a dissertation - georgetown university
TRANSCRIPT

INTERACTION OF ATTENTION AND EMOTION ACROSS DEVELOPMENT AND DISORDER
A Dissertation submitted to the Faculty of the
Graduate School of Arts and Sciences of Georgetown University
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy in Neuroscience
By
Ericka D. Ruiz, M.A.
Washington, DC July 6, 2012

ii
INTERACTION OF ATTENTION AND EMOTION
ACROSS DEVELOPMENT AND DISORDER
Ericka D. Ruiz, M.A.
Thesis Advisor: Chandan J. Vaidya, Ph.D.
ABSTRACT
This dissertation examines how attention and emotion interact in early visual
processing across typical development and in Attention Deficit/Hyperactivity Disorder
(ADHD). To investigate early visual processing, we utilized the attentional blink (AB)
paradigm. The AB is the loss of awareness of a target immediately following detection
of a first target. However, the AB does not occur if the second target contains
emotional information. This modulation of the AB indicates that emotional
information has privileged access to early visual attention. Chapter 2 reports that
independent of the emotional expression, children showed a larger AB than adults for
human faces, suggesting reduced attentional resources in early visual processing.
There was, however, no developmental difference in the extent to which the AB was
reduced for angry faces, indicating that early visual attention to emotionally salient
information is mature in children. Further, typically developing children who were
more anxious, displayed a reduced AB for faces with neutral expression, indicating
heightened early visual attention to socially important information. Chapter 3 reports
that children with ADHD were similar to controls in the magnitude of the AB for
neutral faces and its reduction for angry faces. Thus, early visual attention to human

iii
faces and its modulation by emotional expression is intact in ADHD. Further, ADHD
children with more inattention symptoms showed a larger modulation of the AB by
emotional information. These results have implications for models of how attention
and emotion interact in early visual attentional processing in typical development and
ADHD.

iv
ACKNOWLEDGEMENTS
This dissertation is first dedicated to God, whom I owe everything to. He
opened the doors so that I could continue my education at Georgetown University,
allowed me to find favor and receive financial support to complete my doctoral
training, and has given me the strength, courage and perseverance to complete this
great endeavor.
It brings me great pleasure to acknowledge the generosity, dedication and
guidance of the many individuals who contributed to my thesis work. I cannot
overstate my gratitude to my mentor, Chandan Vaidya. I greatly admire and appreciate
her knowledge, patience, support, and kindness. Many special thanks to my committee
members, Guinevere Eden, Monique Ernst, Ludise Malkova, and Barbara Schwartz,
who supported me in so many ways through this process. I am also grateful to
Professor Merry Sleigh, for introducing me to research and teaching me to love
neuroscience.
I also thank the Interdisciplinary Program in Neuroscience faculty at
Georgetown, and in particular those who taught me more about this expanse ocean we
call neuroscience. This dissertation would not have been possible without the team of
students and research staff in the Developmental Cognitive Neuroscience Lab. Most
notably, I am thankful to those who worked closely with me in the last years, including
Jane DeWire, Bri Flores, Helen Burton, Kelly Barnes, Melanie Stollstorff, and Philip

v
Lee. I give a heartfelt thanks to all the Georgetown graduate and undergraduate
students with whom I shared this experience.
My very deepest thanks goes to my family, whose unconditional love and
endless support motivates my work and yet reminds me of all other things that are
important in life. Thank you to my best friend and love, my husband Christian Ruiz,
who has been my anchor and helped me to balance our family while continuing my
education. To my children, Franco, Esteban, Christian (Kiki) and Joaquin, for being
my motivation to complete this dissertation. To my proud parents, Erick and Angela,
who taught me that education is far greater than riches. To my sister, Clarissa, who has
helped me unconditionally. To my in-laws, Axires and Alicia, whose kindness and
love have infused me with strength. To my Aunt Rina, my cousin Ericka, and friend
Rosita, who selflessly cared for my children so that I could focus on my work. Last
but not least of all, thanks to my Pastor and friend, Dr. Nestor Alvarado, his wife
Ericka and daughters, Michelle and Elizabeth, who have always been there for all
kinds of support. I know there are others who have helped so that I may complete my
degreee, to those I say thank you and give witness that their prayers were not in vain.

vi
TABLE OF CONTENTS
CHAPTER I: INTRODUCTION............................................................................................. 1
CHAPTER II: DEVELOPMENTAL DIFFERENCES IN THE EMOTIONAL MODULATION OF
ATTENTIONAL FUNCTION
Abstract………………................................................................................................. 16
Introduction................................................................................................................... 17
Method…...................................................................................................................... 22
Results…....................................................................................................................... 29
Discussion..................................................................................................................... 35
CHAPTER III: EMOTIONAL MODULATION OF THE ATTENTIONAL FUNCTION IN
CHILDHOOD ATTENTIONAL DEFICIT/HYPERACTIVITY DISORDER
Abstract………………................................................................................................. 42
Introduction................................................................................................................... 43
Method…...................................................................................................................... 48
Results…....................................................................................................................... 54
Discussion..................................................................................................................... 60
CHAPTER IV: GENERAL DISCUSSION............................................................................. 66
REFERENCES.................................................................................................................. 75

vii
LIST OF FIGURES
Figure 1. Rapid serial visual presentation (RSVP) stream for the Attentional Blink
Task………………………………………………………………………….…....….. 28
Figure 2. Mean percentage correct scores as a function of group (adult vs. child),
emotional face of T2 (angry vs. neutral), and lag (2, 4, 8)........................................... 30
Figure 3. Mean percentage correct scores for emotional face of T2 (angry vs. neutral)
and group (adult vs. child)............................................................................................ 33
Figure 4. Correlation between STAI Trait Anxiety and the size of the attentional blink
in children……………................................................................................................. 35
Figure 5. Rapid serial visual presentation (RSVP) stream for the Attentional Blink
Task………………………………………………………………………….…....….. 53
Figure 6. Mean percentage correct scores as a function of group (Control vs. ADHD),
emotional face of T2 (angry vs. neutral), and lag (2, 4, 8)........................................... 55
Figure 7. Correlation between the AB Size with Neutral Faces and Trait Anxiety based
on STAI scores…………………………………………….......................................... 58
Figure 8. Correlation between the Influence of Emotion on Early Attentional
Processing and Inattention symptoms as measured by the ADHD Rating Scale......... 60

1
CHAPTER 1: INTRODUCTION
Emotional events capture our attention instantly. A mundane drive home from
work can become memorable when there is a car accident on the side of the road. A
stern face looking at us in a crowd immediately draws our attention to that person and
makes us more likely to remember that person more than anyone else in that crowd.
When emotional value is added to a stimulus, that stimulus becomes more significant.
The increased significance allows us to remember it more. Attention can be facilitated
by emotion such that emotionally charged stimuli can be consolidated into our memory
faster. The facilitation by emotion may arise from the evolutionary need to identify
important stimuli in the environment, whether it is negative, like a snake, or positive,
like a mother’s face. In this dissertation, I will investigate how emotion influences
attention at the perceptual level across typical development and in a common
attentional disorder of development, Attention Deficit Hyperactivity Disorder
(ADHD).
ATTENTION Attention is the process by which we reduce the flow of information into the
sensory system of the brain by enhancing the relevant or important components of the
input stream while eliminating the distracting ones (Taylor & Fragopanagos, 2005).
Attention can be voluntary and controlled by either top-down or bottom-up processes
(Corbetta & Shulman, 2002). Top down processes are more goal-directed and rely on
a dorsal brain network (Corbetta, Patel, & Shulman, 2008; Corbetta & Shulman, 2002).
Bottom up processes on the other hand are reflexive, sensory driven and rely on a

2
ventral brain network. The ventral network underlies emotional arousal and
motivational processes (Weissman & Prado, 2012). When we study for an exam we
engage our dorsal network to focus on only the material that is necessary for us to
know for the exam. The ventral network is engaged when we see bright neon shirt or
when we see someone crying in a crowd. The difference between these two networks
is the effort required, when we engage our dorsal network, it is more effortful-we must
direct our attention voluntarily, (endogenous). Engaging the ventral network is less
effortful; stimuli capture our attention even if we do not want to direct our attention to
it (exogenous). There has been an increasing interest among researchers to understand
the types of stimuli that capture attention but also how they do it as well.
Attention processing is believed to have two components, an early and late
stage of processing (Corbetta et al., 2008; Corbetta & Shulman, 2002). In the early
stage, information enters through sensory inputs and is selected without full perceptual
analysis. An example of this would be the cocktail party effect, where in a busy and
confusing environment we are alerted by our name being called out; however, we do
not know who or where our name was called from (Kuyper, 1972). In the late stage of
attention processing, information can only be selected on after full perceptual analysis
and recoding into a semantic or categorical representation. A gating mechanism is
believed to be present between early and late stage processing that determines what
information pursues later stage processing (D. Broadbent & Broadbent, 1990).

3
Attention processing is limited in its capacity to process information (Johnston,
1986; Posner, 1993). Information cannot be simultaneously processed if multiple
inputs are receiving large amounts of information. When this does happen, a
bottleneck is believed to occur while determining what is selected for extended
processing to gain awareness and what is not selected (D. Broadbent & Broadbent,
1990). In the presence of high priority information, a mechanism has been established
early on to overcome the limited capacity of attention processing (Anderson, 2005).
Information that is identified as high priority like emotional information, can be
selected or filtered rather easily so that it gain access and pass the bottleneck efficiently
(Pratto & John, 1991). The bottleneck is manifested in the attentional blink (AB)
phenomenon when one target is being processed and subsequent target is presented
immediately after the first target. There is a quick influx of information and the second
target cannot be detected. However if the second target is of high priority like one
containing emotional information, the AB effect can be reduced or even abolished.
Therefore, it is important to understand what makes emotional information gain a
priority status in attention processing.
Emotion
Emotion can be thought of as states that coordinate homeostasis in a complex
environment (Kuyper, 1972). The experience of emotions or the states of emotional
consciousness are what we commonly call “feelings” (Taylor & Fragopanagos, 2005).
When studying selective attention “emotion” can refer to the emotional quality of a

4
stimulus or emotional state of an individual (Yiend, 2010). In this dissertation,
emotion refers to the emotional quality of the stimulus.
Emotion has been densely studied and certain structures and circuits have
emerged as areas important in the processing of emotion. Both cortical and subcortical
structures have been implicated in processing emotions. Higher order sensory cortices
are involved in the perceptual representation of the stimuli and its features. Cortical
structures like the ventrolateral (VLPFC), orbitofrontal (OFC), and prefrontal cortices
are involved in modulating emotional responses in subcortical structures like the
amygdala and ventral striatum. The VLPFC cortex is a modulator of the emotional
responses in the amygdala. When there is reduced activation in the VLPFC it is
thought to reflect a reduced top-down control of the amygdala (Marsh, Gerber, &
Peterson, 2008). The OFC along with the amygdala and ventral striatum mediate the
association of the perceptual representation with an emotional response, cognitive
processing and behavioral motivation (Adolphs, 2003). The amygdala in particular is
suggested to be the encoder of emotional value before or even without the full object-
level perception of the stimuli that permits the conscious awareness of a stimuli
(Taylor & Fragopanagos, 2005). Therefore an extensive circuit is utilized in
processing emotion.
Faces have been used ubiquitously in researching emotion as experimental
stimuli for several reasons. Faces contain not only information about identity, gender,
or age but also convey information about emotion (Ekman & Oster, 1979; Patrik

5
Vuilleumier & Pourtois, 2007). A primary means of communication is done through
the emotional expressions on the face (Ekman & Oster, 1979; Somerville, Fani, &
McClure-Tone, 2011). Basic emotions like anger, fear, disgust, happiness, sadness and
surprise are easily identified across cultures and ages (Ekman & Friesen, 1971). Faces
can be controlled for visual characteristics and because of this there are several
normalized sets readily available to researchers (Somerville et al., 2011). Emotional
faces can be easily used across different age groups and clinical populations because
they do not require semantic processing and do not contain graphic images that can be
disturbing for younger research participants. The availability and ease of using faces
has produced a large literature that allows us to further understand emotion with
different techniques from behavioral to electrophysiological to neuroimaging.
The human brain has developed a way to process emotional faces because of
their evolutionary and social significance early in development. Animal research
shows that among primate species there is a relationship between the size of their
neocortex and their social group (Adolphs, 2003; Dunbar, 2009). The evolutionary
hypothesis is that the larger the social group the greater the need for social skills and
cognitive processes and thus greater brain development to subserve social cognition
(Dunbar, 2009; Whiten, 1997). Human developmental research shows the importance
of faces from infancy in the development of early preference for faces and face-like
stimuli (Batty & Taylor, 2006; Cohen Kadosh & Johnson, 2007). At birth, infants rely
on high contrast features to detect face-like shapes (Lamb & Sherrod, 1981). Infants at

6
3 months, cannot discriminate between emotional expressions, (Nelson, 1987) but will
change their behavior in response to different emotional expressions (Montague &
Walker-Andrews, 2001). By 4 months, infants are able to discriminate between
expressions (T. M. Field, Woodson, Greenberg, & Cohen, 1982) . Longer looking
times were seen in 7 month olds in response to fearful faces, (de Haan, Belsky, Reid,
Volein, & Johnson, 2004). Discrimination of emotional expression improves
dramatically throughout the first year of life, (Nelson, 1987; Somerville et al., 2011).
Emotional faces are evolutionarily and developmentally important stimuli in the
environment.
Facial expressions of emotion are so inherently salient that the human brain has
developed a way to process faces that is independent of basic object recognition.
Components of a face are initially processed individually and subsequently integrated
to perceive a face and its emotional expression (Haxby, Hoffman, & Gobbini, 2000).
Neuroimaging studies have outlined an extensive network that supports face
processing, comprised of visual object processing regions: inferior occipital gyrus,
superior temporal sulcus, anterior temporal pole and the fusiform face area, (LeDoux,
2000; Pessoa, 2002; Patrik Vuilleumier, 2005). Regions traditionally associated with
the ventral network of attention, like the amygdala, posterior cingulate and
orbitofrontal cortex, are also believed to process some aspects of faces (Blagrove &
Watson, 2010; Blasi et al., 2009; Hariri, Bookheimer, & Mazziotta, 2000; LeDoux,
2000). Electrophysiological studies have identified certain potentials in the parts of

7
these regions to be indicative of face processing (Darque, Del Zotto, Khateb, & Pegna,
2012; M. J. Taylor, Batty, & Itier, 2004). Event related potentials (ERPs) are
measured using electroencephalography (EEG) that measure the electrical activity over
time using electrodes placed on the scalp. ERPs have different waveforms that are
either positive or negative which are called components. In the early stages of face
processing, intracranial ERPs recorded from the cortical surface of the inferior
temporal and fusiform gyri have shown that a negative component occurs 200 ms after
the presentation of faces (N200). Another potential has been found with intracranial
depth recordings that is specific to faces, the P180, in the basal occiptotemporal cortex.
One more signal is the N170 that is recorded at the scalp from the occipito-temporal
cortex. The N170 in both adults and children is largest at parieto-temporal locations,
however the latencies decrease from childhood to adulthood. The decrease in latency
with increasing age suggests a faster and more efficient face processing mechanism. In
addition to the N170 there is a positive component that is involved in early face
processing, the “P1”. In children it is rather large and can be found earlier than the
N170. Like the N170, the P1 decreases with age. The amplitudes of these potentials
also differ between adulthood and childhood. The P1’s amplitude decreases with age
while the N170 decreases from early childhood (approximately 4 years of age) to mid
childhood and then increases again from the teenager to adulthood. Collectively all
these results suggest an extensive face processing network that matures over time to be
able to detect finer details, (Somerville et al., 2011; Taylor et al., 2004).

8
It is therefore important to understand how the emotional face processing
mechanism develops and how it might be affected in a developmental disorder.
Attention Deficit Hyperactivity Disorder
Attention Deficit/Hyperactivity Disorder, ADHD, is one of the most common
neuropsychiatric disorders in childhood, affecting 3-7% of school-aged children.
ADHD was first described as “hyperkinesis disorder of childhood,” then Attention
Deficit Disorder, (ADD) and as it currently identified in the DSM-IV, ADHD. ADHD
is characterized by symptoms of inattention, hyperactivity and impulsivity. Depending
on the ratio of symptoms, the DSM-IV recognizes three subtypes, a hyperactive-
impulsive (more hyperactive/impulsive symptoms), inattentive (more symptoms of
inattention), and combined (presence of both hyperactive/impulsive and inattention
symptoms) subtype. ADHD’s symptoms interfere with scholastic and social
functioning and significantly impact quality of life. For over 20 years, ADHD has
been conceptualized as a disorder of the prefrontal cortex and is still diagnosed on
behavioral reports from educators, parents, and clinicians. Children with ADHD are
known to have problems filtering information, selective visual attention, (the ability to
focus on relevant information while ignoring irrelevant distractors) (Mason,
Humphreys, & Kent, 2005) all related to a core cognitive deficit in a top down process,
executive control (Schachar, Tannock, Marriott, & Logan, 1995). Executive control is
ascribed to a nigrostriatal network, which utilizes a similar structure like the dorsal
network of attention, the lateral frontal cortex. Recently, researchers have questioned

9
the belief that ADHD is exclusively a result of the deficit in executive control.
Evaluation of early studies of selective visual attention, showed that children with
ADHD despite being distracted by irrelevant stimuli were no more vulnerable to
irrelevant information than controls (Castel, Lee, Humphreys, & Moore, 2011).
Children with ADHD have a stronger preference for small immediate rewards over
larger delayed rewards (E. J. S. Sonuga-Barke, 2002) and high-rate intrusive and
impulsive behavior (excessive talking, interruptions, obnoxious behavior), deficient
communication skills (including eye contact and motor regulation), biased and
deficient social cognitive skills (decreased self-awareness, deficient social problem-
solving skills, biased attributions of others’ intentions, inattentiveness to social cues),
and poor emotional regulation (Guevremont & Dumas, 1994; Hoza, 2007; Nixon,
2001; Soliva et al., 2009; Wheeler & Carlson, 1994). All these behaviors cannot be
explained exclusively from a deficit in executive control, therefore a deficit or atypical
process must be present in an area outside of executive control.
Current ADHD research has begun to consider investigating bottom-up
processes like those involved in the ventral network of attention as a potential site of a
deficit for children with ADHD. The mesolimbic network, comprised of the
orbitofrontal cortex, anterior cingulate, ventral striatum, and amygdala, are believed to
contribute to an emotional and motivation dysfunction in ADHD (E. J. S. Sonuga-
Barke, 2003). Of particular concern for this dissertation is the amygdala because of its

10
role both in the ventral network of attention and mesolimbic emotional dysfunction in
ADHD.
ADHD is mainly known by its executive control deficits but there is also
behavioral evidence for emotional dysfunction in ADHD. Boys with ADHD have
more difficulty on tasks that match emotions to situations even when impulsive
responding was controlled (Yuill & Lyon, 2007). This suggests emotional
dysregulation above and beyond executive dysfunction. Social functioning and
emotional regulation vary across ADHD subtypes, such that the combined subtype is
characterized by being less popular, more aggressive, and being more negative after
being disappointed (Maedgen & Carlson, 2000). A performance measure of
impulsivity (Stop Signal Task) accounted for only a small amount of variability in
emotional regulation when a child with ADHD performed a task that was designed to
make the child frustrated (Kühle et al., 2007). In addition, Stop-signal performance did
not change from before to after performing the frustration-inducing task even when the
children with ADHD were told to regulate their emotions explicitly. In a different
study, medication treatment with either methylphenidate or atomoxetine (whose action
influences both the top down and bottom up networks) improved positive emotion
expression on a parent-rated Expression and Emotion Scale for Children (Kratochvil et
al., 2007). Together these studies provide evidence that emotional dysfunction in
ADHD is not accounted for by executive control mediated by dorsal top down

11
processes. Direct investigation of emotional processing in ADHD is necessary to
further provide a comprehensive view of ADHD pathophysiology.
Studies assessing the bottom-up processes directly in ADHD have mainly
utilized rewards. Tangible rewards like money, tokens, or food incentives (Konrad,
Gauggel, Manz, & Schöll, 2000; Luman, Oosterlaan, & Sergeant, 2005), have been
used to improve ADHD’s executive control deficits. However recently there is an
emerging literature indicating the increased salience of socio-emotional information
and its effect on executive control tasks. One group directly compared the effect of
non-social (money) and social (smiling faces) rewards in typically developing children
and adolescents on a response inhibition task. Interestingly they found that both types
of reward increased performance on the response inhibition task and the degree of
influence of the social reward depended on how empathetic a child or adolescent is
(Kohls, Peltzer, Herpertz-Dahlmann, & Konrad, 2009). This provides one of the first
pieces of evidence that socio-emotional information is salient and can enhance
performance in children. Among ADHD children, when a social motivation condition
(children were made to believe they were competing with other children) was
introduced in an interference suppression task (a child adapted flanker), children with
ADHD were able to perform equally well as controls (Geurts, Luman, & van Meel,
2008). Lastly when both monetary and social (positive faces) rewards were directly
compared in children with ADHD, social rewards were found to be more salient and
produced greater improvements than monetary rewards on a response inhibition task

12
(Kohls, Herpertz-Dahlmann, & Konrad, 2009). Socio-emotional information is just as
salient as rewards that engage bottom up processes that recruit the mesolimbic network
in ADHD. By using socio-emotional stimuli we also have insight into how emotional
information is processed by children with ADHD. It is therefore, necessary to study
early emotional processing in ADHD to understand whether the emotional dysfunction
is a by-product of increased bottom up processes that overload the top-down process of
executive control of children with ADHD or if bottom up processes are intact and the
dysfunction present is the result of a weak executive control.
Attentional Blink Paradigm
When the visual system is presented with a large amount of information, it is
only capable of perceiving and consciously remembering a small selection. Think of
the last time you went to a wedding, you can remember parts of the party, the dress the
bride wore, the cake, the tossing of the garter and bouquet. But can you remember:
whether the groom wore a bow tie or tie, whether the flower girl had a colored sash,
whether the bouquet had colored ribbon or not. The selection of the information
involves attention, and if allocation of attentional resources is insufficient,
“inattentional blindness” can occur. If it was a large elaborate wedding you may not
have paid enough attention to the little details because there were too many. A task
that elucidates this phenomenon is the rapid serial visual presentation paradigm
(RSVP), also known as the attentional blink paradigm (Raymond, Shapiro, & Arnell,
1992). In this task, stimuli are presented quickly at about a rate of 10 stimuli per

13
second (100ms). In the RSVP stream, there are two targets, T1 and T2 that are
presented among distractors, with T2 appearing always after T1 at different time lags.
There is a reduced number of T2 targets that are consciously reported when they occur
between 200 and 500ms after T1, resulting in the attentional blink. The attentional
blink demonstrates the temporal limitation of the attentional resources readily available
for information to reach consciousness.
The attentional blink is believed to reflect the transient impairment of post
perceptual attentional mechanisms active at a late stage (after the identification of the
stimulus) at the top-down process level. Attentional resources are depleted while T1 is
being processed. However if there is a salient T2, attentional capture occurs via the
reflexive allocation of attention toward the salient T2 reallocating attentional resources
so that it is processed (Anderson, 2005). Bottom-up processes like that of the ventral
network of attention are believed to be responsible for this enhanced early attentional
processing. Emotional T2s are an example of salient T2s that can be perceived at
early lag intervals, because of the allocation of attentional resources available that
reduce the attentional blink effect. The amygdala has been implied for the preferential
processing of emotion with the attentional blink paradigm (Anderson & Phelps, 2001),
when the amygdala has been compromised due to brain insult, there is no allocation of
attentional resources so that T2 can be fully processed. The attentional blink paradigm
through the use of emotional stimuli can provide insight into how attentional capture
occurs across development and ADHD.

14
Faces can convey various emotions making them even more efficient in
capturing visuo-spatial attention, to the extent that early stages of face processing are
considered to be automatic (Pessoa, 2002; Patrik Vuilleumier, 2005). Because of the
influence that emotional faces exert in early visual processing, it is believed that they
have a preferred status in the perceptual processing stream. Relative to other visual
objects, faces elicit an early enhanced negative component in the lateral occipital scalp
at about 170ms after stimulus presentation which is said to be face specific (Darque et
al., 2012). Therefore for this dissertation, faces will be used in the emotional
attentional blink paradigm.
The use of emotional faces in the attentional blink paradigm has facilitated the
study of how anxiety influences early perceptual processing. Angry schematic faces
reduce the AB effect in normal adults (Maratos, Mogg, & Bradley, 2008). Among
anxious individuals there is heightened sensitivity to negative emotions like anger and
fear (de Jong & Martens, 2007; B. De Martino, Kalisch, Rees, & Dolan, 2008; Fox,
Russo, & Georgiou, 2005). Highly anxious individuals show a reduced AB effect with
fearful faces compared to happy faces (Fox et al., 2005). Women with higher trait
anxiety also show a reduced attentional blink when T2 targets are faces with anger
compared to happy faces (de Jong & Martens, 2007; Fox et al., 2005). Anxious
individuals therefore perceive emotional faces as more salient and thus are processed
more efficiently because less competition is present for attentional resources (B. De
Martino et al., 2008; Fox et al., 2005; Maratos et al., 2008). It is unknown, however,

15
how anxiety influences early emotional perceptual processing across development and
in a disorder that suffers from attentional problems.
The attentional blink has been successfully adapted to investigate early
perceptual processing in children and in ADHD (Garrad-Cole, Shapiro, & Thierry,
2011; Li, Lin, Chang, & Hung, 2004). However to date there is no study investigating
the role emotion plays in early perceptual processing in children and in ADHD.
Research Goals & Rationale
The influence of emotion on attention and how they interact has been a major
focus in cognitive neuroscience. While there is a considerable amount of literature on
how emotion and attention interact across development, little is known on how
emotion captures attention in the perceptual stage of processing. Even less is known
about how this process may be affected in a developmental disorder in which
attentional function is disrupted, such as ADHD. In Study 1, we used the Attentional
blink paradigm to examine the effect of emotional faces on early visual attention in
children and adults. We also investigated whether these effects are associated with
anxiety in children and adults. Study 2 used the Attentional blink paradigm to examine
the effect of emotional faces on early visual attention in children with ADHD. We also
examined whether these effects are associated with ADHD symptoms and anxiety.

16
CHAPTER II: DEVELOPMENTAL DIFFERENCES IN THE EMOTIONAL MODULATION OF
THE ATTENTIONAL BLINK
Abstract
We examined developmental differences in early stages of attention processing
for socio-emotional information. Adults and children (8–14 years) performed an
attentional blink task that required identification of a dog face (T1) and either an angry
or neutral face (T2) among scrambled face distractors. The anticipated attentional
blink phenomenon was found with both neutral faces and angry faces, reduced
detection at earlier lags that increased across later lags, in both adults in children. We
found a developmental difference such that children took more time to detect faces
than adults. Among children, those with higher trait anxiety showed a smaller
magnitude of the attentional blink. Emotional stimuli reduced the attentional blink, as
expected, but this effect did not differ developmentally. However, overall detection of
emotional stimuli was higher for emotional than neutral stimuli, to a larger extent in
children relative to adults. Together, these findings are the first to reveal
developmental differences in early stages of attention processing and its modulation by
emotion.

17
Introduction
When a person blinks, there is information that the visual system will not perceive
and therefore not process; like an eye blink, attention appears to blink as well. When a
person is asked to search for a specific target within a rapid series visual presentation
(RSVP) of irrelevant stimuli (~100ms), upon successful detection of that target, a
second subsequent target will not be perceived nor processed, therefore resulting in an
attentional blink (D. E. Broadbent & Broadbent, 1987; Raymond et al., 1992).
Detection of the second subsequent target improves when there is more time in
between the presentation of the first and second target (e.g. 600ms) than less time (e.g.
200ms). The attentional blink is a natural phenomenon that occurs and has been
instrumental in studying early stages of attention processing.
Theoretical models of the attentional blink suggest a two-stage process that has its
limitations. Detection of a second subsequent target decreases because attentional
resources are depleted when the first target is being processed (Shapiro, Caldwell, &
Sorensen, 1997). All items in the RSVP stream are processed at stage 1, in order to
identify target features. Target selection occurs at Stage 2 and requires short-term
memory. Decreased T2 detection occurs because Stage 1 has priority and therefore
Stage 2 processing cannot begin until Stage 1’s is complete. A bottleneck effect is said
to occur at Stage 2 because it cannot process additional items from Stage 1 until it is
done processing (Arnell, Helion, Hurdelbrink, & Pasieka, 2004; Marvin M. Chun &
Potter, 1995; Jolicoeur, 1998; Martens & Wyble, 2010). It is believed that more

18
efficient consolidation of the first target should reduce the wait at the bottleneck,
resulting in a smaller attentional blink (Arnell, Howe, Joanisse, & Klein, 2006). The
more unique a target is among the distractors, the easier and thus faster processing that
occurs at Stage 2, which results in increased T2 detection at earlier lags (M M Chun,
1997; Marvin M. Chun & Potter, 1995). Recent research has begun to focus on using
salient stimuli like emotions as targets in the attentional blink to investigate early
perceptual processes.
Emotional information has been shown to influence the attentional blink. When
emotional information is introduced as the second subsequent target in the attentional
blink paradigm, the blink is reduced such that the second emotional stimuli can be
detected at earlier time points, (e.g. before 500ms), (Anderson & Phelps, 2001). One
hypothesis for the attenuated attentional blink with emotion is that the increased
salience of an emotional face among non-emotional face results in a low threshold for
detecting the emotional face, reducing the bottleneck effect (de Jong & Martens, 2007).
Another explanation for the reduced attentional blink is evidence suggesting an
alternate pathway that is faster in processing emotional stimuli that engages the
amygdala to reduce the need for attentional resources (Anderson & Phelps, 2001;
Pessoa, 2002). The reduced attentional blink has been confirmed with various types of
emotional stimuli: words (Benedetto De Martino, Strange, & Dolan, 2007), scenes (B.
De Martino et al., 2008), schematic faces (Maratos et al., 2008) and photographs of

19
faces (Stein, Peelen, Funk, & Seidl, 2010). The reduced attentional blink for emotional
information suggests emotion is prioritized in early stages of attention processing.
Developmental studies confirm that emotional information is prioritized in later
stages of attention processing, however little is known about earlier stages. Infants
prefer faces over other stimuli (Grossmann & Johnson, 2007; Johnson, 2005) and are
biased towards happy faces (Faroni, et al., 2007) indicating that full brain maturation is
not necessary to detect and prefer an emotional stimuli. A bias for angry faces
emerges later in early childhood, between 1-3 years, (Vaish, Grossman, & Woodward,
2008), suggesting that a bias for negative emotions develops from a very young age.
In older children (ages 10 to 13 years old) the bias for angry faces created more
interference, on an executive control task that used emotion as irrelevant stimulus, than
in young children (ages 6 to 9 years old) and adults (K. A. Barnes, Kaplan, & Vaidya,
2007). This study suggests that emotional information will compete for attentional
resources even when it is not relevant to the task. This competition for attentional
resources by emotion appears to be more vulnerable in older children than in adults.
Therefore it is necessary to examine developmentally whether emotional information
has a preferred status at early stages of attention processing.
While a few studies have examined early stages of attention processing using the
attentional blink phenomenon in children, none have investigated whether the
magnitude of the attentional blink changes across development with socio-emotional
information. Only one study has looked at how the attentional blink changes across

20
age (Garrad-Cole et al., 2011). In this study, children of seven, twelve, and
adolescents of fifteen years of age were compared to adults on an attentional blink task
that used shapes as targets. All but children of seven years of age recovered from the
attentional blink by 600 ms, these children took over 1000 ms to recover. This study
demonstrated that with increasing age, the attentional blink is reduced. This study
suggests that the profound blink found amongst younger children is due to inefficient
processing at Stage 2, where the bottleneck occurs, and that by 12 years of age children
are showing a similar attentional blink pattern to adolescents and adults. We expect to
replicate with our emotional attentional blink paradigm, that with increasing age the
attentional blink is reduced.
More insight about the attentional blink in children can be drawn from studies that
have focused upon two developmental disorders, Attention Deficit Hyperactivity
Disorder (ADHD; (Mason et al., 2005) and dyslexia (Visser, Boden, & Giaschi, 2004).
Children with ADHD show mixed results with one study showing no differences in the
attentional blink relative to gender-matched controls when letters were used as both
targets and distractors (Mason et al., 2005). Another study has demonstrated evidence
for a prolonged blink using letters as target and distractors as well (Li et al., 2004).
Relative to their age-matched controls, children with dyslexia had a larger attentional
blink when shapes were used as targets among random dot distractors (Visser et al.,
2004). Thus, the attentional blink phenomenon occurs in children as young as 8 years
and developmental disorders with attentional and language problems appear to alter it.

21
In healthy children, individual differences in the magnitude of the attentional blink are
associated with impulsivity, a trait associated with ADHD (Chhabildas, Pennington, &
Willcutt, 2001; Gomez, 2003; Oades, Slusarek, Velling, & Bondy, 2002). Specifically,
higher levels of impulsivity in adolescence were related to two characteristics of the
attentional blink: worse subsequent target detection at early lags and a prolonged blink
(Ray Li, Chen, Lin, & Yang, 2005). Thus, children with higher levels of impulsivity
were characterized as having a larger and more long-lasting attentional blink.
Impulsivity is characteristic of immaturity and therefore, it is possible that younger
children will show a larger and prolonged attentional blink relative to older children
and/or adults. Currently, there is no study investigating differences in early stages of
attention processing using the emotional attentional blink in children.
Differences seen in early stages of attention processing, through the use of the
emotional attentional blink paradigm, have been attributed to individual differences
among healthy adults in their levels of anxiety. One study assessed healthy females
who either scored high or low on measures of anxiety and revealed a bias for a specific
emotion among anxious individuals(Fox et al., 2005). Highly anxious females have a
shortened attentional blink for fearful faces while less anxious females have a
shortened attentional blink for both fearful and happy faces. In another similar study,
selecting high and low socially anxious healthy females, both high and low socially
anxious women had a shortened attentional blink in the presence of an angry face, (de
Jong & Martens, 2007). This study further established the anger superiority effect,

22
which is the tendency for healthy individuals to selectively attend to angry faces,
(Hansen & Hansen, 1988), seen in other studies with other paradigms (Bradley, Mogg,
White, Groom, & de Bono, 1999; Schutter, Putman, Hermans, & van Honk, 2001). It
is unknown whether individual differences in anxiety may also explain the differences
in early stages of attention processing in healthy children with emotional information.
In the present study, we examined developmental differences in the attentional
blink for faces with and without emotional expressions. We hypothesized that children
would have a larger attentional blink with neutral faces than adults because their brains
are still developing and thus are less efficient than adult brains to process a subsequent
target. With regards to the influence of emotion, we hypothesize that children will
benefit the most from the emotional information resulting in a smaller attentional blink
than adults. Lastly, we hypothesize that both state and trait anxiety will be related to
the size of emotional attentional blink, such that more anxious individuals will be more
sensitive to emotional faces and thus have a smaller attentional blink.
Methods
Participants
Thirty eight 7-14 year old children were recruited from the Washington DC
metropolitan area through advertisements. Thirty five undergraduate students (16
female; mean age = 19.3, SD = 1.1 years) were recruited from Georgetown University
Psychology classes. The final sample included 31 children (15 female; mean age =

23
11.5, SD = 1.9 years, Range= 8-14; mean IQ = 111, SD = 14.3, Range= 91-138) and 33
adults (16 female; mean age = 19.4, SD = 1.1 years, Range= 18-22) following
exclusion of 7 children and 2 adults who did not meet performance criteria (see below
in results). All participants were paid $10 for participation. Written informed consent
from adults and parents of children, and assent from children were obtained according
to Georgetown University Institutional Review Board guidelines.
Participants with neurological and psychiatric conditions were excluded by
self-report for adults and standardized measures for children. Exclusion criteria for
children was: 1) Full scale IQ below 85 (estimated using Vocabulary and Block Design
subtests of the Wechsler Abbreviated Scale of Intelligence, (WASI; (Weschler, n.d.).
2) Presence of neurological disorder by parent-report and psychiatric disorder based
upon the Behavior Assessment System for Children (BASC; (Reynolds & Kamphaus,
1992) including conduct disorder. 3) Score below 50 on the Word Attack and Letter
Word Identification subtests of the Woodcock-Johnson reading test to screen for
reading problems (WJ III; (R.W. Woodcock, K.S. McGrew, & N. Mather, 2000).
Stimulus materials
All stimuli were displayed on a white background at a viewing distance of 60 cm on a
17 inch monitor using E-Prime version 1.1 (Psychology Software Tools Inc., Pittsburg,
PA). Twenty color face photographs of adult faces (10 with neutral facial expressions
and 10 with angry facial expressions) were obtained from the NimStim set,
www.macbrain.org; faces did not overlap across tasks. The Emotional Identification

24
Task consisted of six colored photographs of adults with angry and neutral facial
expressions (three of each gender). The Attentional Blink Task consisted of three
classes of stimuli: 1) Two color photographs of dogs, from the website
www.dogbreedinfo.com, were used as the first target stimuli (T1). 2) Twenty eight
colored photographs of scrambled faces were used for the distractor stimuli. Faces of
seven females and seven males with neutral expressions were scrambled with the face
outline preserved and the interior portion scrambled using the scramble feature from
the website http://www.faceresearch.org ). 3) Fourteen colored photographs of faces
with angry and neutral expressions (seven of each gender) were used for the second
target stimuli (T2).
Procedure
First, participants completed the State-Trait Anxiety Inventory questionnaire
[STAI Form Y for adults, and STAI-CH for children; (Spielberger, Edwards,
Montuori, & Lushene, 1983)]. Then, participants completed the Emotional
Identification Task followed by 20 practice trials and 188 test trials of the Attentional
Blink (AB) task.
Emotional Identification Task
The goal of the task was to assess whether participants recognized the
emotional valence of faces accurately. The task consisted of 6 trials. Participants
began the task by pressing the space bar in response to the words “Get Ready!” on the
screen. Either an angry face or neutral face was presented centrally for 2000 ms,

25
following which the participant had to press the number “1” key for “angry” or the
number “2” key for “no emotion” in answer to the question “Is this person:”.
Participants could take as long as they needed to record their response. Recognition
accuracy of 70% or higher was required to move on to complete the AB task. All
subjects achieved this criterion at first attempt.
Attentional Blink task
The AB task consisted of 188 trials divided into four blocks of 47 trials each.
Participants pressed the space bar in response to the words “Get Ready!” on the screen
to start each block; they could take a self-paced break in between blocks. Each trial
began with the presentation of a fixation cross for 750 ms followed by a rapid serial
visual presentation (RSVP) stream of 15 stimuli presented for 100 ms each (see Figure
1). Participants were instructed to look for a dog and informed that sometimes a face
would appear and to pay attention because the pictures came on the screen very fast.
Each trial ended with the a question on the screen “Did you see a dog?”. Participants
pressed “1“ for “yes” or “2” for “no” on the number pad of the keyboard. Following
their response, a second question “Did you see a face?” appeared on the screen. Once
again, participants indicated their response by using the number pad. For each
question, participants had 5000 ms to respond.
The AB task consisted of eight types of trials, two types of catch trials, and six
types of AB trials varying in the facial expression of the T2 stimulus, at each of three
lags. Specifically, catch trials included: 1) 30 T1-only trials, in which only the T1, a

26
dog, appeared in the RSVP stream but no T2 appeared; 2) 32 T2-only trials, in which a
T2 (either an angry or neutral face) appeared in the RSVP stream without a preceding
T1. These trials were included in order to ‘catch’ children with a bias to respond with
“yes” to both questions at the end of the RSVP stream.
On the remaining AB trials, a T2 was either a face with a neutral or angry
expression and followed the T1 with three lags: 1) 21 trials with a neutral T2
appearing 200 ms after the T1, (termed Neutral-Lag 2); 4) 21 trials with an angry T2
appearing 200 ms after the T1, (termed Angry-Lag 2); 5) 21 trials with a neutral T2
appearing 400 ms after the T1 (termed Neutral-Lag 4); 6) 21 trials with an angry T2
appearing 400 ms after the T1, (termed Angry-Lag 4); 7) 21 trials with a neutral T2
appearing 800 ms after the T1 (termed Neutral-Lag 8); 8) 21 trials with an angry T2
appearing 800 ms after the T1 (termed Angry-Lag 8). Further, within the RSVP
stream, the serial position of T1 was varied among stimuli 4, 5, and 6 in the trial
sequence. The position of T1 was varied at these points to minimize missing the T1
because it occurred too early in the sequence that individuals would not have enough
time to adapt to the RSVP presentation and to minimize working memory demands of
remembering they saw the T1 by the end of the trial. Thus, T2 followed each of these
positions at the three lags described above such that it was either the second (Lag 2),
fourth (Lag 4), or eighth (Lag 8) stimulus after the T1. All the trials were
pseudorandomized into blocks so that no more than two successive trials were of the
same type within each block. Blocks were created to allow breaks after 47 trials, and

27
were balanced to include the same number of each trial types. Two versions of the task
were created to counterbalance block presentation order.

28
Figure 1. Rapid serial visual presentation (RSVP) stream for the Attentional Blink
Task. Participants monitored two target stimuli (T1 & T2) embedded among 13
distractor stimuli, each presented for 100ms with no interstimulus interval. The first
target stimulus (T1) was a dog face and the second target stimulus (T2) was a human
face (either neutral or angry). The distractors were scrambled faces. a) T1-only catch
trials in which only the dog face was presented, b) T2-only catch trials in which only
an angry or neutral face was presented, c) Double target trials in both T1, the dog face,
and T2, the human face, were presented. The two targets were separated by either 2, 4
or 8 distractors, (200ms, 400ms, or 800ms, respectively). At the end of the RSVP
stream, participants indicated by key press whether they had seen saw a dog and a face

29
Results
Participants who performed worse than chance (50%) on the T2 catch trial were
excluded from analysis. This criterion resulted in the exclusion of seven children and
two adults from the total cohort of participants. A response was scored as correct if the
participant correctly identified first the T1 and then the T2, as for T1 incorrect trials the
source of T2 errors is unknown (Marvin M. Chun & Potter, 1995). For each
participant, percentage of accurate T2 trials was computed for each of the six types of
AB trials. Percent accuracy was analyzed in a mixed analysis of variance with Group
(Children, Adults) as the between-subjects factor and emotion (T2 - angry, neutral) and
lag (2, 4, 8) as within-subjects factors. Measures of effect size (partial eta-squared for
analyses of variance [ANOVA], Cohen’s d for means, r2 for correlations) are presented
for the attentional blink effects. Figure 2 summarizes the results by trial type and
groups.

30
Figure 2. Mean percentage correct scores as a function of group (adult vs. child),
emotional face of T2 (angry vs. neutral), and lag (2, 4, 8).
We found three main effects. Overall, T2 accuracy was higher in adults than in
children (main effect of group), F(1,62) = 21.77, p< .001, η2=.26 and both groups were
more accurate when the T2 was a face with angry than neutral expression (main effect
of emotion), F(1,62) = 39.84, p < .001, η2=.10. Further, T2 accuracy differed by lag
(main effect of lag), F(2,61) = 25.99, p< .001, η2=.17. Post-hoc tests showed that T2
detection at lag 2 was lower than at lag 4, t(63) = 4.51, p < .001, d= 0.80, and lag 8,

31
t(63), = 6.75, p < .001, d= 1.76. T2 detection at lag 4 was also lower than lag 8, t(63),
= 4.48, p < .001, d= 0.82. These findings indicate that our paradigm produced the
expected attentional blink phenomenon, decreased accuracy at the shortest lag relative
to longer lags.
In addition to these main effects, several key interactions were observed. First,
a significant lag X group interaction was obtained, F(2,61) = 5.39, p < .01, η2=.03,
suggesting that the magnitude of the overall attentional blink (neutral and emotional)
differed by age. Paired t-tests revealed that both groups showed improvement in
overall T2 detection with each increasing lag: Adults - lag 2 versus lag 4, t(32), = 3.45,
p < .005, d= 1.53; lag 4 versus lag 8, t(32), = 3.22, p < .001, d= 0.68, and lag 2 versus
lag 8, t(32), = 4.70, p < .005, d= 2.18; Children - lag 2 versus lag 4, t(30), = 3.06, p <
.01, d= 0.78; lag 4 versus lag 8, t(30), = 4.02, p < .001, d= 1.13, and lag 2 versus lag 8,
t(30), = 5.44, p < .001, d= 2.11. These results demonstrate that both adults and
children had an attentional blink effect, poor T2 accuracy at shorter lags that improved
as the lag increased. However, adults’ accuracy was better at each lag. Independent
samples t-test revealed that adults had higher T2 detection at lag 2, t(62), = 4.52, p <
.001, d= 1.15, lag 4, t(62)= 4.18, p < .001, d= 1.02 , and lag 8, t(62)= 3.84, p < .001, d=
.97. When a difference score was calculated to assess the magnitude of the blink, (i.e.
lag 8 performance minus lag 2), there was a larger magnitude of blink in children than
in adults, t(62), = 2.63, p < .05, d= 0.67, such that children had a more profound blink,
(lower T2 accuracy) than adults.

32
Second, there was also a significant lag X emotion interaction, F(2,61) = 10.97,
p<.001, η2=.05, suggesting that the magnitude of the attentional blink differed by the
nature of the T2. Collapsing across groups, paired t-tests showed a higher T2 accuracy
for angry trials relative to neutral trials at lags 2 and 4 but not lag 8: lag 2, t(63), =
6.37, p < .001, d= 1.52, lag 4, t(63)= 3.25, p < .005, d= 1.02 , and lag 8, t(63)= 1.72, p
= .09, d= .23. Thus, better T2 detection for emotional faces was limited to earlier lags.
Among the angry trials, T2 detection improved from lag 2 to lag 8, t(63)= 3.05, p <
.005, d= 0.93 and also from lag 4 and lag 8, t(63)= 2.74, p < .01, d= 0.59, but not from
lag 2 to lag 4, t(63)= 1.47, p = .15, d= 0.36. Among the neutral trials, T2 detection
improved with increasing lag length: lag 2 and lag 4, t(63), = 5.18, p < .001, d= 1.06;
lag 4 and lag 8, t(63), = 4.29, p < .001, d= 0.89, and lag 2 and lag 8, t(63), = 7.45, p <
.001, d= 2.14. This finding was further substantiated when a difference score was
calculated to assess the magnitude of blink (lag 8 minus lag 2), a significantly larger
blink was found on neutral trials than on angry trials, t(63), = 4.71, p < .001, d= 1.56.
These findings indicate that we observed the expected effect of emotional stimuli on
the attentional blink phenomenon, a shorter blink for angry than neutral faces.
Third, a significant Emotion by Group interaction was found, F(1, 62) = 5.67,
p< .05, η2=.01. Independent sample t-tests revealed better T2 detection (when
collapsing across all lags) in adults than children for angry trials, t(62)= 4.18, p < .001,
d= 1.06 and neutral trials, t(62)= 4.61, p < .001, d= 1.17. Paired t-tests revealed a
significantly higher T2 accuracy for angry than neutral trials in both adults, t(32)=

33
3.61, p < .001, d= 1.47, and children, t(30)= 5.09, p < .001, d= 1.26. When a
difference score was calculated to assess the influence of emotion , (i.e. angry face
trials’ performance minus neutral face trials), children had a larger difference score
than adults, t(62), = 2.38, p < .05, d= 0.60, indicating that accuracy improved much
more with the emotional face in children. Figure 3 summarizes graphically that adults
had better detection of T2 for both angry and neutral faces than children, however
children had a greater improvement in accuracy when an emotional face was presented
as T2. Most importantly, the Group X Lag X Emotion interaction was not significant,
F(2,61) = .632, p =.54, η2=.033 indicating that the effect of the facial expression of the
T2 stimulus did not influence the magnitude of the attentional blink differently in the
two age groups.
Figure 3. Means percentage correct scores for emotional face of T2 (angry vs neutral)
and group (adult vs. child).

34
In order to assess individual differences in state and trait anxiety, both the adult
and child STAI, raw scores were converted to Z-scores. There was a difference in trait
anxiety between adults (raw: mean = 37.4, SD = 12.2; Z-score: mean = 0.27, SD =
1.23) and children (raw: mean = 31.6, SD = 5.14; Z-score: mean = -0.31, SD = .52),
t(60)= 2.37, p < 0.05, d= 0.96) such that adults were more anxious than children. In
lieu of the difference, we then assessed individual differences in the size of the
attentional blink, (indexed by subtracting T2 accuracy for neutral at lag 8 from T2
accuracy for neutral at lag 2), separately in each group. A negative correlation between
the size of the attentional blink and trait anxiety level was shown in children, such that
more anxious children had a smaller attentional blink, (r= -0.41, p < .05, r2= 0.16); this
relationship was not significant in adults (r= 0.21, p = 0.25, r2= 0.04). Lastly to assess
whether trait anxiety was associated with the emotional attentional blink (indexed by
subtracting T2 accuracy for neutral from T2 accuracy for angry faces at Lag 2). No
significant correlations were found in either group with trait anxiety. Furthermore,
state anxiety was also higher in adults (raw: mean = 33.0, SD = 8.68; Z-score: mean=
0.28, SD= 1.18) than children (raw: mean = 28.2, SD= 3.57; Z-score: mean= -0.38,
SD= .49), t(55)= 2.55, p < .001, d= 0.70, but was not associated with either the size of
the attentional blink or its modulation by emotion.
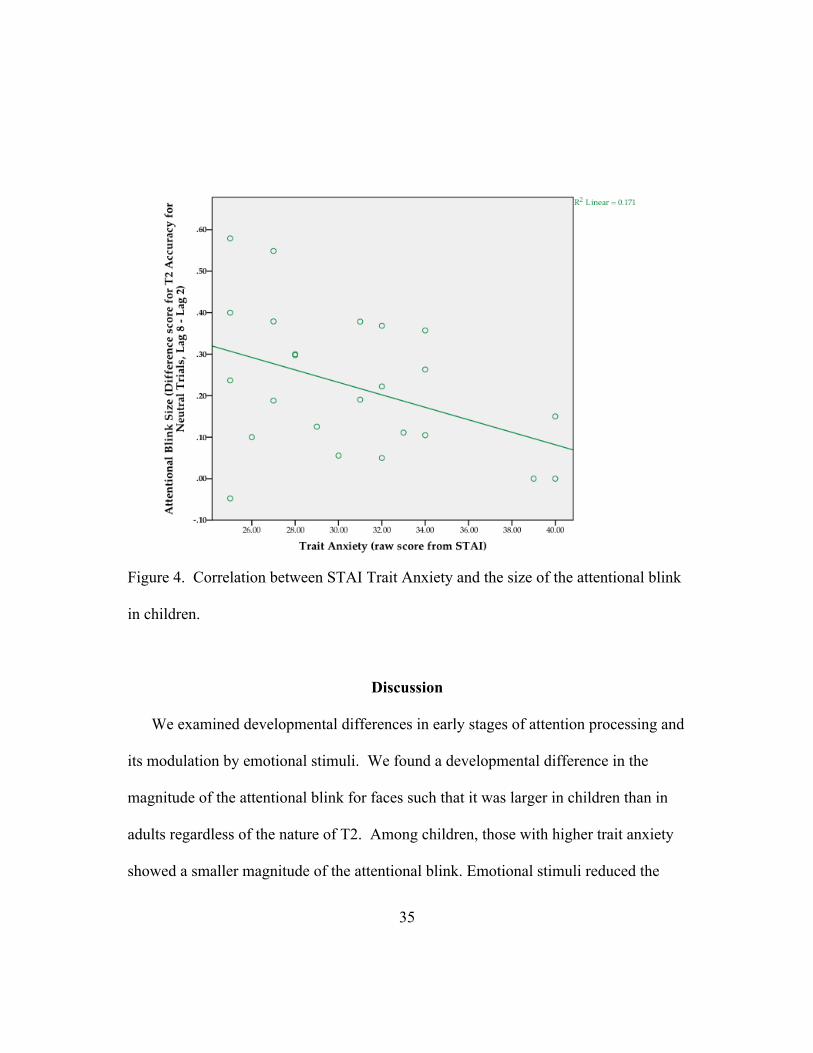
35
Figure 4. Correlation between STAI Trait Anxiety and the size of the attentional blink
in children.
Discussion
We examined developmental differences in early stages of attention processing and
its modulation by emotional stimuli. We found a developmental difference in the
magnitude of the attentional blink for faces such that it was larger in children than in
adults regardless of the nature of T2. Among children, those with higher trait anxiety
showed a smaller magnitude of the attentional blink. Emotional stimuli reduced the

36
attentional blink, as expected, but this effect did not differ developmentally. However,
overall detection of emotional stimuli was higher for emotional than neutral stimuli, to
a larger extent in children relative to adults. Together, these findings are the first to
reveal developmental differences in early stages of attention processing and its
modulation by emotion.
The introduction of socio-emotional information in our paradigm is central to
understanding our findings. Attentional blink paradigms that have been used in
children to date have used non-social stimuli: words, (Anderson & Phelps, 2001),
chinese characters, (Ray Li et al., 2005), letters, (Marois & Ivanoff, 2005), numbers,
(Colzato, Slagter, Spapé, & Hommel, 2008), shapes, (Garrad-Cole et al., 2011). We do
see a larger more profound blink with children than adults as the previous
developmental study that used non-social stimuli, shapes, (Garrad-Cole et al., 2011).
We do not however see a developmental difference between neutral and emotional
faces for the magnitude of the attentional blink, despite seeing the attentional blink
phenomenon (main effect of lag) and a difference among adults and children in the
nature of the emotional face, (significant Emotion x Group interaction). Therefore it is
important to look at our study more closely to understand the influence of emotion
across development in early stages of attention processing. The introduction of social
information, as is the case of with neutral faces, reveals a developmental difference in
early stages of attention processing seen in children having a larger attentional blink
than adults, (a significant Group X Lag interaction is observed when a repeated

37
measures ANOVA is analyzed with the neutral face trials alone). However when
emotional information is introduced, as in the case of the angry faces, we see no
difference in the size of the attentional blink between children and adults, (there is also
no longer a Group x Lag interaction present when a repeated measures ANOVA is run
with only the angry face trials). It maybe that neutral faces are salient and the
introduction of an emotion is not as salient to produce a strong effect or difference in
early stages of attention processing, when comparing neutral and angry faces together.
In order to better assess the influence of emotion a non-social stimulus like shapes,
(that have been previously used), could be additionally incorporated to the task to
better distinguish between the influence of social and emotional information. Another
alternative to assess the influence of emotion in the attentional blink would be to use
the emotional information as the first target. This “reverse” version of the attentional
blink has been used more recently to assess how long an emotional stimulus engages
early attentional processes (de Jong & Martens, 2007). In conclusion the specifics of
our paradigm may in fact explain our lack of a specific developmental difference in the
emotional modulation of early stages of attention processing.
The developmental difference found in the attentional blink can be explained by
recent research showing developmental differences in event-related potential (ERPs)
indicators of early face processing. Developmental ERP studies have shown the
presence of two components that are indicative of early face processing, the N170 and
the P1 (M. J. Taylor et al., 2004). The N170, in young children between 4 and 7 years

38
of age, is found to be bifid, (two peaks) and between 10 and 13 years of age, (our
sample included children ranging from 8 to 14 years of age), the bifid pattern merges
to one, looking more like the single peak adult N170. The bifid N170 in childhood is
reflective of a slower inefficient process that is distributed across different areas, and
as the brain matures, the processing of faces is localized to one region. The P1, the
positive component preceding the N170, is an indicator of earlier stage processing than
the N170. The P1 latency has been shown to decrease with age among a group of
children ranging from 4 to 13 and adults. The larger latency in young children (4 years
to 10 years old) reflects a slower processing speed. In adulthood, myelination,
facilitates neurotransmission that results in faster and more efficient processing (Luna
& Sweeney, 2004; Rubia et al., 2000; Toga, Thompson, & Sowell, 2006). The
presentation of a larger blink by children maybe a result of a larger distributed set of
regions trying to encode each face and thus require more time to not only encode but
communicate to each other in order to successfully identify T2, indicating an immature
face processing network.
Our study continues to show converging evidence for the preferential treatment for
emotion in early attention processing in both adults and children. Emotion modulated
the attentional blink at early lags but its effect is reduced at later lags. The modulation
of emotion was an expected finding since it has been previously shown with other
emotional attentional blink studies (Jefferies, Smilek, Eich, & Enns, 2008; Maratos et
al., 2008; Srivastava & Srinivasan, 2010; Stein et al., 2010). One key difference

39
between our paradigm and previous studies’ is the high accuracy present on the lag 2
trials in the adult group, approximately 87% for the neutral faces and 95% for angry
faces, other studies using faces have ranged from 30%-50% for neutral faces and 65%-
88% for angry faces (de Jong & Martens, 2007; Maratos et al., 2008). In the later lags,
adults perform at near ceiling, over 90%. The higher accuracy could be a result of an
adaptation to our paradigm, the initial target, T1, is a dog face, that is salient enough to
facilitate faster processing of T1 allowing attentional resources to become readily
available for a subsequent target to be processed. The use of a dog face was done to
make the task child-friendly. Both adults and children improved with the presentation
of an angry face as the T2 at any time point, this finding is consistent with other
literature showing a benefit from angry or threatening faces in performance on various
tasks in children (Hadwin et al., 2003; Heim-Dreger, Kohlmann, Eschenbeck, &
Burkhardt, 2006; Perez-Edgar et al., 2010) and adults (Mogg, Bradley, de Bono, &
Painter, 1997; Schub, Meinecke, Abele, & Gendolla, 2006; Schutter et al., 2001).
Lastly, our study provides correlational evidence for a vigilant-avoidant bias
among more anxious individuals for ambiguous emotion (Amir, Foa, & Coles, 1998;
MacLeod & Cohen, 1993; Stopa, 2000). In our study, the neutral faces appeared to be
more salient to highly anxious children, than angry faces in our sample, as indicated by
a smaller attentional blink with neutral faces. Emerging literature suggests that a
critical discriminate among anxious individuals is in how they process ambiguous
information. Anxious individuals appear to perceive ambiguity as threatening and

40
therefore avoid any ambiguous information and it is evident from childhood, (A. P.
Field & Lester, 2010). Neutral faces are considered to provide ambiguous information
in previous child studies, (for review(Leppanen & Nelson, 2009). In our study of early
attentional processing, we see that the neutral faces are very salient to highly anxious
children, since their attentional blink is reduced. It maybe the case that in early stages
of attention processing children are vigilant towards ambiguous stimuli and when
assessed in later stages of attention, they actually display the avoidant bias traditionally
seen in emotion studies (Perez-Edgar et al., 2010). Important to note is that adults did
not show the heightened response to neutral faces. One explanation is that adults
understood that these were neutral faces and thus did not perceive any ambiguity.
Most of the adult studies assessing the role of anxiety in early attentional processing
have selected those adults who score higher on measures of anxiety. In our study we
did not select the more anxious individuals, none of our subjects met the criteria for an
anxiety disorder. In conclusion, further research should be done to confirm this early
vigilant, later avoidant pattern of behavior in more anxious children and investigate if
this pattern might be present in adults as well.
Further studies should continue to investigate early automatic attentional processes.
It is important to differentiate whether the differences we see with emotion are due to
overall salience difference or truly a preference for emotion. The reason we must
distinguish this is because the size of the attentional blink varies depending on the
relationship between the targets and the distractors in the trials, the greater difference

41
between the targets and distractors, the smaller the blink, and increased sensitivity
(Anderson, 2005; Marvin M. Chun & Potter, 1995). Lastly, within the realm of
individual differences, presence of a short allele for the serotonin transporter
corresponds with an increased reactivity to threatening stimuli in an individual on an
emotional go-no-go task (Hariri, 2002; Hariri et al., 2000). Therefore genetic
differences in the serotonin transporter may help explain individual differences in early
automatic attention processing. In conclusion, our study is one of the first pieces of
evidence for developmental differences in the early automatic attention processing and
the increased saliency of both social and emotional information in early attention
processing.

42
CHAPTER III: EMOTIONAL MODULATION OF THE ATTENTIONAL FUNCTION IN
CHILDHOOD ATTENTION DEFICIT/HYPERACTIVITY DISORDER
Abstract
The attentional blink (AB) phenomenon was used to assess the modulation of
emotion in early visual attention in children with Attention Deficit/Hyperactivity
Disorder (ADHD). The AB effect is the temporary loss of perceptual awareness that
follows successful target identification in a rapid serial visual presentation. Emotional
stimuli can either attenuate or abolish the AB effect, which suggests that there is a
preferential access to early visual attention processing for emotional information. In
the present study, we examined the AB effect with neutral and angry faces in 7-14 year
old children with and without an ADHD diagnosis. Children with ADHD exhibited an
AB effect for neutral faces and it was reduced for angry faces to the same extent as
control children. Children with ADHD who were more inattentive benefitted the most
from the presentation of an angry face in early lags. Control children who were more
anxious had a smaller AB effect with neutral faces. Thus, early visual attentional
processing and its modulation by emotion are intact in ADHD, however individual
differences in anxiety and inattention symptoms influence it.

43
Introduction
Attention Deficit Hyperactivity Disorder (ADHD) is one of the most common
neuropsychiatric disorders in childhood, affecting 3-7% of school-aged children and
continues to affect between 30% and 60% of adults (Wender, Wolf, & Wasserstein,
2001). ADHD is characterized by symptoms of inattention, hyperactivity and
impulsivity, which are believed to be indicative of a core cognitive deficit in executive
functioning (Barkley, 1997; Cubillo, Halari, Smith, Taylor, & Rubia, 2012). More
recently, emotional problems exhibited by children with ADHD have gained attention
and are thought to contribute to ADHD pathophysiology (Johansen, Aase, Meyer, &
Sagvolden, 2002; Nigg & Casey, 2005; Sonuga-Barke, 2005). Individuals with
ADHD have difficulties in establishing and maintaining social interactions and
relationships (Biederman et al., 1996; Friedman et al., 2003), and the same medications
that aid executive dysfunction in ADHD also reduce emotional dysfunction (Reimherr
et al., 2005; Williams et al., 2008). However, emotional dysfunction in ADHD is not
well characterized currently.
There are differences in the perception of emotional stimuli by individuals with
ADHD. When a positive word was read in a dichotic listening task, children with
ADHD did not respond faster when the word was presented in the right ear, indicating
reduced reactivity to positive stimuli (Becker, Doane, & Wexler, 1993). Adults with
ADHD showed a reduced startle response to pleasant picture scenes, suggesting that
positively valenced stimuli are not salient in ADHD (Conzelmann et al., 2009).

44
Adolescent ADHD boys had more signs of emotional impairment in that they had
higher anxiety and depression levels and difficulty correctly identifying negative (e.g.,
angry and fearful) emotional faces than controls (Williams et al., 2008) . These
children also exhibited an atypical ERP response: reduced occipital lobe activity during
early perceptual analysis of the emotional expression (within 120ms), followed by an
exaggerated amount of activity associated with structural encoding (120-220ms) and
lastly a slowed and reduced amount of temporal brain activity indicative of context
processing (300-400ms). Upon receiving methylphenidate treatment, the boys’ ability
to identify the negative emotions improved and a more normalized ERP response to
emotional faces was observed. Further, adults with ADHD who self reported a
heightened negative response to the emotional face which subsequently interfered with
their ability to successfully categorize an emotional face (Rapport, Friedman, Tzelepis,
Van Voorhis, & Friedman, 2002). Together, these studies suggest that negative
emotional stimuli produce a greater response than positive emotional stimuli in
ADHD.
The attentional blink paradigm is a tool that allows the assessment of early visual
attentional processing with minimal executive control demands. The paradigm
requires a person to search for a specific target within a rapid serial visual processing
(RSVP) stream of irrelevant stimuli (~100ms). Upon successful detection of that
target, a second subsequent target will not be perceived; this effect is referred to as an
attentional blink, AB, (D. E. Broadbent & Broadbent, 1987). Detection of the second

45
subsequent target improves when there is more time in between the presentation of the
first and second target (e.g. 600ms) than less time (e.g. 200ms).
Theoretical models posit that the AB effect indexes information processing
limitations in a two-stage process. Detection of a second subsequent target decreases
because attentional resources are depleted when the first target is being processed
(Shapiro et al., 1997). All items in the RSVP stream are processed at stage 1, in order
to identify target features. Target selection occurs at Stage 2 and requires short-term
memory. Decreased T2 detection occurs because Stage 1 has priority and therefore
Stage 2 processing cannot begin until Stage 1’s is complete. A bottleneck effect is said
to occur at Stage 2 because it cannot process additional items from Stage 1 until it is
done processing (Arnell, Helion, Hurdelbrink, & Pasieka, 2004; Chun & Potter, 1995;
Jolicoeur, 1998; Martens & Wyble, 2010). It is believed that more efficient
consolidation of the first target should reduce the wait at the bottleneck, resulting in a
smaller AB (Arnell et al., 2006). Newer models that incorporate neuroimaging
evidence believe that the bottleneck is caused by an attentional filter under executive
control in frontal regions (Maratos et al., 2008; Martens & Wyble, 2010).
Emotional information has been shown to influence the AB. When emotional
information is introduced as the second subsequent target in the AB paradigm, the
blink is reduced such that the second emotional stimuli can be detected at earlier time
points, before 500ms (Anderson & Phelps, 2001). The reduced AB has also been
confirmed with socio-emotional stimuli like schematic faces (Maratos et al., 2008) and

46
photographs of faces (Stein et al., 2010). Reduction of the AB for socio-emotional
information suggests a prioritization in early attentional processing that eliminates the
bottleneck effect.
Studies of the AB in ADHD show mixed results. There is evidence in both
children and adults with ADHD that the AB is intact with non-social stimuli like letters
(Carr, Henderson, & Nigg, 2006; Mason, Humphreys, & Kent, 2005). Other studies
report an atypical AB with non-social stimuli. In adults with ADHD, a prolonged
blink has been reported with letters in which there is below chance detection of T2 and
an increased amount of lags before recovering from the blink (Hollingsworth,
McAuliffe, & Knowlton, 2001). Increased gaze instability has been proposed to
explain the prolonged blink with letters in adults with ADHD (Armstrong & Munoz,
2003). Children with ADHD have also shown a prolonged blink with numbers and
Chinese characters (Li et al., 2004). Increased symptoms of impulsivity have been
related to a prolonged AB in control children with numbers and Chinese characters (Li,
Chen, Lin, & Yang, 2005) and children with ADHD (Chhabildas et al., 2001; Gomez,
2003; Oades et al., 2002). In contrast, adolescents with the inattentive subtype of
ADHD did not show the expected AB effect with letters such that they had increased
detection at early lags but as the lag interval increased, detection of T2 did not increase
suggesting attentional filtering problems (Carr et al., 2006). Therefore differences in
the AB effect are related to symptoms of ADHD. To date there is no study
investigating the influence of socio-emotional information on the AB in ADHD.

47
Anxiety is a common symptom that is comorbid with ADHD and is associated with
differences in the emotional modulation of the AB. Individuals with increased anxiety
levels have consistently shown more sensitivity to emotional information and thus
exhibited a smaller AB for emotional stimuli (Amir, Taylor, Bomyea, & Badour, 2009;
de Jong & Martens, 2007; Fox et al., 2005; Srivastava & Srinivasan, 2010). It is also
important to investigate the influence of anxiety in ADHD because up to 25% of the
child ADHD population display one or more anxiety conditions (Brown, 2009).
Deficits in the self-regulation of emotion are evident in a substantial number of
children with ADHD and these deficits play an important role in determining
functional impairment and the risk of developing comorbid anxiety disorders
(Anastopoulos et al., 2010).
In the present study, we investigated the influence of socio-emotional information
on the AB in children with ADHD and age and IQ matched controls. It is
hypothesized that both controls and children with ADHD would have a reduced AB
with emotional faces. Children with ADHD would be more sensitive to the emotional
face and thus exhibit a smaller AB effect than controls, since they are more susceptible
to attend to negatively valenced faces (Williams et al., 2008) and are more vulnerable
to salient information with the AB as seen with Mason et al. (2005). In addition, as
reported by Li et al. (2005), symptoms of impulsivity would be related to a larger AB
with neutral faces. Based on findings from Carr, et. al. (2010), symptoms of
inattention would be related to an increased detection of emotional information at early

48
lags and therefore, a smaller emotional AB. Lastly symptoms of anxiety would be
related to increased detection of emotional faces at the early lags and a smaller
emotional AB as well as previously found with De Martino et. al. (2008) and Fox et al.
(2005).
Methods
Participants
Thirty-one children with a diagnosis of ADHD and 35 control children, aged 7-
14 years were recruited from the Washington DC metropolitan area through
advertisements. The final sample included 31 control children (15 female; mean age =
11.5, SD = 1.9 years; mean IQ = 110, SD = 13.9, Range= 91-138) and 27 children with
ADHD (12 female; mean age = 11.0 , SD = 1.8 years; mean IQ = 114, SD =14.9,
Range= 87-140) following exclusion of 7 children who did not meet performance
criteria (see below in results). Children with ADHD consisted of ten inattentive and
seventeen combined subtypes. All participants were paid $10 for participation.
Written informed consent from parents of children, and assent from children were
obtained according to Georgetown University Institutional Review Board guidelines.
Participants with neurological and psychiatric conditions were excluded by
parent-report and the Behavior Assessment System for Children (BASC; (Brown,
2009) including conduct disorder. Additional exclusion criteria for children was: 1)
Full scale IQ below 85 (estimated using Vocabulary and Block Design subtests of the

49
Wechsler Abbreviated Scale of Intelligence, (WASI; (Weschler, n.d.). 2) Score below
50 on the Word Attack and Letter Word Identification subtests of the Woodcock-
Johnson reading test to screen for reading disability (WJ III; (R.W. Woodcock et al.,
2000). All participants with ADHD provided documentation from the diagnosing
clinician (either a physician or a psychologist) which was reviewed by a clinical
psychologist (PL) to confirm DSM-IV criteria were met (i.e., presence of 6 out of 9
symptoms of inattention and 6 out of 9 symptoms of hyperactivity/impulsivity in more
than one setting, present before age 7 years and causing impairment). Further, a parent
for each child completed the ADHD Rating Scale (DuPaul, 1998) to characterize
ADHD symptoms of hyperactivity and inattention.
Age (p = 0.30) and Full Scale IQ (p = 0.43) did not differ between ADHD
(Mean age = 11.0 years, SD = 1.76; Mean IQ = 114.11, SD = 14.94) and control
children (Mean age = 11.5 years, SD = 1.91; Mean IQ = 110.45, SD = 13.86). Relative
to controls, ADHD children had higher symptoms of hyperactivity (ADHD: Mean =
12.5, SD= 5.26; Controls: Mean = 4.05, SD = 3.70; t(40)= 6.04, p < .01) and
inattention (ADHD: Mean = 16.2, SD= 5.39; Controls: Mean = 5.62, SD = 3.92; t(45)=
7.73, p < .001). At the time of testing, six ADHD children were stimulant medication
naïve and the remaining withheld medication for over 24 hours.
Stimulus materials
All stimuli were displayed on a white background at a viewing distance of 60
cm on a 17 inch monitor using E-Prime version 1.1 (Psychology Software Tools Inc.,

50
Pittsburg, PA). Twenty color face photographs of adult faces (10 with neutral facial
expressions and 10 with angry facial expressions) were obtained from the NimStim set,
www.macbrain.org.
All face stimuli were drawn from the NimStim set and did not overlap across
tasks. The Emotional Identification Task consisted of six colored photographs of
adults with angry and neutral facial expressions (three of each gender). The AB Task
consisted of three classes of stimuli: 1) Two color photographs of dogs, from the
website www.dogbreedinfo.com, were used as the first target stimuli (T1). 2) Twenty
eight colored photographs of scrambled faces were used for the distractor stimuli.
Faces of seven females and seven males with neutral expressions were scrambled with
the face outline preserved and the interior portion scrambled using the scramble feature
from the website http://www.faceresearch.org). 3) Fourteen colored photographs of
faces with angry and neutral expressions (seven of each gender) were used for the
second target stimuli (T2).
Procedure
First, participants completed the State-Trait Anxiety Inventory questionnaire
[STAI-CH for children; (Spielberger et al., 1983)] to assess both state and trait anxiety.
Then, participants completed the Emotional Identification Task followed by 20
practice trials and 188 test trials of the AB task.
Emotional Identification Task

51
The goal of the task was to assess whether participants recognized the
emotional valence of faces accurately. The task consisted of 6 trials. Participants
began the task by pressing the space bar in response to the words “Get Ready!” on the
screen. Either an angry face or neutral face was presented centrally for 2000 ms,
following which the participant had to press the number “1” key for “angry” or the
number “2” key for “no emotion” in answer to the question “Is this person:”.
Participants could take as long as they needed to record their response. Upon
successful recognition of at least 70% of the emotional faces, they went on to complete
the AB task.
Attentional Blink task
The AB task consisted of 188 trials divided into four blocks of 47 trials each.
Participants pressed the space bar in response to the words “Get Ready!” on the screen
to start each block; they could take a self-paced break in between blocks. Each trial
began with the presentation of a fixation cross for 750 ms followed by a rapid serial
visual presentation (RSVP) stream of 15 stimuli presented for 100 ms each (see Figure
1). Participants were instructed to look for a dog and informed that sometimes a face
would appear and to pay attention because the pictures came on the screen very fast.
Each trial ended with the a question on the screen “Did you see a dog?”. Participants
pressed “1“ for “yes” or “2” for “no” on the number pad of the keyboard. Following
their response, a second question “Did you see a face?” appeared on the screen. Once

52
again, participants indicated their response by using the number pad. For each
question, participants had 5000 ms to respond.
The AB task consisted of eight types of trials, two types of catch trials, and six
types of AB trials varying in the facial expression of the T2 stimulus, at each of three
lags. Specifically, catch trials included: 1) 30- T1-only trials, in which only the T1, a
dog, appeared in the RSVP stream but no T2 appeared; 2) 32- T2-only trials, in which
a T2 (either an angry or neutral face) appeared in the RSVP stream without a T1
preceding it. These trials were included in order to ‘catch’ any response biases to
answering “yes” to both questions at the end of the RSVP stream.
On the remaining AB trials, a T2 was either a face with a neutral or angry
expression and followed the T1 with three lags: 1) 21 trials with a neutral T2
appearing 200 ms after the T1, (termed Neutral-Lag 2); 4) 21 trials with an angry T2
appearing 200 ms after the T1, (termed Angry-Lag 2); 5) 21 trials with a neutral T2
appearing 400 ms after the T1 (termed Neutral-Lag 4); 6) 21 trials with an angry T2
appearing 400 ms after the T1, (termed Angry-Lag 4); 7) 21 trials with a neutral T2
appearing 800 ms after the T1 (termed Neutral-Lag 8); 8) 21 trials with an angry T2
appearing 800 ms after the T1 (termed Angry-Lag 8). Further, within the RSVP
stream, the serial position of T1 was varied among stimuli 4, 5, and 6 in the trial
sequence. Thus, T2 followed each of these positions at the three lags described above
such that it was either the second (Lag 2), fourth (Lag 4), or eighth (Lag 8) stimulus
after the T1. All the trials were pseudorandomized into blocks so that no more than

53
two successive trials were of the same type within each block. Blocks were created to
allow breaks after 47 trials, and were balanced to include the same number of each trial
types. Two versions of the task were created to counterbalance block presentation
order.
Figure 5. Rapid serial visual stream presentation (RSVP) stream for the AB Task.
Participants monitored two target stimuli (T1 & T2) embedded among 13 distractor
stimuli, each presented for 100ms with no interstimulus interval. The first target
stimulus (T1) was a dog face and the second target stimulus (T2) was a human face
(either neutral or angry). The distractors were scrambled faces. a) T1-only catch trials
in which only the dog face was presented, b) T2-only catch trials in which only an
angry or neutral face was presented, c) Double target trials in both T1, the dog face,
and T2, the human face, were presented. The two targets were separated by either 2, 4

54
or 8 distractors, (200ms, 400ms, or 800ms, respectively). At the end of the RSVP
stream, participants indicated by key press whether they had seen saw a dog and a face.
Results
Participants who performed worse than chance (50%) on the T2 catch trial were
excluded from analysis. This criterion resulted in the exclusion of seven children and
two children with ADHD from the total cohort of participants. A response was scored
as correct if the participant correctly identified first the T1 and then the T2, as for T1
incorrect trials the source of T2 errors is unknown (Marvin M. Chun & Potter, 1995).
For each participant, percentage of accurate T2 trials was computed for each of the six
types of AB trials. Percent accuracy was analyzed in a mixed analysis of variance with
Group (Children, Adults) as the between-subjects factor and emotion (T2 - angry,
neutral) and lag (2, 4, 8) as within-subjects factors. Measures of effect size (partial eta-
squared for analyses of variance [ANOVA], Cohen’s d for means) are presented for the
AB effects. Figure 2 summarizes the results by trial type and groups.

55
Figure 6. Mean percentage correct scores as a function of group (Control vs. ADHD),
emotional face of T2 (angry vs. neutral), and lag (2, 4, 8).
We found two main effects. Both groups were more accurate when the T2 was
a face with angry than neutral expression (main effect of emotion), F(1,56) = 36.77, p
< .001, η2=.04. Further, T2 accuracy differed by lag (main effect of lag), F(2,56) =
21.42, p< .001, η2=.08. Post-hoc tests showed that T2 detection at lag 2 was lower
than at lag 4, t(58) = 3.90, p < .001, d= 1.71, and lag 8, t(58), = 6.66, p < .001, d= 1.93.
T2 detection at lag 4 was also lower than lag 8, t(58), = 3.89, p < .001, d= 0.98. These

56
differences by lag indicate that our adapted paradigm produced the expected AB
phenomenon, decreased accuracy at the shortest lag relative to longer lags.
In addition to these main effects, one key interaction was observed, a
significant Lag X Emotion interaction, F(2,56) = 3.51, p<.001, η2=.01, indicating that
the magnitude of the AB differed by the nature of the T2. Collapsing across groups,
paired t-tests showed a higher T2 accuracy for angry trials relative to neutral trials at
each lag: lag 2, t(58), = 4.89, p < .001, d= 1.40, lag 4, t(58)= 2.97, p < .005, d= 0.72 ,
and lag 8, t(58)= 2.82, p < .01, d= 0.79. Among the angry trials, there was an
improvement in T2 detection from lag 2 to lag 8, t(58)= 3.97, p < .001, d= 1.31 , and
another from lag 4 and lag 8, t(58)= 3.54, p < .001, d= 1.13. Lag 2 and lag 4 did not
show an improvement for the angry faces, t(58)= 1.59, p = .12, d= 0.46. Among the
neutral trials, all the lags were different from each other: lag 2 and lag 4, t(58), = 4.10,
p < .001, d= 1.10; lag 4 and lag 8, t(58), = 6.06, p < .001, d= 0.81, and lag 2 and lag 8,
t(58), = 2.81, p < .01, d= 2.01, showing improved T2 detection with increasing lag
length. This finding was further substantiated when a difference score was calculated
to assess the magnitude of the AB, (i.e. lag 8 performance minus lag 2) which was
significantly larger on neutral trials than on angry trials, t(58), = 2.49, p < .05, d= 0.94.
This finding indicates that we observed the expected effect of emotional stimuli on the
AB phenomenon, a shorter blink for angry than neutral faces.

57
Most importantly, the Group X Lag X Emotion interaction was not significant,
F(2,56) = 0.72, p =.49, η2=.001 indicating that the effect of the facial expression of the
T2 stimulus did not influence the magnitude of the AB differently in both groups.
We examined whether the magnitude of the AB was associated with anxiety. In
order to assess the magnitude of the AB, the neutral face performance (T2 accuracy)
from lag 2 was subtracted from lag 8. In order to assess the influence of emotion in
early attentional processing, neutral face performance (T2 accuracy) for lag 2 was
subtracted from angry face performance at lag 2.
Trait anxiety, as measured with the STAI-C, was higher in children with
ADHD (mean = 37.6, SD = 5.69) relative to control children (mean = 31.6, SD = 5.14,
t(41)= 3.99 p < .001, d= 1.11); no child met the criteria for the presence of a clinical
anxiety disorder. A negative correlation between the magnitude of the AB and trait
anxiety level was shown across control and ADHD children combined, such that more
anxious children had a smaller AB (r= -0.24, p < .05). Examining the groups
separately, trait anxiety correlated significantly with the magnitude of the AB in
control children, (r= - .41, p <.05), but not ADHD children, (r= - .04, p =.43).
Therefore, the correlation in the full group was driven by the control children. Results
are shown in Figure 3. There were no correlations between trait anxiety and the
influence of emotion on the AB across both groups, (r= -.13, p =.18), or within each
group, (ADHD: r= .05, p=.40; Controls: r = -.25, p=.09).

58
Figure 7. Correlation between the AB Size with Neutral Faces and Trait Anxiety based
on STAI scores
No significant correlations were found with state anxiety and the magnitude of
the AB across both groups (r= .16, p =.13) or within ADHD (r= .30, p=.09) or
Controls: r = .06, p=.38. No significant correlations were found between state anxiety
and the influence of emotion on the AB across both groups (r= .11, p =.21) or within
ADHD (r= .25, p=.13) or Controls (r = .01, p=.48).
Lastly, we investigated whether the magnitude of the AB or its modulation with
emotion was associated with symptoms of hyperactivity/impulsivity and inattention

59
measured with the ADHD Rating Scale. A positive correlation was found between
symptoms of inattention and the size of the emotional AB (r= 0.38, p < .05) in children
with ADHD, such that the more inattentive children benefitted more from an angry
face at lag 2 relative to a neutral face. Results are shown in Figure 4. This finding was
exclusive to the ADHD group since it did not appear in control children (r= - .25, p
=.12) and across both groups (r= .02, p =.43). The magnitude of the AB did not
correlate with inattention symptoms, (r= - .14, p =.17). Further, symptoms of
hyperactivity/impulsivity did not show association with either the magnitude of the AB
(r= - .11, p =.23) or the influence of emotion on the AB (r= - .02, p =.43) across both
groups. Neither group alone showed a relationship with hyperactive/impulsivity
symptoms for the magnitude of the AB, (ADHD: r= .09, p =.35,; Controls: r= - .18, p
=.19), and influence of emotion on the AB (ADHD: r= .35, p =.05; Controls: r= - .09, p
=.35).

60
Figure 8. Correlation between the Influence of Emotion on Early Attentional
Processing and Inattention symptoms as measured by the ADHD Rating Scale.
Discussion
We assessed early attentional processing and its modulation by emotional
stimuli in children with ADHD and age and IQ matched controls using the AB
paradigm. The magnitude of the AB was similar in both groups with neutral faces.
Both groups also showed a similar reduction of the AB with angry faces. Thus, early

61
attentional processing was preserved for socio-emotional stimuli in ADHD. The
magnitude of the AB varied by individual differences in anxiety such that children who
were more anxious had a smaller AB with neutral faces. The influence of emotion on
the AB effect varied by individual differences in symptoms of inattention among
children with ADHD such that more inattentive children showed a larger effect of
angry faces on the AB. Together, these findings further the understanding of early
attentional processing in ADHD and typical development.
The present study extends our knowledge of early attention processing in
ADHD in four ways. First, our study is the first to examine the AB effect for socio-
emotional stimuli in ADHD. Prior to this study, the AB had been investigated with
non-social stimuli such as letters (Carr et al., 2006), letters with asterisks, (Mason et
al., 2005), and numbers among Chinese characters (Li et al., 2004), with mixed results.
Our study demonstrates that the AB effect with socio-emotional information is similar
in children with ADHD and controls.
Second, we demonstrate preserved AB effect for both neutral and emotional
faces in both controls and ADHD. One probable explanation for the absence of
differences between groups with our study is our task design. A bottleneck effect
normally occurs at Stage 2 because it cannot process additional items from Stage 1
until it is done processing (Arnell, Helion, Hurdelbrink, & Pasieka, 2004; Chun &
Potter, 1995; Jolicoeur, 1998; Martens & Wyble, 2010). When the first target can be
more efficiently processed because its uniqueness amongst distractors, a smaller AB is

62
seen because there is no bottleneck effect (Arnell et al., 2006). Thus, the more unique
a target is among the distractors, like in our study that required a dog and human face
to be detected among scrambled faces, the easier and thus faster processing that occurs
at both Stage 1 and 2, which results in increased T2 detection at earlier lags (Chun,
1997; Chun & Potter, 1995). We see how the increased contrast between targets and
non-targets improved performance, since our control children had better T2 detection,
above 70% for neutral faces and above 85% for angry faces at lag 2, than previous
non-emotional AB studies that have performance for lag 2 between 50% and 60%,
(Carr, Henderson, & Nigg, 2010; Mason et al., 2005). In conclusion, out task design
may have facilitated early attentional processing of both neutral and emotional stimuli
in both controls and ADHD.
An alternative explanation for our lack of group differences in the emotional
AB arises from the networks responsible for processing emotional information.
Preferential emotion processing is believed to be dependent on the amygdala, since
patients with amygdala lesions do not show a reduction in the AB when emotional
words are presented as the second target (Anderson & Phelps, 2001). The amygdala is
part of the mesolimbic network involved in emotional processing (Sonuga-Barke,
Bitsakou, & Thompson, 2010) and has been highlighted recently in ADHD research.
Increased amygdala activation has been found during making judgments regarding
nose width and perceived hostility of a face. The altered amygdala functioning may
relate to later stages of emotional attention processing since the task required an

63
evaluative judgement (Brotman et al., 2010). In our study, we only required detection
of a face without any evaluation of valence. We can only infer from our study that
behaviorally, amygdala functioning is intact in children and ADHD during early
attentional processing.
Third, our study shows a reduced AB with neutral faces in more anxious
control children. This finding is different from previous studies revealing individuals
with higher anxiety levels have a smaller AB with emotional faces (de Jong &
Martens, 2007; B. De Martino et al., 2008; Fox et al., 2005). We propose that the
neutral faces were perceived as ambiguous, and thus, those who were more anxious
perceived them more negatively and were more vigilant towards these faces.
Emerging literature suggests that anxious individuals process ambiguous information
differently (Becker & Detweiler-Bedell, 2009; Heim-Dreger, Kohlmann, Eschenbeck,
& Burkhardt, 2006; MacLeod & Cohen, 1993; Stopa, 2000). Anxious children appear
to perceive ambiguity as threatening and therefore avoid any ambiguous information
(A. P. Field & Lester, 2010). Faces with neutral expressions are perceived as
ambiguous in children (for review(Leppanen & Nelson, 2009). It maybe the case that
in early stages of attention processing children are more vigilant towards ambiguous
stimuli and when assessed in later stages of attention, they actually display the
avoidant bias traditionally seen in emotion studies (Perez-Edgar et al., 2010). Children
with ADHD despite reporting higher levels of trait anxiety did not show the same
phenomenon as controls. Controls’ anxiety scores seem more evenly distributed across

64
the full range while children with ADHD’s range is smaller if the outliers are dropped,
as seen in Figure 3. Therefore in ADHD, the absence of correlation between anxiety
and the magnitude of the attentional blink is the result of a reduced range of scores.
Fourth, children with ADHD who are more inattentive had a larger effect of
emotion on the AB (i.e., higher accuracy for detecting angry relative to neutral faces
immediately after detection of T1). The inattentive subtype is believed to have a
defective attentional filter (gate) and therefore, poor interference suppression
(Diamond, 2005). In our study, children with ADHD with more inattentive symptoms
benefitted the most from an emotional T2. Emotional stimuli are more salient and
therefore, may have been more difficult to suppress for those with more severe
inattention. This is inline with a previous study indicating that inattention in ADHD is
related more to early-stage information control (attention-gating) instead of response
inhibition (Carr et al., 2006). There are neuroimaging studies suggesting that the
deficit in attention filtering seen in the inattentive subtype is related to a dysfunction in
fronto-parietal cortical attention networks, while the combined and
hyperactive/impulsive subtypes possess late stage control deficits subserved by frontal-
subcortical circuits related to executive control (Adams, Derefinko, Milich, &
Fillmore, 2008; Diamond, 2005; Milich, Balentine, & Lynam, 2006).
It is important to note the limitations of our study. First, ADHD is a
heterogenous disorder comprised of different subtypes and it maybe important to study
more closely subtype differences especially in lieu of our finding that inattentive

65
symptoms relate to the emotional AB effect and the previous inattentive subtype study
that only found a difference in the AB effect among the inattentive subtype (Carr et al.,
2006). Second, our task was manipulated to enhance child engagement and
subsequently performance. It is possible that by making our task too child-friendly, we
lost the ability to detect more subtle differences between controls and ADHD. Lastly,
we only investigate negative and neutral faces, it maybe the case that positive emotions
may should a greater AB since there is evidence that positive emotions are not as
salient as negative emotions in ADHD (Becker et al., 1993; Conzelmann et al., 2009).
In summary, early visual attention processes were sensitive to socio-emotional
information in children with ADHD. Furthermore, both anxiety and inattention
influence early visual attention processes, anxiety reduces the overall magnitude of the
blink while in ADHD inattention symptoms were related to greater sensitivity to angry
faces.

66
CHAPTER IV: GENERAL DISCUSSION
This dissertation presented results that extend the understanding of the role of
socio-emotional information in early visual attention processing in typical development
and in a developmental disorder, Attention Deficit/Hyperactivity Disorder (ADHD).
Chapter 2 and 3 reported the results from studies, which are the first to use the
attentional blink (AB) paradigm to examine early visual attention to human faces with
neutral and negative emotional expression. We found two novel findings: First,
independent of emotional expression, typically developing children showed a larger
attentional blink than adults, indicating reduced attentional resources in early visual
processing. However, the extent to which early visual attention is influenced by
emotionally salient information did not differ between typically developing children
and adults as there were no developmental differences in the attentional blink effect for
angry faces. This finding suggests that processing of emotional stimuli as salient is
mature in late childhood. Second, early visual attention to neutral faces and to
emotionally salient information was intact in ADHD as children with ADHD showed a
similar magnitude of the attentional blink for neutral and angry faces as age and IQ
matched control children. Therefore, attentional deficits observed in ADHD do not
appear to stem from early visual attention, at least, for socio-emotional information.
Anxiety was found to enhance early attention processing in typically
developing children. Prior adult work had shown a reduced AB effect among anxious
individuals with emotional words, faces and scenes (Jefferies et al., 2008; Maratos et

67
al., 2008; Tibboel, Van Bockstaele, & De Houwer, 2011). Our study did not replicate
the reduced AB effect with faces in anxious adults but did find a reduced AB effect
using neutral faces in more anxious children. Neutral faces are high priority
information as there is evidence that they are perceived as ambiguous and therefore
negative for anxious individuals (MacLeod & Cohen, 1993; Leppanen & Nelson,
2009). In this dissertation we suggest that vigilant bias is present among anxious
children for neutral (ambiguous) faces and it would disappear as the brain matures into
adulthood. The perception of the neutral faces as negative among the anxious children
reduced the bottlenecking in attentional processing because they were considered to be
highly important information to process.
In children with ADHD, symptoms of inattention were related to early
attention processing of emotional information. Inattentive subtype of ADHD had been
characterized as showing a lack of the AB effect because of an inefficient attentional
filter (Carr, Henderson, & Nigg, 2010). We show that in the presence of high priority
information like faces, the AB effect is not only intact but in the early lags (early stages
of attentional processing) angry faces reduce it. Our study suggests that the attentional
filter is aided possibly by a subcortical structure like the amygdala to normalize the AB
effect and reduces the bottlenecking effect at the early lags when a high priority
stimulus like an emotional face is presented. Collectively from both studies, individual
differences in anxiety and inattention can influence early attentional processing in the
presence of socio-emotional information.

68
Neural substrates of the Attentional Blink effect
Attentional processing has been shown to be mediated via a dorsal and ventral
pathway. The AB task has furthered our understanding of early attention processing to
include more specific neural substrates. A distributed network across the occipital,
parietal and frontal areas is implicated in the non-emotional AB effect (Martens &
Wyble, 2010). Early visual areas like V1 are activated by both T1 and T2 (Hein,
Alink, Kleinschmidt, & Müller, 2009). When T2 is not reported, there is reduced
activity in the fronto-parietal cortex, specifically the anterior cingulate, lateral
prefrontal, and parietal regions (Kranczioch, Debener, Schwarzbach, Goebel, & Engel,
2005; Marois & Ivanoff, 2005; Marois, Yi, & Chun, 2004). The lateral frontal and
parietal regions are believed to be the neural substrates for the AB bottleneck (Marois
& Ivanoff, 2005) and under top-down control. Our findings from Chapter II suggest
selection for high priority information in early attention processing is critical for
humans therefore a mechanism is established early in childhood in order to detect
threatening or important information in the environment.
The emotional AB recruits other neural substrates than the nonemotional AB.
When faces are used in the emotional AB, the fusiform face area (FFA) is more
activated for correct identification of a face (B. De Martino et al., 2008). The
recruitment of the FFA was related more to the presence of a face in T2 than to
whether the face had a positive or negative emotion. Interestingly the ventral striatum
is more active when T2 is a fearful face than with a neutral face T2, consistent with

69
previous research showing ventral striatum activation during aversive or unpleasant
visual stimuli (Paradiso et al., 1999). When attentional resources are limited, rostral
anterior cingulate cortex (rACC) shows increased activation when T2 is an emotional
face (B. De Martino et al., 2008). The rACC activation correlated with successful
identification of T2. Interesting to note is that the rACC has been shown previously to
have reciprocal connections with areas of emotion processing (ventral striatum,
anterior insula, orbitofrontal cortex, and amygdala) that are also part of the ventral
pathway of attention processing (Bush, Luu, & Posner, 2000; B. De Martino et al.,
2008). Therefore, the rACC is an important structure in the selection of high priority
emotional information under limited attentional resources (B. De Martino et al., 2008).
One critical structure in the interaction of attention and emotion is the
amygdala (Phelps, 2006). The amygdala is strategically positioned in the brain to
communicate with various structures and thus has influence on various stages and
types of processing. The amygdala has reciprocal connections with sensory cortical
processing areas, like the visual cortex (Amaral, Behniea, & Kelly, 2003). Because of
its location it is able to receive emotional information input before gaining awareness
of the emotional information (Romanski & LeDoux, 1992; Whalen, 1998). In ADHD
the amygdala is also a key structure in the mesolimbic pathway responsible for
emotional and motivational dysfunctioning (E. J. S. Sonuga-Barke, 2003). Early
attentional processing requires the amygdala (Anderson & Phelps, 2001) to be intact
for the attentional capture of emotion to occur. From our findings, we suspect that the

70
amygdala is working efficiently to detect emotional information in the environment
quickly. Both emotional influences from the amygdala and attentional influences from
fronto-parietal areas appear to act to amplify emotion or task-relevant information in a
stimulus-specific manner, producing similar increases in fMRI and EEG responses
(Lang et al., 1998; Sabatinelli, Lang, Bradley, Costa, & Keil, 2009). Emotion and
attention effects have distinct sources, they may occur in a parallel or competitive
manner and produce additive (or occasionally interactive) effects on sensory responses
(Brosch, Pourtois, Sander, & Vuilleumier, 2011; Keil, Moratti, Sabatinelli, Bradley, &
Lang, 2005; Pourtois, Schettino, & Vuilleumier, 2012; Rossi & Pourtois, 2012; P
Vuilleumier & Schwartz, 2001). Therefore in our emotional AB effect with adults,
children and ADHD we see an enhanced bottom-up signal that facilitates detection of
emotional information in very early stages of attention processing. Therefore, we
continue to confirm behaviorally an important status for emotional information in early
attention processing.
Future Directions
Of central importance to this dissertation was the behavioral investigation of
the early attentional processing of socio-emotional information. A logical next step is
to investigate the neural mechanism of how socio-emotional information is processed
in early attentional stages. By investigating the neural mechanism, we can enhance our
understanding of brain development by shedding light on how early attentional

71
processing may be an immature process in childhood and evidenced as such by a
distributed network of regions. We can also continue to provide evidence for the
emotional dysfunction seen in ADHD. There are several pieces of evidence that the
amygdala is not normal in ADHD, however our behavioral study in Chapter III would
suggest otherwise. For normal behavioral functioning of the amygdala in ADHD,
certain neural compensations may be made such as the development of a
volumetrically smaller amygdala when compared to controls (Frodl & Skokauskas,
2012) or the hyper activation of the amygdala when rating subjective fear of neutral
faces (Brotman et al., 2010)).
To address the notion that salient information has a preferred status of
attentional processing that utilizes a ventral network of attention, a direct investigation
of non-social and social-emotional stimuli in early attentional processing should be
conducted. There is behavioral evidence emotional value is not the sole contributor to
an enhanced early attentional processing, but aspects of arousal and salience can
produce similar results (Anderson, 2005). Therefore, bottom-up processes that utilize
the ventral network of attention may in fact not be coding for emotion but instead a
more global feature like arousal or salience.
Another potential avenue for future research is to investigate the role of
genetics in early attentional processing. Serotonin (5-hydroxytrptamine; 5-HT), is a
monoamine neurotransmitter synthesized in the raphe nucleus and released all through
out the brain (N. M. Barnes & Sharp, 1999). The serotonin transporter protein (5-

72
HTT) is located on the pre-synaptic terminal, and is responsible terminating the action
of 5-HT. A polymorphism in the serotonin transporter long promoter region, 5-
HTTLPR, of the serotonin transporter gene (SLC6A4) influences 5-HTT mRNA
transcription. A short “S” allele is linked with reduced expression of the serotonin
transporter mRNA relative to the long “l” allele (Hu et al., 2006). Given that
polymorphisms in the serotonin transporter gene have been found to contribute to
individual differences in emotional reactivity, it is likely that the individual differences
we see in anxiety may have its origin in the genetic expression of the serotonin
transporter gene. Expression of the S allele is associated with heightened emotional
reactivity (Pérez-Edgar et al., 2010), higher scores of anxiety-related behaviors (Lesch
et al., 1996), and increased amygdala activation to more negative stimuli (Munafò,
Brown, & Hariri, 2008). Therefore the serotonin transporter gene may help
differentiate between those individuals who are more susceptible to socio-emotional
information in earlier lags of the attentional blink. Another gene of interest would be
the catechol-O-methyltransferase (COMT) gene. COMT is the enzyme that degrades
catecholamines like dopamine in the synapse. A single nucleotide polymorphism
(SNP) in the COMT gene, Val158Met, which substitutes a methionine for valine,
results in lower levels of COMT in the synapse, which in turn leads to less dopamine
breakdown. Individuals with two inherited copies of the met allele (met/met) have less
COMT and more dopamine functioning in the synapse compared to those with two
copies of the val allele (val/val) (Chen et al., 2004). Children and adults who are

73
homozygous for the met allele tend to have better performance on attentional tasks
while also possessing reduced control for emotionally salient stimuli (Blasi et al.,
2005; Diamond, Briand, Fossella, & Gehlbach, 2004). Therefore by investigating the
role of the COMT gene, we can determine if dopamine has a role in the gating
mechanism of the attentional filter in ADHD in early attentional processing. Therefore
genetic studies of early attentional processing should be conducted.
CONCLUSION
Emotional information has the ability to capture attention quickly. We know
that emotion can draw our attention through a ventral pathway of attention that utilizes
bottom-up processing. The amygdala appears to be the center for the attentional
capture of emotion because of its location in the brain and its reciprocal connections
with sensory cortices and prefrontal cortices. The AB paradigm has allowed us to
gauge what and how emotion captures attention. Through the use of the AB we can
continue to further understand the mechanism by which emotion and other salient
information captures our attention.
To our knowledge, these are the first studies to utilize an AB with neutral and
angry faces to assess the early visual attention processing across development and in
ADHD. We found that independent of the emotional expression of a face, children
show a larger AB than adults. We also found that children with ADHD have an intact
AB like control children with both angry and neutral faces. We also found that

74
individual differences can influence early attention processing. Based on our findings,
more research should investigate the development of the AB in children and its neural
substrates. Individual differences in gene expressions should be also investigated to
further understand how one’s genetic code can make one more sensitive to emotional
information.

75
REFERENCES
Adolphs, R. (2003). Cognitive neuroscience of human social behaviour. Nature
Reviews. Neuroscience, 4(3), 165–178. doi:10.1038/nrn1056
Amaral, D. G., Behniea, H., & Kelly, J. L. (2003). Topographic organization of
projections from the amygdala to the visual cortex in the macaque monkey.
Neuroscience, 118(4), 1099–1120.
Amir, N., Foa, E. B., & Coles, M. E. (1998). Automatic activation and strategic
avoidance of threat-relevant information in social phobia. Journal of Abnormal
Psychology, 107(2), 285–290. doi:10.1037/0021-843X.107.2.285
Amir, N., Taylor, C. T., Bomyea, J. A., & Badour, C. L. (2009). Temporal allocation
of attention toward threat in individuals with posttraumatic stress symptoms.
Journal of Anxiety Disorders, 23(8), 1080–1085. doi:doi: DOI:
10.1016/j.janxdis.2009.07.010
Anastopoulos, A. D., Smith, T. F., Garrett, M. E., Morrissey-Kane, E., Schatz, N. K.,
Sommer, J. L., Kollins, S. H., et al. (2010). Self-Regulation of Emotion,
Functional Impairment, and Comorbidity Among ChildrenWith AD/HD.
Journal of Attention Disorders, 15(7), 583–592.
doi:10.1177/1087054710370567

76
Anderson, A. K. (2005). Affective Influences on the Attentional Dynamics Supporting
Awareness. Journal of Experimental Psychology: General, 134(2), 258–281.
doi:10.1037/0096-3445.134.2.258
Anderson, A. K., & Phelps, E. A. (2001). Lesions of the human amygdala impair
enhanced perception of emotionally salient events. Nature, 411(6835), 305–
309. doi:10.1038/35077083
Armstrong, I. T., & Munoz, D. P. (2003). Attentional blink in adults with attention-
deficit hyperactivity disorder. Experimental Brain Research, 152(2), 243–250.
doi:10.1007/s00221-003-1535-0
Arnell, K. M., Helion, A. M., Hurdelbrink, J. A., & Pasieka, B. (2004). Dissociating
sources of dual-task interference using human electrophysiology. Psychonomic
Bulletin & Review, 11(1), 77–83.
Arnell, K. M., Howe, A. E., Joanisse, M. F., & Klein, R. M. (2006). Relationships
between attentional blink magnitude, RSVP target accuracy, and performance
on other cognitive tasks. Memory & Cognition, 34(7), 1472–1483.
Barkley, R. A. (1997). Behavioral inhibition, sustained attention, and executive
functions: constructing a unifying theory of ADHD. Psychological bulletin,
121(1), 65–94.
Barnes, K. A., Kaplan, L. A., & Vaidya, C. J. (2007). Developmental differences in
cognitive control of socio-affective processing. Developmental
Neuropsychology, 32(3), 787–807. doi:10.1080/87565640701539576

77
Barnes, N. M., & Sharp, T. (1999). A review of central 5-HT receptors and their
function. Neuropharmacology, 38(8), 1083–1152.
Batty, M., & Taylor, M. J. (2006). The development of emotional face processing
during childhood. Developmental Science, 9(2), 207–220. doi:10.1111/j.1467-
7687.2006.00480.x
Becker, D. F., Doane, J. A., & Wexler, B. E. (1993). Effects of emotion on perceptual
asymmetry in adolescent inpatients with attention-deficit hyperactivity
disorder. Journal of the American Academy of Child and Adolescent
Psychiatry, 32(2), 318–321; discussion 322–323. doi:10.1097/00004583-
199303000-00012
Becker, M., & Detweiler-Bedell, B. (2009). Early detection and avoidance of
threatening faces during passive viewing. The Quarterly Journal of
Experimental Psychology, 62(7), 1257–1264. doi:10.1080/17470210902725753
Biederman, J., Faraone, S., Milberger, S., Guite, J., Mick, E., Chen, L., Mennin, D., et
al. (1996). A prospective 4-year follow-up study of attention-deficit
hyperactivity and related disorders. Archives of general psychiatry, 53(5), 437–
446.
Blagrove, E., & Watson, D. G. (2010). Visual Marking and Facial Affect: Can an
Emotional Face Be Ignored? Emotion, 10(2), 147–168. doi:doi: DOI:
10.1037/a0017743

78
Blasi, G., Hariri, A. R., Alce, G., Taurisano, P., Sambataro, F., Das, S., Bertolino, A.,
et al. (2009). Preferential Amygdala Reactivity to the Negative Assessment of
Neutral Faces. Biological Psychiatry, 66(9), 847–853. doi:doi: DOI:
10.1016/j.biopsych.2009.06.017
Blasi, G., Mattay, V. S., Bertolino, A., Elvevåg, B., Callicott, J. H., Das, S., Kolachana,
B. S., et al. (2005). Effect of catechol-O-methyltransferase val158met genotype
on attentional control. The Journal of neuroscience: the official journal of the
Society for Neuroscience, 25(20), 5038–5045. doi:10.1523/JNEUROSCI.0476-
05.2005
Bradley, B. P., Mogg, K., White, J., Groom, C., & de Bono, J. (1999). Attentional bias
for emotional faces in generalized anxiety disorder. The British Journal of
Clinical Psychology / the British Psychological Society, 38 ( Pt 3), 267–278.
Broadbent, D., & Broadbent, M. H. (1990). Human attention: the exclusion of
distracting information as a function of real and apparent separation of relevant
and irrelevant events. Proceedings. Biological sciences / The Royal Society,
242(1303), 11–16. doi:10.1098/rspb.1990.0096
Broadbent, D. E., & Broadbent, M. H. (1987). From detection to identification:
response to multiple targets in rapid serial visual presentation. Perception &
Psychophysics, 42(2), 105–113.
Brosch, T., Pourtois, G., Sander, D., & Vuilleumier, P. (2011). Additive effects of
emotional, endogenous, and exogenous attention: behavioral and

79
electrophysiological evidence. Neuropsychologia, 49(7), 1779–1787.
doi:10.1016/j.neuropsychologia.2011.02.056
Brotman, M. A., Rich, B. A., Guyer, A. E., Lunsford, J. R., Horsey, S. E., Reising, M.
M., Thomas, L. A., et al. (2010). Amygdala activation during emotion
processing of neutral faces in children with severe mood dysregulation versus
ADHD or bipolar disorder. The American Journal of Psychiatry, 167(1), 61–
69. doi:10.1176/appi.ajp.2009.09010043
Brown, T. E. (2009). ADHD comorbidities : handbook for ADHD complications in
children and adults. Washington, D.C: American Psychiatric Publishing.
Bush, Luu, & Posner. (2000). Cognitive and emotional influences in anterior cingulate
cortex. Trends in cognitive sciences, 4(6), 215–222.
Carr, L. A., Henderson, J. M., & Nigg, J. T. (2006). Attentional Versus Motor
Inhibition in Adults With Attention-Deficit/Hyperactivity Disorder.
Neuropsychology, 20(4), 430–441. doi:doi: DOI: 10.1037/0894-4105.20.4.430
Carr, L., Henderson, J., & Nigg, J. T. (2010). Cognitive Control and Attentional
Selection in Adolescents with ADHD Versus ADD. Journal of Clinical Child
& Adolescent Psychology, 39(6), 726–740. doi:10.1080/15374416.2010.517168
Castel, A. D., Lee, S. S., Humphreys, K. L., & Moore, A. N. (2011). Memory capacity,
selective control, and value-directed remembering in children with and without
attention-deficit/hyperactivity disorder (ADHD). Neuropsychology, 25(1), 15–
24. doi:10.1037/a0020298

80
Chen, J., Lipska, B. K., Halim, N., Ma, Q. D., Matsumoto, M., Melhem, S., Kolachana,
B. S., et al. (2004). Functional analysis of genetic variation in catechol-O-
methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in
postmortem human brain. American journal of human genetics, 75(5), 807–
821. doi:10.1086/425589
Chhabildas, N., Pennington, B. F., & Willcutt, E. G. (2001). A comparison of the
neuropsychological profiles of the DSM-IV subtypes of ADHD. Journal of
Abnormal Child Psychology, 29(6), 529–540.
Chun, M M. (1997). Temporal binding errors are redistributed by the attentional blink.
Perception & Psychophysics, 59(8), 1191–1199.
Chun, Marvin M., & Potter, M. C. (1995). A two-stage model for multiple target
detection in rapid serial visual presentation. Journal of Experimental
Psychology: Human Perception and Performance, 21(1), 109–127.
doi:10.1037/0096-1523.21.1.109
Cohen Kadosh, K., & Johnson, M. H. (2007). Developing a cortex specialized for face
perception. Trends in Cognitive Sciences, 11(9), 367–369. doi:doi: DOI:
10.1016/j.tics.2007.06.007
Colzato, L. S., Slagter, H. A., Spapé, M. M. A., & Hommel, B. (2008). Blinks of the
eye predict blinks of the mind. Neuropsychologia, 46, 3179–3183.
doi:10.1016/j.neuropsychologia.2008.07.006

81
Conzelmann, A., Mucha, R. F., Jacob, C. P., Weyers, P., Romanos, J., Gerdes, A. B.
M., Baehne, C. G., et al. (2009). Abnormal affective responsiveness in
attention-deficit/hyperactivity disorder: subtype differences. Biological
Psychiatry, 65(7), 578–585. doi:10.1016/j.biopsych.2008.10.038
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The Reorienting System of the
Human Brain: From Environment to Theory of Mind. Neuron, 58(3), 306–324.
doi:10.1016/j.neuron.2008.04.017
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven
attention in the brain. Nat Rev Neurosci, 3(3), 201–215. doi:10.1038/nrn755
Cubillo, A., Halari, R., Smith, A., Taylor, E., & Rubia, K. (2012). A review of fronto-
striatal and fronto-cortical brain abnormalities in children and adults with
Attention Deficit Hyperactivity Disorder (ADHD) and new evidence for
dysfunction in adults with ADHD during motivation and attention. Cortex; a
Journal Devoted to the Study of the Nervous System and Behavior, 48(2), 194–
215. doi:10.1016/j.cortex.2011.04.007
Darque, A., Del Zotto, M., Khateb, A., & Pegna, A. J. (2012). Attentional modulation
of early ERP components in response to faces: evidence from the attentional
blink paradigm. Brain topography, 25(2), 167–181. doi:10.1007/s10548-011-
0199-5
de Haan, M., Belsky, J., Reid, V., Volein, A., & Johnson, M. H. (2004). Maternal
personality and infants’ neural and visual responsivity to facial expressions of

82
emotion. Journal of child psychology and psychiatry, and allied disciplines,
45(7), 1209–1218. doi:10.1111/j.1469-7610.2004.00320.x
de Jong, P. J., & Martens, S. (2007). Detection of emotional expressions in rapidly
changing facial displays in high- and low-socially anxious women. Behaviour
Research and Therapy, 45(6), 1285–1294. doi:doi: DOI:
10.1016/j.brat.2006.10.003
De Martino, B., Kalisch, R., Rees, G., & Dolan, R. J. (2008). Enhanced Processing of
Threat Stimuli under Limited Attentional Resources. Cerebral Cortex, 19(1),
127–133. doi:10.1093/cercor/bhn062
De Martino, Benedetto, Strange, B. A., & Dolan, R. J. (2007). Noradrenergic
neuromodulation of human attention for emotional and neutral stimuli.
Psychopharmacology, 197(1), 127–136. doi:10.1007/s00213-007-1015-5
Diamond, A. (2005). Attention-deficit disorder (attention-deficit/ hyperactivity
disorder without hyperactivity): A neurobiologically and behaviorally distinct
disorder from attention-deficit/hyperactivity disorder (with hyperactivity).
Development and Psychopathology, 17(03). doi:10.1017/S0954579405050388
Diamond, A., Briand, L., Fossella, J., & Gehlbach, L. (2004). Genetic and
neurochemical modulation of prefrontal cognitive functions in children. The
American journal of psychiatry, 161(1), 125–132.

83
Dunbar, R. I. M. (2009). The social brain hypothesis and its implications for social
evolution. Annals of Human Biology, 36(5), 562–572.
doi:10.1080/03014460902960289
DuPaul, G. (1998). ADHD rating scale-IV : checklists, norms, and clinical
interpretation. New York: Guilford Press.
Ekman, P., & Friesen, W. V. (1971). Constants across cultures in the face and emotion.
Journal of personality and social psychology, 17(2), 124–129.
Ekman, P., & Oster, H. (1979). Facial Expressions of Emotion. Annual Review of
Psychology, 30(1), 527–554. doi:10.1146/annurev.ps.30.020179.002523
Field, A. P., & Lester, K. J. (2010). Is There Room for “Development” in
Developmental Models of Information Processing Biases to Threat in Children
and Adolescents? Clinical Child and Family Psychology Review, 13(4), 315–
332. doi:10.1007/s10567-010-0078-8
Field, T. M., Woodson, R., Greenberg, R., & Cohen, D. (1982). Discrimination and
imitation of facial expression by neonates. Science (New York, N.Y.),
218(4568), 179–181.
Fox, E., Russo, R., & Georgiou, G. A. (2005). Anxiety modulates the degree of
attentive resources required to process emotional faces. Cognitive, Affective, &
Behavioral Neuroscience, 5(4), 396–404. doi:10.3758/CABN.5.4.396

84
Friedman, S. R., Rapport, L. J., Lumley, M., Tzelepis, A., VanVoorhis, A., Stettner, L.,
& Kakaati, L. (2003). Aspects of social and emotional competence in adult
attention-deficit/hyperactivity disorder. Neuropsychology, 17(1), 50–58.
Frodl, T., & Skokauskas, N. (2012). Meta-analysis of structural MRI studies in
children and adults with attention deficit hyperactivity disorder indicates
treatment effects. Acta Psychiatrica Scandinavica, 125(2), 114–126.
doi:10.1111/j.1600-0447.2011.01786.x
Garrad-Cole, F., Shapiro, K. L., & Thierry, G. (2011). Developmental Aspects of
Temporal and Spatial Visual Attention: Insights from the Attentional Blink and
Visual Search Tasks. Child Neuropsychology, 17, 118–137.
doi:10.1080/09297049.2010.509716
Geurts, H. M., Luman, M., & van Meel, C. S. (2008). What’s in a game: the effect of
social motivation on interference control in boys with ADHD and autism
spectrum disorders. Journal of Child Psychology and Psychiatry, and Allied
Disciplines, 49(8), 848–857. doi:10.1111/j.1469-7610.2008.01916.x
Gomez, R. (2003). Underlying processes in the poor response inhibition of children
with Attention-Deficit/Hyperactivity Disorder. Journal of Attention Disorders,
6(3), 111–122.
Grossmann, T., & Johnson, M. H. (2007). The development of the social brain in
human infancy. European Journal of Neuroscience, 25(4), 909–919.
doi:10.1111/j.1460-9568.2007.05379.x

85
Guevremont, D., & Dumas, M. (1994). Peer relationship problems and disruptive
behavior disorders. Journal of Emotional and Behavioral Disorders, 2(3), 164–
173.
Hadwin, J. A., Donnelly, N., French, C. C., Richards, A., Watts, A., & Daley, D.
(2003). The influence of children’s self-report trait anxiety and depression on
visual search for emotional faces. Journal of Child Psychology and Psychiatry,
44(3), 432–444. doi:10.1111/1469-7610.00133
Hansen, C. H., & Hansen, R. D. (1988). Finding the Face in the Crowd: An Anger
Superiority Effect. Journal of Personality and Social Psychology, 54(6), 917–
924. doi:doi:
Hariri, A. R. (2002). Serotonin Transporter Genetic Variation and the Response of the
Human Amygdala. Science, 297(5580), 400–403. doi:10.1126/science.1071829
Hariri, A. R., Bookheimer, S. Y., & Mazziotta, J. C. (2000). Modulating emotional
responses: effects of a neocortical network on the limbic system. Neuroreport,
11(1), 43–48.
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The distributed human neural
system for face perception. Trends in Cognitive Sciences, 4(6), 223–233.
doi:10.1016/S1364-6613(00)01482-0
Heim-Dreger, U., Kohlmann, C.-W., Eschenbeck, H., & Burkhardt, U. (2006).
Attentional Biases for Threatening Faces in Children: Vigilant and Avoidant
Processes. Emotion, 6(2), 320–325. doi:doi: DOI: 10.1037/1528-3542.6.2.320

86
Hein, G., Alink, A., Kleinschmidt, A., & Müller, N. G. (2009). The attentional blink
modulates activity in the early visual cortex. Journal of cognitive neuroscience,
21(1), 197–206. doi:10.1162/jocn.2008.21026
Hollingsworth, D. E., McAuliffe, S. P., & Knowlton, B. J. (2001). Temporal
Allocation of Visual Attention in Adult Attention Deficit Hyperactivity
Disorder. Journal of Cognitive Neuroscience, 13(3), 298–305.
doi:10.1162/08989290151137359
Hoza, B. (2007). Peer functioning in children with ADHD. Journal of pediatric
psychology, 32(6), 655–663. doi:10.1093/jpepsy/jsm024
Hu, X.-Z., Lipsky, R. H., Zhu, G., Akhtar, L. A., Taubman, J., Greenberg, B. D., Xu,
K., et al. (2006). Serotonin transporter promoter gain-of-function genotypes are
linked to obsessive-compulsive disorder. American journal of human genetics,
78(5), 815–826. doi:10.1086/503850
Jefferies, L. N., Smilek, D., Eich, E., & Enns, J. T. (2008). Emotional Valence and
Arousal Interact in Attentional Control. Psychological Science, 19(3), 290–295.
doi:10.1111/j.1467-9280.2008.02082.x
Johansen, E. B., Aase, H., Meyer, A., & Sagvolden, T. (2002). Attention-
deficit/hyperactivity disorder (ADHD) behaviour explained by dysfunctioning
reinforcement and extinction processes. Behavioural brain research, 130(1-2),
37–45.

87
Johnson, M. H. (2005). Subcortical face processing. Nature Reviews Neuroscience,
6(10), 766–774. doi:10.1038/nrn1766
Johnston, W. (1986). Selective Attention. Annual Review of Psychology, 37(1), 43–75.
doi:10.1146/annurev.psych.37.1.43
Jolicoeur, P. (1998). Modulation of the attentional blink by on-line response selection:
evidence from speeded and unspeeded task1 decisions. Memory & Cognition,
26(5), 1014–1032.
Keil, A., Moratti, S., Sabatinelli, D., Bradley, M. M., & Lang, P. J. (2005). Additive
effects of emotional content and spatial selective attention on electrocortical
facilitation. Cerebral cortex (New York, N.Y.: 1991), 15(8), 1187–1197.
doi:10.1093/cercor/bhi001
Kohls, G., Herpertz-Dahlmann, B., & Konrad, K. (2009). Hyperresponsiveness to
social rewards in children and adolescents with attention-deficit/hyperactivity
disorder (ADHD). Behavioral and Brain Functions, 5(1), 20.
doi:10.1186/1744-9081-5-20
Kohls, G., Peltzer, J., Herpertz-Dahlmann, B., & Konrad, K. (2009). Differential
effects of social and non-social reward on response inhibition in children and
adolescents. Developmental Science, 12(4), 614–625. doi:10.1111/j.1467-
7687.2009.00816.x
Konrad, K., Gauggel, S., Manz, A., & Schöll, M. (2000). Lack of inhibition: a
motivational deficit in children with attention deficit/hyperactivity disorder and

88
children with traumatic brain injury. Child Neuropsychology: A Journal on
Normal and Abnormal Development in Childhood and Adolescence, 6(4), 286–
296.
Kranczioch, C., Debener, S., Schwarzbach, J., Goebel, R., & Engel, A. K. (2005).
Neural correlates of conscious perception in the attentional blink. NeuroImage,
24(3), 704–714. doi:10.1016/j.neuroimage.2004.09.024
Kratochvil, C. J., Faries, D., Vaughan, B., Perwien, A., Busner, J., Saylor, K., Kaplan,
S., et al. (2007). Emotional expression during attention-deficit/hyperactivity
disorders treatment: initial assessment of treatment effects. Journal of child and
adolescent psychopharmacology, 17(1), 51–62. doi:10.1089/cap.2006.0018
Kühle, H. J., Kinkelbur, J., Andes, K., Heidorn, F. M., Zeyer, S., Rautzenberg, P., &
Jansen, F. (2007). Self-regulation of visual attention and facial expression of
emotions in ADHD children. Journal of attention disorders, 10(4), 350–358.
doi:10.1177/1087054706289944
Kuyper, P. (1972). The cocktail party effect. Audiology: official organ of the
International Society of Audiology, 11(5), 277–282.
Lamb, M. E., & Sherrod, L. R. (1981). Infant social cognition : empirical and
theoretical considerations. Hillsdale, N.J.: L. Erlbaum Associates.
Lang, P. J., Bradley, M. M., Fitzsimmons, J. R., Cuthbert, B. N., Scott, J. D., Moulder,
B., & Nangia, V. (1998). Emotional arousal and activation of the visual cortex:
an fMRI analysis. Psychophysiology, 35(2), 199–210.

89
LeDoux, J. E. (2000). Emotion Circuits in the Brain. Annual Review of Neuroscience,
23(1), 155–184. doi:10.1146/annurev.neuro.23.1.155
Leppanen, J. M., & Nelson, C. A. (2009). Tuning the developing brain to social signals
of emotions. Nat Rev Neurosci, 10(1), 37–47. doi:10.1038/nrn2554
Lesch, K. P., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., Benjamin,
J., et al. (1996). Association of anxiety-related traits with a polymorphism in
the serotonin transporter gene regulatory region. Science (New York, N.Y.),
274(5292), 1527–1531.
Li, R., C.-shan, Chen, S.H., Lin, W.hao, & Yang, Y.-Y. (2005). Attentional blink in
adolescents with varying levels of impulsivity. Journal of Psychiatric
Research, 39(2), 197-205. doi:doi: DOI: 10.1016/j.jpsychires.2004.06.003
Li, C. R., Lin, W., Chang, H., & Hung, Y. (2004). A Psychophysical Measure of
Attention Deficit in Children With Attention-Deficit/Hyperactivity Disorder.
Journal of Abnormal Psychology, 113(2), 228–236. doi:doi:
Luman, M., Oosterlaan, J., & Sergeant, J. A. (2005). The impact of reinforcement
contingencies on AD/HD: a review and theoretical appraisal. Clinical
Psychology Review, 25(2), 183–213. doi:10.1016/j.cpr.2004.11.001
Luna, B., & Sweeney, J. A. (2004). The emergence of collaborative brain function:
FMRI studies of the development of response inhibition. Annals of the New
York Academy of Sciences, 1021, 296–309. doi:10.1196/annals.1308.035

90
MacLeod, C., & Cohen, I. L. (1993). Anxiety and the interpretation of ambiguity: A
text comprehension study. Journal of Abnormal Psychology, 102(2), 238–247.
doi:10.1037/0021-843X.102.2.238
Maedgen, J. W., & Carlson, C. L. (2000). Social functioning and emotional regulation
in the attention deficit hyperactivity disorder subtypes. Journal of clinical child
psychology, 29(1), 30–42. doi:10.1207/S15374424jccp2901_4
Maratos, F., Mogg, K., & Bradley, B. (2008). Identification of angry faces in the
attentional blink. Cognition & Emotion, 22(7), 1340–1352.
doi:10.1080/02699930701774218
Marois, R., & Ivanoff, J. (2005). Capacity limits of information processing in the brain.
Trends in Cognitive Sciences, 9, 296–305. doi:10.1016/j.tics.2005.04.010
Marois, R., Yi, D.-J., & Chun, M. M. (2004). The neural fate of consciously perceived
and missed events in the attentional blink. Neuron, 41(3), 465–472.
Marsh, R., Gerber, A. J., & Peterson, B. S. (2008). Neuroimaging Studies of Normal
Brain Development and Their Relevance for Understanding Childhood
Neuropsychiatric Disorders. Journal of the American Academy of Child &
Adolescent Psychiatry, 47(11), 1233–1251.
doi:10.1097/CHI.0b013e318185e703
Martens, S., & Wyble, B. (2010). The attentional blink: Past, present, and future of a
blind spot in perceptual awareness. Neuroscience & Biobehavioral Reviews,
34(6), 947–957. doi:doi: DOI: 10.1016/j.neubiorev.2009.12.005

91
Mason, D. J., Humphreys, G. W., & Kent, L. (2005). Insights into the control of
attentional set in ADHD using the attentional blink paradigm. Journal of Child
Psychology and Psychiatry, 46(12), 1345–1353. doi:10.1111/j.1469-
7610.2005.01428.x
Mogg, K., Bradley, B. P., de Bono, J., & Painter, M. (1997). Time course of attentional
bias for threat information in non-clinical anxiety. Behaviour Research and
Therapy, 35(4), 297–303.
Montague, D. P., & Walker-Andrews, A. S. (2001). Peekaboo: a new look at infants’
perception of emotion expressions. Developmental psychology, 37(6), 826–838.
Munafò, M. R., Brown, S. M., & Hariri, A. R. (2008). Serotonin transporter (5-
HTTLPR) genotype and amygdala activation: a meta-analysis. Biological
psychiatry, 63(9), 852–857. doi:10.1016/j.biopsych.2007.08.016
Nelson, C. A. (1987). The recognition of facial expressions in the first two years of
life: mechanisms of development. Child development, 58(4), 889–909.
Nigg, J. T., & Casey, B. J. (2005). An integrative theory of attention-deficit/
hyperactivity disorder based on the cognitive and affective neurosciences.
Development and psychopathology, 17(3), 785–806.
doi:10.1017/S0954579405050376
Nixon, E. (2001). The social competence of children with attention deficit
hyperactivity disorder: a review of the literature. Child Psychology and
Psychiatry Review, 6, 172–180.

92
Oades, R. D., Slusarek, M., Velling, S., & Bondy, B. (2002). Serotonin platelet-
transporter measures in childhood attention-deficit/hyperactivity disorder
(ADHD): clinical versus experimental measures of impulsivity. The World
Journal of Biological Psychiatry: The Official Journal of the World Federation
of Societies of Biological Psychiatry, 3(2), 96–100.
Paradiso, S., Johnson, D. L., Andreasen, N. C., O’Leary, D. S., Watkins, G. L., Ponto,
L. L., & Hichwa, R. D. (1999). Cerebral blood flow changes associated with
attribution of emotional valence to pleasant, unpleasant, and neutral visual
stimuli in a PET study of normal subjects. The American journal of psychiatry,
156(10), 1618–1629.
Perez-Edgar, K., Bar-Haim, Y., McDermott, J. M., Chronis-Tuscano, A., Pine, D. S., &
Fox, N. A. (2010). Attention Biases to Threat and Behavioral Inhibition in
Early Childhood Shape Adolescent Social Withdrawal. Emotion, 10(3), 349–
357. doi:doi: DOI: 10.1037/a0018486
Pérez-Edgar, K., Bar-Haim, Y., McDermott, J. M., Gorodetsky, E., Hodgkinson, C. A.,
Goldman, D., Ernst, M., et al. (2010). Variations in the serotonin-transporter
gene are associated with attention bias patterns to positive and negative
emotion faces. Biological psychology, 83(3), 269–271.
doi:10.1016/j.biopsycho.2009.08.009

93
Pessoa, L. (2002). Attentional control of the processing of neutral and emotional
stimuli. Cognitive Brain Research, 15(1), 31–45. doi:10.1016/S0926-
6410(02)00214-8
Phelps, E. A. (2006). Emotion and cognition: insights from studies of the human
amygdala. Annual review of psychology, 57, 27–53.
doi:10.1146/annurev.psych.56.091103.070234
Posner, M. I. (1993). Foundations of cognitive science. Cambridge, Mass.: MIT Press.
Pourtois, G., Schettino, A., & Vuilleumier, P. (2012). Brain mechanisms for emotional
influences on perception and attention: What is magic and what is not.
Biological psychology. doi:10.1016/j.biopsycho.2012.02.007
Pratto, F., & John, O. P. (1991). Automatic vigilance: the attention-grabbing power of
negative social information. Journal of personality and social psychology,
61(3), 380–391.
Rapport, L. J., Friedman, S. R., Tzelepis, A., Van Voorhis, A., & Friedman, S. L.
(2002). Experienced emotion and affect recognition in adult attention-deficit
hyperactivity disorder. Neuropsychology, 16(1), 102–110.
Ray Li, C., Chen, S.-H., Lin, W., & Yang, Y.-Y. (2005). Attentional blink in
adolescents with varying levels of impulsivity. Journal of Psychiatric
Research, 39(2), 197–205. doi:doi: DOI: 10.1016/j.jpsychires.2004.06.003
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992). Temporary suppression of
visual processing in an RSVP task: an attentional blink? Journal of

94
Experimental Psychology. Human Perception and Performance, 18(3), 849–
860.
Reimherr, F. W., Marchant, B. K., Strong, R. E., Hedges, D. W., Adler, L., Spencer, T.
J., West, S. A., et al. (2005). Emotional dysregulation in adult ADHD and
response to atomoxetine. Biological psychiatry, 58(2), 125–131.
doi:10.1016/j.biopsych.2005.04.040
Reynolds, C. R., & Kamphaus, R. W. (1992). Behavior Asseement System for
Children, BASC. San Antonio, TX: Pearson Education, Inc.
Romanski, L. M., & LeDoux, J. E. (1992). Equipotentiality of thalamo-amygdala and
thalamo-cortico-amygdala circuits in auditory fear conditioning. The Journal of
neuroscience: the official journal of the Society for Neuroscience, 12(11),
4501–4509.
Rossi, V., & Pourtois, G. (2012). State-dependent attention modulation of human
primary visual cortex: A high density ERP study. NeuroImage, 60(4), 2365–
2378. doi:10.1016/j.neuroimage.2012.02.007
Rubia, K., Overmeyer, S., Taylor, E., Brammer, M., Williams, S. C. R., Simmons, A.,
Andrew, C., et al. (2000). Functional frontalisation with age: mapping
neurodevelopmental trajectories with fMRI. Neuroscience & Biobehavioral
Reviews, 24(1), 13–19. doi:doi: DOI: 10.1016/S0149-7634(99)00055-X
Sabatinelli, D., Lang, P. J., Bradley, M. M., Costa, V. D., & Keil, A. (2009). The
timing of emotional discrimination in human amygdala and ventral visual

95
cortex. The Journal of neuroscience: the official journal of the Society for
Neuroscience, 29(47), 14864–14868. doi:10.1523/JNEUROSCI.3278-09.2009
Schachar, R., Tannock, R., Marriott, M., & Logan, G. (1995). Deficient inhibitory
control in attention deficit hyperactivity disorder. Journal of abnormal child
psychology, 23(4), 411–437.
Schub, A., Meinecke, C., Abele, A. E., & Gendolla, G. H. E. (2006). Detecting
Emotional Faces and Features in a Visual Search Paradigm: Are Faces Special?
Emotion, 6(2), 246-256. doi:doi: DOI: 10.1037/1528-3542.6.2.246
Schutter, D. J., Putman, P., Hermans, E., & van Honk, J. (2001). Parietal
electroencephalogram beta asymmetry and selective attention to angry facial
expressions in healthy human subjects. Neuroscience Letters, 314(1-2), 13-16.
Shapiro, K L, Caldwell, J., & Sorensen, R. E. (1997). Personal names and the
attentional blink: a visual “cocktail party” effect. Journal of Experimental
Psychology. Human Perception and Performance, 23(2), 504-514.
Soliva, J. C., Carmona, S., Fauquet, J., Hoekzema, E., Bulbena, A., Hilferty, J., &
Vilarroya, O. (2009). Neurobiological Substrates of Social Cognition
Impairment in Attention-Deficit Hyperactivity Disorder. Annals of the New
York Academy of Sciences, 1167(1), 212–220. doi:10.1111/j.1749-
6632.2009.04604.x
Somerville, L. H., Fani, N., & McClure-Tone, E. B. (2011). Behavioral and Neural
Representation of Emotional Facial Expressions Across the Lifespan.

96
Developmental Neuropsychology, 36(4), 408–428.
doi:10.1080/87565641.2010.549865
Sonuga-Barke, E. J. S. (2005). Causal models of attention-deficit/hyperactivity
disorder: from common simple deficits to multiple developmental pathways.
Biological psychiatry, 57(11), 1231–1238. doi:10.1016/j.biopsych.2004.09.008
Sonuga-Barke, E. J. S. (2003). The dual pathway model of AD/HD: an elaboration of
neuro-developmental characteristics. Neuroscience and biobehavioral reviews,
27(7), 593–604.
Sonuga-Barke, E. J. S. (2002). Psychological heterogeneity in AD/HD--a dual pathway
model of behaviour and cognition. Behavioural brain research, 130(1-2), 29–
36.
Sonuga-Barke, E., Bitsakou, P., & Thompson, M. (2010). Beyond the dual pathway
model: evidence for the dissociation of timing, inhibitory, and delay-related
impairments in attention-deficit/hyperactivity disorder. Journal of the American
Academy of Child and Adolescent Psychiatry, 49(4), 345–355.
Spielberger, C. D., Edwards, C. D., Montuori, J., & Lushene, R. (1983). State-Trait
Anxiety Inventory for Children. Menlo Park, CA: Mind Garden, Inc.
Srivastava, P., & Srinivasan, N. (2010). Time course of visual attention with emotional
faces. Attention, Perception & Psychophysics, 72(2), 369-377.
doi:10.3758/APP.72.2.369

97
Stein, T., Peelen, M. V., Funk, J., & Seidl, K. N. (2010). The Fearful-Face Advantage
Is Modulated by Task Demands: Evidence From the Attentional Blink.
Emotion, 10(1), 136-140. doi:doi: DOI: 10.1037/a0017814
Stopa, L. (2000). Social phobia and interpretation of social events. Behaviour Research
and Therapy, 38(3), 273-283. doi:10.1016/S0005-7967(99)00043-1
Taylor, J. G., & Fragopanagos, N. F. (2005). The interaction of attention and emotion.
Neural Networks, 18(4), 353–369. doi:doi: 10.1016/j.neunet.2005.03.005
Taylor, M. J., Batty, M., & Itier, R. J. (2004). The faces of development: a review of
early face processing over childhood. Journal of Cognitive Neuroscience,
16(8), 1426–1442. doi:10.1162/0898929042304732
Tibboel, H., Van Bockstaele, B., & De Houwer, J. (2011). Is the emotional modulation
of the attentional blink driven by response bias? Cognition & Emotion, 25(7),
1176–1183. doi:10.1080/02699931.2010.524192
Toga, A. W., Thompson, P. M., & Sowell, E. R. (2006). Mapping brain maturation.
Trends in Neurosciences, 29(3), 148–159. doi:doi: DOI:
10.1016/j.tins.2006.01.007
Visser, T. A. W., Boden, C., & Giaschi, D. E. (2004). Children with dyslexia: evidence
for visual attention deficits in perception of rapid sequences of objects. Vision
Research, 44(21), 2521-2535. doi:doi: 10.1016/j.visres.2004.05.010
Vuilleumier, P. (2005). How brains beware: neural mechanisms of emotional attention.
Trends in Cognitive Sciences, 9(12), 585–594. doi:10.1016/j.tics.2005.10.011

98
Vuilleumier, P., & Pourtois, G. (2007). Distributed and interactive brain mechanisms
during emotion face perception: Evidence from functional neuroimaging.
Neuropsychologia, 45(1), 174–194. doi:doi: DOI:
10.1016/j.neuropsychologia.2006.06.003
Vuilleumier, P, & Schwartz, S. (2001). Beware and be aware: capture of spatial
attention by fear-related stimuli in neglect. Neuroreport, 12(6), 1119–1122.
Weissman, D. H., & Prado, J. (2012). Heightened activity in a key region of the ventral
attention network is linked to reduced activity in a key region of the dorsal
attention network during unexpected shifts of covert visual spatial attention.
NeuroImage, 61(4), 798–804. doi:10.1016/j.neuroimage.2012.03.032
Wender, P. H., Wolf, L. E., & Wasserstein, J. (2001). Adults with ADHD. An
overview. Annals of the New York Academy of Sciences, 931, 1–16.
Weschler, D. (n.d.). Wechsler Intelligence Scale for Children® — Fourth Edition
(WISC®-IV). Retrieved July 6, 2010, from
http://psychcorp.pearsonassessments.com/HAIWEB/Cultures/en-
us/Productdetail.htm?Pid=015-8979-044
Whalen, P. J. (1998). Fear, Vigilance, and Ambiguity: Initial Neuroimaging Studies of
the Human Amygdala. Current Directions in Psychological Science, 7(6), 177–
188. doi:10.1111/1467-8721.ep10836912
Wheeler, J., & Carlson, C. (1994). The social functioning of children with ADD with
hyperactivity and ADD without hyperactivity: A comparison of their peer

99
relations and social deficits. Journal of Emotional and Behavioral Disorders,
2(1), 2–13.
Whiten, A. (1997). Machiavellian intelligence. Cambridge ;;New York: Cambridge
University press.
Williams, L. M., Hermens, D. F., Palmer, D., Kohn, M., Clarke, S., Keage, H., Clark,
C. R., et al. (2008). Misinterpreting emotional expressions in attention-
deficit/hyperactivity disorder: evidence for a neural marker and stimulant
effects. Biological psychiatry, 63(10), 917–926.
doi:10.1016/j.biopsych.2007.11.022
Woodcock, R.W., McGrew, K.S., & Mather, N. (2000). Woodcock-Johnson III.
Rolling Meadows, IL: Riverside Publishing.
Yiend, J. (2010). The effects of emotion on attention: A review of attentional
processing of emotional information. Cognition & Emotion, 24(1), 3–47.
doi:10.1080/02699930903205698
Yuill, N., & Lyon, J. (2007). Selective difficulty in recognising facial expressions of
emotion in boys with ADHD. General performance impairments or specific
problems in social cognition? European child & adolescent psychiatry, 16(6),
398–404. doi:10.1007/s00787-007-0612-5