a clearing picture of phytophthora evolution: from the ... · a clearing picture of phytophthora...
TRANSCRIPT

A clearing picture of Phytophthora evolution: from
the wide-angle to the zoom lens for optimal phylogenetic focus
David Cooke, SCRI

• Introduction – aims of paper
• Early evolution of oomycetes
• Lessons learned from genomics
• Current status of Phytophthora phylogeny
• Tools and drivers: fine-scale population shifts
• Conclusions
• Acknowledgements
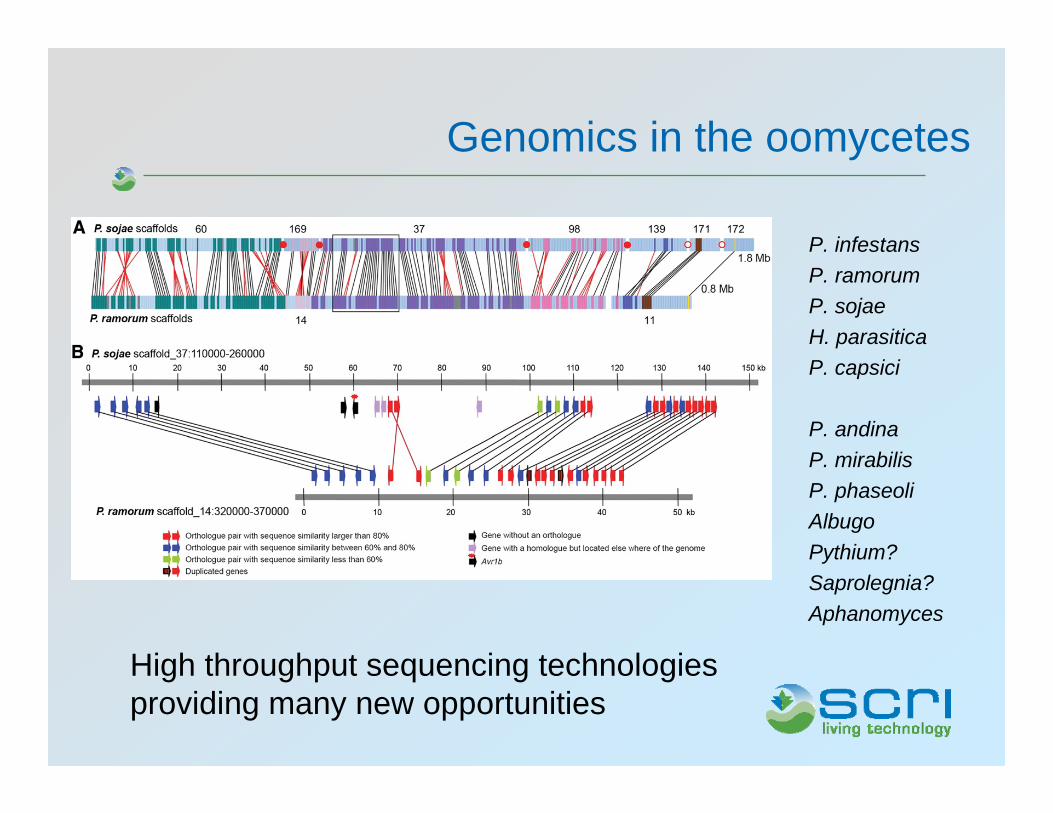
Genomics in the oomycetes
P. infestansP. ramorumP. sojaeH. parasiticaP. capsici
P. andinaP. mirabilisP. phaseoliAlbugoPythium?Saprolegnia?Aphanomyces
High throughput sequencing technologies providing many new opportunities

Eukaryote tree of life
Lane & Archibald, TREE 2008

Contributions of genomics
Lane & Archibald, TREE 2008
Tyler et al., 2006 ScienceP. ramorum and P. sojae genomesFound 855 Phytophthoragenes with similarity to red algae or cyanobacteria
Whisson et al 2007 NatureShared mechanisms of effector transport into host cells – oomycete and Apicomplexans
RxLR class of effectors considered key to pathogenicity: a reduced repertoire noted in Hy. parasitica genome (Warwick HRI). Also an absence of genes involved in zoospore formation…
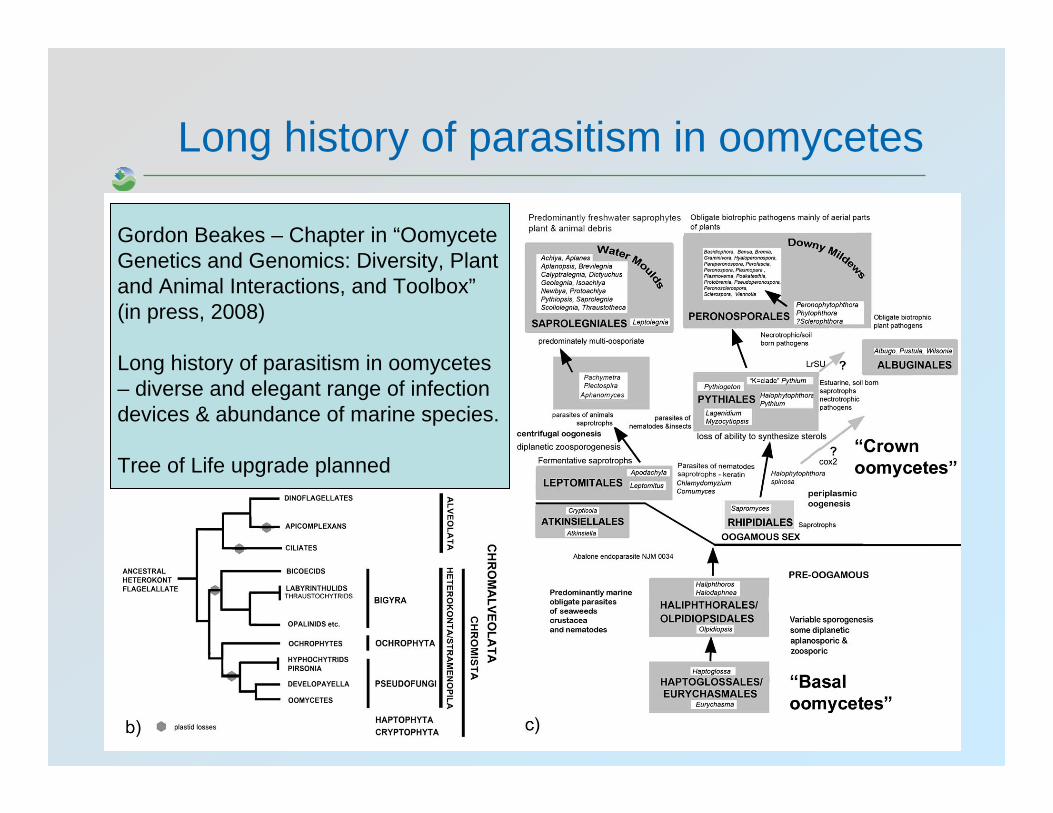
Long history of parasitism in oomycetes
Gordon Beakes – Chapter in “OomyceteGenetics and Genomics: Diversity, Plant and Animal Interactions, and Toolbox”(in press, 2008)
Long history of parasitism in oomycetes– diverse and elegant range of infection devices & abundance of marine species.
Tree of Life upgrade planned

Basal oomycetesThe Development, Ultrastructural Cytology, and Molecular Phylogeny of the Basal OomyceteEurychasma dicksonii …
Satoshi Sekimoto, Gordon W. Beakes, Claire M.M. Gachon, Dieter G. Müller, Frithjof C. Küpper, Daiske Honda Protist, 2008
Parasites of plants across oomycete lineages
rDNA 18S tree

Cooke et al 2000 c.f. Blair et al 2007
• Multigeneapproach more robust
• Clades shared
• Basal branch structure similar

ITS Tree 2008• Scan of GenBank Taxonomy
browser for ITS seq.
• 146 seq but some redundancy
• Clade 8 position of P. cryptogea & drechsleri
• Clade 7 P. fragaefolia on deep root
• Clade 5 P. novaeguinee and a NZ isolate
• Clade 2 P. frigida in basal position
• Clade 4 ‘tropical and australasian flavour?’
• Clade 6 dramatic expansion & appearance of aerial habit
0.1
PquinineaPmacrochl
PrichardEU301150Psp.TIB20079PfallaxPcaptiosaEU644726Psp.BFAEU196368Psp.167008EU196369Psp.167011Pgallica PboehmeriaeDQ066921Psp.Drimys winteriDQ002008Psp.cc2124Pkernoviae PpolonicaPinsol cookeEF590254PcuyabensisEF680320PnapoensisEU419612Psp.6216900EU419615Psp.8021391EU334634Pirrigata23J7EU644721Psp.BF3EU644715Psp.BF4EU644714Psp.BFKEF680321PthermophilumEU644725Psp.BF1EU583798PhydropathicaSCRP237Rf6EU164426.1PzentmyeriiEF590256PlagoarianaEU644720Psp.BFREU644722Psp.BF36Rf17 PbrassicaePprimulaePporriPfoliorum PhibernalisPramorumPlateralisPsyringaeP. austrocedraePdrechsleriPmedicaginisPtrifoliiPsansomeaPkelmaniaPcryptogeaPerythrosepticaP. fragaefoliaPsojaePniederhauseriiPpistaciaePmelonis
PcajaniPvignae
PcinnamomiPcinnparvi
PfvrPuliginosa
PeuropaeaPalnimultPcambivora
PquercinaPilicisPpsychrophila
PpseudosyringaePnemorosa
PheveaePkatsurae
EF067922Psp.NZICMPP. novaeguineePpseudotsugae
PcactPidaei
PhedraiandraPnicotianae
PtentaculataPclandestinaPiranica
Pinfestans & andinaPipomoeae
PmirabilisPphaseoli
P. frigidaEU644723Psp.BLW71
PmultivesiculataEU301125Psp.TIB20072 strain
PbisheriaPcolocasiae
PbotryosaEU419614Psp.204BPcitrophthoraEU419613Psp.1211
EF523387PsiskiyouensisPtropicalis
PcapsiciPglovera
DQ821185Psp.BR514EU301132Psp.TIB20074
PcitricolaPinflataEU877749Psp.59EU877753.1Psp.63EU877755Psp.65EU877754Psp.64
PmegakaryaPelitch vogl
EU419617Psp.1931PpalmivoraParecae
P. alticolaP. quercetorumEU301118Psp.TIB20071 strain Psp banksia
PasparaEF590257Psulawesiensis
EU593265Psp.IMI329669EU301170Psp.TIB20076
PinundataPhumicola
EU000108Psp.77231EU594601Psp.1FFL2008EF153672Psp.P12745
PmegaspermaEU301174Psp.TIB20077
DQ396412Psp.AG34UASWS0200EU000143Psp.AG43EU106591Psp.92209C
EU594600Psp.2FFL2008EU106590Psp.92207EU301176Psp.TIB20078
EU869199PaustralisVHSEU301158.1Psp.TIB20073 strai
EF152514Psp.GD2fEF522140Psylvatica
EU725809PpinifoliaEU594598PhungaricaEU594599Psp.4FFL2008
PgonapodyidesEU419616Psp.B6EU000124Psp.91112AY787023Psp.AG52
6
4
2
153
7
8
9 & 10

Expansion of clade 9 & 10
• Discrete lineage from majority of genus
• No obvious common morphological/behavioural traits
• Distinct geographical origin but blurred by migration?
P. quinineaP. macrochlamydospora
P. richardiaeEU301150Psp.TIB20079
P. fallaxP. captiosa
EU644726Psp.BFAEU196368Psp.167008EU196369Psp.167011
P. gallicaP. boehmeriae
P. kernoviae
P. polonicaP. insolita
P. cuyabensisP. napoensis
EU419612Psp.6216900EU419615Psp.8021391
P. irrigataEU644721Psp.BF3
EU644715Psp.BF4EU644714Psp.BFK
P. thermophilumEU644725Psp.BF1
P. hydropathicaP. parsiana
P. zentmyeriiP. lagoarianaEU644720Psp.BFR
EU644722Psp.BF36Rf17
10
9
9

Status of Phytophthora phylogenetics?
• Move from single gene to multigene phylogenies
• ITS remains good starting point
• Number of species increasing but main structure unchanged
• Expansion of clades 9 & 10 most marked
• Ever expanding range of taxa in clade 6
• Monitoring methods by Silvia Scibetta – powerful tool
• Still issues with GenBank – reference collection better in independent source (Phytophthora database)

Oomycete pathogen population analysis
• Neutral markersAFLPsSSRs
P. infestans, P. ramorum, Pl. viticolaAccelerated discovery examined by Schena and Cooke (BMC Genomics submitted)
SequencingCoalescent analysis
• Functional markersHost rangeVirulenceAggressiveness & FitnessFungicide resistanceSequencing genes under strong selection
effector genes e.g. RxLRs P. infestans, Hy. parasitica
Carbone web page
• Mutation • Recombination • Natural selection • Gene flow• Random genetic drift • Migration

Simple Sequence Repeats – P. infestans
ABI 3730 capillary sequencer
Marker G11
Isolate 2
Isolate 1
Isolate 4
Isolate 3
Multiplex PCR – protocols on www.eucablight.org

Changes in British P. infestans population
• Eucablightdatabase - powerful
• Increase in A2 mating type seen in GB
• What about genotypes?
Scotland
England

Derived data plotted on web
• Differences in allele frequency between countries
• Expansion to South and Central America complete
• Further expansion under discussion

Great Britain SSR genotypes 1982-2007
• Major change in 2006 and 2007• Some genotypes very durable, others not. Why?
A2
A1
misc.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Freq
uenc
y
1982 1995 1996 1997/8 2003 2004 2005 2006 2007
GB genotypes
7,8
6
512
2
4
3
10
1371%
10%
US1old
85 581 73 899Isolate no. 145229026418834
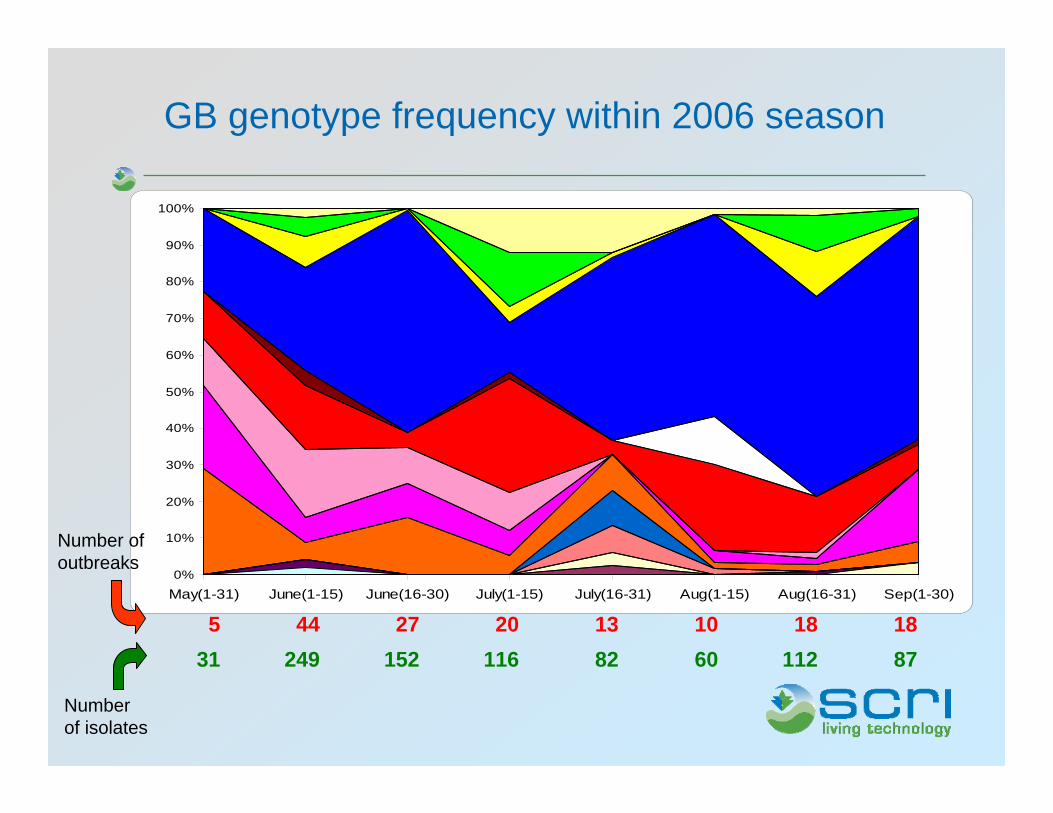
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
May(1-31) June(1-15) June(16-30) July(1-15) July(16-31) Aug(1-15) Aug(16-31) Sep(1-30)
GB genotype frequency within 2006 season
Number of outbreaks
Number of isolates
87112608211615224931181810132027445
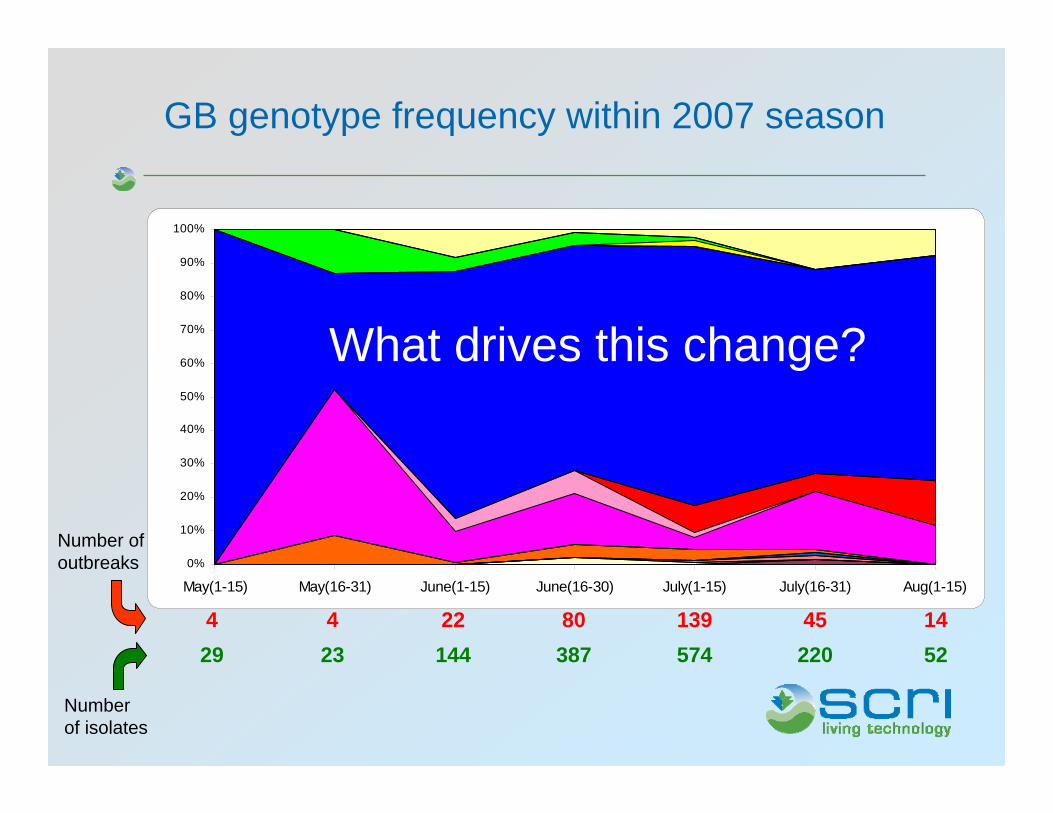
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
May(1-15) May(16-31) June(1-15) June(16-30) July(1-15) July(16-31) Aug(1-15)
GB genotype frequency within 2007 season
Number of outbreaks
Number of isolates
5222057438714423291445139802244
What drives this change?

Lesion area (mean of all varieties)
0
2
4
6
8
10
12
14
16
6_A1
_410
0ANL0
6269
13_A
2_38
84B
6_A1
_392
0ANL0
4246
13_A
2_39
64A
13_A
2_39
28A
17_A
2_43
88D
7_A1_
4168
C7_
A1_41
68B
10_A
2_44
40C
13_A
2_41
32B
10_A
2_39
36C2
SE0305
8SE0
3087
2_A1_
3888
A3b
_A2_
4244
E2_
A1_40
68B
LD 15
1 PMP62
2
8a_A
1_42
56B
3_A2
_401
2FMP61
8 C4
8_2a
_A1_
4232
E C2Le
sion
are
a
13 oC
• On average, genotype 13 isolates result in larger lesions than other genotypes at 13oC
• Genotype 6 largest lesions of A1 genotypes
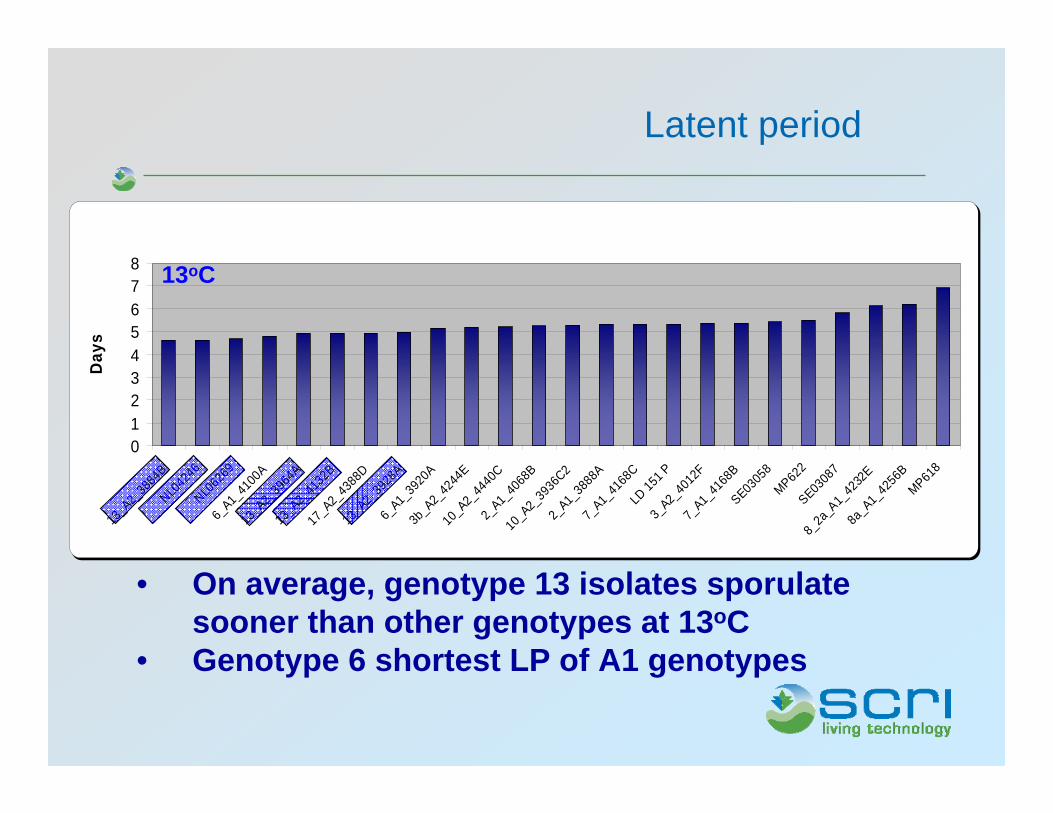
Latent period
• On average, genotype 13 isolates sporulatesooner than other genotypes at 13oC
• Genotype 6 shortest LP of A1 genotypes
012345678
13_A2_
3884B
NL042
46NL0
6269
6_A1_
4100
A13
_A2_
3964
A13_
A2_413
2B17_
A2_438
8D13
_A2_
3928
A6_
A1_392
0A3b
_A2_42
44E
10_A2_4
440C
2_A1_
4068
B10
_A2_
3936
C22_
A1_388
8A7_
A1_41
68C
LD 15
1 P3_
A2_401
2F7_
A1_416
8BSE030
58MP62
2SE03
087
8_2a_
A1_423
2E8a_
A1_425
6BMP61
8D
ays
13oC

Field trial results
• All isolates were pathogenic in lab test at D0• Domination of genotype 13 clear• Four other genotypes (1 alien) rare
0.00%
10.00%
20.00%
30.00%
40.00%
50.00%
60.00%
70.00%
80.00%
90.00%
100.00%Pe
rcen
tage
of e
ach
geno
type
D1 D2 D3 D4 D5 D6
x728613
No. lesions 67 67 99 100 17 20

Virulence of genotype 8- A1
R1 R2 R6R3 R5R4 R7 R8 R10 R11
BPC_06_4256B = 8_A1 1,4,7,10,11
Less complex and less aggressive (note 7,10&11)

Virulence of genotype 13 - A2
R1 R2 R6R3 R5R4 R7 R8 R10 R11
BPC_06_3928A = 13_A2 1,2,3,4,5,6,7,9,10,11Complex virulence and aggressive
Resistant to metalaxyl

• Study of evolutionary forces on key pathogen traits Panel of 30 GB isolates• Avr 3a
• 3 SNPs (3 replacement changes)• 23 EM homozygous (virulent) • 6 EM/KI heterozygous (avirulent)• 1 KI homozygous (avirulent)
• avirulent strains confined to a specific lineage which explains decline of this avirulence• R3a not deployed widely (3.5% area) – low selection pressure implies no ‘cost’ to virulence for avr3a
• PEX 161• 9 SNPs (1 outside ORF, 2 silent, 6 replacement)• 5 haplotypes
• 7 TTGAATCGC A• 2 YTGMWWYRY B• 2 YWRMWWYRY C• 14 YWRAAWYRY D• 1 CAAAAATAT E
• Haplotypes correspond to SSR lineages
6
2
2
4
3
3
2
7/1
3_A210_A2
13_A2
1_A1
6_A1
2_A1
7&8_A1
rare
Structured screening of RxLR diversity
C
B
A
D
D
A
A/B/D/E
D

Conclusions
• Genomics
High throughput, affordable (£5K per 100 Mb) DNA sequencing offering insights into mechanisms of pathogenicity, host range and ultimately host resistance responses. Post-genomics is going to be a ‘long haul’.
• Phylogenetics
Exciting prospects for environmental monitoring of Phytophthora diversity in support of conventional approaches. Evaluating risks. Exploring what happens in ‘balanced’ natural ecosystems as an aid to understanding invasive species. Phylogeography to improve our understanding of origins of species. Phylogenomics to understand mechanisms conserved across genus.
• Population analysis
Tracking populations – co-ordinated responses needed – tracking using standardised tools – sharing ‘intelligence’ on local problems. Predicting risk? How does population structure influence management? Examining selection pressures on key genes.

Acknowledgments
• Eucablight team Jens Hansen, Poul Lassen & Alison Lees
• SCRI Team: Alison Lees, Naomi Williams and Louise Sullivan,
• Italian team: Silvia Scibetta, Leonardo Schena, SantinaCacciola and others
• BPC team: David Shaw, Ruairidh Bain, Nick Bradshaw, Moray Taylor
• Many, many people who have contributed inspiration, friendship & cultures
• Potato Council & RERAD for funding