document
TRANSCRIPT

© 2001 Macmillan Magazines Ltd
articles
NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com 221
Ran stimulates spindle assembly byaltering microtubule dynamics andthe balance of motor activities
Andrew Wilde*†‡, Sofia B. Lizarraga*‡§, Lijun Zhang*, Christiane Wiese*, Neal R. Gliksman¶, Claire E. Walczak# and Yixian Zheng*§**
*Howard Hughes Medical Institute, Department of Embryology, Carnegie Institution of Washington, Baltimore, Maryland 21210, USA§Graduate program in Biology, Johns Hopkins University, Baltimore, Maryland 21210, USA
¶Universal Imaging Corporation, West Chester, Pennsylvania 19380, USA#Medical Sciences Program, Indiana University, Bloomington, Indiana 47405, USA
†Current address: Department of Medical Genetics and Microbiology, University of Toronto, Kings College Circle, Toronto, M5S 1A8, Canada‡These authors contributed equally to this work.
**e-mail: [email protected]
The guanosine tri-phosphatase Ran stimulates assembly of microtubule spindles. However, it is not known whataspects of the microtubule cytoskeleton are subject to regulation by Ran in mitosis. Here we show that Ran–GTPstimulates microtubule assembly by increasing the rescue frequency of microtubules three- to eightfold. In additionto changing microtubule dynamics, Ran–GTP also alters the balance of motor activities, partly as a result of anincrease in the amount of motile Eg5, a plus-end-directed microtubule motor that is essential for spindle formation.Thus, Ran regulates multiple processes that are involved in spindle assembly.
Amicrotubule spindle is required for equal segregation of chro-mosomes in mitosis and meiosis. Spindle assembly requiresthe coordinated action of chromosomes, centrosomes,
microtubule-based motor proteins, and changes in microtubuledynamics1,2. Microtubules are dynamic polymers that alternateabruptly and stochastically between phases of polymerization anddepolymerization. This behaviour, known as ‘dynamic instability’3,is characterized by four parameters — polymerization rate, depoly-merization rate, frequency of transition from growth to shrinkage(catastrophe frequency), and frequency of transition from shrink-age to growth (rescue frequency). Upon the transition from inter-phase to mitosis, microtubules become more dynamic as the resultof increased catastrophe frequency4,5 and reduced rescue frequen-cy6,7. These changes in microtubule dynamics correspond to anincrease in Cdc2 kinase activity4,6, and result in the replacement oflong, relatively stable interphase microtubules with short, highly
dynamic mitotic microtubules8. However, spindle assemblyrequires the selective stabilization of these highly dynamic micro-tubules in the vicinity of mitotic chromosomes9–11.
Mitotic chromosomes have been shown to be important in micro-tubule polymerization and spindle assembly in several systems12–15.For example, in Xenopus eggs, chromosomes or nuclei can inducelarge microtubule asters from nearby centrosomes12. Moreover, in theabsence of centrosomes, artificial chromatin tethered to beads hasbeen shown to stimulate microtubule assembly and spindle forma-tion in Xenopus egg extracts13. By detailed dynamic analysis, chro-matin has been found to have a stabilizing effect on microtubules16.This provides a possible explanation for the observed effects of chro-matin on microtubule polymerization and spindle assembly.
In addition to changes in microtubule dynamics, microtubule-based motors have been shown to be essential for establishmentand maintenance of spindles2,17. Motor proteins generate forces to
Table 1 Summary of the effects of Ran(L43E) on microtubule dynamics
Extract Vg Vs fcat fres fpg fps n(µm min-1) (µm min-1) (events per s) (events per s) (events per s) (events per s)
1Ran(L43E) 8.3 ± 2.9 5.8 ± 2.2 0.023 0.017 0.01 0.004 49Control 6.9 ± 2.8 6.9 ± 3.1 0.030 0.004 0.01 0.004 52
2Ran(L43E) 7.4 ± 2.7 7.3 ± 3.2 0.026 0.016 0.004 0.006 54Control 7.9 ± 2.7 7.5 ± 2.6 0.044 0.002 0.006 0.003 94
3Ran(L43E) 10.6 ± 4.6 8.4 ± 3.8 0.023 0.022 0.009 0.005 73Control 10.1 ± 4.7 11.3 ± 4.8 0.024 0.006 0.009 0.004 83
Avg.Ran(L43E) 8.8 ± 3.4 7.2 ± 3.1 0.024 0.018 0.008 0.005 –Control 8.3 ± 3.4 8.6 ± 3.4 0.033 0.004 0.008 0.004 –
Analysis from three independent experiments using different fresh extracts. Growth (Vg) and shrinkage (Vs) are shown as mean rates ± s.d. Frequencies of catastrophe (fcat), rescue (fres), pausingwhile growing (fpg) and pausing while shrinking (fps) are expressed as events per second. n represents number of dynamic microtubules analyzed. Similar results were obtained in three independ-ent experiments with frozen extracts (data not shown).
© 2001 Macmillan Magazines Ltd
articles
NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com222
move cargo uni-directionally, towards either the plus or the minusend of the microtubule, depending on the motor family18. In mito-sis, motors are important for organizing the minus ends of micro-tubules at the spindle pole and the plus ends of microtubules at thespindle mid-zone2. Several studies have shown that removal ofeither a plus- or a minus-end-directed motor disrupts spindle for-mation17,19–21. However, removal of a certain combination of bothplus- and minus-end-directed motor activity allows spindle forma-tion17,19,20. It was therefore proposed that plus- and minus-end-directed motors work in opposition to one another within a spin-dle. Consequently, balancing these motor activities is crucial forspindle formation and maintenance.
The central question in spindle assembly is how microtubulepolymerization and motor activities are regulated to ensure properalignment of chromosomes on the spindle. Recently, Ran–GTP andits chromatin-bound nucleotide-exchange factor, RCC1, werefound to stimulate microtubule polymerization and spindle assem-bly in Xenopus egg extracts22–26. This gave rise to the hypothesis that,in vivo, chromatin-bound RCC1 generates high local concentra-tions of Ran–GTP, which then stimulates microtubule polymeriza-tion and spindle assembly around the chromosomes. Here we
report that Ran–GTP is capable of carrying out chromatin-inducedstabilization of microtubules. We also show that Ran–GTP is ableto regulate the balance of microtubule-motor activities. These find-ings indicate that Ran–GTP may be a central regulator of spindleassembly and maintenance.
ResultsRanGTP increases the frequency of microtubule rescue. To deter-mine whether Ran–GTP alters microtubule dynamics, we analysed
20
15
10
5
0
Leng
th (
µm)
Leng
th (
µm)
Time (s)Time (s)
c dControl Ran(L43E)
0 50 100 150 200 250
20
15
10
5
0 50 100 150 200 2500
a Control
b Ran(L43E)
0:00 0:18 1:08
0:00 0:18 0:38
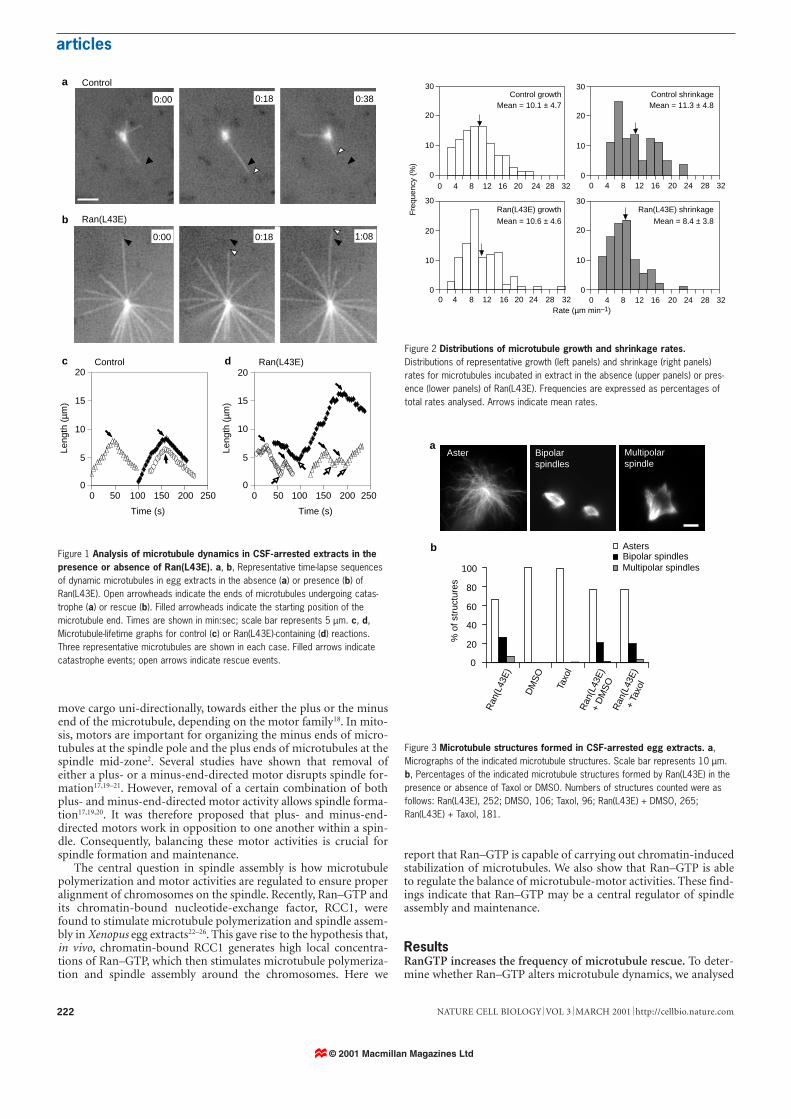
Figure 1 Analysis of microtubule dynamics in CSF-arrested extracts in thepresence or absence of Ran(L43E). a, b, Representative time-lapse sequencesof dynamic microtubules in egg extracts in the absence (a) or presence (b) ofRan(L43E). Open arrowheads indicate the ends of microtubules undergoing catas-trophe (a) or rescue (b). Filled arrowheads indicate the starting position of themicrotubule end. Times are shown in min:sec; scale bar represents 5 µm. c, d,Microtubule-lifetime graphs for control (c) or Ran(L43E)-containing (d) reactions.Three representative microtubules are shown in each case. Filled arrows indicatecatastrophe events; open arrows indicate rescue events.
Freq
uenc
y (%
)
Control growth
Rate (µm min–1)
0 4 8 12 16 20 24 28 32
Mean = 10.1 ± 4.7
Ran(L43E) growth
Mean = 10.6 ± 4.6
Control shrinkageMean = 11.3 ± 4.8
Ran(L43E) shrinkage
Mean = 8.4 ± 3.8
0 4 8 12 16 20 24 28 32
30
20
10
0
0
10
20
30
0 4 8 12 16 20 24 28 32
0 4 8 12 16 20 24 28 32
30
20
10
0
30
20
10
0
Figure 2 Distributions of microtubule growth and shrinkage rates.Distributions of representative growth (left panels) and shrinkage (right panels)rates for microtubules incubated in extract in the absence (upper panels) or pres-ence (lower panels) of Ran(L43E). Frequencies are expressed as percentages oftotal rates analysed. Arrows indicate mean rates.
Aster Bipolarspindles
Multipolarspindle
% o
f str
uctu
res
100
0
20
40
60
80
Ran
(L43
E)
DM
SO
Taxo
l
Ran
(L43
E)
+ D
MS
OR
an(L
43E
)+
Taxo
l
AstersBipolar spindlesMultipolar spindles
a
b
Figure 3 Microtubule structures formed in CSF-arrested egg extracts. a,Micrographs of the indicated microtubule structures. Scale bar represents 10 µm.b, Percentages of the indicated microtubule structures formed by Ran(L43E) in thepresence or absence of Taxol or DMSO. Numbers of structures counted were asfollows: Ran(L43E), 252; DMSO, 106; Taxol, 96; Ran(L43E) + DMSO, 265;Ran(L43E) + Taxol, 181.
© 2001 Macmillan Magazines Ltd
articles
223NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com
the behaviour of individual microtubules nucleated from exoge-neously added centrosomes in cytostatic factor (CSF)-arrested eggextracts using time-lapse fluorescence microscopy. Ran(L43E) (aconstitutively active Ran allele bound to GTP), stimulated the for-mation of dense microtubule asters within 10–15 min of incuba-tion in the egg extract23. Therefore, to visualize individual micro-tubules, we obtained time-lapse images within the first 10 min ofthe experiment. To examine the behaviour of individual micro-tubules, we measured microtubule lengths as a function of timefrom sequential images of centrosomes incubated in the extract inthe presence or absence of Ran(L43E) (Fig. 1a, b). We analysedmicrotubules in fresh extracts in three independent experiments,each involving measurement of at least 45 microtubules (Table 1),and plotted the length of each microtubule against time.Microtubule lifetime plots (Fig. 1) show that the most strikingeffect of Ran(L43E) on microtubules is an increase in the frequen-cy of microtubule rescue.
To assess the dynamic parameters, we chose criteria thatrequired microtubule growth or shrinkage to persist for at leastfour consecutive frames (6 s), and the net change in length to be atleast 0.5 µm. Otherwise, microtubules were considered to be ‘paus-ing’. The distribution of growth rates for one representative exper-iment (Fig. 2), and the mean rates for each of the three independ-ent experiments (Table 1), show that the microtubule growth ratewas unaffected by Ran(L43E). The mean rate of microtubuleshrinkage was slightly reduced in two out of three experiments(Table 1), which is also reflected in a change of the distribution ofshrinkage rates (Fig. 2). This analysis shows that Ran(L43E) doesnot markedly alter the rates of microtubule growth or shrinkage inCSF-arrested extracts.
Some microtubules exhibited short pausing periods whilegrowing or shrinking in the presence or absence of Ran(L43E)(data not shown). Because changes in the time microtubules spentpausing could also affect microtubule stability, we determined thefrequencies with which microtubules paused either during growth(fpg) or during shrinkage (fps). Each pausing event was defined by alack of change in microtubule length (<0.5 µm) over at least fiveconsecutive frames. We calculated fpg and fps by dividing the num-ber of pausing events during growth or shrinkage by the timespent in each phase, respectively. We found that Ran(L43E) did notsignificantly affect the pausing frequency (Table 1).
To determine catastrophe and rescue frequencies, we divided thenumber of catastrophe or rescue events by the total time thedynamic microtubules spent growing or shrinking, respectively.Ran(L43E) caused a small reduction in the catastrophe frequencyin two out of three experiments (Table 1). However, the most strik-ing and consistent effect of Ran(L43E) on microtubule dynamicswas a three- to eightfold increase in rescue frequency (Table 1).Similar results were obtained using frozen extracts (three inde-pendent experiments, data not shown).
To investigate whether the effect of Ran–GTP on microtubuledynamics is direct, we used a purified system with centrosomes andpure tubulin in the presence of Ran(L43E) or Ran(T24N) (a Ranallele bound to GDP). We found no difference in total mass ofmicrotubule polymer, mean microtubule length or length distribu-tion in three different experiments in the presence or absence ofRan–GTP (data not shown). This indicates that Ran does notdirectly affect microtubule dynamics. On the basis of these studies,we propose that Ran–GTP may mediate the stabilizing effect of thechromosome on microtubules, primarily by increasing the fre-quency of microtubule rescue.Taxol and dimethylsulphoxide (DMSO) do not inhibit spindleassembly. Microtubule stabilization alone may not be sufficientfor Ran-induced spindle assembly. In CSF-arrested egg extracts,Ran(L43E) stimulates formation of microtubule asters and bipolarspindles, whereas the microtubule-stabilizing reagents Taxol andDMSO induce formation of microtubule asters only23,24. To deter-mine whether Taxol and DMSO inhibit spindle assembly, we initi-
130.4 s87.7 s56.3 s
4.7 s 36.1 s 45.1 s
% o
f mov
ing
seed
s%
of t
otal
mic
rotu
bule
stru
ctur
es s
core
d
0
20
40
60
80
100
Asters Bipolarspindles
Multipolarspindles
Control antibody
Anti-Eg5 antibody
Monastrol
0
20
40
60
80
100
% o
f mov
ing
seed
s
0
20
40
60
80
100
a
b
c
d
Ran
(L43
E)
DM
SO
Taxo
l
Ran
(L43
E)
+ D
MS
O
Ran
(T24
N)
+ Ta
xol
Ran
(L43
E)
+ Ta
xol
Taxo
l
Ant
i-Eg5
+Ta
xol
Ran
(L43
E)
+ an
ti-E
g5+
Taxo
l
Ran
(L43
E)
+ an
ti-X
CTK
2+
Taxo
l
Ran
(L43
E)
+ Ta
xol
Ran
(L43
E)
+ M
A +
Tax
ol
Minus-end-directedPlus-end-directed
Minus-end-directedPlus-end-directed
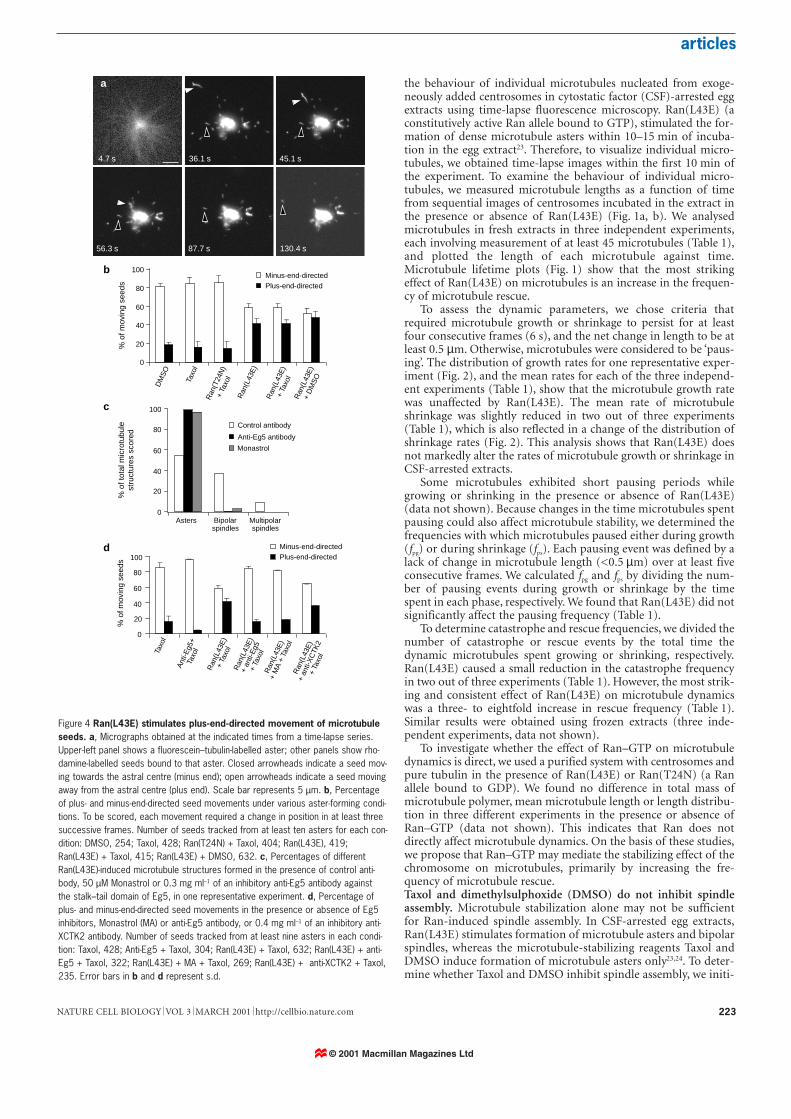
Figure 4 Ran(L43E) stimulates plus-end-directed movement of microtubuleseeds. a, Micrographs obtained at the indicated times from a time-lapse series.Upper-left panel shows a fluorescein–tubulin-labelled aster; other panels show rho-damine-labelled seeds bound to that aster. Closed arrowheads indicate a seed mov-ing towards the astral centre (minus end); open arrowheads indicate a seed movingaway from the astral centre (plus end). Scale bar represents 5 µm. b, Percentageof plus- and minus-end-directed seed movements under various aster-forming condi-tions. To be scored, each movement required a change in position in at least threesuccessive frames. Number of seeds tracked from at least ten asters for each con-dition: DMSO, 254; Taxol, 428; Ran(T24N) + Taxol, 404; Ran(L43E), 419;Ran(L43E) + Taxol, 415; Ran(L43E) + DMSO, 632. c, Percentages of differentRan(L43E)-induced microtubule structures formed in the presence of control anti-body, 50 µM Monastrol or 0.3 mg ml–1 of an inhibitory anti-Eg5 antibody againstthe stalk–tail domain of Eg5, in one representative experiment. d, Percentage ofplus- and minus-end-directed seed movements in the presence or absence of Eg5inhibitors, Monastrol (MA) or anti-Eg5 antibody, or 0.4 mg ml–1 of an inhibitory anti-XCTK2 antibody. Number of seeds tracked from at least nine asters in each condi-tion: Taxol, 428; Anti-Eg5 + Taxol, 304; Ran(L43E) + Taxol, 632; Ran(L43E) + anti-Eg5 + Taxol, 322; Ran(L43E) + MA + Taxol, 269; Ran(L43E) + anti-XCTK2 + Taxol,235. Error bars in b and d represent s.d.

© 2001 Macmillan Magazines Ltd
articles
NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com224
ated Ran-dependent spindle assembly in the presence of Taxol orDMSO. In the presence or absence of Taxol or DMSO, Ran(L43E)induced formation of similar numbers of bipolar spindles relativeto the total number of microtubule structures formed, indicatingthat neither Taxol nor DMSO inhibited spindle assembly (Fig. 3).These results inicate that microtubule production in CSF-arrestedextract may be insufficient for spindle assembly. Therefore, inaddition to stabilizing microtubules, Ran–GTP seems to activateother processes that are required for spindle formation.RanGTP alters the balance of motor activities on microtubuleasters. Motor proteins have been found to have important func-tions in spindle assembly and maintenance2,17,21. Therefore, todetermine whether Ran–GTP affects the activity of motor proteins,we monitored the movement of stabilized, rhodamine-labelledmicrotubule seeds on fluorescein-labelled microtubule asters, bytime-lapse fluorescence microscopy (Fig. 4a). We induced forma-tion of microtubule asters with Taxol or DMSO in the presence ofeither Ran(L43E) or Ran(T24N) in CSF-arrested egg extracts.Ran(L43E) did not affect the velocity (26.6 ± 18 µm min–1 towardsthe astral centre; 6.34 ± 3.34 µm min–1 away from the astral centre)or duration of seed movement. However, analysis of the relativeproportion of seeds translocating to either the plus or minus end ofastral microtubules revealed significant differences. Under non-spindle-forming conditions (in the absence of Ran(L43E)), 80% ofmotile seeds translocated towards the minus end (astral centre),whereas 20% translocated towards the plus end (astral periphery;Fig. 4b). This indicates that under these conditions, minus-end-directed motor activity is predominant. In contrast, under spindle-forming conditions (in the presence of Ran(L43E)), ~40% ofmotile seeds translocated towards the plus end, a twofold increaseover non-spindle-forming conditions (Fig. 4b). This increase in therelative amount of plus-end-directed seed movement indicates thatby changing the balance of motor activities, Ran–GTP reduces thedominance of minus-end-directed motors on the microtubuleaster. This could provide the driving force for spindle formation.The change in the balance of motor activity requires Eg5. Eg5, atetrameric kinesin consisting of two pairs of motor domains separat-ed by stalk domains, is a plus-end-directed motor that is essential forspindle assembly27,28. To determine whether Eg5 is involved in theRan–GTP-dependent increase in plus-end-directed motor activity,we inhibited Eg5 activity either with an inhibitory antibody againstthe Eg5 stalk–tail domain27 or with Monastrol, a chemical inhibitorof Eg5 (ref. 29). Each of these reagents blocked Ran(L43E)-inducedspindle formation without affecting aster assembly in CSF-arrestedegg extract (Fig. 4c). Inhibiting Eg5 also blocked the increase in plus-
end-directed seed movement induced by Ran(L43E) (Fig. 4d). Incontrast, inhibiting XCTK2 (ref. 30), a minus-end-directed motor,had no effect (Fig. 4d). We were unable to assess the contribution ofdynein, another minus-end-directed motor, because inhibitingdynein severely disrupts aster formation23,24.Plus-end-moving microtubules are antiparallel to astral micro-tubules. Eg5 is thought to move microtubules in an antiparallel man-ner; seeds moved by Eg5 towards the plus ends of astral microtubulesshould therefore move with their minus ends leading. We determineethe orientation of motile microtubule seeds using polarity-markedmicrotubule seeds that have a bright minus end and a dim plus end.In the seed-translocation assay in the presence of Ran(L43E), 94% ofseeds (n = 31) moving towards the plus ends of microtubules ledwith their bright minus ends, indicating that they were antiparallel tothe astral microtubules upon which they moved (Fig. 5a). Consistentwith a previous report31, all seeds moving towards the minus ends ofasters led with their minus ends (Fig. 5b). This analysis indicates thatEg5 may drive plus-end-directed seed translocation by slidingantiparallel microtubules against one another.Ran–GTP promotes movement of Eg5 to the plus ends of micro-tubule asters. We found that Ran–GTP caused no apparent changein the phosphorylation level of Eg5, or in the amount of Eg5 boundto microtubules in CSF-arrested egg extracts (data not shown).Moreover, using an in vitro assay for Eg5-mediated microtubuletranslocation27, we found that Ran–GTP did not alter the velocity,duration or distance of Eg5-dependant microtubule gliding (datanot shown). These data indicate that Ran–GTP does not directlyaffect Eg5, but may instead act through intermediary effectors. Todetermine whether Ran–GTP affects the behaviour of Eg5 onmicrotubule asters, we used a labelled antibody against the Eg5stalk domain to monitor Eg5 by time-lapse fluorescencemicroscopy. At 25 µg ml–1, this antibody did not inhibitRan(L43E)-induced spindle formation (data not shown). Thelabelled anti-Eg5 antibody appeared as bright speckles concentrat-ed at the centre of microtubule asters in the presence or absence ofRan(L43E) (Fig. 6a, a′, b, b′). A fraction of the speckles moved awayfrom the astral centre, towards the microtubule plus end, withvelocities of 2.75 ± 0.9 µm min–1 (n = 107) for Taxol plusRan(L43E) and 2.71 ± 0.8 µm min–1 (n = 111) for Taxol plusRan–GDP (inactive, wild-type Ran bound to GDP23). The motilityof speckles was dependent on Eg5, as Monastrol markedly reducedthe amount of speckle movement (data not shown).
The velocity of seeds moving towards the plus end(6.34 ± 3.34 µm min–1) was roughly twice that of the speckles(2.75 ± 0.9 µm min–1). One possible explanation for this is that Eg5
197.0 s 201.0 s 204.0 s 206.0 s
115.5 s 122.7 s 127.5 s 232.3 sa
b
Figure 5 Microtubule seeds lead with their minus ends when moving towardsthe plus or minus end of the aster. Micrographs obtained at the indicated timesfrom a time-lapse series depicting stabilized, polarity-marked microtubule seeds on
Ran(L43E)-induced microtubule asters. a, Seed moving towards the plus end. b,Seed moving towards the minus end. Open arrows show the initial position of theseed; filled arrows show subsequent positions. Scale bars represent 5 µm.
© 2001 Macmillan Magazines Ltd
articles
225NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com
simultaneously uses the motor domains at both ends of the Eg5molecule. The motor domains at one end of the molecule movealong the astral microtubules, whereas motor domains at the oppo-site end move the seed. As the seed is being moved by an alreadymotile Eg5, the apparent velocity of the seed would be twice that ofEg5 itself. This process would require enough Eg5 to be localizedalong the length of the microtubules to allow microtubule seeds tobe passed from one motor to another. Immunostaining revealedthat Eg5 localized to the astral centre, as well as along the length ofthe microtubule (data not shown), which is consistent with previ-ous studies32. Therefore, there may be enough Eg5 along the lengthof the microtubule to allow seeds to be passed from one motor to
another irrespective of their size, thereby creating the appearance ofseeds that are moving with twice the speed of Eg5.
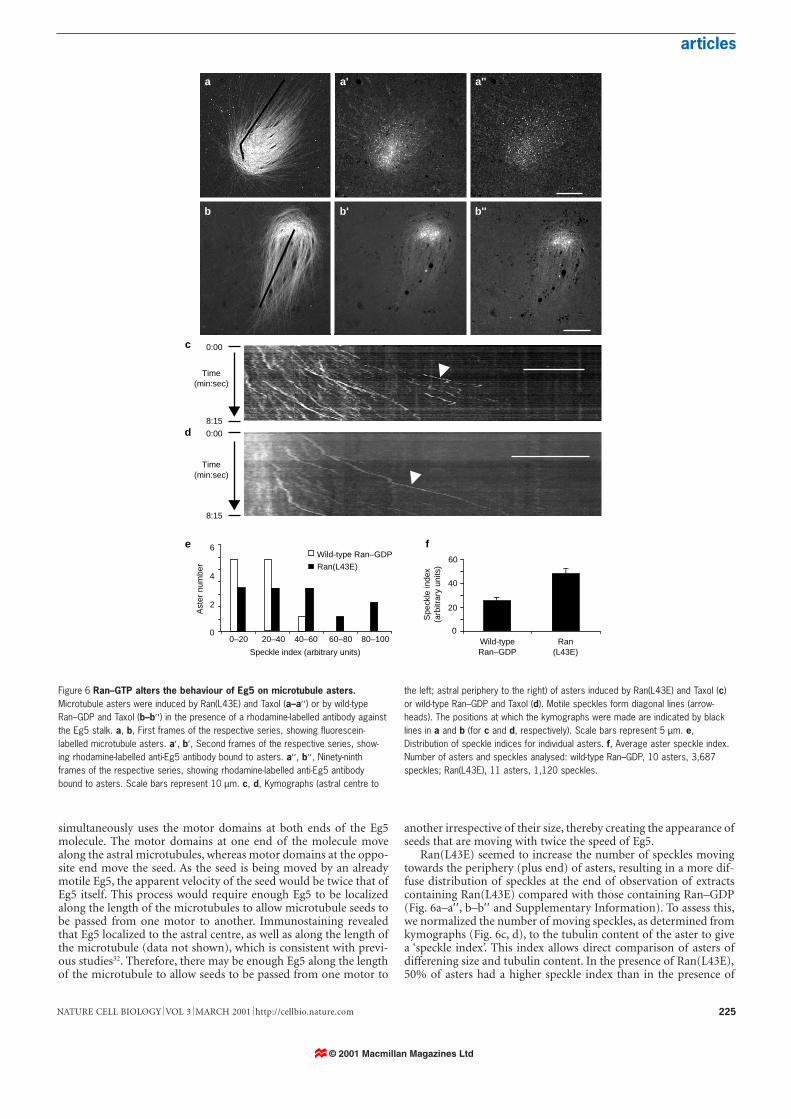
Ran(L43E) seemed to increase the number of speckles movingtowards the periphery (plus end) of asters, resulting in a more dif-fuse distribution of speckles at the end of observation of extractscontaining Ran(L43E) compared with those containing Ran–GDP(Fig. 6a–a′′ , b–b′′ and Supplementary Information). To assess this,we normalized the number of moving speckles, as determined fromkymographs (Fig. 6c, d), to the tubulin content of the aster to givea ‘speckle index’. This index allows direct comparison of asters ofdifferening size and tubulin content. In the presence of Ran(L43E),50% of asters had a higher speckle index than in the presence of
0
2
4
6
0–20 20–40 40–60 60–80 80–100
Wild-type Ran–GDP
Ran(L43E)
0
20
40
60
Wild-typeRan–GDP
Ran(L43E)
0:00
8:15
Time(min:sec)
Time(min:sec)
8:15
0:00
Speckle index (arbitrary units)
Spe
ckle
inde
x(a
rbitr
ary
units
)
Ast
er n
umbe
r
a
b
a'
b'
a''
b''
c
d
e f
Figure 6 Ran–GTP alters the behaviour of Eg5 on microtubule asters.Microtubule asters were induced by Ran(L43E) and Taxol (a–a′′ ) or by wild-typeRan–GDP and Taxol (b–b′′ ) in the presence of a rhodamine-labelled antibody againstthe Eg5 stalk. a, b, First frames of the respective series, showing fluorescein-labelled microtubule asters. a′, b′, Second frames of the respective series, show-ing rhodamine-labelled anti-Eg5 antibody bound to asters. a′′ , b′′ , Ninety-ninthframes of the respective series, showing rhodamine-labelled anti-Eg5 antibodybound to asters. Scale bars represent 10 µm. c, d, Kymographs (astral centre to
the left; astral periphery to the right) of asters induced by Ran(L43E) and Taxol (c)or wild-type Ran–GDP and Taxol (d). Motile speckles form diagonal lines (arrow-heads). The positions at which the kymographs were made are indicated by blacklines in a and b (for c and d, respectively). Scale bars represent 5 µm. e,Distribution of speckle indices for individual asters. f, Average aster speckle index.Number of asters and speckles analysed: wild-type Ran–GDP, 10 asters, 3,687speckles; Ran(L43E), 11 asters, 1,120 speckles.

© 2001 Macmillan Magazines Ltd
articles
NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com226
Ran–GDP. This is reflected in the fact that the average aster speck-le index was twofold greater (P < 0.025) in the presence ofRan(L43E) than Ran–GDP. These findings demonstrate thatRan–GTP increases the amount of Eg5 moving towards the plusends of microtubule asters and that this change could lead toincreased plus-end-directed microtubule translocation and spindleassembly.
DiscussionRecent studies have indicated that the observed stabilizing effect ofchromosomes on mitotic microtubules may be mediated by a highlocal concentration of Ran–GTP generated by chromatin-associat-ed RCC1. However, the mechanism of the activity of Ran in mito-sis is completely unknown. Here we have shown that one of theprincipal effects of Ran–GTP on microtubules is a three- to eight-fold increase in rescue frequency. Thus, Ran–GTP creates a distinctstate of dynamic behaviour of microtubules that differs from thedynamics of interphase microtubules, as well as those of mitoticmicrotubules (in the absence of chromatin). In this dynamic state,microtubules undergo frequent catastrophes and rescues, resultingin the formation of a large number of microtubules that can beused for spindle assembly.
In animal cells, spindles are assembled by one of two differentmorphogenetic processes, represented by mitotic spindle forma-tion and, in certain cases (such as Drosophila), female meiotic spin-dle formation. Formation of the mitotic spindle begins with radialarrays of microtubules that are nucleated by two duplicated cen-trosomes. As the two centrosomes move away from each other andmicrotubules capture chromosomes, a bipolar spindle evolves witha centrosome at each pole. In contrast, the morphogenesis of, forexample, the Drosophila female meiotic spindle33, which is assem-bled in the absence of centrosomes, begins with a random arrayof microtubules that are polymerized near to condensed chro-mosomes. Several studies have given rise to the hypothesis thatchromosomes induce spindle formation12,13. This indicates thatchromosomes in both mitotic and meiotic spindles may imposea localized microtubule-stabilizing environment upon a generaldestabilizing environment. Therefore, the two seemingly differ-ent spindle assembly processes may share some common princi-ples. We propose that the high local concentration of Ran–GTPgenerated by chromatin-bound RCC1 could drive the growth ofmicrotubules towards chromosomes, primarily by increasing therescue frequency of microtubules. Thus, chromosomes would pro-vide a positional cue to guide the reorganization of astral micro-tubule arrays, in the case of mitotic-spindle assembly, or randomarrays, in the case of meiotic-spindle assembly, into a bipolar array.
In addition to regulating microtubule dynamics, we found thatRan–GTP alters the balance of motor activities on microtubuleasters, partly by affecting the behaviour of Eg5, such that in thepresence of Ran–GTP more Eg5 translocates to the plus ends ofasters. One possible explanation is that Ran–GTP, through itsimmediate downstream effectors, causes Eg5 tethered at the astralcenter to be released and to move to microtubule plus ends. Byreleasing Eg5 from the astral centre the balance of motor activity ischanged, allowing a spindle to form. This observation could haveprofound implications for spindle morphogenesis. In vitro, astralarrays of microtubules can still undergo Ran-induced bipolar reor-ganization even in the absence of chromosomes. This indicates thatspindle morphogenesis per se can be self-propagating, requiringonly the microtubule cytoskeleton and its associated proteins. The question is how a self-propagating process can acquire polarityinformation that would allow the reorganization of microtubulesinto a bipolar spindle. In our simplified in vitro system, Ran–GTPfirst induces microtubule asters from which spindles subsequentlyevolve. We found that the relative amount of minus-end-directedmicrotubule translocation is much higher in asters formed in anon-spindle-forming environment than in those formed in a
spindle-forming environment (in the presence of Ran–GTP). Thisindicates that when minus-end-directed motor activity is dominant(in the absence of Ran–GTP), microtubule asters cannot break theirradial symmetry and are therefore unable to form a bipolar spindle.Ran–GTP reduces the dominance of minus-end-directed motors,thereby providing the physical basis for bipolar-spindle formation.The resulting change in the balance of motor activities could gener-ate the key driving force behind this self-propagated reorganization.
In vitro, in the presence of Ran–GTP, spindle morphogenesiscould be a self-propagated process. However, in vivo, the functionof the spindle is to ensure equal segregation of chromosomes,which requires chromosomes to be aligned on a single spindle. Bytethering RCC1 to chromosomes, the cell has evolved a mechanismto use Ran–GTP to ensure that chromosomes are in the centre ofthe spindle. Thus, Ran–GTP seems to be a central regulator of spin-dle assembly.Note added in proof: A role for Ran-GTP in coordinating micro-tubule nucleation and dynamics is described in Carazo-Salas et al.39
MethodsAnalysis of microtubule dynamics.Microtubules were nucleated from centrosomes34 in Xenopus CSF-arrested egg extracts35 containing
rhodamine-labelled tubulin (0.3 µM final concentration) and an oxygen-scavenger system36 in the
presence of 25 µM Ran(L43E) or CSF-XB buffer35 at ~25 °C (using a temperature-controlled stage).
Microtubules were imaged every 2 s with a 250-ms exposure for ~5 min using a Nikon E800 micro-
scope equipped with a ×100 oil-immersion objective, a CCD (charge-coupled device) camera
(Princeton Scientific Instruments) and an automatic shutter (Sutter Instruments) driven by the
Metamorph software (Universal Imaging System). Dynamic parameters were as follows: microtubule
growth or shrinkage had to persist for at least four consecutive frames (~6 s), with a net change in
length of at least 0.5 µm. Growth and shrinkage rates were determined using the least-squares linear-
regression method in Microsoft Excel. Catastrophe and rescue frequencies were determined by divid-
ing the number of catastrophe or rescue events by the total time microtubules spent growing or
shrinking, respectively.
Assays for microtubule-spindle assembly.Ran(L43E) (25 µM final concentration), Taxol (1 µM final concentration), or DMSO (5% v/v final
concentration) were added to 10 µl of CSF-arrested egg extract containing rhodamine-labelled tubulin
and incubated at room temperature for 30 min. Microtubule structures were analysed as described23.
Microtubule-translocation assay.Asters were carried out as above but using fluorescein-labelled tubulin, and the reaction was incubated
for 10 min at room temperature. Before imaging, 1 µl of a saturated haemoglobin solution and 1 µl of
rhodamine-labelled ethylene glycol-bis-succinimidylsuccinate seeds37 were added to the reaction.
Images were obtained using 300-ms exposures at 2 s intervals and a ×60 oil-immersion lens. Polarity-
marked seeds were made as described38 and then stabilized as described37.
Anti-Eg5 antibodies.A GST–Eg5 stalk-domain fusion protein was used to purify stalk-domain-specific rabbit antibodies
raised against the stalk–tail domain of Eg5. The antibody was then labelled with rhodamine using a
tetramethyl–rhodamine succinimidyl ester (Molecular Probes) and dialysed into CSF-XB buffer35.
When added to Xenopus egg extract at a final concentration of 25 µg ml–1, the antibody did not inhibit
spindle formation (data not shown), but localized predominantly to astral centres and spindle poles,
with some staining along individual microtubules.
Kymograph analysis.Kymographs (cross-sectional views of the intensity values of a defined region of the time lapse) were
obtained using the kymograph function in Metamorph set to a width of 10, by drawing a line tracing a
microtubule bundle, beginning at the astral centre and finishing at the astral periphery. This was
repeated across the aster to ensure complete, non-overlapping coverage. A moving speckle forms a
sharply defined continuous diagonal line. To count speckles moving towards the aster periphery,
kymographs were enlarged by 200%.
Quantification of aster intensity.The tubulin content of asters in the first image of the time-lapse series was determined from the fluo-
rescence intensity of the fluorescein–tubulin signal detected by a cooled CCD camera using
Metamorph software (Universal Imaging System). The number of moving speckles was then divided
by the fluorescein–tubulin intensity value to give a speckle index for each aster.
RECEIVED 8 SEPTEMBER 2000; REVISED 1 NOVEMBER 2000; ACCEPTED 20 DECEMBER 2000;PUBLISHED 31 JANUARY 2001.
1. Waters, J. C. & Salmon, E. Pathways of spindle assembly. Curr. Opin. Cell Biol. 9, 37–43 (1997).
2. Heald, R. & Walczak, C. E. Microtubule-based motor function in mitosis. Curr. Opin. Struc. Biol. 9,
268–274 (1999).
3. Desai, A. & Mitchison, T. J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 13,

© 2001 Macmillan Magazines Ltd
articles
227
83–117 (1998).
4. Cassimeris, L. Accessory protein regulation of microtubule dynamics throughout the cell cycle.
Curr. Opin. Cell Biol. 11, 134–141 (1999).
5. Belmont, L. D., Hyman, A. A., Sawin, K. E. & Mitchison, T. J. Real-time visualization of cell cycle-
dependent changes in microtubule dynamics in cytoplasmic extracts. Cell 62, 579–589 (1990).
6. Verde, F., Dogterom, M., Stelzer, E., Karsenti, E. & Leibler, S. Control of microtubule dynamics and
length by cyclin A- and cyclin B-dependent kinases in Xenopus egg extracts. J. Cell Biol. 118,
1097–108 (1992).
7. Gliksman, N. R., Parsons, S. F. & Salmon, E. D. Okadaic acid induces interphase to mitotic like
microtubule dynamic instability by inactivating rescue. J. Cell Biol. 119, 1271–1276 (1992).
8. Zhai, Y., Kronebusch, P. J., Simon, P. M. & Borisy, G. G. Microtubule dynamics at the G2/M transi-
tion: abrupt breakdown of cytoplasmic microtubules at nuclear envelope breakdown and implica-
tions for spindle morphogenesis. J. Cell Biol. 135, 201–214 (1996).
9. Tournebize, R., Heald, R. & Hyman, A. Role of chromosomes in assembly of meiotic and mitotic
spindles. Prog. Cell Cycle Res. 3, 271–84 (1997).
10. Holy, T. E. & Liebler, S. Dynamic instability of microtubules as an efficient way to search in space.
Proc. Natl Acad. Sci. USA 91, 5682–5685 (1994).
11. Andersen, S. S. L. Balanced regulation of microtubule dynamics during the cell cycle: a contempo-
rary view. Bioessays 21, 53–60 (1999).
12. Karsenti, E., Newport, J., Hubble, R. & Kirschner, M. W. Interconversion of metaphase and inter-
phase microtubule arrays as studied by the injection of centrosomes and nuclei in eggs. J. Cell Biol.
98, 1730–1745 (1984).
13. Heald, R. et al. Self-organization of microtubules into bipolar spindles around artificial chromo-
somes in Xenopus egg extracts. Nature 382, 420–5 (1996).
14. Zhang, D. & Nicklas, R. Chromosomes initiate spindle assembly upon experimental dissolution of
the nuclear envelope in grasshopper spermatocytes. J. Cell Biol. 131, 1125–31 (1995).
15. Zhang, D. & Nicklas, R. The impact of chromosomes and centrosomes on spindle assembly as
observed in living cells. J Cell Biol 129, 1287–300 (1995).
16. Dogterom, M., Felix, M., Guet, C. & Leibler, S. Influence of M-phase chromatin on the anisotropy
of microtubule asters. J. Cell Biol. 133, 125–40 (1996).
17. Hoyt, M. A. & Giese, J. R. Genetic analysis of mitotic spindles. Annu. Rev. Genet. 30, 7–33 (1996).
18. Vale, R. D. & Milligan, R. A. The way things move: looking under the hood of molecular motors.
Science 288, 88–95 (2000).
19. Mountain, V., Simerly, C., Howard, L., Schatten, G. & Compton, D. A. The kinesin-related protein,
HSET, opposes the activity of Eg5 and cross-links microtubules in the mammalian mitotic spindle.
J. Cell Biol. 147, 351–365 (1999).
20. Sharp, D. J. et al. Functional coordination of three mitotic motors in Drosophila embryos. Mol. Biol.
Cell 11, 241–253 (2000).
21. Walczak, C. E., Vernos, I., Mitchison, T. J., Karsenti, E. & Heald, R. A model for the proposed roles
of different microtubule-based motor proteins in establishing spindle bi-polarity. Curr. Biol. 8,
903–913 (1998).
22. Kalab, P., Pu, R. T. & Dasso, M. C. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9,
481–484 (1999).
23. Wilde, A. & Zheng, Y. Stimulation of microtubule aster and spindle assembly by the small GTPase
Ran. Science 284, 1359–1362 (1999).
24. Ohba, T., Nakamura, M., Nishitani, H. & Nishimoto, T. Self-organization of microtubule asters
induced in Xenopus egg extracts by GTP-bound Ran. Science 284, 1356–1358 (1999).
25. Carazo-Salas, R. E. et al. Generation of GTP-bound Ran by RCC1 is required for chromatin-
induced mitotic spindle formation. Nature 400, 178–181 (1999).
26. Zhang, C., Hughes, M. & Clarke, P. R. Ran–GTP stabilises microtubule asters and inhibits nuclear
assembly in Xenopus egg extracts. J. Cell Sci. 112, 2453–2461 (1999).
27. Sawin, K. E., LeGuellec, K., Philippe, M. & Mitchison, T. J. Mitotic spindle organization by a plus-
end directed microtubule motor. Nature 359, 540–543 (1992).
28. Blangy, A. et al. Phosphorylation of p34 cdc2 regulates spindle association of human Eg5, a
kinesin-related motor essential for bi-polar spindle formation in vivo. Cell 83, 1159–1169 (1995).
29. Meyer, T. U. et al. Small molecule inhibitor of mitotic spindle polarity identified in a phenotype-
based screen. Science 286, 971–974 (1999).
30. Walczak, C. E., Verma, S. & Mitchison, T. J. XCTK2: a kinesin-related protein that promotes mitotic
spindle assembly in Xenopus laevis egg extracts. J. Cell Biol. 136, 859–870 (1997).
31. Heald, R., Tournebize, R., Habermann, A., Karsenti, E. & Hyman, A. Spindle assembly in Xenopus
egg extracts: respective roles of centrosomes and microtubule self-organization. J. Cell Biol. 138,
615–28 (1997).
32. Sawin, K. E. & Mitchison, T. J. Mutations in the kinesin-like protein Eg5 disrupting localization to
the mitotic spindle. Proc. Natl Acad. Sci. USA 92, 4289–4293 (1995).
33. Theurkauf, W. & Hawley, R. Meiotic spindle assembly in Drosophila females: behavior of nonex-
change chromosomes and the effects of mutations in the nod kinesin-like protein. J. Cell Biol. 116,
1167–80 (1992).
34. Mitchison, T. J. & Kirschner, M. W. Isolation of mammalian centrosomes. Methods Enzymol. 134,
261–268 (1986).
35. Murray, A. W. Cell cycle extracts. Methods Cell Biol. 36, 581–605 (1991).
36. Tournebize, R. et al. Distinct roles of PP1 and PP2A-like phosphatases in control of microtubule
dynamics during mitosis. EMBO. J 16, 5537–5549 (1997).
37. Fanara, P., Oback, B., Ashman, K., Podtelejnikov, P. & Brandt, R. Identification of MINUS, a small
polypeptide that functions as a microtubule nucleation suppressor. EMBO J. 18, 565–577 (1999).
38. Hyman, A. A. Preparation of marked microtubules for the assay of the polarity of microtubule
based motors by fluorescence. J. Cell Sci. (Suppl.) 14, 125–127 (1991).
39. Carazo-Salas, R. E., Gruss, O. J., Mattaj, I. W. & Karsenti, E. Ran–GTP coordinates regulation of
microtubule nucleation and dynamics during mitotic-spindle assembly. Nature Cell Biol. 3, 228-234
(2001).
ACKNOWLEDGEMENTSWe thank T. Kapoor and T. Mitchison (Harvard Medical School) for Monastrol, O. Martin for purify-ing Ran and for critical reading of the manuscript, L. Cassimeris (Lehigh Univ.) for advice onanalysing microtubule dynamics, E. Salmon (Univ. North Carolina) for advice on image analysis, A.Khodjakov (Wadsworth Center) for PKG cells used for centrosome isolation, and R. Gunawardane forcomments on the manuscript. This work was supported by a postdoctoral fellowship from theAmerican Cancer Society (to C.W.), NIH grants (to Y.Z. and C.E.W.), and a Pew Scholar’s award (toY.Z.).Correspondence and requests for materials should be addressed to Y.Z. Supplementary Information isavailable at http://www.ciwemb.edu/labs/zheng/zhenglab.html.
NATURE CELL BIOLOGY VOL 3 MARCH 2001 http://cellbio.nature.com