a histological variant of white spot syndrome virus (wssv) from the kingdom of saudi arabia
TRANSCRIPT

Journal of Invertebrate Pathology 113 (2013) 82–85
Contents lists available at SciVerse ScienceDirect
Journal of Invertebrate Pathology
journal homepage: www.elsevier .com/ locate / j ip
Short Communication
A histological variant of white spot syndrome virus (WSSV) from the Kingdomof Saudi Arabia
Kathy F.J. Tang ⇑, Carlos R. Pantoja, Rita M. Redman, Donald V. LightnerDepartment of Veterinary Science and Microbiology, University of Arizona, Tucson, AZ 85721, United States
a r t i c l e i n f o
Article history:Received 24 October 2012Accepted 18 February 2013Available online 27 February 2013
Keywords:White spot syndrome virusWSSVHistology variantImmunohistochemistryIn situ hybridizationPenaeid shrimp
0022-2011/$ - see front matter � 2013 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.jip.2013.02.004
⇑ Corresponding author. Fax: +1 520 6214899.E-mail address: [email protected] (K.F.J. Tang
a b s t r a c t
White spot syndrome virus (WSSV) is highly pathogenic to penaeid shrimp. The major targets of WSSVinfection are tissues of ectodermal and mesodermal embryonic origin, predominantly the cuticular epi-thelium and subcuticular connective tissues. Recently, we discovered a WSSV variant in Penaeus indicusthat heavily infects the subcuticular connective tissue, with very slight indications in the cuticular epithe-lium. The variant was also unusual in that WSSV accumulations were found in the interstitial spaces ofboth the subcuticular connective tissue and the lymphoid organ. This WSSV variant was confirmedthrough immunohistochemistry with an anti-WSSV VP28 monoclonal antibody, and also by in situhybridization with a VP28 DNA probe. By in situ hybridization, shrimp with variant and typical histologywere shown a deletion in ORF94, which is characteristic of a new type of WSSV found in Saudi Arabia;apparently, the loss of this ORF is not associated with the variant’s reduced capability of infecting thecuticular epithelium cells.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
White spot syndrome virus (WSSV) has caused high mortalitiesin many species of penaeid shrimp and other aquatic crustaceansthroughout the world (Lightner, 2011).
The clinical signs of WSSV disease in penaeids include theappearance of white spots inside the carapace and a reddish discol-oration of the body (Lightner, 1996). Histological analysis revealsdistinct hypertrophied nuclei within the tissues of ectodermaland mesodermal embryonic origin. WSSV is a large (70–150 nm� 275–380 nm), enveloped, dsDNA virus with sizes ranged from292 to 307 kb (Van Huten et al., 2001; Yang et al., 2001).
During 2010–2012, WSSV was detected in the Kingdom of SaudiArabia (OIE, 2011) where it has severely affected shrimp produc-tion. Through PCR and DNA sequencing, we discovered a newgenotype of WSSV with a 1.5 kb deletion in ORF94/95 (Tanget al., 2012). In this report, we describe a Saudi Arabia isolateshowing atypical histopathology. This variant showed conspicu-ously reduced infection in the cuticular epithelium while other tar-geted tissues and organs presented with heavy infections. Inaddition, shrimp showing the variant histology had substantialaccumulations of WSSV in the interstitial spaces; these interstitialWSSV accumulations were found both in the subcuticular connec-tive tissue and in the lymphoid organ.
ll rights reserved.
).
2. Materials and methods
Samples of Indian white shrimp, Penaeus indicus (taxonomyaccording to Holthius, 1980) were collected from a shrimp farm(designated as farm B in Tang et al., 2012) 160 km north of Jazancity in Saudi Arabia during 2012. A total of 22 shrimp from 5grow-out ponds were fixed in Davidson’s solution and sent toAquaculture Pathology Laboratory at University of Arizona for his-tological examination. These samples were processed according toconventional techniques for paraffin embedding and sectioning.Paraffin sections were stained with Mayer–Bennett’s hematoxy-lin/eosin-phloxine (H&E) and examined under a light microscope.This submission also included parallel samples (hemolymphdrawn from the same shrimp) to be analyzed by PCR and RT-PCRfor the following pathogens: infectious hyperdermal and hemato-poietic necrosis virus (IHHNV), yellow head virus (YHV), Taura syn-drome virus (TSV) and WSSV. Only WSSV was detected.
For immunohistochemistry (IHC), the tissue sections (4 lmthick) were deparaffinized, rehydrated, then blocked with PBS con-taining 10% normal goat serum (NGS) and 2% NFM (non-fat milk)for 15 min at 37 �C. A volume of 100 ll of anti-WSSV MAb (AquaticDiagnostics Ltd., Scotland, UK) was applied to the tissue sectionsfor 1 h at room temperature. After 3 washes in PBS, the sectionswere reacted with a goat anti-mouse IgG, F(ab0)2 antibody conju-gated to alkaline phosphatase (Kirkegaard & Perry Laboratories,Maryland) for 30 min at 37 �C. The reactions were then developedwith nitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indoyl
K.F.J. Tang et al. / Journal of Invertebrate Pathology 113 (2013) 82–85 83
phosphate (X-phos). The sections were counterstained with Bis-marck brown and examined under light microscopy.
For in situ hybridization (ISH), WSSV VP28 and ORF94 probeswere used, these probes were prepared by PCR labeling with digox-igenin-11-dUTP as described by Mari et al. (1998). For VP28 probe(0.6-kb), the template was from a plasmid with an insert of VP28fragment, the primers are VP28-600F (50-TTCACTCTTTCGGTCGTGTC) and VP28-600R (50-CTCGGTCTCAGTGCCAGAGT). For ORF94probe (1.1-kb), the template was a plasmid with an insert ofORF94, the primers were ORF94-F (50-TCTACTCGAGGAGGTGAC-GAC) and ORF94-R (50-AGCAGGTGTGTACACATTTCATG). For ISH,the sections on slides were overlaid with a hybridization solution(4� SSC, 50% formamide, 1� Denhardt’s, 5% dextran sulfate,0.5 lg/ml salmon sperm DNA) containing a WSSV probe (0.2 lg/ml). Slides were placed on a heated surface at 90 �C for 10 min,and hybridized overnight at 37 �C. Final color development wasperformed with anti-digoxigenin antibody conjugated with analkaline phosphatase, NBT and X-phos.
3. Results and discussion
We describe here the first known instance of a peculiar mani-festation of WSSV histology in penaeid shrimp. During routinemonitoring in 2012, WSSV was detected in samples from a shrimpfarm culturing Penaeus indicus in Saudi Arabia. We found that
12-072B/1
12-072B/1
1
1
A
C
Cep
sCn
Lym
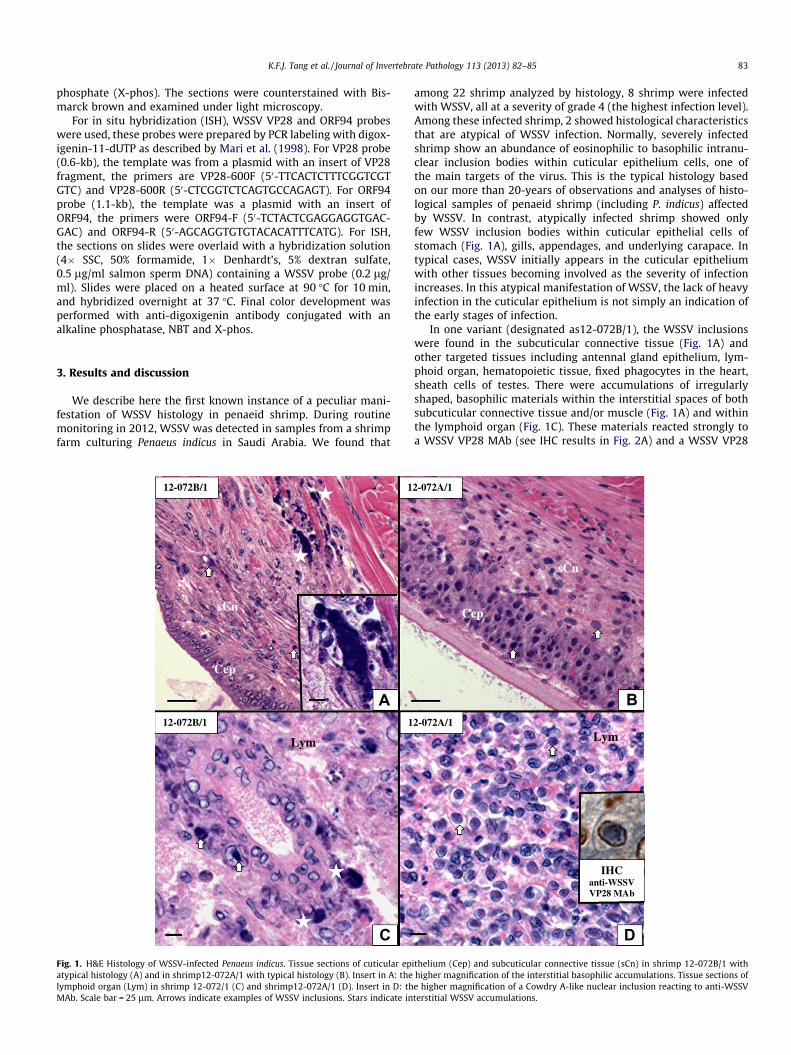
Fig. 1. H&E Histology of WSSV-infected Penaeus indicus. Tissue sections of cuticular epatypical histology (A) and in shrimp12-072A/1 with typical histology (B). Insert in A: thelymphoid organ (Lym) in shrimp 12-072/1 (C) and shrimp12-072A/1 (D). Insert in D: thMAb. Scale bar = 25 lm. Arrows indicate examples of WSSV inclusions. Stars indicate in
among 22 shrimp analyzed by histology, 8 shrimp were infectedwith WSSV, all at a severity of grade 4 (the highest infection level).Among these infected shrimp, 2 showed histological characteristicsthat are atypical of WSSV infection. Normally, severely infectedshrimp show an abundance of eosinophilic to basophilic intranu-clear inclusion bodies within cuticular epithelium cells, one ofthe main targets of the virus. This is the typical histology basedon our more than 20-years of observations and analyses of histo-logical samples of penaeid shrimp (including P. indicus) affectedby WSSV. In contrast, atypically infected shrimp showed onlyfew WSSV inclusion bodies within cuticular epithelial cells ofstomach (Fig. 1A), gills, appendages, and underlying carapace. Intypical cases, WSSV initially appears in the cuticular epitheliumwith other tissues becoming involved as the severity of infectionincreases. In this atypical manifestation of WSSV, the lack of heavyinfection in the cuticular epithelium is not simply an indication ofthe early stages of infection.
In one variant (designated as12-072B/1), the WSSV inclusionswere found in the subcuticular connective tissue (Fig. 1A) andother targeted tissues including antennal gland epithelium, lym-phoid organ, hematopoietic tissue, fixed phagocytes in the heart,sheath cells of testes. There were accumulations of irregularlyshaped, basophilic materials within the interstitial spaces of bothsubcuticular connective tissue and/or muscle (Fig. 1A) and withinthe lymphoid organ (Fig. 1C). These materials reacted strongly toa WSSV VP28 MAb (see IHC results in Fig. 2A) and a WSSV VP28
2-072A/1
2-072A/1
B
D
Cep
sCn
Lym
IHCanti-WSSVVP28 MAb
ithelium (Cep) and subcuticular connective tissue (sCn) in shrimp 12-072B/1 withhigher magnification of the interstitial basophilic accumulations. Tissue sections ofe higher magnification of a Cowdry A-like nuclear inclusion reacting to anti-WSSVterstitial WSSV accumulations.

84 K.F.J. Tang et al. / Journal of Invertebrate Pathology 113 (2013) 82–85
probe (see ISH results in Fig. 2C). The detailed structure of theseinterstitial WSSV accumulations cannot be determined withoutTEM examination (no samples were fixed for TEM). It is possiblethat they are the result of the lysis of infected cells. The remaining6 infected shrimp were shown to have a typical WSSV histology,that both cuticular epithelial and subcuticular connective tissueswere found with abundant inclusions (Fig. 1B), which were also de-tected in the lymphoid organ (Fig. 1D). No basophilic accumula-tions were found within the interstitial spaces in these tissues.There were abundant eosinophilic, centronuclear, Cowdry-A-likeinclusion bodies in the lymphoid organ (Fig. 1D). These viral inclu-sions were determined by IHC (see the inserted photograph) andISH (data not shown) to be WSSV. Cowdry-A-like inclusions are of-ten found at the early stages of WSSV infection (Lightner, 1996).
To determine if the atypical histology is due to WSSV infection,we applied the IHC with an anti-WSSV VP28 MAb. In this assay, theWSSV infected cells showed blue-black color precipitates in theantibody reacting sites after colorimetic detection. In the shrimp12-072B/1, strong signals were observed in the subcuticular con-nective tissues (Fig. 2A). The MAb reacted to the inclusions andto the interstitial basophilic accumulations found in subcuticularconnective tissue and in lymphoid organ. Some inclusions didnot react to the WSSV antibodies; it is possible that these werenot fully formed such that VP28 proteins had not been incorpo-rated into the viral envelopes. The cuticulum epithelium appearednot react to anti-WSSV antibodies. In shrimp 12-072A/1 withtypical histology, the inclusions in the cuticular epithelial and
sCn
sCn
sCn
A
C
E
Cep
Cep
Cep
12-072B/1
12-072B/1
12-072B/1
Fig. 2. Immunohistochemistry and in situ hybridization of WSSV-infected Penaeus indicculticular epithelium (Cep) and subcuticular connective tissue (sCn): (A) shrimp 12-07072B1, (D) 12-072A/1; with ORF94 probe: (E) 12-072B/1, (F) 12-072A/1. Scale bar = 25accumulations.
subcuticular connective tissues were reacted intensely to theanti-VP28 MAb (Fig. 2B).
The presence of WSSV was further confirmed with ISH using aVP28 probe.
In shrimp 12-072B/1, positive reactions were found in the sub-cuticular connective tissues (Fig. 2C), but almost none were foundwithin the cuticular epithelial cells. The ISH signals were strong inintranuclear inclusions and much weaker in the interstitial accu-mulations which were likely to be washed away during the pro-teinase K digestion in the pre-hybridization step. In the shrimp12-072A/1, strong reactions were found in the cuticular epitheliumand subcuticular connective tissue (Fig. 2D).
A new WSSV genotype lacking ORF94 was recently discoveredfrom some farms in Saudi Arabia. However, from the farm sampledin this study we previously found a WSSV isolate that had a com-plete ORF94 with 13-repeat units (Tang et al., 2012). This suggeststhat the new genotype is spreading, and led us also to suspect thatthe variant infection described here may be associated with thedeletion of ORF94. However, a probe targeting ORF94 did not reactto infected tissues exhibiting either variant (12-072B/1; Fig. 2E) ortypical (12-072A/1; Fig. 2F) histology. So, the variant histology isapparently not a consequence of the deletion. The probe reactedto a tissue section infected with a Thailand WSSV isolate knownto have an ORF94 and serving as a positive control (see inserts inFig. 2E and F).
Our finding increases the spectrum of WSSV histopathology,and this information will be useful for WSSV diagnosis.
sCn
sCn
sCn
B
D
F
Cep
Cep
Cep
12-072A/1
12-072A/1
12-072A/1
us. Immunohistochemical reaction using anti-WSSV VP28 MAb in tissue sections of2B/1, (B) shrimp 12-072A/1. In situ hybridization with WSSV VP28 probe: (C) 12-lm. Arrows indicate examples of WSSV inclusions. Stars indicate interstitial WSSV

K.F.J. Tang et al. / Journal of Invertebrate Pathology 113 (2013) 82–85 85
References
Holthuis, L.B., 1980. Shrimp and prawn of the world: an annotated catalogue ofspecies of interest to fisheries. FAO species catalog. FAO Fisheries Synopsis 125(1), Food and Agriculture Organization of the United Nations, Rome.
Lightner, D.V., 1996. A Handbook of Shrimp Pathology and Diagnostic Proceduresfor Diseases of Cultured Penaeid Shrimp. World Aquaculture Society, BatonRouge, LA.
Lightner, D.V., 2011. Virus diseases of farmed shrimp in the Western Hemisphere(The Americas): a review. J. Invertebr. Pathol. 106, 110–130.
Mari, J., Bonami, J.R., Lightner, D.V., 1998. Taura syndrome of penaeid shrimp:cloning of viral genome fragments and development of specific gene probes.Dis. Aquat. Organ. 33, 11–17.
OIE (Office International des Epizooties/World Animal Health Organization), 2011.Immediate notification, submitted date: 04/10/2011. Office International desEpizooties, Paris, France.
Tang, K.F.J., Navarro, S.A., Pantoja, C.R., Aranguren, F.L., Lightner, D.V., 2012. Newgenotype of white spot syndrome virus (WSSV) and Taura syndrome virus (TSV)from the Kingdom of Saudi Arabia. Dis. Aquat. Organ. 99, 179–185.
Van Hulten, M.C.W., Witeveldt, J., Peters, S., Kloosterboer, N., Tarchini, R., Fiers, M.,Sandbrink, H., Lankhorst, R.K., Vlak, J.M., 2001. The white spot syndrome virusDNA genome sequence. Virology 286, 7–22.
Yang, F., He, J., Lin, X., Li, Q., Pan, D., Zhang, X., Xu, X., 2001. Complete genomesequence of the shrimp white spot bacilliform virus. J. Virol. 75, 11811–11820.