document
TRANSCRIPT

Zbl. Bakt. Hyg., 1. Abt, O rig. C 3, 101-109 (1982)
Institute of Microbiology, University of Heidelberg, Federal Republic of Germany
Polypeptide Synthesis in Escherichia coli Directed by Cloned
Methanobrevibacter arboriphilus DNA*
CLAUS BOLLSCHWEILER and ALBRECHT KLEIN
Received October 16, 1981
Summary
Methano brevibacter arboriph ilus DNA was isolated after treatment of the cells withbacitracin in a sucrose containing growth medium. HindIlI restriction fragments of theDNA were inserted into an expression vector plasmid. The newly constructed plasmidswhen introduced into E. coli gave rise to the synthesis of polypeptides coded for by themethanogen DNA. The expression of genetic information from the MethanobrevibacterDNA in E. coli is efficient as judged by the total molecular weight of the polypeptidesfound as compared to the length of the Me tha nobrevibacter DNA carried on the plasmids.
Key words: Methanobrevibacter DNA - Cloning in E. coli - Heterologous expression - T ranscription signals
Introduction
Methanogen ic bacteria form a subgroup of the archaeba cteria (Balch et aI.,1979) which strikingly differ from the other procar yotes in a num ber of respects,notably in cell wall and cell memb rane constituents and stru cture (Kandler andKonig, 1978; Kandler, 1979; M akula and Singer , 1978; T ornabene and Langworthy, 1978) RNA pol ymerase (Z illig et aI., 1980) 5S (pieler et aI., 1981) and165 (Fox et aI., 1977) ribosomal RNA sequences. On the other hand, it is remarkabl e th at a common sequence at the 3' -end of the 165 rRNA has been preservedamo ng eubacteria and archaebacteri a, which is thought to be involved in bindingof the messenger RNA to the ribo som e (Fox et aI., 1977; Steitz, 1978).
Methanogens play a key role in anaerobic digestion of organic matter. Theyperform the terminal step leadi ng from metabolic end pr oducts of fermentativeorgan isms like acetic or formic acid, carbon dioxide, hydrogen or methylamineto methane, thu s transferring electron s to Cj-units.
~ Paper given at the First International Workshop on Archaebacteria, Miinchen, June 27to July 1, 1981.
Abbreviation : kb = kilobases.

102 C. Bollschweiler and A. Klein
In natural environment methanogenic and eubacteria live in syntrophy forminga food chain starting from the depolymerization of biopolymers to methane formation. This occurs, e. g. in the rumen and in anaerobic digesters during wastetreatment.
Syntrophy of pure cultures of an acetogenic fermentative eubacterium andmethanogens has been described (Winter and Wolfe, 1979; 1980). More recentlysyntrophic cultures of mixed acid producing eubacteria among them Escherichiacoli, and various methanogens have been reported to shift the metabolism of thefermenting bacteria due to the removal of electrons from the equilibrium in thecourse of methanogenesis (Winter, 1980).
We are interested in the possibility of establishing the methanogenic pathwayin a fermentative eubacterium by introducing genetic material from a methanogeninto it. As a first step we have introduced genomic material from Methanobrevibacter arboriphilus into E. coli and have looked for the synthesis of polypeptidescoded for by the Methanobrevibacter DNA. We show that such polypeptide formation does occur and that the Methanobrevibacter genes might be transcribedfrom their own promoters.
Materials and Methods
Bacteria and Plasmid
Methanobrevibacter arboriphilus AZ (DSM 744) was obtained from the Deutsche Sammlung fiir Mikroorganismen, G6ttingen. E. coli C600 cm+ is a nonrestricting strain fromour collection which confers K12 specific DNA modification to incoming DNA. E. coliC600 cm+Fiq was obtained by F-duction from a mating with BMH408Fi\ a gift fromDr. B. Muller-Hill, Cologne. The Fiq plasmid leads to a high level of lac repressor in itshost cell. E. coli CSR603Fiq was similarly constructed using E. coli CSR603 uvrA recA
pEx150
Fig. 1. Relevant features of the expression vector pEx150 (Weiher, 1980) used in this study.The amp and tet genes confer ampicillin and tetracycline resistance, respectively, to thecarrier cells. The tet gene expression depends on the intactness of the promoter ptet whichis destroyed upon insertion of DNA into the HindIII restriction site. Alternatively the inducible lac promoter (piac) can yield transcription of the tet gene in the absence of interrupting signals on the insert DNA. p1 is a promoter recently discovered (Stuber and Bujard,1981) whose transcripts reach beyond the HindIII site in the direction opposite to that ofplac.

Methanobrevibacter Polypeptide Synthesis in E. coli 103
phd (Sancar et a!', 1979) obtained from the E. coli Genetic Stock Center, New Haven,Conn. USA.
Plasmid pEx150 (Fig. 1) (Weiher, 1980) given to us by Dr. H.Schaller, is a derivative ofpBR322 (Bolivar et aI., 1977) carrying the inducible lac promoter next to the EcoRI site.The promoter points in the direction of the tet gene.
Chemicals and Enzymes
The chemicals and enzymes are listed with their suppliers in parentheses: Ampicillin(Serva, Heidelberg), tetracycline (Serva), 3liS-methionine (Amersham-Buchler, Braunschweig), DNAse I (Serva), lysozyme (Sigma, Miinchen), DNA polymerase I (gift ofDr. H.Schaller), T4 polynucleotide ligase (gift of Dr. R.Herrmann), pronase (Serva),RNAse (Sigma).
Methanobrevibacter DNA preparation
Methanobrevibacter arboriphilus was grown anaerobically at 37°C according to theprocedures of Balch et a!' (1979). At a density of 1 X 10' cells/ml sucrose was added at afinal concentration of 20%. The cells were then grown to 10B/ml, bacitracin was addedat 700 ,ug/ml and growth continued for another 15 h. The cells were harvested, resuspendedin 10 mmol/I Tris, 1 mmol/I EDTA pH 7.8 at 200-fold concentration and lysed by theaddition of 1% sodium dodecyl sulfate at 60°C for 15 min. DNA was then prepared byroutine techniques after digestion with 200,ug/ml pronase for 1 hour and subsequentphenol extraction.
Restriction, ligation and transformation
HindlII restriction of DNA followed the procedure given by the enzyme supplier. DNAligation and transformation has been described previously (Kuypers et aI., 1980).
DNA/DNA hybridization
The technique described by Southern (1975) was used with minor modifications. Radioactive labeling of the DNA by combined DNAse/DNA polymerase treatment followed themethod of Rigby et al. (1977).
3liS-labeling
The method of Sancar et al. (1979) was used for labeling plasmid coded proteins. Thelabeled cells were lysed and analyzed by sodium dodecyl sulfate polyacrylamide gclelccrrophoresis and fluorography as described earlier (Kuypers et aI., 1980).
Results
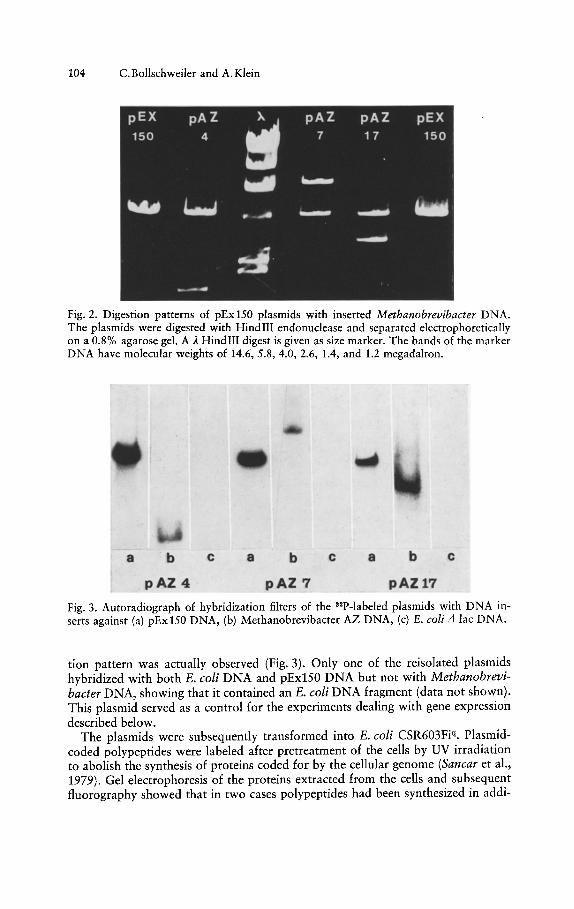
Insertion of HindIII fragments of Methanobrevibacter DNA into the HindIIIrestriction site of pEx150 and subsequent transformation of E. coli e600 r" cellsyielded ampicillin resistant tetracycline sensitive clones. The plasmids found inthese cells were analyzed by HindIII restriction endonuclease digestion. Fig. 2shows that fragments of various lengths had been incorporated into the vector.
In order to verify that the inserts were indeed Methanobrevibacter DNA andnot contaminating E. coli DNA left in the plasmid preparation, the isolatedplasmids were labeled with 32p and hybridized against HindIII fragments of totalMethanobrevibacter DNA, E. coli LI lac DNA, or pEx150 DNA. pEx150 derivatives containing MethanobrevibacterDNA were expected to hybridize with pEx150and Methanobrevibacter DNA and not with E. coli LI lac DNA. This hybridiza-
104 C.Bollschweiler and A. Klein
pEX pAZ
~pAZ pAZ pEX
150 4 7 17 150... ----.. ~ .... - ..... ....-::j.
_..Fig. 2. Digestion patterns of pEx 150 plasmids with inserted M ethanobrevibacter DNA .Th e plasmids were digested with HindIII endonuclease and separated electrophoretic allyon a 0.8% agaro se gel. A A. HindIII digest is given as size marker. Th e bands of the markerDNA have molecular weights of 14.6, 5.8,4.0, 2.6, 1,4, and 1.2 megadalron.
a b
pAZ 4
c a b
pAZ 7
c a b
pAZ17
c
Fig. 3. Autoradiograph of hybridization filters of the 32P-labeled plasmids with DNA inserts against (a) pEx1 50 DNA, (b) Methanobrevibacter AZ DNA, (c) E. coli LJ lac DNA .
tion pattern was actually observed (Fig. 3). Only one of the reisolated plasmidshybridized with both E. coli DNA and pEx150 DNA but not with M ethanobrevibacter DNA, showing that it cont ained an E. coli DNA fragment (data not shown).This plasmid served as a control for the experiments dealing with gene expressiondescribed below.
The plasmids were subsequently transformed into E. coli CSR603FiQ. Plasmidcoded polypeptides were labeled after pretreatment of the cells by UV irradiationto abolish the synthesis of proteins coded for by the cellular genome (Sancar et a1. ,1979). Gel electrophoresis of the proteins extracted from the cells and subsequentfluorography showed that in two cases polypeptides had been synthesized in add i-

Methanobrevibacter Polypeptide Synthesis in E. coli 105
tion to those found in cells containing the vector plasmid pEx150 alone withoutMethanobrevibacter DNA inserts (Fig.4). The insert length of the two plasmidspAZ7 and pAZ17 was 7 and 4.4 kb respectively. A third plasmid pAZ4 with aninsert of 0.5 kb did not exhibit a Methanobrevibacter specific polypeptide synthesisin E. coli (Fig. 4). Induction or repression of the lac promoter adjacent to the insert(compare Fig. 1) did not have an influence on the expression of the methanogengenes (compare lanes a and b in Fig. 4).
43- - -
a b
pAZ17
a b
pEX150
..
a b
pAZ 7
a b
pAZ4
12-
20
17-
30-
Fig. 4. Fluorograph of asS-labeled polypeptides synt hesized by E. coli CSR 603 Fiq cellscarrying pEx 150 or pEx 150 plasmids with different Me thanobrevibacter DNA inserts(compare Figs. 2 and 3). The pol ypeptides were separated by pol yacryl amide gel electrophoresis in the presence of sodium dodecyl sulfate after extraction from the cells. Label ing was performed with (a) or without (b) induction of the lac promoter by 0.1 mmol /lisopropyl-ji-th iogalactoside. Molecular weights of marker pr oteins used on the same gelare given in kilodalton.
Next we tried to find out whether the signals used for transcnpnon of theMethanobrevibacter DNA, the promoters recognized by the E. coli RNA polymerase, were located on the pEx150 or on the M ethanobrevibacter insert part ofthe new plasmids. For this purpose the inserts from pAZ7 and pAZ17 wereisolated and reinserted in inverse orient ation into the same restriction sites ofpEx150. All four different plasmids were then transformed into E. coli C600Fiqand the clones checked for tetracycline resistance after induction of the lac promoter. In one case tetr acycline resistance did reappear (Table 1), showing thattranscription through the insert was possible. In order to find out whether in this

106 C. Bollschweiler and A.Klein
Table 1. Tetracycline sensitivityof cells carrying pEx ISO-derivatives containing Methanobrevibacter DNA
plasmid plac induction tet sensitivity
pAZ7 sensitive+ sensitive
pAZ7/2 sensitive+ resistant
pAZ 17 sensitive+ sensitive
pAZ 17/2 sensitive+ sensitive
The growth of cells carrying pAZ7, pAZ17 or their derivatives with inverted insertspAZ7/2 or pAZ 17/2, respectively, was tested on solid medium containing 10 .ug/ml tetracycline under induction of plac with 0.1 mrnol/l isopropyl-ji-thiogalacroside or withoutinduction.
case the Methanobrevibacter DNA coded polypeptide synthesis was also dependent on the induction of the lac promoter the plasmids were transformed intoE. coli CSR603Fiq and polypeptide synthesis with and without induction wasmeasured. It was seen that, again, expression of the Methanobrevibacter polypeptides did not depend on the induction of the lac promoter even in the case whichhad shown an inducible tetracycline resistance (data not shown).
Discussion
Due to fundamental differences in the protein synthesizing apparatus of archaebacteria and eubacteria, one might expect the expression of genomic materialfrom methanobacteria in E. coli to be impossible. Our results show that Methanebrevibacter genomic material cloned in E. coli is transcribed and translated in thishost. This requires recognition signals for the E. coli RNA polymerase on theDNA and for the E. coli ribosomes on the mRNA level. The construction of theplasmid used in this study allowed for the use of E. coli promoters adjacent tothe inserted Methanobrevibacter DNA for its transcription.
Our finding (Table 1) that induction of the lac promoter leads to tetracyclineresistance in plasmid pAZ7/2, i. e. to an expression of a gene separated from thepromoter by a 7 kb Methanobrevibacter DNA insert, shows that this DNA couldbe transcribed from an E. coli promoter. However, the constitutive expression ofthe Methanobrevibacter specific polypeptides in all cases studied indicates that thelac promoter did not playa role in their synthesis. Therefore signals on the Methanobrevibacter DNA must be recognized as transcription start signals by E. coliRNA polymerase. This is symbolized in Fig. 5, which gives one possible interpretation of the results represented in Table 1. The scheme does not discriminatebetween termination on the transcriptional and translational level, i, e. polareffects. We must postulate that the Methanobrevibacter genes are transcribedfrom the opposite strand compared to the ret gene in pAZ7/2 which yields indu-

Methanobrevibacter PolypeptideSynthesis in E. coli 107
pAZ 7plae - - - - --t>
• I to iJ Ip1..
--------
pAZ7/2<3------
~ ~ " ..------------..
..----~
~ I b ~'-II------+--
-------pAZ17
•<3---- --
~ I ~i__----C3C3-+--_-_+_-----pAZ17/2
--·-tMbr.DNA insert -+-- tet --t---
Fig. 5. Schematic interpretation of the results presented in Fig. 4 and Table 1. The figureshows the region of pEx150 in between plac and p1 with Methallobrevibacter (Mbr.)DNA inserted in either orientation into the HindIII site. The induced state of plac is represented. Since transcription starting from p1 does not influence the interpretation transcripts starting at this promoter are not shown.-'-plasmid promoter -t>-- promoter on Mbr. DNA,----zJ- transcription/translation stop signal,--.. transcript starting at plasmid promoter,---t> transcript starting at Mbr. DNA promoter.The directionsof the arrow heads indicate the directions in which the signals are functional.
cible tetracycline resistance. Consequently a promoter on the insert DNA must berecognized on pAZ7 which shows constitutive expression of genes on the insert.The tetracycline sensitivity in this case indicates that the mRNA is terminatedbefore it reaches the tet gene or translation of this gene is prevented. For plasmidspAZ17 and pAZ17/2 we have to assume that transcription and/or translation stopsoccur within the insert downstream of the lac promoter and in front of the tet gene,irrespective of the orientation of the inserted DNA. Once again, the simplest explanation for the expression of the Methanobrevibacter genes in this case is theassumption of transcription start signals on the insert DNA. Further investigationsare being performed to directly demonstrate the expression of a gene functionunder the control of such a promoter element. Preliminary results indicate thatsuch promoters occur quite frequently (Klein and Bollschweiler, unpublishedresults).
From Figs. 1 and 3 it is possible to estimate the proportion of the genetic information contained on the insert actually translated into polypeptides. The totalapparent molecular weight of the polypeptide chains formed from pAZ17 amountsto 44 kilodalton or approximately 400 amino acids coded by a DNA polynucleotide chain of approximately 1.5 megadalton molecular weight or 2300 basepairs.This accounts for roughly 50 per cent of the maximal coding capacity based onthe assumption that the total insert would consist of structural genes. Therefore,the given percentage is certainly a minimal estimate. The same figures for pAZ7are 110 kilodalton polypeptide chains corresponding to 103 amino acids per 4.5

108 C.Bollschweiler and A.Klein
megadalton or approximately 7000 nucleotide pairs of methanogen DNA or 45per cent of the maximal coding capacity. These figures compare to 64 per centof theoretical coding capacity expressed in the plasmid unintentionally isolatedwith an E. coli 2.5 megadalton insert mentioned in the results. Thus the Metbanobrevibacter DNA appears to be almost equally well expressed as E. coli DNA.
As mentioned in the introduction ribosome binding signals on messenger RNAmolecules may be similar in eubacteria and archaebacteria allowing for the translation of polypeptides from archaebacterial messenger in eubacteria. The apparentexistence of transcription start signals on Methanobrevibacter DNA recognizableby E. coli RNA polymerase had not been expected due to the known structuraldifference between the RNA polymerases. It remains to be seen whether thenucleotide sequences recognized by the E. coli RNA polymerase represent trueMethanobrevibacter promoters also used by the homologous RNA polymerase.
References
Balch, W.E., Fox, G.E., Magrum, L.]., Woese, C.R., Wolfe, R.S.: Methanogens: Reevaluation of a Unique Biological Group. Microbiol. Rev. 43, 260-293 (1979)
Bolivar, F., Rodriguez, R. L., Greene, P. Y., Betlacb, M. C., Heynecker, H. L., Boyer, H. W.,Crosa, Y.H., Falkow, S.: Construction and characterization of new cloning vehicles.II. A multiple cloning system. Gene 2,95-113 (1977)
Fox, G. F., Magrum, L.J., Balch, W. E., Wolfe, R. S., Woese, C. R.: Classification of methanogenic bacteria by 16S ribosomal RNA characterization. Proc. nat. Acad. Sci. (Wash.) 74,4537-4541 (1977)
Kandler, 0.: Zellwandstrukturen bei Methanbakterien. Zur Evolution der Prokaryonten.Naturwissenschaften 66, 95-105 (1979)
Kandler, 0., Konig, H.: Chemical composition of peptidoglycan-free cell walls of methanogenic bacteria. Arch. Microbiol. 118, 141-152 (1978)
Kuypers, B., Reiser, W., Klein, A.: Cloning of the replication gene 0 of E. coli bacteriophage Lambda and its expression under the control of the lac promoter. Gene 10, 195203 (1980)
Makula, R. A., Singer, M. E.: Ether-containing lipids in methanogenic bacteria. Biochem,Biophys. Res. Commun. 82, 716-722 (1978)
Pieler, T., Kumagai, I., Erdmann, V.A.: Phylogenetic diversity reflected in the threedirnensional structure of ribosomal 5 s RNA. Zbl. Bakt. Hyg., I. Abt. Orig. C (1982) in press
Rigby, P. W.]., Dieckmann, M., Rhoades, c., Berg, P.: Labeling Deoxyribonucleic Acid toHigh Specific Activity in vitro by Nick Translation with DNA Polymerase I. J. molec.BioI. 113, 237-251 (1977)
Sancar, A., Hack, A.M., Rupp, W.D.: Simple Method for Identification of Plasmid-CodedProteins. J. Bact. 137, 692-693 (1979)
Southern, E.M.: Detection of Specific Sequences Among DNA Fragments Separated byGel Electrophoresis. J. molec. BioI. 98, 503-517 (1975)
Steitz, ].A.: Methanogenic bacteria. Nature 273,10 (1978)Stuber, D., Bujard, H.: Organization of transcriptional signals in plasmids pBR322 and
pACYC184. Proc. nat. Acad. Sci. (Wash.) 78, 167-171 (1981)Tornabene, T. G., Langworthy, T. A.: Diphytanyl and dibiphytanyl glycerol ether lipids in
methanogenic archaebacteria. Science 203, 51-53 (1978)Weiher, H.: Untersuchungen zur Struktur und Funktion von E. coli Promotoren: Variation
des lac Promoters durch gezielte Neukombination von Teilsequenzen und Mutagenese.Ph. D. Thesis, University of Heidelberg (1980)

Methanobrevibacter Polypeptide Synthesis in E. coli 109
Winter, J.U.: Glucose fermentation to methane and CO 2 by defined mixed cultures. Zbl.Bakt. Hyg., I.Abt. Orig. C 1, 293-305 (1980)
Winter, J., Wolfe, R. S.: Complete degradation of carbohydrate to carbon dioxide andmethane by syntrophic cultures of Acetobacterium woodii and Methanosarcina barkeri.Arch. Microbiol. 121, 97-102 (1979)
Winter, J.U., Wolfe, R.S.: Methane formation from fructose by syntrophic associations ofAcetobacterium woodii and different strains of methanogens. Arch. Microbiol. 124,73-79 (1980)
Zillig, W., Stetter, K.O., Schulz, W., ]anekovic, D.: Comparative studies of structure andfunction of DNA-dependent RNA polymerase from eubacteria and archaebacteria.I: Enzyme regulation and mechanism of action, Vol. 60 (P.Mildner, BiRies, eds.) Oxford-New York, Pergamon Press 1980
Professor Dr. A. Klein, Mikrobiologie, Universitat Heidelberg, Im Neuenheimer Fe1d230,D-6900 Heidelberg 1