. Weissenbach, S. Mansour, I. D. Young, P. N. Goodfellow, et al.1994. Campomelic dysplasia and autosomal sex reversal caused by mutationsin an SRY-related gene. Nature 372:525–530.
13. Friedman, J. R., W. J. Fredericks, D. E. Jensen, D. W. Speicher, X. P. Huang,E. G. Neilson, and F. J. Rauscher III. 1996. KAP-1, a novel corepressor forthe highly conserved KRAB repression domain. Genes Dev. 10:2067–2078.
14. Furuto, D. K., and E. J. Miller. 1983. Different levels of glycosylation con-tribute to the heterogeneity of alpha 1(II) collagen chains derived from atransplantable rat chondrosarcoma. Arch. Biochem. Biophys. 226:604–611.
15. Gray, S., and M. Levine. 1996. Transcriptional repression in development.Curr. Opin. Cell Biol. 8:358–364.
16. Grimes, H. L., T. O. Chan, P. A. Zweidler-McKay, B. Tong, and P. N.Tsichlis. 1996. The Gfi-1 proto-oncoprotein contains a novel transcriptionalrepressor domain, SNAG, and inhibits G1 arrest induced by interleukin-2withdrawal. Mol. Cell. Biol. 16:6263–6272.
17. Harley, V. R., R. Lovell-Badge, and P. N. Goodfellow. 1994. Definition of aconsensus DNA binding site for SRY. Nucleic Acids Res. 22:1500–1501.
18. Hirota, S., A. Ito, E. Morii, A. Wanaka, M. Tohyama, Y. Kitamura, and S.Nomura. 1992. Localization of mRNA for c-kit receptor and its ligand in thebrain of adult rats: an analysis using in situ hybridization histochemistry.Brain Res. Mol. Brain Res. 15:47–54.
19. Jheon, A. H., B. Ganss, S. Cheifetz, and J. Sodek. 2001. Characterization ofa novel KRAB/C2H2 zinc finger transcription factor involved in bone de-velopment. J. Biol. Chem. 276:18282–18289.
20. Johnson, A. D. 1995. The price of repression. Cell 81:655–658.21. Kim, S. S., Y. M. Chen, E. O’Leary, R. Witzgall, M. Vidal, and J. V. Bon-
ventre. 1996. A novel member of the RING finger family, KRIP-1, associateswith the KRAB-A transcriptional repressor domain of zinc finger proteins.Proc. Natl. Acad. Sci. USA 93:15299–15304.
22. Kimura, T., K. S. Cheah, S. D. Chan, V. C. Lui, M. G. Mattei, M. van derRest, K. Ono, E. Solomon, Y. Ninomiya, and B. R. Olsen. 1989. The humanalpha 2(XI) collagen (COL11A2) chain. Molecular cloning of cDNA andgenomic DNA reveals characteristics of a fibrillar collagen with differences ingenomic organization. J. Biol. Chem. 264:13910–13916.
23. Klug, A., and J. W. Schwabe. 1995. Protein motifs 5. Zinc fingers. FASEB J.9:597–604.
23a.Kozak, C. A., M. Peyser, M. Krall, T. M. Mariano, C. S. Kumar, S. Pestka,
and B. A. Mock. 1990. Molecular genetic markers spanning mouse chromo-some 10. Genomics 8:519–524.
24. Lee, P. L., T. Gelbart, C. West, M. Adams, R. Blackstone, and E. Beutler.1997. Three genes encoding zinc finger proteins on human chromosome6p21.3: members of a new subclass of the Kruppel gene family containing theconserved SCAN box domain. Genomics 43:191–201.
25. Lefebvre, V., W. Huang, V. R. Harley, P. N. Goodfellow, and B. de Crom-brugghe. 1997. SOX9 is a potent activator of the chondrocyte-specific en-hancer of the proá1(II) collagen gene. Mol. Cell. Biol. 17:2336–2346.
26. Li, J. J., and I. Herskowitz. 1993. Isolation of ORC6, a component of theyeast origin recognition complex by a one-hybrid system. Science 262:1870–1874.
27. Li, Y., D. A. Lacerda, M. L. Warman, D. R. Beier, H. Yoshioka, Y. Ninomiya,J. T. Oxford, N. P. Morris, K. Andrikopoulos, F. Ramirez, et al. 1995. Afibrillar collagen gene, Col11a1, is essential for skeletal morphogenesis. Cell80:423–430.
28. Liu, Y., H. Li, K. Tanaka, N. Tsumaki, and Y. Yamada. 2000. Identificationof an enhancer sequence within the first intron required for cartilage-specifictranscription of the alpha2(XI) collagen gene. J. Biol. Chem. 275:12712–12718.
29. Margolin, J. F., J. R. Friedman, W. K. Meyer, H. Vissing, H. J. Thiesen, andF. J. Rauscher III. 1994. Kruppel-associated boxes are potent transcriptionalrepression domains. Proc. Natl. Acad. Sci. USA 91:4509–4513.
30. Mendler, M., S. G. Eich-Bender, L. Vaughan, K. H. Winterhalter, and P.Bruckner. 1989. Cartilage contains mixed fibrils of collagen types II, IX, andXI. J. Cell Biol. 108:191–197.
31. Moosmann, P., O. Georgiev, B. Le Douarin, J. P. Bourquin, and W.Schaffner. 1996. Transcriptional repression by RING finger protein TIF1beta that interacts with the KRAB repressor domain of KOX1. NucleicAcids Res. 24:4859–4867.
32. Morris, N. P., and H. P. Bachinger. 1987. Type XI collagen is a heterotrimerwith the composition (1 alpha, 2 alpha, 3 alpha) retaining non-triple-helicaldomains. J. Biol. Chem. 262:11345–11350.
33. Mukhopadhyay, K., V. Lefebvre, G. Zhou, S. Garofalo, J. H. Kimura, and B.de Crombrugghe. 1995. Use of a new rat chondrosarcoma cell line to delin-eate a 119-base pair chondrocyte-specific enhancer element and to defineactive promoter segments in the mouse pro-alpha 1(II) collagen gene.J. Biol. Chem. 270:27711–27719.
34. Peng, H., G. E. Begg, D. C. Schultz, J. R. Friedman, D. E. Jensen, D. W.Speicher, and F. J. Rauscher III. 2000. Reconstitution of the KRAB–KAP-1repressor complex: a model system for defining the molecular anatomy ofRING–B box–coiled-coil domain-mediated protein-protein interactions. J.Mol. Biol. 295:1139–1162.
35. Pengue, G., V. Calabro, P. C. Bartoli, A. Pagliuca, and L. Lania. 1994.Repression of transcriptional activity at a distance by the evolutionarilyconserved KRAB domain present in a subfamily of zinc finger proteins.Nucleic Acids Res. 22:2908–2914.
36. Pengue, G., V. Calabro, P. Cannada-Bartoli, P. De Luca, T. Esposito, P.Taillon-Miller, S. LaForgia, T. Druck, K. Huebner, M. D’Urso, et al. 1993.YAC-assisted cloning of transcribed sequences from the human chromo-some 3p21 region. Hum. Mol. Genet. 2:791–796.
37. Pengue, G., and L. Lania. 1996. Kruppel-associated box-mediated repressionof RNA polymerase II promoters is influenced by the arrangement of basalpromoter elements. Proc. Natl. Acad. Sci. USA 93:1015–1020.
38. Petit, B., M. C. Ronziere, D. J. Hartmann, and D. Herbage. 1993. Ultra-structural organization of type XI collagen in fetal bovine epiphyseal carti-lage. Histochemistry 100:231–239.
39. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Extraction, purification,and analysis of messenger RNA from eukaryotic cells, p. 7.1–7.87. In C.Noran (ed.), Molecular cloning: a laboratory manual. Cold Spring HarborPress, New York, N.Y.
40. Schiestl, R. H., and R. D. Gietz. 1989. High efficiency transformation ofintact yeast cells using single stranded nucleic acids as a carrier. Curr. Genet.16:339–346.
41. Shukunami, C., C. Shigeno, T. Atsumi, K. Ishizeki, F. Suzuki, and Y. Hiraki.1996. Chondrogenic differentiation of clonal mouse embryonic cell lineATDC5 in vitro: differentiation-dependent gene expression of parathyroidhormone (PTH)/PTH-related peptide receptor. J. Cell Biol. 133:457–468.
42. Tanaka, K., Y. Matsumoto, F. Nakatani, Y. Iwamoto, and Y. Yamada. 2000.A zinc finger transcription factor, áA-crystallin binding protein 1, is a neg-ative regulator of the chondrocyte-specific enhancer of the á1(II) collagengene. Mol. Cell. Biol. 20:4428–4435.
43. Tsumaki, N., and T. Kimura. 1995. Differential expression of an acidicdomain in the amino-terminal propeptide of mouse pro-alpha 2(XI) collagenby complex alternative splicing. J. Biol. Chem. 270:2372–2378.
44. Tsumaki, N., T. Kimura, Y. Matsui, K. Nakata, and T. Ochi. 1996. Separablecis-regulatory elements that contribute to tissue- and site- specific alpha2(XI) collagen gene expression in the embryonic mouse cartilage. J. CellBiol. 134:1573–1582.
45. Tsumaki, N., T. Kimura, K. Tanaka, J. H. Kimura, T. Ochi, and Y. Yamada.1998. Modular arrangement of cartilage- and neural tissue-specific cis-ele-
4266 TANAKA ET AL. MOL. CELL. BIOL.
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

ments in the mouse alpha2(XI) collagen promoter. J. Biol. Chem. 273:22861–22864.
46. Vaughan, L., M. Mendler, S. Huber, P. Bruckner, K. H. Winterhalter, M. I.Irwin, and R. Mayne. 1988. D-Periodic distribution of collagen type IX alongcartilage fibrils. J. Cell Biol. 106:991–997.
47. Vikkula, M., E. C. Mariman, V. C. Lui, N. I. Zhidkova, G. E. Tiller, M. B.Goldring, S. E. van Beersum, M. C. de Waal Malefijt, F. H. van den Hoogen,H. H. Ropers, et al. 1995. Autosomal dominant and recessive osteochondro-dysplasias associated with the COL11A2 locus. Cell 80:431–437.
48. Vissing, H., W. K. Meyer, L. Aagaard, N. Tommerup, and H. J. Thiesen.1995. Repression of transcriptional activity by heterologous KRAB domainspresent in zinc finger proteins. FEBS Lett. 369:153–157.
49. Wagner, T., J. Wirth, J. Meyer, B. Zabel, M. Held, J. Zimmer, J. Pasantes,F. D. Bricarelli, J. Keutel, E. Hustert, et al. 1994. Autosomal sex reversal andcampomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell 79:1111–1120.
50. Wang, M. M., and R. R. Reed. 1993. Molecular cloning of the olfactoryneuronal transcription factor Olf-1 by genetic selection in yeast. Nature364:121–126.
51. Williams, A. J., L. M. Khachigian, T. Shows, and T. Collins. 1995. Isolationand characterization of a novel zinc-finger protein with transcription repres-sor activity. J. Biol. Chem. 270:22143–22152.
52. Wirth, J., T. Wagner, J. Meyer, R. A. Pfeiffer, H. U. Tietze, W. Schempp, andG. Scherer. 1996. Translocation breakpoints in three patients with cam-
pomelic dysplasia and autosomal sex reversal map more than 130 kb fromSOX9. Hum. Genet. 97:186–193.
53. Witzgall, R., E. O’Leary, A. Leaf, D. Onaldi, and J. V. Bonventre. 1994. TheKruppel-associated box-A (KRAB-A) domain of zinc finger proteins medi-ates transcriptional repression. Proc. Natl. Acad. Sci. USA 91:4514–4518.
54. Wright, E., M. R. Hargrave, J. Christiansen, L. Cooper, J. Kun, T. Evans, U.Gangadharan, A. Greenfield, and P. Koopman. 1995. The Sry-related geneSox9 is expressed during chondrogenesis in mouse embryos. Nat. Genet.9:15–20.
55. Yang, X. W., R. Zhong, and N. Heintz. 1996. Granule cell specification in thedeveloping mouse brain as defined by expression of the zinc finger transcrip-tion factor RU49. Development 122:555–566.
56. Yokoyama, M., M. Nakamura, K. Okubo, K. Matsubara, Y. Nishi, T. Mat-sumoto, and A. Fukushima. 1997. Isolation of a cDNA encoding a widelyexpressed novel zinc finger protein with the LeR and KRAB-A domains.Biochim. Biophys. Acta 1353:13–17.
57. Zheng, L., H. Pan, S. Li, A. Flesken-Nikitin, P. L. Chen, T. G. Boyer, andW. H. Lee. 2000. Sequence-specific transcriptional corepressor function forBRCA1 through a novel zinc finger protein, ZBRK1. Mol. Cell 6:757–768.
58. Zhu, Z., B. Ma, R. J. Homer, T. Zheng, and J. A. Elias. 2001. Use of thetetracycline-controlled transcriptional silencer (tTS) to eliminate transgeneleak in inducible overexpression transgenic mice. J. Biol. Chem. 276:25222–25229.
VOL. 22, 2002 REPRESSOR FOR Col11a2 PROMOTER 4267
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

MOLECULAR AND CELLULAR BIOLOGY, Nov. 2006, p. 8215–8216 Vol. 26, No. 210270-7306/06/$08.00�0 doi:10.1128/MCB.01723-06
RETRACTION
A Kruppel-Associated Box–Zinc Finger Protein, NT2, Represses Cell-Type-Specific Promoter Activity of the �2(XI) Collage Gene
Kazuhiro Tanaka, Noriyuki Tsumaki, Christine A. Kozak, Yoshihiro Matsumoto, Fumihiko Nakatani,Yukihide Iwamoto, and Yoshihiko Yamada
Craniofacial Developmental Biology and Regeneration Branch, National Institute of Dental and Craniofacial Research, andLaboratory of Molecular Microbiology, National Institute of Allergy and Infectious Diseases, National Institutes of Health,
Bethesda, Maryland 20892, and Department of Orthopaedic Surgery, Graduate School of Medical Sciences,Kyushu University, Fukuoka 812-8582, Japan
Volume 22, no. 12, p. 4256–4267, 2002. Figures 2D, 4, 5 [panel �1(X)], 6B, and 8B are not faithful representations of theiroriginal source data and are therefore being retracted. Corrected figures are shown below. Use of the incorrect figures had noeffect on the other data or the basic results and conclusions of the paper.
Page 4259, legend to Fig. 2D: “The small arrow marks supershifted NT2-promoter complexes by anti-NT2 antibodies (lanes 4and 6)” should be replaced by “Anti-NT2 antibody reduced NT2-promoter complex formation (lanes 4 and 6).”
Page 4265, legend to Fig. 8B: A new sentence, “The asterisks indicate protein bands with expected sizes,” should be includedat the end of the legend.
FIG. 2D
Ant
S
nt
lb
nt
r v
nt
rv
ED
CB
GF
FIG. 4
FIG. 5
8215
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from
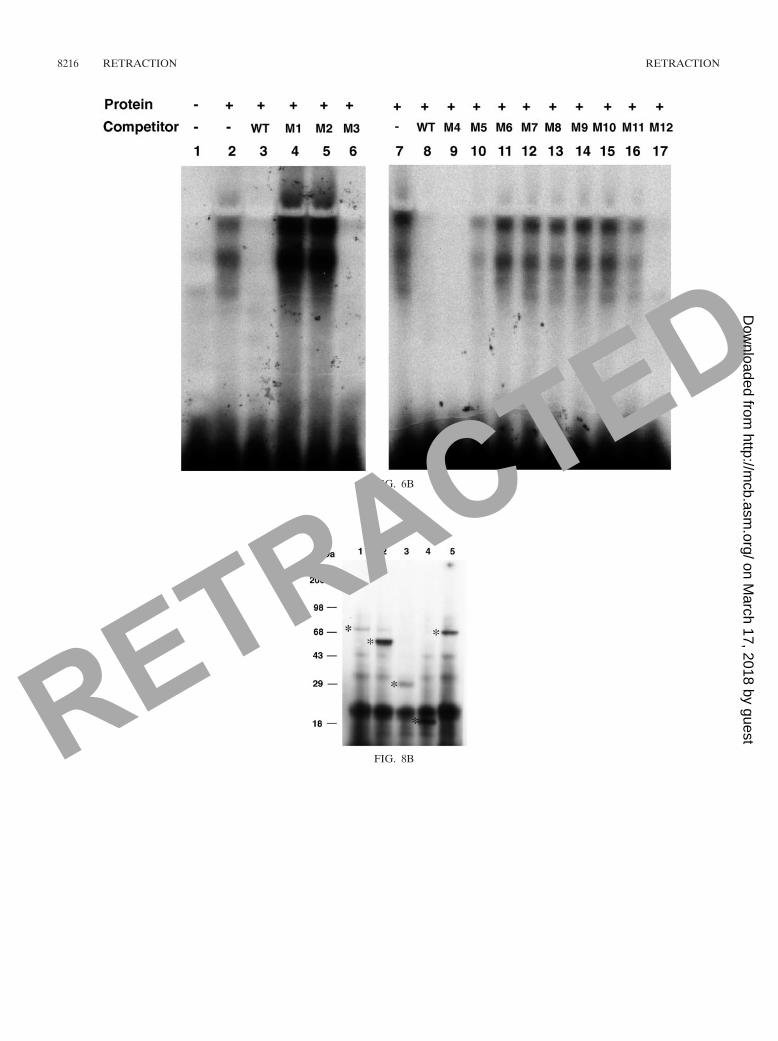
FIG. 6B
FIG. 8B
8216 RETRACTION RETRACTION
RETRACTED
on March 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

MOLECULAR AND CELLULAR BIOLOGY, Nov. 2006, p. 8215–8216 Vol. 26, No. 210270-7306/06/$08.00�0 doi:10.1128/MCB.01723-06
RETRACTION
A Kruppel-Associated Box–Zinc Finger Protein, NT2, Represses Cell-Type-Specific Promoter Activity of the �2(XI) Collage Gene
Kazuhiro Tanaka, Noriyuki Tsumaki, Christine A. Kozak, Yoshihiro Matsumoto, Fumihiko Nakatani,Yukihide Iwamoto, and Yoshihiko Yamada
Craniofacial Developmental Biology and Regeneration Branch, National Institute of Dental and Craniofacial Research, andLaboratory of Molecular Microbiology, National Institute of Allergy and Infectious Diseases, National Institutes of Health,
Bethesda, Maryland 20892, and Department of Orthopaedic Surgery, Graduate School of Medical Sciences,Kyushu University, Fukuoka 812-8582, Japan
Volume 22, no. 12, p. 4256–4267, 2002. Figures 2D, 4, 5 [panel �1(X)], 6B, and 8B are not faithful representations of theiroriginal source data and are therefore being retracted. Corrected figures are shown below. Use of the incorrect figures had noeffect on the other data or the basic results and conclusions of the paper.
Page 4259, legend to Fig. 2D: “The small arrow marks supershifted NT2-promoter complexes by anti-NT2 antibodies (lanes 4and 6)” should be replaced by “Anti-NT2 antibody reduced NT2-promoter complex formation (lanes 4 and 6).”
Page 4265, legend to Fig. 8B: A new sentence, “The asterisks indicate protein bands with expected sizes,” should be includedat the end of the legend.
FIG. 2D
Ant
S
nt
lb
nt
r v
nt
rv
ED
CB
GF
FIG. 4
FIG. 5
8215

FIG. 6B
FIG. 8B
8216 RETRACTION RETRACTION

MOLECULAR AND CELLULAR BIOLOGY, June 2009, p. 3453 Vol. 29, No. 120270-7306/09/$08.00�0 doi:10.1128/MCB.00411-09
RETRACTION
A Zinc Finger Transcription Factor, �A-Crystallin Binding Protein 1,Is a Negative Regulator of the Chondrocyte-Specific
Enhancer of the �1(II) Collagen GeneKazuhiro Tanaka, Yoshihiro Matsumoto, Fumihiko Nakatani,
Yukihide Iwamoto, and Yoshihiko YamadaCraniofacial Developmental Biology and Regeneration Branch, National Institute of Dental and Craniofacial Research,
National Institutes of Health, Bethesda, Maryland 20892, and Department of Orthopaedic Surgery,Graduate School of Medical Sciences, Kyushu University, Fukuoka 812-8582, Japan
A Kruppel-Associated Box–Zinc Finger Protein, NT2, Represses Cell-Type-Specific Promoter Activity of the �2(XI) Collagen Gene
Kazuhiro Tanaka, Noriyuki Tsumaki, Christine A. Kozak, Yoshihiro Matsumoto, Fumihiko Nakatani,Yukihide Iwamoto, and Yoshihiko Yamada
Craniofacial Developmental Biology and Regeneration Branch, National Institute of Dental and Craniofacial Research, andLaboratory of Molecular Microbiology, National Institute of Allergy and Infectious Diseases, National Institutes of Health,
Bethesda, Maryland 20892, and Department of Orthopaedic Surgery, Graduate School of Medical Sciences,Kyushu University, Fukuoka 812-8582, Japan
Volume 20, no. 12, p. 4428–4435, 2000, and volume 22, no. 12, p. 4256–4267, 2002. These two articles were subject to previouspartial retractions (volume 26, no. 13, p. 5202, 2006, and volume 26, no. 21, p. 8215–8216, 2006, respectively). Since our originalpartial retractions, questions have arisen concerning the validity of other figures in these articles that were provided by one of theauthors (K.T.). We therefore retract fully these two articles as well as the previous partial retractions.
3453