document
TRANSCRIPT

news and views
192 nature structural biology • volume 8 number 3 • march 2001
The expression of eukaryotic genes in spe-cific tissues and at particular times ofdevelopment is determined by regulatorysequences called enhancers and silencers.Enhancers bind transcription factors,which then help initiate transcription ofthe RNA at the promoter. Enhancersaccomplish this independently of theirdistance and location with respect to thegene. This flexibility in enhancer functionalso gives these sequences the potential ofbeing highly promiscuous. Since thispromiscuity is rarely observed, eukaryotesmust have developed mechanisms toinsure that genes are not activated in thewrong tissue or at the wrong time ofdevelopment by enhancers from a neigh-boring gene. Chromatin insulators mayplay the role of gatekeepers that curtailenhancer promiscuity.
Boundaries or insulator elements havebeen experimentally identified based ontwo defining characteristics. Insulatorsinterfere with interactions betweenenhancers and promoters and inhibitenhancer-activated transcription, specifi-cally when interposed between the affect-ed enhancer and the promoter. Insulatorsalso buffer transgenes, inserted in thegenome at random sites, from chromoso-mal position effects1. Based on the firstproperty, insulators can be viewed as anadditional class of regulatory sequences inthe arsenal of controlling elements at theservice of genes to ensure their properspatial and temporal regulation. But thelatter property seems to suggest a morecomplex role, perhaps involving the orga-nization and compartmentalization of thechromatin fiber within the nucleus. Twopapers recently published in Scienceappear to support this interpretation2,3.
Properties of an insulatorInsulator elements have been described inmany organisms, including yeast,Drosophila and vertebrates1. In yeast, insu-lators appear to delimit the boundaries ofsilenced chromatin at telomeres and themating-type loci HML and HMR.Drosophila insulators were the first to becharacterized and this organism displays
the largest collection of these sequences sofar. The specialized chromatin structure(scs and scs′ elements) flank one of theheat shock protein 70 (hsp70) gene loci,where they seem to delimit the region thatbecomes transcriptionally active (orpuffed) in response to heat shock. OtherDrosophila insulators include those foundin the so-called gypsy retrovirus and theFab-7 and Fab-8 elements of the bithoraxcomplex, a complex containing three ofthe Hox genes that encode importantdevelopmental control proteins in flies.Insulators have also been studied in verte-brates, where they have been found in theribosomal RNA genes of Xenopus, thechicken β-globin genes and the human Tcell receptor (TCR)-α/δ locus among oth-ers. The widespread distribution of insu-lators is suggestive of an important rolefor these sequences in the biology of thenucleus.
An idea of the role insulators could playin the cell can be gleaned from the analysisof their distribution in the bithorax com-plex of Drosophila. The various genes ofthis complex are expressed in a paraseg-ment-specific pattern (a parasegment is ametameric unit along the anterior-posteri-or axis of flies that is out of phase with seg-ments) by a complex set of regulatorysequences arranged in a linear fashion over300 kb of DNA, corresponding to the orderof expression of the genes along the anteri-or-posterior axis of the animal. Theseparasegment-specific regulatory sequencesappear to be separated by insulator ele-ments, one of which is called Fab-7. TheFab-7 insulator is located between twoenhancers named iab-6 and iab-7 that con-trol expression of the Abdominal-B(Abd-B) gene in parasegments PS11 andPS12, respectively. Deletion of the Fab-7insulator eliminates the gatekeeper nor-mally ensuring that the iab-6 and iab-7enhancers activate transcription of theAbd-B gene in the appropriate tissues.Without the Fab-7 insulator, the iab-7enhancer activates expression of Abd-B inthe wrong cells, causing homeotic pheno-types in the adult fly. Homeotic pheno-types are those in which homologous
structures, such as legs and antennae, arechanged into one another. These resultssuggest that the Fab-7 insulator is requiredfor proper gene expression in order toavoid interference among the different reg-ulatory sequences of the Abd-B gene4–6. Asecond example that illustrates the role ofinsulators in vivo is that of imprinting inmice. Imprinting is an epigenetic traitoften seen as differences between paternaland maternal inheritance. The parent-of-origin-specific expression of the mouseH19 and Igf2 genes is controlled by aninsulator element located between the twogenes7,8.
Two possible models Much of the work on chromatin insulatorshas centered on the characterization ofassociated proteins with the goal ofexplaining how these sequences affectenhancer function, however, progress inthe field has been hampered by the lack ofunderstanding of how enhancers activatetranscription. Enhancers are currentlyviewed as entry points for transcriptionfactors which, once bound, can interactwith the transcription complex by loopingout intervening sequences9. More recently,enhancer-bound transcription factorshave been shown tethered to proteins atthe promoter region that are involved inhistone modification or other types ofchromatin remodeling complexes10. Basedon these findings, the effects of insulatorson enhancers could be explained byassuming that the insulator acts as a barri-er that inhibits the processivity of a signaltraveling from the enhancer to the pro-moter, without inactivating either. Theleitmotiv of this scenario, sometimesdubbed the ‘transcriptional model’, is thatthe insulator acts as a promoter decoy,confusing the enhancer-bound transcrip-tion factor into interacting with the insu-lator instead of the transcriptioncomplex11. This would result in the insula-tor trapping the enhancer, which thenwould not be able to interact with the pro-moter, thus precluding transcriptionalactivation. Another explanation, termedthe ‘structural model’ envisions insulators
Two insulators are not better than oneFabien Mongelard and Victor G. Corces
Chromatin insulators seem to control eukaryotic gene expression by regulating interactions between enhancersand promoters. Mounting evidence suggests that these sequences play a complex role in the regulation oftranscription, perhaps mediated by changes in chromatin structure and nuclear organization.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://s
tru
ctb
io.n
atu
re.c
om
© 2001 Nature Publishing Group http://structbio.nature.com
news and views
nature structural biology • volume 8 number 3 • march 2001 193
as sequences that organize the chromatinfiber within the nuclear space, creatingtranscriptionally independent domains12
by providing chromatin with anchoringpoints able to interact with some kind offixed substrate, perhaps the nuclearmatrix (see below). A way of explainingboth the effect of insulators onenhancer–promoter interactions and theirability to buffer transgenes from positioneffects is to assume that the barrier is aconsequence of the involvement of insula-tors in the establishment of domains ofhigher-order chromatin structure withinthe nucleus. The formation of thesedomains would then be the primary func-tion of insulators, and their effect onenhancer–promoter interactions would bea secondary consequence of the establish-ment of these domains. Recent articles byCai and Shen2 and by Muravyova et al.3
seem to support this view.
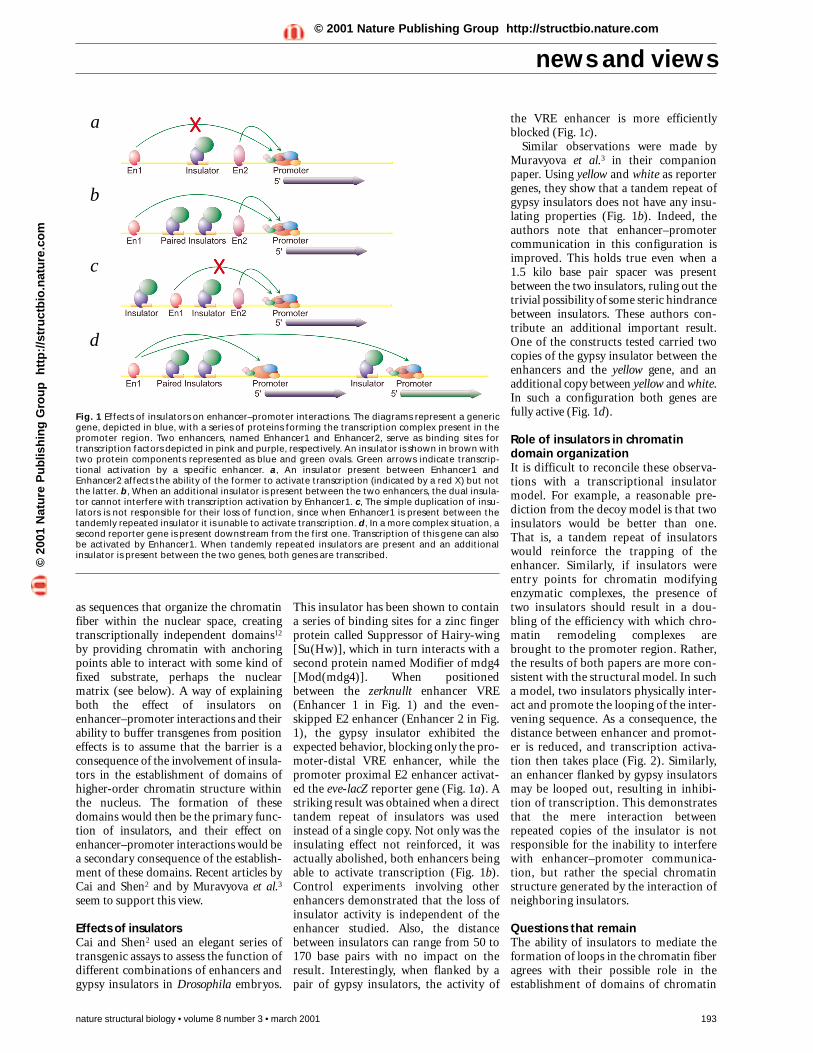
Effects of insulatorsCai and Shen2 used an elegant series oftransgenic assays to assess the function ofdifferent combinations of enhancers andgypsy insulators in Drosophila embryos.
This insulator has been shown to containa series of binding sites for a zinc fingerprotein called Suppressor of Hairy-wing[Su(Hw)], which in turn interacts with asecond protein named Modifier of mdg4[Mod(mdg4)]. When positionedbetween the zerknullt enhancer VRE(Enhancer 1 in Fig. 1) and the even-skipped E2 enhancer (Enhancer 2 in Fig.1), the gypsy insulator exhibited theexpected behavior, blocking only the pro-moter-distal VRE enhancer, while thepromoter proximal E2 enhancer activat-ed the eve-lacZ reporter gene (Fig. 1a). Astriking result was obtained when a directtandem repeat of insulators was usedinstead of a single copy. Not only was theinsulating effect not reinforced, it wasactually abolished, both enhancers beingable to activate transcription (Fig. 1b).Control experiments involving otherenhancers demonstrated that the loss ofinsulator activity is independent of theenhancer studied. Also, the distancebetween insulators can range from 50 to170 base pairs with no impact on theresult. Interestingly, when flanked by apair of gypsy insulators, the activity of
the VRE enhancer is more efficientlyblocked (Fig. 1c).
Similar observations were made byMuravyova et al.3 in their companionpaper. Using yellow and white as reportergenes, they show that a tandem repeat ofgypsy insulators does not have any insu-lating properties (Fig. 1b). Indeed, theauthors note that enhancer–promotercommunication in this configuration isimproved. This holds true even when a1.5 kilo base pair spacer was presentbetween the two insulators, ruling out thetrivial possibility of some steric hindrancebetween insulators. These authors con-tribute an additional important result.One of the constructs tested carried twocopies of the gypsy insulator between theenhancers and the yellow gene, and anadditional copy between yellow and white.In such a configuration both genes arefully active (Fig. 1d).
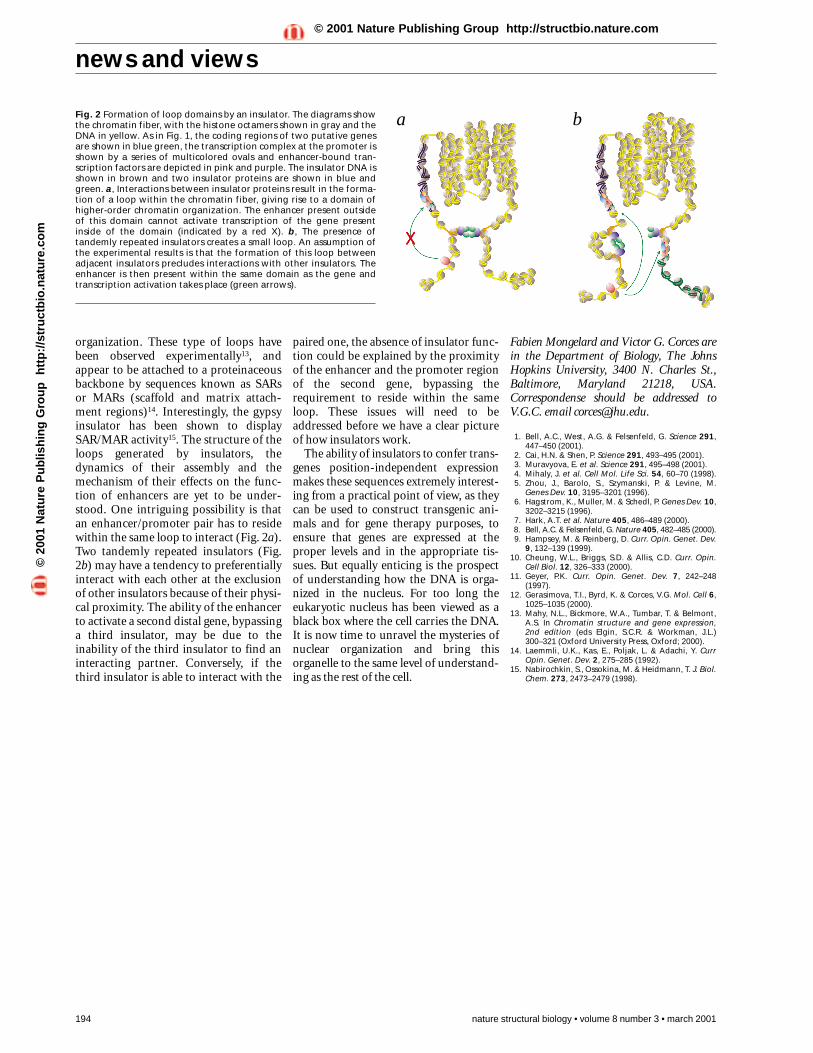
Role of insulators in chromatindomain organizationIt is difficult to reconcile these observa-tions with a transcriptional insulatormodel. For example, a reasonable pre-diction from the decoy model is that twoinsulators would be better than one.That is, a tandem repeat of insulatorswould reinforce the trapping of theenhancer. Similarly, if insulators wereentry points for chromatin modifyingenzymatic complexes, the presence oftwo insulators should result in a dou-bling of the efficiency with which chro-matin remodeling complexes arebrought to the promoter region. Rather,the results of both papers are more con-sistent with the structural model. In sucha model, two insulators physically inter-act and promote the looping of the inter-vening sequence. As a consequence, thedistance between enhancer and promot-er is reduced, and transcription activa-tion then takes place (Fig. 2). Similarly,an enhancer flanked by gypsy insulatorsmay be looped out, resulting in inhibi-tion of transcription. This demonstratesthat the mere interaction betweenrepeated copies of the insulator is notresponsible for the inability to interferewith enhancer–promoter communica-tion, but rather the special chromatinstructure generated by the interaction ofneighboring insulators.
Questions that remainThe ability of insulators to mediate theformation of loops in the chromatin fiberagrees with their possible role in theestablishment of domains of chromatin
Fig. 1 Effects of insulators on enhancer–promoter interactions. The diagrams represent a genericgene, depicted in blue, with a series of proteins forming the transcription complex present in thepromoter region. Two enhancers, named Enhancer1 and Enhancer2, serve as binding sites fortranscription factors depicted in pink and purple, respectively. An insulator is shown in brown withtwo protein components represented as blue and green ovals. Green arrows indicate transcrip-tional activation by a specific enhancer. a, An insulator present between Enhancer1 andEnhancer2 affects the ability of the former to activate transcription (indicated by a red X) but notthe latter. b, When an additional insulator is present between the two enhancers, the dual insula-tor cannot interfere with transcription activation by Enhancer1. c, The simple duplication of insu-lators is not responsible for their loss of function, since when Enhancer1 is present between thetandemly repeated insulator it is unable to activate transcription. d, In a more complex situation, asecond reporter gene is present downstream from the first one. Transcription of this gene can alsobe activated by Enhancer1. When tandemly repeated insulators are present and an additionalinsulator is present between the two genes, both genes are transcribed.
a
b
c
d
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://s
tru
ctb
io.n
atu
re.c
om
© 2001 Nature Publishing Group http://structbio.nature.com
news and views
194 nature structural biology • volume 8 number 3 • march 2001
organization. These type of loops havebeen observed experimentally13, andappear to be attached to a proteinaceousbackbone by sequences known as SARsor MARs (scaffold and matrix attach-ment regions)14. Interestingly, the gypsyinsulator has been shown to displaySAR/MAR activity15. The structure of theloops generated by insulators, thedynamics of their assembly and themechanism of their effects on the func-tion of enhancers are yet to be under-stood. One intriguing possibility is thatan enhancer/promoter pair has to residewithin the same loop to interact (Fig. 2a).Two tandemly repeated insulators (Fig.2b) may have a tendency to preferentiallyinteract with each other at the exclusionof other insulators because of their physi-cal proximity. The ability of the enhancerto activate a second distal gene, bypassinga third insulator, may be due to theinability of the third insulator to find aninteracting partner. Conversely, if thethird insulator is able to interact with the
paired one, the absence of insulator func-tion could be explained by the proximityof the enhancer and the promoter regionof the second gene, bypassing therequirement to reside within the sameloop. These issues will need to beaddressed before we have a clear pictureof how insulators work.
The ability of insulators to confer trans-genes position-independent expressionmakes these sequences extremely interest-ing from a practical point of view, as theycan be used to construct transgenic ani-mals and for gene therapy purposes, toensure that genes are expressed at theproper levels and in the appropriate tis-sues. But equally enticing is the prospectof understanding how the DNA is orga-nized in the nucleus. For too long theeukaryotic nucleus has been viewed as ablack box where the cell carries the DNA.It is now time to unravel the mysteries ofnuclear organization and bring thisorganelle to the same level of understand-ing as the rest of the cell.
Fabien Mongelard and Victor G. Corces arein the Department of Biology, The JohnsHopkins University, 3400 N. Charles St.,Baltimore, Maryland 21218, USA.Correspondense should be addressed toV.G.C. email [email protected].
1. Bell, A.C., West, A.G. & Felsenfeld, G. Science 291,447–450 (2001).
2. Cai, H.N. & Shen, P. Science 291, 493–495 (2001).3. Muravyova, E. et al. Science 291, 495–498 (2001).4. Mihaly, J. et al. Cell Mol. Life Sci. 54, 60–70 (1998).5. Zhou, J., Barolo, S., Szymanski, P. & Levine, M.
Genes Dev. 10, 3195–3201 (1996).6. Hagstrom, K., Muller, M. & Schedl, P. Genes Dev. 10,
3202–3215 (1996).7. Hark, A.T. et al. Nature 405, 486–489 (2000).8. Bell, A.C. & Felsenfeld, G. Nature 405, 482–485 (2000).9. Hampsey, M. & Reinberg, D. Curr. Opin. Genet. Dev.
9, 132–139 (1999).10. Cheung, W.L., Briggs, S.D. & Allis, C.D. Curr. Opin.
Cell Biol. 12, 326–333 (2000).11. Geyer, P.K. Curr. Opin. Genet. Dev. 7, 242–248
(1997).12. Gerasimova, T.I., Byrd, K. & Corces, V.G. Mol. Cell 6,
1025–1035 (2000).13. Mahy, N.L., Bickmore, W.A., Tumbar, T. & Belmont,
A.S. In Chromatin structure and gene expression,2nd edition (eds Elgin, S.C.R. & Workman, J.L.)300–321 (Oxford University Press, Oxford; 2000).
14. Laemmli, U.K., Kas, E., Poljak, L. & Adachi, Y. CurrOpin. Genet. Dev. 2, 275–285 (1992).
15. Nabirochkin, S., Ossokina, M. & Heidmann, T. J. Biol.Chem. 273, 2473–2479 (1998).
a bFig. 2 Formation of loop domains by an insulator. The diagrams showthe chromatin fiber, with the histone octamers shown in gray and theDNA in yellow. As in Fig. 1, the coding regions of two putative genesare shown in blue green, the transcription complex at the promoter isshown by a series of multicolored ovals and enhancer-bound tran-scription factors are depicted in pink and purple. The insulator DNA isshown in brown and two insulator proteins are shown in blue andgreen. a, Interactions between insulator proteins result in the forma-tion of a loop within the chromatin fiber, giving rise to a domain ofhigher-order chromatin organization. The enhancer present outsideof this domain cannot activate transcription of the gene presentinside of the domain (indicated by a red X). b, The presence oftandemly repeated insulators creates a small loop. An assumption ofthe experimental results is that the formation of this loop betweenadjacent insulators precludes interactions with other insulators. Theenhancer is then present within the same domain as the gene andtranscription activation takes place (green arrows).
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://s
tru
ctb
io.n
atu
re.c
om
© 2001 Nature Publishing Group http://structbio.nature.com