a gene expression profile that defines colon cell...
TRANSCRIPT

[CANCER RESEARCH 62, 4791–4804, August 15, 2002]
A Gene Expression Profile That Defines Colon Cell Maturation in Vitro1
John M. Mariadason,2 Diego Arango, Georgia A. Corner, Maria J. Aranes, Kylie A. Hotchkiss, Wancai Yang, andLeonard H. AugenlichtDepartment of Oncology, Albert Einstein Cancer Center, Montefiore Medical Center, Bronx, New York 10467
ABSTRACT
Colonic epithelial cells undergo cell cycle arrest, lineage specific differ-entiation, and apoptosis, as they migrate along the crypt axis toward thelumenal surface. The Caco-2 colon carcinoma cell line models many ofthese phenotypic changes, in vitro. We used this model system and cDNAmicroarray analysis to characterize the genetic reprogramming that ac-companies colon cell differentiation. The analyses revealed extensive yetfunctionally coordinated alterations in gene expression during the differ-entiation program. Consistent with cell differentiation reflecting a morespecialized phenotype, the majority of changes (70%) were down-regula-tions of gene expression. Specifically, Caco-2 cell differentiation was ac-companied by the coordinate down-regulation of genes involved in cellcycle progression and DNA synthesis, which reflected the concomitantreduction in cell proliferation. Simultaneously, genes involved in RNAsplicing and transport, protein translation, folding, and degradation, werecoordinately down-regulated, paralleled by a reduction in protein synthe-sis. Conversely, genes involved in xenobiotic and drug metabolism wereup-regulated, which was linked to increased resistance of differentiatedcells to chemotherapeutic agents. Increased expression of genes involvedin extracellular matrix deposition, lipid transport, and lipid metabolismwere also evident. Underlying these altered profiles of expression, com-ponents of signal transduction pathways, and several transcription factorswere altered in expression.
INTRODUCTION
Cellular proliferation, lineage-specific differentiation, migration,and apoptosis are highly coordinated processes that occur in a sequen-tial and spatially organized manner along the colonic crypt axis. Stemcells at the base of the crypt give rise to progenitor cells, whichexpand by rapid proliferation before undergoing cell cycle arrestfollowed by differentiation along one of three cell lineages, absorp-tive, goblet, or enteroendocrine, as they migrate along the crypt axistoward the lumenal surface (1). The factors that regulate these pro-cesses are only partially understood, although a number of biochem-ical events including E-cadherin-mediated cell-cell and integrin-mediated cell-substratum adhesion, chemotactic gradients, ECM3 andmesenchymal components, cytokines, hormones and growth factors,have been implicated (2–5). Downstream of these stimuli, signalingpathways and transcription factors, including Tcf-4, MATH-1, thehomeobox genes cdx-1 and cdx-2, kruppel-like factor 4, and severalmembers of the forkhead family of transcription factors, have beenshown to play a role in the coordination of colonic cell maturation(5–9).
Whereas the genetic reprogramming induced by these stimuli to
bring about the differentiated phenotype are likely to be extensive andcomplex, understanding these events would enhance both our knowl-edge of the mechanisms of maintenance of colonic epithelial cellhomoeostasis, as well as the pathogenesis of colorectal tumorigenesis.Differentiation of colonic epithelial cells also remains a poorly de-fined process at the biochemical and molecular level. Typically,differentiation of absorptive cells is described as a cessation of DNAsynthesis, followed by morphological changes such as cellular polar-ization, and the formation of well-developed tight junctions andmicrovilli (1, 10). On a biochemical level, increased expression ofbrush border hydrolases such as alkaline phosphatase, sucrase isoma-ltase, and dipeptidylpeptidase IV have been described (11). However,other changes likely to occur as colonic epithelial cells migrateupwards along the crypt axis, including changes in signaling path-ways, interactions with the extracellular matrix, metabolic processes,adaptive responses to lumenal contents, and apoptotic pathways,remain only partially characterized.
Advances in cDNA microarray technologies have enabled the def-inition of global changes in gene expression (12). In the present studywe have used this technology, in combination with a unique modelsystem, the Caco-2 colon carcinoma cell line, to gain additionalinsight into the genetic reprogramming that accompanies colonicepithelial cell maturation. The Caco-2 cell line spontaneously under-goes contact inhibition-dependent cell cycle arrest and differentiationalong the absorptive cell lineage as a function of time in culture,modeling the phenotypic changes normal colonic epithelial cells un-dergo as they migrate along the crypt axis. This model has been usedextensively in the study of colon cell maturation (13–16).
Microarray analysis of 17,280 sequences revealed that maturationof Caco-2 cells is characterized by extensive reprogramming at themolecular level. Changes in expression occurred in an organizedmanner, with genes in subsets of functional categories highly enrichedin patterns of coordinate regulation. Consistent with the induction ofcellular differentiation, several previously defined markers of absorp-tive cell differentiation were up-regulated in expression. Classificationof named genes in the database into functional categories revealedthat, in general, genes involved in cell cycle regulation and nucleicacid synthesis were down-regulated, consistent with the parallel in-duction of cell cycle arrest. Coordinate down-regulation of genesinvolved in RNA processing, translation, protein folding, and degra-dation was also evident. Conversely, genes involved in xenobiotic anddrug detoxification, extracellular matrix deposition, and lipid metab-olism were up-regulated. A number of signal transduction pathwaysand transcription factors potentially involved in the coordination ofthese responses were identified. The complete database can be ac-cessed at our laboratory website.4
MATERIALS AND METHODS
Cell Culture. The Caco-2 human colon cancer cell line was obtained fromthe American Type Culture Collection. Cells were cultured in MEM (LifeTechnologies, Inc., Grand Island, NY), supplemented with 10% FCS, 0.1 mM
Received 3/21/02; accepted 6/19/02.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported in part by National Cancer Institute Grants CA88104 and PO 13330, anda Fellowship from the American Institute for Cancer Research (to J. M. M.).
2 To whom requests for reprints should be addressed, at Department of Oncology,Albert Einstein Cancer Center, Montefiore Medical Center, 111 East 210th Street, Bronx,NY 10467. Phone: (718) 920-2093; Fax: (718) 882-4464; E-mail: [email protected].
3 The abbreviations used are: ECM, extracellular matrix; EST, expressed sequence tag;PCNA, proliferating cell nuclear antigen; HUVEC, human umbilical vascular endothelialcell; MAP, microtubule-associated protein; UBP, ubiquitin-specific protease; RXR, reti-noid X receptor; PPAR, peroxisome proliferator-activated receptor; LXR, Liver X recep-tor; MAPK, mitogen-activated protein kinase; TCF, T cell factor.
4 Internet address: http://sequence.aecom.yu.edu/bioinf/Augenlicht/default.html. In ad-dition, the supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org).
4791
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

nonessential amino acids, 10 mM HEPES buffer, 100 units/ml penicillin, 100�g/ml streptomycin, and 0.25 �g/ml amphotericin (Life Technologies, Inc.).Cell cultures were maintained at 37°C in 95% air and 5% CO2.
RNA and Probe Preparation. Caco-2 cells were cultured to confluence(day 0), or for 2, 5, 14, or 21 days post confluence, with medium changed everysecond day. Cells were harvested and RNA isolated using the RNeasy kit(Qiagen, Valencia, CA). For each hybridization two separate probes wereprepared: one labeled with Cy3 (day 0) and the other with Cy5 (days 2, 5, 14,and 21). The probe preparation, hybridization conditions, and slide scanningprocedure were as described previously (17). Arrays used in this report wereprepared by the microarray facility at the Albert Einstein College of Medicine(18). For each time point, arrays were performed on two independent chipsgenerated from independent clone sets. The first chip contained 8,064 se-quences and the second 9,216 sequences, resulting in 17,280 sequences ana-lyzed for each time point.
Data Analysis. The Scanalyze software of Eisen et al. (19) was used togenerate signal and background intensities for each channel at each spot on themicroarray. Data were transferred to a Microsoft Excel spreadsheet where thesignal:background ratio for each channel was calculated as well as the ratiobetween these ratios (i.e., green signal:background ratio was divided by the redsignal:background ratio). Data were then normalized among arrays by express-ing this value relative to the average of these values for all of the expressedgenes on the array. The data were log (2) transformed and transferred toMicrosoft Access where genes changing in expression during the maturationprogram were identified. For graphical representation, data were clustered anddisplayed using the Eisen’s Cluster and Treeview programs (19). A gene wasincluded for additional analysis if the signal:background ratio was �1.25 foreither the red or green channel for at least two of the four time points. A genewas considered differentially expressed if the ratio of the red:green signal was�25% in at least two of the final three time points. These criteria are similarto those used previously (17) and are based on the 95% confidence intervaldetermined from self-hybridization experiments.
Gene Classification and Functional Group Analysis. Of the 17,280genes on the arrays, 39% were named genes, and 61% genes of unknownfunction or ESTs. Named genes were sorted in alphabetical order and catego-rized into 1 or more of 25 functional categories. Gene classification was basedon searches of the Genecards5 or Medline6 databases. Functional group anal-ysis was performed as described previously by Muller et al. (20) and is basedon the hypothesis that if a particular biological process is modulated during thedifferentiation program, a disproportionately higher percentage of genes in-volved in the regulation of that process will be altered in expression (20). Thelevel of significance was calculated from the binomial distribution, using thefollowing algorithm in Microsoft Excel: BINOMDIST (X, Y, Z, FALSE),where X is the number of genes regulated in a given functional group, Y is thenumber of genes in that functional group, and Z is the overall proportion ofgenes regulated in the experiment.
Measurement of Apoptosis. Caco-2 cells cultured to confluence (day 0) orfor 2, 5, 7, 14, or 21 days postconfluence were treated with 10 �M of cisplatin(Sigma, St. Louis, MO) or 1 �M camptothecin (Calbiochem, San Diego, CA)for 72 h. Adherent and nonadherent cell populations were pooled and stainedovernight with propidium iodide [50 mg/ml in 0.1% (w/v) sodium citrate and0.1% (v/v) Triton X-100; Sigma]. The extent of apoptosis was determined byquantification of the sub-diploid cell fraction by fluorescence-activated cellsorter analysis, as described previously (21).
Clonogenic Assay. Caco-2 cells cultured to confluence (day 0) or for 2, 5,7, 14, or 21 days postconfluence were treated with 10 �M cisplatin (Sigma) or1 �M camptothecin (Calbiochem) for 9 h. Medium was removed, cellstrysinized, counted, and reseeded in triplicate into six-well plates at a densityof 500 cells/well. Colonies formed over �3 weeks and were visualized bystaining with 1% crystal violet for 30 min.
Western Blotting. Protein isolation, Western blotting, and signal detectionwere performed as described previously (16). Anti-p21WAF-1/CIP-1, cyclin A,cyclin E, c-myc, PCNA, HMG-1, topoisomerase I, eIF2�, eIF4E, and E2Awere obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-thymi-dylate synthase was obtained from Lab Vision Corporation (Fremont, CA), andanti-�-actin from Sigma.
[3H]Thymidine and [14C]Leucine Uptake Experiments. For measure-ment of [3H]thymidine and [14C]leucine uptake, Caco-2 cells were grown toconfluence (day 0), or for 2, 5, 7, 14, and 21 days postconfluence, in 24-wellplates. At each time point, cells were pulsed with 0.5 �Ci/well [14C]leucine or0.5 �Ci/well [3H]thymidine (Amersham, Piscataway, NJ) for 8 h. Cells werewashed three times in cold PBS, harvested, and centrifuged at 1,500 rpm for5 min at 4°C. Pellets were resuspended in 0.5 ml of 10% trichloroacetic acidand 1% phosphotungstic acid (Sigma), vortexed, and centrifuged at 15,000 rpmfor 15 min. The supernatant was discarded, the process repeated on two moreoccasions, and the pellet air-dried overnight. The dried pellet was resuspendedin 0.3 ml of NCS tissue solubilizer (Amersham), mixed with 5 ml of liquidscintillant (Sigma), and counted in a � counter. Counts were corrected for totalcellular protein measured in parallel samples.
Boyden Chamber Assay of Chemotactic Endothelial Cell Migration.HUVECs were isolated from umbilical cords and cultured as described pre-viously (22). Confluent HUVEC monolayers (passages 2–4) were harvestedwith cell dissociation solution and suspended at 1 � 106/ml in M199 medium(Life Technologies, Inc.) supplemented with 1% serum. HUVECs (105 cells)were seeded into 8.0-�m pore transwell inserts (Costar, Cambridge, MA)precoated with 10 �g/ml fibronectin. Inserts containing HUVECs were placedinto a 24-well plate (Costar) containing M199 medium supplemented with 1%serum and incubated for 1 h at 37°C. HUVEC migration was stimulated bycoculture with Caco-2 cells grown to confluence (day 0), or for 2, 5, 7, 14, and21 days postconfluence. After 5 h, HUVECs were fluorescently stained with10 �M of cell tracker green (Molecular Probes, Eugene, OR) and the uppersurface of the insert swabbed to remove nonmigrated cells. Inserts werewashed three times with PBS, fixed in 3.7% formaldehyde, and mounted onmicroscope slides. HUVEC migration was quantified by counting the numberof cells in three random fields (100� total magnification) per insert. Data areexpressed as endothelial cells per field (mean � SE)/1 � 106 Caco-2 cells.
RESULTS
Changes in Gene Expression during Caco-2 Cell Differentia-tion. Caco-2 cells spontaneously undergo cell cycle arrest and differ-entiation along the absorptive cell lineage as a function of time inculture (15). Initial analysis revealed that 13,638 of the 17,280 se-quences analyzed (78.9%) were expressed to a significant degree inCaco-2 cells (ratio of signal:background was �1.25 in at least one ofthe two channels). By our criteria (see “Materials and Methods”),2,286 of these sequences (13.2%) were altered in expression duringthe maturation program. This comprised 697 genes that were in-creased and 1,589 genes that were down-regulated in expression. Thepercentage of genes expressed in Caco-2 cells (78.9%) is consistentwith what we have observed in a number of other colon cancer celllines,7 whereas the percentage of genes altered in expression (13.2%)confirms previous reports that the extent of alteration of gene expres-sion during differentiation or transformation is on the order of 10%(23, 24).
Functional Group Analysis. To understand the biological signif-icance of the overall changes in gene expression, genes in the databasewere categorized into 1 or more of 25 predefined functional groups(Table 1). As expected, the number of genes comprising the differentfunctional categories varied significantly. For example, 504 geneswere linked to a role in metabolism, whereas only 30 genes with a rolein neurotransmission were identified (Table 1). Functional groups thatshowed significant enrichment during Caco-2 cell maturation weredetermined by comparison of the percentage of genes altered inexpression in a particular functional group to the overall percentage ofchange during the maturation program (13.2%). Functional groupsthat showed highly significant alterations of expression (P � 0.001)included cell cycle, DNA synthesis and repair, RNA processing,translation, protein processing and transport, protein degradation,
5 Internet address: http://nciarray.nci.nih.gov/cards.6 Internet address: http://www.ncbi.nlm.nih.gov/entrez. 7 Unpublished observations.
4792
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

xenobiotic and drug detoxification, ECM-associated factors, metabo-lism, kinases and phosphatases, and transcription factors (Table 1).
In contrast, we observed only a minimal enrichment of apoptosis-related genes consistent with the fact that despite undergoing cellcycle arrest and differentiation, the basal level of apoptosis remainsunchanged during Caco-2 cell differentiation (Ref. 15; Fig. 5I). Fur-thermore, and consistent with the fact that Caco-2 are epithelial cells,we found no enrichment of genes involved in immunoinflammatoryevents or in neurotransmission (Table 1).
Of the 17,280 sequences analyzed, 10,549 were sequences of un-known function or ESTs. Interestingly, only 9% of these sequences(948) were altered in expression, which was significantly less than theoverall level of change (13.2%). These results indicate that namedgenes were more likely to be altered in expression than genes ofunknown function and may reflect the existence of a bias in terms ofgene discovery, with genes with more visible phenotypic effectstending to be discovered and characterized more quickly than thosewith more subtle biological effects.
Finally, to additionally validate the functional group analysis, werandomly chose 222 genes from the database. This number wasselected as it reflected the average size of the functional groups.Thirty-three of the 222 (14.9%) randomly chosen genes were alteredin expression during the maturation program, which was not signifi-cantly different from the overall percentage of genes changed, dem-onstrating that functional groups that showed significant enrichmenttruly reflected an underlying biological change.
Differentiation. The morphological changes that characterize co-lon cell differentiation encompass changes in cell polarization, withthe development of well-formed tight junctions and microvilli (13).Consistent with this and a number of previous reports (16, 25), weobserved increased expression of the intestinal brush border-associ-ated proteins dipeptidylpeptidase IV and villin, the adherens junctionprotein E-cadherin, and the tight junction protein claudin 7 (See Fig.
1).8 Importantly, we also observed reduced expression of the gobletcell-specific markers mucin-2 and intestinal trefoil factor 3 in accord-ance with Caco-2 cell differentiation occurring specifically along theabsorptive cell lineage (26). Increased ion-transport, and associatedwater absorption, is an additional characteristic of differentiatedcolonic epithelial cells and is a feature that manifests itself in differ-entiated Caco-2 cells through the formation of dome-like structures(13). Consistent with these changes, we observed increased expres-sion of a number of genes involved in the regulation of Na�, Cl�, K�,
H� and HCO3� exchange (Fig. 1; supplementary data).
Cell Cycle Genes. We and others have demonstrated previouslythat Caco-2 cell differentiation is accompanied by a parallel decreasein cell proliferation reflected in a reduction of the percentage of cellsin S phase (14, 15). This observation was confirmed by a 67%reduction in the rate of [3H]thymidine incorporation during the mat-uration program (Fig. 1H). To dissect the molecular mechanisms andpathways involved in the induction of cell cycle arrest, we analyzedthe expression profiles of a panel of 148 genes that we had identifiedpreviously as playing a role in cell cycle regulation. Thirty-eight ofthese (25.7%) were altered in expression during Caco-2 cell matura-tion, with the majority down-regulated in expression. Relative to theoverall level of change in gene expression (13.2%), this represented asignificant enrichment in this functional class (P � 0.0001). Specificgenes down-regulated included the cyclins A, B, B1, D1, D2, E, and F,the cell division cycle proteins cdc2, cdc6, cdc10, cdc25C, and cdc28,and the cyclin-dependent kinases, cdk-1 and cdk-2. Conversely,three genes with a role described previously in cell cycle arrest,p21WAF-1/CIP1, transducer of ERBB2 1 (TOB), and the fork head/winged helix family transcription factor checkpoint suppressor 1 (27,28) were up-regulated. The changes in expression of cyclin A, E, andp21WAF1/CIP1 were confirmed by Western blot (Fig. 1, A–C).
Caco-2 cell differentiation was also accompanied by reduced ex-
8 Supplementary data at internet address: http://cancerres.aacrjournals.org.
Table 1 Functional Group Analysis.
Table of functional groups. When applicable, genes in the microarray database were assigned to one of 25 pre-defined functional categories. The number and percentage of genesin each category that were altered in expression during Caco-2 cell maturation were determined, and the P value was determined as described in the methods.
Functional groupNumber of sequences
on arrayNumber altered in expression(upregulated/downregulated)
%changed p
Total 17280 2286 (697/1589) 13.2Angiogenesis 85 18 (8/10) 21.2 0.01382Apoptosis 103 18 (7/11) 17.5 0.04888Cell cycle 148 38 (3/35) 25.7 *P � 0.0001Cytokines & growth factors 137 26 (6/20) 19.0 0.01472Cytoskeleton/microtubules 192 38 (8/30) 19.8 *0.00295DNA synthesis and repair 209 59 (3/56) 28.2 *P � 0.0001Xenobiotic and drug metabolism 122 34 (22/12) 27.9 *P � 0.0001ESTs 10549 948 (320/628) 9.0 *P � 0.0001ECM-associated 209 43 (21/22) 20.6 *0.00092Immune/inflammatory 277 34 (17/17) 12.3 0.06511Intercellular signaling 51 11 (2/9) 21.6 0.03546Kinases/phosphatases 444 85 (18/67) 19.1 *P � 0.0001Membrane-associated 326 60 (17/43) 18.4 *0.00181Membrane channels 206 39 (11/28) 18.9 *0.00513Metabolism 495 102 (44/58) 20.6 *P � 0.0001Mitochondria-associated 121 25 (5/20) 20.7 *0.00698Neurotransmitters 29 3 (1/2) 10.3 0.21138Oncogenes 103 15 (3/12) 14.6 0.10254Protein processing 216 53 (1/52) 25.0 *P � 0.0001Protein degradation 158 37 (6/31) 23.4 *P � 0.0001Redox 167 31 (16/15) 18.6 0.01217RNA processing 121 35 (11/24) 28.9 *P � 0.0001Signal transduction 723 111 (36/75) 15.4 0.01056Transcription 564 108 (33/75) 19.1 *P � 0.0001Translation 194 66 (26/40) 34.0 *P � 0.0001Tumor suppressors 48 9 (3/6) 18.8 0.08223Random set 222 33 (9/24) 14.9 0.05846
4793
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

pression of the E2F transcription factor family members, E2F-1,E2F-3, and E2F-5, as well as the E2F transcriptional coactivator DP-1(Fig. 1D). This family of transcription factors plays a central role inthe regulation of gene expression at the G1-S phase transition of thecell cycle by regulating the expression of genes of which the productsare required for nucleotide biosynthesis, DNA replication, and cellcycle progression (20). Consistent with their down-regulation, therewas a parallel decrease in a number of E2F target genes reportedpreviously including cyclin A, E, and D1, dihydrofolate reductase,DNA polymerase �, minichromosome maintenance deficient proteins2, 3, and 6, cdc 2 and 6, origin recognition complex 1, and c-myc (Ref.29; Figs. 1 and 2).
Finally, and consistent with the inhibition of cell proliferation,
Caco-2 cell maturation was accompanied by a concomitant down-regulation of several M phase phosphoproteins and proto-oncogenes,including K-ras, tpr, c-fes, met (HGF receptor), c-myc, v-ets, andv-yes (Fig. 1, E–G).
Nucleic Acid Synthesis, DNA Replication and Repair, Chromo-some Segregation, and Chromatin Assembly. Consistent with thereduction in expression of cell cycle regulatory genes and the reduc-tion in cell proliferation, there was a concomitant reduction of genesinvolved in DNA synthesis, replication, and repair. Down-regulatedgenes with a role in nucleic acid biosynthesis included thymidylatesynthase, dihydrofolate reductase, UMP kinase, adenosine kinase,adenylosuccinate lyase, and phosphoribosyl PPI synthetase 2 (Fig.2A). Genes with a role in DNA replication included MCM2, MCM3,
Fig. 1. Expression of genes involved in cellcycle progression, during Caco-2 cell differentia-tion (A–G). A, cyclins; B, cell division cycle pro-teins (cdc’s)/cyclin-dependent kinases; C, inhibi-tors of cell cycle progression; D, E2F transcriptionfactor family members; E, M phase phosphopro-teins; F, miscellaneous cell cycle-associated genes;and G, proto-oncogenes. Changes in expression ofp21WAF1/Cip1, cyclin A, cyclin E, and c-myc wereconfirmed by Western blotting. H, reduction in cellproliferation during Caco-2 cell differentiation. Therate of cell proliferation at progressive stages ofcell differentiation (0–21 days postconfluence) wasdetermined by [3H]thymidine uptake; bars, �SD.
4794
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

MCM6, origin recognition complex subunits 1 and 3, topoisomerasesI, and II, DNA ligase 1, DNA polymerase � and �, PCNA, andreplication factor c (activator 1) 4 (Fig. 2B). Similarly, and consistentwith DNA repair being most active in rapidly proliferating cells, DNArepair genes including DNA polymerase epsilon, mutS homologue 3,BRCA1 and BRCA2 were maximally expressed in undifferentiated,proliferating Caco-2 cells (Fig. 2C). Furthermore, genes involved inspindle formation and chromosome segregation, including severaldyneins, microtubule-associated proteins (MAPs), tubulin, and kinesinfamily members, were down-regulated during the maturation program(Fig. 2D). Ran, Ran-GAP1, and several Ran binding proteins,which in addition to their role in nuclear transport, play a role inmicrotubule polymerization and spindle formation (30), were sim-ilarly down-regulated in expression (see Fig. 4E, protein trans-port). Finally, genes involved in chromatin assembly were progres-sively down-regulated including nucleosome assembly protein1-like 1, H4 histone family member G, histone H2A, and H2A
histone family member O (Fig. 2E). The down-regulation of PCNA,thymidylate synthase, and topoisomerase 1 were confirmed byWestern blot analysis (Fig. 2).
RNA Processing, Splicing, and Transport. Consistent with anoverall reduction in gene expression during the maturation program,genes with a role in RNA processing, including splicing, transport,and maturation of RNA, were coordinately down-regulated in expres-sion (Fig. 2; supplementary data). Of the 121 predefined genes in thiscategory, 35 (28.9%) were altered in expression by our criteria, withthe majority down-regulated in expression. This included severalsplicing factors, hnRNP family members, and RNA helicases, includ-ing BAT-1 and the DEAD/H box containing polypeptides 9, and 20(Fig. 4, A–D). Furthermore, poly (rC) binding proteins 1, and 2 (alsoknown as hnRNPE1 and E2), which are RNA binding proteins impli-cated in translational silencing, were up-regulated in expression (31).However, two members of the serine/arginine family of splicingfactors, SRp20 and SC35 were up-regulated during Caco-2 cell dif-
Fig. 2. Expression of genes involved in (A)nucleic acid metabolism, (B) DNA replication, (C)DNA repair, (D) chromosome segregation, and (E)chromatin assembly during Caco-2 cell differenti-ation. Expression patterns of PCNA, DNA topoi-somerase 1, and thymidylate synthase were con-firmed by Western blotting.
4795
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ferentiation, as were the DEAD/H box containing polypeptides 19 andhPrp18 (Fig. 2; supplementary data). These observations raise thepossibility that different splicing factors may regulate the post-tran-scriptional processing of different groups of mRNA transcripts, whichmay be differentially expressed during colon cell maturation.
Translation, Protein Processing, Trafficking, and Degradation.Caco-2 cell maturation was also associated with a significant enrichmentof genes involved in protein translation (Table 1). Paradoxically, how-ever, this was caused by both increased and decreased expression ofgenes involved in protein translation (Fig. 3, A–C). First, with the excep-tion of 3 sequences, a number of genes involved in translation initiation
and elongation were down-regulated. Examples included subunits oftranslation initiation factors 1–5, and translation elongation factor 1,epsilon 1 (Fig. 3A). The down-regulation of eIF4E and eIF2� wasconfirmed by Western blotting. Furthermore, the Pkr gene, the product ofwhich catalyzes the phosphorylation of the � subunit of eIF2 leading toan inhibition of protein synthesis initiation (32), was also down-regulated(Fig. 3A). Finally, genes encoding 9 of the 20 tRNA synthetases, andexportin, which mediates the nuclear export of all of the tRNAs, werecoordinately down-regulated during the maturation program (Fig. 3B).Consistent with these changes, the overall rate of protein synthesis wasreduced by 80% in fully differentiated cells (Fig. 3D).
Fig. 3. Protein translation. Expression of genesinvolved in (A) translation initiation and elonga-tion, (B) tRNA synthetases, and (C) ribosomal pro-teins during Caco-2 cell differentiation. The ex-pression pattern of eIF4E and eIF2� was confirmedby Western blotting. D, concordant down-regula-tion in the rate of protein synthesis during Caco-2cell differentiation, as determined by [14C]leucineuptake experiments; bars, �SD.
4796
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

We also observed reduced expression of RNA polymerase I (Mr
16,000 subunit) and upstream binding transcription factor (see Fig.7C, transcription factors). These enzymes play a key role in ribosomalRNA expression, and their down-regulation is suggestive of a down-regulation of rRNA gene expression (33). However, in apparentcontrast we observed a general, although modest, up-regulation ofribosomal protein genes, a trend that was consistent for 20 familymembers (Fig. 3C). Whether this reflects the continued need fortranslation in differentiated cells, a compensatory response to thedown-regulation of other components of the translational machinery,or simply a discordance in the model system, remains to be deter-mined. Nevertheless, the general up-regulation of genes encodingribosomal proteins is consistent with their coordinate regulation (34).
After translation, nascent polypeptides that exit the ribosome mustbe folded into their correct tertiary structure, a task performed by anumber of proteins collectively termed chaperones (35). In accord-ance with the down-regulation of protein synthesis there was a con-comitant down-regulation of expression of several such genes, includ-ing members of the major chaperone classes, hsp60, hsp70, and hsp90(Fig. 4, A and B). Similarly, several cyclophilins and FK506-bindingprotein 5, involved in accelerating protein folding (36), were alsodown-regulated (Fig. 4C). Interestingly, we observed an increase inexpression of the heat shock transcription factor HSF4 (Fig. 7I),which has been shown to be a negative regulator of heat shock gene
expression (37), suggesting a possible mechanism for the down-regulation of heat shock gene expression.
Upon synthesis and folding, a large percentage of proteins must betranslocated to specific cellular organelles, particularly the nucleus.Transport of proteins �Mr 40,000 into and out of the nucleus occursvia the nuclear pore complex. To do so, proteins containing a nuclearlocalization signal first attach to soluble carriers of the importin-�/karyopherin-� family either directly or via an adapter such as importin� (38). Translocation into the nucleus is an energy-dependent processand requires the cotranslocation of Ran, a Ras family GTPase (38). Anumber of genes involved in the regulation of these processes weredown-regulated, suggesting a reduction in nuclear protein traffickingduring Caco-2 cell differentiation. These included four members ofthe importin-� (2, 3, 4, and 6) and four members of the importin-�family (1, 2, 2b, and 3) of protein carriers (Fig. 4D). Furthermore,Ran, RanGAP1, and 5 Ran binding proteins (1, 2, 2-like-1, 7, and 16)were also concomitantly down-regulated in expression (Fig. 4E).Finally, a number of components of the nuclear pore complex itselfwere down-regulated, including the nucleoporins 88, 153, 155, and214, and the proto-oncogene tpr (Fig. 4F). There was also decreasedexpression of genes involved in protein translocation to other or-ganelles. Examples included, adaptor-related protein complex � 2subunit, and metaxin-1, active in protein trafficking to lysosomes andmitochondria, respectively (Fig. 4G). Overall, of the 216 predefined
Fig. 4. Expression of genes involved in protein processing, transport, and degradation during Caco-2 cell differentiation. A–C, genes with a role in protein folding. A, chaperonins;B, heat shock proteins; and C, cyclophilins. D–G, genes involved in protein transport. D, karyopherins; E, Ran family members; F, components of the nuclear pore complex; and G,genes involved in protein transport to specific organelles. H–K, genes involved in protein degradation. H, ubiquitin-activating enzyme (E-1); I, ubiquitin-conjugating enzymes; J,components of the ubiquitin-ligase complex; K, UBPs; and L, components of the proteasome 26S subunit.
4797
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

genes in this functional class, 54 (25%) were altered in expression(P � 0.0001).
Several components of the ubiquitin-26S proteasome protein deg-radation pathway were also down-regulated (Fig. 4, H–L). This path-way involves the conjugation of ubiquitin to a protein substrate, aprocess executed by a series of well-defined enzymatic reactions,involving ubiquitin-activating enzyme (E-1), ubiquitin-conjugatingenzymes, and ubiquitin ligase complexes (39). Caco-2 cell differen-tiation was characterized by the down-regulation of E-1 enzyme, threeubiquitin-conjugating enzymes (E2G 1, E2L 3, and E2M), and severalmembers of ubiquitin ligase complexes including cullins 1, 3, 4A, andthe 1B F-box protein (Fig. 4, H–J, respectively). There was also adown-regulation of five UBPs, UBP 4, 8, 10, 12, and 14 (Fig. 4K),which help regulate the ubiquitin-26S proteolytic pathway by gener-ating free ubiquitin monomers from their initial translational products,by recycling ubiquitins during the breakdown of ubiquitin-proteinconjugates, and/or by removing ubiquitin from specific targets (40).
Ubiquitinated proteins are recognized by the 26S proteasome andtargeted for destruction. The 26S proteasome comprises two subunits,the 20S proteolytic core and the 19S regulatory subunit. The 20S coreis composed of 7 different � and 7 different � subunits arranged as acylindrical �7�7�7�7 complex in four stacked rings (41, 42). Eight ofthese 14 subunits were present on the arrays, but none were altered inexpression during the maturation program (data not shown). In con-trast, several components of the 19S regulatory subunit were down-
regulated in expression. The 19S regulatory subunit, which deliversprotein substrates to the 20S subunit in an ATP-dependent manner,comprises at least 18 different proteins (42). Of these, 3 ATPases (1,2, and 3) and 3 non-ATPases (5, 11, and 12) were down-regulatedduring the maturation program (Fig. 4L). Furthermore, proteasomeactivator subunit 3 (PA28, �, and Ki), an 11S regulator capable ofactivating the proteolytic activity of the 20S proteasome (43), was alsodown-regulated in expression (Fig. 4L).
Collectively, of the 158 predefined genes in this functional cate-gory, 37 (23.4%) were altered in expression during Caco-2 celldifferentiation, a significant enrichment relative to the overall level ofchange (P � 0.0001). The down-regulation of this pathway is medi-ated both at the level of protein ubiquitination and also at the protea-somal level, particularly through down-regulation of components ofthe 19S subunit.
Xenobiotic and Drug Detoxification. Among the most highlyinduced genes during Caco-2 cell differentiation, and consistent withprevious reports (44), were four members of the glutathione S-trans-ferase family, GST A1-1, A3, A4, and Ha subunit 2 (Fig. 5A). Gluta-thione S-transferases are Phase II conjugating enzymes that play animportant role in xenobiotic efflux and in the prevention of oxidativestress (45). Similarly, and consistent with the presence of commonregulatory elements in the promoter regions of genes encoding PhaseII enzymes (46), members of two other families of Phase II conju-
Fig. 5. Expression of genes involved in xenobiotic/drug metabolism during Caco-2 cell differentiation. A, glutathione S-transferases; B, sulfotransferases; C, UDP glycosyltrans-ferases; D, metallothioneins; E, avidin-biotin complex method subfamily members; F, cytochrome P450 members; G, aldehyde dehydrogenases; and H, miscellaneous enzymes involvedin xenobiotic detoxification. I, effect of camptothecin and cisplatin on apoptosis, at progressive stages of Caco-2 cell differentiation. Caco-2 cell cultured to confluence (day 0), or for2, 5, 7, 14, or 21 days postconfluence were treated with camptothecin or cisplatin for 72 h, and apoptosis measured by propidium iodide staining and FACS analysis. J, effect ofcamptothecin (cpt) and cisplatin (cisp) on clonogenicity of undifferentiated and differentiated Caco-2 cells; bars, �SD.
4798
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
gating enzymes, the sulfotransferases and UDP glycosytransferases,were also up-regulated (Fig. 5, B and C).
Expression of several metallothioneins (which are involved in cellprotection against toxic metals and reactive oxygen species; Ref. 47),ATP binding cassette sub-family C (CFTR/MRP), cytochrome p450,and aldehyde dehydrogenase family members were also up-regulatedduring the maturation program (Fig. 5, C–G). Furthermore, epoxidehydrolase, which catalyzes the hydration of reactive expoxide speciesthat arise from the metabolism of endogenous as well as xenobioticcompounds (48), was also up-regulated (Fig. 5H).
Colonic cell exposure to xenobiotic agents is likely to increase ascells migrate along the crypt axis to the lumenal surface. Therefore,the increased expression of these genes during differentiation mayreflect the activation of defense mechanisms in colonic epithelial cellsas they mature in relation to their altered position in the crypt.Increased expression of many of these genes has also been linked toincreased resistance of tumor cells to chemotherapeutic agents. Forexample, the active metabolite of camptothecin, SN-38, is detoxifiedthrough glucuronidation (49), whereas platinum compounds such ascisplatin are eliminated through glutathione conjugation (50), two setsof enzymes up-regulated during Caco-2 cell maturation. To testwhether the increase in expression of these genes (glycosyltrans-ferases and glutathione S-transferases) is linked to reduced sensitivityof differentiated Caco-2 cells to these agents, we treated Caco-2 cellsat various stages of differentiation with camptothecin and cisplatin. Asshown in Fig. 5I and consistent with the increased expression of genesinvolved in drug detoxification, the ability of camptothecin and cis-
platin to induce apoptosis decreased significantly as Caco-2 cellsunderwent differentiation. However, it remained possible that thisreduced sensitivity was a consequence of the reduced rate of cellproliferation during Caco-2 cell differentiation. Therefore, we per-formed an additional, clonogenic assay in which cells were treatedwith camptothecin or cisplatin for 9 h, after which the drug wasremoved, and cells were trypsinized, reseeded, and allowed to formcolonies. Differentiated Caco-2 cells had a substantially greater abilityto form colonies after drug treatment compared with their undiffer-entiated counterparts. Treatment of undifferentiated Caco-2 cells with1 �M camptothecin or 10 �M cisplatin resulted in 20% and 0% colonyformation, respectively, in comparison to untreated controls. In com-parison, identical treatment of differentiated Caco-2 cells resulted in69% and 33% colony formation after camptothecin and cisplatintreatment, respectively (Fig. 5J).
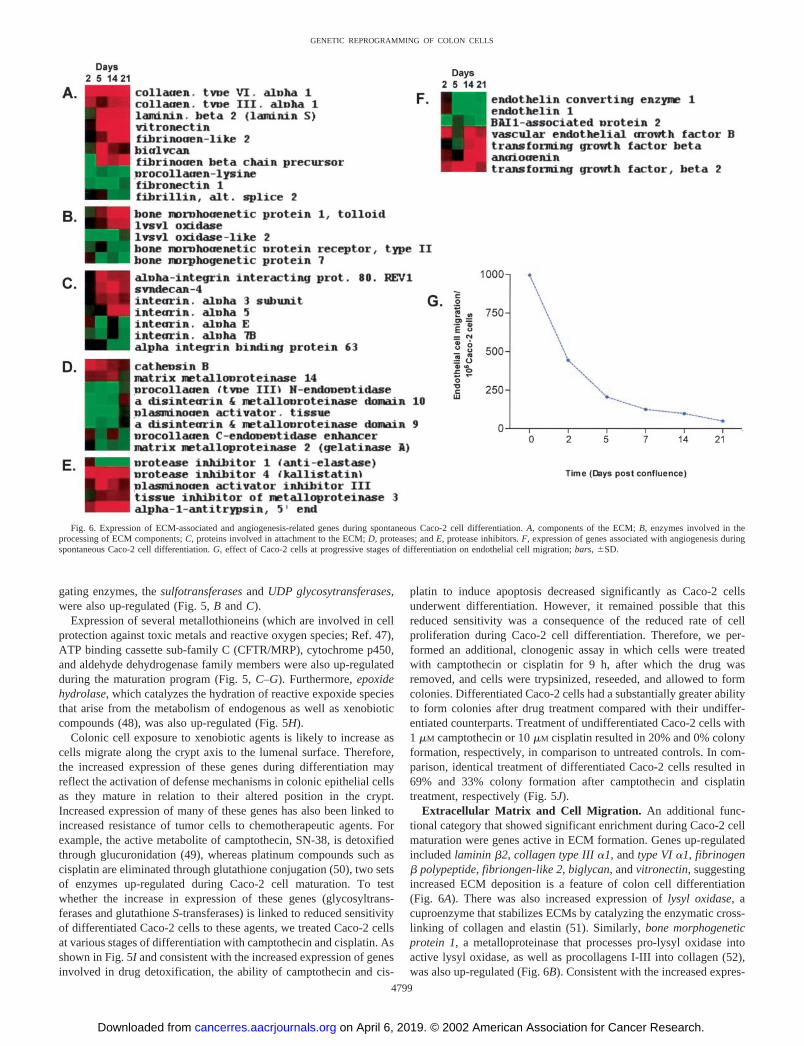
Extracellular Matrix and Cell Migration. An additional func-tional category that showed significant enrichment during Caco-2 cellmaturation were genes active in ECM formation. Genes up-regulatedincluded laminin �2, collagen type III �1, and type VI �1, fibrinogen� polypeptide, fibriongen-like 2, biglycan, and vitronectin, suggestingincreased ECM deposition is a feature of colon cell differentiation(Fig. 6A). There was also increased expression of lysyl oxidase, acuproenzyme that stabilizes ECMs by catalyzing the enzymatic cross-linking of collagen and elastin (51). Similarly, bone morphogeneticprotein 1, a metalloproteinase that processes pro-lysyl oxidase intoactive lysyl oxidase, as well as procollagens I-III into collagen (52),was also up-regulated (Fig. 6B). Consistent with the increased expres-
Fig. 6. Expression of ECM-associated and angiogenesis-related genes during spontaneous Caco-2 cell differentiation. A, components of the ECM; B, enzymes involved in theprocessing of ECM components; C, proteins involved in attachment to the ECM; D, proteases; and E, protease inhibitors. F, expression of genes associated with angiogenesis duringspontaneous Caco-2 cell differentiation. G, effect of Caco-2 cells at progressive stages of differentiation on endothelial cell migration; bars, �SD.
4799
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

sion of extracellular matrix genes, the integrin �3 subunit, whichbinds laminin and collagen (as well as fibronectin), integrin �5, whichbinds fibrinogen, and syndecan-4, which acts as a coreceptor in theadhesion of integrins to a range of ECM components (53), were allup-regulated during Caco-2 cell differentiation (Fig. 6C). In contrast,fibronectin was down-regulated during the maturation program (Fig.6A), an observation consistent with a previous report (54). Therefore,whereas these results indicate increased secretion of ECM compo-nents may be a feature of colon cell differentiation, they also suggestthat proliferating and differentiated cells secrete different combina-tions of ECM components.
Also included in this functional category were genes that have beenimplicated in cell migration. First, a number of proteases were alteredin expression. Those down-regulated included MMP-2 and tPA,whereas MMP-14 and cathepsin B were up-regulated in expression.On the other hand, several protease inhibitors were up-regulated,including TIMP3, PAI III, �-1-antitrypsin, and kallistatin (Fig. 6, Dand E). We have demonstrated previously that the basal rate of cellmigration decreases �2-fold during Caco-2 cell differentiation (21).The general increase in expression of genes associated with ECMdeposition and protease inhibitors are consistent with this reduced rateof cell migration.
The ECM also plays a role in regulating the rate of migration ofother cell types. For example, in the context of a growing tumor, theECM can act as a reservoir for the storage of proangiogenic factors,such as TGF-� and basic fibroblast growth factor, secreted by tumorcells (55). There was increased expression of several such factorsduring Caco-2 cell differentiation. Proangiogenic factors up-regulatedincluded VEGF-B (but not VEGF or VEGF-C), TGF-�, TGF-�2, andangiogenin. We also observed a decrease in BAI-associated protein 1,a gene of which the product interacts with the angiogenesis inhibitorBAI-1 (56). On the other hand, the proangiogenic factor endothelin 1,and endothelin 1 converting enzyme were down-regulated in expres-sion (Fig. 6F).
These observations prompted us to ask whether the ability of tumorcells to induce angiogenesis was dependent on their differentiationstatus. To determine this, we used an experimental coculture system totest the ability of Caco-2 cells at progressive stages of differentiationto induce endothelial cell migration, an initial step in the angiogenicprocess. Endothelial cell migration was maximal when coculturedwith undifferentiated Caco-2 cells, an effect that decreased signifi-cantly as the cells underwent differentiation (Fig. 6G). As expected,therefore, these observations suggest that undifferentiated, proliferat-ing tumor cells have a greater ability to induce angiogenesis than theirdifferentiated counterparts. Therefore, despite the differentiation-associated increase in expression of several proangiogenic genes, thisis outweighed by the reduced expression of others or by the increasedexpression of inhibitors of chemotaxis, resulting in an overall reduc-tion in endothelial cell migration.
Metabolism. In addition to the absorption of water and ions,factors that have escaped absorption in the small intestine, such as bileacids and cholesterol, can be taken up by the colonic epithelium (57).Furthermore, colonic epithelial cells also readily absorb and metabo-lize short-chain fatty acids produced from the bacterial fermentationof dietary carbohydrate (58). Consistent with the likelihood thatexposure of colonic cells to these substrates is likely to vary accordingto location along the crypt axis, several genes involved in lipid uptakeand metabolism were altered in expression during Caco-2 cell matu-ration, including muscle fatty acid binding protein, lipase A, hepaticlipase, acyl-CoA dehydrogenase (very long chain), and C-2 to C-3short chain acyl-CoA dehydrogenase (Fig. 3; supplementary data).
The increased expression of genes involved in lipid catabolismduring colon cell differentiation is consistent with several previous
observations. First, levels of fatty acid binding protein, which isinvolved in fatty acid uptake, increases in vivo as enterocytes migratetoward the lumen along the crypt axis (59). Similarly, alkaline phos-phatase, a brush border protein consistently up-regulated during coloncell differentiation (21), is thought to play a role in fat absorption (60).Second, PPAR�, a member of a family of nuclear hormone receptorsinvolved in the regulation of the catabolism and storage of fatty acids,is expressed in an incremental manner up the crypt axis and duringCaco-2 cell differentiation (61, 62). Finally, increased lipid uptake andmetabolism has been linked to differentiation status of colonic epi-thelial cells in vitro (21, 63).
Dietary fats absorbed by enterocytes are assembled into chylomi-crons and secreted into the mesenteric lymphatics. Consistent with theincreased expression of genes involved in lipid metabolism and up-take, expression of a number of apolipoproteins, a key component ofchylomicrons, was concomitantly up-regulated in expression. Amongthose were apolipoproteins A-1, B, C-III, and H, which are consistentwith previous reports (Refs. 64, 65; See Fig. 3; supplementary data).Collectively, therefore, these observations suggest that increased lipiduptake, metabolism, and packaging are characteristics of differenti-ated colonic epithelial cells.
In contrast, and possibly reflecting the reduced need for newmembrane synthesis after the cessation of cell division, there was ageneral down-regulation of genes with a role in lipid and cholesterolsynthesis, including acetyl CoA carboxylase �, ATP citrate lyase, fattyacid synthase, fatty acid CoA ligase, pyruvate carboxylase, meval-onate decarboxylase, mevalonate kinase, and sterol isomerase, duringthe maturation program (See Fig. 3; supplementary data).
Mechanisms. To gain additional insight into the mechanisms thatunderlie the reprogramming events described, we examined the intra-cellular signaling and transcription factor functional classes. Thesignal transduction class showed only a slight enrichment in thepercentage of genes altered in expression relative to the overallchanges. This may be because in many cases, the activity of signalingmolecules is regulated by post-translational modification events, par-ticularly phosphorylation. Therefore, signaling molecules not alteredin expression at the mRNA level may still play critical roles inmediating the differentiated phenotype. However, one pathway thatwas regulated at the mRNA level was the MAPK signaling pathwaywith a number of components, including Ras, MAPK6, MAP kinasekinase kinase 2, 8, and 12, MAPK8-IP1, and JAK kinase 1 down-regulated during the maturation program (Fig. 7A).
On the other hand, and consistent with the extensive genetic repro-gramming that characterizes Caco-2 cell maturation, a number oftranscription factors were both up and down-regulated. Among thosedown-regulated were members of the high mobility group family oftranscription factors (HMG-1, 2, and 14; Fig. 7B). HMG1 and 2 haveno known specific DNA recognition sequence but are able to inducebends in DNA, suggesting they may function as architectural factorsin processes that require transient alteration of DNA structure such asDNA repair, recombination, replication, and transcription (66). There-fore, their down-regulation is consistent with the concomitant down-regulation of many of these processes during Caco-2 cell maturation.Similarly, a number of genes involved in alteration of chromatinstructure, including several histone acetyltransferases and histonedeacetylases, were down-regulated (Fig. 7C). A role for these proteinsin regulating colon cell maturation is consistent with previous obser-vations that inhibitors of histone deacetylase activity, such as sodiumbutyrate, are potent inducers of colon cell maturation (67).
Several components of the RNA polymerase I, I, and III transcrip-tional complexes were also down-regulated, an observation consistentwith the overall reduction in transcription during Caco-2 cell matu-ration (Fig. 7, D–F). Finally, several basic helix-loop-helix proteins
4800
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

(HTF4A, ITF-2, and E2A; Fig. 7G) and a number of zinc fingerproteins were down-regulated during the maturation program.
Among the transcription factors up-regulated were two members ofthe retinoid receptor family of transcription factors, RXR� and � (Fig.7H). RXRs heterodimerize with activated PPARs and alter the tran-scription of numerous target genes, including several involved in lipidmetabolism and transport. One such target, Apo A-1, was shown to behighly up-regulated in the present study (See Fig. 3; supplementarydata). Previous studies have shown that PPAR� is up-regulated duringCaco-2 cell differentiation and as colonic epithelial cells migrateupwards along the crypt axis (61, 62). Furthermore, overexpression ofPPAR� stimulates markers of cell differentiation in colon cancer celllines, and loss of function mutations in the PPAR� gene is associatedwith colon cancer (68, 69), although studies to the contrary have alsobeen reported (70). LXR� was an additional nuclear receptor up-regulated during Caco-2 cell differentiation. LXRs heterodimerizewith RXRs and regulate the expression of several genes involved incholesterol metabolism and efflux (71). Collectively, these changesmay be responsible for the alterations in expression of genes involvedin lipid and cholesterol metabolism, and transport, during Caco-2 celldifferentiation.
A number of other transcription factors up-regulated during Caco-2cell differentiation were identified (Fig. 7I), each of which may
potentially contribute to the maturation program. Of these, the in-crease in CCAAT/enhancer binding protein-� is particularly interest-ing, as, consistent with the changes in the present study, it has beenimplicated in the regulation of p21WAF-1/Cip-1, and genes involved inlipid uptake and metabolism through induction of PPAR� (72).Among the other transcription factors, the increase in kruppel-likefactor 4 and Id-2 are consistent with their increased expression indifferentiated enterocytes of the intestinal epithelium (8, 73, 74).
DISCUSSION
The study of normal colon cell maturation has been hindered by theinability to maintain normal colonic epithelial cells in culture (5).However, model systems such as the Caco-2 cell line have proven tobe a suitable and highly informative alternatives for such studies (13).On contact inhibition, Caco-2 cells spontaneously undergo cell cyclearrest, with cells accumulating in the G0/G1 phase of the cell cycle,followed by differentiation along the absorptive cell lineage (15). Inthe current study we used microarray analysis to profile the changesin gene expression that accompany these processes.
As expected, the alterations in gene expression during spontaneousCaco-2 cell differentiation were extensive. By our criteria, 13.2% ofall of the genes were altered in expression during the maturation
Fig. 7. Expression of genes involved in signaltransduction and regulation of gene transcriptionduring Caco-2 cell differentiation. A, componentsof the MAPK signaling pathway; B, high mobilitygroup proteins; C, chromatin modifying enzymes;D–F, components of general transcriptional ma-chinery: RNA polymerases I, II, and III, respec-tively; G, helix-loop-helix proteins; H, nuclear re-ceptors and (I) miscellaneous transcription factorsup-regulated during Caco-2 cell maturation. Thechanges HMG-1 and E2A expression were con-firmed by Western blotting; bars, �SD.
4801
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

program. The majority of these changes (70%) were reductions ingene expression, a finding consistent with cell differentiation reflect-ing increased cellular specialization.
The regulation of colon cell differentiation has been suggested to bemodulated by a number of stimuli, including contact inhibition, che-motactic gradients, interactions with the ECM, a variety of growthfactors, cytokines, and luminal contents (5). Whereas all of thesefactors may play significant roles, the extent of the genetic reprogram-ming observed in the present study suggests that contact inhibitionmay be the most fundamental of these, and that the ability to undergodifferentiation is essentially an inherent feature of colon cells, requir-ing only the proper trigger to initiate a preprogrammed cascade ofevents.
One likely trigger is the �-catenin-TCF signaling pathway. Wehave shown recently that this pathway is down-regulated duringCaco-2 cell differentiation as a consequence of both increased E-cadherin and reduced TCF-4 expression (16). Consistent with this, inthe present study we observed a concomitant down-regulation of twowell-characterized �-catenin-TCF signaling genes, cyclin D1 andc-myc. Our previous studies showed that down-regulation of thispathway resulted in increased promoter activities in two of the fourmarkers of cell differentiation tested (16), suggesting at least part ofthe differentiation response can be attributed to the down-regulationof this pathway. There is also considerable in vivo evidence implicat-ing this pathway in the regulation of intestinal cell maturation. First,down-regulation of �-catenin-TCF signaling through targeted inacti-vation of the TCF-4 gene results in reduced cell proliferation and thepremature onset of differentiation in the small intestine (7). Similarly,down-regulation of the pathway through forced expression of E-cadherin results in reduced cell proliferation along the crypt axis (75).
Organization of the microarray database into functional groupsenabled changes in biological processes to be more clearly visualized.This approach revealed that colon cell maturation is a highly orga-nized process. First, in parallel with the cessation of cell proliferation,genes involved in cell cycle progression, DNA synthesis, replication,and repair were down-regulated. Second, and reflecting the reducedsize of the transcriptome in differentiated cells, components of thebasal transcriptional apparatus, and genes involved in RNA splicingand transport, were down-regulated. Third, and consistent with thereduced rate of [14C]leucine uptake, genes involved in protein trans-lation, folding, and degradation were coordinately down-regulated.
Despite the general down-regulation of components of the transla-tional machinery, including translation initiation and elongation fac-tors, translation does continue in differentiated cells, with severalproteins expressed at high levels. One explanation may be the highdegree of variability in translational efficiency that exists amongdifferent mRNA species. Typically, mRNAs coding for proteins pos-itively involved in growth control (e.g., cyclin D1) are poorly trans-lated in resting cells, attributable in part to the presence of a highdegree of secondary structure in the 5� untranslated region in thecyclin D1 mRNA. The translation of such messages is particularlysensitive to the activity of the CAP-dependent unwinding machinery,components of which (e.g., eIF4E) were down-regulated duringCaco-2 cell differentiation (32). Therefore, it is possible that mRNAspecies that are up-regulated in expression and which continue to betranslated in differentiated cells may be less reliant on eIF4E com-plexes because of less highly ordered secondary structures.
Our analyses also revealed that colon cell differentiation was asso-ciated with increased expression of genes encoding ECM-associatedproteins, as well as proteins involved in xenobiotic detoxification,lipid metabolism, and transport. The increase in ECM-associatedgenes suggests increased deposition of an ECM may be a function ofdifferentiated colonic epithelial cells. Indeed, interaction of epithelial
cells with ECM components is known to modulate differentiation(76). For example, culture of Caco-2 cells on different matrix sub-strates, such as laminin or collagen IV, enhanced the expression ofmarkers of cell differentiation (77). The stimulation of cell differen-tiation may be brought about in several ways: integrin-mediatedadhesion of epithelial cells to ECM components can trigger signalingevents leading to alterations in cell growth and differentiation. TheECM can also serve as a reservoir for the sequestration of latent formsof growth factors such as TGF-�, which along with TGF-�2, wasup-regulated during Caco-2 cell differentiation. TGF-� is secreted asan inactive “latent” complex, which can be subsequently activated bya number of proteases contained within the ECM. On activation,TGF-� can induce a number of changes in epithelial cell growth anddifferentiation (78). Whether some of the pathways of Caco-2 cellmaturation are mediated by enhanced TGF-� signaling requires ad-ditional investigation.
The increased expression of xenobiotic and drug detoxificationgenes may reflect an important defense mechanism that colon cellshave developed to manage their increased exposure to lumenal com-pounds as they migrate up the crypt axis. However, in the context ofa tumor, increased expression of these genes may lead to increasedresistance to specific chemotherapeutic agents. Indeed, differentiatedCaco-2 cells showed a high degree of resistance to cisplatin andcamptothecin-induced apoptosis. Importantly, the ability of gene arrayanalysis to simultaneously examine the expression of large numbersof such genes illustrates the potential use of this methodology for theprediction of response of tumor cells to chemotherapeutic agents.
Finally, the analyses revealed a general up-regulation of genesinvolved in lipid metabolism and transport, likely reflecting a role fordifferentiated enterocytes in the absorption and metabolism of lume-nal lipids, and their packaging into chylomicrons for transport to theliver.
The analyses also revealed a number of pathways that may drivethese reprogramming events, including chromatin-modifying en-zymes, signaling pathways such as TGF-�, MAPK, and RXR, as wellas many other transcription factors. The challenge of future studieswill be to dissect the contribution of these individual pathways, andmore important, their interaction to the overall maturation program.
In conclusion, this study demonstrates that the genetic reprogram-ming that accompanies colon cell maturation occurs in a highlyorganized and coordinate manner. Most important, the complexity ofchange, even within some functional groups of genes, and along asingle differentiation lineage emphasizes that cell maturation reflectsan interplay and balance among many pathways.
ACKNOWLEDGMENTS
We thank Dr. Geoff Childs and Aldo Massimi for generation and scanningof the cDNA arrays, and Drs. Anna Velcich and Andrew Wilson for theirhelpful advice and critical review of the manuscript.
REFERENCES
1. Gordon, G. I., and Hermiston, M. L. Differentiation and self-renewal in the mousegastrointestinal epithelium. Curr. Opin. Cell Biol., 6: 795–803, 1994.
2. Wang, T. C., and Dockray, G. J. Lessons from genetically engineered animal models.I. Physiological studies with gastrin in transgenic mice. Am. J. Physiol., 277: G6–11,1999.
3. Hermiston, M. L., and Gordon, G. I. Inflammatory bowel disease and adenomas inmice expressing a dominant negative N-cadherin. Science (Wash. DC), 270: 1203–1207, 1995.
4. Kedinger, M., Lefebvre, O., Duluc, I., Freund, J. N., and Simon-Assmann, P. Cellularand molecular partners involved in gut morphogenesis and differentiation. Philos.Trans. R. Soc. Lond. B. Biol. Sci., 353: 847–856, 1998.
5. Burgess, A. W. Growth control mechanisms in normal and transformed intestinalcells. Philos. Trans. R. Soc. Lond. B. Biol. Sci., 353: 903–909, 1998.
4802
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

6. Yang, Q., Bermingham, N. A., Finegold, M. J., and Zoghbi, H. Y. Requirement ofMath1 for secretory cell lineage commitment in the mouse intestine. Science (Wash.DC), 294: 2155–2158, 2001.
7. Korinek, V., Barker, N., Moerer, P., van Donselaar, E., Huls, G., Peters, P. J., andClevers, H. Depletion of epithelial stem-cell compartments in the small intestine ofmice lacking Tcf-4. Nat. Genet., 19: 379–383, 1998.
8. Shie, J. L., Chen, Z. Y., O’Brien, M. J., Pestell, R. G., Lee, M. E., and Tseng, C. C.Role of gut-enriched Kruppel-like factor in colonic cell growth and differentiation.Am. J. Physiol. Gastrointest. Liver Physiol., 279: G806–G814, 2000.
9. Clatworthy, J. P., and Subramanian, V. Stem cells and the regulation of proliferation,differentiation and patterning in the intestinal epithelium: emerging insights fromgene expression patterns, transgenic and gene ablation studies. Mech. Dev., 101: 3–9,2001.
10. Karam, S. M. Lineage commitment and maturation of epithelial cells in the gut. Front.Biosci., 4: D286–298, 1999.
11. Ho, S. B. Cytoskeleton and other differentiation markers in the colon. J. Cell.Biochem., 16G: 119–128, 1992.
12. Schulze, A., and Downward, J. Navigating gene expression using microarrays–atechnology review. Nat. Cell Biol., 3: E190–E195, 2001.
13. Pinto, M., Robine-Leon, S., Appay, M. D., Kedinger, N., Triadou, N., Dussaulx, E.,Lacroix, P., Simon-Assman, K., Haffen, K., Fogh, J., and Zwiebaum, A. Enterocytelike differentiation and polarization of the human colon carcinoma cell line Caco-2 inculture. Biol. Cell., 47: 323–330, 1983.
14. Hara, A., Hibi, T., Yoshioka, M., Toda, K., Watanabe, N., Hayashi, A., Iwao, Y.,Saito, H., Watanabe, T., and Tsuchiya, M. Changes in proliferative activity andphenotypes in spontaneous differentiation of a colon cancer cell line. Jpn. J. CancerRes., 84: 625–632, 1993.
15. Mariadason, J. M., Rickard, K. L., Barkla, D. H., Augenlicht, L. H., and Gibson, P. R.Divergent phenotypic patterns and commitment to apoptosis of Caco-2 cells duringspontaneous and butyrate-induced differentiation. J. Cell. Physiol., 183: 347–354,2000.
16. Mariadason, J. M., Bordonaro, M., Aslam, F., Shi, L., Kuraguchi, M., Velcich, A.,and Augenlicht, L. H. Down-regulation of �-catenin TCF signaling is linked tocolonic epithelial cell differentiation. Cancer Res., 61: 3465–3471, 2001.
17. Mariadason, J. M., Corner, G. A., and Augenlicht, L. H. Genetic reprogramming inpathways of colonic cell maturation induced by short chain fatty acids: comparisonwith trichostatin A, sulindac, and curcumin and implications for chemoprevention ofcolon cancer. Cancer Res., 60: 4561–4572, 2000.
18. Cheung, V. G., Morley, M., Aguilar, F., Massimi, A., Kucherlapati, R., and Childs,G. Making and reading microarrays. Nat. Genet., 21: 15–19, 1999.
19. Eisen, M. B., Spellman, P. T., Brown, P. O., and Botstein, D. Cluster analysis anddisplay of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA, 95: 14863–14868, 1998.
20. Muller, H., Bracken, A. P., Vernell, R., Moroni, M. C., Christians, F., Grassilli, E.,Prosperini, E., Vigo, E., Oliner, J. D., and Helin, K. E2Fs regulate the expression ofgenes involved in differentiation, development, proliferation, and apoptosis. GenesDev., 15: 267–285, 2001.
21. Mariadason, J. M., Velcich, A., Wilson, A. J., Augenlicht, L. H., and Gibson, P. R.Resistance to butyrate-induced cell differentiation and apoptosis during spontaneousCaco-2 cell differentiation. Gastroenterology, 120: 889–899, 2001.
22. Ashton, A. W., Yokota, R., John, G., Zhao, S., Suadicani, S. O., Spray, D. C., andWare, J. A. Inhibition of endothelial cell migration, intercellular communication, andvascular tube formation by thromboxane A(2). J. Biol. Chem., 274: 35562–35570,1999.
23. Hastie, N. D., and Bishop, J. O. The expression of three abundance classes ofmessenger RNA in mouse tissues. Cell, 9: 761–774, 1976.
24. Augenlicht, L. H., Wahrman, M. Z., Halsey, H., Anderson, L., Taylor, J., and Lipkin,M. Expression of cloned sequences in biopsies of human colonic tissue and in coloniccarcinoma cells induced to differentiate in vitro. Cancer Res., 47: 6017–6021, 1987.
25. Vachon, P. H., Perreault, N., Magny, P., and Beaulieu, J. F. Uncoordinate, transientmosaic patterns of intestinal hydrolase expression in differentiating human entero-cytes. J. Cell. Physiol., 166: 198–207, 1996.
26. Niv, Y., Byrd, J. C., Ho, S. B., Dahiya, R., and Kim, Y. S. Mucin synthesis andsecretion in relation to spontaneous differentiation of colon cancer cells in vitro. Int.J. Cancer, 50: 147–152, 1992.
27. Yanagie, H., Sumimoto, H., Nonaka, Y., Matsuda, S., Hirose, I., Hanada, S.,Sugiyama, H., Mikamo, S., Takeda, Y., Yoshizaki, I., Nakazawa, K., Tani, K.,Yamamoto, T., Asano, S., Eriguchi, M., and Muto, T. Inhibition of human pancreaticcancer growth by the adenovirus-mediated introduction of a novel growth suppressinggene, tob, in vitro. Adv. Exp. Med. Biol., 451: 91–96, 1998.
28. Medema, R. H., Kops, G. J., Bos, J. L., and Burgering, B. M. AFX-like Forkheadtranscription factors mediate cell-cycle regulation by Ras and PKB through p27kip1.Nature (Lond.), 404: 782–787, 2000.
29. Leone, G., DeGregori, J., Yan, Z., Jakoi, L., Ishida, S., Williams, R. S., and Nevins,J. R. E2F3 activity is regulated during the cell cycle and is required for the inductionof S phase. Genes Dev., 12: 2120–2130, 1998.
30. Walczak, C. E. Ran hits the ground running. Nat. Cell Biol., 3: E69–70, 2001.31. Ostareck, D. H., Ostareck-Lederer, A., Wilm, M., Thiele, B. J., Mann, M., and
Hentze, M. W. mRNA silencing in erythroid differentiation: hnRNP K and hnRNP E1regulate 15-lipoxygenase translation from the 3� end. Cell, 89: 597–606, 1997.
32. Clemens, M. J., and Bommer, U. A. Translational control: the cancer connection. Int.J. Biochem. Cell Biol., 31: 1–23, 1999.
33. Paule, M. R., and White, R. J. Survey and summary: transcription by RNA poly-merases I and III. Nucleic Acids Res., 28: 1283–1298, 2000.
34. Li, B., Nierras, C. R., and Warner, J. R. Transcriptional elements involved in therepression of ribosomal protein synthesis. Mol. Cell. Biol., 19: 5393–5404, 1999.
35. Bukau, B., Deuerling, E., Pfund, C., and Craig, E. A. Getting newly synthesizedproteins into shape. Cell, 101: 119–122, 2000.
36. Schiene-Fischer, C., and Yu, C. Receptor accessory folding helper enzymes: thefunctional role of peptidyl prolyl cis/trans isomerases. FEBS Lett., 495: 1–6, 2001.
37. Nakai, A., Tanabe, M., Kawazoe, Y., Inazawa, J., Morimoto, R. I., and Nagata, K.HSF4, a new member of the human heat shock factor family which lacks propertiesof a transcriptional activator. Mol. Cell. Biol., 17: 469–481, 1997.
38. Stewart, M., Baker, R. P., Bayliss, R., Clayton, L., Grant, R. P., Littlewood, T., andMatsuura, Y. Molecular mechanism of translocation through nuclear pore complexesduring nuclear protein import. FEBS Lett., 498: 145–149, 2001.
39. DeSalle, L. M., and Pagano, M. Regulation of the G1 to S transition by the ubiquitinpathway. FEBS Lett., 490: 179–189, 2001.
40. Yan, N., Doelling, J. H., Falbel, T. G., Durski, A. M., and Vierstra, R. D. Theubiquitin-specific protease family from Arabidopsis. AtUBP1 and 2 are required forthe resistance to the amino acid analog canavanine. Plant Physiol., 124: 1828–1843,2000.
41. Hirsch, C., and Ploegh, H. L. Intracellular targeting of the proteasome. Trends CellBiol., 10: 268–272, 2000.
42. Voges, D., Zwickl, P., and Baumeister, W. The 26S proteasome: a molecular machinedesigned for controlled proteolysis. Annu. Rev. Biochem., 68: 1015–1068, 1999.
43. Ahn, J. Y., Tanahashi, N., Akiyama, K., Hisamatsu, H., Noda, C., Tanaka, K., Chung,C. H., Shibmara, N., Willy, P. J., Mott, J. D., and et al. Primary structures of twohomologous subunits of PA28, a �-interferon-inducible protein activator of the 20Sproteasome. FEBS Lett., 366: 37–42, 1995.
44. Vecchini, F., Pringault, E., Billiar, T. R., Geller, D. A., Hausel, P., and Felley-Bosco,E. Decreased activity of inducible nitric oxide synthase type 2 and modulation of theexpression of glutathione S-transferase �, bcl-2, and metallothioneins during thedifferentiation of CaCo-2 cells. Cell Growth Differ., 8: 261–268, 1997.
45. van Bladeren, P. J. Glutathione conjugation as a bioactivation reaction. Chem. Biol.Interact., 129: 61–76, 2000.
46. Kwak, M. K., Egner, P. A., Dolan, P. M., Ramos-Gomez, M., Groopman, J. D., Itoh,K., Yamamoto, M., and Kensler, T. W. Role of phase 2 enzyme induction inchemoprotection by dithiolethiones. Mutat. Res., 480–481: 305–315, 2001.
47. Davis, S. R., and Cousins, R. J. Metallothionein expression in animals: a physiolog-ical perspective on function. J. Nutr., 130: 1085–1088, 2000.
48. Fretland, A. J., and Omiecinski, C. J. Epoxide hydrolases: biochemistry and molec-ular biology. Chem. Biol. Interact., 129: 41–59, 2000.
49. Brangi, M., Litman, T., Ciotti, M., Nishiyama, K., Kohlhagen, G., Takimoto, C.,Robey, R., Pommier, Y., Fojo, T., and Bates, S. E. Camptothecin resistance: role ofthe ATP-binding cassette (ABC), mitoxantrone-resistance half-transporter (MXR),and potential for glucuronidation in MXR-expressing cells. Cancer Res., 59: 5938–5946, 1999.
50. Zhang, K., Chew, M., Yang, E. B., Wong, K. P., and Mack, P. Modulation of cisplatincytotoxicity and cisplatin-induced DNA cross-links in HepG2 cells by regulation ofglutathione-related mechanisms. Mol. Pharmacol., 59: 837–843, 2001.
51. Rucker, R. B., Kosonen, T., Clegg, M. S., Mitchell, A. E., Rucker, B. R., Uriu-Hare,J. Y., and Keen, C. L. Copper, lysyl oxidase, and extracellular matrix proteincross-linking. Am. J. Clin. Nutr., 67: 996S–1002S, 1998.
52. Uzel, M. I., Scott, I. C., Babakhanlou-Chase, H., Palamakumbura, A. H., Pappano,W. N., Hong, H. H., Greenspan, D. S., and Trackman, P. C. Multiple bone morpho-genetic protein 1-related mammalian metalloproteinases process pro-lysyl oxidase atthe correct physiological site and control lysyl oxidase activation in mouse embryofibroblast cultures. J. Biol. Chem., 276: 22537–22543, 2001.
53. Woods, A., and Couchman, J. R. Syndecan-4 and focal adhesion function. Curr. Opin.Cell Biol., 13: 578–583, 2001.
54. Levy, P., Loreal, O., Munier, A., Yamada, Y., Picard, J., Cherqui, G., Clement, B.,and Capeau, J. Enterocytic differentiation of the human Caco-2 cell line is correlatedwith down-regulation of fibronectin and laminin. FEBS Lett., 338: 272–276, 1994.
55. Rooney, P., Kumar, P., Ponting, J., and Kumar, S. The role of collagens andproteoglycans in tumor angiogenesis. In: R. Bicknell, C. E. Lewis, and N. Ferrera(eds.), Tumor Angiogenesis, pp. 141–153. Oxford: Oxford University Press, 1997.
56. Oda, K., Shiratsuchi, T., Nishimori, H., Inazawa, J., Yoshikawa, H., Taketani, Y.,Nakamura, Y., and Tokino, T. Identification of BAIAP2 (BAI-associated protein 2),a novel human homologue of hamster IRSp53, whose SH3 domain interacts with thecytoplasmic domain of BAI1. Cytogenet. Cell Genet., 84: 75–82, 1999.
57. Molina, M. T., Ruiz-Gutierrez, V., Vazquez, C. M., and Bolufer, J. Caecal andcolonic uptake of both linoleic acid and cholesterol in rats following intestinalresection. Lipids, 25: 594–597, 1990.
58. Roediger, W. E. W. Utilization of nutrients by isolated epithelial cells of the rat colon.Gastroenterology, 83: 424–429, 1982.
59. Cohn, S. M., Simon, T. C., Roth, K. A., Birkenmeier, E. H., and Gordon, J. I. Use oftransgenic mice to map cis-acting elements in the intestinal fatty acid binding proteingene (Fabpi) that control its cell lineage-specific and regional patterns of expressionalong the duodenal-colonic and crypt-villus axes of the gut epithelium. J. Cell Biol.,119: 27–44, 1992.
60. Yamagishi, F., Komoda, T., and Alpers, D. H. Secretion and distribution of ratintestinal surfactant-like particles after fat feeding. Am. J. Physiol., 266: G944–952,1994.
61. Lefebvre, M., Paulweber, B., Fajas, L., Woods, J., McCrary, C., Colombel, J. F.,Najib, J., Fruchart, J. C., Datz, C., Vidal, H., Desreumaux, P., and Auwerx, J.Peroxisome proliferator-activated receptor � is induced during differentiation of colonepithelium cells. J. Endocrinol., 162: 331–340, 1999.
62. Mansen, A., Guardiola-Diaz, H., Rafter, J., Branting, C., and Gustafsson, J. A.Expression of the peroxisome proliferator-activated receptor (PPAR) in the mousecolonic mucosa. Biochem. Biophys. Res. Commun., 222: 844–851, 1996.
4803
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

63. Thomson, A. B., Doring, K., Keelan, M., and Armstrong, G. Nutrient uptake intoundifferentiated and differentiated HT-29 cells in culture. Can. J. Physiol. Pharmacol.,75: 351–356, 1997.
64. Ricchi, P., Pignata, S., Di Popolo, A., Memoli, A., Apicella, A., Zarrilli, R., andAcquaviva, A. M. Effect of aspirin on cell proliferation and differentiation of colonadenocarcinoma Caco-2 cells. Int. J. Cancer, 73: 880–884, 1997.
65. Reisher, S. R., Hughes, T. E., Ordovas, J. M., Schaefer, E. J., and Feinstein, S. I.Increased expression of apolipoprotein genes accompanies differentiation in theintestinal cell line Caco-2. Proc. Natl. Acad. Sci. USA, 90: 5757–5761, 1993.
66. Grosschedl, R., Giese, K., and Pagel, J. HMG domain proteins: architectural elementsin the assembly of nucleoprotein structures. Trends Genet., 10: 94–100, 1994.
67. Kim, Y. S., Tsao, D., Siddiqui, B., Whitehead, J. S., Arntein, P., Bennett, J., andHicks, J. Effects of sodium butyrate and dimethylsulfoxide on biochemical propertiesof human colon cancer cells. Cancer (Phila.), 45: 1185–1192, 1980.
68. Sarraf, P., Mueller, E., Jones, D., King, F. J., DeAngelo, D. J., Partridge, J. B.,Holden, S. A., Chen, L. B., Singer, S., Fletcher, C., and Spiegelman, B. M. Differ-entiation and reversal of malignant changes in colon cancer through PPAR�. Nat.Med., 4: 1046–1052, 1998.
69. Sarraf, P., Mueller, E., Smith, W. M., Wright, H. M., Kum, J. B., Aaltonen, L. A., dela Chapelle, A., Spiegelman, B. M., and Eng, C. Loss-of-function mutations in PPAR� associated with human colon cancer. Mol. Cell, 3: 799–804, 1999.
70. Lefebvre, A. M., Chen, I., Desreumaux, P., Najib, J., Fruchart, J. C., Geboes, K.,Briggs, M., Heyman, R., and Auwerx, J. Activation of the peroxisome proliferator-
activated receptor � promotes the development of colon tumors in C57BL/6J-APCMin/� mice. Nat. Med., 4: 1053–1057, 1998.
71. Edwards, P. A., Kast, H. R., and Anisfeld, A. M. BAREing it all. The adoption of lxrand fxr and their roles in lipid homeostasis. J. Lipid Res., 43: 2–12, 2002.
72. Fajas, L., Fruchart, J. C., and Auwerx, J. Transcriptional control of adipogenesis.Curr. Opin. Cell Biol., 10: 165–173, 1998.
73. Chandrasekaran, C., and Gordon, J. I. Cell lineage-specific and differentiation-dependent patterns of CCAAT/enhancer binding protein � expression in the gutepithelium of normal and transgenic mice. Proc. Natl. Acad. Sci. USA, 90: 8871–8875, 1993.
74. Wice, B. M., and Gordon, J. I. Forced expression of Id-1 in the adult mouse smallintestinal epithelium is associated with development of adenomas. J. Biol. Chem.,273: 25310–25319, 1998.
75. Hermiston, M. L., Wong, M. H., and Gordon, J. I. Forced expression of E-cadherinin the mouse intestinal epithelium slows cell migration and provides evidence fornonautonomous regulation of cell fate in a self-renewing system. Genes Dev., 10:985–996, 1996.
76. Streuli, C. Extracellular matrix remodelling and cellular differentiation. Curr. Opin.Cell Biol., 11: 634–640, 1999.
77. Basson, M. D., Turowski, G., and Emenaker, N. J. Regulation of human (Caco-2)intestinal epithelial cell differentiation by extracellular matrix proteins. Exp. CellRes., 225: 301–305, 1996.
78. Derynck, R., Akhurst, R. J., and Balmain, A. TGF-� signaling in tumor suppressionand cancer progression. Nat. Genet., 29: 117–129, 2001.
4804
GENETIC REPROGRAMMING OF COLON CELLS
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

2002;62:4791-4804. Cancer Res John M. Mariadason, Diego Arango, Georgia A. Corner, et al.
in VitroA Gene Expression Profile That Defines Colon Cell Maturation
Updated version
http://cancerres.aacrjournals.org/content/62/16/4791
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2003/04/15/62.16.4791.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/62/16/4791.full#ref-list-1
This article cites 74 articles, 25 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/62/16/4791.full#related-urls
This article has been cited by 17 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/62/16/4791To request permission to re-use all or part of this article, use this link
on April 6, 2019. © 2002 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from