31p nmr studies of spinach leaves and their chloroplasts* · 31p nmr studies of spinach leaves and...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 265, No. 3, Issue of January 25, pp. 1319-1326, 1990 Printed in CJ. S. A.
31P NMR Studies of Spinach Leaves and Their Chloroplasts*
(Received for publication, August 15, 1989)
Richard Bligny, Per Gardestrom+, Claude Roby, and Roland Douceg From the Laboratoire de Physiologie Cellulaire VkgPtale, URA Centre National de la Recherche Scientifique 576, Dipartement de Recherche Fondamentale, Centre d%tudes Nuclkaires de Grenoble et Uniuersitt? Joseph Fourier, 85X, Grenoble-Ckdex F-38041, France
An experimental arrangement is described which enables high quality 31P NMR spectra of compressed spinach leaf pieces to be continuously recorded in which all the resonances observed (cytoplasmic and vacuolar Pi, glycerate-3-P, nucleotides) were sharp and well resolved. 31P NMR spectra obtained from intact chloroplasts showed a distinct peak of stromal Pi. An upfield shift of the stromal Pi resonance was associated with a decrease in the external Pi and vice versa. Nucleotides were largely invisible to NMR in intact chloroplasts, whereas the same nucleotides reap- peared in a typical 31P NMR spectrum of an acid ex- tract of intact chloroplasts. Perfusion of compressed spinach leaf pieces with a medium containing Pi trig- gered a dramatic increase in the vacuolar Pi over 12 h. Addition of choline to the Pi-free perfusate of com- pressed leaf pieces resulted in a steady accumulation of phosphorylcholine in the cytoplasmic compartment at the expense of cytoplasmic Pi. When a threshold of cytoplasmic Pi concentration was attained, Pi was drawn from the vacuole to sustain choline phosphoryl- ation. In spinach leaves, the vacuole represents a po- tentially large Pi reservoir, and cycling of Pi through vacuolar influx (energy dependent) and efflux path- ways is an efficient system that may provide for control over the cytosolic-free Pi and phosphorylated inter- mediate concentrations. ‘lP NMR spectra of neutral- ized perchloric acid extracts of spinach leaves showed well defined multipeak resonances (quadruplet) of in- tracellular phytate. The question of cytosolic Pi con- centration in green cells is discussed.
In photosynthetic higher plant cells, cytosolic inorganic phosphate (Pi) plays a central role in the regulation of respi- ration (1) and photosynthesis (2). Unfortunately, by usual analytical techniques it is almost impossible to distinguish between the Pi contents of the vacuolar and cytoplasmic compartments. This limitation has been partially overcome with the relatively recent expansion of nuclear magnetic resonance (NMR) spectroscopy to studies of intact nongreen plant tissues (3-5) and leaves (6, 7). However, even with this methodology, the investigation of the relative concentrations of ATP and cytosolic Pi in photosynthetic cells, which are of great usefulness and importance in the study of the control of photosynthesis, have been hindered for two major reasons: (a) more than 80% of the volume of mature photosynthetic
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Dept. of Biochemistry, University of Umea, Umea 590187, Sweden.
I To whom correspondence should be addressed.
cells consists of a vacuole (8); and (b) the lacuna between upper and lower epidermis represents a large proportion of the total leaf volume.
Despite the aforementioned difficulties, in the present stud- ies an experimental arrangement is described for monitoring by ‘lP NMR the behavior of spinach leaves over a long period of time under different conditions. Our data shed new light on the cytosolic Pi status and on the controlled flux of Pi into and out of the vacuole in photosynthetic tissues.
EXPERIMENTAL PROCEDURES
Plant Material-Mature spinach (Spinacia oleracea L.) leaves were obtained from the local market and cut into small pieces (approxi- mately 1 mm*) directly into ice-cold perfusion medium (0.28 M mannitol, 0.05% (w/v) insoluble polyvinylpyrrolidone (Kca 25, Serva), 0.05% bovine serum albumin, and 5 mM MES’ buffer (pH 6.5)) containing 20 mM sucrose and 10 pM Pi. The small leaf pieces were infiltrated with perfusion medium under slight vacuum (7-9) and were stored at 10 ‘C under dark conditions until use.
Isolation of Intact Chloroplasts-Infiltrated leaves were introduced into ice-cold chloroplast extraction medium (330 mM mannitol, 30 mM MOPS-NaOH (DH 7.8). 2 mM EDTA. and 0.15% bovine serum
,A I
albumin) with a tissue/medium volume ratio of 1:3. Leaves were homogenized three times for 2 s each with a Waring blender, and intact chloroplasts were prepared as fast as possible according to the method of Nakatani and Barber (10). Chloroplasts thus obtained were purified by isopycnic centrifugation in a nontoxic silica sol (Percoll, Pharmacia LKB Biotechnology Inc.) gradient that main- tained isoosmotic conditions throughout the isolation procedure ac- cording to the method of Mourioux and Deuce (11). The entire isolation procedure could be accomplished in less than 30 min. At this point in the procedure, the thick chloroplast suspension was used for the measurement of CO,-dependent O2 evolution rates and P, content of the stroma (11).
P, Determination-An aliquot (50 ~1; 350-400 pg of chlorophyll) of the chloroplast suspension was suspended in 5 ml of extraction medium. The tube was centrifuged at 3 “C for 1 min at 1500 X g (Sorvall SM 24 rotor). The supernatant was removed by aspiration, and the pellet was resuspended in a low osmolarity medium (10 mM MES (pH 6), 2 mM EDTA; final volume, 1.1 ml). One hundred ~1 of this suspension was used for chlorophyll determination. The remain- ing suspension was centrifuged 10 min at 7000 X g (Sorvall SM 24 rotor) after addition of 200 ~1 of 20% trichloroacetic acid. A fraction of the clear supernatant (1 ml) devoid of mannitol, which interferes with phosphomolybdate complex formation, was used for the colori- metric determination of P, after isobutyl alcohol extraction (12).
31P NMR of Spinach Leaues-A Bruker NMR spectrometer (AM 400) operating in the pulsed Fourier transform mode at 162 MHz was used to characterize the various intracellular pools of Pi and phos- phorus compounds known to be present in leaves. In order to get a better signal-to-noise ratio, NMR experiments on leaves required small leaf pieces densely packed within a limited volume approxi- mating that of the NMR receiver coil (volume within the detection
’ The abbreviations used are: MES, 4-morpholineethanesulfonic acid; MOPS, 4-morpholinepropanesulfonic acid, FID(s), free induc- tion decay; CDTA, trans-1,2-diaminocyclohexane-N,N,N’,N’-tetra- acetic acid; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid; UDPG, uridine 5’-diphosphate glucose.
1319
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

1320 31P NMR Studies of Spinach Leaves and Their Chloroplasts
coil, 4.5 ml). To this end, small leaf pieces infiltrated with perfusion medium (7) were slightly compressed by hand between two circular polymer filters from a volume of 20 ml (150 pg of chlorophyll/ml) to a volume of 4.5 ml (666 pg of chlorophyll/ml) fitting exactly into a standard 15mm NMR tube. Under our condition, 1 g (wet weight) of leaf tissue corresponded to 1 mg of chlorophyll. Details of this assembly and its operation have been described previously by Roby et al. (13). A reference capillary containing methylenediphosphonate (pH 8.9 in 30 mM Tris) was inserted inside the inlet tube along the symmetry axis of the tissue sample. The perfusate consisted of the perfusion medium containing 20 mM sucrose. Its pH was constantly monitored and adiusted to 6.5 using an automatic device. The flow rate routinely provided more than twice the measured oxygen con- sumption by 3 g (wet weight) of spinach leaves at 25 “C. Such a system prevented gas bubbles from collecting in the intercellular space sandwiched between upper and lower epidermis, which would have otherwise decreased the homogeneity of the magnetic field. For kinetic experiments on respiration, the gas flow (see Ref. 13) was switched alternately from O2 to NP. Furthermore, by modifying the composition of the circulating medium (introduction of P,, choline) it was possible to perturb the leaf metabolism and to monitor the spectra1 changes simultaneously, obtaining several successive spectra from the same sample. For example, changes from aerobic to anaer- obic conditions and vice versa could be performed at intervals as short as every 2 min.
The assignment of nucleotides and phosphate esters to specific peaks was carried out according to N&on-et al. (14), Evans and Kaplan (15). and Roberts and Jardetzkv (3) and from snectra of the perchloric extracts that contained the soluble low molecular weight constituents (13). A 5-s recycling time was used to obtain fully relaxed spectra (for P, determination), but for qualitative purposes, FIDs were accumulated using a 0.3-s repetition time and a 30’ pulse angle.
Neutralized Perchloric Acid Extracts-For nerchloric acid extracts. spinach leaves (5 g, wet weight) or intact-chloroplasts (8 mg of chlorophyll) were quickly frozen at liquid nitrogen temperature to avoid ATP destruction and then finely ground with 1 ml of 70% (w/ v) perchloric acid. The frozen powder was then placed at -10 “C and thawed. The thick suspension thus obtained was centrifuged at 10,000 x g for 10 min to remove particulate matter. The supernatant was neutralized with 2 M KHCOS to about pH 6.0 and centrifuged at 10,000 X a for 10 min to remove nrecinitated KClO,. Next. 120 umol of’ trnns-i,2-diaminocyclohexane-~,~,~~,N’-tetraacetic acid (CD- TA), a strong chelating agent (16), was added. CDTA prevented the precipitation of specific P, esters, such as phytate, as magnesium salts. It was also more effective than EDTA in eliminating free divalent metal ions and led to sharp lines for the phosphorus reso- nances of metabolites such as ATP. The resulting supernatant was lyophilized and stored at liquid nitrogen temperature. For the NMR measurements, this freeze-dried material containing 120 rmol of CDTA was redissolved in 2 ml of 40 mM HEPES buffer (pH 7.8) containing 10% D20 (perchloric acid extract).
The 31P NMR spectra of neutralized perchloric acid extracts were measured on a Bruker NMR spectrometer (AM 400) equipped with a lo-mm multinuclear probe tuned at 162 MHz according to Rohy et al. (13). Spectra of standard solutions of known phosphate compounds at pH 7.8 were compared with that of a perchloric acid extract of spinach leaves. The resonances of the standards (ATP, ADP, AMP, CTP, GTP, UTP, UMP, CMP, TMP, NAD, NADP; glucose-6-P, elucose-1-P. UDP-xlucose. UDP-ealactose. ADP-alucose. nlvcerate- i-P, glyceraldehyde-3-P, ‘sn-glycerol-3-P,’ pyrophosphate, glycero- phosphorylcholine, phosphorylcholine, and phytate) were resolvable. Definitive attribution of resonances to specific compounds was made after running a series of spectra obtained by successive addition of the authentic compounds to the perchloric acid extracts. In the case of phosphorylcholine and phytate, these compounds were added to the extracts, and the pH titrations were carried out to further sub- stantiate the assignments.
RESULTS
Conditions for Incubation--Small leaf pieces were prepared in perfusion medium without sucrose and Pi as described under “Experimental Procedures.” The leaf pieces were then divided into four parts and incubated in the dark at room temperature with added 20 mM sucrose and 1 mM Pi (Table I). At different times during incubation, the capacity to per- form photosynthesis at saturating light and COZ was meas-
TABLE I Effect of sucrose feeding of spinach leaf pieces on the photosynthesis
rate Leaves were cut into small pieces and infiltrated in the ice-cold
perfusion medium (see “Plant Material”). At time 0, the leaf pieces were transferred to medium at 22 “C with additions as indicated. 02 evolution was followed polarographically at 25 “C by using a Clark- type O2 electrode system (47). The reaction medium devoid of sucrose and containing 5 mM HCO, was gassed with argon before the addition of leaf pieces equivalent to 50-100 pg of chlorophyll. Light was provided by a 150-watt xenon arc lamp source (Oriel corporation), giving an irradiance of 300 watts. me2 at the surface of the vessel.
Addition Time 0, evolution
h rmol 01. h-‘.I& chlorophyll
No addition 1 51 3 28
24 8 20 mM sucrose 3 85
5 116 24 142
lmMPi 3 18 24 4
20 mM sucrose + 1 mM Pi 3 95 24 120
ured. In order to prevent a possible inhibition of photosyn- thesis by the carbohydrate in leaf pieces (48), photosynthetic rates were measured in the absence of sucrose in the medium. When sucrose was omitted from the incubation medium, the leaf pieces rapidly lost the capacity to perform photosynthesis (Table I). When sucrose was added alone or together with Pi, a high rate of photosynthesis was observed even after 36 h at room temperature using a sterile perfusion medium. Pi alone was, however, not sufficient to maintain a high photosynthetic activity. The initial response of leaf pieces to sucrose feeding varied depending on the size of the intracellular carbohydrate pools (starch, sucrose). For some leaves, the initial rate of photosynthesis was high in the absence of added sucrose; but to get good rates of photosynthesis after longer incubation times, addition of sucrose was always necessary. By using 31P NMR (see below) we have observed that the concentrations of the major intracellular metabolites (glucose-6-P, UDP- glucose, and glycerate-3-P) declined progressively once the intracellular sucrose pool had disappeared (not shown; for an explanation, see Ref. 13). In marked contrast, in the presence of sucrose in the perfusion medium, little change occurred. We have observed that a high rate of photosynthesis was always correlated with high intracellular metabolite levels. Interestingly, Stitt et al. (17) have shown clearly that higher levels of phosphorylated intermediates in leaf cells shorten the length of the induction phase of photosynthesis.
31P NMR of Compressed Small Leaf Pieces-slP NMR spec- tra obtained from compressed leaf pieces supplied with oxygen and at pH 6.5 showed two distinct peaks of intracellular Pi (Cyt-Pi and Vat-Pi, Fig. 1) at approximately 2.4 and 0.5 ppm. These positions correspond roughly to pH 7.5 and 5, respec- tively. These values reflect the presence of the vacuolar Pi pool at the acidic pH and the cytoplasmic Pi pool at the slightly alkaline pH (18). In the spectra obtained from spinach leaves, no other Pi peaks were identified unambiguously (see also Refs. 7-9). Such a result is surprising considering that in spinach mesophyll cells, plastids take up a much higher pro- portion of the cytoplasmic volume than in nonchlorophyllian cells (6). This means that in the dark, either the amount of Pi present in the chloroplast stroma of spinach leaf is below the level of sensitivity of NMR and/or the pH difference between chloroplasts and the cytosol is too small for the signals to be discriminated. The first hypothesis is most
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

31P NMR Studies of Spinach Leaves and Their Chloroplasts 1321
. -5 -10 -15 -20
Chemical shift O), PPm
FIG. 1. Representative 31P NMR spectrum (162 MHz) of compressed spinach leaf pieces. Leaf pieces (3 g, wet weight) were first maintained in a well aerated perfusion medium containing 20 mM sucrose to increase the intracellular pools of phosphoesters. After 10 h, leaf pieces were compressed by hand between two filters. The cell volume comprises approximately 66% of the total volume. The perfusion medium was pumped through the compressed leaf pieces and was circulated via a 300-ml reservoir that was oxygenated by 02 bubbling. The FID recorded at 25 “C is the result of 96,000 transients obtained with a 30” pulse angle and 0.3-s repetition time. Data treatments include Lorentz-Gauss transformation and zero filling. Peak assignments: Glc-6-P, glucose-6-P; a, position of glycerate-3-P; b, position of phosphorylcholine; Cyt-Pi, cytoplasmic Pi; Vat-P,, vac- uolar P,.
unlikely because the concentrations of Pi we have found in the stroma of freshly prepared chloroplasts was approximately 12 mM (we have verified that during the course of chloroplast preparation carried out at 0 “C, the slow leakage of Pi from intact isolated chloroplasts was almost negligible (11); con- sequently, the Pi content of intact chloroplasts reflected the in viuo situation). To determine accurately the cytoplasmic and vacuolar Pi pools in leaf tissue, a calibration of the peak intensity of the Pi resonance with known amounts of external Pi was first performed. In our experimental conditions, we have calculated that within the receiving coil the cell volume comprised approximately 66% of the total volume. By consid- ering the ratio of vacuolar volume to cytoplasmic volume (roughly 8.5), the curve thus generated gave estimates of cytoplasmic Pi levels ranging from 6 to 9 mM for the average concentration in the chloroplast, cytosol, and other cell or- ganelles in the dark. Assuming that the chloroplasts comprise more than 60% of the cytoplasmic volume (40 bl/mg of chlorophyll) (17) and that the concentration of mobile Pi in the chloroplast is approximately 12 mM, our results suggest strongly that in the dark the concentration of cytosolic Pi is much lower than that of chloroplast stroma. Owing to obvious technical problems associated with the illumination of packed mesophyll cells, we were unable to follow the fluctuation of cytoplasmic P, concentration which might be expected in the dark/light transition and vice versa (2, 17).
In the representative 31P NMR spectrum of spinach leaves (Fig. l), there were also signals from cytoplasmic glucose-6- P, p- and (Y-UDPG (with contribution of some NAD(P) to the P-peak of UDPG), y-, o(-, and P-phosphorus of NTP. The region of the spectrum to high field of glucose-6-P corre- sponded to glycerate-3-P (peak a). However, other minor peaks resided underneath it. The resonance at 3.5 ppm was attributable to NMP and phosphorylcholine (peak b).
However, meaningful interpretations of in uiuo spectra re- quire the rigorous identification of resonances corresponding
to specific phosphorus-containing metabolites. Numerous studies have shown that the region of the 31P NMR spectrum, generally ascribed to sugar phosphates in previous investiga- tions of a variety of plant tissues or cells (19), may contain other phosphomonoesters including lipid metabolites. These considerations point to the need for unambiguous determi- nation of the original peaks.
3’P NMR of Neutralized Perchloric Acid Extracts-Fig. 2 shows a typical undecoupled 31P NMR spectrum of a perchlo- ric acid extract of spinach leaf pieces (5 g, wet weight; 5 mg of chlorophyll) at pH 7.8 in the presence of a large excess of CDTA. The resonances that were observed were sharp insofar as the concentration of phosphorylated metabolites was not too high. By adding known compounds to the extract, the peaks obtained were identified as signals from phosphomon- oesters including dihydroxyacetone phosphate, gluconate-6- P, glucose-6-P, glycerate-3-P, sn-glycerol-3-P, and phospho- rylcholine; Pi; y-, (Y-, and ,&ATP; y-, a-, and P-UTP; fi-, and a-ADP; @-, and by-UDPG; @- and a-UDP-galactose. As ex- pected, ATP was the most abundant triphosphate. The NAD and NADP peaks, which yield a quadruplet centered at -10.7, -10.9, -11.1, and -11.2 ppm on the high field site of (r-NTP peak, were well identified in contrast with what was observed previously in nongreen tissues (19). However, NADH and NADPH are acid labile (20), and consequently, these reduced coenzymes are certainly destroyed during the course of per- chloric acid treatment of intact tissues. We observed large amounts of phytate in the perchloric acid extracts from spin- ach leaves when they contained very high concentrations of CDTA (this chelating agent prevented the precipitation of phytate which is highly insoluble as magnesium salt). Phytate appeared as a complex multipeak resonance (quadruplet) showing absorption bands at approximately l-2 ppm in our t experimental conditions. The exact position of this quadruplet
0 -10 -20 Chemical shift (~1, ppm
FIG. 2. Proton-decoupled 31P NMR spectrum of perchloric acid extract of spinach leaves. Perchloric acid extract was pre- pared from leaf pieces (3 g, wet weight). Freeze-dried extracts were solubilized and supplemented with CDTA as described under “Ex- perimental Procedures.” The sample (2.5 ml) containing 300 ~1 of *HZ0 was analyzed at 25 “C with a lo-mm multinuclear probe tuned at 162 MHz. 2048 FIDs were acquired with a 90” pulse angle and a recycling time of 0.4 s under proton decoupling. Two levels of broad band ‘H decoupling were employed: 2.5 watts during the data acqui- sition, and 0.5 watt during the delay period. The resulting FID memorized over 8,000 data was Fourier transformed over 32,000 after gaussian multiplication. All the interesting regions are shown on an expanded scale. Peak assignments: a, unidentified peak; b, position of fructose-6-P, ribose-5-P, and phosphorylethanolamine; P,, total inorganic phosphate; c, unidentified peak (phosphodiester?); DHAP, dihydroxyacetone phosphate; PGA, glycerate-3-P, GPC, glycerophos- phorylcholine; PEP, phosphoenolpyruvate.
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from
1322 31P NMR Studies of Spinach Leaves and Their Chloroplasts
was highly sensitive to pH and to the chemical environment, making its correct identification difficult. Since this phospho- rus compound is not easily NMR detectable in uiuo (Mg2 phytate is insoluble), it was not possible to determine in which compartment (vacuole or cytoplasm) it was accumulated. The peak at 1 ppm (peak c in Fig. 2) close to that of phytate was not assigned. Since the position of this peak did not change when pH was varied from near 6 to near 8, it was very likely a phosphodiester. Its characterization is under investigation in our laboratory. The peak at 0 ppm has been ascribed to glycerophosphorylcholine. An additional resonance was seen at 0.5 ppm and was tentatively identified as phosphoenolpy- ruvate. There are many metabolites that are known to be present in the NMR spectra of the extracts. Among these are fructose 1,6-bisphosphate, pyrophosphate, and glyceralde- hyde-3-P. We estimate that in our conditions using 5 g of spinach leaves, the concentration of phosphorylated com- pounds lower than 100 WM in the cytoplasmic compartment was not distinguishable from the background noise.
The high NTP/NDP ratio as seen in uiuo (Fig. 1) (the /3- NDP peak is almost invisible) was not observed for cell perchloric acid extracts (Fig. 2). Likewise, in the case of maize (Zea muys L.) root tips, NTP/NDP ratios determined in uiuo by 31P NMR were always higher than ratios observed in extracts (21). Since little if any hydrolysis of ATP had oc- curred during the extraction procedure, these results indicate that ADP became visible to NMR during the process of HClO, extraction and strongly suggest that ADP in uiuo is invisible to NMR because it is either bound to some paramagnetic metals or sequestered in a large protein-rich compartment (e.g. chloroplast stroma). In order to confirm this hypothesis, a careful analysis of the 31P NMR spectra of spinach chloro- plasts has been carried out.
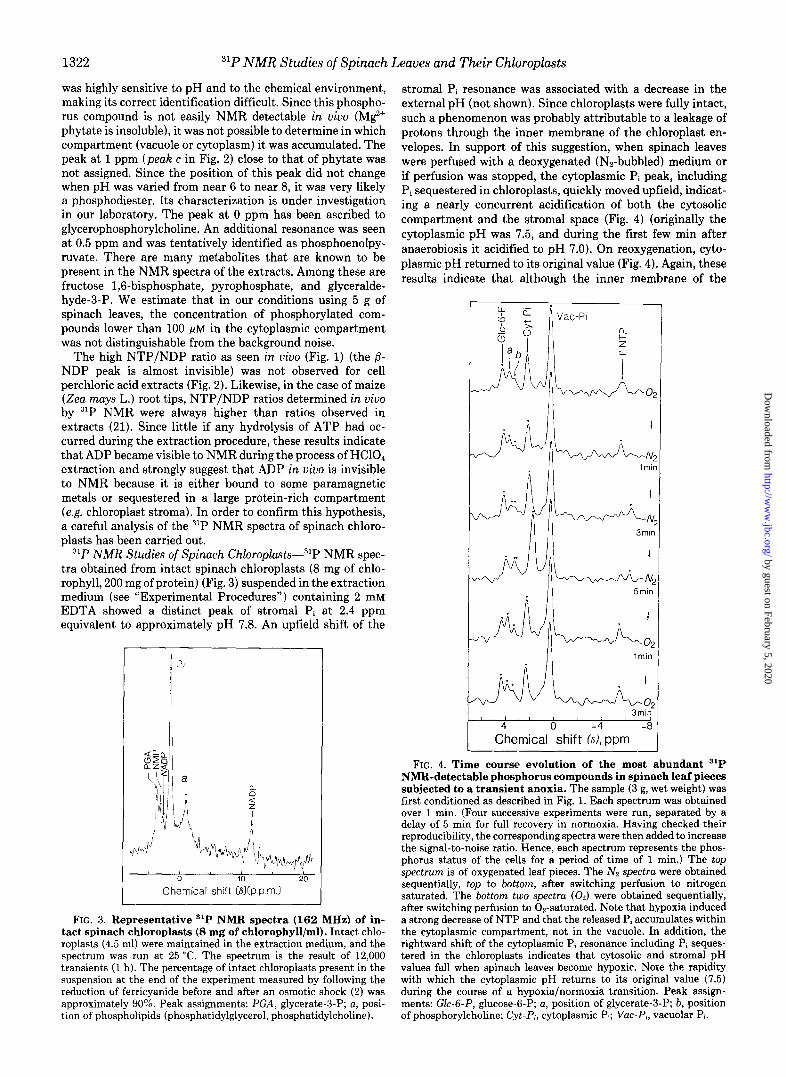
31P NMR Studies of Spinach Chloroplasts-31P NMR spec- tra obtained from intact spinach chloroplasts (8 mg of chlo- rophyll, 200 mg of protein) (Fig. 3) suspended in the extraction medium (see “Experimental Procedures”) containing 2 mM EDTA showed a distinct peak of stromal Pi at 2.4 ppm equivalent to approximately pH 7.8. An upfield shift of the
I Pi
Chemical shift (8b.p.m.)
FIG. 3. Representative ‘lP NMR spectra (162 MHz) of in- tact spinach chloroplasts (8 mg of chlorophyll/ml). Intact chlo- roplasts (4.5 ml) were maintained in the extraction medium, and the spectrum was run at 25 “C. The spectrum is the result of 12,000 transients (I h). The percentage of intact chloroplasts present in the suspension at the end of the experiment measured by following the reduction of ferricyanide before and after an osmotic shock (2) was approximately 90%. Peak assignments: PGA, glycerate-3-P; a, posi- tion of phospholipids (phosphatidylglycerol, phosphatidylcholine).
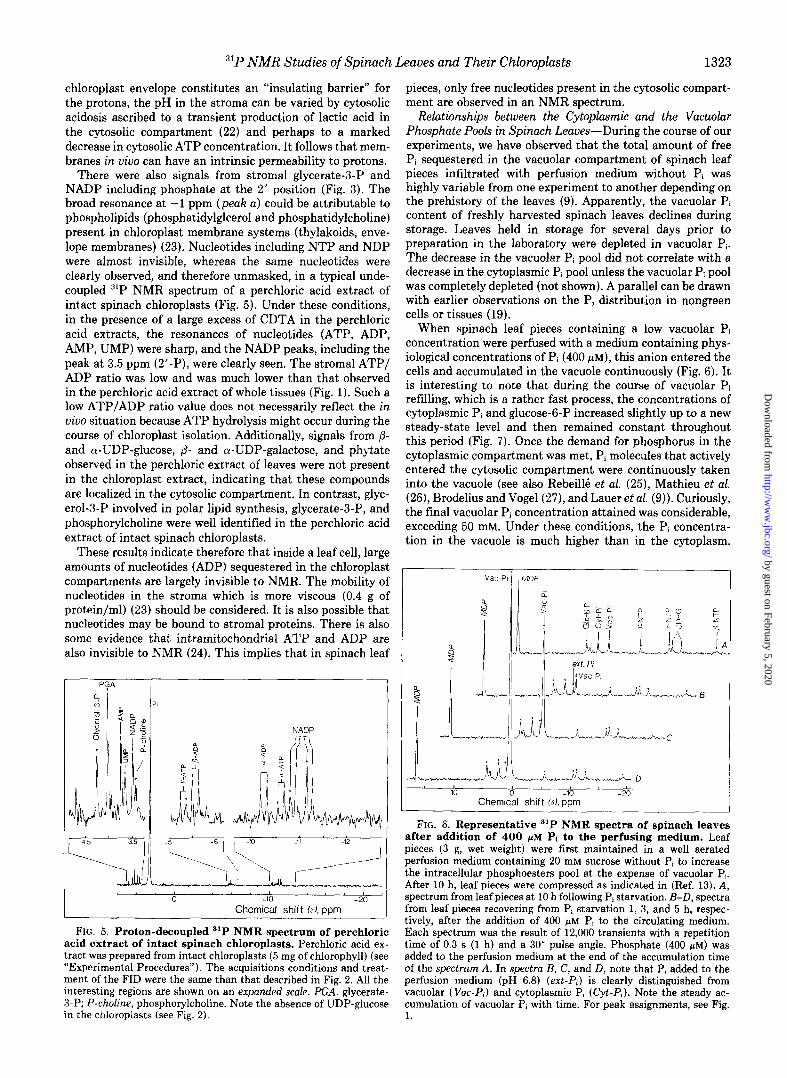
stromal Pi resonance was associated with a decrease in the external pH (not shown). Since chloroplasts were fully intact, such a phenomenon was probably attributable to a leakage of protons through the inner membrane of the chloroplast en- velopes. In support of this suggestion, when spinach leaves were perfused with a deoxygenated (Nz-bubbled) medium or if perfusion was stopped, the cytoplasmic Pi peak, including Pi sequestered in chloroplasts, quickly moved upfield, indicat- ing a nearly concurrent acidification of both the cytosolic compartment and the stromal space (Fig. 4) (originally the cytoplasmic pH was 7.5, and during the first few min after anaerobiosis it acidified to pH 7.0). On reoxygenation, cyto- plasmic pH returned to its original value (Fig. 4). Again, these results indicate that although the inner membrane of the
lmin
4
Chemical ‘shift &yppm -8
FIG. 4. Time course evolution of the most abundant 31P NMR-detectable phosphorus compounds in spinach leaf pieces subjected to a transient anoxia. The sample (3 g, wet weight) was first conditioned as described in Fig. 1. Each spectrum was obtained over 1 min. (Four successive experiments were run, separated by a delay of 5 min for full recovery in normoxia. Having checked their reproducibility, the corresponding spectra were then added to increase the signal-to-noise ratio. Hence, each spectrum represents the phos- phorus status of the cells for a period of time of 1 min.) The top spectrum is of oxygenated leaf pieces. The Nz spectra were obtained sequentially, top to bottom, after switching perfusion to nitrogen saturated. The bottom two spectra (02) were obtained sequentially, after switching perfusion to 0,.saturated. Note that hypoxia induced a strong decrease of NTP and that the released P, accumulates within the cytoplasmic compartment, not in the vacuole. In addition, the rightward shift of the cytoplasmic Pi resonance including Pi seques- tered in the chloroplasts indicates that cytosolic and stromal pH values fall when spinach leaves become hypoxic. Note the rapidity with which the cytoplasmic pH returns to its original value (7.51 during the course of a hypoxia/normoxia transition. Peak assign- ments: G&3-P, glucose-6-P; a, position of glycerate-3-P; b, position of phosphorylcholine; Cyt-P,, cytoplasmic Pi; VW-P,, vacuolar Pi.
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from
3iP NMR Studies of Spinach Leaves and Their Chloroplasts 1323
chloroplast envelope constitutes an “insulating barrier” for the protons, the pH in the stroma can be varied by cytosolic acidosis ascribed to a transient production of lactic acid in the cytosolic compartment (22) and perhaps to a marked decrease in cytosolic ATP concentration. It follows that mem- branes in uiuo can have an intrinsic permeability to protons.
There were also signals from stromal glycerate-3-P and NADP including phosphate at the 2’ position (Fig. 3). The broad resonance at -1 ppm (peak a) could be attributable to phospholipids (phosphatidylglcerol and phosphatidylcholine) present in chloroplast membrane systems (thylakoids, enve- lope membranes) (23). Nucleotides including NTP and NDP were almost invisible, whereas the same nucleotides were clearly observed, and therefore unmasked, in a typical unde- coupled 31P NMR spectrum of a perchloric acid extract of intact spinach chloroplasts (Fig. 5). Under these conditions, in the presence of a large excess of CDTA in the perchloric acid extracts, the resonances of nucleotides (ATP, ADP, AMP, UMP) were sharp, and the NADP peaks, including the peak at 3.5 ppm (2’-P), were clearly seen. The stromal ATP/ ADP ratio was low and was much lower than that observed in the perchloric acid extract of whole tissues (Fig. 1). Such a low ATP/ADP ratio value does not necessarily reflect the in uivo situation because ATP hydrolysis might occur during the course of chloroplast isolation. Additionally, signals from /3- and Ly-UDP-glucose, /3- and a-UDP-galactose, and phytate observed in the perchloric extract of leaves were not present in the chloroplast extract, indicating that these compounds are localized in the cytosolic compartment. In contrast, glyc- erol-3-P involved in polar lipid synthesis, glycerate-3-P, and phosphorylcholine were well identified in the perchloric acid extract of intact spinach chloroplasts.
These results indicate therefore that inside a leaf cell, large amounts of nucleotides (ADP) sequestered in the chloroplast compartments are largely invisible to NMR. The mobility of nucleotides in the stroma which is more viscous (0.4 g of protein/ml) (23) should be considered. It is also possible that nucleotides may be bound to stromal proteins. There is also some evidence that intramitochondrial ATP and ADP are also invisible to NMR (24). This implies that in spinach leaf
PGA I
NADP
-10 -20 Chemical shift OJ, PPm
pieces, only free nucleotides present in the cytosolic compart- ment are observed in an NMR spectrum.
Relationships between the Cytoplasmic and the Vacuolar Phosphate Pools in Spinach Leaues-During the course of our experiments, we have observed that the total amount of free Pi sequestered in the vacuolar compartment of spinach leaf pieces infiltrated with perfusion medium without Pi was highly variable from one experiment to another depending on the prehistory of the leaves (9). Apparently, the vacuolar Pi content of freshly harvested spinach leaves declines during storage. Leaves held in storage for several days prior to preparation in the laboratory were depleted in vacuolar Pi. The decrease in the vacuolar Pi pool did not correlate with a decrease in the cytoplasmic Pi pool unless the vacuolar Pi pool was completely depleted (not shown). A parallel can be drawn with earlier observations on the P, distribution in nongreen cells or tissues (19).
When spinach leaf pieces containing a low vacuolar Pi concentration were perfused with a medium containing phys- iological concentrations of Pi (400 PM), this anion entered the cells and accumulated in the vacuole continuously (Fig. 6). It is interesting to note that during the course of vacuolar Pi refilling, which is a rather fast process, the concentrations of cytoplasmic Pi and glucose-6-P increased slightly up to a new steady-state level and then remained constant throughout this period (Fig. 7). Once the demand for phosphorus in the cytoplasmic compartment was met, Pi molecules that actively entered the cytosolic compartment were continuously taken into the vacuole (see also Rebeill6 et al. (25), Mathieu et al. (26), Brodelius and Vogel (27), and Lauer et al. (9)). Curiously, the final vacuolar Pi concentration attained was considerable, exceeding 50 mM. Under these conditions, the Pi concentra- tion in the vacuole is much higher than in the cytoplasm
V8C.F MDP
10 Chemic\l shift f;*J, bb9n
-20
FIG. 5. Proton-decoupled 31P NMR spectrum of perchloric acid extract of intact spinach chloroplasts. Perchloric acid ex- tract was prepared from intact chloroplasts (5 mg of chlorophyll) (see “Experimental Procedures”). The acquisitions conditions and treat- ment of the FID were the same than that described in Fig. 2. All the interesting regions are shown on an expanded scale. PGA, glycerate- 3-P; P-choline, phosphorylcholine. Note the absence of UDP-glucose . . . . - ~. m the chloroplasts (See Fig. 2).
after addition of 400 I.~M Pi to the perfusing medium. Leaf pieces (3 g, wet weight) were first maintained in a well aerated perfusion medium containing 20 mM sucrose without Pi to increase the intracellular phosphoesters pool at the expense of vacuolar Pi. After 10 h, leaf pieces were compressed as indicated in (Ref. 13). A, spectrum from leaf pieces at 10 h following Pi starvation. B-D, spectra from leaf pieces recovering from Pi starvation 1, 3, and 5 h, respec- tively, after the addition of 400 @M Pi to the circulating medium. Each spectrum was the result of 12,000 transients with a repetition time of 0.3 s (1 h) and a 30” pulse angle. Phosphate (400 pM) was added to the perfusion medium at the end of the accumulation time of the spectrum A. In spectra B, C, and D, note that P, added to the perfusion medium (pH 6.8) (at-P,) is clearly distinguished from vacuolar ( Vat-P,) and cytoplasmic P, (@t-P;). Note the steady ac- cumulation of vacuolar P; with time. For peak assignments, see Fig. 1
FIG. 6. Representative 31P NMR spectra of spinach leaves
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

1324 31P NMR Studies of Spinach Leaves and Their Chloroplasts
NTP nj.-,..
0 5 10 Time, hours
FIG. 7. Time course evolution of the most abundant a1P NMR-detectable phosphorus compounds in spinach leaves perfused with a medium containing 400 pM Pi. The concentra- tions of mobile phosphorus compounds in the leaf tissue (average over the total sample within the detector) were determined as de- scribed under “Experimental Procedures.” Note the dramatic accu- mulation of Pi in the vacuole ( Vuc-I’,). Cyt-Pi, cytoplasmic P,.
i vat-PI
10 0 -10 -20 Chemical shift (A), ppm
FIG. 8. Representative 31P NMR spectra of spinach leaves after addition of 50 pM choline in the perfusing medium. Leaf pieces (3 g, wet weight) were compressed as indicated in Ref. 13 0 h, standard spectrum; 2 h and 8 h, spectra obtained after 2 and 8 h, respectively, of perfusion of the compressed leaf pieces with a medium containing 50 fiM choline. Each spectrum was the result of 12,000 transients with a repetition time of 0.3 s (1 h) and a 30” pulse angle. Note the steady accumulation of phosphorylcholine (P-choline) with time. Peak assignments are the same as in Fig. 1.
Such a result indicates that the sequestration of Pi in the vacuole is an energy-dependent process. In support of this suggestion, we have observed that whenever the cytosolic ATP concentration is substantial, excess cytosolic Pi is preferen- tially expelled toward the vacuole (19). On the contrary, when the ATP concentration is low (i.e. during transition from aerobic to anaerobic conditions) (19) (Fig. 4), Pi derived from nucleotide and glucose-6-P pools stays in the cytoplasmic compartment. It is clear therefore that in the case of spinach leaf cells, the vacuole represents a potentially large Pi reser- voir.
Conversely, we also studied the availability of vacuolar Pi for cellular metabolism (8, 19, 28) in spinach leaves. To do this, we examined the effects that choline, a powerful new Pi- sequestering agent (29), has on the fluctuation of the intra- cellular Pi pool in spinach leaves. This compound is phos- phorylated by a cytosolic choline kinase to form phosphoryl- choline, which is not subsequently metabolized. Fig. 8 illus- trates the changes that occurred in the compressed leaf pieces when choline (50 pM) was added to the perfusion medium
devoid of Pi. At first, compressed leaf pieces were maintained for 10 h in a continuously oxygenated circulating medium containing sucrose in order to fill the cytoplasmic compart- ment with phosphorylated compounds. Then, the compressed leaf pieces were carefully washed with a medium devoid of Pi. Addition of choline (50 FM) in the circulating medium led to a marked increase in the resonance at 3.3 ppm (Fig. 1, peak b), which was essentially attributable to phosphorylcholine as confirmed by perchloric acid extracts (see Fig. 2). The NMR titration curve of phosphorylcholine indicated that the posi- tion of the phosphorylcholine peak (Fig. 8) corresponded to phosphorylcholine above pH 7.0 (not shown). This result suggests that phosphorylcholine was derived from exoge- nously added choline and accumulated in the cytoplasmic compartment, pH 7.5, and not in the vacuole, pH 5.5. Identical results were obtained using nongreen material (sycamore cells) (29). A careful analysis of Figs. 8 and 9 indicates that after 2 h of choline perfusion, the cytoplasmic Pi concentra- tion decreased from 8 to 5 mM. No further decrease was observed even when high concentrations of choline were sup- plied (not shown). Although the total amount of cytoplasmic Pi declined noticeably, the NTP level remained almost con- stant (Figs. 8 and 9) (see, however, Ref. 8). In spinach leaves, when choline was used as a Pi-sequestering agent instead of mannose (8), only a part of the cytoplasmic Pi pool was used for choline phosphorylation because a large part of it remained sequestered in the chloroplast compartment devoid of choline kinase activity. The phosphorylcholine that appeared during the first 2 h after the addition of choline in the circulating system corresponded approximately to the total amount of cytoplasmic Pi, vacuolar Pi, and glucose-6-P which disap- peared within the same period of time (Fig. 9). When a threshold of cytoplasmic Pi was attained, the phosphorylation of choline was sustained almost exclusively by the continuous release of Pi from the vacuole (Fig. 9). In this particular experiment, when almost all of the vacuolar Pi had been utilized, the final cytoplasmic phosphorylcholine concentra- tion attained was considerable, exceeding 40 mM (about six times the concentration of glucose-6-P, the most abundant phosphate ester measured in uiuo by 31P NMR). We have also observed that phosphorylcholine remained in the cytoplasmic fraction and did not significantly leak out of the leaf pieces (not shown). On the other hand, when 100 pM Pi was present
P-Chol
0 5 10 Time, hours
FIG. 9. Time course evolution of the most abundant ‘lP NMR-detectable phosphorus compounds in spinach leaves perfused with a medium containing 50 I.~M choline. The concen- trations of mobile phosphorus compounds in the leaf sample were determined as in Fig. 7. Note that after the addition of choline there is a small lag phase followed by a steady decrease of the vacuolar Pi ( Vuc-P,) to sustain phosphorylcholine (P-chol) synthesis in the cy- toplasmic (Cyt) compartment.
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

31P NMR Studies of Spinach Leaves and Their Chloroplasts 1325
in the perfusion medium at the beginning of the experiment, the addition of 50 pM choline led to the same initial velocity of phosphorylcholine synthesis which was observed in the absence of Pi (not shown). However, under these conditions, the cytoplasmic Pi concentration was maintained constant while the concentration of vacuolar Pi increased steadily (see Figs. 6 and 7). These results together demonstrate that exter- nally added Pi was preferentially used to sustain Pi consump- tion in the cytosolic compartment. These results also dem- onstrate that inside a green cell, the level of phosphorylated intermediates in the cytoplasm can depend upon the amount of Pi accumulated in the vacuole.
DISCUSSION
Our results emphasize that special precautions are required for monitoring by 31P NMR the behavior of leaf tissues over a long period of time under different conditions. In mature spinach leaves, the lacuna between upper and lower epidermis represents a large proportion of the total leaf volume. More than 80% of the volume of a mature photosynthetic cell consists of a vacuole. Furthermore, the NMR resonances for the cytoplasmic metabolites are weak. Consequently, in order to increase both the homogeneity of the magnetic field and the signal-to-noise ratio for cytoplasmic metabolites in the NMR spectra, it is absolutely necessary to eliminate gas bubbles trapped in the intercellular space by vacuum infiltra- tion as shown previously by Waterton et al. (6) and to com- press small leaf pieces between two circular porous plates (13). Under these conditions, leaf pieces can survive at least 4 days as long as a well aerated nutrient medium is pumped through the system under slight pressure. Such a system enabled high quality 31P NMR spectra of leaf tissue to be continuously recorded in which all the resonances observed (cytoplasmic and vacuolar Pi, nucleotides) were sharp and well resolved. However, direct measurement of changes be- tween light and dark were hampered by obvious technical difficulties, and future work should focus on this problem.
31P NMR spectrum of spinach leaf pieces under dark con- ditions was almost identical to those of nongreen plant tissues or cells (for review, see Refs. 5 and 19). The same resonances were observed with varying intensities. For example, all plant materials examined so far contain high concentrations of UDP-glucose. As pointed out by Bieleski (30), UDP-glucose is frequently underestimated in plant tissues because it hy- drolyzes rapidly under alkaline conditions. The presence of large amounts of UDP-glucose in plant cells is probably related to the initial breakdown of sucrose catalyzed by su- crose synthase (EC 2.4.1.13) to give UDP-glucose (31). In addition, the pH values of the vacuolar and cytoplasmic compartments of different plant materials are very similar. The pH is approximately 7.5 for the cytoplasmic compartment and 5.5 for the vacuole.
Although 31P NMR cannot discriminate among all the Pi pools present in the cytoplasmic compartment, the results presented in this paper strongly suggest that oxygenated leaf tissues maintained in the dark contain a low cytosolic Pi concentration and that the bulk of the cytoplasmic Pi is sequestered in the chloroplasts. We appreciate that a portion of cytoplasmic P, is sequestered in the mitochondria but suggest that the small volume of the matrix (6) means that this proportion is relatively small. The arguments that favor a low cytosolic Pi concentration are numerous. First, the Pi concentration found in isolated chloroplasts (12 mM) is higher than that measured in the cytoplasmic compartment (6-9 mM) using ‘lP NMR. Given that the chloroplast comprises more than 60% of the cytoplasmic volume, such a result
strongly suggests that the cytosolic Pi concentration is much lower than that of chloroplast stroma. Second, one of the most surprising findings in our studies was that the total vacuolar Pi pool could fluctuate considerably, whereas the cytoplasmic pool remained almost constant (Figs. 6-9) (see also Refs. 25 and 27). It is difficult to understand how Pi movement is regulated between the vacuole and the cytosol if there is a continuous high cytosolic Pi concentration. Again, these observations are best explained if one assumes that the cytosolic Pi concentration is low. Under these conditions, any small 31P NMR changes in the concentration of free cytosolic Pi would directly or indirectly affect Pi efflux and influx from or into the vacuole. Third, it has been shown clearly that external Pi strongly influences the rate of photosynthesis by isolated chloroplasts (2). If Pi concentrations are above 1 mM
in the medium, photosynthesis is inhibited because triose phosphate molecules are withdrawn from the stroma too rapidly. This classical observation strongly suggests that in a leaf, the cytosolic Pi concentration, a key factor determining the cellular partitioning of carbon between sucrose and starch synthesis (32), is low. In addition, a high cytosolic Pi concen- tration would be utterly inappropriate to the precise regula- tion of sucrose functioning pathway in the cytosolic fraction (32-34). Fourth, in nongreen cells or tissues such as sycamore cells (13) or maize root tips (35), calibration of the peak intensity of the cytoplasmic Pi resonance with known amounts of external Pi gave estimates of cytoplasmic Pi levels of 0.6-1.2 mM including P, present in the cytosol and various cell organelles (mitochondria and amyloplastids). Thus, when organelles comprised a small part of the cytoplasmic volume, the cytoplasmic Pi concentration measured using 31P NMR was low. This is another indirect proof that in leaf tissues the cytosolic Pi concentration is much lower than the values that have been suggested previously (28,36; see, however, Ref. 8).
The results presented in this paper indicated that owing to the utilization of choline, a new cytoplasmic Pi-sequestering reagent (29), vacuolar Pi efflux can occur in response to the depletion of Pi from the cytoplasmic compartment. Exoge- nously supplied D-mannose to maize and tomato leaves can also reduce the vacuolar P, level in some circumstances (8). These observations raise, therefore, the question of the prob- lem of Pi movement between the vacuole and the cytoplasm and vice versa. Although no clear answers are available to such questions at this time, the cycling of Pi through vacuolar influx (energy dependent) and efflux (passive?) pathways is a good system that may provide (on the time scale of several min) for control over the cytosolic-free Pi and phosphorylated intermediate concentrations and therefore may be involved in the regulation of photosynthesis. For example, according to Stitt et al. (17), the change of esterified phosphate during the day may represent a decrease of the cytosolic Pi by about 10 mM. Conversely, the change of the esterified phosphate in the cytosol during the night may indicate an increase of the cytosolic Pi. Since our results suggest strongly that the con- centration of Pi in the cytosol is insufficient to sustain the accumulation of esterified phosphate observed during the day, this implies that part of the phosphorylated intermediates are synthesized at the expense of Pi deriving either from the external medium or the vacuole.
In contrast to the cytosol, the present results demonstrate that the stroma of the chloroplasts contain a high concentra- tion of free Pi. Such a high concentration of P, is required to sustain the rapid synthesis of the Benson-Calvin cycle inter- mediates, especially ribulose 1,5-bisphosphate after the onset of illumination (2). Our results strongly suggest that a one- way Pi transport, distinct from the one-for-one exchange of
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

31P NMR Studies of Spinach Leaves and Their Chloroplasts
triose phosphate and free Pi via the phosphate translocator, may be the means by which the total stromal Pi pool is normally maintained. In support of this suggestion, we have observed recently that pyrophosphate is transported through the chloroplast envelope leading to a marked increase in the stromal Pi concentration (37). Such a suggestion is attractive because the cytosolic fraction of all the plant cells examined so far contains a pyrophosphate:fructose-6-P l-phosphotrans- ferase capable of working in the reverse direction to produce PPi (38).
12. 13.
14.
The results presented in this paper also demonstrate that intrachloroplastic ATP and ADP are invisible to NMR. In- deed, upon making a perchloric extract of intact chloroplasts and treating it with CDTA to chelate metal ions, all the resonance peaks of ADP and ATP reappeared in the extract medium. Thus, these data indicated that inside the chloro- plasts the nucleotides may be bound to proteins or paramag- netic substances that render the nucleotides invisible to NMR (39). Under these conditions, the high ATP/ADP ratio meas- ured in uiuo should reflect the cytosolic status in good agree- ment with biochemical data (40, 41).
Our results demonstrate for the first time that leaf cells contain nonnegligible amounts of phytate. Its estimated cel- lular concentration was about 1 mM. The precise role of phytate in the cells remains unknown, but it may serve as a store of Pi, cations (Mg*+ and perhaps free iron), or of high energy phosphoryl groups that can be metabolized by phytase and phytate-nucleotide diphosphate transferase during cell metabolism (42). Thus, phytate may buffer the variations of free Mg2+ and Pi concentrations in the cytosol. It is also possible that phytate in the cytoplasmic compartment pre- vents the catalysis by iron of radical formation and subse- quent oxidative damage. As a matter of fact, phytate by virtue of chelating free iron could be a potent inhibitor of iron- driven hydroxyl radical (OH) formation (43, 44).
Finally, the results presented here demonstrate that intact chloroplasts contain nonnegligible amounts of sn-glycerol-3- P. The estimated concentration of sn-glycerol-3-P was about 0.5 mM. on the basis of the whole chloroplasts, an amount sufficient to sustain polar lipid synthesis (45). The presence of this metabolite in the chloroplast stroma is probably cor- related with significant levels of dihydroxyacetone phosphate reductase activity in the chloroplast (46).
15.
16.
17.
18.
19.
20.
21.
22.
23. 24. 25.
26.
27.
28.
29.
30. 31.
32.
33.
REFERENCES
1. Deuce, R. (1985) Mitochondria in Higher Plants: Structure, Func- tion and Biogenesis, Academic Press, New York
2. Walker, D. (1976) in The Intact Chloroplust (Barber, J., ed). pp. 235-278, Elsevier Scientific Publishing Co., Amsterdam
3. Roberts, J. K. M., and Jardetzky, 0. (1981) Biochim. Biophys. Acta 639,53-76
4. Ratcliffe, R. G. (1987) Methods Enzymol. 148, 683-700 5. Pfeffer, P. E., and Gerasimowicz, W. V. (1989) in Nuclear Mug-
netic Resonance in Agriculture (Pfeffer, P. E., and Gerasimow- icz, W. V., eds) pp. 3-70, CRC Press, Boca Raton, FL
6. Waterton, J. C., Bridges, I. G., and Irving, M. P. (1983) B&him. Biophys. Actu 763, 315-320
7. Loughman, B. C., Ratcliffe, R. G., and Southon, T. E. (1989) FEBS Lett. 242,279-284
34. 35.
36. 37.
38.
39.
40.
41. 42.
43.
44.
45. 8. Gunning, B. E. S., and Steer, M. V. I. (1975) Ultrastructure and
the Biology of Plant Celki, Edward Arnold, London 9. Lauer, M. J., Blevins, D. G., and Sierzputowska-Gracz, H. (1989)
Plant Physiol. 89,1331-1336 46.
10. Nakatani, H. Y., and Barber, J. (1977) Biochim. Biophys. Actu 47. 461,510-512 48.
11. Mourioux, G., and Deuce, R. (1981) Plant Physiol. 67,470-473
Yanagita,T. (1964)J. Biochem. 55,260-268 Roby, C., Martin, J.-B., Bligny, R., and Deuce, R. (1987) J. Biol.
Chem. 262,5000-5007 Navon, G., Shulman, R. G., Yamane, T., Eccleshall, T. R., Lam,
K.-B., Baronofsky, J. J., and Marmur, J. (1979) Biochemistry l&4487-4499
Evans, F. E., and Kaplan, N. 0. (1977) Proc. N&l. Acud. Sci. U. S.A. 74,4909-4913
Bass, M. B., and Fromm, M. J. (1985) Anal. &o&m. 145,192- 303
Stitt. M., Wirtz, W., Gerhardt, R., Heldt, H. W., Spencer, C. A., Walker, D., and Foyer, C. (1985) Planto (Berl.) 166, 354-364
Roberts, J. K. M., Ray, P. M., Wade-Jardetzky, N., and Jardetzky, 0. (1980) Nature 283,870-872
Bligny, R., Roby, C., and Deuce, R. (1989) in Nuclear Magnetic Resonance in Agriculture (Pfeffer, P. E., and Gerasimowicz, W. V., eds) pp. 71-89, CRC Press, Boca Raton, FL
Klingenberg, M. (1974) in Methods of Enzymatic Analysis (Berg- meyer, H. U., ed) Vol. 4, pp. 2045-2059, Academic Press, New York
Hooks, M. A., Clark, R. A., Nieman, R. H., and Roberts, J. K. M. (1989) Plant Physiol. 89,963-969
Roberts, J. K. M., Callis, J., Wemmer, D., Walbot, V., and Jardetzky, 0. (1984) Proc. N&Z. Acud. Sci. U. S. A. 81, 3379- 3382
Deuce, R., and Joyard, J. (1979) Adu. Bot. Res. 7, l-116 Oaawa. S.. and Lee. T.-M. (1982) Biochemistrv 21, 4467-4473 RibeilIe, F., Blignyj R., Martin, J. B., and Do&e, R. (1983) Arch.
Biochem. BioDhvs. 225.143-148 Mathieu, Y., cue&, J., Kurkdjian, A., Manigault, P., Manigault,
J.. Zielinska. T.. Gillet. B.. Beloeil. J. C.. and Lallemand. J. Y. (1989) Pl;ntPiysioZ.~89, i9-26 ’ ’
Brodelius, P., and Vogel, H. J. (1985) J. Biol. Chem. 260, 3556- 3560
Foyer, C., Walker, D., Spencer, C., and Mann, B. (1982) Biochem. J. 202,429-434
Bligny, R., Foray, M. F., Roby, C., and Deuce, R. (1989) J. Biol. Chem. 264,4888-4895
Bieleski, R. L. (1973) Anna Rev. Plunt Physiot. 24,225-252 ap Rees, T. (1987) in The Biochemistry of Plants, Vol. 12, Physi-
ology of Metabolism (Davies, D. D., ed) pp. 87-115, Academic Press, New York
Heldt, H. W., and Fliigge, U. I. (1987) in The Biochemistry of Plants, Vol. 12, Physiology of Metabolism (Davies, D. D., ed) on. 49-85. Academic Press. New York
Czgke, C., Balogh, A., Wong, J. H., Buchanan, B. B., Stitt, M., Herzoa. B.. and Heldt. H. W. (1984) Trends Biochem. Sci. 9. 533-5% '
Huber, S. C. (1986) Annu. Reu. Plant Physiol. 37, 233-246 Pfeffer, P. E., Tu, S. I., Gerasimowicz, W. V., and Cavanaugh, J.
R. (1986) Plant Physiol. 80, 77-84 Foyer, C., and Spencer, C. (1986) Pluntu (Berl.) 167,369-375 Dumas, R., Joyard, J., and Deuce, R. (1989) Biochem. J. 259,
769-774 Carnal, N. W., and Black, C. C., Jr. (1983) Plant Physiol. 71,
150-155 Stubbs, M., Freeman, D., and Ross, B. D. (1984) Biochem. J.
224,241-246 Stitt, M., Lilley, R. M., and Heldt, H. W. (1982) Plant Physiol.
70,971-977. Gardestrom, P., and Wigge, B. (1988) Plant Physiol. 88, 69-76 Biswas, B. B., Ghosh, B., Majumder, A. L. (1984) Subcell.
Biochem. 10,237-280 Loewus. F. A.. and Loewus. M. W. (1983) Anna Reu. Plant
Physibl. 34, i37-161 ' Graf, E., Empson, K. L., and Eaton, J. W. (1987) J. Biol. Chem.
262,11647-11650 Jovard. J.. and Deuce. R. (1987) in The Biochemistrv of Plants,
Vol. 9, Lipids: Structure and l&n&ion (Stumpf, P.-K.: ed) pp: 215-274, Academic Press, New York
Gee, R. W., Byerrum, R. U., Gerber, D. W., and Tolbert, N. E. (1988) Plant Phvsiol. 86.98-103
Delieu, T., and Walker, D..A. (1972) New Phytol. 71, 201-225 Blechschmidt-Schneider. S.. Ferrar. P., and Osmond, C. B. (1989)
Pluntu (Berl.) 177,515~$25
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from

R Bligny, P Gardestrom, C Roby and R Douce31P NMR studies of spinach leaves and their chloroplasts.
1990, 265:1319-1326.J. Biol. Chem.
http://www.jbc.org/content/265/3/1319Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/3/1319.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 5, 2020http://w
ww
.jbc.org/D
ownloaded from