6801 01 p443-454eea.spaceflight.esa.int/attachments/retrievablecapsules/id4ced... · height) that...
TRANSCRIPT

ASTROBIOLOGYVolume 7, Number 3, 2007© Mary Ann Liebert, Inc.DOI: 10.1089/ast.2006.0046
Research Paper
Lichens Survive in Space: Results from the 2005 LICHENS Experiment
LEOPOLDO G. SANCHO,1 ROSA DE LA TORRE,2 GERDA HORNECK,3CARMEN ASCASO,4 ASUNCIÓN DE LOS RIOS,4 ANA PINTADO,1 J. WIERZCHOS,5
and M. SCHUSTER6
ABSTRACT
This experiment was aimed at establishing, for the first time, the survival capability of lichensexposed to space conditions. In particular, the damaging effect of various wavelengths of ex-traterrestrial solar UV radiation was studied. The lichens used were the bipolar species Rhi-zocarpon geographicum and Xanthoria elegans, which were collected above 2000 m in themountains of central Spain and as endolithic communities inhabiting granites in the Antarc-tic Dry Valleys. Lichens were exposed to space in the BIOPAN-5 facility of the EuropeanSpace Agency; BIOPAN-5 is located on the outer shell of the Earth-orbiting FOTON-M2 Russ-ian satellite. The lichen samples were launched from Baikonur by a Soyuz rocket on May 31,2005, and were returned to Earth after 16 days in space, at which time they were tested forsurvival. Chlorophyll fluorescence was used for the measurement of photosynthetic parame-ters. Scanning electron microscopy in back-scattered mode, low temperature scanning elec-tron microscopy, and transmission electron microscopy were used to study the organizationand composition of both symbionts. Confocal laser scanning microscopy, in combination withthe use of specific fluorescent probes, allowed for the assessment of the physiological stateof the cells. All exposed lichens, regardless of the optical filters used, showed nearly the samephotosynthetic activity after the flight as measured before the flight. Likewise, the multimi-croscopy approach revealed no detectable ultrastructural changes in most of the algal and fun-gal cells of the lichen thalli, though a greater proportion of cells in the flight samples hadcompromised membranes, as revealed by the LIVE/DEAD BacLight Bacterial Viability Kit.These findings indicate that most lichenized fungal and algal cells can survive in space afterfull exposure to massive UV and cosmic radiation, conditions proven to be lethal to bacteriaand other microorganisms. The lichen upper cortex seems to provide adequate protection
443
1Departamento de Biología Vegetal II, Universidad Complutense, Madrid, Spain.2Instituto Nacional de Técnica Aeroespacial, Torrejón, Madrid, Spain.3Institute of Aerospace Medicine, German Aerospace Center DLR, Köln, Germany.4Centro de Ciencias Medioambientales, CSIC, Madrid, Spain.5Servei de Microscopia Electrónica, Universitat de Lleida, Spain.6Universität Erlangen-Nürnberg, Ökophysiologie der Pflanzen, Erlangen, Germany.

INTRODUCTION
EXOBIOLOGY EXPERIMENTS are directed towardthe possibility of the presence or survival of
life beyond the confines of Earth. Interplanetaryspace is especially interesting because of the po-tential for testing Panspermia. The Panspermiatheory holds that reproductive bodies of livingorganisms can exist throughout the universe anddevelop wherever the environment is favorable(Richter, 1865; Arrhenius, 1903). In recent years,the Panspermia theory has been reformulated interms of the Lithopanspermia hypothesis, whichpostulates that microorganisms that reside withinimpact ejecta could be transported between plan-ets (Fajardo-Cavalzos et al., 2005). Spores of Bacil-lus subtilis can survive years of exposure to spacein low-Earth orbit (Horneck, 1993; Horneck et al.,2001; Rettberg et al., 2002), shock and heat of sim-ulated impact (Horneck et al., 2001; Burchell et al.,2004), and hypervelocity atmospheric entry (Fa-jardo-Cavalzos et al., 2005). However, a majorcriticism of interplanetary transfer of life is thatthe extremely harsh conditions of interplanetaryspace, mainly massive UV and cosmic radiation,are lethal to all forms of life. Experiments led bythe European Space Agency (ESA), however,have shown that some terrestrial organisms cansurvive this highly extreme environment if thereis sufficient shielding from UV radiation (Hor-neck et al., 1984; Mancinelli et al., 1998; Mancinelliand Klovstad, 2000). In a study by Rothschild andMancinelli (2001), however, only 25%–40% ofhalophilic Archaea and cyanobacteria survivedafter two weeks of exposure to the Sun in outerspace, and Bertolani et al. (2001) found that mul-ticellular eukaryotic organisms like nematodesperish when directly exposed to the Sun.
Many lichen species are regarded as ex-tremophiles in terms of their tolerance to tem-perature, radiation, and desiccation. In highmountains and polar regions, lichens are welladapted to long-term desiccation, temperaturesbetween �40 °C and 60 °C, and high radiation,including UV (Green et al., 1999; de la Torre et al.,2002; Heber et al., 2006). Dry lichens can recoverafter 10 years of inactivity and are able to survive
after immersion in liquid nitrogen (Kappen andValladares, 1999; Kappen, 2000). Dry lichens alsohave a high tolerance for radioactivity and can bethe first organisms to colonize––or the longest tosurvive––in areas of high radioactive contamina-tion (Purvis et al., 2004). Therefore, lichens seemto be pre-adapted to contend with the extremeconditions of space, including vacuum, abrupttemperature changes, and massive UV and cos-mic radiation.
The experiment LICHENS was aimed at estab-lishing for the first time the survival capability oflichens when exposed to space conditions, in par-ticular, the damaging effect of solar UV. Thelichens used were the bipolar species Rhizocarpongeographicum and Xanthoria elegans. Because theseorganisms are desiccation tolerant and survive inregions with extremely high amounts of UV ra-diation, we hypothesized that these organismswould be uniquely suited to survival in the harshconditions of space. Moreover, lichens can be con-sidered as simple ecosystems made up of au-totrophic and heterotrophic partners. Chloro-phycean Trebouxia colonies are the autotrophs ofboth lichens tested, though cyanobacteria areusually also present with endolithic communities(Friedmann, 1982). The heterotrophic fungi in as-sociation with the lichens fall within the Divisionof Ascomycota.
MATERIALS AND METHODS
Thalli of Rhizocarpon geographicum on naturalrock were collected in the mountains of centralSpain (La Plataforma, Sierra de Gredos, Avila,Spain; 2020 m above sea level, 40°16� N, 5°14�19�W), and thalli of Xanthoria elegans were collectedfrom their rock habitat in the Sierra NevadaMountains of southern Spain (Peñones de SanFrancisco, Sierra Nevada, Granada, Spain; 2400 mabove sea level, 37°06� N, 03°23� W). Pieces ofgranite rock that contained endolithic lichenswere collected in continental Antarctica (GraniteHarbour, Ross Sea Coast, Antarctica; 150 m abovesea level, 77°00� S, 162°34� E). Cylindrical rocksamples (10 mm in diameter and 6.9 mm in
SANCHO ET AL.444
against solar radiation. Moreover, after extreme dehydration induced by high vacuum, thelichens proved to be able to recover, in full, their metabolic activity within 24 hours. KeyWords: Lithopanspermia—Exobiology—Lichens—BIOPAN experiment. Astrobiology 7,443–454.

height) that contained epilithic or endolithiclichens were cut out of larger rocks with a dia-mond point saw continuously cooled with run-ning water. Only those areas of the rock thatshowed a rich population of lichens were selectedfor the LICHENS experiment. The diameter ofthese samples was chosen so that they coincidedwith the diameter of the optic fiber of the fluo-rometer system, which would allow for homoge-neous measurements of the whole thallus duringsuccessive experiments.
Experimental design
As part of the FOTON-M2 mission, launchedfrom Baikonur on May 31, 2005, on board a Soyuzrocket, material for the LICHENS experiment wasincluded in the ESA facility BIOPAN. BIOPAN isa multi-user exposure facility, designed for exo-biology, radiation biology, radiation dosimetry,and material science investigations in space (Fig.1). BIOPAN is installed on the external surface ofthe FOTON descent capsule that protrudes from
LICHENS SURVIVE IN SPACE 445
FIG. 1. BIOPAN-5 when it was openedimmediately after the flight. Arrowshows the position of the experimentLICHENS.
FIG. 2. LICHENS experimental hardware:The lichen samples were accommodated inthe 12 cells of plates no. 2 and 3. The topplate (no. 1) gave support to screw heads.Plate no. 2 contained the cells for the flightsamples exposed to solar UV and VIS radi-ation and plate no. 3 the cells for the in-flightdark controls. Ventilation holes passthrough all three plates.

the thermal blanket that covers the satellite. Themission profile of the facility, described inDemets et al. (2005) and Baglioni et al. (2007), in-cludes late installation at the launch site and earlyretrieval after landing. BIOPAN is equipped witha motor-driven hinged lid that, during nominaloperations, opens 180° in orbit to expose the sam-ples within to the harsh space environment. InBIOPAN, the test samples are exposed to a basicmenu of microgravity and cosmic radiation, withexposure to space vacuum and/or solar UV ra-diation as optional additions.
The hardware to which lichens were fixed wasdesigned and constructed by INTA (SpanishAerospace Establishment, Madrid, Spain). Thesample carrier consisted of two sample plates(78 � 50 � 23 mm) for the accommodation oflichen samples (nos. 2 and 3 in Fig. 2) and a coverplate (80 � 50 � 3 mm), which gave support toscrew heads and the optical filters below its cir-cular holes (no. 1 in Fig. 2). Each sample plate wascomprised of 12 cells that housed the lichen sam-ples; vent holes made for a direct interaction withthe space vacuum. Ventilation holes passedthrough all three plates and were left open dur-ing the entire space flight. The top layer (no. 2 inFig. 2) allowed for exposure of the lichen samplesto the full space environment, which included se-lected wavelength ranges of extraterrestrial solarUV and VIS radiation. To expose the samples toa discrete number of wavelengths, each samplecell was covered with an optical filter (13 mm di-ameter and 2 mm thick). Assembly was as fol-lows: 3 cells with a Suprasil SQ0 quartz windowallowed transmission of wavelengths at � � 170nm; 3 cells with a long pass filter allowed trans-mission of UV at � � 280 nm; 3 cells with a longpass filter allowed transmission of UV at � � 320nm; and 3 cells exposed the samples to VIS (� �400 nm). The bottom layer (no. 3 in Fig. 2) ac-commodated an identical set of lichen samples,which experienced the same space exposure con-ditions as those in layer 2 except for exposure tosolar UV and VIS radiation. They served as in-flight dark controls. For thermal control, the coverplate (plate no. 1 in Fig. 2) was painted white (SG-121 FD®) to reflect radiation. We used standardspace materials, ISO AlMgSi, an aluminum alloy,for fabrication of the hardware, which had a fi-nal area of 78 � 50 mm and a total mass of 250 g.Cylindrical biological membranes of polysulfon(Berghof und Anlagetechnik GmbH), with a poresize of 100,000 daltons, were positioned inside the
sample cells to avoid contamination between theexperiments integrated in BIOPAN. For fixationof the lichen samples in the hardware cells, weused Scotchweld® and Silicone RTV566® gluemixed with Primer SS41565® (General Electric Sil-icones Europe).
The lichens were exposed to the space envi-ronment for two weeks while in Earth orbitaboard the BIOPAN facility. An identical set ofhardware with lichens was produced and keptfor the same period in the laboratory at ambientconditions (air, 20 °C) and in darkness to serve aslaboratory ground control (Earth control). Duringthe mission all samples were in an anabiosis statecaused by natural dehydration.
Fluorescence analysis
On June 16, immediately after landing in theregion of Kostanay (Kazakhstan), the BIOPANwas detached from the FOTON capsule and sentto the European Space Research and TechnologyCentre (Nordwijk, the Netherlands). TheBIOPAN was opened on June 19 in the ESTECclean room, and the experiments were returnedto the corresponding researchers. The followingday, we started the rehydration and revitalizationprocess under controlled conditions in our labo-ratory facilities in Madrid. These consisted ofmaintaining probes in a climatic chamber at atemperature of 10 °C and a 12 h dark / 12 h lightcycle treatment for 72 h. This photosyntheticallyactive light (400–700 nm) was obtained from amercury lamp that reached the probes at around100 �mol m�2 s�1 photosynthetic photon fluxdensity (PPFD). The sample chambers weresprayed with 25 ml of deionized water twice aday. Fluorescence measurements were carriedout two or three hours after spraying the lichenswith water.
Survivability was determined first by measur-ing the quantum yield activity of photosystem II,assessed by fluorescence measurements ofchlorophyll a, using a MiniPAM (Walz, GmbH,FRG) (Maxwell and Johnson, 2000). The measur-ing and saturation light passed through a fiberoptic cable held at an angle of 60° to the lichensurface. The cable had nearly the same diameteras the lichen sample (10 mm) to allow better repli-cates of the same measurements. Potential maxi-mal photosystem II (PSII) quantum yield(Fv/Fm � variable fluorescence / maximal fluo-rescence)—for nomenclature see van Kooten and
SANCHO ET AL.446

Snel (1990)—was measured after the samples hadbeen kept for 20 min in darkness, so that all re-action centers were open (Schreiber et al., 1994).Due to physical constraints, fluorescence cannotbe obtained from the rock samples that containedthe endolithic algae. Fluorescence and electronmicroscopy techniques allowed us to evaluate theintegrity of the algae and fungi of both endolithicand epilithic lichens.
Microscopy analysis
Confocal scanning laser microscopy. Sam-ples were analyzed with a Zeiss LSM 310 con-focal microscope. Fluorescence assays and de-termination of the proportion of living and deadcells were performed on fragments of lichenthalli immediately after the photosynthetic ex-periments were finished according to the pro-tocol described in de los Ríos et al. (2004). TheLIVE/DEAD BacLight Bacterial Viability Kit L-13152 (Molecular Probes) was used to distin-guish living cells from cells with compromisedcell membranes, the latter interpreted as deadmycobiont and photobiont cells. This kit con-tains two proprietary nucleic acid stains thatdiffer in their ability to penetrate bacterial cellmembranes. The green fluorescence nucleic acidstain, SYTO 9, labels all cells, and the red fluo-rescence nucleic acid stain, propidium iodide,only penetrates cells with damaged membranesand quenches the green SYTO 9 stain. As noted,we interpreted cells that were fluorescing greenas viable, and those that were fluorescing red asdead, though the test only gives an indicationof the integrity of the cell membrane. The fluo-rescence intensity of living and dead microor-ganisms was determined by CSLM double-channel or three-channel scanning. An argonlaser was used to generate an excitation wave-
length of 488 nm, whereas the resultant emis-sion of living microorganisms (SYTO 9) was fil-tered through a 515–545 nm band pass filter andthat of dead ones (propidium iodide) through a575–640 nm band pass filter. On occasion, thesignal of algal autofluorescence was also col-lected by filtration with a �570 nm long passfilter. Each channel was displayed in a singlecolor (green, red, and blue, respectively).
Low-temperature scanning electron mi-croscopy. Untreated lichens were examined us-ing the low-temperature scanning electron mi-croscope (LTSEM). Small lichen fragments weremechanically fixed onto the specimen holder ofthe cryo-transfer system (Oxford CT1500),plunged into sub-cooled liquid nitrogen, andthen transferred to the preparation unit via an air-lock transfer device. The frozen specimens werecryo-fractured and transferred directly via a sec-ond air lock to the microscope cold stage, wherethey were etched for 2 min at �90 °C. After icesublimation, the etched surfaces were gold-sput-ter coated in the preparation unit. Samples weresubsequently transferred onto the cold stage ofthe SEM chamber. Fractured surfaces were ob-served using a Zeiss DSM960 SEM microscopeoperating at �135 °C with 15 kV acceleration po-tential, 10 mm working distance, and 5–10 nAprobe current.
Transmission electron microscopy. Smalllichen fragments were fixed in glutaraldehydeand post-fixed in osmium tetroxide solution, de-hydrated in a graded ethanol series, and embed-ded in Spurr’s resin following the protocol de-scribed in de los Ríos and Ascaso (2002). Ultrathinsections were post-stained with lead citrate(Reynolds, 1963) and observed in a Zeiss EM910scanning transmission electron microscope(STEM) operating in transmission electron mi-croscope mode at 100 kV.
LICHENS SURVIVE IN SPACE 447
TABLE 1. FLIGHT AND ENVIRONMENT DATA OF THE LICHENS EXPERIMENT DURING THE ESA BIOPAN-5 MISSION
LASTING 14.8 DAYS ON THE RUSSIAN SATELLITE FOTON-M2
Solar Solar IonizationTemperature constant Solar PARa UV � 170 nm Solar UV-Ab Solar UV-Bc Solar UV-Cd radiationrange (°C) hours (kJ m-2) (kJ m-2) (kJ m-2) (kJ m-2) (kJ m-2) (�Gy/day)
� 21.7 to � 21.8 58.78 133,144 22,473 18,893 2,908 671 213.36
aPhotosynthetically active radiationbUV-A: 315–400 nmcUV-B: 280–315 nmdUV-C: 200–280 nm
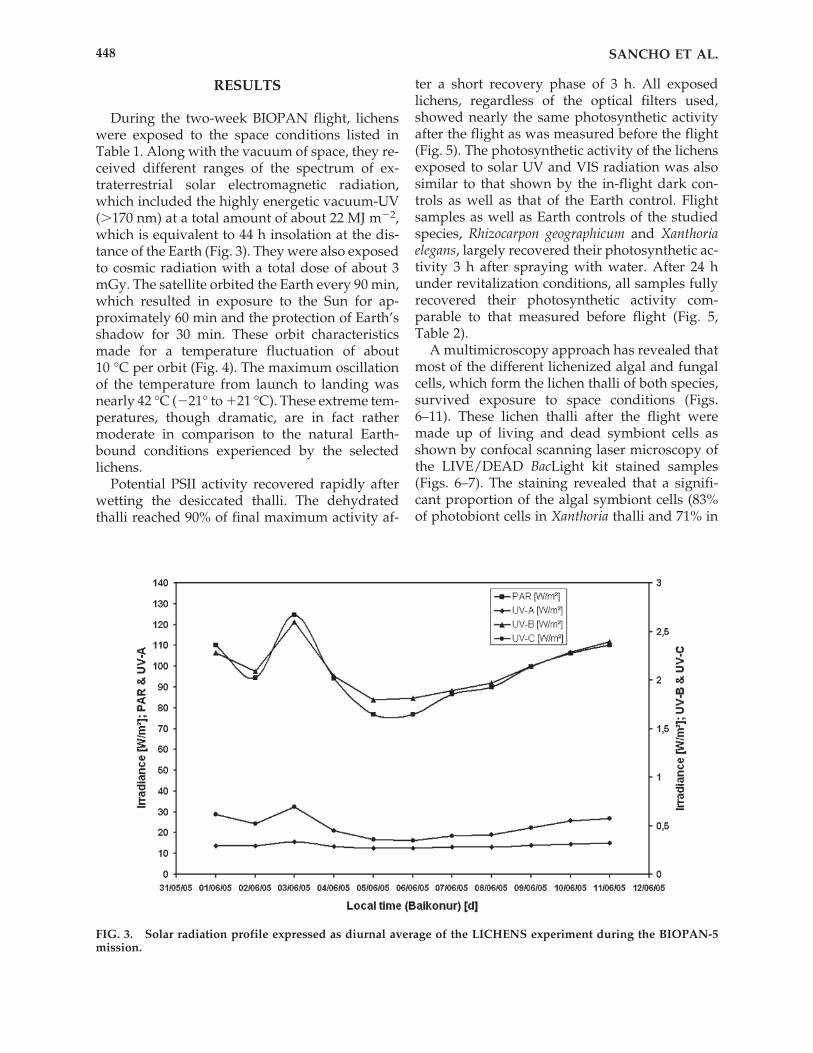
RESULTS
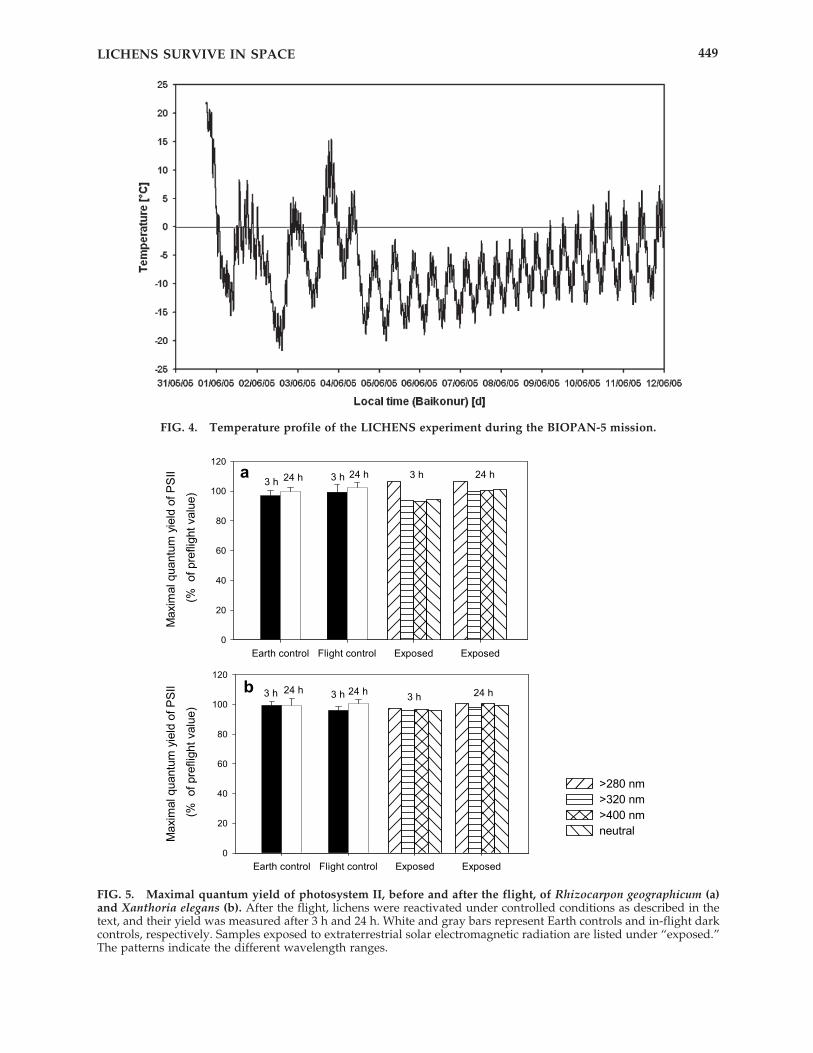
During the two-week BIOPAN flight, lichenswere exposed to the space conditions listed inTable 1. Along with the vacuum of space, they re-ceived different ranges of the spectrum of ex-traterrestrial solar electromagnetic radiation,which included the highly energetic vacuum-UV(�170 nm) at a total amount of about 22 MJ m�2,which is equivalent to 44 h insolation at the dis-tance of the Earth (Fig. 3). They were also exposedto cosmic radiation with a total dose of about 3mGy. The satellite orbited the Earth every 90 min,which resulted in exposure to the Sun for ap-proximately 60 min and the protection of Earth’sshadow for 30 min. These orbit characteristicsmade for a temperature fluctuation of about 10 °C per orbit (Fig. 4). The maximum oscillationof the temperature from launch to landing wasnearly 42 °C (�21° to �21 °C). These extreme tem-peratures, though dramatic, are in fact rathermoderate in comparison to the natural Earth-bound conditions experienced by the selectedlichens.
Potential PSII activity recovered rapidly afterwetting the desiccated thalli. The dehydratedthalli reached 90% of final maximum activity af-
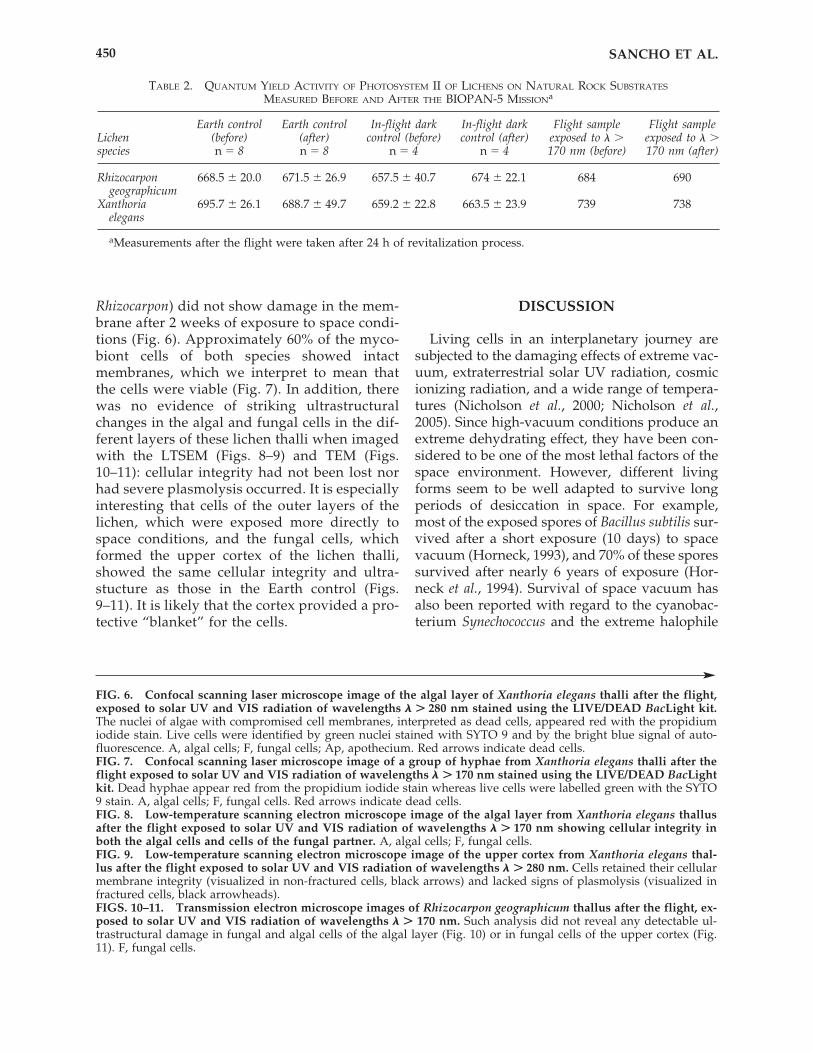
ter a short recovery phase of 3 h. All exposedlichens, regardless of the optical filters used,showed nearly the same photosynthetic activityafter the flight as was measured before the flight(Fig. 5). The photosynthetic activity of the lichensexposed to solar UV and VIS radiation was alsosimilar to that shown by the in-flight dark con-trols as well as that of the Earth control. Flightsamples as well as Earth controls of the studiedspecies, Rhizocarpon geographicum and Xanthoriaelegans, largely recovered their photosynthetic ac-tivity 3 h after spraying with water. After 24 hunder revitalization conditions, all samples fullyrecovered their photosynthetic activity com-parable to that measured before flight (Fig. 5,Table 2).
A multimicroscopy approach has revealed thatmost of the different lichenized algal and fungalcells, which form the lichen thalli of both species,survived exposure to space conditions (Figs.6–11). These lichen thalli after the flight weremade up of living and dead symbiont cells asshown by confocal scanning laser microscopy ofthe LIVE/DEAD BacLight kit stained samples(Figs. 6–7). The staining revealed that a signifi-cant proportion of the algal symbiont cells (83%of photobiont cells in Xanthoria thalli and 71% in
SANCHO ET AL.448
FIG. 3. Solar radiation profile expressed as diurnal average of the LICHENS experiment during the BIOPAN-5mission.
LICHENS SURVIVE IN SPACE 449
FIG. 4. Temperature profile of the LICHENS experiment during the BIOPAN-5 mission.
FIG. 5. Maximal quantum yield of photosystem II, before and after the flight, of Rhizocarpon geographicum (a)and Xanthoria elegans (b). After the flight, lichens were reactivated under controlled conditions as described in thetext, and their yield was measured after 3 h and 24 h. White and gray bars represent Earth controls and in-flight darkcontrols, respectively. Samples exposed to extraterrestrial solar electromagnetic radiation are listed under “exposed.”The patterns indicate the different wavelength ranges.
Rhizocarpon) did not show damage in the mem-brane after 2 weeks of exposure to space condi-tions (Fig. 6). Approximately 60% of the myco-biont cells of both species showed intactmembranes, which we interpret to mean thatthe cells were viable (Fig. 7). In addition, therewas no evidence of striking ultrastructuralchanges in the algal and fungal cells in the dif-ferent layers of these lichen thalli when imagedwith the LTSEM (Figs. 8–9) and TEM (Figs.10–11): cellular integrity had not been lost norhad severe plasmolysis occurred. It is especiallyinteresting that cells of the outer layers of thelichen, which were exposed more directly tospace conditions, and the fungal cells, whichformed the upper cortex of the lichen thalli,showed the same cellular integrity and ultra-stucture as those in the Earth control (Figs.9–11). It is likely that the cortex provided a pro-tective “blanket” for the cells.
DISCUSSION
Living cells in an interplanetary journey aresubjected to the damaging effects of extreme vac-uum, extraterrestrial solar UV radiation, cosmicionizing radiation, and a wide range of tempera-tures (Nicholson et al., 2000; Nicholson et al.,2005). Since high-vacuum conditions produce anextreme dehydrating effect, they have been con-sidered to be one of the most lethal factors of thespace environment. However, different livingforms seem to be well adapted to survive longperiods of desiccation in space. For example,most of the exposed spores of Bacillus subtilis sur-vived after a short exposure (10 days) to spacevacuum (Horneck, 1993), and 70% of these sporessurvived after nearly 6 years of exposure (Hor-neck et al., 1994). Survival of space vacuum hasalso been reported with regard to the cyanobac-terium Synechococcus and the extreme halophile
SANCHO ET AL.450
TABLE 2. QUANTUM YIELD ACTIVITY OF PHOTOSYSTEM II OF LICHENS ON NATURAL ROCK SUBSTRATES
MEASURED BEFORE AND AFTER THE BIOPAN-5 MISSIONa
Earth control Earth control In-flight dark In-flight dark Flight sample Flight sampleLichen (before) (after) control (before) control (after) exposed to � � exposed to � �species n � 8 n � 8 n � 4 n � 4 170 nm (before) 170 nm (after)
Rhizocarpon 668.5 20.0 671.5 26.9 657.5 40.7 674 22.1 684 690geographicum
Xanthoria 695.7 26.1 688.7 49.7 659.2 22.8 663.5 23.9 739 738elegans
aMeasurements after the flight were taken after 24 h of revitalization process.
FIG. 6. Confocal scanning laser microscope image of the algal layer of Xanthoria elegans thalli after the flight,exposed to solar UV and VIS radiation of wavelengths � � 280 nm stained using the LIVE/DEAD BacLight kit.The nuclei of algae with compromised cell membranes, interpreted as dead cells, appeared red with the propidiumiodide stain. Live cells were identified by green nuclei stained with SYTO 9 and by the bright blue signal of auto-fluorescence. A, algal cells; F, fungal cells; Ap, apothecium. Red arrows indicate dead cells.FIG. 7. Confocal scanning laser microscope image of a group of hyphae from Xanthoria elegans thalli after theflight exposed to solar UV and VIS radiation of wavelengths � � 170 nm stained using the LIVE/DEAD BacLightkit. Dead hyphae appear red from the propidium iodide stain whereas live cells were labelled green with the SYTO9 stain. A, algal cells; F, fungal cells. Red arrows indicate dead cells.FIG. 8. Low-temperature scanning electron microscope image of the algal layer from Xanthoria elegans thallusafter the flight exposed to solar UV and VIS radiation of wavelengths � � 170 nm showing cellular integrity inboth the algal cells and cells of the fungal partner. A, algal cells; F, fungal cells.FIG. 9. Low-temperature scanning electron microscope image of the upper cortex from Xanthoria elegans thal-lus after the flight exposed to solar UV and VIS radiation of wavelengths � � 280 nm. Cells retained their cellularmembrane integrity (visualized in non-fractured cells, black arrows) and lacked signs of plasmolysis (visualized infractured cells, black arrowheads).FIGS. 10–11. Transmission electron microscope images of Rhizocarpon geographicum thallus after the flight, ex-posed to solar UV and VIS radiation of wavelengths � � 170 nm. Such analysis did not reveal any detectable ul-trastructural damage in fungal and algal cells of the algal layer (Fig. 10) or in fungal cells of the upper cortex (Fig.11). F, fungal cells.

LICHENS SURVIVE IN SPACE 451
FIGS. 6–11.

Haloarcula (Mancinelli et al., 1998). Pluricellularorganisms such as nematodes have also beenshown to be desiccation resistant in exobiologyexperiments (Bertolani et al., 2001). This ability tosurvive desiccation may be related to the pres-ence of sugars and polyalcohols, which stabilizethe cellular macromolecules during extreme de-hydration processes (Crowe and Crowe, 1992).Lichens are rich in sugar alcohols, which are as-sumed to be the basis of their remarkable desic-cation tolerance (Kappen and Valladares, 1999).In endolithic Antarctic biofilms, extracellularpolymeric substances (EPS) contribute signifi-cantly to the intercellular structure and have beenproposed as a strategy to avoid desiccation stress(de los Ríos et al., 2004; de los Ríos et al., 2005).
Ultraviolet radiation has been considered themost damaging factor for living organisms inouter space. The highly desiccation-tolerantspores of Bacillus subtilis are rapidly killed whenexposed to the full spectrum of extraterrestrialUV radiation (Horneck et al., 1984; Horneck et al.,2001; Nicholson et al., 2005). Similar results havebeen obtained for other organisms tested in exo-biology experiments, including viruses, bacteria,fungi, and nematodes (Nicholson et al., 2000;Rothschild and Mancinelli, 2001). The green al-gae Chlamydomonas reinhardtii tested in BIOPANshowed a very low tolerance to UV radiation. Allthe wild types in that experiment died, even be-hind a 5% transmission filter. Only when the so-lar UV was blocked or the total transmission wasreduced to not more than 1% were survivorsfound (René Demets, personal communication).Therefore, the observed full recovery of the pho-tosynthetic activity of the lichenized green algae,even in those exposed to 99% of the extraterres-trial sunlight, was unexpected. The recovery ofphotosynthetic activity can be partially explainedby the combined effects of a thick and dense up-per cortex and a high concentration of lichen sub-stances, which also efficiently screened the harm-ful radiation. A direct relationship betweenpigment production and light intensity has beenwell established for alpine and polar lichens (Ah-madjian and Jacobs, 1987; Kappen, 1993). Thebright colors of the studied species Rhizocarpongeographicum and Xanthoria elegans are due to rhi-zocarpic and parietin phenolic acids that accu-mulate in the upper cortex and may be importantin filtering UV radiation (Solhaug et al., 2003;Gauslaa and Solhaug, 2004). The high tolerance
of some lichens to UV exposure in a desiccatedstate has been recently attributed to an efficientthermal dissipation mechanism of light energy(Heber et al., 2006).
Cosmic radiation, mainly the heavy ions ofhigh charge Z and high-energy particles (HZE),could easily penetrate even the thick lichen cor-tex and damage or kill cells (Nicholson et al.,2000). However, due to their low fluence, theirdeleterious effect strongly depends on exposuretime. It has been calculated that, with regard tocosmic radiation, spores of Bacillus subtilis couldsurvive in space for several hundred thousandyears (Horneck et al., 2001).
The temperature range of the BIOPAN duringthe flight fluctuated from �20 to �20 °C. Sincepolar-alpine lichens are tolerant of a much widernatural temperature range (Kappen, 1993), nodeleterious effect of temperature was expected.
These findings suggest that some lichenspecies may be pre-adapted to cope with the ex-treme conditions of outer space. Lichen chem-istry and thallus structure seem to provide anadequate shield to the effects of both UV radia-tion and high vacuum. Even though membraneswere damaged in some cells, as determined byLIVE/DEAD staining, the rest of the cells re-mained capable of maintaining the photosyn-thetic productivity of the lichen. Moreover, af-ter extreme dehydration induced by highvacuum, lichens proved to be able to recover, infull, their metabolic activity in a remarkablyshort time (24 h). It is difficult, however, to ex-trapolate these short-term experimental resultsto a long-term scenario of thousands or millionsof years. Accumulated galactic cosmic radiationeffects in a dose of 10–50 kGy have been postu-lated as sufficient to completely sterilize the sur-face of a meteor (Clark, 2001). The value of thiscalculation based on laboratory measurementsin a long-term space scenario is equivocal (Hor-neck et al., 2001).
Since we did not detect any clear physiologicalor structural degradation in the lichens after thetwo-week exposure to space conditions, we arenot able to give any estimate about the half-lifeof these lichen species in space. Our findings in-dicate, therefore, that these types of lichens havethe capability to survive an ephemeral ejectioninto space. How long they will retain their via-bility in the interplanetary environment remainsa challenge for future experiments.
SANCHO ET AL.452

ACKNOWLEDGMENTS
Our research was supported by grantsREN2003-07366-C01/02, CGL2006-12179, andBOS2003-02418 of the Plan Nacional I�D. The authors would like to thank Fernando Pinto(CCMA), Charo Santos (CCMA), Sara Paniagua(CCMA), and Xavier Calomarde (Udl) for tech-nical assistance. Logistical and technical supportwas kindly provided by INTA (Madrid, Spain)and DLR (Cologne, Germany). We are indebtedto René Demets and Pietro Baglioni (ESA) for con-tinuous support and critical reading of the man-uscript and to Lillian and Alejandra Mora for re-vising the English text.
REFERENCES
Ahmadjian, V. and Jacobs, J.B. (1987) Studies on the de-velopment of synthetic lichens. Bibl. Lichenol. 25, 47–58.
Arrhenius, S. (1903) Die Verbreitung des Lebens im Wel-tenraum. Die Umschau 7, 481–485.
Baglioni, P., Sabbatini, M., and Horneck, G. (2007) Astro-biology experiments in low earth orbit—facilities, in-strumentation and results. In Complete Course in Astro-biology, edited by P. Rettberg and G. Horneck,Wiley-VCH, Berlin, pp. 273–319.
Bertolani, R., Rebecchi, L., Jönsson, K.I., Guidetti, R., Bor-sari, S., and Altiero, T. (2001) Tardigrades as a modelfor experiences of animal survival in space. MSSU: Mi-crogravity and Space Station Utilization 2, 211–212.
Burchell, M.J., Mann, J.R., and Bunch, A.W. (2004) Sur-vival of bacteria and spores under extreme shock pres-sures. Mon. Not. R. Astron. Soc. 352, 1273–1278.
Clark, B. (2001) Planetary interchange of bioactive mate-rial: probability factors and implications. Orig. Life Evol.Biosph. 31, 185–197.
Crowe, L.M. and Crowe, J.H. (1992) Anhydrobiosis: astrategy for survival. Adv. Space Res. 12, 239–247.
de la Torre, R., Horneck, G., Sancho, L.G., Scherer, K.,Facius, R., Urlings, T., Rettberg, P., Reina, M., and Pin-tado, A. (2002) Photoecological characterization of anepilithic ecosystem at high mountain locality (centralSpain). In Proceedings of the Second European Workshopon Exo/Astrobiology, Graz, Austria, ESA SP-518,ESA_ESTEC, Noordwijk, the Netherlands, pp. 443–444.
de los Ríos, A., and Ascaso, C. (2002) Preparative tech-niques for transmission electron microscopy and con-focal laser scanning microscopy of lichens. In Protocolsin Lichenology, edited by I. Kranner, R.P. Beckett, andA.K. Varma, Springer, Berlin, pp. 87–151.
de los Ríos, A., Wiezchos, J., Sancho, L.G. and Ascaso, C.(2004) Exploring the physiological state of continentalAntarctic endolithic microorganisms by microscopy.FEMS Microbiol. Ecol. 50, 143–152.
de los Ríos, A., Wierzchos, J., Sancho, L.G., Green, T.G.A.and Ascaso, C. (2005) Ecology of endolithic lichens col-onizing granite in continental Antarctica. Lichenologist37, 383–395.
Demets, R., Schulte, W., Baglioni, P. (2005) The past, pre-sent and future of Biopan, Adv. Space Res. 36, 311–316.
Fajardo-Cavalzos, P., Link, L., Melosh, H.J., and Nichol-son, W.L. (2005) Bacillus subtillis spores on artificialmeteorites survive hypervelocity atmospheric entry:implications for Lithopanspermia. Astrobiology 5,726–736.
Friedmann, E.I. (1982) Endolithic microorganisms in theAntarctic cold desert. Science 215, 68–74.
Gauslaa, Y. and Solhaug, K.A. (2004) Photoinhibition inlichens depends on cortical characteristics and hydra-tion. Lichenologist 36, 133–143.
Green, T.G.A., Schroeter, B., and Sancho, L.G. (1999) Plantlife in Antarctica. In Handbook of Functional Plant Ecol-ogy, edited by F.I. Pugnaire and F. Valladares, MarcelDekker, New York, Basel, pp. 495–543.
Heber, U., Lange, O.L., and Shuvalov, V.A. (2006) Con-servation and dissipation of light energy as comple-mentary process: homoiohydric and poikilohydric au-totrophs. J. Exp. Bot. 57, 1211–1223.
Horneck, G. (1993) Responses of Bacillus subtilis spores tothe space environment: results from experiments inspace. Orig. Life Evol. Biosph. 23, 37–52.
Horneck, G., Bücker, H., Reitz, G., Requardt, H., Dose, K.,Martens, K.D., and Weber, P. (1984) Microorganisms inthe space environment. Science 225, 226–228.
Horneck, G., Bücker, H., and Reitz, G. (1994) Long-termsurvival of bacterial spores in space. Adv. Space Res. 14,41–45.
Horneck, G., Rettberg, P., Reitz, G., Wehner, J., Es-chweiler, U., Strauch, K., Panitz, C., Starke, V., andBaumstark-Khan, C. (2001) Protection of bacterialspores in space, a contribution to the discussion onPanspermia. Orig. Life Evol. Biosph. 31, 527–547.
Kappen, L. (1993) Lichens in the Antarctic region. InAntarctic Microbiology, edited by E.I. Friedman, Wiley,New York, pp. 433–490.
Kappen, L. (2000) Some aspects of the great success oflichens in Antarctica. Antarct. Sci. 12, 314–324.
Kappen, L. and Valladares, F. (1999) Opportunisticgrowth and desiccation tolerance: the ecological successof poikilohydrous autotrophs. In Handbook of FunctionalPlant Ecology, edited by F.I. Pugnaire and F. Valladares,Marcel Dekker, New York, Basel, pp. 121–194.
Mancinelli, R.L. and Klovstad, M. (2000) Martian soil andUV radiation: microbial viability assessment on space-craft surfaces. Planet. Space Sci. 48, 1093–1097.
Mancinelli, R.L., White, M.R., and Rothschild, L.J. (1998)Biopan survival I: exposure of the osmophiles Syne-chococcus sp. (Nageli) and Haloarcula sp. to the space en-vironment. Adv. Space Res. 22, 327–334.
Maxwell, K. and Johnson, G.N. (2000) Chlorophyll fluo-rescence—a practical guide. J. Exp. Bot. 51, 659–668.
Nicholson, W.L., Munakata, N., Horneck, G., Melosh, H.J.,and Setlow, P. (2000) Resistance of Bacillus endospores
LICHENS SURVIVE IN SPACE 453

to extreme terrestrial and extraterrestrial environments.Microbiol. Mol. Biol. Rev. 64, 548–572.
Nicholson, W.L., Schuerger, A.C., and Setlow, P. (2005)The solar UV environment and bacterial spore UV re-sistance: considerations for Earth-to-Mars transport bynatural processes and human spaceflight. Mutat. Res.571, 249–264.
Purvis, O.W., Bailey, E.H., McLean, J., Kasama, T., andWilliamson, B.J. (2004) Uranium biosorption by thelichen Trapelia involuta at a uranium mine. Geomicrobiol.J. 21, 159–167.
Rettberg, P., Eschweiler, U., Strauch, K., Reitz, G., Hor-neck, G., Wänke, H., Brack, A., and Barbier, B. (2002)Survival of microorganisms in space protected by me-teorite material: results of the experiment EXOBIOLO-GIE of the PERSEUS mission. Adv. Space. Res. 30,1539–1545.
Reynolds, S. (1963) The use of lead citrate at high pH asan electron-opaque stain in electron microscopy. J. CellBiol. 17, 200–211.
Richter, H. (1865) Zur Darwinschen Lehre. Schmidts Jahrb.Ges. Med. 126, 243–249.
Rothschild, L.J. and Mancinelli, R.L. (2001) Life in extremeenvironments. Nature 409, 1092–1101.
Schreiber, U., Bilger, W., and Neubauer, C. (1994) Chloro-phyll fluorescence as a nonintrusive indicator for rapidassessment of in vivo photosynthesis. In Ecophysiologyof Photosynthesis, edited by E-D. Schulze and M.M. Cad-well, Springer, Berlin, Heidelberg, New York, Vol. 1,pp. 49–70.
Solhaug, K.A., Gauslaa, Y., Nybakken, L., and Bilger, W.(2003) UV-induction of sun-screening pigments inlichens. New Phytol. 158, 91–100.
van Kooten, O. and Snel, J.F.H. (1990) The use of chloro-phyll fluorescence nomenclature in plant stress physi-ology. Photosynth. Res. 25, 147–150.
Address reprint requests to:L.G. Sancho
Departamento de Biología Vegetal IIFacultad de Farmacia
Universidad ComplutensePlaza Ramón y Cajal s/n
28040 Madrid, Spain
E-mail: [email protected]
SANCHO ET AL.454