2002-ag-1672 (m. asif iqbal) ph.d. thesis...
TRANSCRIPT

Response of maize (Zea mays) to auxin producing plant growth promoting rhizobacteria under saline conditions
By
Muhammad Asif Iqbal
M.Sc. (Hons.) Agriculture 2002-ag-1672
A thesis submitted in partial fulfillment of the requirements for the
Degree of
DOCTOR OF PHILOSOPHY In
Soil Science
Institute of Soil & Environmental Sciences FACULTY OF AGRICULTURE
UNIVERSITY OF AGRICULTURE, FAISALABAD
2014

To,
The Controller of Examinations, University of Agriculture, Faisalabad.
We, Supervisory Committee certify that the contents and form of thesis
submitted by Muhammad Asif Iqbal, Regd. No. 2002-ag-1672 have been found
satisfactory and recommend that it be processed for evaluation by the External
Examiner (s) for the award of degree.
SUPERVISORY COMMITTEE
CHAIRMAN (Dr. Muhammad Khalid)
MEMBER (Dr. Zahir Ahmad Zahir)
MEMBER (Dr. Rashid Ahmad)

DEDICATED
TO
My graceful and polite father
My most loving mother
My brothers and sister
Who live in my mind and soul
Whose love is more precious
Than pearls and diamonds
Who are those whom I say my own
Whose love will never change
Whose prayers will never die
&
Who are nearest, dearest and deepest

ACKNOWLEDGEMENTS
All praise to ALMIGHTY ALLAH alone, the most merciful and the most compassionate and his Holy Prophet ‘MUHAMMAD’ (Peace be upon him) the most perfect and exalted among and even of ever born on the surface of earth, who is, forever a torch of guidance and knowledge for the humanity as a whole.
The work presented in this manuscript was accomplished under the inspiring guidance, gorgeous assistance, constructive criticism and enlightened supervision of Dr. Muhammad Khalid, Professor, Institute of Soil & Environmental sciences, University of Agriculture, Faisalabad. His efforts toward the inculcation of the spirit of constant work and the maintenance of professional integrity besides other invaluable words of advice will always serve as beacon of light throughout the course of my life. I take this humblest opportunity to express my deepest sense of gratitude and thankfulness to him.
It is matter of great pleasure and honor for me to express my gratitude and appreciation to the members of my Supervisory Committee, Dr. Zahir Ahmad Zahir, Professor, Institute of Soil & Environmental sciences, University of Agriculture, Faisalabad and Dr. Rashid Ahmad, Department of Crop Physiology, University of Agriculture, Faisalabad, under whose kind and scholastic guidance, keen interest and constant encouragement, I completed my research. I am thankful to all my School teachers especially, Muhammad Mumtaz who helped and guided me during the entire course of my studies.
A deep sense of appreciation and special thanks to Dr. M. Usman, Dr. S.M. Shahzad, Dr. Naveed Iqbal and Dr. Shahid Hussain, without whom generous guidance, encouragement and immense help, I would have not been able to complete my work. Friends are the comrades of the battle, the battle to generate knowledge, sift myths and facts and to remove ambiguity. They were co–sharer of my struggle and my work. I express my thankful feelings for my friends, Dr. Ahmad Nawaz, Dr. Sana Ullah, Dr. Jawad ul Hassan, Habib Ali, Kashif Anjum, Muhammad Aon, M. Hamid Rasool, Mujahid Manzoor, Sami Ullah, Zeeshan Aslam and M. Kashif Malana.
In the end I want to convey my love and affection to the apples of my eye (Areeba, Hunzla, Shaheera, Minahil, Amna, Aisha, Uswa, Laiba, Meher, Affan, Mahad, Hassan, Abubakar, Ibrahim, Abdullah Talat, Usama and Ahmad Afroz). They were always been a source of inspiration for me. My mind always refreshed in their company and I was able to shed the burden of sorrows, distress and hopelessness. They were my cutest companions during this journey. I regard them as my deariest and precious findings. They will always remain in my heart and will regulate its beat when pressure of life will try to stop it and will serve as a source of light when shadows of time will try to blind my eyes. May Allah bless all these people with long, happy and peaceful lives (Ameen).
Muhammad Asif Iqbal

DECLARATION
I hereby declare that the contents of the thesis “Response of Maize (Zea mays) to Auxin
Producing Plant Growth Promoting Rhizobacteria under Saline Conditions” are product of
my own research and no part has been copied from any published source (except the
references, standard mathematical or genetic models/formulae/protocols, etc.). I further
declare that this work has not been submitted for award of any diploma/degree. The
university may take action if the information provided is found inaccurate at any stage. In
case of any default the scholar will be proceeded against as per HEC plagiarism policy.
Signature of the student: ___________________
Name: Muhammad Asif Iqbal
Regd. No. 2002-ag-1672

LIST OF CONTENTS
CH# TITLE Page#
1. INTRODUCTION 1
2. REVIEW OF LITERATURE 5
3. MATERIALS AND METHODS 33
4. RESULTS 48
5. DISCUSSION 133
6. SUMMARY 147
LITERATURE CITED 150

LIST OF TABLES
No. TITLE Page1. Physico-chemical characteristics of soil used for pot trial 37 2. Physico-chemical characteristics of soil used for field trial 38 3 Growth of rhizobacterial strains in broth tubes after 3 days
under salt stress conditions 49
4 Growth of rhizobacterial strains in broth tubes after 3 days under salt stress conditions
50
5 Amount of IAA equivalents (µg mL-1) produced by the selected rhizobacterial strains
52
6 Effect of rhizobacterial inoculation on total plant dry weight of maize seedlings under salt stressed axenic conditions
60
7 Effect of rhizobacterial inoculation on shoot root ratio of maize seedlings under salt stressed axenic conditions
61
8 Effect of rhizobacterial inoculation (with and without L-TRP) on shoot root ratio of maize at different salinity levels
73
9 Characterization of selected rhizobacterial strains 132

LIST OF FIGURES
No. TITLE PAGE
1 Effect of rhizobacterial inoculation on shoot length of maize seedlings under salt
stressed axenic conditions. 54
2 Effect of rhizobacterial inoculation on root length of maize seedlings under salt
stressed axenic conditions 56
3 Effect of rhizobacterial inoculation on shoot dry weight of maize seedlings under salt
stressed axenic conditions 57
4 Effect of rhizobacterial inoculation on root dry weight of maize seedlings under salt
stressed axenic conditions 59
5 Effect of rhizobacterial inoculation on plant height of maize under salt stressed pot
conditions 64
6 Effect of rhizobacterial inoculation on fresh biomass of maize under salt stressed pot
conditions 66
7 Effect of rhizobacterial inoculation on root length of maize under salt stressed pot
conditions 68
8 Effect of rhizobacterial inoculation on root weight of maize under salt stressed pot
conditions 71
9 Effect of rhizobacterial inoculation on grains yield of maize under salt stressed pot
conditions 72
10 Effect of rhizobacterial inoculation on 1000-grain weight of maize under salt stressed
pot conditions 75
11 Effect of rhizobacterial inoculation on cob biomass of maize under salt stressed pot
conditions 76
12 Effect of rhizobacterial inoculation on chlorophyll content (SPAD values) of maize
under salt stressed pot conditions 78
13 Effect of rhizobacterial inoculation on electrolyte leakage of maize under salt
stressed pot conditions 80
14 Effect of rhizobacterial inoculation on photosynthetic rate in shoot of maize under
salt stressed pot conditions 82
15 Effect of rhizobacterial inoculation on transpiration rate of maize under salt stressed
pot conditions 83

16 Effect of rhizobacterial inoculation on stomatal conductance of maize under salt
stressed pot conditions 85
17 Effect of rhizobacterial inoculation on substomatal CO2 concentation of maize under
salt stressed pot conditions 86
18 Effect of rhizobacterial inoculation on proline content of maize under salt stressed
pot conditions 88
19 Effect of rhizobacterial inoculation on total solube sugars of maize under salt
stressed pot conditions 89
20 Effect of rhizobacterial inoculation on total phenolics of maize under salt stressed
pot conditions 91
21 Effect of rhizobacterial inoculation on ascorbate peroxidase vales of maize under salt
stressed pot conditions 93
22 Effect of rhizobacterial inoculation on K+/Na+ of maize under salt stressed pot
conditions 95
23 Effect of rhizobacterial inoculation on nitrogen concentration in shoot of maize
under salt stressed pot conditions 96
24 Effect of rhizobacterial inoculation on phosphorous concentration in grains of maize
under salt stressed pot conditions 98
25 Effect of rhizobacterial inoculation on crude protein content in grains of maize
under salt stressed pot conditions 100
26 Effect of soil salinity on grain yield per plant of maize in pots 101
27 Effect of rhozobacterial inoculation on grain yield per plant of maize
in pots
101
28 Effect of soil salinity on total chlorophyll contents (SPAD values) of
maize in pots
102
29 Effect of rhozobacterial inoculation on total chlorophyll contents
(SPAD values) of maize in pots
102
30 Effect of soil salinity on proline contents of maize in pots 103
31 Effect of rhozobacterial inoculation on proline contents of maize in
pots
103
32 Effect of soil salinity on electrolyte leakage of maize in pots 104

33 Effect rhizobacterial inoculation on electrolyte leakage of maize in
pots
104
34 Effect of rhizobacterial inoculation on plant height of maize under salt stressed field
conditions 106
35 Effect of rhizobacterial inoculation on grain yield of maize under salt stressed field
conditions 107
36 Effect of rhizobacterial inoculation on fresh biomass of maize under salt stressed
field conditions 109
37 Effect of rhizobacterial inoculation 1000-grain weight of maize under salt stressed
field conditions 111
38 Effect of rhizobacterial inoculation on straw yield of maize under salt stressed field
conditions 112
39 Effect of rhizobacterial inoculation on cob length of maize under salt stressed field
conditions 114
40 Effect of rhizobacterial inoculation on cob weight of maize under salt stressed field
conditions 115
41 Effect of rhizobacterial inoculation chlorophyll contnt (SPAD values) of maize under
salt stressed field conditions 117
42 Effect of rhizobacterial inoculation on proline content of maize under salt stressed
field conditions 118
43 Effect of rhizobacterial inoculation on crude protein contents of maize under salt
stressed field conditions 120
44 Effect of rhizobacterial inoculation on phosphorous in grain of maize under salt
stressed field conditions 122
45 Effect of rhizobacterial inoculation on K+/Na+ of maize under salt stressed field
conditions 123
46 Effect of rhizobacterial inoculation on phenol content of maize under salt stressed
field conditions 125
47 Effect of rhizobacterial inoculation on total soluble sugar of maize under salt
stressed field conditions 126
48 Effect of rhizobacterial inoculation on electrolyte leakage of maize under salt
stressed field conditions 128
49 Effect of rhizobacterial inoculation on SOD values of maize under salt stressed field
conditions 130

S1 Exprimental layout of field trials 39
S2 Pictorial representation of rhizobacterial inoculation at different salinity levels
under axenic conditions 62
S3 Pictorial representation of rhizobacterial inoculation on root growth at different
salinity levels in pot experiment. 69

ABSTRACT
Soil salinity is one of the most widespread agricultural problems which reduce the field and
crop productivity. Salinity disturbs the hormonal balance in plants which results in poor
growth. Use of plant growth promoting rhizobacteria (PGPR) is considered an economical
and environment-friendly approach to combat salinity stress. This study was carried out to
investigate the effect of PGPR on the growth, antioxidant status, physiological parameters
and mineral content of maize (Zea mays) in salt affected soils. Bacterial strains were isolated
from the rhizosphere of maize growing under salt affected soil conditions. These strains were
screened on the basis of auxin production and their ability to withstand salinity stress.
Nineteen rhizobacterial strains producing auxin were further screened for their growth
promoting activity under axenic conditions at 0, 4, 8, 12 and 15 dS m-1 salinity levels. Two
strains were selected after this study and their ability of growth promotion in the presence and
absence of L-TRP was tested alone and in combination under salt affected pot and field
conditions. Results showed that soil salinity reduced the plant growth, physiological
parameters, mineral nutrient uptake and yield of maize while antioxidant activity and proline
concentration was increased. Inoculation with PGPR strains under saline soils alleviated the
salinity effects on the antioxidant enzymes (APX and SOD), along with those on
photosynthesis, mineral content and growth in the absence and presence of L-TRP
application. Dual inoculation showed better results than sole inoculation. The results of the
present study highlight the significance of PGPR strains to alleviate the adverse effects of
salinity stress.

1
CHAPTER-1
INTRODUCTION
Maize (Zea mays L.) is a major food crop after wheat and rice in Pakistan. It is a
multiuse crop as it provides raw material to industry and feed to livestock and poultry. It
contributes more than 10% to agricultural production and 15% to agricultural employment in
the country (Khaliq et al., 2004). Maize is one of the most significant cereal crops both for
human food and animal feed and is grown under varying conditions throughout the world.
Being an important cereal crop, maize is grown on about one million hectares with a total
production of about 3.13 million tons and an average yield of 3.3 ton ha-1 (Anonymous,
2008). Due to its important role in food and feed of humans and animals it is vital to cause
improvements in agronomic characteristics to get good and high quality produce under salt
affected soil conditions.
Around the world, crop farming has to deal with varying biotic and climatic stresses
which limit the crop growth and productivity (Nemati et al., 2011). Among such hazards is
soil salinization that reduces the productivity of fertile lands (Lawlor, 2002). There is
progressive increase in barren lands due to accumulation of salts (Azooz, 2004) and more
than half of the cultivated land is expected to be affected by soil salinization till 2050
(Vinocur and Altman, 2005). Problems associated with soil salinity not only affect agriculture
but also disturb the biodiversity of the environment (Parida et al., 2003; Fernandez-Aunión et
al., 2010). Due to low rain fall and dry climatic conditions the problem of soil salinity is more
vigorous in arid and semi-arid environments. Presence of higher levels of sodium and
chloride cause negative effects on plants growth. Soil salinity causes two types of stresses
namely osmotic stress and ion toxicity in plant cells, thus upsetting the growth and
metabolism. Plants suffer various physiological changes due to soil salinity which include
changes in nutrient and water uptake (Munns, 2005; Parida and Das, 2005), reduction in
photosynthetic activity (Chavez et al., 2011), production of reactive oxygen species
(Dionisio-Sese and Tobita, 1998; Sgherri et al., 2007; Pérez-López et al., 2009, 2010) and
modification nitrogen uptake and metabolism (Munns, 2005). All these changes ultimately
affect the plant growth and performance (Gouia et al., 1994). The osmotic effects brought by
soil salinity can at first stage cause the disintegration of root membranes (Carvajal et al.,

2
1999) and lower the transpiration rate and photosynthetic activity (Parida and Das, 2005).
Along with the induction of the osmotic stress, ionic toxicity is caused by the salinity stress.
These toxic ions negatively affect the NO3- transporters, nitrogen reductase (NR) activity and
glutamine synthase (GS) activity (Flores et al., 2000).
To surmount this problem, numerous approaches of defense against salt stress have
been adopted. Some of these strategies include the development of salt tolerant plants through
conventional and genetic engineering tools, reclamation and amelioration of salt affected soils
(Vinocur and Altman, 2005; Valliyodan and Nguyen, 2006; Ashraf and Akram, 2009; Mittler
and Blumwald, 2010). During current years, a new biological approach has been developed to
shelter plants from salt stress in soil, is the use of plant growth-promoting rhizobacteria
(PGPR) (Yue, 2007). Such beneficial microorganisms have been renowned in the
maintenance of healthy root, facilitation of nutrient uptake and helping the plants to withstand
environmental stresses and increase plant growth (Reed and Glick, 2004). Positive impacts of
these PGPR include the production of phytohormones; auxin, cytokinins and gibberellins
(Garcia de Salamone et al., 2001), enhancing release of the nutrients (Nautiyal et al., 2000) as
well as preventing the deleterious effects of environmental stresses (Kloepper and Schroth,
1978). Against using higher doses of pesticides and fertilizers the use of PGPR is an
economic and environmental friendly approach that promises the improvement in crop
growth and productivity on sustained basis. Different inter cellular and intra cellular
microorganism colonize the plant roots (Gray and Smith, 2005). Beneficial bacteria and fungi
present in the rhizosphere have the potential to directly and indirectly enhance the plant
growth under stress conditions (Dimkpa et al., 2009). The direct mechanisms of plant growth
stimulation by these microorganisms include the production of phytohormones, biological
nitrogen fixation, production of siderophores and phosphate solubilization (Hayat et al., 2010;
Rodriguez and Fraga, 1999). By an indirect way, these bacteria protect the plants against
diseases through antibiotics production (Lugtenberg and Kamolova, 2009).
Plants commonly adapt to various types of biotic and abiotic stress by causing
changes in their root morphology (Potters et al., 2007). The plant hormones play a vital role
in altering the root morphology (Spaepen and Vanderleyden, 2011; Spaepen et al., 2007).
Hayat et al. (2010) reported that most of the plant growth-promoting bacteria possess the
ability to produce indole acetic acid (IAA). Different plants inoculated with such bacteria

3
showed an increase in root growth, lateral root formation, root hairs, which helped them to
withstand abiotic stress. Activity of ACC-deaminase enzyme that degrades the ACC, the
precursor of ethylene, is stimulated by the IAA produced by the bacteria (Glick, 2005). Under
salt affected soil conditions, ACC-deaminase activity of bacteria can be helping for better
growth and yield of crops. Positive effects of such phytohormone producing PGPR have been
documented in different crops such as wheat (Khalid et al., 2004; Egamberdieva, 2009) and
rice (Mirza et al., 2006).
Plant hormones play a vital role in regulating the protective response against different
abiotic and biotic stresses (Ansari et al., 2012). These phytohormones are involved in the
expression of various genes via synergistic and/or antagonistic actions (Reed and Glick,
2004). This phenomenon is known as signaling cross talk (Kraft et al., 2007). Plants can slot
in core developmental programs and swiftly adjust growth to mutable situations by
persuading hormone homeostasis (Wolters and Jurgens, 2009). Indole acetic acid is known to
play a vital role in growth and developmental aspects of plants such as development of
vascular tissues, lateral root formation, gravitropism, cell differentiation apical dominance as
well as directive of gene expression (Malamy and Ryan, 2001; Wang et al., 2001). Current
proofs point out that auxin also responds to salinity in crop plants and its signaling is down-
regulated under different stresses (Kazan and Manners, 2009). When plants faces various
environments stresses, the internal levels of phytohormones are decreased which are mostly
unsatisfactory for prime growth (De Salamone et al., 2005). The negative effect of salinity on
the germination of seed may be due to drop in the internal levels of phytohormones
(Zholkevich and Pustovoytova, 1993; Debez et al., 2001). Naqvi and Ansari (1974) reported
that transport of cytokinin from the root to shoot was decreased due to salinity stress.
Similarly, Itai et al. (1968) reported that from the maize coleoptiles tip the recovery of
diffusible auxins was decreased. Therefore, under soil salinity reduction in growth of plants
could be the result of changes in hormonal balance. The negative effects of salinity were
alleviated by the exogenous application of such plant growth regulators (Dhingra and
Varghese, 1985; Khan and Weber 1986; Prakash and Parthapasenan, 1990; Gul et al., 2000;
Saimbhi, 1993; Gul et al., 2000; Khan et al., 2004; Afzal et al., 2005). A significant
reduction in IAA contents was noted in rice plants due to soil salinity by Prakash and
Prathapasenan (1990). They reported application of GA3 partly mitigate the effect of salinity

4
on reducing IAA levels. This shows that salinity may influence hormone balances by
affecting plant growth and development. Nilsen and Orcutt (1996) also reported marked
reduction in IAA concentration of in five days old rice after salinity application. In tomato the
level of IAA was also decreased due to salinity (Dunlap and Binzel, 1996). Sakhabutdinova et
al. (2003) documented that soil salinization causes a progressive reduction in IAA contents of
plant roots. The growth inhibiting effect of salt stress have been found to be alleviated by the
priming of wheat seed with various plant growth regulators by other researchers (Sastry and
Shekhawa, 2001; Afzal et al., 2005). Increasing soil salinity caused a reduction in seed
germination while application of IAA and/or NAA proved helpful in lowering the adverse
effects of soil salinity on germination (Balki and Padole, 1982; Gulnaz et al., 1999).
Keeping in view the above mentioned discussion it is hypothesized that auxin (IAA)
producing PGPR may facilitate maize growth in saline conditions. This study was planned
with the following objectives:
o Isolation of PGPR from rhizosphere of maize growing under salt affected conditions
o Determination of in-vitro biosynthesis of auxin by these microbes
o Screening of isolates for their growth promoting ability for maize under salt affected
axenic conditions
o Evaluation of PGPR potential to promote growth and yield of maize in pot and field
under saline conditions
o Characterization and identification of the selected strains

5
CHAPTER-2
REVIEW OF LITERATURE
Soil salinity is worldwide agricultural problem. Most of the water present on earth
contains about 3% sodium chloride salt. Such high concentration of salts has resulted in vast
range of salt affected soils. Approximately 900 Mha of the land is affected by this menace.
Soil salinity is considered a major threat to agricultural productivity and food security
(Flowers and Yeo, 1995; Munns, 2002).
To get good quality and high produce of crop, people have been using different
selection and breeding techniques since ages. Different crops are selected with special
reference to environment and soil conditions. Shift in cultivation of crops has been noted in
the Tigris-Euphrates basin of ancient Mesopotamia (from wheat to barley, a relatively salt
tolerant crop) as those soils gradually turned into salt affected soils (Jacobsen and Adams,
1958). This problem will cause more serious threats to food security as the world population
increases especially in developing countries like Pakistan.
2.1 Problem of salinity in Pakistan and at world level
Naturally soil salinization is caused due to the upward movement of groundwater
which is then evaporated on the surface of soils leaving behind the salts which cause soil
salinity. Alongside natural causes of soil salinity, human activities also add to this problem.
For example, irrigation practices in arid and semi-arid regions of the world are major source
of soil salinization (Supper, 2003). These practices elevate the water level and ultimately the
evaporation. All over the world, approximately more than 800 million hectares of soil is
affected by this problem (FAO, 2005). This accounts for almost 6% of the total land area.
Major source of this soil salinity and/or sodication is natural, but a reasonable part of land is
salt affected due to irrigation practices and clearing of land.
Out of 1500 million ha land that is cultivated under dry land agriculture, 2% (32
million ha) is affected by soil salinity to different levels. Forty-five million ha land of
irrigated agriculture are also subjected to this menace (FAO, 2005). Soil salinity either caused
naturally or by human activities reduces the 1/3rd yield of world’s arable lands. High amounts

6
of salts in soils, taking into account both human made and naturally occurring salinization,
are responsible for yield reduction on one third of the global arable land.
Approximately 6.30 Mha of Pakistani land are under soil salinity problem (Alam et
al., 2000). Irrigation in Pakistan is mainly done through a vast network of canals which
extends up to over 62,400 km. Although 15% of the cultivated land of Pakistan receives
irrigation, yet the production from this area is more than twice of that of rain fed land. These
lands contribute to the world’s food production. These lands face a serious issue of soil
salinity which reduces their production capacity (Rahman, 1998). It is estimated that the
population of the world is going to increase 1.5 billion people in next two decades. With
increase in population and urbanization, the issue of food productivity and maintenance
becomes a serious threat to agriculture (Owen, 2001). There is need to find ways to ensure
food security on sustained basis but not at the cost of new land area which would ultimately
cause deforestation and changes in biodiversity. It is estimated that developed and developing
countries will need to boost their production up to 20 and 60%, respectively (Gruhn et al.,
2000; Cakmak, 2002). Under these situations, the problem of soil salinity becomes more
important.
2.2 Impact of salinity stress on plant growth and development
Plants face four types of stresses due to soil salinity which adversely affects the plant
growth and development (Greenway and Munns, 1980) i.e.
1. Osmotic stress (limited water supply)
2. Ionic stress Ion toxicity (Ion toxicity)
3. Impairment of nutritional balance (caused by elevated concentration of toxic ions such as
Na+and Cl-)
4. Damage caused by reactive oxygen species (ROS)

7
2.2.1 Osmotic stress (Limited water supply)
At first instance, uptake of water is reduced due to soil salinity. This situation
ultimately leads to reduced growth. This effect of salinity stress is referred to as osmotic
stress. The effects of drought and soil salinity are similar regarding osmotic stress. The
production of new plant leaves primarily depends upon the soil water potential. In the
growing parts of plants, salts are not accumulated to such a high concentration that can inhibit
the growth, but they are continuously translocated into the expending vacuoles. So the growth
of newly formed leaves is not directly affected by the accumulation of salts (Munns, 2005).
Decrease in root/shoot growth is mainly caused by the water shortage caused by salinity
stress (Munns, 2002). This hypothesis is supported by the fact that in the growing cells, the
Na+ and Cl- concentrations are below toxic levels. Hu and co-workers (Hu et al., 2005)
reported that Na+ concentration in growing cells (of wheat sown in 120 mM sodium chloride)
was 20 mM at maximum and 10 mM in fastly expanding tissues. The concentration of Cl-
was recorded 50 mM only. Similar findings regarding concentration of toxic ions have been
documented by other researchers in maize (Neves-Piestun and Bernstein, 2005) and barley
(Fricke, 2004). It was also reported (Munns et al., 2000) that osmotic stress caused by the soil
salinity outside the roots causes change in root/shoot water relations which induce hormonal
signals that control the elongation of cells. Severity of salinity mainly directs the growth
inhibition rate. At low levels of osmotic stress, growth of leaves and stem is inhibited while
roots keep on growing and elongating (Hsiao and Xu, 2000). Extent of growth retardation
caused by osmotic stress depends upon the particular plant part, response time and plant
species under consideration as well as nature of salt stress treatment (Ashraf, 1994).
2.2.2 Ionic Stress
When certain ions are taken up from the soil or water and accumulated in the plants
itself, they cause toxicity. This ion toxicity may be caused even if the severity of salinity is
less. Na+, Cl- and SO42- are mainly considered as the toxic ions from salinity point of view
which are responsible for the low crop yield. Crops vary in their sensitivity to soil salinity e.g.
most of the woody perennial plants and crops are sensitive (Abrol et al., 1988). Taken up salts
are accumulated in older leaves and they are continuously transported to the transpiring
leaves where the concentration of Na+ and Cl- rises to a toxic level and ultimately cause the

8
death of leaves. This injury is mainly caused because these ions are accumulated to such a
high level that the cells are unable to translocate them into vacuoles. This buildup of salts in
the cytoplasm hinders the activity of enzymes. On the other hand, these ions can also be
deposited in the cell walls and deprives them of water (Munns, 2005) but no such effect was
observed by Muhling and Lauchli (2002) in two maize cultivars having different salt
tolerance abilities. Tester and Davenport (2003) reported that concentration of Na+ in root
cytosole was 10-30 m M. Cytosolic concentration of leaves are not clear but are thought to be
less than 100 mM (Wyn Jones and Gorham, 2002). The toxic levels of Cl- are not well
understood. Salts must be excluded from the roots into the soil solution otherwise they will
accumulate to toxic levels. Amount of water that is transpired is about 50 times higher than
that retained in leaves of plants (Munns, 2005). Another study (Hussain et al., 2003)
compared the two wheat verities to exclude Na+ and prevent from leaf chlorosis and
increasing their yield. They reported on the basis of their results that in high sodium-variety,
older leaves showed injury symptoms and died earlier than low-sodium genotypes. This
genotype also showed an increase in yield of 20% at moderate salinity levels. They concluded
that Na+ exclusion is a beneficial trait of genotypes to withstand salinity stress. Accumulation
of Na+ ions negatively affects the seed germination. This is mainly due to that it disturbs the
water content of plants. It may also displace Ca2+ and take its place in vital cell wall binding
sites, which ultimately affect the cell wall formation and growth of plants (Xue et al., 2004).
Similarly Cl- concentration higher than 80 mM also cause negative effect on morphology of
plants, function of stomata is disturbed and leaves become thinner (Loreto and Bongi, 1987).
Cl- is highly mobile in water and taken up by the plants and ultimately transported to leaves
when Cl- is built up to a toxic level in leaves, it causes burning and drying of leaves. This
situation may lead to loss of early leaves and plants become defoliated as whole Marschner
(1995).
2.2.3. Impairment of nutritional balance
Buildup of high amount of salts in the vicinity of roots results in osmotic stress. This
condition causes alteration in water relation of plants, impairment of essential nutrients
uptake and buildup of toxic ions in plants. Owing to this situation, the metabolism of plants
and activity of various enzymes is badly disturbed (Lacerda et al., 2003). Negative effect of

9
toxic ions on the uptake of essential nutrients by plants causes serious imbalance of nutrients
within plants (McCue and Hanson, 1990; Karimi et al., 2005). The activity of various
enzymes is altered due to high Na+: K+ caused by the accumulation of high levels of Na+ than
K+ (Booth and Beardall, 1991). The least uptake of K+ due to high Na+ uptake leads to
potassium deficiency symptoms in plants which are at first noted as chlorosis and after time
as necrosis of plant parts (Gopal and Dube, 2003). Potassium plays a vital role in protein
synthesis, osmoregulation, enhancing photosynthesis and maintenance of turgor pressure
(Ashraf, 2004). Calcium and K+ are essential for proper functioning and integrity of cell
membranes (Wenxue et al., 2003). Selective uptake of K+ and its compartmentalization in
cells and transportation in roots plays a main role in the maintenance of appropriate levels of
K+ in plants under salt stress conditions (Carden et al., 2003). Soil salinity tolerance of plants
is judged by their ability to take up and maintain adequate amount of Ca2+ under salinity
stress (Unno et al., 2002). Kinraide (1998) reported that under salinity stress, Ca2+/Na+ ratio
decreased which lead to membrane disintegration and selectivity. Hasegawa et al. (2000)
reported that exogenic application of Ca2+ resulted better growth of plants under salinity
stress. This might be due to improved Ca2+/Na+ ratio. Calcium is also important as it helps in
osmotic adjustment by enhancing the accumulation of compatible solutes (Girija et al., 2002).
Calcium also protects the plants from salinity stress by maintaining the integrity of
membranes and protecting the plants against oxidative damage (Larkindale and Knight,
2002). Khadri et al. (2007) found an increase in K+/Na+ ratio due to the application of ABA.
2.2.4. Reactive oxygen species
When plants are exposed to certain biotic and abiotic stresses, the activity of reactive
oxygen species (ROS) is increased. These species cause peroxidation of lipids, mutation of
DNA and degradation of proteins (Pitzschke and Hirt, 2006). These species are produced
mainly in the mitochondria and chloroplast of the plant cells (Mittler, 2002). Although these
ROS cause oxidative damage to plant cells, they also cause signal transduction during stresses
and in response to pathogenic attacks (Torres and Dangle, 2005).
Increased production of these species during salinity stress causes injury to the
membranes (Shalala et al., 2001). Decrease in CO2 uptake due to the closure of stomata
results in the production of ROS in higher amounts. This increased production of ROS causes

10
a decrease in the concentration of NADP+ (Foyer and Noctor, 2003). These ROS degrade
photosystem II and D1 proteins which ultimately result in inhibition of photosynthetic
activity. Hydrogen peroxide is also accumulated due to NADPH activity and photorespiration
caused by stress and may decrease the activity of enzymes. H2O2 in the presence of metal
ions is changed into highly active (.OH) which has the ability to degrade the biomolecules
(Halliwell and Gutteridge, 1989).
2.3 Measures to enhance salt tolerance
Crop’s salt tolerance can be enhanced by causing variations in present crops. Notable
enhancements in salinity tolerance have been achieved through conventional selection and
breeding approaches (Ashraf, 2002), as well as through agronomic practices. Such agronomic
strategies include the use of exogenic use of phytohormones and inoculation of plant seeds
with plant growth promoting rhizobacterial.
2.3.1 Conventional crop breeding
Breeding of crops by using conventional breeding techniques provides an economic
and efficient way of inducing stress tolerance in crops than soil amelioration methods.
Response of plants to salt stress is influenced by the interaction of various environmental
factors and salt stress. That’s why it is of great importance to develop such genotypes that
have the ability to cope with adverse conditions of soil and climate (Ashraf and McNeilly,
2004; Sairam and Tyagi, 2004). Such crop genotypes have been established and are
commercially available for cultivation. Such varieties of wheat and rice have been developed
which are tolerant to salinity stress (Ashraf and McNeilly, 1990). A limited success is
achieved in developing such salt tolerant crops. This is due to the fact that these cultivars
have only been tested under ideal conditions by the breeders, and if these genotypes are not
competitive then there is no benefit in developing such tolerant crops (Yamaguchi and
Blumwald, 2005). Although many high yielding cultivars have been developed by using this
approach but only a few cultivars are known that are tolerant to salinity (Ashraf, 1994). The
main drawback of this approach is that it requires a long period of time and undesirable genes
are also transferred alongside the desired one. The transfer of desired alleles from

11
interspecific and inter generic sources is limited due to reproductive barriers (Apse and
Blumwald, 2000).
2.3.2 Genetic Engineering
Methods of conventional breeding are useful for inducing stress tolerance in different
crops but due to complex trait and environmental impacts, this technique is not very much
successful and effective. As compared with conventional breeding techniques, genetic
engineering is faster and attractive tool for imposing salinity stress tolerance. The induced
genes are directly or indirectly protected against the stresses (Bohnert and Sheveleva, 1998).
In transgenic plants, a few genes have been tried to test stress tolerance (Winicov,
1998). Different genes that encode antioxidant enzymes have been cloned and found in
transgenic plants (Allen, 1995). Torsethaugen et al. (1997) also reported enhanced oxidative
stress tolerance in plants of tobacco that overexpressed cytosolic ascorbate peroxidase. Zhang
et al. (2001) incorporated AtNHXl gene that encodes for Na+/H+ antiport in canola plants and
it showed enhanced growth compared with wild type plants subjected to salinity stress. The
betA gene from E-coli was used to transform a maize line. This gene encodes an important
enzyme i.e. choline dehydrogenase in the biosynthesis of glycine betaine (Quan et al., 2004).
The disadvantage of this technique may be that the character improvement is done at cell
which may not appear in whole plant (Ashraf, 1994).
2.3.3 Exogenous application of phytohormones
When plants are subjected to salinity stress, their indigenous level of hormones is
disturbed. Younis et al. (2003) reported that salinity stress resulted in the decreased level of
IAA and increase in ABA in maize shoots. Similarly, Fricke et al. (2004) found that salinity
stress caused the accumulation of ABA in barley leaves. Akhiyarova et al. (2005) also found
increased contents of ABA in all plant parts due to salinity stress. Boucaud and Unger (1976)
reported decreased in gibberellic acid and cytokinin contents and increased ABA contents
owing to salt stress. Similar changes in level of IAA in maize have also been reported (Ribaut
and Pilet, 1994). It can be deduced from all these results that retardation in growth of plants
due to salinity stress is caused by changes in internal hormonal balance of plants. The
balancing of hormonal levels in plants under salinity stress can be achieved by the exogenic

12
application of phytohormones. Parasher and Varma (1988) reported positive effect of GA3
application in wheat under salinity stress. Afroz et al. (2005) reported that GA3 application
enhanced the photosynthetic efficiency and nitrogen uptake of mustard plants and improved
the growth. Improvement in seed germination and plant seedling by the application of GA3
and kinetin has been reported by Satvir et al. (1998). IAA application enhanced the uptake of
mineral nutrients and lowered the uptake of Na+ which ultimately resulted in better growth of
maize plants (Darra and Saxena, 1973). Datta et al. (1997) also found seed treatment with
kinetin and IAA effective in mitigating the salinity stress in plants.
2.3.4 Use of plant growth-promoting rhizobacteria (PGPR)
It is well established plant hormones play a vital role in controlling plant growth and
development. Many workers have reported that inoculation with auxin producing plant
growth promoting rhizobacterial can improve the growth of crop plants under salt affected
soil conditions. Zahir et al. (2010) conducted a pot trial to investigate the effect of Rhizobium
phaseoli to improve the growth of mung bean under salt affected conditions. They reported
that salinity badly reduced the growth of mung bean while application of rhizobium and L-
TRP were found very effective in mitigating the negative effects of salinity. Their combined
application was more effective than their sole application. They concluded from their results
that this improvement in growth might be due to auxin production ability of these microbes.
They suggested that inoculation with rhizobium producing auxins may be a useful tool to
induce the salt tolerance in mung bean.
Nia et al. (2012) conducted a greenhouse experiment to test the effectiveness of
inoculation with Azospirillum strains isolated from saline or non-saline soil in alleviating the
salinity stress in wheat plants grown with irrigation water with different electrical
conductivities (ECw) of 0.7, 4, 8 and 12 dS m-1. Inoculation with the two isolates increased
salinity tolerance of wheat plants; the saline-adapted isolate significantly increased shoot dry
weight and grain yield under severe water salinity. The component of grain yield most
affected by inoculation was grains per plant. Plants inoculated with saline-adapted
Azospirillum strains had higher N concentrations at all water salinity levels.

13
Bacteria producing 1-aminocyclopropane-carboxylate (ACC) deaminase can be
helpful in mitigating salinity stress in plants (Glick et al., 1997). Ethylene, a plant hormone,
has been found to inhibit the elongation of plant roots (Abeles et al., 1992). ACC-deaminase
produced by PGPR can breakdown the ACC, the precursor of ethylene and in this way, lower
the ethylene concentration in plants under stress situation (Glick, 1995; Glick et al., 1998). By
lowering ACC levels in plants, these bacteria help those plants in ameliorating the stress and
to improve their growth (Shah et al., 1997; Glick et al., 2007).
The symbiotic relationship between rhizobia and legumes is disturbed due to salinity.
It is documented in many studies that nodulation and N2 fixation was significantly decreased
due to salinity stress (Rabie et al., 2005). Other authors reported a decrease of rhizobial
colonization and shrinkage of root hairs of soybean, common bean, and chickpea in salt stress
(Singleton and Bohlool, 1984; Zahran and Sprent, 1986; Elsheikh and Wood, 1990).
The study of Marcar et al. (1991) indicated that symbiotic properties were more
sensitive to salinity than plant growth. Thus, the development of salt-tolerant symbioses is an
absolute necessity to enable cultivation of leguminous crops in salt-affected soils (Lauter et
al., 1981; Yousef and Sprent, 1988; Velagaleti and Marsh, 1989).
In the rhizosphere, the synergism between various bacterial genera, such as between
Bacillus, Pseudomonas, and Rhizobium has been demonstrated to promote plant growth and
development (Bolton et al., 1990; Halverson et al., 1993). The coinoculation with
Pseudomonas spp. and Rhizobium spp. had enhanced nodulation and nitrogen fixation, plant
biomass, and grain yield in various leguminous species (Bolton et al., 1990; Dashti et al.,
1998; Goel et al., 2002; Tilak et al., 2006). We have also observed that root-colonizing
Pseudomonas strains improve rhizobia-legume interactions. Combined inoculations could be
an option to improve plant growth and increase nodule numbers and N content of Galega
species (personal communications). Plant growth-promoting bacteria can prevent the
deleterious effects of stressors from the environment (Lugtenberg et al., 2001; Egamberdieva,
2009). Hasnain and Sabri (1996) reported that inoculation of plants with Pseudomonas sp.
stimulated plant growth by reduction of toxic ion uptake and increases in auxin contents.
Another explanation for enhancement of nodule formation by the rhizobia in legumes might
be the production of hydrolytic enzymes such as cellulases by root colonizing Pseudomonas

14
strains, which could make penetration of rhizobia into root hairs or intercellular spaces of root
cells easier, leading to increased numbers of nodules (Sindhu and Dadarwal, 2001).
2.4 Auxins
Auxins are a very important group of plant hormones which regulate different aspect
of plant growth and development. There are different types of auxins present in plants among
them indole-3-acetic acid (IAA), indole-3-propanic acid (IPA) and indole-3-butyric acid
(IBA) are the common examples of naturally occurring auxins. In tissues of plants simple
auxins can bind with other organic molecules to form conjugates which can be stored in
plants (Frankenberger and Arshad, 1995). Tryptophan is an essential amino acid which can
play the role of precursor of auxins (IAA). Through different pathways tryptophan undergoes
into various biochemical changes which result in wide variety of intermediates indole-3-
acetamide (IAM), indole-3-acetonitrile (IAN), tryptamine (TAM), tryptophol (TOL) and
indole-3-ethanol. Among different types of auxins in indole-3-acetic acid (IAA) is the
functional unit and most active form of auxins (Davies, 1995; Frankenberger and Arshad,
1995; Yasmin et al., 2007; Ali et al., 2008; Sridevi and Mallaiah, 2008).
2.4.1 Sources of auxins
Like higher plants, different soil microbes also possess the ability to biosynthesize the
auxins. Owing to presence of various substrates as root exudates microbes present in the
rhizosphere are more active and efficient in producing auxin than those present in the bulk
(Asghar et al., 2004; Khalid et al., 2004; Zahir et al., 2005). In the rhizosphere, some of the
auxins are released by the plants in the form of root exudates, where rest of the auxins might
have been synthesized by the active rhizospheric microbes. Rhizobia are endogenous
microbes and due to close contact with the tissues of plants are very active in producing
auxins. Etesami et al. (2009) isolated 100 rhizobial isolates belonging to genera Rhizobium
leguminosarum var. phaseoli and Rhizobium leguminosarum var. viciae and selected the five
rhizobial isolates superior in IAA production in vitro. These isolates were used as inoculants
alone and in combination with L-TRP to study their effect on growth parameters of wheat.
Ghosh et al. (2008) reported the presence of higher amounts of indole-3-acetic acid (IAA) in
root nodules of Phaseolus mungo. Rhizobia biosynthesized large quantities of auxin (142 μg
mL-1) upon the addition of L-tryptophan to the inoculation medium. An increase in auxin

15
production was observed when mannitol (1%), L-asparagine (0.3 %) and thiamine
hydrochloride (1 μg mL-1) were added in the inoculation medium in addition to L-tryptophan
(2 mg mL-1). Erum and Bano (2008) isolated rhizobial strains from high altitudes of northern
Pakistan and studied their phytohormone production capabilities (auxins and gibberillins).
They reported that rhizobia isolated from five different locations of higher altitudes of
Pakistan were capable of producing IAA and GA. They also studied the beneficial effects of
these isolates upon inoculation on soybean crop. Sridevi and Mallaiah (2007) isolated twenty
six rhizobium strains from root nodules of Sesbania sesban collected from different regions
of Andhra Pradesh. All the rhizobium strains produced auxins but the maximum auxin
production was observed by only five strains in yeast extract mannitol (YEM) medium upon
the addition of 2.5 mg mL-1 L-tryptophan.
Bhowmick and Basu (1987) reported that Rhizobium sp. isolated from legume host
produced auxin but a significant difference in auxin biosynthesis by the strains was observed
depending upon the culture conditions. Bhowmick and Basu (1986) reported that large
amounts of auxin were produced by the Rhizobium sp. from the root nodules of Sesbania
grandiflora upon the addition of L-tryptophan in the culture medium. Auxin production could
be promoted more than 76% by supplementing the medium with mannitol, sodium molybdate
and Triton X-100. Basu and Ghosh (1998) found that root nodules of a monocotyledonous
tree, Roystonea regia, produced high quantities (16.9 μg g-1 fresh mass) of auxin. Root
nodules contained higher amounts of tryptophan (1555.1 μg g-1 fresh mass) which might have
been microbially converted into auxin. The Rhizobium sp. isolated from the root nodules of R.
regia produced high amount of auxin in tryptophan supplemented medium. Dullart (1970)
quantified the auxin present in root nodules and roots of Alnus glutinosa (L.)
spectroflurometrically. More auxins were present in nodules than in roots. Kaneshiro et al.
(1990) conducted experiments on two different strains of Bradyrhizobium japonicum (USDA
110 and 26) which are different in biological nitrogen fixation and auxin-biosynthesis in
soybean. These strains produced effective nodules and symbiont 26 contained 0.3–1.1 μg
IAA g-1 nodular mass. Infection of nodules by tryptophan catabolic variants 4b and 20d,
isolated from B. japonicum strain also extracted two to four folds more auxin (2–3.9 μg g–1)
while nodules infected with B. japonicum strains USDA 110 and 123 produced significantly
less auxin. Yasmin et al. (2007) screened fifteen rhizobacterial isolates from sweet potato

16
rhizosphere for in vitro auxin biosynthesis with and without L-TRP, solubilization of
phosphates and N-fixing ability and reported that all the isolates efficiently produced auxin in
L-TRP supplemented medium. Most of the isolates were effective in biological nitrogen
fixation and phosphate solubilization while differed in plant growth promoting activities.
Asghar et al. (2000) tested 100 rhizobacterial isolates from rapeseed for in vitro auxin
biosynthesis which ranged between 0.13 to 14.4 μg IAA mL-1 of the culture medium. Auxin
production increased many folds in supplementation with filter sterilized L-tryptophan.
Sarwar and Kremmer (1995a) tested the in vitro auxin-producing capabilities of 16 cultures
belonging to Xanthomonas, Pseudomonas, Agrobacterium spp. and Alcaligens and four
Pseudomonas spp. They observed more auxin biosynthesis in rhizosphere isolates than non-
rhizosphere isolates. Sarwar and Kremmer (1995) also concluded from another study that 16
out of 70 isolates were capable of metabolizing L-TRP into auxin in the range of 4.0-86.1 μg
IAA mL-1 of the liquid medium. Spaepen et al. (2009) reported that interaction of rhizobia
with rhizosphere results in the production of plant hormones like auxins in addition to many
other compounds. Although most of the rhizobia have the ability to produce auxins (IAA), yet
the role of auxin in the legume rhizobium symbiosis is not clear. To test the effect of auxin on
rhizobial gene expression, a transposon (mTn5gusA-oriV) mutant library of Rhizobium etli,
enriched for mutants that show differential gene expression under microaerobiosis and/or
addition of nodule extracts as compared with control conditions, was screened for altered
gene expression upon addition of auxin. Four genes were regulated by auxin which appeared
to be involved in plant signal processing, motility or attachment to plant roots, clearly
demonstrating a distinct role for auxin in legume-Rhizobium symbiosis.
2.4.2 Substrate dependent biosynthesis of auxins by other soil micro biota
Other than rhizobia some other microbes such as bacteria, fungi and actinomycetes
when provided with suitable precursor also possess the ability to produce the auxin. Certain
rhizobacterial strains isolated from wild plants have been reported to produce auxins and
these strains have also potential to enhance the growth of wheat (Ali et al., 2009). These
strains when supplemented with precursor L-TRP produced auxin in the range of 0.6-8.22 μg
mL-1 IAA. These researchers proposed that growth enhancement of wheat by these strains
were due to their ability to produce auxin. Gas chromatography and mass spectrometric (GC-

17
MS) analysis revealed that auxin was produced by bacterial strains in the range of IAA in
supplementation with L-Trp. Auxin produced as a result of plant microbe interactions
significantly increased growth of T. aestivum. Nassar et al. (2005) isolated auxin producing
yeast (Williopsis saturnus) from the rhizosphere of maize. These yeasts were used to
investigate their ability to produce auxin in vitro and their potential to enhance the growth of
maize in axenic and green house conditions. These strains produced many fold higher level of
auxins when supplemented with L-TRP as compared to controls where no augmentation with
L-TRP was done. Growth of maize seedlings when inoculated with these strains was also
significantly improved in growth room and green house experiments. Significant increases in
shoot and root dry weight and lengths were observed when maize seedlings were inoculated
with W. saturnus in the presence of L-TRP. These strains did not produce detectable amounts
of gibberellins or cytokinins. In another experiment of the same study another strain of
endophytic fungi R. glutinis which had similar root colonization ability but did not possess
the ability of auxin production was unable to promote growth of maize seedlings. These
findings highlight the significance of substrate dependent auxin biosynthesis and its role in
plant growth promotion.
Tami and Yaacov (1992) documented that higher concentrations of L-TRP in the
culture of Azospirillum brasilense Sp7 produced toxic effects which were revealed in the
form of decrease in growth, changes in colour of colonies, changes in DNA and alterations in
path of protein synthesis. IAA biosynthesis is affected by the age of culture and concentration
of added L-TRP. It was found that IAA production was linear vs. time after the addition of
tryptophan by pulse injection and younger cultures produced higher amounts of IAA as
compared to older cultures. Thus IAA production from tryptophan and its eventual release in
growth medium of A.brasilense Sp7 might be a way out through which this bacterial strain
overcomes the toxic concentrations of tryptophan. Tao et al. (2008) demonstrated the
enhanced production of indole-3-pyruvic acid from an, amino transferase, Trp transaminase
(TAA1) from L-TRP.
Although auxin production by soil microbes positively influence the growth and
development of plants in most cases, but there are some pathogenic species of microbes
which have auxin production abilities but they cause highly negative effect on plant growth

18
and health. Vandeputte et al. (2005) reported that an auxin producing gram positive
pathogenic bacterial strain Rhodococcus fascians, produced leaf galls in plants possibly due
to auxin production. Regulation of auxin production is controlled by plasmid encoded genes.
Intermediates of indole -3-acetic acid present in cells of Rhodococcus fascians indicate that
this strain used IPA pathway as their biosynthesis while possibly limited the indole-3-ethanol
pathway. In this study, auxin production is directly correlated with leaf galls by these
pathogenic bacteria.
Ali et al. (2008) isolated 16 bacterial strains from different plant species.
Identification of these strains was carried out by using 16S rRNA gene sequencing. After this
these strains were tested for their abilities to produce auxin and for their potential to enhance
the growth of Vigna radiata (L.). Auxin production by these strain ranged from 0.60 to 3.0 μg
IAA mL-1 in the presence of L-TRP. Auxin production was confirmed by the gas
chromatography and mass spectrometric GC-MS analysis. Inoculation with B. megaterium
MiR-4 (61%) and B. pumilus NpR-1 strains resulted in significant improvements in all tested
growth parameters. Positive correlation of auxin production and various parameters of growth
was drawn such as shoot length (r = 0.495*, P = 0.05), number of pods (r = 0.498*, P = 0.05)
and grain weight (r = 0.537*, P = 0.05) at full maturity under greenhouse conditions. Results
of this study suggested that auxin production by these strains was such a trait that can be
exploited for increased production of Vigna radiata (L.).
2.4.3 Biochemistry of microbial production of auxins
On the basis of their intermediates i.e. indole-3- acetamide (IAM), indole-3-pyruvate
(IPyA), tryptamine, and indole-3-acetonitrile (Patten and Glick, 1996), plants and
microorganisms produce auxin by using different pathways that may be tryptophan dependent
or independent (Bartel, 1997). Disease causing bacteria cause hyperplasia by auxin
production which alters the appearance and development of plants. Morris (1995) reported
that Agrobacterium tumefaciens, Agrobacterium rhizogenes, Pseudomonas savastanoi and
Erwinia herbicola carry out biosynthesis of auxin through the IAM pathway this was
confirmed by the analysis of iaaM gene from DNA sequencing.

19
Disease induction can be controlled by reducing the supply of tryptophan and
inhibiting the activity of iaaM as IAA production requires iaaM gene. E. herbicola pv.
gypsophilae uses IAM and IPyA pathways for the biosynthesis of auxin. IAM pathway
controls the size of gall while IPyA pathway regulates the epiphytic fitness (i.e. plant growth
monitoring) (Hutcheson and Kosuge, 1985). It is like a universal truth those bacteria that
cause diseases in plants and those that help the plants in better growth (i.e. PGPR) both
biosynthesize the auxin. Auxin production by the plant growth promoting rhizobacteria
improves the nutrient uptake of the inoculated plants which helps in better growth of these
plants. Beneficial bacteria mostly synthesize the auxins by IPyA pathway (Costacurta and
Vanderleyden, 1995). Azospirillum brasilense a plant growth promoting rhizobacterial strain
can produce auxin by one tryptophan-independent pathway and two tryptophan-dependent
pathways (IAM and IPyA) (Prinsen et al., 1993).
Badenoch-Jones et al. (1983) reported that most species of Rhizobium can produce
auxin. It is documented in many studies that alterations in levels of phytohormones such as
auxin and cytokinins are the basic requirements for nodulation (Mathesius et al., 1998). On
the roots of alfalfa pseudo nodules are produced due to inhibition of auxin polar transport
(Hirsch and Fang, 1994). In rhizobia nod gene is expressed by plant produced flavonoids
which produce Nod factors (lipo-chitin oligosaccharides or LCOs) and results in inhibition of
auxin transport in legumes (Mathesius et al., 1998). In a transgenic white clover plant,
expression of the auxin-responsive reporter constructs GH3: gusA was rapidly and transiently
down regulated after inoculation by Rhizobium leguminosarum bv. trifolii which was
subsequently up regulated at the site of nodule initiation (Mathesius et al., 1998). These
studies predicted that local lowering of the auxin balance is a prerequisite for the initiation of
nodule primordia. Prinsen et al. (1991) reported that auxins production by Rhizobium is
stimulated by flavonoids to make Nod factors (lipochitino-oligosaccahride compounds). Co-
inoculation of Rhizobium and Azospirillum can increase nodulation (Dullaart, 1970; Volpin et
al., 1996). Inoculation with A. brasilense resulted in significant increase in activity of nod-
inducer of crude alfalfa root exudates because these exudates possessed flavonoid compounds
compared with those of un-inoculated plants (Volpin et al., 1996). Root nodules possess more
auxin compared with un-inoculated roots and active auxin biosynthesis takes place in
functional nodules (Badenoch-Jones et al., 1983). It is concluded that increased auxin levels

20
in nodules are resulted from rhizobia because a mutant of Bradyrhizobium japonicum that
produces 30-fold more auxin than the wild-type strain exhibited higher nodulation efficiency
(Kaneshiro and Kwolek 1985). Inoculation with mutant B.japonicum strains overproducing
IAA produced higher quantities of auxin in comparison with wild-type bacteroids (Hunter,
1989) predicted that increased auxin biosynthesis in nodules is of bacterial origin.
Badenoch-Jones et al. (1983) detected the radioactivity in all tissues after hours of
high specific activity [(3)H]IAA application to the apical bud of intact pea (Pisum sativum L.
cv Greenfeast) plants. More radio activity was detected in the nodules than root and more in
functional nodules than in non-functional nodules. Thin layer chromatography (TLC) analysis
confirmed the presence of IAA and indole-3-acetylaspartic acid (IAAsp) in effective nodules
and also the involvement of auxin metabolism in functional root nodules. Williams and
Signer (1990) demonstrated the biosynthesis of auxin by Rhizobium meliloti Rm1021 in
alfalfa when supplemented with tryptophan. Mutants which did not utilize tryptophan analogs
still produced indole-3-acetic acid but were Fix (-) as these bacteria did not fully differentiate
into the nitrogen fixing bacteriods. These mutants possessed essential genes which were
tightly linked to a dominant streptomycin resistance locus. Kepinski (2008) concluded from
experiments that auxin, in spite of being simple molecule, plays a crucial role in plant growth
and development. Auxin regulated gene expression by promoting the ubiquitin-dependent
proteolysis of the Aux/IAA family of transcriptional repressor proteins. Mode of action of
auxin is binding the TIR1/AFB auxin receptor F-box proteins thereby enhancing their
recruitment of Aux/IAA repressors for SCF-mediated ubiquitination. TIR1 is an integral
constituent of an SCF-type E3 ubiquitin ligase and is thus the prototype of an entirely
different receptor in which the generic mechanism for ubiquitination is regulated by the
binding of a small molecule. The auxin-mediated destabilization of Aux/IAAs genes mediates
the expression of Auxin Response Factors (ARF) family of DNA-binding transcription
factors.
2.5 Indole acetic acid (IAA) producing rhizobacteria
Many important plant-microbial interactions center on the production of auxins, IAA
being the main plant auxin. The IAA is responsible for the division, expansion and
differentiation of plant cells and tissues and stimulates root elongation. The ability to

21
synthesize IAA has been detected in many rhizobacteria as well as in pathogenic, symbiotic
and free living bacterial species (Costacurta et al., 1995; Tsavkelova et al., 2006).
At present, auxin synthesizing rhizobacteria are the most well-studied phytohormone
producers (Tsavkelova et al., 2006; Spaepen et al., 2007). These rhizobacteria synthesize IAA
from tryptophan by different pathways, although it can also be synthesized via tryptophan-
independent pathways, though in lower quantities (Spaepen et al., 2007). Phytopathogenic
bacteria mainly use the indole acetamide pathway to synthesize IAA, which has been
implicated in tumor induction in plants. It is not clear whether it is used by beneficial
bacteria. In contrast, the indole pyruvic acid pathway appears to be the main pathway present
in plant growth promoting beneficial bacteria (Patten and Glick, 2002).
Azospirillum is the most studied indole acetic acid producer among all plant growth
promoting rhizobacteria (Dobbelaere et al., 1999). Other bacteria belonging to different
genera i.e. Aeromonas (Halda-Alija, 2003), Azotobacter (Ahmad et al., 2008), Bacillus
(Swain et al., 2007), Burkholderia (Halda-Alija, 2003), Enterobacter (Shoebitz et al., 2009),
Pseudomonas (Hariprasad and Niranjana, 2009) and Rhizobium (Ghosh et al., 2008) have
also been found to possess IAA production ability. Inlole acetic acid producing bacteria on
inoculation increase the germination of seed, stimulate growth of root, alter the root system
architecture and improve the biomass of root. Tsavkelova et al. (2007) have reported
improvement in seed germination of orchid seed (Dendrobium moschatum) by inoculating
with an auxin producing Mycobacterium sp. Along with stimulation of root growth, these
bacterial species also enhance the growth of tubers in tuber crops. Swain et al. (2007) applied
Bacillus subtilis as a suspension on the surface of Dioscorea rotundata L. and found
improvements in growth parameters as compared with control plants.
2.5.1 Impact of microbially derived auxins on plant growth and development under
normal soil conditions
Auxins produced by the microbes augment the suboptimal levels of auxins in plants
and help in regulating normal functioning of plants. Ali et al. (2009) investigated the effect of
different auxin producing PGPR on the growth and development of wheat crop (Inqalab-91)
auxin production by these strains was confirmed by GC-MS analysis. It was found that auxin

22
production these strains ranged 0.6-8.22 μg IAA mL-1 in L-TRP supplemented media. A
significant positive correlation was observed between auxin production by bacterial strains
and endogenous IAA content of T. aestivum for GC-MS (r = 0.62; P = 0.05) and colorimetric
analysis (r = 0.69; P = 0.01). Likewise, highly significant positive correlation for shoot length
(r = 0.637; P = 0.01) and shoot fresh weight (r = 0.63; P = 0.01) was observed with auxin
production under gnotobiotic conditions. Inoculation with these bacterial isolates increased
shoot length (up to 29.16%), number of tillers (97.35%), spike length (25.20%) and seed
weight (13.70%) at harvesting.
IAA producing rhizobacterial strains had the potential to increase the endogenous
levels of auxin in wheat and its growth. Bacterial strains isolated from the rhizosphere of wild
plants can be used for enhancing the growth and production of agronomic crops. Patten and
Glick (2002) reported that beneficial auxin producing PGPR had the potential to improve
plant growth while IAA producing pathogenic bacteria caused tumors in plants by using the
indole-3-acetamide pathway. In order to investigate the role of IAA produced by bacteria on
enhancing the host plant root growth, an IAA deficient mutant Pseudomonas putida GR12-2
was developed. For this purpose ipde gene was isolated. Ipde gene is involved in encoding an
important enzyme in indole-3-pyruvic pathway via indole pyruvate decarboxylase was
developed by using bacterial cultures were potentially capable of increasing endogenous IAA
content and growth of T. aestivum var. Inqalab-91. Microbial strains associated with wild
herbaceous flora can be effectively used for improving growth and yield of agronomically
important crops. Pattens and Glick (2002) reported that many plant-associated bacteria
synthesize IAA while tumors (out growths which are formed due to auxin overproduction)
are induced in plants as a result of IAA production by phytopathogenic bacteria, mainly via
the indole-3-acetamide pathway. Upon inoculation with wild type Pseudomonas putida
GR12-2 canola seedlings roots were increased 30-50% as compared to those seedlings that
were inoculated with IAA deficient bacteria.
It is highlighted from these studies that IAA produced by bacterial strains affects the
growth and development of roots of host plants. In hydroponic study, Gravel et al. (2007)
used different bacterial and fungal strains to investigate their effect on the growth of tomato
plants. Pseudomonas putida and T. atroviride improved fruit yields in rock wool and in

23
organic medium as a result of microbial biosynthesis of auxins in supplementation of L-Trp,
tryptamine and tryptophol @ 200 mg mL-1 in the culture medium. With increasing
concentration of L-TRP the root/shoot fresh weights of tomato plants were increased by
inoculation with both of these strains. It was also found that IAA (0-10 mg mL-1) application
along with bacterial inoculation significantly improve the root growth. The detrimental effect
of auxin (IAA) on root elongation might had been reduced due to reduced ethylene
biosynthesis from its precursor ACC by microbial transformation of IAA in the rhizosphere
and/or by ACC deaminase activity contained in both microorganisms.
Perrine et al. (2004) tested the IAA biosynthesis by using an isotope dilution GCMS
assay of IAA and L-Trp in the culture supernatant of Rhizobium leguminosarum bv. trifolii
strains for increasing or inhibiting rice plant growth. Results revealed higher quantities of
auxin by rhizobial strains supplemented with tryptophan in BIII minimal medium compared
to BIII media alone. Rhizobial strains synthesized very low quantities of auxin required for
inhibiting rice growth thus proving that auxin produced by rhizobial strains had no role in
limiting rice growth and development. Pedraza et al. (2006) used nine strains of nitrogen
fixing bacteria efficient in IAA biosynthesis in tryptophan supplemented media. Auxin
production was the highest (16.5-38 µg IAA mg-1 protein) by Azospirillum strains while
Glucon acetobacter and P. stutzeri biosynthesized lower quantities of auxins (1 - 2.9 µg mg-1
protein) in supplementation with tryptophan. Inoculation with B. megaterium MiR-4, B.
pumilus NpR-1 and B. subtilis TpP-1, under axenic conditions increased shoot length by 56,
47 and 46% over un-inoculated control. Bacillus megaterium MiR-4 and B. pumilus NpR-1
inoculation increased shoot fresh weight by 61 and 38%, respectively, over control. There
was highly significant positive correlation between auxin production and growth parameters
of Vigna radiata (L.) under axenic and greenhouse conditions. This implies that auxin-
producing Bacillus spp. can be effectively utilized for enhancing the yield and of Vigna
radiate (L.).
Bellamine et al. (1998) demonstrated the effect of exogenous application of auxin on
poplar shoots, raised in vitro, induced to root by incubation on an auxin (NAA) containing
medium for 7h. Shoots were kept in an auxin-free medium for 13 days post treatment.
Ninteen-seven percent (97%) of the treated shoots produced roots. Application of auxin

24
inhibitors (POAA-phenoxyacetic acid; PBA-2-phenoxy-2-methylpropionic acid; PCIB-2-
(pchlorophenoxy)-2-methylpropionic acid) into auxin-free medium after the 7h induction by
NAA (α-naphthaleneacetic acid), completely or severely inhibited rooting. (PCIB, PBA and
POB are anti-auxin compounds). The exclusion of calcium from the auxin-free medium
reduced the percentage of rooting by 42%. The inhibition was still higher in the presence of a
calcium chelator EGTA (ethyleneglycol-O, Ό-bis (2-aminoethyl ether)-N, N, Ń, Ń, tetraacetic
acid). Application of lanthanum chloride (calcium channel blocker) to the auxin-free medium
completely inhibited rooting. These results confirmed the involvement of endogenous auxin
and calcium application in the later stages of the adventitious root formation.
Hussain et al. (1988) used 10 different concentrations of IAA sprays and investigated
their effect on the growth and nodulation of Leucaena leucocephala. They used 10 sprays per
week. After each spray they observed the girth and height of plants. After eight and half
months plants were harvested and weights of roots, nodules and numbers were recorded. The
plant species were also analyzed for their nutrient concentrations. At 50 ppm IAA the
parameters were significantly enhanced. Higher doses of IAA caused negative effects on the
growth and their values were found even lower as compared with controlled plants. Rafael et
al. (1989) used root stocks of dwarf apple verities and studied their root response and levels
of indole-3-acetic acid. They used Linsmaier-Skoog medium which was supplemented with
benzyladenine (BA), indole-3-butyric acid (IBA), and 1, 3, 5-trihydroxybenzoic acid (PG) to
grow the apple root stock. Lepoivre medium was used to investigate the effect of IBA and PG
on root growth and development. M9 and M26 produced 80-100% root response respectively
by using IBA. Before root inducing treatment in shoots of both root stocks free and conjugate
auxins concentrations were evaluated by using HPLC. These results were reconfirmed by
GC-MS. It was revealed from the results that the lower part of M26 contained 2.8 times
higher contents of free auxins than similar tissues in M9. 298 and 263.7 ng per gram fresh
weight of free auxins was recorded in epical parts of M26 and M9. M9 contained
significantly higher amounts of conjugated auxins than M26, which means that most of the
auxins M9 were found in conjugated form. Due to different free auxins concentrations both
strains showed variable rooting response.

25
2.5.2 The role of bacterial phytohormones in plant growth promotion in saline soils
The mechanism through which plant growth promoting rhizobacteria improve the
growth and development of host plant include the mobilization of nutrients (Lugtenberg et al.,
2002; Lugtenberg and Kamilova, 2004) and production of phytohormones (Lifshitz et al.,
1987; Frankenberger and Arshad, 1995). The microbial synthesis of plant growth regulators is
an important factor in soil fertility. Gibberellin and indole acetic acid are secondary
metabolites, which are important biotechnological products, obtained commercially from
bacteria and fungi for the agricultural and horticultural industry (Patten and Glick, 2002).
Indole acetic acid (IAA) is the most common natural auxin found in plants. IAA is involved
in physiological processes such as cell elongation and tissue differentiation (Taiz and Zeiger,
1991; Frankenberger and Arshad, 1995) and has also been associated with the plant growth-
promoting effect of numerous rhizospheric microorganisms (Patten and Glick, 2002).
It is hypothesized that negative effect of salinity on seed germination may be due to
impairment of levels of plant growth hormones (Zholkevich and Pustovoytova, 1993;
Jackson, 1997; Debez et al., 2001). In some earlier studies, it is reported that movement of
cytokinins from root to shoot and from coleoptile tips of maize and the recovery of diffusible
auxins is limited due to salinity stress (Itai et al., 1968). Use of PGRs such as gibberellins,
auxins and cytokinins has been found very effective in removing the deleterious effects of
salinity stress (Gul et al., 2000; Khan et al., 2001, 2004; Afzal et al., 2005; Prakash and
Parthapasenan, 1990). Exogenous application of these PGRs caused improvements in growth
of plants under salinity stress (Saimbhi, 1993). Priming of wheat seed with these PGRs have
been found very effective and alleviating the growth retarding effects of salinity stress (Singh
and Dara, 1971; Datta et al., 1998; Sastry and Shekhawa, 2001; Afzal et al., 2005). In a study
conducted by Sakhabutdinova et al. (2003) showed that salinity stress caused a progressive
decline in the IAA contents in plant root system. Under such conditions exogenous
application of phytohormones supplies additional natural hormones that help the plants for
normal growth and development under salt affected situations (Kabar, 1987; Afzal et al.,
2005).
It is reported in many studies that plant growth promoting rhizobacteria (PGPR)
which can produce phytohormones on inoculation with plants can alleviate the dangerous

26
effects of salinity stress and thus improve the crop growth (Lindberg et al., 1985;
Frankenberger and Arshad, 1995). Egamberdieva (2009) reported that IAA producing strains
improved the germination percentage up to 79% and mitigated the negative effects of salinity
on wheat due to their ability to produce auxin. These strains had the ability to produce auxin
even in saline conditions. In another study Hasnain and Sabri (1996) found that Pseudomonas
sp. reduced the uptake of toxic ions such as Na+ and increased the auxin level of wheat plants.
Phytohormone production by other PGPR has also been demonstrated by other researchers
(Zimmer et al., 1995) and their on growth promotion has been reported (Frankenberger and
Arshad 1995). Plant growth promoting rhizobacteria help the plants by supplying the
additional phytohormes which can mitigate the harmful effects of salty (Egamberdieva,
2009). Keeping in view such results it can be concluded that such phytohormones producing
rhizobacteria can be used for better growing of crops in saline conditions without doing any
genetic changes in plants.
Along with acting as plant growth regulators and stimulating the growth of plant, IAA
also increases the activity of ACC synthase. ACC synthase produces more ACC, which is
transformed by the action of ACC oxidase into ethylene (Mayak et al., 1999). The ACC
produced at higher levels in plants may be exuded out due to concentration gradient and is
hydrolyzed by the ACC-deaminase produced by PGPR. It can concluded from this
mechanism the final effect of ethylene production and its effect on root growth primarily
depends upon balance ACC deaminase and IAA produced by bacterial strain. Plants show
different responses to IAA application. Its effects are determined by the initial root
elongation. The growth of plants which have slow root elongation is restricted by the
application of phytohormones at any concentration, while plants which have fast elongating
roots show increase in elongation by the use of plant growth regulators at low concentrations
(Pilet and Saugy, 1987).
It is conclude that different species of ACC-deaminase producing plant growth
promoting rhizobacteria produce variable amounts of IAA. Interaction of ethylene and IAA
was explained in a model proposed by Glick et al. (1998). Such bacteria not only lower the
concentration of ethylene and relieve the plant from stress but also minimize the synthesis of
auxin response factor (ARF). In this way the bacteria mitigate the effect of stresses and

27
improve the growth and development of plants. However as the synthesis of ARF is
enhanced, production of ACC is also increased which ultimately checks the synthesis of
ARF. Through this feedback mechanism, ethylene limits its production by its own.
Plants are immovable creation, but they can highly modify their physiology which
enables them to withstand different environmental situations. This is due to that, they have
highly active meristem and they can also produce different organs due to embryogenesis
(Wolter and Jurgens, 2009). Plants use different defensive mechanisms such as changes in
root morphology to cope stresses (Malamy, 2005). Hormonal homeostasis is involved in
morphogenesis, cell division and elongation. IAA is the physiologically most active auxin in
plants and its importance in plant growth and development is reflected by the fact that still no
fully auxin deficient mutant in plants is developed. Auxin is mainly produced in meristems of
aerial parts and roots (Teale et al., 2006). Minute changes in auxin concentration play a vital
role in plant growth and development. Especially under stress situations two phenotypic traits
such as formation of lateral roots and branching are altered by changes in auxin level (Potters
et al., 2007).
Higher number of indole acetic acid producing plant growth promoting rhizobacterial
strains was found in plant tissues as revealed by analysis for bacterial existence (Spaepen et
al., 2007). A significant increase in lateral root formation and root hair was recorded in
different species of plants owing to inoculation with such beneficial bacterial strains (Dimkpa
et al., 2009). The beneficial effects of Azospirillum on the growth and development of roots
are reported in many studies. Plants roots inoculated with Azospirillum show morphological
changes mainly due to the production of plant growth regulators such as auxins. Auxin is the
most important hormone among all the hormones (Spaepen et al., 2007). Evidences of
involvement of auxin in root proliferation are documented in many studies. El-Khawas and
Adachi (1999) conducted a hydroponic study to investigate the effect of A. brasiliense on the
root growth of rice plants. They reported on the basis of their results that root length, surface
area of roots, root dry matter, lateral roots and number of root hairs was increased compared
with control plants. IAA applied exogenously to plants also showed similar responses as of
inoculation with Azospirillum (Remans et al., 2008). Low phytohormone producing and

28
having nitrogen fixing ability mutant of A. brasiliense could not increase the root growth as
compared with control plants (Kundu et al., 1997).
Depending upon the relationship between indole acetic acid and ACC, the positive
effects of IAA on root growth can be either direct or indirect through the reduction of
ethylene levels (Lugtenberg and Kamilova, 2009). When plant faces any stress such as
salinity, ethylene regulates the plant growth and causes reduction in root shoot growth. It is
found that under stress conditions ethylene is produced in two phases in plants. At first phase
minute amount of ethylene stimulates the stress response genes in plants. After one to three
days of stress application higher amount of ethylene is produced which results in retardation
of growth caused negative effect on plants such as chlorosis, senescence and abscission
(Glick et al., 2007).
ACC-deaminase producing rhizobacteria lower the ethylene production in plants by
degrading its precursor ACC into 2-oxybutanate and ammonia. Thus in this way these
bacteria reduce the synthesis if auxin response factor (Glick et al., 2007; Kang et al., 2010).
Under stress conditions ACC may be released from the plants due to concentration gradient,
which is consumed by the ACC-deaminase producing rhizobacteria for their growth. ACC-
deaminase production by plant growth promoting rhizobacteria is a beneficial trait for the
plants and PGPR itself for the normal growth, as the concentration of ACC is lowered in
plants and PGPR can use ACC as a metabolite (Glick et al., 1998). Under various
environmental stresses these ACC-deaminase producing bacteria have been practically found
efficient in promoting the growth and development of plants. Mayak et al. (2004) found that
ACC-deaminase producing PGPR Achromobacter piechaudii significantly improved the root
and shoot fresh/dry matter of tomato plants. Saravanakumar and Samiyappen (2007)
compared the effect of ACC deaminase producing PGPR strain and those lacking ACC
deaminase activity. They found that ACC deaminase producing PGPR strains improve the
salinity resistance in growth of the plants as compared to the control plants inoculated with
ACC deaminase lacking strain. In another study, Siddikee et al. (2010) also reported that 14
halo tolerant bacterial strains caused amelioration of salinity stress in canola plant mainly due
to activity of ACC-deaminase. Maize plants inoculated with Pseudomonas fluorescens
showed increased root and shoot growth. Effect of drought stress on the ripening, growth and

29
yield of pea was also revealed due to the ACC deaminase production. Nadeem et al. (2010)
documented improved growth of wheat under salt stress due to inoculation with ACC
deaminase producing plant growth rhizobacteria. Bianco and Defez (2009) observed the
response of Medicago plants inoculation with Sinorhizobium meliloti strains under high salt
stress condition. They reported that IAA overproduction showed remodulation of
phytoharmones in different plant tissues. Higher levels of auxins were found in roots and
nodules of plants with reduction in IAA level in shoot was compared in relation to those
plants which were inoculated with wild type 1021 strain (Mt–1021). Ethylene resisting genes
were also observed through transcriptional analysis. The results showed that Mt – RD64 did
not show induction of this pathway under salt stress which is an indication that plants were
rescued from stress damages.
2.6 Other mechanisms of growth promotion by PGPR
2.6.1 Accumulation of protective compounds
Accumulation of nitrogen containing compounds (NCC) such as amino acids,
proteins, quaternary ammonium compounds and polyamines have been correlated with
improved growth and yield under different stresses such as salinity and drought stress.
Szabados and Savoure (2009) reported a very high concentration of proline in cells under salt
and drought stress as compared to normal soil conditions. This is due to enhanced production
and less breakdown of such compounds under stressful conditions. In different studies in
which transgenic plants were used elaborate that metabolism of proline has a complex effect
on development of stress responses. Proline acts as a compatible solute in plants and thereby
reserves the carbon and nitrogen. In some other studies it is also reported that proline acts has
an anti-oxidative activity and serves as a scavenger of ROS. Proline also stabilizes the
architecture of proteins and increases the activity of various enzymes by acting as molecular
chaperones. When inoculated in the plants it controls the cystolic pH and controls the redox
potential of cells. Increased biosynthesis and accumulation of proline due to the inoculation
with various rhizobacterial strains was observed in different plants. The production and
accumulation of such solutes is carried out as expanse of energy (41 moles of ATP) and
reduction in growth. These alterations help the plant to survive and recover under different
stress conditions such as salinity stresses. Bianco and Defez (2009) reported 2 fold increase in

30
proline contents of shoots of Medicago plant as a result of inoculation with Mt-64 strain.
These increased levels of proline were positively correlated with reduced symptoms of
senescence as necrosis, chlorosis and drying.
2.6.2 Biosynthesis of antioxidative enzymes
Plants produce different reactive oxygen species such as super oxide (O2-), hydrogen
peroxide (H2O2), hydroxyl radical (OH) and singlet oxygen (102) in different metabolic
activities as byproducts in various cellular compartments (Apel and Hirt 2004). These
reactive oxygen species are harmful and cause oxidative damage to the DNA, proteins and
lipids. As during photosynthesis in the chloroplasts internal concentrations of oxygen are
very high so these portions are highly prone to oxidative damage by reactive oxygen species.
Under normal conditions these oxygen species are continuously removed by antioxidant
component and plant faces no damage. But under stressful environment the production of
these reactive species is elevated and they are accumulated in the different parts due to
imbalance between their productions and scavenge. Among different ROS scavenges
mechanism, SOD, APX and CAT are the major mechanisms. Different antioxidants such as
ascorbic acid and glutathione are present at high concentration in chloroplasts and other
compartments of cells. These play a vital role in defending the plant from oxidative damage
caused by ROS (Miller et al., 2010). Balance between antioxidants is necessary for the
detoxification of different reactive oxygen species (Mittler, 2002). Kohler et al. (2010)
reported that salinity stress was mitigated due to the induction of antioxidant enzymes in
plants inoculated with PGPR strains. Under medium and high salt stress plants inoculated
with strain showed increased biomass than the control plants. Bianco and Defez (2009) also
reported that plants inoculated with Mt-RD64 strain showed less signs of oxidative damage.
This improvement was mainly due to the enhanced activity of antioxidant enzymes.
Sandhya et al. (2010) conducted a study to investigate the effect of five drought
tolerant PGPR strains on the growth and antioxidative activity of maize. The results of the
study revealed that the activity of antioxidant enzymes was lower in inoculated plants as
compared with the uninoculated plants. In barley plants the antioxidative activity was also
reduced. A study conducted by Omer et al. (2009) revealed that under salinity stress activity

31
of antioxidant enzymes such as catalase and peroxidase increase significantly in two
cultivars of barley. On the other hand when these plants were inoculated with Azospirillum
baselines, they showed lowering in the activity of such enzymes, which removed the
deleterious effects of salinity and enhanced the growth and yield of barley plants under salt
effected conditions.
2.6.3 Enhancement of nutrients uptake
Different adaptive mechanisms that plants adopt to avoid or tolerate different stresses
define the survival and productivity of these plants under stresses. Many research studies
report that mineral status plays a crucial role in adapting the harsh environmental stress
conditions. Abiotic stresses disturb the mineral uptake and then accumulation in plants and
external application of these nutrients can alleviate the stress symptoms on plants.
Phosphorus after the nitrogen is the 2nd most important mineral nutrient for growth of plants.
It is the part of various biochemical compounds such as nucleotides, nucleic acid,
phosphoproteins and phospholipids. Under various conditions of salinity stress uptake and
accumulation of phosphorus is reduced and plants develop deficiency symptoms (Martinez
and Lauchlie 1994). Under salt affected soil conditions lowering the availability of P may be
due to the ionic strength that results in the reduced activity of phosphate. Reduced solubility
of Ca-P minerals adsorption of P may be other processes that lower the P availability under
salt effected soil conditions. In soil amount of soluble P is very minimum (less than 1ppm)
(Hinsinger, 2001). Cells can uptake different forms of P but HPO4-2 or HPO4
-1 are the main
forms of P that are adsorbed. In soil P is in the organic and inorganic forms. Like other
mineral nutrients such as K+1, Fe+2, Zn+2 and Cu+2, its mobility in soil is very low (Hayat et
al., 2010). Plant growth promoting rhizobacteria convert the insoluble form of P, both present
in organic and inorganic forms into soluble forms that are available to the plants. Different
plant growth promoting rhizobacterial strains that belong to different genera can solubilize
insoluble compounds of phosphorous. Generally these strains produce some organic acids
that can solubilize the mineral phosphates (Rodriguez and Fraga, 1999). These organic acids
cause acidification in the cells of microbes and thus surrounding environment. In this way P
is released by proton substitution for Ca+2. Many plant growth promoting genera such as

32
Pseudomonas, Erwinia, Rhizobium and Bacillus have been found to produce such organic
acids (Rogriguez and Fraga, 1999).
Soil salinity poses serious threat to crop farming throughout the world. A number of
physical, chemical and biological approaches have been employed to combat this menace.
Soil inhabiting bacteria are also recently being used for this purpose. These bacteria use
different mechanisms to help the plants to withstand salinity stress. Among such bacteria are
those which are capable of producing auxin. These bacteria have been tested on different
crops under normal soil conditions. Very scarce information is available that reports the
efficacy of these bacteria under salt affected soil conditions. Keeping in view the findings of
previous research a comprehensive study was planned to evaluate the effectiveness of auxin
producing rhizobacteria for enhancing the growth, physiology and yield of maize in salt
affected soil conditions.

33
CHAPTER-3
MATERIALS AND METHODS
The research work comprised of series of experiments. These were conducted in the
laboratory, growth room, wire house and farmer fields. Large numbers of rhizobacterial
strains were isolated from the rhizosphere of maize plants growing under salt affected soil
conditions. Axenic trials were carried out to screen the strains of rhizobacteria on account of
growth improving potential with maize as test crops. The performances of the screened
isolates were further evaluated under salt affected soil conditions in pots and fields. The
procedures adopted during the whole experimentation are discussed in detail as under:
3.1 Isolation of PGPR
Plant growth promoting rhizobacteria were isolated from the rhizosphere of maize (cv.
Neelam) growing in the salt affected soil (Sandy clay loam, ECe; 7.2 d Sm-1) from the semi-
arid region of Faisalabad, Punjab, Pakistan by dilution plate technique using general purpose
media (GPM). Fast growing colonies were selected and further purified. Isolated
rhizobacteria were stored in refrigerator at 4 0C for further experimentation.
3.2 Preparation of inoculum
Sterilized general purpose media was inoculated with each rhizobacterial culture and
incubated at 28 + 1 0C for 3 days in shaking incubator at 100 rpm. Fresh inoculum were
prepared for each experiment and diluted to get uniform population (108-109 CFU mL-1) by
measuring optical density (OD 0.5) using spectrophotometer at wavelength 535 nm.
3.3 Seed inoculation
The inoculum for PGPR strain was prepared by culturing the strain on 250 mL of liquid
medium in a 500-mL Erlenmeyer flask incubated for 72 h at 29 oC and 100 rpm. Maize seeds
(Monsanto-919) were surface sterilized with 70% ethanol for 2 min, treated with 2% sodium
hypochlorite for 5 min, and followed by three washings with sterilized distilled water. For
inoculation, PGPR inoculum of selected strains, having cell suspension of 107-108 CFU mL-1,
were injected into sterilized peat (150 mL kg-1 of seed, seed to peat ratio 4:1 w/w). Maize
seed dressing was done with inoculated peat mixed with 10% sterilized sugar solution. In

34
case of un-inoculated control, the seeds were coated with the sterilized peat treated with
sterilized broth and 10% sterilized sugar solution but without bacterial inocula. While for co-
inoculation (1:1 ratio) equal volume of both strains was used.
3.4 Osmotic adjustment test
The isolated strains were tested according to the method as described by Zahir et al.
(2010) for their ability to withstand salinity stress. For this purpose, strains were grown in test
tubes on different salinity levels (original, 10, 20, 30 dS m-1). Fifteen milliliters of broth was
taken in test tubes and autoclaved. The tubes were inoculated with the isolated strains and
incubated at 28 + 1 0C and the absorbance was measured at (λ) 540 nm by using
spectrophotometer.
3.5 Measurement of Auxin Production
Auxin production measurement was carried out according to the method described by
the Sarwar et al. (1992). Twenty milliliters of sterilized General Purpose media (GPM) was
taken in Erlenmeyer flasks of 100 mL. Five milliliters solution of 5% L-TRP after filter
sterilization (0.2 μm membrane filter, Whatman No.2) was applied to the medium to prepare
a media with final concentration of 1.0 g L-1
. Inoculation of the flask contents was done with
0.1 mL of 3-day old broth culture (OD 0.5 (108
-109
CFU mL-1
). After proper plugging, the
flasks were incubated for 48 hours using shaking incubator adjusted at 28 + 2 °C and 100
rpm. A flask containing all the materials except inocula was kept as control for comparison.
The experiment was carried out with three replications. After incubation, the contents in the
flask were filtered using Whatman No.2 filter paper. For measuring IAA equivalents, 3 mL of
filtrate was taken in test tubes and 2 mL of Salkowski reagent (2 mL 0.5 M FeCl2 + 98 mL
35% HClO4) was added to it. The mixture in the tubes was allowed to stand for 30 minutes
for colour development. Standard curve was used for the comparison to calculate auxin
production by the selected isolates. Colour was also developed in the standard solutions of
IAA. The intensity of the colour was measured at (λ) 535 nm by using a spectrophotometer.
Rhizobacterial cultures giving higher auxin production were selected for jar experiments.

35
3.6 Screening of rhizobacterial strains for growth promoting potential
A jar experiment was conducted in the growth room under controlled conditions to
screen rhizobacterial strains for their root/ shoot growth promoting ability. For these
experiments 500g sieved sand was filled in the jars. Jars were wrapped with paper and
autoclaved twice. Different salinity levels (EC 0, 4, 8, 12 and 15 dS m-1) were developed in
the jars by using sodium chloride (NaCl) salt. Surface disinfection of maize seeds was done
by rinsing and stirring maize seeds for 3 minutes in a sterile flask with 5% sodium
hypochlorite. After three minutes the seeds were removed from the sodium hypochlorite
solution and washed with sterile water. Surface disinfected seeds were dipped for 10 minutes
in the broth of respective rhizobacterial strains for inoculation. Three inoculated seeds of each
strain were sown in each jar. To meet the water and nutritional requirements of the seedlings,
half-strength Hogland solution (Hogland and Arnon, 1950) was applied to jars. The jars were
placed randomly following completely randomized design in growth room with three repeats.
Jars were placed in a growth chamber at 25 ± 1 8C set at a 16 h light and 8 h dark period, and
the light intensity was set to 350 mmol m–2 s–1.After twenty days of germination, the
seedlings were measured for root/shoot length, fresh and dry weight.
3.7 Pot experiment
On the basis of results from the ‘‘Jar trial,’’ two strains (M4 and M7) giving high mean
values for the growth parameters of seedlings were selected and further evaluated for
improving the growth and yield of maize (Hybrid Monsantto-919) plants with and without L-
TRP by conducting a pot trial in a wire house. Soil used to pot the seeds was analyzed for
physicochemical characteristics (Table 1). Surface-disinfected seeds of maize were inoculated
(as discussed above) with 4 days old cultures of selected rhizobacterial strains. L-tryptophan
solution was applied in the inoculation media. For the uninoculated control, seeds were
coated with the sterilized (autoclaved) peat treated with sterilized broth. Salinity levels of 4,
8, 12 and 15 dS m–1 were maintained (using NaCl salt) in addition to the non-saline control
(original EC, 1.63 dS m–1). For this, the calculated amount of solid salt (NaCl) was mixed in
the soil with a mechanical mixture before filling the pots. Three inoculated/uninoculated,
non-germinated maize seeds were sown in each pot containing 12 kg of soil per pot at all the
5 EC levels which was thinned to one plant, 15 days after germination. There were 3

36
replications of each treatment. Pots were arranged in the wire house at ambient light and
temperature according to a completely randomized design. Recommended doses of N, P, and
K fertilizers (175:100:50 kg ha–1) were applied in each pot. Phosphorus and potassium were
applied as a basal dose, while nitrogen was applied in splits. Tube well water was applied to
the pots for irrigation whenever needed. Data regarding growth and yield parameters were
collected using standard procedures.
3.8 Field experiments
Field experiments were conducted to further confirm the efficacy of selected rhizobacterial
strains for improving growth and yield of maize at farmer fields, 72/GB, Faisalabad-Pakistan
(73-74 E, 30-31.5 N,) for two consecutive years. Soil samples were collected, air dried, mixed
thoroughly, sieved (2.0 mm), and analyzed for physico-chemical characteristics (Table 2).
The soil was analyzed for saturation percentage, pH, electrical conductivity (ECe), cation
exchange capacity (CEC), sodium adsorption ratio (SAR), organic matter, total N, available
P, and extractable K (Table 2) using standard procedures. Inoculated and non-inoculated
(non-germinated) (as described above) seeds of maize (Hybrid Monsantto-919) were sown in
the well-prepared field at 120 kg ha-1 by maintaining a row to row distance of 75 cm with a
plot size of 20 m2. L-tryptophan solution was applied in the inoculation media. The
composition of microbial community of maize was not tested at any time during the
experiment. Phosphorus and potassium were applied as a basal dose, while nitrogen was
applied in splits. The experiments were laid out in randomized complete block design with 3
replications. Plants were irrigated with good quality canal water (electrical conductivity, 0.08
dS m–1; sodium adsorption ratio, 0.2 (mmol L–1)1/2, and residual sodium carbonates, 0.04
mequiv. L–1) meeting the irrigation quality criteria for crops in the area. Growth and yield
contributing parameters were recorded after maturity. Grain and straw samples were analyzed
for nitrogen, phosphorus, and potassium content.
3.9 Soil analyses
Various physical and chemical characteristic of the soil samples were determined
using the procedures illustrated by US Salinity Laboratory Staff (1954) if not mentioned
afterward.
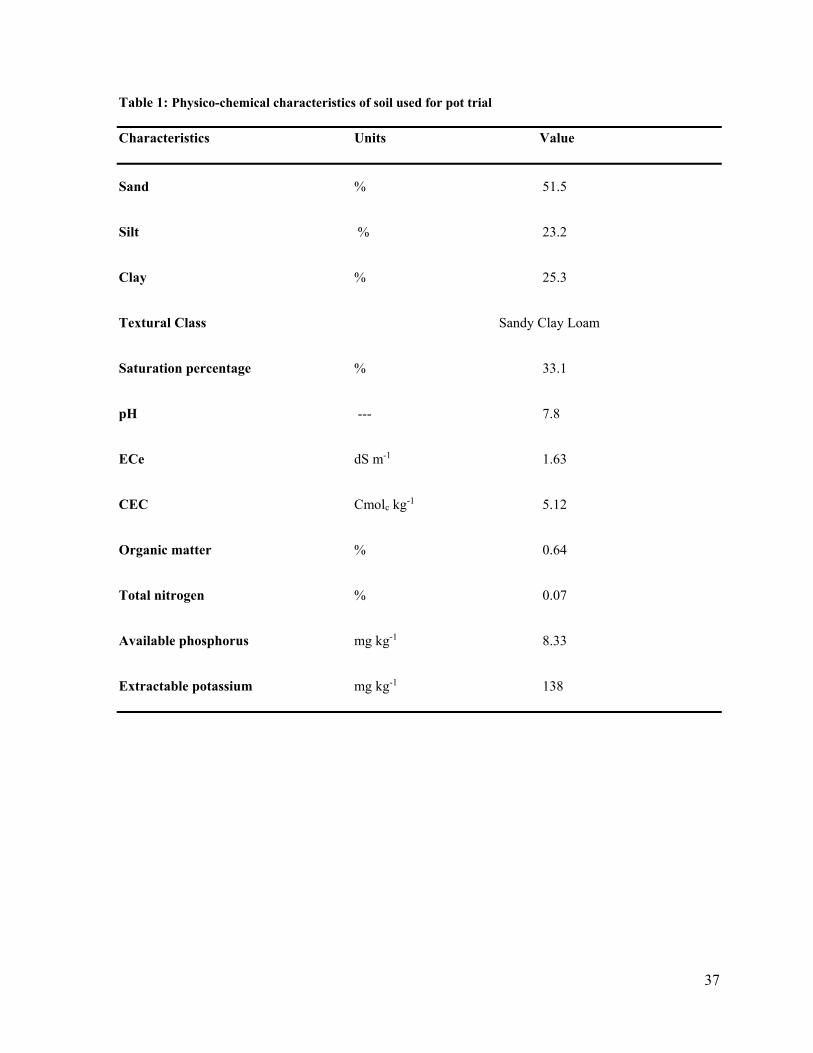
37
Table 1: Physico-chemical characteristics of soil used for pot trial
Characteristics Units Value
Sand % 51.5
Silt % 23.2
Clay % 25.3
Textural Class Sandy Clay Loam
Saturation percentage % 33.1
pH --- 7.8
ECe dS m-1 1.63
CEC Cmolc kg-1 5.12
Organic matter % 0.64
Total nitrogen % 0.07
Available phosphorus mg kg-1 8.33
Extractable potassium mg kg-1 138

38
Table 2: Physico-chemical characteristics of soil used for field trial
Year I Year II Textural Class Sandy clay loam Sandy clay loam
pH 7.6 7.8
ECe (dS m-1
) 6.7 6.8
Organic matter (%) 0.63 0.65
Total nitrogen (%) 0.05 0.04
Available phosphorus (mg kg-1
) 3.93 3.82
Extractable potassium (mg kg-1
) 84.2 78.8

39
Figure S1: Exprimental layout of field trials

40
3.9.1 Particle size analysis
Forty gram of air-dried soil passed through a 2 mm sieve was taken in a 600-mL
plastic beaker. Sodium hexa meta phosphate (2%) was added at 40 mL as dispersing agent
and the soil was left overnight. The suspension of soil was stirred with mechanical stirrer,
poured in a 1-L calibrated cylinder and brought to volume with water. Readings for sand and
silt plus clay were taken by means of Bouyoucos hydrometer following Moodie et al. (1959).
By using International Textural Triangle, soil textural class was dogged.
3.9.2 Saturation percentage (SP)
Saturated paste of soil was made by taking 250 g in a beaker and putting distilled
water whilst stirring the sample using spatula. A portion of the prepared saturated paste of
soil was taken in a tarred china dish. Sample was weighed, dried to constant weight at 1050C
by using an oven and then again weighed. Saturation percentage of the soil sample was
computed using following formula (Method 27a).
Saturation percentage (SP) = (Loss in weight on drying) X 100/ (Weight of the oven dry soil)
3.9.3 Electrical conductivity (ECe)
Saturated paste of soil was transferred into a filter funnel. Vacuum was applied using
a vacuum pump. The saturated extract was collected in a bottle (Method 3a). Using digital
Conductivity Meter, electrical conductivity of the extract was dogged by (Method 3a and 4b).
3.9.4 pH of saturated soil paste (pHs)
After preparation, the saturated soil paste was allowed to stand for 1 h. pH Meter
(Kent EIL Model 7015) was employed to find out the pH of the paste after carefully
standardizing it with standards of 4.0 and 9.2 pH (Method 21a).
3.9.5 Organic matter
One gram of air dried soil sample was weighed in an Erlenmeyer flask of 500-mL
capacity. Solution of 1N potassium dichromate at 10 mL per sample was added by means of a
pipette. Twenty milliliter of sulfuric acid (concentrated) was added to it. The sample was
mixed by shaking and left for 30 minutes. Distilled water at150 mL and 0.5 N ferrous sulfate
solution at 25 mL was added into the sample and the excess was titrated using 0.1N solution

41
of potassium permanganate to pink end point. The same procedure was followed for running
a blank (Moodie et al., 1959).
3.9.6 Total nitrogen
For the determination of total nitrogen, sulfuric acid digestion method of Gunning and
Hibbard’s was used for soil sample digestion and for the distillation of ammonia into 4%
boric acid, macro Kjeldhal’s apparatus was used (Jackson, 1962).
3.9.7 Available phosphorus
Five grams of soil were taken in 250 mL Erlenmeyer flask. Solution of 0.5 M Sodium
bicarbonate was added at 100 mL and the suspension was given 30 minutes shaking. The
extract was taken by filtering the solution and clear extract was taken in volumetric flask at 5
mL Color developing reagent was also added to the flask at 5 mL and completed the volume
using distilled water. By using spectrophotometer, the readings of the soil samples and
standards were recorded at 880 nm wavelengths. Actual P concentration was detected using
standard curve prepared for standards following Watanabe and Olsen (1965).
3.9.8 Extractable potassium
Five grams of soil were taken in centrifuge tube (50 mL capacity) accompanied by 33
mL of ammonium acetate (1N of pH 7.0) solution. After centrifugation, the supernatant were
filtered and taken in 100 mL volumetric flask. The same procedure was repeated two more
times. Jenway PFP-7 Flame photometer was used to determine the potassium (Method 1la).
3.10 Plant analyses
After 60 days fresh leaf samples were collected from the pot and field studies and
analyzed for proline, sodium, potassium and relative water content. At maturity, plants were
harvested and data about growth and yield parameters were recorded. Grain samples were
analyzed for nitrogen and phosphorous.
3.10.1 Measurement of physiological parameters
The physiological parameters i.e. photosynthetic rate (A), stomatal conductance of
water (gs) and transpiration rate (E) was measured by portable infra-red gas analyzer (IRGA).
These parameters were taken in morning (09.00-12.00) at photosynthetic photon flux density

42
of 1200-1400 µ mol m-2 s-1 (Ben-Asher et al., 2006) two fully expanded leaves from one plant
in each experiment unit were selected for the measurement of data regarding above
parameters.
3.10.2 Chlorophyll content (SPAD value)
Chlorophyll content (SPAD value) in leaf of maize plant were measured using
chlorophyll meter SPAD-502 (Minolta, Minolta Co, Ltd, Uk) as described by Hussain et al.
(2000).
3.10.3 Relative water content
Relative water content (RWC) of leaves was determined using the following formula
as described by Mayak et al. (2004).
Relative water content (RWC) = Fresh weight- Dry weight/Fully turgid weight-Dry weight
The fully turgid weight of leaf was taken after putting it in 100 % humidity in the dark
at 4 oC for 48 h.
3.10.4 Electrolyte leakage
Electrolyte leakage (EL) was measured following the protocol described by
Jambunathan (2010). For this purpose, leaf discs were transferred in 5 mL de-ionized water
and electrical conductivity (EC) (R1) was recorded with EC meter (Jenway Conductivity
Meter Model 2052) after 4 h incubation at 28 ± 1 0C and 100 rpm in orbital shaking
incubator. The same samples were autoclaved at 121 0C for 20 min to determine EC (R2).
%EL = (EC before autoclaving R1/ EC after autoclaving R2) X 100
3.11 Analysis of stress-related metabolites 3.11.1 Total soluble sugars
Total soluble sugars were measured by anthrone reagent (0.2 %) following Sadasivam
and Manickam (1992). A reaction mixture (10 mL) consisting of 200 µL leaf extract (1 g leaf
homogenized in de-ionized water), 1,800 µL DI water and 8 mL anthrone reagent was heated

43
for 10 min in boiling water and cooled in ice bath to stop the reaction. Absorbance was
measured at 630 nm and total soluble sugar concentration (µg mL-1) was calculated using
glucose standard curve.
3.11.2 Proline content
Proline content was measured following Bates et al. (1973). A reaction mixture
consisting of leaf extract (1 g leaf homogenized with 3 % sulphosalicylic acid), ninhydrin
acid and glacial acetic acid were heated at 100 0C for 1 h in water bath. The reaction was
stopped by cooling in ice bath and absorbance was recorded at 520 nm after mixture
extraction with toluene. Proline content (µmol g-1) was calculated following standard curve of
L-proline.
3.12Enzymatic/non enzymatic antioxidant activity assays
Ascorbate peroxidase (APX) activity was determined by tracking the ascorbate
reduction through H2O2 with decrease in spectrophotometer absorbance at 290 nm (Nakano
and Asada, 1981). Two milliliters reaction mixture consisting of 20 µL crude leaf extract, 660
µL ascorbic acid solution, 660 µL potassium phosphate buffer (pH 7.0, 50 mM) and 660 µL
H2O2 was used to measure APX activity. Decrease in absorbance was monitored for 3 min
just after the addition of H2O2. Enzyme activity was calculated in the form of µmol ascorbate
min-1 g-1 leaf fresh weight. (Elavarthi and Martin, 2010)
3.13 NP analyses
3.13.1 Digestion
The method of Wolf (1982) was followed for digestion of the plant samples. Plant
sample (dried and ground) was taken in digestion tubes at 0.1 g. Two milliliters of
concentrated sulfuric acid were poured into the tubes. The sample was placed in laboratory at
room temperature and left over night. Then 1 mL H2O
2 (A.R. grade 35%) was added into
digestion tube by the side and was swiveled. The tube was heated up to 350 0C after
mounting it in a digestion block for 20 minutes. After cooling, 1 mL of H2O
2 was added and
again heated the tube for 20 minutes. Same procedure was continued awaiting colorlessness

44
of the material. Clear extract was made 50 mL by means of distilled water. The acquired
extract was stored and used for subsequent estimation of N, P and K as under:
3.13.3 Nitrogen determination
Kjeldhal’s method was used to determine the total nitrogen from the plant samples.
Five milliliter of digested sample was taken in Kjeldhal flask. Forty percent solution of
sodium hydroxide was added at 10 mL and the flask was connected to the Kjeldhal
distillation unit immediately and started distillation. Distillate was collected in 100 ml conical
flask already having 5 mL solution of 2% boric acid accompanied by some mixed indicator’s
drops. Distillation was stopped when the distillate was about 40-50 mL in the collecting flask
which was then titrated with standardized 0.01 N sulfuric acid. The titration volume was
recorded.
3.13.4 Phosphorus determination
Volumetric flask (50 mL capacity) was taken and 5 mL of the digested material was
added to it along with Barton Reagent at 10 mL. After making 50 mL total volume, the
sample was allowed to stay for half an hour. Readings of the blank, standards and samples
were recorded by spectrophotometer at 410-nm wavelength. Calibration curve was prepared
for standards and the phosphorus contents of the samples were determined from this
calibration curve.
Barton Reagent
The reagent was acquired by combining A and B solutions and making volume up to
1 L by following the procedure of Ashraf et al. (1992).
a) Solution A
Ammonium molybdate at 25 gram was dissolved in 400 mL of distilled water.
b) Solution B
Three hundred milliliter of boiling water was used to dissolve 1.25 grams of
ammonium metavenadate. After cooling the solution, HNO3
(concentrated) was added at 250
mL and again cooled at room temperature.

45
3.13.5 Potassium determination
Flame photometer (Jenway PFP-7) was used to determine potash from the extracted
samples. First of all, KCl standards of 2, 4, 6, 8 and ppm were prepared and then by plotting
the flame photometer reading against the respective K concentration there developed a
standard curve. Flame photometer readings of the samples were compared with calibration
curve and the actual K concentration in the samples were computed.
3.14 Characterization of the selected isolates
3.14.1 Root colonization assay
Surface sterilized seeds of maize and wheat inoculated with 3-day-old rhizobacterial
broth culture of the selected strains were sown in sterilized sand filled glass jars system
modified from Simon et al. (1996). The jars were sited under controlled conditions for 10
days in growth room at 25±1 0C. The sand was soaked with autoclaved half-strength
Hoagland solution. All the treatments were replicated four times. After 10 days the seedlings
were uprooted and roots were detached. To estimate root colonization by the selected strains,
one centimeter of root tip from each jar was cut off and placed in sterilized conical flask
having sterilized water. The flasks containing root suspension were shaken vigorously for 30
minutes in shaking incubator at 100 rpm. One milliliter material of each treatment was taken
in sterilized Petri plate and autoclaved Yeast Manitol Agar (YMA) media was spread onto it.
The Petri plates were incubated for 4 days at 28±1 0C. Bacterial colony forming units were
counted in each Petri plate by using colony counter.
3.14.2 Phosphate solubilization assay
The method of Mehta and Nautiyal (2001) was used to assess the potential of the
selected rhizobial strains to solubilize inorganic phosphate qualitatively. Phosphate growth
media i.e. NBRI-PBB (National Botanical Research Institute Phosphate Bromophenol Blue)
was used. A loop full of each culture (OD 0.5 (108
– 109
CFU mL-1
) was placed at three
places on petri plates and incubated for 7 days at 28 ± 1 °C. After seven days the colonies
showing clearing zone around them were characterized as positive for phosphate
solublization. Each treatment was repeated three times in the assay.

46
3.14.3 Exopolysaccharide (EPS) production assay
Broth culture (3-day-old) of the strains (OD 0.5 108
– 109
CFU mL-1
) was applied on
the Petri plates having RCV-glucose media (Ashraf et al., 2004) at five places. The plates
were incubated for four days at 28°C ± 1 after which the Petri plates were examined for
mucoid growth of the inoculated strains. Mucoid colonies were considered as positive in EPS
production.
3.14.4 Chitinase activity assay
The strains were characterized for chitinase activity following Chernin et al. (1998).
Yeast mannitol agar media containing 0.2% (w/v) chitin was prepared, autoclaved and poured
into Petri plates. Loop full of the broth culture of the selected strains (OD 0.5 108
– 109
CFU
mL-1
) was applied on Petri plates at five places. Petri plates were incubated for 5 days at 28 ±
1 °C. Halos formation around rhizobacterial colonies was judged and the strains showing
these halos were characterized as positive for chitinase activity.
3.15 Identification of selected strains
The selected strains were identified by using the BIOLOG® identification system
(Microlog System release 4.2 Biolog Inc., USA). The BIOLOG® identification system has
been found equally as reliable for identificstion as 16s RNA (Flores-Vargus and O, Hara,
2006). For identification on Biolog system the isolates were grown on Biolog universal
growth medium. Cells from the agar plates were separated with a sterile swab. The swab was
twirled and pressed against the inside surface of the tube on the dry glass above the fluid line
to break up clumps and release cells. The swab was moved up and down the wall of the tube
until the organisms mixed with the fluid and became a homogenous, clump free suspension.
The turbidity was adjusted to appropriate value for each isolate. This suspension was poured
into the multi-channel pipette reservoir. Eight sterile tips were fastened securely onto the 8-
channel repeating pipette. Tips were primed and 150 µL inoculum was poured into each well.
Micro plate was covered with its lid and incubated at 30 0C for 6 hours. After incubation,
micro plate was loaded to micro plate reader (Biolog) and identified putatively by microlog
software.

47
3.16 Statistical analysis
Treatment data were compared with ANOVA ( analysis of variance) according to completely
randomized design in pot trial and randomized complete block design for field trial using the
software SPSS version 18.0, ( Steel et al., 1996) and treatment means were compared using
Duncan’s Multiple Range Tests (p≤0.05) (Duncan, 1955). Selection of efficient rhizobacterial
strains was done by using the principal component scoring method using MIITAB software.

48
CHAPTER-4
RESULTS
When plants are subjected to abiotic stress such as soil salinity, their endogenous level of
hormones is disturbed which results in poor growth of plant. In the present study, we isolated
number of rhizobacterial strains from the rhizosphere of maize plants growing under salt-
affected soil conditions. These strains were tested for their ability to tolerate salinity stress by
using osmoadaptation assay. Out of these strains, most tolerant strains were selected and their
auxin production ability in the presence and absence of L-TRP was measured. Strains which
tolerated salinity and produced sufficient amounts of auxin were selected to test their ability
of growth promotion of maize seedlings under salt stressed axenic conditions. Two most
efficient strains (M4 and M7) were selected for further evaluation in pot and field trials to
improve the growth and yield of maize under salt stressed conditions.
4.1 Osmoadaptation assay
The ability of isolated rhizobacterial strains to produce auxin was tested by carrying
out the osmoadaptation assay. The selected strains were subjected to grow at four salinity
levels i.e. original, 10, 20 and 30 dS m-1 developed by using sodium chloride. The growth of
rhizobacterial strains was assessed by measuring the optical density (OD at 540 nm) after
three days of incubation. It was found from the results that all the strains have variable ability
to withstand salinity stress. Most of the strains were tolerant of salinity stress and did not
showed significant decrease in growth with increasing salinity. Maximum optical density at
highest salinity level was observed by the strain MA10 and followed by MA12 and MA9.

49
Table 3: Growth of rhizobacterial strains in broth culture after 3 days under salt stress
conditions
Strain Original 10 dS m-1 20 dS m-1 30 dS m-1
MA-1 1.21 ± 0.10* 0.89 ± 0.04 0.97 ± 0.02 0.97 ± 0.23
MA-2 1.26 ± 0.20 1.05 ± 0.06 1.00 ±0.12 0.75 ± 0.12
MA-3 1.08 ± .01 1.11 ± 0.01 0.98 ± 0.12 1.00 ± 0.08
MA-4 1.00 ± 0.08 1.11 ± 0.36 1.08 ± 0.02 0.78 ± 0.04
MA-5 0.70 ± 0.03 1.40 ± 0.12 0.70 ± 0.05 0.87 ± 0.03
MA-6 0.81 ±0.03 0.94 ± 0.05 0.89 ± 0.04 0.83 ± 0.07
MA-7 0.97 ±0.02 0.85 ± 0.02 0.84 ± 0.17 0.77 ± 0.26
MA-8 0.63 ±0.01 1.14 ± 0.06 0.93 ± 0.11 0.78 ± 0.17
MA-9 0.52 ± 0.01 0.64 ± 0.12 0.40 ± 0.15 0.63 ± 0.21
MA-10 0.87 ± 0.07 0.51 ± 0.03 0.50 ± 0.03 0.68 ± 0.02
MA-11 1.05 ± 0.04 0.62 ± 0.21 0.69 ± 0.02 0.79 ± 0.15
MA-12 0.95 ± 0.03 0.60 ± 0.20 0.64 ± 0.03 0.75 ± 0.01
MA-13 1.48 ± 0.04 1.38 ±0.06 1.04 ± 0.04 0.98 ± 0.23
MA-14 1.43 ± 0.06 0.77 ± 0.08 1.50 ± 0.23 0.65 ± 0.08
MA-15 1.04 ± 0.09 0.50 ± 0.03 1.02 ± 0.14 1.25 ± 0.01
Values are means of three replicates ± Standard error (SE)
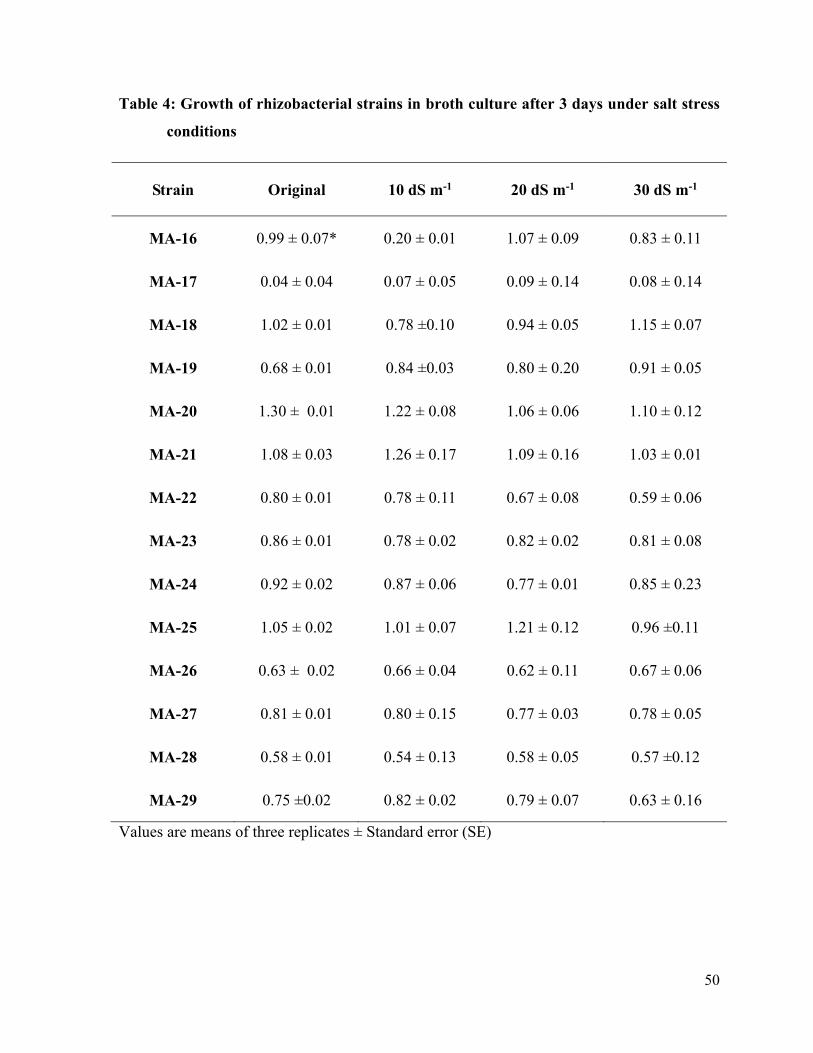
50
Table 4: Growth of rhizobacterial strains in broth culture after 3 days under salt stress
conditions
Strain Original 10 dS m-1 20 dS m-1 30 dS m-1
MA-16 0.99 ± 0.07* 0.20 ± 0.01 1.07 ± 0.09 0.83 ± 0.11
MA-17 0.04 ± 0.04 0.07 ± 0.05 0.09 ± 0.14 0.08 ± 0.14
MA-18 1.02 ± 0.01 0.78 ±0.10 0.94 ± 0.05 1.15 ± 0.07
MA-19 0.68 ± 0.01 0.84 ±0.03 0.80 ± 0.20 0.91 ± 0.05
MA-20 1.30 ± 0.01 1.22 ± 0.08 1.06 ± 0.06 1.10 ± 0.12
MA-21 1.08 ± 0.03 1.26 ± 0.17 1.09 ± 0.16 1.03 ± 0.01
MA-22 0.80 ± 0.01 0.78 ± 0.11 0.67 ± 0.08 0.59 ± 0.06
MA-23 0.86 ± 0.01 0.78 ± 0.02 0.82 ± 0.02 0.81 ± 0.08
MA-24 0.92 ± 0.02 0.87 ± 0.06 0.77 ± 0.01 0.85 ± 0.23
MA-25 1.05 ± 0.02 1.01 ± 0.07 1.21 ± 0.12 0.96 ±0.11
MA-26 0.63 ± 0.02 0.66 ± 0.04 0.62 ± 0.11 0.67 ± 0.06
MA-27 0.81 ± 0.01 0.80 ± 0.15 0.77 ± 0.03 0.78 ± 0.05
MA-28 0.58 ± 0.01 0.54 ± 0.13 0.58 ± 0.05 0.57 ±0.12
MA-29 0.75 ±0.02 0.82 ± 0.02 0.79 ± 0.07 0.63 ± 0.16
Values are means of three replicates ± Standard error (SE)

51
4.2 Auxin production assay
The data in Table 5 shows auxin production ability of the rhizobacterial strains. It is
evident from the data that all rhizobacterial strains were able to produce auxin (IAA
equivalents) to varying degrees, both in the presence and absence of L-TRP. However, the
auxin production was enhanced by applying the precursor L-TRP.
Rhizobacterial strain MA-11 produced maximum IAA equivalents when
supplemented with L-TRP. The minimum auxin production was recorded by strain MA-21.
The data also suggested that these strains were not over auxin producing, but produced
moderate amounts of IAA equivalents.
After conducting auxin production assay, eleven strains were selected for further screening in
growth room (axenic) experiment. The selected strains were renamed as follows.
MA1 → M1
MA2 → M2
MA3 → M3
MA4 → M4
MA5 → M5
MA7 → M6
MA11 → M7
MA13 → M8
MA14 → M9
MA19 → M10
MA20 → M11

52
Table 5: Amount of IAA equivalents (µg mL-1) produced by the selected rhizobacterial
strains
Strains With
L-TRP
Without
L-TRP Strains
With
L-TRP
Without
L-TRP
MA-1 9.23 d* 6.11 gh MA-14 5.56 a 3.09 op
MA-2 7.25 b 5.61 lm MA-15 4.61 c 0.53 q
MA-3 10.19 g 7.07 jk MA-18 2.25 j 2.02 k
MA-4 13.26 g 7.19 g MA-19 5.12 b 2.08 jk
MA-5 6.76 e 5.68 i MA-20 4.61 c 1.63 l
MA-6 2.53 i 2.48 i MA-21 1.18 no 0.91 p
MA-7 8.13 d 4.53 f MA-23 2.95 h 1.40 mn
MA-8 3.07 gh 0.98 op MA-24 2.06 jk 1.11 op
MA-11 19.19 g 12.08 jk MA-25 3.25 g 1.45 lm
MA-13 6.12 b 3.52 q
Means sharing similar letter(s) do not differ significantly

53
4.3 Jar trial (Growth room study)
The selected rhizobacterial strains which were found having the ability of auxin
production were further screened to access their ability to promote growth of maize seedlings
under salt affected axenic conditions. The results of the experiment showed that salinity had a
repressive effect on the growth of maize but inoculation with auxin producing PGPR strains
improved the growth of maize with varying degree.
4.3.1 Shoot length
The Figure 1 demonstrates the effect of salinity and inoculation with rhizobacterial
strains on the shoot length of maize. It was found that shoot length significantly decreased
with increasing salinity levels. Maximum decrease in shoot length was recorded at EC 15 dS
m-1. At this level maximum increase in shoot length was observed by the strain M4 followed
by strain M7 both in the absence and presence of L-TRP as compared to their respective
uninoculated control. At EC 4, 8, and 12 dS m-1 strain M4 was the best performer regarding
improvement in shoot length, where it caused 88.12, 57.67 and 45.43% increase in the
absence and 49.89, 56.71 and 51.73% increase in the presence of L-TRP as compare to their
uninoculated controls. Other strains were also effective in improving the shoot length but
most of them showed non-significant increases in shoot length as compared to their control.
Under normal conditions all the strains improved the shoot length both in the absence
and presence of L-TRP. However along with L-TRP application the performance of strains
was enhanced. Under these conditions strain M6 was among the front runners in improving
the shoot length in the absence and presence of L-TRP. Strains M2, M3, M5, M8, M9, M10
and M11 in the absence of L-TRP showed non-significant results as compared to
uninoculated control at normal conditions.
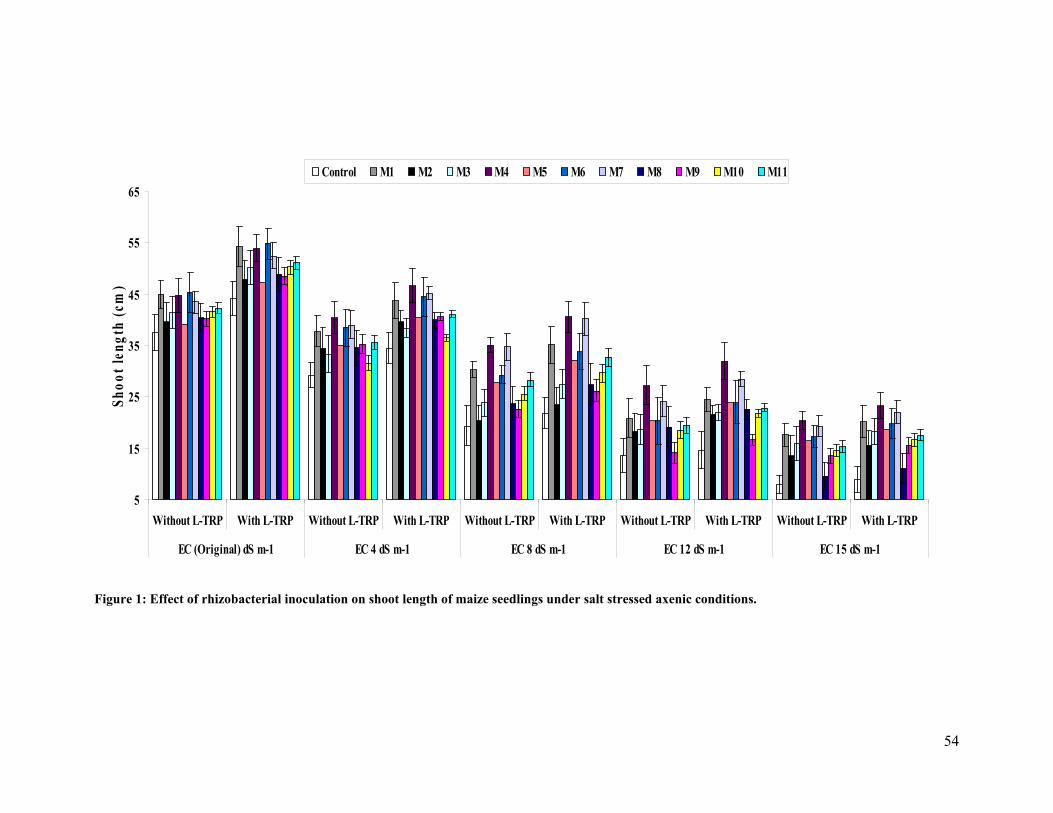
54
5
15
25
35
45
55
65
Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP
EC (Original) dS m-1 EC 4 dS m-1 EC 8 dS m-1 EC 12 dS m-1 EC 15 dS m-1
Shoo
t le
ngth
(cm
)
Control M1 M2 M3 M4 M5 M6 M7 M8 M9 M10 M11
Figure 1: Effect of rhizobacterial inoculation on shoot length of maize seedlings under salt stressed axenic conditions.

55
4.3.2 Root length
Figure 2 demonstrate the results regarding root length of maize seedlings. A marked
reduction in root length of maize seedlings subjected to salinity stress was recorded. A 22, 32,
35 and 55% decrease in root length was observed at EC 4, 8, 12 and 15 dS m-1. However,
rhizobacterial inoculation was found effective in mitigating the deleterious effects of salinity
and caused significant improvements in root length at all salinity levels.
At 15 dS m-1, strain M4 performed better than the rest of the strains and caused 57 and
69% increase in root length in the absence and presence of L-TRP, respectively, over
uninoculated control. Most of the other strains showed non-significant increases in the root
length. At EC 4, 8 and 12 dS m-1, strains M4, M7 and M1 were the leaders in enhancing the
root length both in the presence and absence of L-TRP. Under normal conditions, strains M1
and M3 performed better than the rest of the strains. In the absence of L-TRP, M2, M6 and
M10 showed non-significant increases in root length as compared to uninoculated control.
4.3.3 Shoot dry weight
The Figure 3 shows that salinity significantly reduced the shoot dry weight at all
salinity levels. At EC 15 dS m-1 maximum reduction in shoot dry weight was recorded.
Rhizobacterial inoculation was found effective in improving the shoot dry weight to varying
degrees, but most of the strains showed non-significant increases in shoot dry weight.
Maximum increase in shoot dry weight was recorded by strain M4 both in the absence and
presence of L-TRP application, respectively, followed by strain M7. In the presence of L-
TRP strain M4 also performed better than the rest of the strains at other salinity levels and
caused 25, 36 and 49% increase in shoot dry weight of maize seedlings at 4, 8 and 12 dS m-1,
respectively, as compared to respective uninoculated control.
Under normal conditions all the strains significantly improved the shoot dry weight
both in the absence and presence of L-TRP. Strains M1, M4 and M6 were among the leaders
in improving the shoot dry weight of maize under normal conditions. Maximum increase in
this case was recorded by strain M6 which caused 45 and 52% increase in shoot dry weight of
maize seedlings over its respective uninoculated control.

56
0
10
20
30
40
50
Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP
EC (Original) dS m-1 EC 4 dS m-1 EC 8 dS m-1 EC 12 dS m-1 EC 15 dS m-1
Roo
t le
ngt
h p
er p
lan
t (c
m)
Control M1 M2 M3 M4 M5 M6 M7 M8 M9 M10 M11
Figure 2: Effect of rhizobacterial inoculation on root length of maize seedlings under salt stressed axenic conditions.

57
0.05
0.15
0.25
0.35
0.45
0.55
0.65
Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP
EC (Original) dS m-1 EC 4 dS m-1 EC 8 dS m-1 EC 12 dS m-1 EC 15 dS m-1
Shoo
t dr
y pe
r pl
ant
(g)
Control M1 M2 M3 M4 M5 M6 M7 M8 M9 M10 M11
Figure 3: Effect of rhizobacterial inoculation on shoot dry weight of maize seedlings under salt stressed axenic conditions.

58
4.3.4 Root dry weight
It is evident from the Figure 4 that salinity stress had a repressive effect on root dry
weight and it declined with increasing salinity levels. Inoculation with different rhizobacterial
strains improved the root dry weight to varying levels at all tested salinity levels. Under
normal conditions maximum increase in root dry weight was recorded with strain M6 both in
the absence and presence of L-TRP. This strain was followed by strain M1 and M4. Strain
M9 was found least effective among all the strains in improving the root dry weight of maize
seedlings.
Maximum reduction in root dry weight was observed at the highest salinity level i.e.
15 dS m-1. At this level, strain M7 was found most effective in enhancing the root dry weight
of maize seedlings which caused 45 and 57% increase in the absence and presence of L-TRP,
respectively, as compared to its uninoculated control. Strain M7 was followed by strain M4,
M1 and M6 regarding improvement in root dry weight. Rest of the strains showed non-
significant increase in root dry weight as compared to uninoculated control.
4.3.5 Total plant dry weight
Total plant dry weight significantly decreased with increasing salinity levels (Table
6). At 15 dS m-1, maximum increase in total plant dry weight was observed by the strains M4
and M7. At EC 4, 8, and 12 dS m-1, strain M4 performed better than rest of the strains
regarding improvement in total plant dry weight. Other strains were also effective in
improving the total plant dry weight but most of them showed non-significant increases in
total plant dry weight as compared to their control.
Under normal conditions all the strains improved the total plant dry weight both in the
absence and presence of L-TRP. However along with L-TRP application the performance of
strains was enhanced. Under these conditions strain M6 was among the front runners to
improve the total plant dry weight in the absence as well as in the presence of L-TRP.

59
0.10
0.20
0.30
0.40
0.50
0.60
Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP Without L-TRP With L-TRP
EC (Original) dS m-1 EC 4 dS m-1 EC 8 dS m-1 EC 12 dS m-1 EC 15 dS m-1
Roo
t dr
y pe
r pl
ant
(g)
Control M1 M2 M3 M4 M5 M6 M7 M8 M9 M10 M11
Figure 4: Effect of rhizobacterial inoculation on root dry weight of maize seedlings under salt stressed axenic conditions.

60
Bacterial strains
Total plant dry weight (g)
Normal (1.6 dS m-1) 4 dS m-1 8 dS m-1 12 dS m-1 15 dS m-1
L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+)
Control 0.61±0.013 0.66±0.010 0.47±0.017 0.51±0.012 0.32±0.009 0.37±0.019 0.26±0.032 0.30±0.008 0.20±0.063 0.23±0.059
M1 0.92±0.018 1.03±0.015 0.74±0.063 0.86±0.059 0.53±0.060 0.63±0.005 0.43±0.003 0.48±0.015 0.36±0.032 0.41±0.010
M2 0.79±0.007 0.88±0.026 0.61±0.004 0.71±0.003 0.46±0.008 0.54±0.026 0.40±0.017 0.44±0.009 0.25±0.001 0.28±0.019
M3 0.84±0.032 0.94±0.009 0.57±0.015 0.66±0.032 0.43±0.029 0.52±0.017 0.34±0.004 0.38±0.059 0.24±0.029 0.27±0.003
M4 0.99±0.011 1.11±0.060 0.84±0.017 0.96±0.053 0.61±0.063 0.72±0.012 0.49±0.008 0.56±0.001 0.41±0.032 0.46±0.015
M5 0.90±0.011 1.01±0.017 0.71±0.019 0.81±0.004 0.53±0.017 0.63±0.009 0.33±0.009 0.36±0.019 0.26±0.060 0.29±0.004
M6 1.05±0.027 1.17±0.059 0.76±0.003 0.88±0.017 0.52±0.059 0.61±0.008 0.42±0.004 0.48±0.017 0.35±0.009 0.39±0.017
M7 0.91±0.063 1.01±0.060 0.78±0.017 0.90±0.026 0.55±0.017 0.66±0.032 0.48±0.029 0.54±0.012 0.42±0.001 0.47±0.012
M8 0.84±0.004 0.94±0.032 0.66±0.005 0.76±0.010 0.50±0.001 0.59±0.053 0.39±0.060 0.43±0.059 0.27±0.010 0.31±0.059
M9 0.73±0.059 0.81±0.001 0.61±0.017 0.70±0.019 0.43±0.008 0.51±0.012 0.34±0.019 0.39±0.005 0.27±0.009 0.30±0.005
M10 0.76±0.015 0.85±0.004 0.55±0.008 0.63±0.029 0.42±0.017 0.50±0.060 0.36±0.008 0.40±0.063 0.28±0.026 0.31±0.029
M11 0.79±0.010 0.88±0.026 0.63±0.003 0.72±0.001 0.46±0.015 0.54±0.026 0.35±0.003 0.40±0.017 0.27±0.009 0.30±0.012
Table 6: Effect of rhizobacterial inoculation on total plant dry weight of maize seedlings under salt stressed axenic conditions.

61
Bacterial strains
Shoot root ratio
Normal (1.6 dS m-1) 4 dS m-1 8 dS m-1 12 dS m-1 15 dS m-1
L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+) L-TRP (-) L-TRP (+)
Control 1.90 2.01 2.23 2.34 2.07 1.66 1.92 1.60 1.87 1.77
M1 1.41 1.47 1.61 1.61 1.46 1.29 1.39 1.33 1.57 1.60
M2 1.89 1.96 2.06 2.06 1.72 1.45 2.88 2.77 2.66 2.71
M3 1.30 1.35 2.08 2.08 2.36 2.05 2.59 2.43 2.99 3.05
M4 1.47 1.53 1.55 1.55 1.55 1.35 1.49 1.43 1.57 1.59
M5 1.47 1.53 1.64 1.64 1.67 1.52 1.49 1.42 2.82 2.88
M6 1.46 1.52 1.64 1.64 1.55 1.35 1.37 1.31 1.65 1.68
M7 1.46 1.52 1.53 1.53 1.64 1.44 1.41 1.35 1.58 1.61
M8 1.88 1.96 2.57 2.58 1.64 1.38 1.90 1.82 1.25 1.28
M9 1.37 1.42 1.58 1.58 1.47 1.32 1.10 1.06 1.69 1.73
M10 1.97 2.05 1.52 1.52 1.53 1.32 1.82 1.74 1.67 1.70
M11 1.68 1.75 1.82 1.82 2.69 2.38 2.70 2.56 2.54 2.60
Table 7: Effect of rhizobacterial inoculation on shoot root ratio of maize seedlings under salt stressed axenic conditions.

62
Figure S2: Pictorial representation of rhizobacterial inoculation at different salinity levels under axenic conditions

63
4.4 Pot trial
4.4.1 Plant height
The results of the pot trial revealed that salinity had a repressive effect on plant height
(Figure 5), regardless of inoculation treatment. It was evident from the results that with
increasing salinity levels there was progressive decline in plant height. However,
rhizobacterial inoculation with M4, M7 or both seemed promising in alleviating the salinity
stress which was reflected in case of improvement in plant height. Rhizobacterial inoculation
in single and dual combinations improved plant height at all salinity levels. The Effect of
rhizobacterial inoculation was pronounced when strains were tested along with L-TRP
application.
Salinity stress significantly reduced the plant height at all levels. Maximum reduction
in plant height (71%) was noted at 15 dS m-1. In case of single strain inoculation strain M4
performed better than strain M7; M4 and M7 increased plant height up to 51 and 27%,
respectively, over their respective uninoculated controls. When both these strains were tested
in the presence of L-TRP, further increase in plant height was recorded i.e. M4 and M7
increased up to 54 and 29%, respectively, over control. However, combined application of
strain M4 and M7 gave the best results regarding the improvement in plant height at all
salinity levels both in the presence and absence of L-TRP application. At 15 dS m-1, an
increase up to 74% by M4 and M7 was recorded, in the absence and presence of L-TRP,
respectively, over uninoculated control regardless of L-TRP.

64
ik
m
r
s
ghj
m
r
s
d-g d-gi
m
q
d degh
n
q
f-h gh
k
n
r
d-f e-g
j
o
r
c
a
c
l
p
ba
c
m
p
0
20
40
60
80
100
120
140
160
180
200
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Pla
nt H
eigh
t (c
m)
Control M4 M7 M4XM7
Figure: 5 Effect of rhizobacterial inoculation (with and without L-TRP) on plant height of maize at different salinity levels (original = 1.63,
4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of
three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

65
4.4.2 Fresh biomass
The data (Figure 6) about the Effect of rhizobacterial inoculation on fresh biomass of
maize grown under salt affected soil conditions revealed that salinity had adverse effect on
the fresh biomass and it decreased with increasing salinity. However, improvement in fresh
biomass was observed due to inoculation with rhizobacterial strains.
Sole inoculation was effective under normal and at all salinity levels. Maximum
reduction (45%) in fresh biomass was recorded at highest salinity level. Maximum increase in
fresh biomass at salinity 15 dS m-1 was observed by strain M4 in the presence and absence of
L-TRP and that increase was 39 and 31%, respectively, over its respective uninoculated
control. Inoculation with stain M7 gave non-significant results with control.
Co-inoculation of M4xM7 performed even better than sole inoculation of strain M4
and M7 at all salinity levels. At the highest salinity level i.e.15 dS m-1 an increase of 35 and
33% in fresh biomass as compared to respective uninoculated control was recorded by
inoculation with M4xM7 in the absence and presence of L-TRP application, respectively. The
results of co-inoculation were statistically non-significant as compared to single inoculation
treatments. Co-inoculation was also effective at lower salinity levels and under normal soil
conditions. At 12 dS m-1, increase in fresh biomass due to co-inoculation with M4xM7 was
46 and 43%, over respective uninoculated control when it was tested with and without L-TRP
application, respectively. At normal soil conditions, combination M4xM7 resulted 22.17%
increase in the absence of L-TRP and 34% increase in fresh biomass over respective
uninoculated control along with L-TRP application.
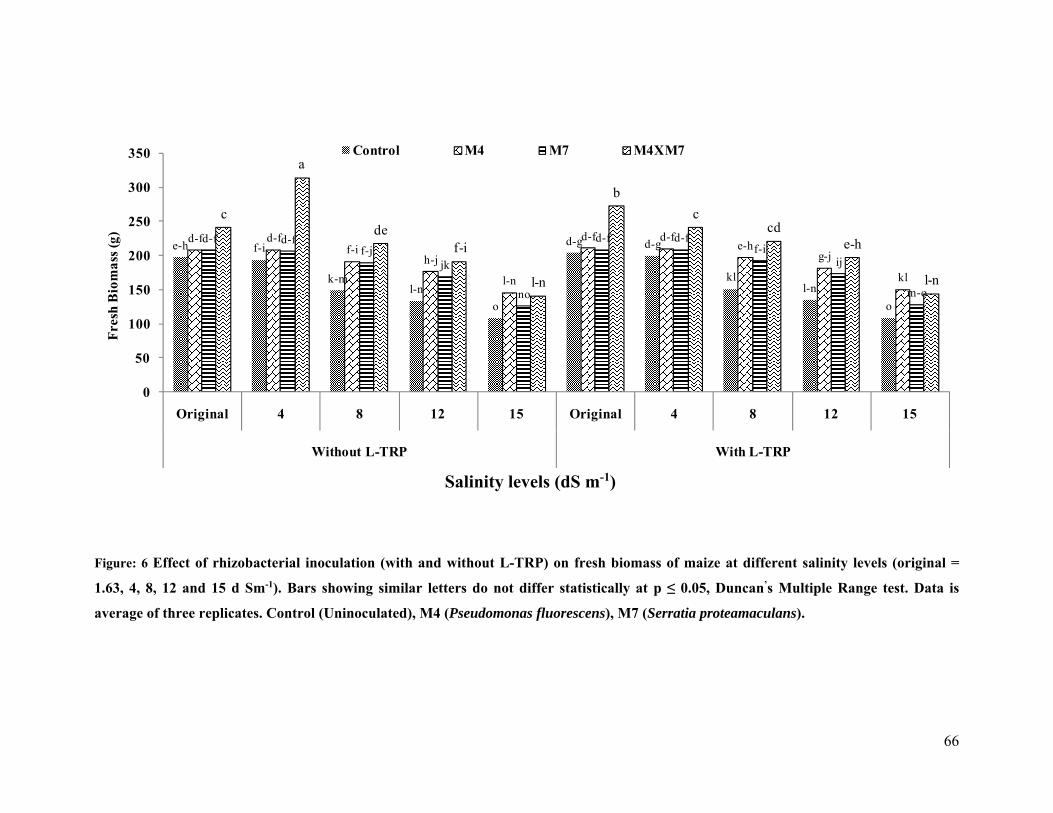
66
e-h f-i
k-ml-n
o
d-g d-g
kll-n
o
d-f d-ff-i
h-j
l-n
d-f d-fe-h
g-j
kl
d-f d-ff-j
jk
no
d-f d-ff-i
ij
m-o
c
a
def-i
l-n
b
ccd
e-h
l-n
0
50
100
150
200
250
300
350
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Fre
sh B
iom
ass
(g)
Control M4 M7 M4XM7
Figure: 6 Effect of rhizobacterial inoculation (with and without L-TRP) on fresh biomass of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

67
4.4.3 Root length
The results regarding root length (Figure 7) demonstrate that soil salinity
progressively decreased the root length with increasing soil salinity level, however, the
rhizobacterial inoculation with and without L-TRP application was found effective in
reducing the adverse effects of salinity on root length. At EC 15 dS m-1, 75% reduction in root
length was recorded. Sole inoculation with strain M4 without L-TRP caused 81% increase in
root length over respective uninoculated control at EC 15 dS m-1.
Inoculation with strain M4 Along with L-TRP application caused 6, 30, 35 and 39%
increase in root length at 4, 8, 12 and 15 dS m-1, respectively. Sole inoculation with strain M7
also significantly improved root length at all salinity levels. Combined application of M4 and
M7 (M4xM7) performed better than their sole inoculation at both L-TRP applications. An
increase of 33 and 35% in root length at 15 dS m-1 was recorded by the inoculation of
M4xM7 in presence and absence of L-TRP application, respectively.

68
lmn
op
tu
v
klm
no
tu
v
e-gg-i
jk
q
tu
b-dd-f
ij
p
tu
ghij
k
qr
u
c-fg-i
k
q
tu
b bc
fg
mn
st
a a
b-e
n
rs
0
10
20
30
40
50
60
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Roo
t Len
gth
(cm
)
Control M4 M7 M4XM7
Figure: 7 Effect of rhizobacterial inoculation (with and without L-TRP) on root length of maize at different salinity levels (original = 1.63,
4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of
three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
.
Salinity levels (dS m-1)

69
Figure S3: Pictorial representation of rhizobacterial inoculation on root growth at different salinity levels in pot experiment.

70
4.4.4 Root weight
Data regarding the root weight (Figure 8) reveal that soil salinity progressively
decreased the root weight with increasing soil salinity level. However, the rhizobacterial
inoculation effectively improved the root weight under saline conditions. Inoculation
performed even better when it was tested along with L-TRP application. At EC 15 dS m-1,
up to 69% reduction in root weight was recorded. Sole inoculation with strain M4, without
L-TRP application caused 44% increase in root length over respective uninoculated
control at EC 15 dS m-1. L-TRP application alone also improved the root weight.
In the presence of L-TRP application, inoculation with stain M4 showed up to
32% increase in root weight at EC 15 dS m-1. Similar improvement in root weight was
also observed at other salinity levels. Sole inoculation with strain M7 also significantly
improved root weight at all salinity levels both with and without L-TRP application. As
compared with single strain inoculation, combined application of M4 and M7 (M4xM7)
performed better at both L-TRP applications. An increase up to 58 and 78% in root weight
at 15 dS m-1 was recorded by M4xM7 in presence and absence of L-TRP application,
respectively, over their respective uninoculated controls.
4.4.5 Grain yield
Figure 9 demonstrates, that the rhizobacterial inoculation with and without L-TRP
application was found effective in improving the grain yield of maize under salt stress. It
is evident from the data that salinity adversely reduced the grain yield. At EC 15 dS m-1 no
grain formation was observed. At 12 dS m-1, 83% reduction in grain yield was recorded.
Sole inoculation with strain M4 without L-TRP caused 20.6, 31.1 and 41% increase in
grain yield over respective uninoculated control at EC 4, 8 and 12 dS m-1, respectively.
Along with L-TRP application, up to 33% increase in grain yield was recorded by
inoculation with strain M4 at 12 dS m-1. In case of single strain inoculation strain M4
performed better than M7. At both L-TRP applications, combined application of M4 and
M7 (M4xM7) performed better than their sole inoculation. An 89 and 77% increase in
grain yield at 12 dS m-1 was recorded by M4xM7 in presence and absence of L-TRP
application respectively, over their respective uninoculated controls.

71
ijk
mn
st
u
gh
i-k
lm
rstu
cdcd
h-j
n-p
st
bccd
g-i
m-o
q-s
deef
jk
o-q
st
cdde
i-k
n-p
st
ab a
fg
lm
p-r
a a
de
l
o-q
0
5
10
15
20
25
30
35
40
45
50
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Roo
t wei
ght
(g)
Control M4 M7 M4XM7
Figure: 8 Effect of rhizobacterial inoculation (with and without L-TRP) on root weight of maize at different salinity levels (original = 1.63,
4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of
three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

72
ghi
j
o
fghi
j
o
c-e c-efg
m
cd cdde
l
de egh
n
c-e c-egh
mn
aba
c
l
ab ab
k
0
10
20
30
40
50
60
70
80
90
100
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Gra
in Y
ield
(g)
Control M4 M7 M4XM7
Figure 9: Effect of rhizobacterial inoculation (with and without L-TRP) on grain yield of maize at different salinity levels (original = 1.63, 4,
8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of
three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)
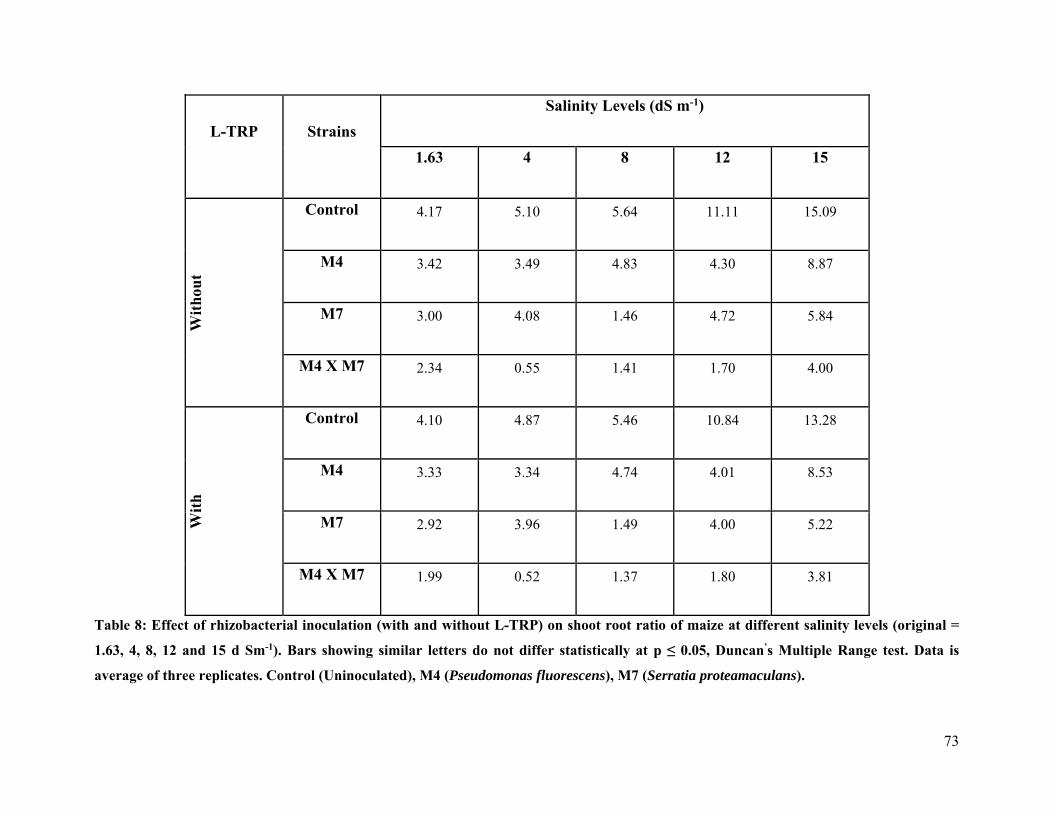
73
L-TRP Strains
Salinity Levels (dS m-1)
1.63 4 8 12 15
Wit
hou
t Control 4.17 5.10 5.64 11.11 15.09
M4 3.42 3.49 4.83 4.30 8.87
M7 3.00 4.08 1.46 4.72 5.84
M4 X M7 2.34 0.55 1.41 1.70 4.00
Wit
h
Control 4.10 4.87 5.46 10.84 13.28
M4 3.33 3.34 4.74 4.01 8.53
M7 2.92 3.96 1.49 4.00 5.22
M4 X M7 1.99 0.52 1.37 1.80 3.81
Table 8: Effect of rhizobacterial inoculation (with and without L-TRP) on shoot root ratio of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).

74
4.4.6 1000-grain weight
The results showing the effect of salinity and rhizobacterial inoculation on 1000-grain
weight of maize are presented in the Figure 10. The data demonstrates that soil salinity
progressively decreased the 1000-grain weight with increasing soil salinity level, however the
rhizobacterial inoculation with and without L-TRP application improved the 1000-grain
weight. At EC 12 dS m-1, 51% reduction in 1000-grain weight was recorded. In the absence
of L-TRP, sole inoculation with strain M4 caused up to 8, 22 and 37% increase in 1000-grain
weight over respective uninoculated control at EC 4, 8 and 12 dS m-1, respectively.
The results of inoculation improved when it was carried out along with L-TRP
application. In the presence of L-TRP strain M4 caused 10, 21 and 34% increase in 1000-
grain weight at 4, 8 and 12 dS m-1, respectively. Both in the presence and absence of L-TRP,
sole inoculation with M7 also positively improved 1000-grain at all salinity levels. Combined
application of M4 and M7 (M4xM7) performed better than their sole inoculation at both L-
TRP applications. Up to 59 and 57% increase in 1000-grain at 12 dS m-1 was recorded by
M4xM7 in presence and absence of L-TRP application, respectively.
4.4.7 Cob biomass
Figure 11 shows the effects of salinity and rhizobacterial inoculation on cob biomass
of maize. The data demonstrates that soil salinity caused a significant reduction in the cob
biomass however, the rhizobacterial inoculation significantly increased the cob biomass.
Combined application of both strains showed better results than their sole inoculation. The
effect of inoculation was enhanced when it was used along with L-TRP application. Only L-
TRP application also positively improved the cob biomass. At EC 12 dS m-1, 8% reduction in
cob biomass was recorded. Sole inoculation with strain M4 without L-TRP caused up to 6, 10
and 38% increase in cob biomass over respective uninoculated control at EC 4, 8 and 12 dS
m-1, respectively.
At 4, 8 and 12 dS m-1, M4 strain along with L-TRP application caused up to 7, 10 and
39% increases in cob biomass, respectively. Sole inoculation with M7 also positively
improved cob biomass at all salinity levels both with and without L-TRP application. M4 and

75
h-k j-l
pq
r
h-k i-l
p
r
eff-h
mn
pq
c-efg
m
pq
efg-j
m-o
q
de
g-i
m-o
q
abcd
l
no
a a
kl
o
0
50
100
150
200
250
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
1000
Gra
in W
eigh
t (g
)
Control M4 M7 M4XM7
Figure 10: Effect of rhizobacterial inoculation (with and without L-TRP) on 1000-grain weight of maize at different salinity levels
(original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

76
fgh-j
k
m
cd deg-i
k
de efh-j
k
cdef
ij
l
defg
j
l
a ac
f-h
ap
cd
g-j
c cdg-i
k
0
20
40
60
80
100
120
140
160
180
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Cob
bio
mas
s (g
)
Control M4 M7 M4XM7
Figure: 11 Effect of rhizobacterial inoculation (with and without L-TRP) on cob biomass of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

77
M7 applied in combination (M4xM7) showed better results than their sole inoculation at
both L-TRP applications. An increase up to 55 and 57% in cob biomass at 12 dS m-1 was
recorded by M4xM7 in presence and absence of L-TRP application, respectively.
4.4.8 Chlorophyll contents (SPAD Values)
The results regarding chlorophyll contents (Figure 12) indicate that the salinity
significantly reduced the chlorophyll contents at all levels. However inoculation/co-inoculation
reduced the effects of salinity and improved the chlorophyll contents both in the presence and
absence of L-TRP application. At 4, 8, 12 and 15 dS m-1, a significant decrease of 8, 23, 24 and
26%, respectively, in chlorophyll contents was recorded as compared with uninoculated controls.
Single inoculation improved the chlorophyll contents at all tested salinity levels both in
the presence and absence of L-TRP. In the absence of L-TRP, stain M4 performed better than
M7 and increased the chlorophyll contents up to 17, 35, 11 and 14% at 4, 8, 12 and15 dS m-1
respectively, over its respective uninoculated control. Maximum increase in chlorophyll contents
in the absence of L-TRP was recorded by coinoculation with M4XM7 which gave up to 34.86,
64.74, 30.84 and 10.58% increase at 4, 8, 12, and 15 dS m-1 respectively, over its respective
uninoculated control. Separate application of L-TRP also caused positive improvement in
chlorophyll contents but these results were non-significant as compared to uninoculated control.
Along with L-TRP application co-inoculation performed better than sole inoculation with
M4 and M7. Maximum increase in chlorophyll contents was recorded up to 24, 55, 28 and 13%
at 4, 8, 12 and 15 dS m-1, respectively, by M4xM7 over uninoculated control. Single inoculation
with M4 and M7 also significantly improved the chlorophyll contents at all salinity levels as
compared with uninoculated control. The results of M4 and M7 inoculation along with L-TRP
were statistically non-significant as compared to one another at all tested salinity levels except at
EC 15 dS m-1.
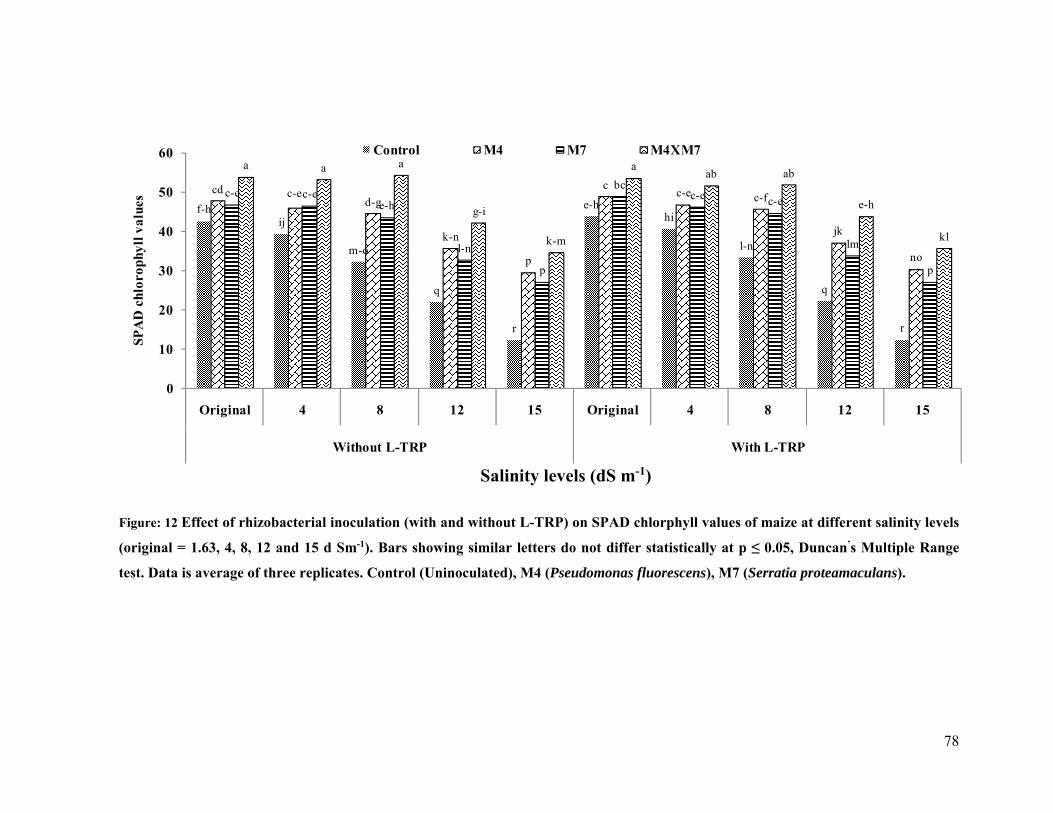
78
f-hij
m-o
q
r
e-hhi
l-n
q
r
cd c-ed-g
k-n
p
cc-e c-f
jk
no
c-e c-ee-h
l-n
p
bcc-e c-e
lm
p
a a a
g-i
k-m
aab ab
e-h
kl
0
10
20
30
40
50
60
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
SPA
D c
hlor
ophy
ll v
alue
s Control M4 M7 M4XM7
Figure: 12 Effect of rhizobacterial inoculation (with and without L-TRP) on SPAD chlorphyll values of maize at different salinity levels
(original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

79
4.4.9 Electrolyte leakage
The effect of inoculation/combined inoculation on the electrolyte leakage of maize
grown under salt affected soil conditions is shown in Figure 13. The data demonstrate that
electrolyte leakage increased due to exposure to salinity. At EC 15 dS m-1, up to 120%
increase in electrolyte leakage was noted, as compared with uninoculated control.
Electrolyte leakage decreased due to single inoculation/coinoculation with
rhizobacterial strains both in the absence and presence of L-TRP. However, inoculation/co-
inoculation gave better results when it was carried out along with L-TRP application.
Maximum reduction in electrolyte leakage was recorded by coinoculation with M4xM7 that
was up to 39, 37, 38 and 82% at 4, 8, 12 and 15 dS m-1, respectively over respective un-
inoculated control. Along with L-TRP application sole inoculation also significantly reduced
the electrolyte leakage at all salinity levels. As compared with strain M7 and M4 showed
greater reduction in electrolyte leakage. The results of single inoculation were statistically
non significant with each other.
Inoculation/coinoculation in the absence of L-TRP application significantly reduced
the electrolyte leakage. In this case M4xM7 combination gave maximum reduction in
electrolyte leakage that was up to 40, 35, 40 and 43% at 4, 8, 12 and 15 dS m-1, respectively
as compared with uninoculated control. L-TRP application as alone treatment was also found
effective in reducing the electrolyte leakage at all salinity levels.

80
k-m
hi
ef
b
a
o-q
jk
ji
c
a
rs
n-pk-m
hi
e
t
p-rm-o
i-j
ef
qr
k-n k-m
hi
c
t
p-rl-o
f-h
d
trs
n-p
jk
gh
u
tpr
k-n
hi
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Ele
ctro
lyte
leak
age
(dS/
m)
Control M4 M7 M4XM7
Figure 13: Effect of rhizobacterial inoculation (with and without L-TRP) on electrolyte leakage of maize at different salinity levels (original
= 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

81
4.4.10 Photosynthetic rate
Photosynthetic rate was adversely affected by the soil salinization (Figure 14).
However inoculation/coinoculation was found promising in improving the photosynthetic rate
of maize plants. Application of L-TRP further enhanced the efficiency of inoculation. Up to
73% decrease in photosynthetic rate was noted at EC 15 dS m-1. Similar decrease in
photosynthetic rate was also observed at other salinity levels.
In the presence of L-TRP, M4xM7 caused maximum increase up to 143% in
photosynthetic rate at EC 15dS m-1. Single inoculation with M7 in the presence of L-TRP
caused up to 57, 92, 86 and 98% increase at 4, 8, 12 and 15dS m-1 respectively, as compared
to respective uninoculated control. Single inoculation/coinoculation in the absence of L-TRP
also significantly improved the photosynthetic rate but combined inoculation gave better
results than sole inoculation. M4xM7 combination enhanced photosynthetic rate up to 90% at
15 dS m-1 in the absence of L-TRP, respectively over respective un-inoculated control.
Separate application of L-TRP also caused significant improvement in photosynthetic rate as
compared to control.
4.4.11 Transpiration rate
Figure 15 reveals that soil salinization had a negative effect on transpiration rate.
However, inoculation/coinoculation mitigated the adverse effects of salinity and improved the
transpiration rate at all salinity levels. A 9, 21, 51 and 69% decrease in transpiration rate was
observed at EC 4, 8, 12 and 15 dS m-1, respectively. Separate application of L-TRP also
caused significant improvement in transpiration rate compared to control.
Maximum increase up to 90, 96, 92 and 125% increase in transpiration rate at 4, 8, 12
and 15 dS m-1, respectively, was recorded by M4xM7 in the presence of L-TRP. Single
inoculation with M7 in the presence of L-TRP caused 73, 75, 95 and 100% increase at 4, 8,
12 and 15 dS m-1, respectively, as compared to respective un-inoculated control. Inoculation
in the absence of L-TRP also significantly improved the transpiration rate. M4xM7
combination in the absence of L-TRP enhanced transpiration rate up to 90% at 15 dS m-1,
respectively over respective un-inoculated control. Separate application of L-TRP also caused
significant improvement in transpiration rate compared to control.

82
kl
m-o
p-r
tu u
i-k
l-n
o-r
s-u s-u
c-e
h-j
kl
m-q
r-t
b-d
f-h
jk
m-o
o-r
e-g
g-j
kl
qr rs
d-f
g-i
i-k
n-r o-r
ab
b-e
g-j
lm
n-r
aa-c
e-g
jk
m-p
0
5
10
15
20
25
30
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Pho
tosy
nthe
tic
rate
Control M4 M7 M4XM7
Figure 14: Effect of rhizobacterial inoculation (with and without L-TRP) on photosynthetic rate (µmol m-2 S-1) of maize at different salinity levels
(original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test.
Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

83
n-qp-r
r-t
uv
v
m-p n-qp-r
s-u
v
g-ji-k
l-n
r-t
v
d-he-i
i-k
n-q
tu
c-gf-i
j-l
p-s
uv
a-db-e
e-i
l-o
r-t
a-c
c-g
h-k
o-r
tu
aab
b-f
k-m
q-s
0
1
2
3
4
5
6
7
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Tra
nspi
rati
on r
ate
Control M4 M7 M4XM7
Figure 15: Effect of rhizobacterial inoculation (with and without L-TRP) on transpiration rate (mmol m-2 S-1) of maize at different salinity
levels (original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

84
4.4.12 Stomatal conductance
Severe reduction in the stomatal conductance of maize was recorded due to soil
salinity (Figure 16). On the other hand, inoculation was found promising in improving the
stomatal conductance of maize plants. Combined application of strains performed better than
their separate inoculation. L-TRP application further enhanced the performance of
inoculation. A decrease up to 9, 21, 51 and 69% in stomatal conductance was recorded at EC
4, 8, 12 and 15 dS m-1, respectively.
In the presence of L-TRP maximum increase up to 125% in stomatal conductance
was observed by M4xM74 at 15 dS m-1. In the absence of L-TRP, inoculation/combined
inoculation also significantly improved the stomatal conductance. M4xM7 combination
enhanced stomatal conductance up to 90% at 15 dS m-1, over respective un-inoculated
control. Similarly at EC 8 and 12 dS m-1, an increase up to 81 and 87% was recorded by
inoculation with M4xM7.
4.4.13 Substomatal CO2 concentration
Soil salinization caused a negative effect on substomatal CO2 concentration (Figure
17). However inoculation/combined-inoculation in the absence and presence of L-TRP was
found promising in improving the transpiration rate of maize plants. At EC 4, 8, 12 and 15 dS
m-1, up to 5, 18, 32 and 40% decrease in transpiration rate as compared with control was
recorded, respectively.
M4xM7 in the presence of L-TRP caused maximum increase up to 9, 3, 9, and 8%
increase in substomatal CO2 concentration at 4, 8, 12 and 15 dS m-1. Single inoculation with
strain M4 in the presence of L-TRP caused up to 6, 3, 7, 6 and 7% increase at 4, 8, 12 and
15dS m-1 respectively, as compared with respective un-inoculated control.
Inoculation/coinoculation in the absence of L-TRP also significantly improved the
transpiration rate.

85
e-ih-k
k-p
j-o
p-r
b-e
b-f
e-i
j-n
qr
a-d
d-g h-m
l-p
o-r
ab
b-gf-l
k-r
qr
e-k
d-i
h-o
l-r
o-r
ab
b-g
f-l
k-r
qr
a-c
b-g
f-l
j-r
m-r
a
b-f
d-j
j-r
n-r
0
0.05
0.1
0.15
0.2
0.25
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Stom
atal
con
duct
ance
Control M4 M7 M4XM7
Figure 16: Effect of rhizobacterial inoculation (with and without L-TRP) on stomatal conductance (mmol m-2 S-1) of maize at different salinity
levels (original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

86
d-hf-i
k-m
p-r
t
c-f
e-g
h-j
o-qr-t
c-f
e-g j-l
n-p
st
c-f
d-ff-h
o-q
h-k
c-e
e-h
h-j
m-or-t
ab
c-e
i-k
o-q
i-k
a-c
d-f
h-j
m-o
r-t
a
b-d
g-j
l-n
g-j
0
0.05
0.1
0.15
0.2
0.25
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Subs
tom
atal
CO
2co
ncen
trat
ion
(vpm
)Control M4 M7 M4XM7
Figure 17: Effect of rhizobacterial inoculation (with and without L-TRP) on substomatalCO2 concentration of maize at different salinity levels
(original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test.
Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

87
4.4.14 Proline contents
Proline acts as an osmolyte, so increased concentration of proline is produced in
plants under stress condition. Figure 18 represents that proline concentration increased in
plants subjected to salt stress. The maximum increase (96%) in proline contents as compared
with control was recorded in plants subjected to salinity at 15 dS m-1. Inoculation with
rhizobacterial strains caused a significant reduction in proline contents over uninoculated
controls both in the presence and absence of L-TRP. Along with L-TRP sole inoculation with
strain M4 caused maximum reduction (15%) in proline contents at EC 15 dS m-1 over
respective uninoculated control. At 4, 8 and 12 dS m-1, strain M4 caused 24, 21 and 17%
decrease in proline contents, respectively, over respective uninoculated control. M4xM7
along with L-TRP caused 33, 45, 23 and 22% decrease in proline contents at 4, 8, 12 and 15
dS m-1, respectively, over respective uninoculated control. Without L-TRP application
maximum decrease in proline contents at EC 15 dS m-1 was recorded by M4xM7 which was
24% less as compared to respective uninoculated control. Sole inoculation with M4 and M7
caused positive reduction in proline contents, both in the presence and absence of L-TRP.
4.4.15 Total soluble sugars
Figure 19 shows the effect of inoculation/combined-inoculation on the total soluble
sugars of maize grown under salt affected soil conditions. The data demonstrate that
concentration of soluble sugars was increased due to exposure to salinity. Upton 5, 7, 13 and
24% increase in soluble sugars was noted at 4, 8, 12 and 15 dS m-1, respectively, as compared
to uninoculated control.
Soluble sugars contents were decreased due to inoculation/co-inoculation with
rhizobacterial strains both in the absence and presence of L-TRP. However inoculation/co-
inoculation gave better results when it was carried out along with L-TRP application.
Maximum reduction in total soluble contents was recorded by co-inoculation with M4xM7
that was up to 30, 30, 34 and 32% at 4, 8, 12 and 15 dS m-1, respectively over respective un-
inoculated control. Along with L-TRP application sole inoculation also significantly reduced
the total soluble sugars at all salinity levels. As compared with strain M7, M4 showed greater
reduction in soluble sugar contents.

88
r
e-gcd
b
a
rs
f-hde
bca
tu
mni-l
h-j
d-f
tu
o-ql-n
h-j
d-f
u
k-mh-k gh
cd
tu
m-oi-l
gh
de
u
qn-p
l-n
gh
v
rpq
lmh-j
0
0.5
1
1.5
2
2.5
3
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Prl
ine
Con
tent
s (µ
mol
g-1
FW
)Control M4 M7 M4XM7
Figure 18: Effect of rhizobacterial inoculation (with and without L-TRP) on proline contents of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

89
c-fb-d bc
b
a
j-m j-mh-k
g-jd-g
h-mg-k g-j
e-h
c-e
op l-pm-o m-o
i-mg-j f-i f-h
d-g
b-d
m-o m-ok-o k-o
h-l
m-o l-o l-oj-m
h-m
q q pq pqo-q
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Tot
al so
lubl
e su
gars
(µg
g-1F
W)
Control M4 M7 M4XM7
Figure 19: Effect of rhizobacterial inoculation (with and without L-TRP) on total soluble sugars of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

90
Inoculation/co-inoculation also significantly reduced the soluble sugar contents in the absence
of L-TRP application. In this case combination M4xM7 gave maximum reduction in soluble
sugars that was up to 29, 31, 31 and 30% at 4, 8, 12 and 15 dS m-1, respectively as compared
to uninoculated control. L-TRP application as alone treatment was also found effective in
reducing the total soluble sugar concentration at all salinity levels as compared to control.
4.4.16 Total phenol contents
Data regarding the total phenol contents is presented in Figure 20. The data
demonstrate that concentration of phenol contents was increased due to exposure to salinity.
However, inoculation/coinoculation lowered the phenol contents were decreased due to with
rhizobacterial strains. Inoculation gave better results when it was carried out along with L-
TRP application. Maximum reduction in phenol contents (42%) over respective un-inoculated
control were recorded by M4xM7 at 15 dS m-1. Along with L-TRP application sole
inoculation also significantly reduced the phenol contents at all salinity levels. Strain M4
showed greater reduction in phenol contents as compared with strain M7.
In the absence of L-TRP application, M4xM7 combination gave maximum reduction
in soluble sugars that was up to 45% at 15 dS m-1, as compared with respective uninoculated
control. L-TRP application as alone treatment was also found effective in reducing the phenol
contents at all salinity levels as compared to control.

91
b-d b-d bc
c-f
a
d-g c-f c-gb-e
b
f-k e-j e-id-e
b
j-o i-o i-oh-m
d-h
i-n h-m h-mb-e
b-e
n-p n-p m-o l-og-l
l-o k-o j-nb-d
b-g
q pq pq o-ql-o
0
0.5
1
1.5
2
2.5
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Tot
al p
heno
lics
(µg
g-1F
W)
Control M4 M7 M4XM7
Figure 20: Effect of rhizobacterial inoculation (with and without L-TRP) on total phenolics of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

92
4.4.17 Ascorbate peroxidase (cAPX)
Figure 21 shows the effect of inoculation/coinocualtion on the ascorbate peroxidase of
maize grown under salt affected soil conditions. The data demonstrate that activity of
ascorbate peroxidase increased under salinity stress. An increase of 5, 7, 13 and 24% in
ascorbate peroxidase was noted at 4, 8, 12 and 15 dS m-1, respectively as compared to
uninoculated control.
Ascorbate peroxidase activities were decreased due to inoculation/co-inoculation with
rhizobacterial strains both in the absence and presence of L-TRP. However
inoculation/coinoculation gave better results when it was carried out along with L-TRP
application. Maximum reduction in activity was recorded by co-inoculation with M4xM7 that
was 30, 30, 34 and 32% at 4, 8, 12 and 15 dS m-1, respectively over respective un-inoculated
control. Along with L-TRP application sole inoculation also significantly reduced the
ascorbate peroxidase at all salinity levels. Strain M4 showed greater reduction in ascorbate
peroxidase activity as compared with strain M7. Inoculation/combined-inoculation also
significantly reduced the ascorbate peroxidase activity in the absence of L-TRP application.
In this case M4xM7 combination gave maximum reduction that was up to 30, 31, 31 and 30%
at 4, 8, 12 and 15 dS m-1 respectively, as compared to uninoculated control. L-TRP
application as alone treatment was also found effective in reducing the ascorbate peroxidase
activity.

93
e-hd-g
c-ea-c
ab
f-h e-g
e-gd-f
d-f
g-ie-g
d-f
b-d
a-c
h-j h-j h-j h-j g-if-h f-h
d-f
a-c
a
0.610.55
0.65 0.67 0.69
0
0.2
0.4
0.6
0.8
1
1.2
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
cAP
X (m
M a
scor
bate
min
-1g-1
)Control M4 M7 M4XM7
Figure 21: Effect of rhizobacterial inoculation (with and without L-TRP) on cAPX activity of maize at different salinity levels (original =
1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is
average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

94
4.4.18 K:Na
The results of the inoculation on the K:Na are demonstrated in Figure 22. It is clear
from the data that salinity stress significantly decreased the K:Na in maize plants at all
salinity levels. Upto 9, 42, 57 and 61% decrease in K:Na was recorded at 4, 8, 12 and 15 dS
m-1, respectively, as compared with control. However, inoculation/coinoculation enhanced the
K:Na both in the absence and presence of L-TRP application. Maximum improvement in
K:Na was caused by co-inoculation with M4xM7 along with L-TRP application. This
increase in K:Na was up to 52, 69, 90 and 87% at 4, 8, 12 and 15 dS m-1 respectively, over
respective uninoculated control. Sole inoculation with M4 and M7 along with L-TRP also
performed better and maximum increase in this case was recorded by strain M4 which caused
up to 31, 46, 60 and 75% increase in K:Na at 4, 8, 12 and 15 dS m-1 respectively, over
uninoculated control.
Inoculation/coinoculation also improved the K:Na even in the absence of L-TRP
application. Co-inoculation with M4xM7 caused an increase up to 47, 75, 88 and 87% at 4, 8,
12 and 15 dS m-1, respectively as compared to respective uninoculated control. Separate
application of L-TRP also positively improved the K:Na but this improvement was
statistically non significant as compared to respective control.
4.4.19 Nitrogen in shoot (%)
The results regarding the Effect of rhizobacterial inoculation on the nitrogen content
in shoot of maize of maize under salt affected soil conditions revealed that salinity had a
repressive effect on nitrogen content (Figure 23). It was evident from the results that with
increasing salinity levels there was progressive decline in nitrogen content. However
rhizobacterial inoculation seemed promising in alleviating the salinity stress which was
reflected in case of improvement in nitrogen content. Rhizobacterial inoculation in single and
dual combinations improved nitrogen content at all salinity levels. The Effect of
rhizobacterial inoculation was pronounced when strains were tested along with L-TRP
application.
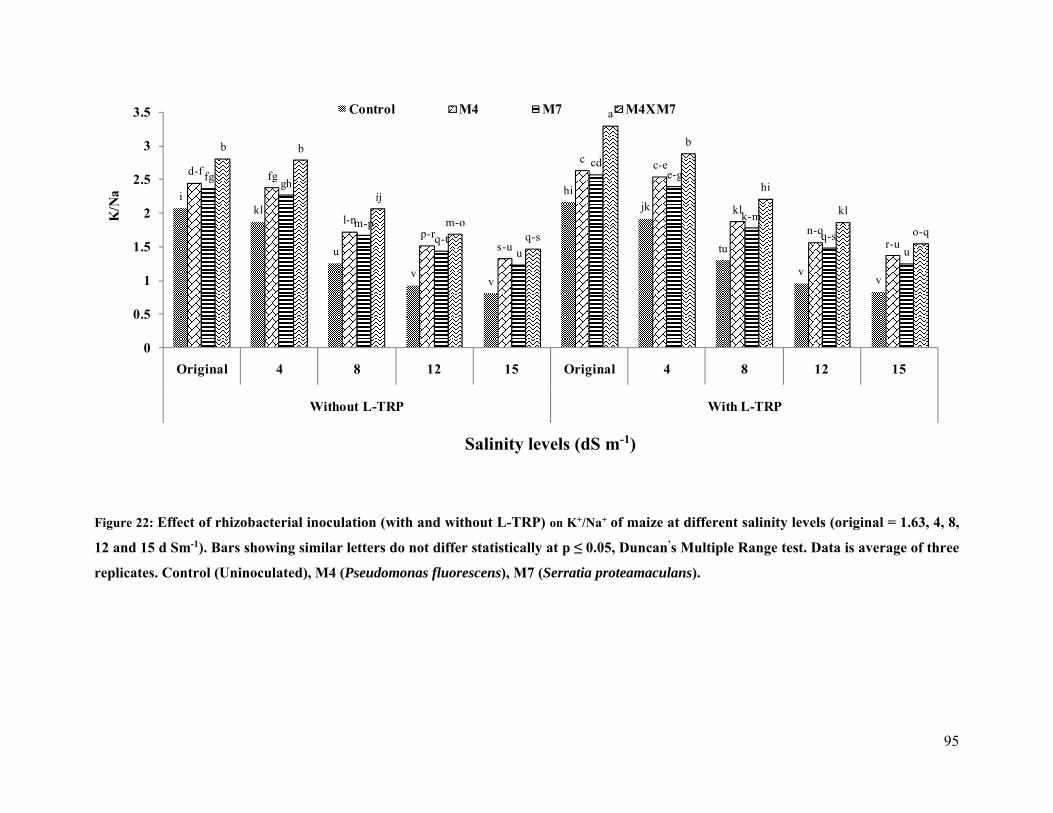
95
ikl
u
vv
hijk
tu
vv
d-f fg
l-np-r
s-u
c c-e
kl
n-qr-u
fggh
m-pq-t
u
cde-g
k-m
q-su
b b
ij
m-oq-s
a
b
hi
kl
o-q
0
0.5
1
1.5
2
2.5
3
3.5
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
K/N
aControl M4 M7 M4XM7
Figure 22: Effect of rhizobacterial inoculation (with and without L-TRP) on K+/Na+ of maize at different salinity levels (original = 1.63, 4, 8,
12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

96
i
k
p
qr
f-i
j
n
pr
d-f a e-i
jk
op
c-e cd d-h
j
no
d-h d-h g-i
kl
p
c-e d-g d-g
k
p
b b bc
hi
a
b bc-e
lm
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Nit
roge
n in
sho
ot (
%)
Control M4 M7 M4XM7
Figure 23: Effect of rhizobacterial inoculation (with and without L-TRP) on nitrogen concentration in shoot of maize at different salinity levels
(original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test.
Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

97
Salinity stress significantly reduced the nitrogen content at all tested salinity levels.
Maximum reduction in nitrogen content (71%) was noted at 15 dS m-1. In case of single strain
inoculation, M4 performed better than strain M7 and increased nitrogen content up to 51 and
27%, respectively. But when both these strains were tested in the presence of L-TRP further
increase in nitrogen content was noted i.e. M4 and M7 caused increase up to 53 and 29%,
respectively. However combined application of M4 and M7 gave the best results regarding
the improvement in plant height at all salinity levels both in the presence and absence of L-
TRP application. At 15dS m-1, an increase up to 74 and 73% was recorded in the absence and
presence of L-TRP respectively over respective uninoculated control by inoculation with
M4xM7.
4.4.20 Phosphorous concentration in grains (%)
As shown by the Figure 24 the phosphorous contents of maize plants were
significantly reduced by the soil salinity. Up to 17.54, 35.08 and 45.61% decrease in
phosphorous concentration was observed by salinity at 4, 8, and 12 dS m-1.
Sole and combined inoculation with M4 and M7 resulted significant increase in
phosphorous concentration under normal as well as salt affected soil conditions both in the
presence and absence of L-TRP application. Co-inoculation with M4xM7 caused maximum
increase in phosphorous concentration in grain when it was used in the absence and presence
of L-TRP application. In the presence of L-TRP 43, 57 and 53% increase and in the absence
of L-TRP a 42, 51 and 48% increase in phosphorous concentration was recorded at 4, 8 and
12 dS m-1, respectively, as compared with uninoculated control by this inoculation.
Sole inoculation with M4 and M7 also significantly enhanced the phosphorous contents.
Maximum increase in this case recorded by inoculation with M4 when it was tested along
with L-TRP, which resulted up to 25, 31 and 28% more phosphorous contents in maize grain
at 4, 8 and 12 dS m-1, respectively as compared with uninoculated control. Separate
application of L-TRP was also found effective in enhancing the P contents but this increase
was statistically non significant as compared to control at all tested salinity levels.

98
f
gh
i
j
d-f
gh
i
j
deef
gh
i
cdd-f
g
i
def
h
i
cdd-f
gh
i
a
bc
f
h
a
b
d-f
gh
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
P in
gra
in (%
)Control M4 M7 M4XM7
Figure 24: Effect of rhizobacterial inoculation (with and without L-TRP) on phosphorous concentration in grains of maize at different salinity
levels (original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

99
4.4.21 Crude protein contents
As shown by the Figure 25 the protein contents of maize grains were significantly
reduced by the soil salinity. Upto 7, 24 and 53% decrease in crude protein concentration as
compared with control was observed by salinity at 4, 8, and 12 dS m-1.
Sole and combined inoculation with M4 and M7 resulted a significant increase in
protein contents under normal as well as salt affected soil conditions both in the presence and
absence of L-TRP application. Co-inoculation with M4M7 caused maximum increase in
protein concentration in grain when it was used in the absence and presence of L-TRP
application. In the presence of L-TRP a 12, 11 and 60% increase and in the absence of L-
TRP, a 17, 28 and 56% increase in crude protein concentration was recorded at 4, 8 and 12 dS
m-1, respectively as compared with unioculated control by this inoculation.
4.5 Correlation analysis
The different coefficient of correlation between salinity, inoculation and different parameters
were correlated and significance of correlation was performed at p < 0.05 by using SPSS
software. A significant negative correlation was found between salinity and grain yield (Fig.
26) while a positive correlation was found between salinity and proline contents at different
salinity levels (Fig. 30). A positive correlation between inoculation and grain yield (Fig. 27)
was recorded while a negative correlation between inoculation and proline contents was
observed at different salinity levels (Fig. 31).

100
ghj
m
q
de
j
l
q
cd ef
j
o
bcde
h
n
ef ef
k
p
cdfg
ij
o
b b
hi
n
a bef
l
0
2
4
6
8
10
12
14
Original 4 8 12 15 Original 4 8 12 15
Without L-TRP With L-TRP
Cru
de p
rote
in c
onte
nts
(%)
Control M4 M7 M4XM7
Figure 25: Effect of rhizobacterial inoculation (with and without L-TRP) on crude protein contents in grains of maize at different salinity
levels (original = 1.63, 4, 8, 12 and 15 d Sm-1). Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range
test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans).
Salinity levels (dS m-1)

101
Figure 26: Effect of soil salinity on grain yield per plant of maize in pots
Figure 27: Effect of rhozobacterial inoculation on grain yield per plant of
maize in pots
R²=0.3727
R²=0.286
R²=0.3095
R²=0.1683
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
100.00
0 2 4 6 8 10
1.63
4
8
12
15
R²=0.1933R²=0.1729
R²=0.1864
R²=0.1758
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
100.00
0 5 10 15
Control
M4
M7
M4xM7

102
Figure 28: Effect of soil salinity on total chlorophyll contents (SPAD
values) of maize in pots
Figure 29: Effect of rhizobacterial inoculation on total
chlorophyllcontents (SPAD values) of maize in pots
R²=0.3189
R²=0.2164
R²=0.2138
R²=0.2441
R²=0.2
0.0
10.0
20.0
30.0
40.0
50.0
60.0
0 2 4 6 8 10
1.63
4
8
12
15
R²=0.206
R²=0.1582
R²=0.1808
R²=0.2116
0.0
10.0
20.0
30.0
40.0
50.0
60.0
0 2 4 6 8 10 12
Control
M4
M7
M4xM7

103
Figure 30: Effect of soil salinity on proline contents of maize in pots
Figure 31: Effect of rhizobacterial inoculation on proline contents of
maize in pots
R²=0.3248
R²=0.3835
R²=0.3557
R²=0.2208
R²=0.2801
0.00
0.50
1.00
1.50
2.00
2.50
3.00
0 2 4 6 8 10
1.63
4
8
12
15
R²=0.1543
R²=0.1474
R²=0.147
R²=0.1588
0.00
0.50
1.00
1.50
2.00
2.50
3.00
0 2 4 6 8 10 12
Control
M4
M7
M4xM7

104
Figure 32: Effect of soil salinity on electrolyte leakage of maize in pots
Figure 33: Effect of rhizobacterial inoculation on electrolyte leakage of
maize in pots
R²=0.6303
R²=0.5117
R²=0.4527
R²=0.2925
R²=0.2303
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
0 2 4 6 8 10
1.63
4
8
12
15
R²=0.1188
R²=0.1131
R²=0.1197
R²=0.0909
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
1.80
2.00
0 2 4 6 8 10 12
Control
M4
M7
M4xM7

105
4.5 Field trial
In order to confirm the results of the pot trial under natural salt affected soil
conditions field trial were conducted for two consecutive years at a farmer field 72 GB,
Satiana Road, Faisalabad. The results of the field trial are described below.
4.5.1 Plant height
The results regarding the Effect of rhizobacterial inoculation on plant height of maize
are demonstrated in Figure 34. It was revealed from the data that plant height was improved
with bacterial inoculation in the absence and presence of L-TRP application. In the year I,
maximum increase in plant height (43%) was found in case of M4xM7 combination over un-
inoculated control. Single strain inoculation with M4 and M7 also significantly increased the
plant height up to 31 and 28%, respectively as compared as compared with uninoculated
control. Rhizobacterial inoculation along with L-TRP further enhanced the plant height. In
this case M4xM7 combination showed 37% increase over respective un-inoculated control.
Sole application of M4 and M7 along with L-TRP significantly improved plant height up to
26 and 25% respectively, over respective un-inoculated control. In 2nd year experiment,
maximum increase in plant height was recorded by M4xM7 combination both in the presence
and absence of L-TRP. Combined application of rhizobacterial strains improved plant height
up to 31 and 33% in the absence and presence of L-TRP respectively, as compared with
uninoculated control.
4.5.2 Grain yield
The results of Effect of rhizobacterial inoculation on the grain yield of maize grown
under salt affected soil condition are represented in Figure 35. In the first year experiment, a
maximum increase in grain yield was recorded up to 41 and 35% by M4xM7 when it was
tested with out and along with L-TRP application respectively, as compared with
uninoculated control. Single inoculation with M4 and M7 also significantly enhanced the
grain yield up to 33 and 30% respectively, over respective un-inoculated control. These
strains along with L-TRP application improved grain yield up to 23 and 20% respectively,
over respective un-inoculated control. Sole L-TRP application also significantly improved the
grain yield.
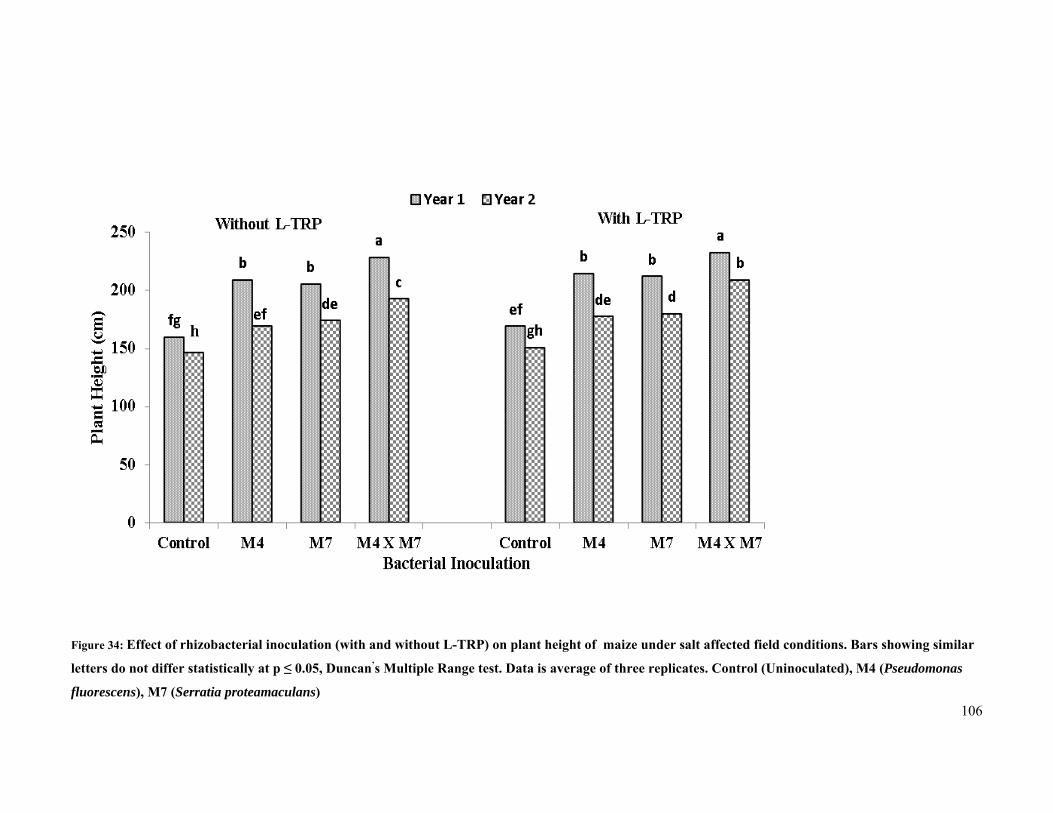
106
Figure 34: Effect of rhizobacterial inoculation (with and without L-TRP) on plant height of maize under salt affected field conditions. Bars showing similar
letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control (Uninoculated), M4 (Pseudomonas
fluorescens), M7 (Serratia proteamaculans)
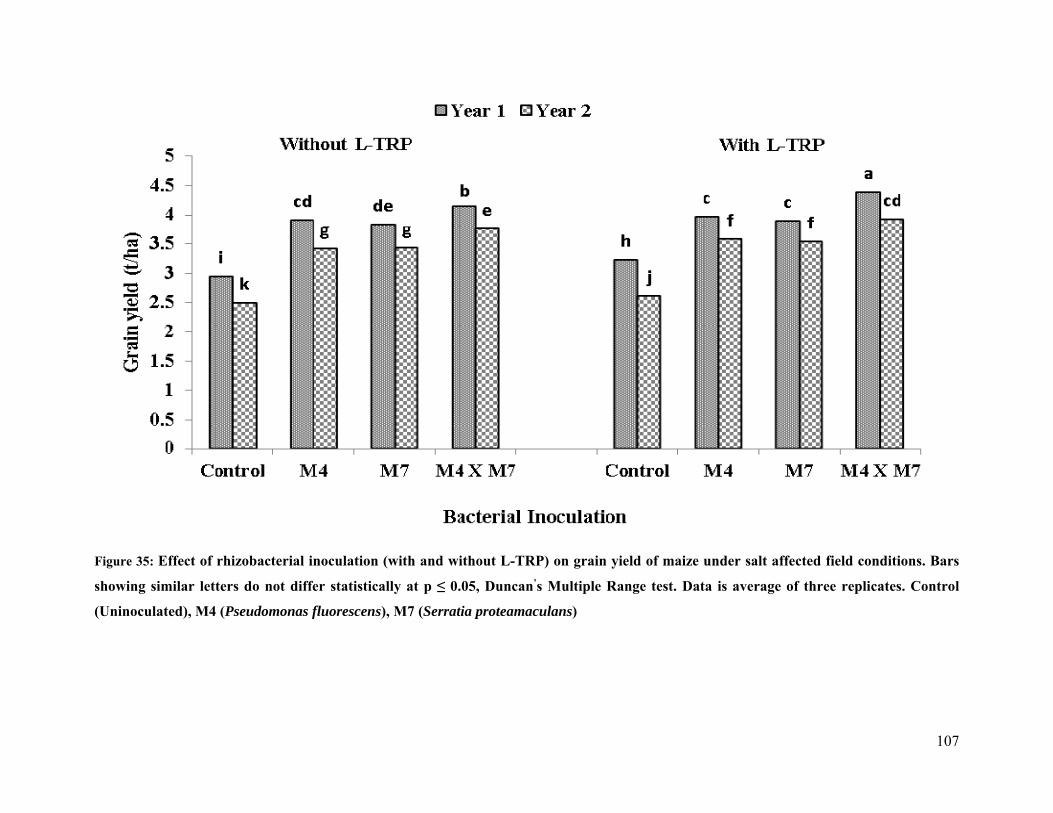
107
Figure 35: Effect of rhizobacterial inoculation (with and without L-TRP) on grain yield of maize under salt affected field conditions. Bars
showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control
(Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

108
In the second year trial co-inoculation with M4xM7 without L-TRP application
showed 49% improvement in grain yield over un-inoculated control while up to 50% increase
in grain yield over un-inoculated control was observed when M4xM7 combination was used
along with L-TRP. Single inoculation with M4 and M7 without L-TRP improved grain yield
up to 36 and 30% respectively, as compared with uninoculated control. Along with L-TRP
application sole inoculation with M4 and M7 showed increase up to 36 and 35% respectively,
over their respective uninoculated control.
4.5.3 Fresh biomass
Data (Figure 36) showed that single and dual inoculation significantly improved the
fresh biomass both in the presence and absence of L-TRP application.
In the year I, experiment, single inoculation with M4 and M7 in the absence of L-TRP
enhanced fresh biomass up to 44 and 43%, respectively, over un-inoculated control. While
M4xM7 showed a maximum increase (50%) when used in the absence of L-TRP. Along with
L-TRP application M4 and M7 improved fresh biomass of maize up to 37 and 25%
respectively, while in this case maximum increase (35%) was recorded by M4xM7, as
compared with uninoculated control.
In the second year trial, maximum increase in fresh biomass up to 51 and 36% was
recorded by the M4xM7 combination in the absence and presence of L-TRP respectively,
over their respective un-inoculated controls. Sole inoculation with M4 and M7 also
significantly improved the fresh biomass.

109
Figure 36: Effect of rhizobacterial inoculation (with and without L-TRP) on fresh biomass of maize under salt affected field conditions.
Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates.
Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

110
4.5.4 1000 grain weight
The data regarding 1000 grain weight is presented in Figure 37. Maximum
improvement in 1000 grain weight up to 24 and 26% over their respective controls was seen
in case of M4xM7 in 1st and 2nd year respectively, when it was applied along with L-TRP.
Without L-TRP application M4xM7 showed an increase in 1000 grain weight up to 25% in
year I and up to 26% in year II. In year I, sole inoculation with M4 and M7 without L-TRP
application improved 1000 grain weight up to 19% and 25% respectively, over respective un-
inoculated control. L-TRP application further enhanced the inoculation and both strains (i.e.
M4 and M7) shoed up to 18 and 19% increase in 1000 grain weight, respectively, over their
respective controls.
In the second year experiment, sole inoculation with M4 and M7 in the absence of L-TRP
gave up to 20 and 21% significant increase in 1000 grain weight respectively, over their
respective un-inoculated control. While along with L-TRP application up to 18 and 19%
increase in 1000 grain weight was recorded by inoculation with M4 and M7, respectively,
over uninoculated control.
4.5.5 Straw yield
Results of straw yield (Figure 38) depicted that sole inoculation and dual application
of rhizobacterial strains significantly enhanced the straw yield both in the absence and
presence of L-TRP in both years. Sole inoculation with M4 and M7 in the absence of L-TRP
significantly enhanced the straw yield up to 52 and 53% respectively, in first year and up to
51 and 49% respectively, over their respective un-inoculated control in the second year
experiment. Along with L-TRP application, the maximum increase (29%) in straw yield was
observed by single inoculation with M4 over its respective control in year I. while in 2nd year
experiment maximum increase (21%) was also recorded by inoculation with M4.
In first year co-inoculation with M4xM7 in the absence and presence of L-TRP
application resulted in a significant increase up to 56 and 35%, respectively, over their
respective un-inoculated control. While in 2nd year, an increase up to 50 and27% was
observed by M4xM7 when it was tested in the presence and absence of L-TRP, respectively.
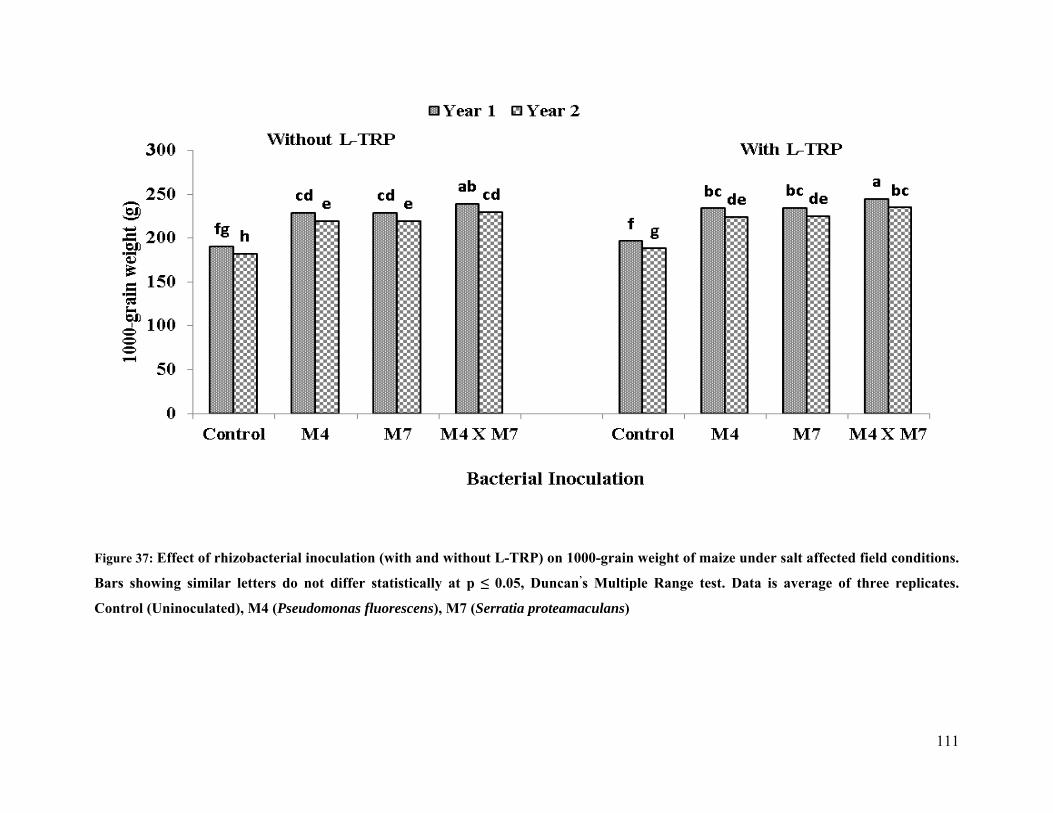
111
Figure 37: Effect of rhizobacterial inoculation (with and without L-TRP) on 1000-grain weight of maize under salt affected field conditions.
Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates.
Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)
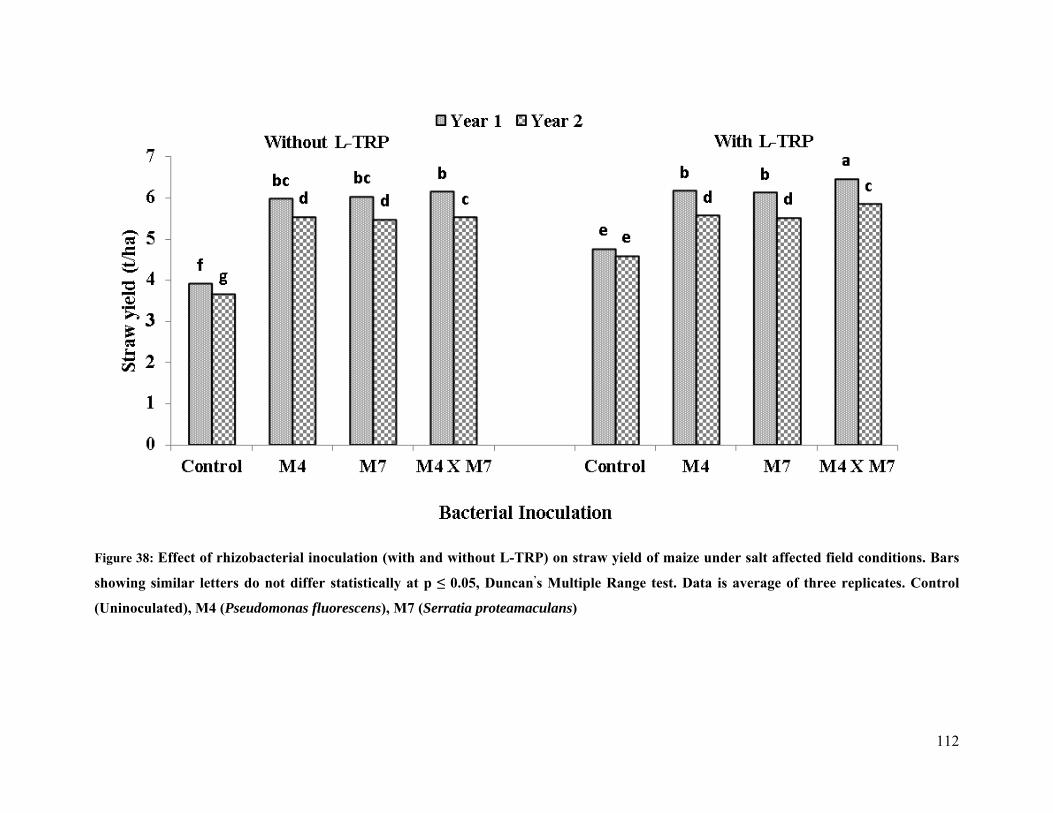
112
Figure 38: Effect of rhizobacterial inoculation (with and without L-TRP) on straw yield of maize under salt affected field conditions. Bars
showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control
(Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

113
4.5.6 Cob length
The results (Figure 39) of cob length showed that single and combined inoculation
with M4 and M7 significantly improved the cob length both in the presence and absence of L-
TRP application.
Without L-TRP application increase in cob length varied from 27.24 to 40.95% in
year I experiment and from 30 to 44% in the year II experiment. The maximum increase in
cob length without the application of L-TRP was recorded up to 40 and 44% by M4xM7 co-
inoculation over respective un-inoculated control in year I and year II, respectively.
Along with L-TRP application the improvement in cob length ranged from 14 to 29%
in year I and from 14 to 25% in year II. M4xM7 co-inoculation gave maximum improvement
in cob length over its respective control.
4.5.7 Weight per cob
Results regarding the Effect of rhizobacterial inoculation on cob weight are demonstrated in
the Figure 40.
In the year I trial, the maximum increase (53 and 46%) in cob weight over respective
un-inoculated control was recorded in case of M4xM7 co-inoculation in the absence and
presence of L-TRP, respectively. Sole inoculation with M4 and M7 in the absence of L-TRP
gave 42 and 41% increase in cob weight over their respective un-inoculated control. Along
with L-TRP application maximum increase (32%) was recorded by inoculation with strain
M4, as compared with uninoculated control.
In the 2nd year experiment, single inoculation with M4 and M7 in the absence of L-
TRP gave up to 40 and 36% increase in cob weight respectively, over respective un-
inoculated control. Along with L-TRP application M4 and M7 showed significant increase in
cob weight up to 33 and 31% respectively, over their un-inoculated control. The maximum
increase in cob weight up to 50 and 47% was recorded in case of M4xM7, when it was tested
without and along with L-TRP application, respectively.

114
Figure 39: Effect of rhizobacterial inoculation (with and without L-TRP) on cob length of maize under salt affected field conditions. Bars
showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control
(Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

115
Figure 40: Effect of rhizobacterial inoculation (with and without L-TRP) on weight per cob of maize under salt affected field conditions.
Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates.
Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

116
4.5.8 Total chlorophyll contents (SPAD values)
The results regarding total chlorophyll contents of maize grown under salt affected
soil condition are presented in Figure 41. It is clear from the results that the inoculation/ co-
inoculation significantly improved the total chlorophyll contents of maize both in the absence
and presence of L-TRP application.
In 1st year experiment, co-inoculation with MxXM7 showed maximum improvement
in total chlorophyll content (23 and 25%) over its respective un-inoculated control in the
presence and absence of L-TRP application, respectively. Single strain inoculation with M4
and M7 also significantly improved the total chlorophyll contents with and without L-TRP
application. The mere application of L-TRP also improved chlorophyll contents but this
improvement was statistically non-significant as compared to control where no L-TRP was
applied.
In the 2nd year experiment, the improvement in chlorophyll ranged from 15 to 27%
over un-inoculated control in case when inoculation/co-inoculation was tested without L-TRP
application. On the other hand, inoculation/co-inoculation along with L-TRP application
improved chlorophyll from 15 to 25% over respective uninoculated control. Separate
application also non- significantly improved total chlorophyll contents of maize.
4.5.9 Proline contents
Under stress condition plants produce osmolytes such as proline. Data regarding
proline contents (Figure 42) showed that inoculation with rhizobacterial strains both in the
presence and absence of L-TRP significantly decreased the proline contents in maize plants
grown under salt stressed soil conditions.

117
Figure 41: Effect of rhizobacterial inoculation (with and without L-TRP) on total chlorophyll contents (SPAD values) under salt affected
field conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)
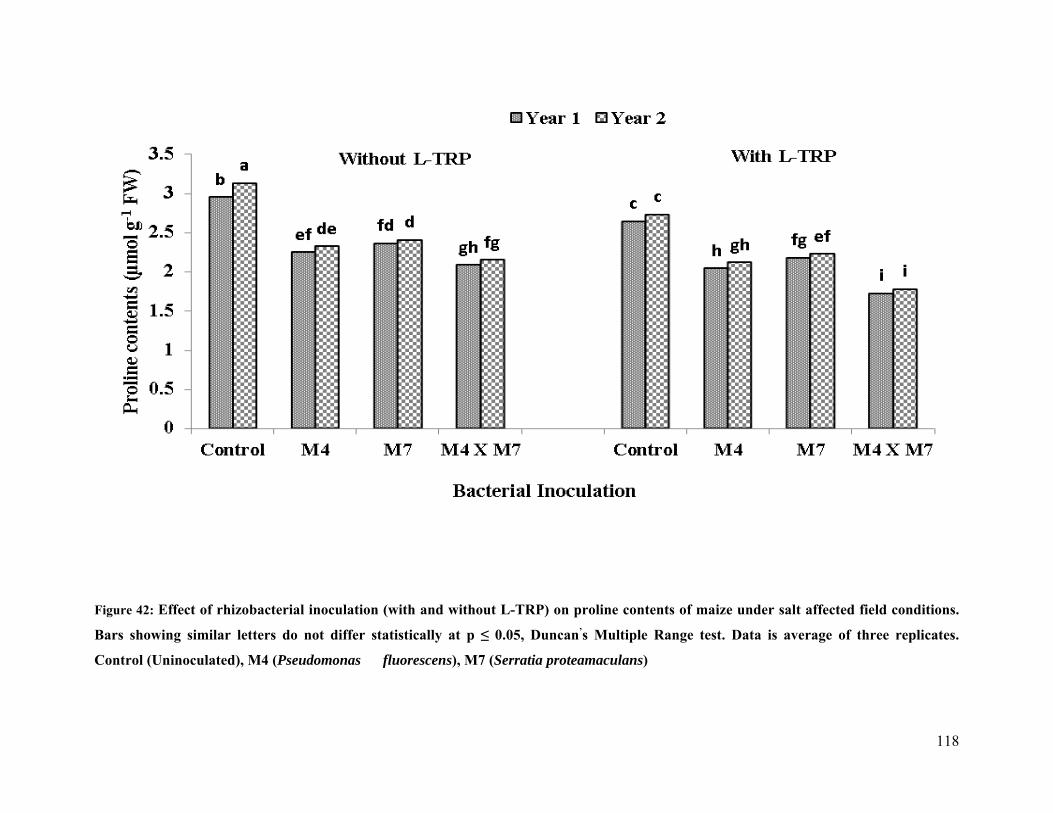
118
Figure 42: Effect of rhizobacterial inoculation (with and without L-TRP) on proline contents of maize under salt affected field conditions.
Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates.
Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

119
In the year I experiment reduction in proline contents ranged from 23to 29% over the
control when rhizobacterial strains were tested in the absence of L-TRP. While along with L-
TRP application reduction in proline contents by these strains ranged from 22 to 34% over
their respective un-inoculated controls. Minimum proline contents were recorded by the co-
inoculation with M4xM7 both in the absence and presence of L-TRP application.
In the year II experiment the maximum reduction (31 and 34%) was recorded by
M4xM7 over respective un-inoculated control in the absence and presence of L-TRP
application, respectively. Sole inoculation with M4 and M7 also significantly reduced the
proline contents both in the presence and absence of L-TRP application. Just an application of
L-TRP also decreased the proline contents but this decrease was non significant as compared
to control.
4.5.10 Crude protein contents in grains
The results regarding crude protein contents in grains (Figure 43) indicate that
rhizobacterial inoculation significantly improved the protein contents in the presence and
absence of L-TRP application. In the year I trial, the increase in protein contents due to
inoculation with M4 and M7 along with L-TRP application was 9 and 8% over respective
control, respectively. In the 2nd year trial these stain showed an enhancement in protein
concentration by 11 and 10% over respective un-inoculated control. These strains applied in
the absence of L-TRP also significantly improved the protein contents.
The combined application of these strains i.e. M4xM7 gave more promising results
than their sole application. An increase up to 15% in the first year experiment and 16% in the
second year experiment over respective un-inoculated control was recorded by M4xM7 in the
presence of L-TRP application. Combined inoculation in the absence of L-TRP application
also significantly improved the protein contents. Alone application of L-TRP also improved
the protein concentration in both years.
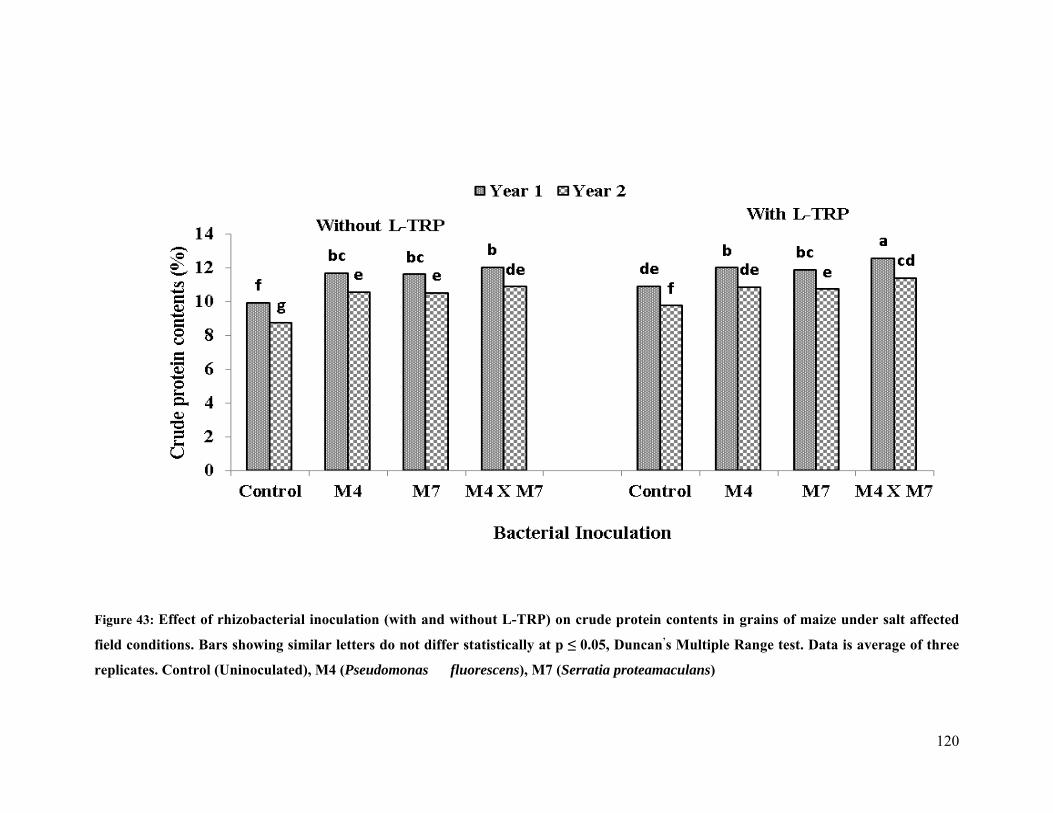
120
Figure 43: Effect of rhizobacterial inoculation (with and without L-TRP) on crude protein contents in grains of maize under salt affected
field conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

121
4.5.11 Phosphorous concentration in grains
The results regarding phosphorous concentration showed that inoculation/co-
inoculation significantly improved the P contents in grains of maize grown under salt affected
field conditions (Figure 44).
In the absence of L-TRP application, sole inoculation with M4 and M7 caused up to
16 and 11% increase in phosphorous concentration over respective control in year I and up to
19 and 14% in year II as compared with their respective control. Along with L-TRP
application these strains caused up to 17 and 21% increase in P contents over their respective
control in year I and up to 17 and 19% increase in year II experiment.
In the absence of L-TRP application co-inoculation with M4xM7 caused up to 21%
increase in phosphorous concentration in first year experiment and up to 25% increase over
its respective uninoculated control in second year trial. While along with L-TRP application
M4xM7 caused an increase of 24% in the 1st year and 24% increase in the 2nd year over its
respective uninoculated control. Improvement in phosphorous concentration was also noted
by just application of L-TRP over uninoculated control.
4.5.12 K:Na in leaves
Data in Figure 45 highlighted that inoculation/coinoculation significantly improved K:Na in
both year experiments.
In year I, maximum increase in K:Na in the presence of L-TRP was recorded when
M4 and M7 (M4xM7) were combined in combination. The increase was 410% over the
respective uninoculated control. M4 and M7 in single inoculation along with L-TRP gave
415% and 399% increase over their respective uninoculated controls.
Inoculation/coinoculation in the absence of L-TRP also significantly improved the K/Na. Non
significant improvement as compared with control was also noted by the sole application of
L-TRP application.

122
Figure 44: Effect of rhizobacterial inoculation (with and without L-TRP) on phosphorous concentration iin grains of maize under salt
affected field conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average
of three replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

123
Figure 45: Effect of rhizobacterial inoculation (with and without L-TRP) on K+/Na+ of maize under salt affected field conditions. Bars
showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control
(Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

124
In the second year experiment M4xM7 in the absence and presence of L-TRP showed
up to 361 and 511% increase in K:Na over their respective un-inoculated control. M4 and M7
applied as single inoculation along with L-TRP showed up to 4188 and 402% increase over
their respective control. These strains in the absence of L-TRP also significantly improved the
K/Na. sole application of L-TRP as a treatment also positively improved the potassium ratio.
4.5.13 Total phenol contents
The results regarding total phenol contents are shown in Figure 46. It is evident from
the data that phenolic contents improved by inoculation/coinoculation. The improvement in
total phenolic contents in presence of L-TRP ranged from 36 to56% in first year and from 29
to36% in second year as compared with respective un-inoculated control. In the absence of L-
TRP maximum improvement in total phenolic contents up to 32% in the first year and 25% in
the second year, over respective un-inoculated controls was recorded by MxM7. Sole
inoculation with M4 and M7 in the absence of L-TRP also improved the total phenolic
contents in both years. The application of L-TRP alone was also found effective in improving
the total phenolic contents. Overall inoculation/ coinoculation along with L-TRP performed
better than without L-TRP.
4.5.14 Total soluble sugars
The data in the Figure 47 shows the effect of inoculation/coinoculation on total
soluble sugars of maize grown salt affected field conditions.
In the year I sole inoculation with M4 and M7 showed 41 and 45% increase in total
soluble sugars in the absence of L-TRP and 35 and 39% increase in total soluble sugars as
compared to their respective uninoculated controls. Maximum increase (50%) in total soluble
sugars was recorded by M4xM7 in the absence and presence of L-TRP, respectively.

125
Figure 46: Effect of rhizobacterial inoculation (with and without L-TRP) on total phenol content of maize under salt affected field
conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

126
Figure 47: Effect of rhizobacterial inoculation (with and without L-TRP) on total soluble sugars of maize under salt affected field
conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

127
In the second year, M4xM7 increase in the total soluble sugars up to 32 and 25% over
the respective uninoculated control in the absence and presence of L-TRP, respectively. Sole
inoculation with M4 and M7 also significantly improved the total soluble sugars both in the
presence and absence of L-TRP. Non significant increase in total soluble sugars was also
observed with the sole application of L-TRP.
4.5.15 Electrolyte leakage
The results of the Effect of rhizobacterial inoculation on electrolyte leakage are shown
in Figure 48. In the year I experiment, inoculation/co-inoculation significantly reduced the
electrolyte leakage both in the presence and absence of L-TRP. The combined application of
both strains was found more effective than their sole inoculation. Along with L-TRP
application the maximum reduction (35%) in electrolyte leakage was recorded by M4xM7.
Sole inoculation with M4 and M7 also reduced the electrolyte leakage up to 32 and 30%
respectively, as compared to uninoculated control. Without L-TRP M4xM7 showed
maximum reduction in electrolyte leakage up to 31% followed by sole inoculation with M4
and M7, which showed a 27 and 27% reduction in electrolyte leakage as compared to un-
inoculated control.
In second year trial, coinoculation showed maximum reduction in electrolyte leakage
both in the presence and absence of L-TRP. This increase was up to 33% without L-TRP and
37% when co-inoculation was tested in the presence of L-TRP. Single inoculation also
reduced the electrolyte leakage significantly. Strain M4 was found more effective than M7
and caused 29 and 33% decrease in electrolyte leakage in the presence and absence of L-TRP
respectively, as compared with uninoculated control.

128
Figure 48: Effect of rhizobacterial inoculation (with and without L-TRP) on electrolyte leakage of maize under salt affected field
conditions. Bars showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three
replicates. Control (Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

129
4.5.16 SOD
The results of the effect of inoculation/coinoculation on the SOD values are demonstrated in
Figure 49. It is clear from the results that the rhizobacterial inoculation/co-inoculation
significantly reduced the SOD values both in the presence and absence of L-TRP.
In the 1st year trial, M4xM7 caused a maximum reduction in SOD values up to 64 and
72% in the absence and presence of L-TRP respectively, as compared with respective
uninoculated control. Single inoculation with M4 and M7 also caused significant reduction in
SOD values up to 71 and 69% in the presence of L-TRP and up to 58 and 54% in the absence
of L-TRP respectively, over their respective un-inoculated control.
In the 2nd year trial, co-inoculation with M4xM7 resulted in a maximum decrease of
up to 65% and 75% in the absence and presence of L-TRP respectively over its respective un-
inoculated control. In the absence of L-TRP single inoculation with M4 and M7 caused 45
and 52% decrease in SOD values over their respective uninoculated control. Along with L-
TRP application these strains caused 55 and 61% decrease in SOD values.

130
Figure 49: Effect of rhizobacterial inoculation (with and without L-TRP) on SOD activity of maize under salt affected field conditions. Bars
showing similar letters do not differ statistically at p ≤ 0.05, Duncan’s Multiple Range test. Data is average of three replicates. Control
(Uninoculated), M4 (Pseudomonas fluorescens), M7 (Serratia proteamaculans)

131
4.6 Identification and characterization of selected strains
Both selected strains were characterized (Table 9) for different biochemical
characters. The results of these tests are summarized below.
It is evident from the results that both of these strains were positive in oxidase
activity, catalase activity, chitinase activity and phosphate solubilization. Both strains were
also positive for ACC-deaminase activity. The results of root colonization showed that both
stains colonized the roots but maximum root colonization (6.11 × 106 cfu g-1) was observed
by the strain M4 follwed by the strain M7. These strains also possesed the exopoly-saccharide
production ability.
The selected strains were identified by BIOLOG® as Pseudomonas fluorescens (M4)
and Serratia proteamaculans (M7).

132
Table 9: Characterization of selected rhizobacterial strains
Strains Percent
match to
Biolog
ACC-
deaminase
Activity
Phosphate
solubilization
Chitinase
activity
Oxidase
activity
Catalase
activity
Exopoly-
saccharide
production
Root
colonization
(cfu g-1)
M4 68% +ve +ve +ve +ve +ve +ve 6.11 × 107
M7 59% +ve +ve +ve +ve +ve +ve 5.32 × 106
M4, Pseudomonas fluorescens; M7, Serratia proteamaculans

133
CHAPTER-5
DISCUSSION
In the present study we isolated a number of rhizobacterial strains from the rhizosphere of
maize plants growing under salt-affected soil conditions. These strains were tested for their
ability to tolerate salinity stress by using osmoadaptation assay. Out of these strains, 19 most
tolerant strains were selected and their auxin production ability in the presence and absence of
L-TRP was measured. Eleven strains which tolerated salinity and produced auxin were
selected to test their ability of growth promotion of maize seedlings under salt stressed axenic
conditions. After this experiment the two most efficient strains (M4 and M7) were selected
for further evaluation in pot and field trials to improve the growth and yield of maize under
salt stressed conditions.
5.1 Salt tolerance of rhizobacterial strains
In this study the isolated rhizobacterial strains were subjected to osmoadaptation
assay. Results of this assay revealed that salinity adversely affected the growth of
rhizobacterial strains. At normal conditions, the growth of strains was optimum but the
growth of rhizobacterial strains varied under salinity stress. The growth of some strains was
improved even at higher salinity levels. This might be due to the ability of these
rhizobacterial strains to tolerate saline conditions. Such variability among bacterial strains to
tolerate salinity has been reported by Loret et al. (1995). Mansah et al. (2006) and Sgroy et al.
(2009) have also reported the variable growth rates of bacteria isolated from the rhizosphere
of salt tolerant plants.
Salinity adversely affects the survival and growth of the bacteria (Tate, 1995; Troman and
Weaver, 2006). The growth of bacteria is inhibited or reduced under saline conditions
(Troman and Weaver, 2006). However, some bacteria are able to withstand salinity stress.
Kaseem et al. (1985) reported that rhizobia had better ability to tolerate higher salinity than
their host plant. The ability of these bacterial strains to tolerate higher levels of salinity may
be due to changes in morphology and metabolism, which makes them able to cope and adopt
saline conditions. Bacterial species vary in their ability to withstand salinity (Bernard et al.,

134
1986; Elsheikh and Wood, 1990). The strains isolated from salt-affected conditions are more
able to tolerate salinity stress than those isolated from normal soil conditions (Hua et al.,
1982). Salt tolerant bacteria adapt to saline conditions by using different strategies. They
expend energy to exclude salt from their cytoplasm to avoid protein aggregation (‘salting
out’). In order to survive the high salinities, these bacteria employ two differing strategies to
prevent desiccation through osmotic movement of water out of their cytoplasm. Both
strategies work by increasing the internal osmolarity of the cell. In the first strategy organic
compounds are accumulated in the cytoplasm. These osmoprotectants are known as
compatible solutes. These can be synthesised or accumulated from the environment The most
common compatible solutes are neutral or zwitterionic and include amino
acids, sugars, polyols, betaines and ectoines, as well as derivatives of some of these
compounds. The second, more radical, adaptation involves the selective influx
of potassium (K+) ions into the cytoplasm (Santos and da Costa, 2002). The bacterial cells
also respond to salinity stress by changing their morphology. The cell ultrastructure and
morphology of salt tolerant strain of cowpea Rhizobium was severely affected under salt
stress. The cells appeared as spiral or filament-like structures, and the cell size greatly
expanded. The cell envelope was distorted, and the homogeneous cytoplasm was disrupted
(Zahran et al., 1992). It has been reported (Busse et al., 1989) that cells of a strain of R.
meliloti appeared with irregular morphology at potentials below -0.5 MPa. Strains of rhizobia
from different species modified their morphology under salt stress, and rhizobia with altered
morphology have been isolated from salt-affected soils in Egypt (Zahran et al., 1992). High
osmotic stress modified the synthesis pattern of extracellular and capsular polysaccharides
of R. leguminosarum (Breedveld, et al., 1991.). The colonies of R. meliloti grown in the
presence salt stress show a decrease in mucoidy and lower level of exopolysaccharides
production (Lloret et al., 1998). The synthesis pattern in SDS-PAGE of lipopolysaccharides
(LPS) from various species of bacteria (Zahran, 1992; Zahran et al., 1994.) was modified by
salt.

135
5.2 Effect of rhizobacterial inoculation on growth of maize in axenic conditions
Abiotic stresses such as salinity cause reduction in the growth and development of
plants by impairing their physiological and biochemical mechanisms. These effects of
stresses can be minimized by the application of bacterial inoculants, which help the plants to
maintain their growth under stressed conditions by provoking natural mechanisms (Yang et
al., 2008; Vardharajula et al., 2011). Plant growth promoting rhizobacteria most commonly
improve the plant growth by increasing the root growth (Lucy et al., 2004) by producing
phytohormones. Plant hormones play a vital role in controlling the root growth. In the root
zone, auxin produced by the bacteria cause changes in root architecture. These changes result
in better root growth and increased root surface area, which ultimately result in better uptake
of nutrients and water leading to enhanced growth of whole plant (Somers et al., 2004;
Principe et al., 2007). On the other hand under stressful conditions plant produced ethylene
results in decreased root and shoots growth. But rhizobacteria containing ACC-deaminase
activity can degrade the precursor of ethylene (ACC), thus lower the internal levels of
ethylene in plants and rescue them from the stress (Mayak et al., 2004). In our growth room
study we observed that salinity caused a negative effect on the growth of maize seedlings. A
progressive decrease in root/shoot length and fresh/dry biomass of maize seedlings was
recorded with increasing salinity levels. However, inoculation with rhizobacterial strains
caused significant improvement in root/shoot growth of maize seedlings. This might be due
to their phytohormone production ability such as auxin and regulating the hormonal status of
plants and lowering the ethylene concentration. Elevated amounts of ethylene production and
alteration in hormonal (auxin) balance in plants have been documented by many researchers
(Mayak et al., 2004; Yue et al., 2007; Zahir et al., 2008). Many researchers have documented
the enhanced growth of different crops by microbial cultures mainly due to their
phytohormone production ability such as auxin (Glick, 1995; Noel et al., 1996; Okon and
Vanderleyden, 1997). The results of this study highlight the importance of screening auxin
producing PGPR for improving the growth and yield of maize under salt stressed conditions.
5.3 Effect of rhizobacterial inoculation on physiological parameters of maize
The ability of plants to cope salinity stress has been accessed on growth, yield,
biochemical and genetic basis. Other than these relative water contents, photosynthetic

136
activity, leaf gas exchange, electrolyte leakage are also good indicators of stresses (Burling et
al., 2013; Maccaferri et al., 2013). In our field and pot studies, it was found that salinity
adversely affected the transpiration rate, stomatal conductance and relative water contents.
The decrease in transpiration rate may be due to reduction in stomatal conductivity (Chedlia
et al., 2007). Plants close their stomata as an adaptive mechanism to withstand osmotic stress
and conserve water for later growth stages (Misra et al., 2002; Tardien, 2005; Hatami et al.,
2010). Due to this closure of stomata carbon dioxide intake is affected and photosynthesis
rate of plants subjected to salinity stress is decreased (Misra et al., 2002). Lawlor and Cornic
(2002) and Ben-Asher et al. (2006) also reported changes in stomatal conductivity and
decrease in photosynthetic activity. Increased concentration of Na+ causes reduction in carbon
dioxide assimilation (Chartzoulakis et al., 2002).
Salinity stress deprives plants from water and alongside cause ionic imbalance which
results in overall reduced growth of plants (Wyn Jones, 1981). The measurement of relative
water contents is an important parameter to assess stress tolerance in plants (Fisher et al.,
2000; Pereyra et al., 2006). In this study, we observed that inoculation /co-inoculation
application significantly enhanced the relative water contents. Our findings are also supported
by the results of other researchers (Nadeem et al., 2009; Naveed et al., 2014) who also found
improvement in relative water contents due to rhizobacterial inoculation. The reason of this
improvement in relative water contents may be due to longer roots caused by rhizobacterial
inoculation, which might enable the plant to absorb water from the deeper layers of soil
where concentration of salts is relatively low (Dodd et al., 2004; Hamida et al., 2004). This
increase in water contents may also be due to production of growth regulators such as auxins.
Cohen et al. (2008) also reported such improvements in relative water contents of leaves and
described it may be due to the bacterial production of plant growth regulators. Nadeem et al.
(2009) also conducted an experiment and reported such improvement in relative water
contents of maize leaves by using rhizobacterial strains under salinity stress. Osmotic stress
caused by salinity results in limited water supply which may lead to reduction in chlorophyll
contents (Paknejad et al., 2007). Higher chlorophyll contents are positively correlated with
increase in yield and under stress conditions elevated chlorophyll contents may result in
improvement in yield of plants.

137
When plants are subjected to osmotic stress, proline is accumulated in plants. Higher
concentration of such organic osmolytes in plants serves as a solute for osmotic adjustment
(Silverita et al., 2003). High concentration of proline is associated with stress conditions (Rai
et al., 2003). Extra energy is utilized to accumulate such osmolytes in addition to normal
metabolic costs. In our study, it was found that salinity stress caused a marked increase in
proline contents however, rhizobacterial inoculation /co-inoculation decreased the proline
contents. This decrease in proline contents might be due to the ameliorative effects of
rhizobacterial strains which helped the plants to withstand salinity stress and decreased the
proline contents. Similar results have been documented where bacterial inoculation was found
effective in lowering the proline concentration (Han and Lee, 2005; Nadeem et al., 2009).
In present study, we observed that salinity markedly decreased the chlorophyll
contents while rhizobacterial inoculation improved the chlorophyll contents. Our results are
also supported by studies of other researchers who found increase in chlorophyll contents due
to rhizobacterial inoculation (Nadeem et al., 2009). This increase in chlorophyll contents can
be attributed to increase in photosynthetic area of leaves due to inoculation/co- inoculation
with rhizobacterial strains which was otherwise decreased by salinity stress (Marcelis and
Van-Hooijdook, 1990). Ion toxicity caused by salinity stress also causes the death of older
leaves which results in the prevention of nutrient and hormonal supply to young leaves
(Munns, 1993; Tamman, 2003) which might lead to reduction in chlorophyll contents. Hamid
and Al-Komy (1998) have also reported such improvement in chlorophyll contents owing to
bacterial inoculation. The increased chlorophyll contents might have resulted in higher
photosynthetic rate and production of more photosynthates which ultimately resulted in
higher yields.

138
5.4 Effect of rhizobacterial inoculation on nutritional balance in maize
Changes in nutrient uptake is the most common phenomenon which plants undergo in
different stresses such as salinity stress (Greenway and Munns, 1980). This results in lower
uptake of essential nutrients due to which plant suffer nutrient deficiencies which result in
poor growth. In our study we observed that salinity stress caused marked decrease in N and P
contents of maize plants. Phosphorous is essential macronutrient which plays a vital role in
normal growth and development of plants. Martenez and Lauchli (1994) reported that
phosphorous uptake by plants was inhibited under salinity stress. We found that
rhizobacterial inoculation significantly improved the mineral nutrient contents (nitrogen,
phosphorous and crude protein contents). This might be due to increased root biomass which
can explore more soil area and results in enhanced uptake of nutrients from larger soil
volume. Our results are in line with Nadeem et al. (2006), Yao (2004) who also reported
enhanced nutrient uptake by microbial inoculation under stressful conditions.
Similarly, Gaballah and Comaa (2005) found decrease in protein contents of faba
bean. However, we observed inoculation /co-inoculation to be very effective in reversing this
pattern and increase the concentration of inorganic nutrients such as N and P in grains and
leaves of maize plant. In our study, crude protein contents of maize grains were also
improved due to rhizobacterial inoculation /co-inoculation. Other scientists have also reported
such improvement in inorganic nutrients and crude protein contents due to rhizobacterial
inoculation/ co-inoculation (Hamidia et al., 2004). Mayak and co-workers (2004) also
reported increase in P contents in tomato plants grown under salt stress conditions. Under
drought conditions, increase in N and P contents of lettuce plant inoculated with Bacillus sp.
have documented by Vivas et al. (2003). Similarly, Han and Lee (2005) also reported
improvement in N, P and Ca concentrations in soya bean plants under salinity stress by
inoculation with PGPR. Dovvlacre and Okon (2007) have correlated this increased uptake of
N and P with root elongation caused by Azospirillum inoculation. It can be stated that
increase in the root area and provision of growth hormones such as auxins would have helped
the plant in taking more nutrients from the soil. It is reported in many studies that salinity
stress causes reduction in uptake and assimilation of essential nutrients in plants, thus their

139
supplementation is required to mitigate the harmful effects of salinity stress (Endris and
Mohammad, 2007; Khayyat at al., 2007; Heider and Jamshid, 2010).
5.5 Effect of rhizobacterial inoculation on ionic balance in plants
Salt tolerance of plants can be indicated by K+/ Na+ (Hamida et al., 2004). It was
found in our study that salinity stress caused the ionic imbalance in plants. It resulted in
higher intake of Na+ than K+. There are two phases of salinity stress in plants (Munns, 1993).
In first phase, salinity causes osmotic stress which results in decrease in water absorption
from soil (Mayak et al., 2004). In the second step, salinity induces ionic stress/toxicity.
Second phase of salinity stress is reported to be more injurious than the first phase as it results
in alteration in uptake, distribution and compartmentalization of ions in the plants. Potassium
acts as an osmoregulator and maintains water balance in plants under stress conditions.
Decreased K+ concentration results in drop in leaf water potential. This decrease in K+
concentration may result in reduction in stomatal conductivity and activity of ribulose
bisphosphate carboxylase which ultimately impairs the photosynthesis (Zhao et al., 2001;
Szepesi et al., 2008).
Salinity also disturbs the physiological activity relating to the ionic and osmolyte
accumulation such as proline (Steward et al., 1974). Ehasanpour and Amini (2003) reported
that the uptake and transport of phosphate within plants was highly sensitive to salinity stress.
Uptake of Na+ and K+ are disturbed due to salinity (Ashraf, 2004). Fortmere and Schubert
(1995) demonstrated that Na+ toxicity was the major cause of stress during the second phase
of salinity in maize. Similarly, Schubert and coworkers (2009) described that grain formation
and reduction in overall yield of maize under saline conditions was caused by osmotic stress
and Na+ toxicity.
In our pot and field studies, plants were exposed to long term salinity stress as they
were grown till maturity and it is highly likely that they have suffered both phases of salinity
stress. We observed higher Na+ concentration in our plants subjected to salinity stress and it
may be the one of the stresses. Higher uptake of Na+ compared to K+ has been reported
previously (Pervaiz et al., 2002; Ashraf, 2005). This increased uptake of Na+ results in
reduction of K+/Na+. Plants uptake Na+ and K+ through common channels thus higher Na+

140
concentration in soil decreased the K+ uptake due to ionic competition (Rains and Epstein,
1997). Similar findings of increased Na+ under salt stress conditions were reported by Saqib
(2000) and Nadeem et al. (2010).
Salinity stress causes not only the disturbances in mineral ion uptake but also their
transport within the plants is altered due to which their natural ratios in different plant parts
are distributed (Glenn et al., 1999). The transport of K+ and Ca2+ to the actively growing plant
parts is inhibited which affects the quality and quantity of produce. Roots of the plant are less
vulnerable to ion toxicity than shoot and leaves as the roots can maintain constant level of
ions by expelling out the extra Na+ and Cl- into soil or into shoot and leaves. So, higher
amounts of these deleterious ions are accumulated in shoots and leaves (Tester and
Davenport, 2003). Sodium causes metabolic toxicity by competing with K+ for binding sites
which is essential for different cellular functions. Potssium is involved in controlling the
activity of more than 50 enzymes and Na+ cannot serve as the alternative of the K+ to carry
out these functions. Thus, higher Na+/K+ cause disturbances in enzyme functions. Higher K+
concentrations are also required for the synthesis of different proteins. Thus, disturbances in
protein synthesis in plants under saline conditions may be caused by Na+ toxicity (Blaha et
al., 2000). Prat and Fathi-Ettai (1990) suggested that salinity stress causes changes in
morphology, ultra-structure and metabolic activities of plant species which ultimately result
in reduction in growth and yield of plants.
Ion homeostasis is the main phenomena which enables the plants to tolerate salinity
stress (Singh et al., 2011). Uptake of higher concentration of K+ as compared to Na+ is vital to
maintain higher K+/Na+ in cells (Kavitha et al., 2012). In our study, we found that
rhizobacterial inoculation /co-inoculation significantly improved the K+/Na+. The interaction
of plant roots with the rhizobacteria reduces the uptake of toxic elements and/or enhances the
uptake of essential nutrients (White, 2003). This might be due to lowered uptake of Na+ than
K+ due to longer roots caused by inoculation/coinoculation with rhizobacterial strains as
reported previously (Yue et al., 2007; Ahmad et al., 2012). Reduction in Na+ concentration
may be due to dilution effect as salt stressed plants inoculated with the rhizobacterial strains
have higher water contents than uninoculated plants. Such salt induced dilution effects have
been reported in different crops e.g. Physalis peviviana (Miranda et al., 2010), Spinacia

141
oleraces (Kaya et al., 2001) and maize (Collado et al., 2010). The increased K+ concentration
have also been correlated with better growth and yield under saline conditions by Giri et al.
(2007). Thus, it can be stated that this improvement in ionic balance caused by rhizobacterial
strains may be one of the reasons of better growth and yield of maize under salt stress.
Membrane permeability (MP) is a good indicator of salt stress and is judged by measuring
electrolyte leakage (Farkhondeh et al., 2012; Mansoor, 2012). In our study, we observed
marked increase in electrolyte leakage in plant subjected to salinity stress. We also observed
that a strong relationship existed between chlorophyll contents and electrolyte leakage. This
type of relationship has been documented in many studies (Chen et al., 1991; Kaya et al.,
2001a). It is likely that membrane integrity somehow is dependent upon chlorophyll contents,
because decrease in chlorophyll contents as caused by salinity stress induces senescence and
membrane become more permeable (Niu et al., 1995; De Araujo et al., 2006)
5.6 Antioxidant enzymes
Under salt stress situations plants accumulate higher levels of antioxidant enzymes
such as SOD, CAT, POD and APX as a defense mechanism to counter oxidative stress
caused by salinity (Munns and Tester, 2008; Ashraf, 2009; Kaye et al., 2011). In our studies
we also found marked increase in antioxidant enzymes under salinity stress. Our results are in
line with other researchers (Apel and Hirt, 2004: Ahmad et al., 2008; Kohler, 2008).
Rhizobacterial inoculation lowers the level of antioxidants (Shahbazi et al., 201; Baniaghil et
al., 2013). Along with production of antioxidant enzymes plants also accumulate some other
compounds such as phenolics and sugars which not only act as osmoregulators but also
scavenge reactive oxygen species (ROS) (Wang et al., 2003; Fernandez et al., 2012;
Theocharis et al., 2012). In our study, we found increase in content of sugars and phenolics
under salt stressed conditions. Our results are in line with Fernendez et al. (2012) who
reported higher sugar contents owing to bacterial inoculation under abiotic stress.
Sandhya et al. (2010) conducted a study to investigate the effect of five drought
tolerant PGPR strains on the growth and antioxidative activity of maize. The results of the
study revealed that the activity of antioxidant enzymes was lower in inoculated plants as
compared with the uninoculated plants. In barley plants the antioxidative activity was also
reduced. A study conducted by Omer et al. (2009) revealed that under salinity stress activity

142
of antioxidant enzymes such as catalase and peroxidase increase significantly in two
cultivars of barley. On the other hand when these plants were inoculated with Azospirillum
baselines, they showed lowering in the activity of such enzymes, which removed the
deleterious effects of salinity and enhanced the growth and yield of barley plants under salt
effected conditions.
5.7 Combined application of L-TRP and rhizobacterial strains
In soil tryptophan dependent biosynthesis of auxin significantly improves the growth and
development of plants (Sarwar et al., 1992). In our study, we observed that sole application of
L-TRP positively improved the maize growth under salt affected soil conditions implying that
native soil microflora was also active in synthesizing auxins from L-TRP or this might be due
to the uptake of L-TRP directly by plant’s root, which have resulted in improved growth
under saline conditions. Hussain et al., (1995) reported the synergistic effect of Rhizobium
inoculation and L-TRP in lentil in terms of 30% more yield than untreated control. The
exogenous provision of phytohormones as a result of microbial activity might affect plant’s
endogenous hormonal levels, either by supplementing the plants own suboptimal levels or by
interacting with the synthesis, translocation, or inactivation of existing hormone levels. Plants
themselves synthesize a diversity of growth-regulating phytohormones but they also respond
to exogenous applications of these plant growth regulators PGRs which, after being taken up
by the plant, may affect its growth directly as growth stimulants or indirectly by modifying
the rhizosphere (Khalid et al., 2006; Zahir et al., 2010). These results are supported by the
work of Datta et al. (1997), who exogenously applied phytohormones and recorded
improvement in wheat growth under salt affected conditions. They found that presoaking
wheat seeds with indole acetic acid mitigated the adverse effects of salinity. We found in our
study that separate application of L-TRP and PGPR strains caused positive effects on plant
growth and development under salt stressed conditions, but their combined application gave
better results regarding growth improvement. The results of Zahir et al. (2000) also support
our findings, which demonstrated that combined use of precursor (L-TRP) and inoculum
(Azotobacter) was better than their sole application. This improvement in growth and yield
can be attributed to the production of plant growth hormone (auxin) in the rhizosphere that
can optimize the internal hormonal level of plant or by increased uptake of essential nutrients

143
due to improved root growth mediated by the phytohormones produced by PGPR. From this
study, it can be suggested that combined application of L-TRP and PGPR strain (precursor-
inoculum interaction) can be very effective technique to induce salt stress tolerance in maize.
5.8 Mechanism used by PGPR for growth promotion in maize grown under salt stress
conditions
Soil salinity is one of the major limiting factors in crop growth and productivity all
over the world (Nemati et al., 2011). Different strategies are in practice to ameliorate the
adverse effects of salinity stress. Among many of the used strategies is the use of plant
growth promoting rhizobacteria (Kohler et al., 2009; Fernandez-Aunion et al., 2010). The
positive effects of such bacteria have been reported by many researchers on different crops
(Creus et al., 1997). In our study we found that both of the rhizozobacterial strains improved
the maize growth under saline conditions. This may be due to the presence of multiple traits
that these strains possessed. Both of the strains were able to coloninize the roots but Strain
M4 had the greater tendency of root colonization than M7 which may be the reason of better
performance of M4 as compared with strain M7 in single inoculation. The combined
application of both strains showed better performance which means that these strains had not
antagonistic effects on each other but developed a synergistic relationship. The enhacement
of maize growth might be due to multiple traits that both of these strains possessed.
Plant growth regulators such as auxins (IAA) are suggested to be involved in the
synthesis/activity of antioxidant enzymes and some of their isoforms also play role in the
metabolism of these growth regulators (Synkova et al., 2004; Tongetti et al., 2012). Reactive
oxygen species are produced at higher level under stress conditions and cause serious damage
to membranes. Despite posing a threat, they also activate a defense mechanism in plants
against stress (Pitzschke et al., 2006). Auxin (IAA) and reactive oxygen species (ROS) play a
critical role in salt stress tolerance. But the actual mechanism that how this cross-talk between
auxins and ROS activate salt tolerance is still to be under stood (Tognetti et al., 2012).
In our study, inoculation with rhizobacterial strains enhanced the growth and yield of
maize most probably due to the production of plant growth hormones such as indole acetic
acid (IAA). Glick (1995) suggested that hormone production is the key mechanism used by

144
the bacterial strains for improving the growth and yield of crop plants. Other researchers have
also documented such growth improvement in plants using the bacterial strains which had the
ability to produce auxins (Fallik et al., 1989; Xie et al., 1996; Dobbelaere et al., 2000; Pati et
al., 1995; Biswas et al., 2000b; Dazzo et al., 2000; Hussain et al., 2009). It is documented that
salinity causes progressively decline in plant hormones (Prakash and Parathapasenan, 1990;
Nilsen and Orcutt, 1996) thus it can be interpreted that reduction in growth and yield caused
by salinity may be due to hormonal imbalance. This imbalance in hormonal level can be
corrected by the exogenous application of hormones (Balki and Padole, 1982; Gulnaz et al.,
1999; Sastry and Shekhawa, 2001; Afzal et al., 2005; Akbari et al., 2007) or by using plant
growth promoting rhizobacteria capable of producing auxins.
Auxin is involved in the transcription of many genes which are known as primary
auxin response genes. For different crops these genes have been identified (Hagen and
Guilfoyle, 2002). Increased production of auxins (IAA) is correlated with root proliferation
which enables the plants to take more water from large soil area (lambrecht et al., 2000;
Steenhoudt and Vanderleyden, 2000). IAA positively affects the root growth directly or
indirectly by lowering the ethylene levels (Dimkpa et al., 2009; Lugtenberg and Kamilova,
2009). Junghans et al. (2006) has also correlated the higher contents of free auxins in
Arabidopsis with root proliferation under salt stress. Bianco and Defez (2009) used IAA
producing rhizobacteria and found that harmful effects of salinity were ameliorated in
Medicago plants.
Plant growth promoting rhizobacteria can affect the growth of plants through the
production of plant growth hormones such as auxins (IAA). These bacteria interact with
plants through the production of auxins (Malhothra and Srivaatava, 2006; Anjum et al.,
2011). In our study, we screened the PGPR on the basis of their auxin production ability and
found that most of the strains were able to produce auxins and this production was even
increased in the presence of L-TRP. Ramiz et al. (1993) have also reported that rhizobacterial
strains produced auxins with varying degrees. Similar to our findings other researchers
(Khalid et al., 2004, 2006; De and Basu, 2007) have reported the auxin production in
rhizobacterial strains.

145
It is highly likely that along with auxin production ability of these strains other
characters of these strains might have helped the plants to tolerate the adverse effects of
salinity. In our laboratory study, we found that all the strains possessed the ACC-deaminase
activity. ACC-deaminase activity in rhizobacterial strains have also been reported by other
researchers (Glick, 2005; Shahroon et al., 2006; Belimov et al., 2009; Nadeem et al., 2009).
Under salinity stress conditions ethylene concentration increased in plants which has
inhibitory effect on plant growth (Ables, 1992). The bacterial strains possessing the ACC-
deaminase activity can lower the ethylene concentration by causing hydrolysis of ACC,
which act as precursor of ethylene (Mayak et al., 2004; Cheng et al., 2007) thus decreasing
the adverse effects of salinity on plants. Many other researchers have reported such
reductions in ethylene by the use of PGPR containing ACC-deaminase activity (Glick, 1998;
Indragandhi et al., 2008; Zahir et al., 2011).
We observed that both the selected strains varied in their growth promotion ability, which
might be due to presence of different other growth promoting characters such as root
colonization, phosphate solubilazation and chitinase activity along with their ability to
produce auxins. These strains possessed phosphate solubilization ability which might have
helped the plants to get extra nutrients. Chitinase activity of these strains might have helped
the plants against their pathogens thus resulting in better growth. Our results are also
supported by the work of Nadeem et al. (2007) who reported that rhizobacterial strains varied
in their ability to improve the plant growth. Salt tolerance in plants also depends on restricted
or controlled uptake of Na+ and continued uptake of K+ (Jeschke and Wolf, 1988). In our
study microbial strains also produced exopolysaccharides. These exopolysaccharides can bind
Na+ (Geddie and Sutherland, 1993), thus decrease the uptake of Na+ in plant and help them in
alleviating salt stress (Ashraf et al., 2004).
Conclusions and future directions
Soil salinity is worldwide problem that poses a serious threat to agricultural production. With
severe and sudden climatic changes this problem is expected to spread at a fast pace. Various
physical, chemical and biological approaches are being used to hammer this problem. But
most of these techniques seem either very expensive or very time consuming. Since near past,

146
soil bacteria have been employed to induce salt stress tolerance in plants. Among such
bacteria are the ones that are capable of producing a hormone, auxin. Many reports document
that endogenous level of plant hormones such as auxin is disturbed due to soil salinization. It
is also well documented that auxin controls the various aspects of plant’s growth and
development. So alteration in hormonal balance is considered one of the main reasons of
growth reduction in plants during salinity stress. The auxin producing rhizobacteria can
supplement the auxin to plants and recover the hormonal alterations caused by salinity. The
results of the present study signify the effectiveness of auxin producing rhizobacteria for
improving the growth and yield of maize in salt affected soils.
However, further research is needed to make the use of this approach more effective in terms
of plant growth enhancement. The future research should be planned giving special emphasis
on these particular aspects,
Population of auxin producing rhizobacteria can be increased by seed or soil
inoculation.
Genetic identification of these strains is needed, so that these strains can be
genetically manipulated for getting better performance
In next phase endophytic auxin producing bacteria be selected to protect the bacterial
population from environmental constraints.
A deep insight into the interaction of these bacteria with the plants can be helpful to
develop statistical models which will help in managing the salt affected soils.
These auxin producing rhizobacteria can be used by seed/fertilizer/pesticide
companies to develop bio-inoculants for maize production in salt affected soils.

147
CHAPTER-6
SUMMARY
Maize (Zea mays) is important cereal crop of the world. Under arid and semiarid
conditions the production of maize is restricted due to number of biotic and abiotic stresses.
Soil salinity is one of the major abiotic stresses that hamper the productivity of the crops
throughout the world especially arid and semiarid regions like those in Pakistan. Soil salinity
adversely affects the growth of plants due to many factors including hormonal imbalance.
Plant growth promoting rhizobacteria have the ability to produce these plant growth
hormones. Inoculation with such bacterial strains can supply adequate amount of plant
growth hormones thus mitigate the adverse effects of salinity. A series of experiments were
carried out to evaluate the beneficial effects of auxin producing PGPR on growth and
performance of maize under salt affected soil conditions.
These experiments and their results are summarized below:
A number of rhizobacterial strains were isolated from the rhizosphere of the maize
grown under salt affected fields.
The isolated strains were subjected to osmoadaptation assay to test their ability to
withstand the salinity stress. The rhizobacterial strains were grown at four salinity
levels i.e. control, 10, 20 and 30 dS m-1 and the optical density was measured. The
results showed that all the strains varied in their ability to withstand salinity.
Twenty selected strains were accessed for their auxin production ability. All the
strains produced varied amounts of indole acetic acid (IAA) in the presence and
absence of L-TRP.
The rhizobacterial strains producing auxin were screened for their ability to promote
growth of maize seedlings under axenic conditions. The results indicated that salinity
treatment significantly reduced the growth of maize seedlings but inoculation with
auxin producing PGPR improved the growth of maize seedlings. These strains varied
in their growth promotion ability.

148
Two most effective strains producing auxin were selected for pot and field
experiments to evaluate their potential for improving the growth and yield of maize in
the presence and absence of L-TRP under salt affected soil conditions. The results of
these studies are summarized below
a) The growth and yield of maize was adversely affected by the salinity stress under pot
and field experiments.
b) The rhizobacterial inoculation alone and in combination significantly improved the
growth and yield of maize under salt affected soil conditions both in the presence and
absence of L-TRP. The results of combined application of bacterial strains along with
L-TRP application were more pronounced than sole application of strains.
c) Physiological parameters like carbon dioxide assimilation, stomatal conductance,
relative water contents, transpiration rate and photosynthetic rate etc. were decreased
by salinity treatment while inoculation with rhizobacterial strains significantly
improved these parameters in the presence and absence of L-TRP in both pot and field
experiments.
d) Salinity stress caused ionic imbalance in the plants and decreased the K+/Na+. The
rhizobacterial inoculation increased the K+ concentration and decreased the Na+ thus
improved the K+ to Na+ ratio in plants.
e) Concentration of osmolytes such as proline and soluble sugars increased under salinity
stress. However, inoculation with auxin producing rhizobacteria decreased the
concentration of these osmolytes in plants.
f) Mineral nutrient concentration was adversely by the salinity as the concentration of N,
P and K decreased under salinity stress. But the concentration of mineral nutrients was
significantly improved by the rhizobacterial inoculation. The concentration of N, P, K
and protein contents was increased due to inoculation thus improving the quality of
produce along with an increased grain yield of maize.
g) Antioxidant activity of the plants was increased under salinity stress, however,
rhizobacterial inoculation significantly lowered the activity of such enzymes showing
their ability to mitigate salinity stress.

149
h) The strains varied in their ability to tolerate the salinity stress. The strains showing
more growth at higher salinity levels also performed better for improving growth and
yield of maize under salinity stress.
i) The strains having more root colonization ability and auxin production ability along
with other characteristics also performed better in improving the growth and yield of
maize under saline conditions.
j) Combined application of auxin producing rhizobacterial strains along with L-TRP can
be an economic option for improving the growth and yield of maize under salt
affected soil conditions.
On overall basis combined application of rhizobacterial strains producing auxin
showed better results than their sole inoculation. The results of the inoculation were
more pronounced when it was carried out along with L-TRP application. Such
bacteria could be very effective as inoculants to improve the growth and yield of
maize under salt affected soil conditions. These results of these experiments support
our hypothesis that auxin producing rhizobacteria can improve maize growth in saline
conditions.

150
Literature cited
Abeles, F.B., P.W. Morghan and M.E. Saltveit Jr. 1992. Ethylene in plant biology. Academic
Press, London, UK.
Abrol, I.P., J.S.P. Yadav and F.I. Massoud. 1988. Salt-affected soils and their management.
Food and Agriculture Org.
Afroz, S., F. Mohammad, S. Hayat and M.H. Siddiqui. 2005. Exogenous Application of
Gibberellic Acid Counteracts the Ill Effect of Sodium Chloride in Mustard. Turk. J.
Biol. 29(4): 744-752.
Afzal, I., S.M.A. Basra and A. Iqbal. 2005. The effects of seed soaking with plant growth
regulators on seedling vigor of wheat under salinity stress. J. Stress Physiol. Biochem.
1(1): 6-14.
Ahmad, F., I. Ahmad and M.S. Khan. 2008. Screening of free-living rhizospheric bacteria for
their multiple plant growth promoting activities. Microbiol. Res. 163(2): 173-181.
Akhiyarova, G.R., I.B. Sabirzhanova, D.S. Veselov and V. Frike. 2005. Participation of plant
hormones in growth resumption of wheat shoots following short-term NaCl treatment.
Russ. J. Plant Physiol. 52(6): 788-792.
Ali, B., S.A. Hasan, S. Hayat, Q. Hayat, S. Yadav, Q. Fariduddin and A. Ahmad. 2008. A
role for brassinosteroids in the amelioration of aluminium stress through antioxidant
system in mung bean (Vigna radiata L.). Environ. Exp. Bot. 62(2): 153-159.
Anjum, S.A., X.Y. Xie, L.C. Wang, M.F. Saleem, C. Man and W. Lei. 2011. Morphological,
physiological and biochemical responses of plants to drought stress. Afr. J. Agric.
Res. 6(9): 2026-2032.
Anonymous, 2008. Regulation of Setting Maximum Levels for Certain Contaminants in
Foodstuffs Official Gazette (26879).
Apel, K. and H. Hirt. 2004. Reactive oxygen species: metabolism, oxidative stress, and signal
transduction. Annul. Rev. Plant Biol. 55: 373-399.

151
Arshad, M., B. Shaharoona and T. Mahmood. 2008. Inoculation with Pseudomonas spp.
Containing ACC-Deaminase Partially Eliminates the Effects of Drought Stress on
Growth, Yield, and Ripening of Pea (Pisum sativum L.). Pedosphere. 18(5): 611-620.
Ashraf, M. 2002. Salt tolerance of cotton: some new advances. Crit. Rev. Plant Sci. 21(1): 1-
30.
Ashraf, M. 2004. Some important physiological selection criteria for salt tolerance in plants.
Flora-Morphology, Distribution, Functional Ecology of Plants. 199(5): 361-376.
Ashraf, M. and N.A. Akram. 2009. Improving salinity tolerance of plants through
conventional breeding and genetic engineering: an analytical comparison. Biotec.
Adv. 27(6): 744-752.
Ashraf, M. and T. McNeilly. 1990. Responses of four Brassica species to sodium chloride.
Environ. Exp. Bot. 30(4): 475-487.
Ashraf, M. and T. McNeilly. 2004. Salinity tolerance in Brassica oilseeds. Crit. Rev. Plant
Sci. 23(2): 157-174.
Ashraf, M. and L. Wu. 1994. Breeding for Salinity Tolerance in Plants. Crit. Rev. Plant Sci.
13(1): 17-42.
Azooz, M.M. 2004. Proteins, sugars and ion leakage as a selection criterion for the salt
tolerance of three sorghum cultivars at seedling stage grown under NaCl and
nicotinamide. Int. J. Agric. Biol. 6: 27-35.
Badenoch-Jones, J., B.G. Rolfe and D.S. Letham. 1983. Phytohormones, Rhizobium Mutants,
and Nodulation in Legumes III. Auxin Metabolism in Effective and Ineffective Pea
Root Nodules. Plant Physiol. 73(2): 347-352.
Baligar, V.C., N.K. Fageria and Z.L. He. 2001. Nutrient use efficiency in plants. Commun.
Soil Sci. Plant Anal. 32(7-8): 921-950.
Balki, A.S. and V.R. Padole. 1982. Effect of Pre-soaking Seed Treatments with Plant
Hormones on Wheat under Conditions, of Soil Salinity. J. Ind. Soc. Soil Sci. 30(3):
361-365.

152
Baniaghil, N., M.H. Arzanesh, M. Ghorbanli and M. Shahbazi. 2013. The effect of plant
growth promoting rhizobacteria on growth parameters, antioxidant enzymes and
microelements of canola under salt stress. J. Appl. Environ. Biol. Sci. 3(1): 17-27.
Bartel, B. 1997. Auxin biosynthesis. Ann. rev. plant bio. 48(1): 51-66.
Basu, P.S. and A.C. Ghosh. 1998. News and Notes Indole Acetic Acid and Its Metabolism in
Root Nodules of a Monocotyledonous Tree Roystonea regia. Curr. microbial. 37(2):
137-140.
Bates, L.S., R.P. Waldren and I.D. Teare. 1973. Rapid determination of free proline for
water-stress studies. Plant Soil. 39(1): 205-207.
Bellamine, J., C. Penel, H. Greppin and T. Gaspar. 1998. Confirmation of the role of auxin
and calcium in the late phases of adventitious root formation. Plant Growth Regul.
26(3): 191-194.
Ben-Asher, J., I. Tsuyuki, B.A. Bravdo and M. Sagih. 2006. Irrigation of grapevines with
saline water: I. Leaf area index, stomatal conductance, transpiration and
photosynthesis. Agric. water manage. 83(1): 13-21.
Bhowmick, P.K. and P.S. Basu. 1986. Production of indole acetic acid by Rhizobium sp.
from root nodules of leguminous tree Sasbania grandiflora Pers. Acta Microbiol. Pol.
35(3): 281-290.
Bhowmick, P.K. and P.S. Basu 1987. Indoleacetic acid production by Rhizobium sp. from a
leguminous tree, Erythrina indica. Folia microbiol. 32(2): 142-148.
Bianco, C. and R. Defez. 2009. Medicago truncatula improves salt tolerance when nodulated
by an indole-3-acetic acid-overproducing Sinorhizobium meliloti strain. J. Exp. Bot.
60(11): 3097-3107.
Bianco, C. and R. Defez. 2010. Improvement of phosphate solubilization and Medicago plant
yield by an indole-3-acetic acid-overproducing strain of Sinorhizobium meliloti. Appl.
Environ. Microbiol. 76(14): 4626-4632.
Bohnert, H.J. and E. Sheveleva. 1998. Plant stress adaptations-making metabolism move.
Curr. Opin. Plant Biol. 1(3): 267-274.

153
Booth, W.A. and J. Beardall. 1991. Effects of salinity on inorganic carbon utilization and
carbonic anhydrase activity in the halotolerant alga Dunaliella salina (Chlorophyta).
Phycologia. 30(2): 220-225.
Boucaud, J. and I.A. Ungar. 1976. Hormonal control of germination under saline conditions
of three halophytic taxa in the genus Suaeda. Physiol. Plantarum. 37(2): 143-148.
Burling, K., Z.G. Cerovic, G. Cornic, J.M. Ducruet, G. Noga, and M. Hunsche. 2013.
Fluorescence-based sensing of drought-induced stress in the vegetative phaseof four
contrasting wheat genotypes. Environ. Exp. Bot. 89: 51-59.
Cakmak, I. 2002. Plant nutrition research: Priorities to meet human needs for food in
sustainable ways, Progress in Plant Nutrition: Plenary Lectures of the XIV
International Plant Nutrition Colloquium. Springer, pp. 3-24.
Camerini, S., B. Senatore, E. Lonardo, E. Imperlini, C. Bianco, G. Moschetti, G.L. Rotino, B.
Campion and R. Defez. 2008. Introduction of a novel pathway for IAA biosynthesis
to rhizobia alters vetch root nodule development. Arch. Microbial. 190(1): 67-77.
Carden, D.E., D.J. Walker, T.J. Flowers and A.J. Miller. 2003. Single-cell measurements of
the contributions of cytosolic Na+ and K+ to salt tolerance. Plant Physiol. 131(2): 676-
683.
Carvajal, M., V. Martnez and A. Cerd, 1999. Influence of magnesium and salinity on tomato
plants grown in hydroponic culture. J. Plant Nutr. 22(1): 177-190.
Chartzoulakis, K., M. Loupassaki, M. Bertaki and I. Androulakis, 2002. Effects of NaCl
salinity on growth, ion content and CO2 assimilation rate of six olive cultivars. Sci.
Hortic. 96(1): 235-247.
Chavez, F.P., M. Messi and J.T. Pennington. 2011. Marine primary production in relation to
climate variability and change. Ann. Rev. Mari. Sci. 3: 227-260.
Chedlia, B.A., B.R. Bechi. and M. Boukhris. 2007. Effect of water deficit onolive trees cv.
Chemlali under field conditions in arid region inTunisia. Sci. Hortic. (1): 235-247.

154
Cohen, A.C., R.n. Bottini and P.N. Piccoli, 2008. Azospirillum brasilense Sp 245 produces
ABA in chemically-defined culture medium and increases ABA content in arabidopsis
plants. Plant Growth Regul. 54(2): 97-103.
Collado, M.B., M.J. Arturi, M.B. Aulicino and M.C. Molina. 2011. Identification of salt
tolerance in seedling of maize (Zea mays L.) with the cell membrane stability trait.
Int. Res. J. Plant Sci. 1(5): 126-132.
Costacurta, A. and J. Vanderleyden. 1995. Synthesis of phytohormones by plant-associated
bacteria. Crit. Rev. microbiol. 21(1): 1-18.
Darra, B.L. and S.N. Saxena. 1973. Role of IAA on the mineral composition of maize (Zea
mays) crop under various osmotic stressed conditions. Plant Soil. 38(3): 657-661.
Dashti, N., F. Zhang, R. Hynes and D.L. Smith. 1998. Plant growth promoting rhizobacteria
accelerate nodulation and increase nitrogen fixation activity by field grown soybean
(Glycine max L.) under short season conditions. Plant Soil. 200(2): 205-213.
Datta, S.R., H. Dudek, X. Tao, S. Masters, H. Fu, Y. Gotoh and M.E. Greenberg. 1997. Akt
phosphorylation of BAD couples survival signals to the cell-intrinsic death
machinery. Cell. 91(2): 231-241.
Davies, P.F. 1995. Flow-mediated endothelial mechanotransduction. Phys. Rev. 75(3): 519.
Debez, A., W. Chaibi and S. Bouzid. 2001. Effect du NaCl et de regulatoeurs de croissance
sur la germination d’ Atriplex halimus L. Cah. Agric. 10:135-138.
De Araujo, I.E., R. Gutierrez, A.J. Oliveira-Maia, A. Pereira Jr, M.A.L. Nicolelis and S.A.
Simon, 2006. Neural ensemble coding of satiety states. Neuron. 51(4): 483-494.
De Lacerda, C.F., J. Cambraia, M.A. Oliva, H.A. Ruiz and J.T.Ì.n. Prisco. 2003. Solute
accumulation and distribution during shoot and leaf development in two sorghum
genotypes under salt stress. Environ. Exp. Bot. 49(2): 107-120.
De Salamone, I.E.G., R.K. Hynes, R.M. Nelson. 2001. Cytokinin production by plant growth
promoting rhizobacteria and selected mutants. Can. J. Microbiol. 47: 404-411.

155
Dhingra, H.R. and T.M. Varghese. 1985. Effect of growth regulators on the in vitro
germination and tube growth of maize (Zea mays) pollen from plants raised under
sodium chloride salinity. New phytol. 100(4): 563-569.
Dimkpa, C.O., D. Merten, A. Svatoa, G. Bchel and E. Kothe. 2009. Metal-induced oxidative
stress impacting plant growth in contaminated soil is alleviated by microbial
siderophores. Soil Biol. Biochem. 41(1): 154-162.
Dionisio-Sese, M.L. and S. Tobita 1998. Antioxidant responses of rice seedlings to salinity
stress. Plant Sci. 135(1): 1-9.
Dobbelaere, S., A. Croonenborghs, A. Thys, A.V. Broek and J. Vanderleyden. 1999.
Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains
altered in IAA production on wheat. Plant Soil. 212(2): 153-162.
Dodd, I.C., A.A. Belimov, W.Y. Sobeih, V.I. Safronova, D. Grierson and W.J. Davies. 2004.
Will modifying plant ethylene status improve plant productivity in water-limited
environments, Proceedings for the 4th International Crop Science Congress, Brisbane,
Australia.
Dullaart, J., 1970. The auxin content of root nodules and roots of Alnus glutinosa (L.). J. Exp.
Bot. 21(4): 975-984.
Duncan, D.B. 1955. Multiple range and multiple F tests. Biometrics.
Dunlap, J.R. and M.L. Binzel. 1996. NaCI reduces indole-3-acetic acid levels in the roots of
tomato plants independent of stress-induced abscisic acid. Plant Physiol. 112(1): 379-
384.
Egamberdieva, D. 2009. Alleviation of salt stress by plant growth regulators and IAA
producing bacteria in wheat. Acta Physiol. Plant. 31(4): 861-864.
Egamberdieva, D. and Z. Kucharova. 2009. Selection for root colonising bacteria stimulating
wheat growth in saline soils. Bio. Fert. Soils. 45(6): 563-571.
Ehsanpour, A.A. and F. Amini. 2003. Effect of salt and drought stress on acid phosphatase
activities in alfalfa (Medicago sativa L.) explants under in vitro culture. Afr. J.
Biotech. 2(5): 133-135.

156
El-Khawas, H. and K. Adachi. 1999. Identification and quantification of auxins in culture
media of Azospirillum and Klebsiella and their effect on rice roots. Bio. Fert. Soil.
28(4): 377-381.
El-Komy, H.M. 1998. Effect of simultaneous inoculation of Azospirillum and Rhizobium
spp. on growth, nodulation and nitrogen fixation of two legumes using the 15 N-
isotope dilution technique [IDT] and the difference method [DM]. Acta Microb. Pol.
47(3):77-82.
Elsheikh, E.A.E. and M. Wood. 1990. Effect of salinity on growth, nodulation and nitrogen
yield of chickpea (Cicer arietinum L.). J. Exp. Bot. 41(10): 1263-1269.
Endris, S. and M.J. Mohammad. 2007. Nutrient acquisition and yield response of Barley
exposed to salt stress under different levels of potassium nutrition. Int. J. Env. Sci.
Technol. 4(3): 323-330.
Ernstsen, A., G. Sandberg, A. Crozier and C.T. Wheeler. 1987. Endogenous indoles and the
biosynthesis and metabolism of indole-3-acetic acid in cultures of Rhizobium
phaseoli. Planta. 171(3): 422-428.
Erum, S. and A. Bano. 2008. Variation in phytohormone production in Rhizobium strains at
different altitudes of northern areas of Pakistan. Int. J. Agri. Biol. 10: 536-40.
Fernandez-Aunion, C., T. Hamouda, F. Iglesias-Guerra, M. Argandoa, M. Reina-Bueno, J.
Nieto, M.E. Aouani and C. Vargas. 2010. Biosynthesis of compatible solutes in
rhizobial strains isolated from Phaseolus vulgaris nodules in Tunisian fields.
Molecular Plant-Microbe Interaction. 10(1): 192.
Flowers, T.J. and A.R. Yeo. 1995. Breeding for salinity resistance in crop plants: where next?
Funct. Plant Biol. 22(6): 875-884.
Foyer, C.H. and G. Noctor. 2003. Redox sensing and signalling associated with reactive
oxygen in chloroplasts, peroxisomes and mitochondria. Phys. Plant. 119(3): 355-364.
Frankenberger Jr, W.T. and M. Arshad. 1995. Phytohormones in soils: microbial production
and function. Marcel Dekker Inc.

157
Gadallah, M.A.A. 2000. Effects of indole-3-acetic acid and zinc on the growth, osmotic
potential and soluble carbon and nitrogen components of soybean plants growing
under water deficit. J. Ar. Env. 44(4): 451-467.
García de Salamone, I.E., R.K. Hynes and L.M. Nelson. 2001. Cytokinin production by plant
growth promoting rhizobacteria and selected mutants. Can. J. Microb. 47(5): 404-411.
Geddie, J.L., and I.W. Sutherland. 1993. Uptake of metals by bacterial polysaccharides.
J. Appl. Bacteriol. 74:467-472.
Ghosh, S., C. Sengupta, T.K. Maiti and P.S. Basu. 2008. Production of 3-indolylacetic acid in
root nodules and culture by a Rhizobium species isolated from root nodules of the
leguminous pulse Phaseolus mungo. Folia Microb. 53(4): 351-355.
Gill, S.S. and N. Tuteja. 2010. Reactive oxygen species and antioxidant machinery in abiotic
stress tolerance in crop plants. Plant Phys. Biochem. 48(12): 909-930.
Giri, S.K. and S. Prasad. 2007. Drying kinetics and rehydration characteristics of microwave-
vacuum and convective hot-air dried mushrooms. J. Food Eng. 78(2): 512-521.
Girija, C., B.N. Smith and P.M. Swamy. 2002. Interactive effects of sodium chloride and
calcium chloride on the accumulation of proline and glycinebetaine in peanut (Arachis
hypogaea L.). Environ. Exp. Bot. 47(1): 1-10.
Glick, B.R. 1995. The enhancement of plant growth by free-living bacteria. Can.J. Microbiol.
41(2): 109-117.
Glick, B.R. 2005. Modulation of plant ethylene levels by the bacterial enzyme ACC
deaminase. FEMS Microb. Let. 251(1): 1-7.
Glick, B.R., Z. Cheng, J. Czarny and J. Duan. 2007. Promotion of plant growth by ACC
deaminase-producing soil bacteria, New Perspectives and Approaches in Plant
Growth-Promoting Rhizobacteria Research. Springer, pp. 329-339.
Glick, B.R., C. Liu, S. Ghosh and E.B. Dumbroff. 1997. Early development of canola
seedlings in the presence of the plant growth-promoting rhizobacterium Pseudomonas
putida GR12-2. Soil Bio. Biochem. 29(8): 1233-1239.

158
Glick, B.R., D.M. Penrose and J. Li. 1998. A model for the lowering of plant ethylene
concentrations by plant growth-promoting bacteria. J. Theo. Biol. 190(1): 63-68.
Gouia, H., M.H. Ghorbal and B. Touraine. 1994. Effects of NaCl on flows of N and mineral
ions and on NO3-reduction rate within whole plants of salt-sensitive bean and salt-
tolerant cotton. Plant Physiol. 105(4): 1409-1418.
Grattan, S.R. and C.M. Grieve. 1998. Salinity-mineral nutrient relations in horticultural crops.
Sci. Hort. 78(1): 127-157.
Gravel, V., H. Antoun and R.J. Tweddell. 2007. Growth stimulation and fruit yield
improvement of greenhouse tomato plants by inoculation with Pseudomonas putida
Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem.
39(8): 1968-1977.
Gray, E.J. and D.L. Smith. 2005. Intracellular and extracellular PGPR: commonalities and
distinctions in the plant–bacterium signaling processes. Soil Biol. Biochem. 37: 395-
412.
Greenway, H. and R. Munns. 1980. Mechanisms of salt tolerance in nonhalophytes. Ann.
Rev. plant physiol. 31(1): 149-190.
Gruhn, P., F. Goletti and M. Yudelman. 2000. Integrated nutrient management, soil fertility,
and sustainable agriculture: current issues and future challenges. Intl Food Policy Res
Inst.
Gul, N., Z.A. Swati, S.M.S. Naqvi and I.U.A. Quraishi. 2000. Magnitude of Somaclonal
variation in Oryza sativa L. cvs. Basmati-385, JP-5, Pakhal and Swat-II. Plant Tissue
Cult. 31(1): 149-190.
Gulnaz, A., J. Iqbal, S. Farooq and F. Azam. 1999. Seed treatment with growth regulators and
crop productivity. I. 2, 4-D as an inducer of salinity-tolerance in wheat (Triticum
aestivum L.). Plant soil. 210(2): 209-218.
Hagen, G. and T. Guilfoyle. 2002. Auxin-responsive gene expression: genes, promoters and
regulatory factors. Plant Mol. Biol. 49(3-4): 373-385.

159
Hajibagheri, M.A. and T.J. Flowers. 1989. X-ray microanalysis of ion distribution within root
cortical cells of the halophyte Suaeda maritima (L.) Dum. Planta. 177(1): 131-134.
Halda-Alija, L. 2003. Identification of indole-3-acetic acid producing freshwater wetland
rhizosphere bacteria associated with Juncus effusus L. Can. J. Microb. 49(12): 781-
787.
Halliwell, B. and J. Gutteridge. 1989. Role of free radicals and catalytic metal ions in human
disease: an overview. Method Enzymol. 186: 1-85.
Han, H.S. and K.D. Lee. 2005. Plant growth promoting rhizobacteria effect on antioxidant
status, photosynthesis, mineral uptake and growth of lettuce under soil salinity. Res. J.
Agric. Biol. Sci. 1(3): 210-215.
Han, S.H. and J.H. Lee. 2005b. An overview of peak-to-average power ratio reduction
techniques for multicarrier transmission. Wireless Commun. IEEE, 12(2): 56-65.
Hare, P.D. and W.A. Cress. 1997. Metabolic implications of stress-induced proline
accumulation in plants. Plant Gr. Reg. 21(2): 79-102.
Hariprasad, P. and S.R. Niranjana. 2009. Isolation and characterization of phosphate
solubilizing rhizobacteria to improve plant health of tomato. Plant Soil. 316(1-2): 13-
24.
Hasegawa, M., S. Sone, S. Takashima, F. Li, Z.G. Yang, Y. Maruyama and T. Watanabe.
2000. Growth rate of small lung cancers detected on mass CT screening. Growth, 73
(876).
Hasegawa, N., N. Yamda and M. Mori. 2003. Powdered green tea has antilipogenic effect on
Zucker rats fed a high fat diet. Phyto. Res. 17(5): 477-480.
Hatami, T., M.A.A. Meireles and G. Zahedi. 2010. Mathematical modeling and genetic
algorithm optimization of clove oil extraction with supercritical carbon dioxide. J.
Supercrit. Fl. 51(3): 331-338.
Hayat, Q., S. Hayat, M. Irfan and A. Ahmad. 2010. Effect of exogenous salicylic acid under
changing environment: a review. Environ. Exp. Bot. 68(1): 14-25.

160
Heidari, M. and P. Jamshid. 2010. Interaction between salinity and potassium on grain yield,
carbohydrate content and nutrient uptake in pearl millet. J. Agr. Biol. Sci. 5: 39-46.
Hinsinger, P. 2001. Bioavailability of soil inorganic P in the rhizosphere as affected by root-
induced chemical changes: a review. Plant soil. 237(2): 173-195.
Hirsch, A.M. and Y. Fang. 1994. Plant hormones and nodulation: what's the connection?
Plant Mol. Boil. 26(1): 5-9.
Hoagland, D.R. and D.I. Arnon. 1950. The water-culture method for growing plants without
soil. Circular. California Agricultural Experiment Station. 347(2nd edit).
Hontzeas, N., S.S. Saleh and B.R. Glick. 2004. Changes in gene expression in canola roots
induced by ACC-deaminase-containing plant-growth-promoting bacteria. Appl.
Environ. Microbiol. 71(11): 7556-7558.
Hsiao, T.C. and L.K. Xu. 2000. Sensitivity of growth of roots versus leaves to water stress:
biophysical analysis and relation to water transport. J. Exp. Bot. 51(350): 1595-1616.
Hua, S.S.T.,V.Y. Tsai, G.M. Lichesn and A.T. Noma. 1982. Accumulation of amino acids in
Rhizobium sp. Strain WR1OO1 in response to NaCl salinity. Appl. Environ.
Microbiol. 44: 135-140.
Hunter, W.J. 1989. Indole acetic acid production by bacteroids from soybean root nodules.
Phys. Plant. 76(1): 31-36.
Hussain, I., M. Arshad, Z. A. Zahir, M. Javed, and A. Hussain. 1995. Substrate dependent
microbial production of auxins and their influence on the growth and nodulation of
lentil. Pakistan J. Agric. Sci. 32: 149-152.
Hutcheson, S.W. and T. Kosuge. 1985. Regulation of 3-indoleacetic acid production in
Pseudomonas syringae pv. savastanoi. Purification and properties of tryptophan 2-
monooxygenase. J. Biol. Chem. 260(10): 6281-6287.
Idris, E.E., D.J. Iglesias, M. Talon and R. Borriss. 2007. Tryptophan-dependent production of
indole-3-acetic acid (IAA) affects level of plant growth promotion by Bacillus
amyloliquefaciens FZB42. Mol. Plant-Micr. Inter. 20(6): 619-626.

161
Itai, C., A.E. Richmond and Y. Vaadia. 1968. The role of root cytokines during water and
salinity stress. Israel J. of Bot. 26(1): 5-9.
J., B.-J., S. R.E., D. M.A., J. Shine, D.S. Letham and B.J. Rolfe. 1982. Mass-spectrometric
quantification of indole-3-acetic and in culture supernatants of Rhizobium strains;
studies in relation to root hair curling and nodule initiation. . J. Appl. Environ.
Microbiol. 44(2 ): 275-280.
Jackson, K.A., 1962. On the origin of dislocations. Phil. Mag. 7(81): 1615-1616.
Jacobsen, T. and R.M. Adams, 1958. Salt and silt in ancient Mesopotamian agriculture.
Science. 128(3334): 1251-1258.
Jambunathan, N. 2010. Determination and detection of reactive oxygen species (ROS), lipid
peroxidation, and electrolyte leakage in plants. In: Sunkar R (edu.) Plant stress
tolerance, methods in molecular biology 639. Humana press, New York, pp 291-297.
Jamil, M., L. Deog Bae, J. Kwang Yong, M. Ashraf, L. Sheong Chun and R. Eui Shik. 2006.
Effect of salt (NaCl) stress on germination and early seedling growth of four
vegetables species. J. Cent. Eur. Agr. 7(2): 273-282.
Jeschke, W.D., and O. Wolf. 1988. External potassium supply is not required for root growth
in saline conditions: Experiments with Ricinus communis L. grown in a reciprocal
split-root system. J. Exp. Bot. 39:1149-1167.
Jha, Y., R.B. Subramanian and S. Patel. 2011. Combination of endophytic and rhizospheric
plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of
osmoprotectant against saline stress. Acta Phys. Plant. 33(3): 797-802.
Jones, G.W. and J. Gorham. 2002. Intra-and inter-cellular compartmentation of ions, Salinity:
environment-plants-molecules. Springer, pp. 159-180.
Junghans, M., T. Backhaus, M. Faust, M. Scholze and L.H. Grimme. 2006. Application and
validation of approaches for the predictive hazard assessment of realistic pesticide
mixtures. Aq. Toxicol. 76(2): 93-110.
Kabar, K. 1987. Alleviation of salinity stress by plant growth regulators on seed germination.
J. Plant physiol. 128(1): 179-183.

162
Kaneshiro, T. and W.F. Kwolek. 1985. Stimulated nodulation of soybeans by Rhizobium
japonicum mutant (B-14075) that catabolizes the conversion of tryptophan to indole-
3-acetic acid. Plant Sci. 42(3): 141-146.
Kaneshiro, T. and J.J. Nicholson. 1990. Bradyrhizobium japonicum subspecies (USDA 110
and 26) characterized by fixed-nitrogen uptake and symbiotic indoleacetic acid
production. Curr. Microbiol. 20(6): 381-385.
Kaneshiro, T., M.E. Slodki and R.D. Plattner. 1983. Tryptophan catabolism to indolepyruvic
and indoleacetic acids byRhizobium japonicum L-259 mutants. Cur. Microbial. 8(5):
301-306.
Kang, B.G., W.T. Kim, H.S. Yun and S.C. Chang. 2010. Use of plant growth-promoting
rhizobacteria to control stress responses of plant roots. Plant Biotech. Rep. 4(3): 179-
183.
Karimi, M., B.r. De Meyer and P. Hilson. 2005. Modular cloning in plant cells. Trend plant
Sci. 10(3): 103-105.
Kausar, R. and S.M. Shahzad. 2006. Effect of ACC-deaminase containing rhizobacteria on
growth promotion of maize under salinity stress. J. Agric. Soc. Sci. 2(4): 216-218.
Kavi Kishor, P.B., S. Sangam and R.N. Amrutha. 2005. Regulation of proline biosynthesis,
degradation, uptake and transport in higher plants: Its implications in plant growth and
abiotic stress tolerance. Curr. Sci. 88: 424-438.
Kavitha, P.G., A.J. Miller, M.K. Mathew and F.J.M. Maathuis. 2012. Rice cultivars with
differing salt tolerance contain similar cation channels in their root cells. J. Exp. Bot.
63(8): 3289-3296.
Kazan, K. and J.M. Manners. 2009. Linking development to defense: auxin in plant pathogen
interactions. Trend. Plant Sci. 14(7): 373-382.
Kerr, I. and M. Bennett. 2007. New insight into the biochemical mechanisms regulating auxin
transport in plants. Biochem. J. 401: 613-622.

163
Khaliq, T., T. Mahmood and A.Masood. 2004. Effectiveness of farmyard manure, poultry
manure and nitrogen for corn (Zea mays) productivity. Inter. J. Agric. Biol. 2: 260-
263.
Khan, J., J.S. Wei, M. Ringner, L.H. Saal, M. Ladanyi, F. Westermann, F. Berthold, M.
Schwab, C.R. Antonescu and C. Peterson. 2001. Classification and diagnostic
prediction of cancers using gene expression profiling and artificial neural networks.
Nature med. 7(6): 673-679.
Khan, M.A. and D.J. Weber. 1986. Factors influencing seed germination in Salicornia
pacifica var. utahensis. Amer. J. Bot. 7(6): 1163-1167.
Khan, S.A., A. Gnther, M.A. Schmidt and K.F. Jensen. 2004. Microfluidic synthesis of
colloidal silica. Langmuir, 20(20): 8604-8611.
Khayyat, M., E. Tafazoli, S. Eshghi, M. Rahemi and S. Rajaee. 2007. Salinity, supplementary
calcium and potassium effects on fruit yield and quality of strawberry (Fragaria
ananassa). Amer. Eurasian J. Agri. Env. Sci. 2: 539-544.
Kirst, H.A. and G.D. Sides. 1989. New directions for macrolide antibiotics: pharmacokinetics
and clinical efficacy. Antimicrob. Agent Chemo. 33(9): 1419-1422.
Kloepper, J.W. and M.N. Schroth. 1978. Plant growth-promoting rhizobacteria on radishes,
Proceedings of the 4th international conference on plant pathogenic bacteria, pp. 879-
882.
Kohler, J., F. Caravaca and A. Roldan. 2010. An AM fungus and a PGPR intensify the
adverse effects of salinity on the stability of rhizosphere soil aggregates of Lactuca
sativa. Soil Biol. Biochem. 42(3): 429-434.
Kohler, J., J.A. Hernndez, F. Caravaca and A. Roldn. 2008. Plant-growth-promoting
rhizobacteria and arbuscular mycorrhizal fungi modify alleviation biochemical
mechanisms in water-stressed plants. Funct. Plant Biol. 35(2): 141-151.
Kohler, J., J.A. Hernandez, F. Caravaca and A. Rold. 2009. Induction of antioxidant enzymes
is involved in the greater effectiveness of a PGPR versus AM fungi with respect to
increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 65(2): 245-
252.

164
Kraft, M., R. Kuglitsch, J. Kwiatkowski, M. Frank and K. Grossmann. 2007. Indole-3-acetic
acid and auxin herbicides up-regulate 9-cisepoxycarotenoid dioxygenase gene
expression and abscisic acid accumulation in cleavers (Galium aparine): interaction
with ethylene. J. Exp. Bot. 58: 1497-503.
Lambrecht, M., Y. Okon, A. Vande Broek and J. Vanderleyden. 2000. Indole-3-acetic acid: a
reciprocal signalling molecule in bacteria plant interactions. Trend. Microbiol. 8(7):
298-300.
Larkindale, J. and M.R. Knight. 2002. Protection against heat stress-induced oxidative
damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid.
Plant Physiol. 128(2): 682-695.
Lauter, D.J., D.N. Munns and K.L. Clarkin. 1981. Salt response of chickpea as influenced by
N supply. Agron. J. 73(6): 961-966.
Lawlor, D.W. 2002. Limitation to Photosynthesis in Water-stressed Leaves: Stomata vs.
Metabolism and the Role of ATP. Annal. Bot. 89(7): 871-885.
Lawlor, D.W. and G. Cornic. 2002. Photosynthetic carbon assimilation and associated
metabolism in relation to water deficits in higher plants. Plant cell Environ. 25(2):
275-294.
Lifshitz, R., J.W. Kloepper, M. Kozlowski, C. Simonson, J. Carlson, E.M. Tipping and I.
Zaleska. 1987. Growth promotion of canola (rapeseed) seedlings by a strain of
Pseudomonas putida under gnotobiotic conditions. Can. J. Microbiol. 33(5): 390-395.
Lloret, J., L. Bolanos, M.M. Lucas, J.M. Peart, N.J. Brewin, I. Bonilla and R. Rivilla. 1995.
Ionic stress and osmotic pressure induce different alterations in the lipopolysaccharide
of a Rhizobium meliloti strain. Appl. Environ. Microbiol. 61(10): 3701-3704.
Lucy, M., E. Reed, B.R. Glick. 2004. Applications of free living plant growth-promoting
rhizobacteria. Antonie van Leeuwenhoek. 86: 1-25.
Ludwig-Muller, J., 2011. Auxin conjugates: their role for plant development and in the
evolution of land plants. J. Exp. Bot. 62(6): 1757-1773.

165
Lugtenberg, B.J.J. and F.D. Kamilova. 2004. Rhizosphere management: microbial
manipulation for biocontrol. Encyclopedia of plant and crop science. Marcel Dekker,
New York: 1098-1101.
Luttenberg and F. Kamilova. 2009. Plant-growth-promoting rhizobacteria. Ann. Rev.
Microbiol. 26(1): 5-9.
Maccaferri, M., M.C. Sanguineti, A. Demontis, A. El-Ahmed, L. Garcia Del Moral, F.
Maalouf, M. Nachit, N. Nserallah, H. Ouabbou, S. Rhouma, C. Royo, D. Villegas and
R. Tuberosa. 2011. Association mapping in durum wheat grownacross a broad range
of water regimes. J. Exp. Bot. 62: 409-438.
Malamy, J.E.. 2005. Intrinsic and environmental response pathways that regulate root system
architecture. Plant cell Environ. 28(1): 67-77.
Malamy, J.E. and K.S. Ryan. 2001. Environmental regulation of lateral root initiation in
Arabidopsis. Plant Physiol. 127(3): 899-909.
Malhotra, M. and S. Srivastava. 2006. Targeted engineering of Azospirillum brasilense SM
with indole acetamide pathway for indoleacetic acid over-expression. Can. J. Microb.
52(11): 1078-1084.
Marcar, N.E., P. Dart and C. Sweeney. 1991. Effect of root-zone salinity on growth and
chemical composition of Acacia ampliceps BR Maslin, A. auriculiformis A. Cunn. ex
Benth. and A. mangium Willd at two nitrogen levels. New phytol. 119(4): 567-573.
Marschner, 1995. Mineral Nutrition of Plants, Ed 2. . Academic Press, Boston
Martinez, V. and A. Lauchli. 1994. Salt-induced inhibition of phosphate uptake in plants of
cotton (Gossypium hirsutum L.). New Phyt. 126(4): 609-614.
Mathesius, U., H.R.M. Schlaman, H.P. Spaink, C. Of Sautter, B.G. Rolfe and M.A.
Djordjevic. 1998. Auxin transport inhibition precedes root nodule formation in white
clover roots and is regulated by flavonoids and derivatives of chitin oligosaccharides.
Plant J. 14(1): 23-34.

166
Mayak, S., T. Tirosh and B.R. Glick. 1999. Effect of wild-type and mutant plant growth-
promoting rhizobacteria on the rooting of mung bean cuttings. J. Plant Growth Regul.
18(2): 49-53.
Mayak, S., T. Tirosh and B.R. Glick. 2004. Plant growth-promoting bacteria that confer
resistance to water stress in tomatoes and peppers. Plant Sci. 166(2): 525-530.
McCue, K.F. and A.D. Hanson, 1990. Drought and salt tolerance: towards understanding and
application. Trend Biotech. 8: 358-362.
McNeil, S.D., M.L. Nuccio and A.D. Hanson. 1999. Betaines and related osmoprotectants.
Targets for metabolic engineering of stress resistance. Plant Physiol. 120(4): 945-949.
Mensah Brown, E.P.K., A. Shahin, M. Allen Shamisi, X. Wei and M.L. Lukic. 2006. AL 23
leads to diabetes induction after subdiabetogenic treatment with multiple low doses of
streptozotocin. Eur. J. Immunol. 36(1): 216-223.
Miller, G.A.D., N. Suzuki, S. Ciftci Yilmaz and R.O.N. Mittler. 2010. Reactive oxygen
species homeostasis and signalling during drought and salinity stresses. Plant cell
Environ. 33(4): 453-467.
Mirza, O., L. Guan, G. Verner, S. Iwata and H.R. Kaback. 2006. Structural evidence for
induced fit and a mechanism for sugar/Handplus; symport in LacY. Embo. J. 25(6):
1177-1183.
Misra, A.N., A.K. Biswal and M. Misra. 2002. Physiological, biochemical and molecular
aspects of water stress responses in plants, and the bio-technological applications.
Proc. Nat. Acad. Sci. India B. 72(2): 115-134.
Mittler, R. 2002. Oxidative stress, antioxidants and stress tolerance. Trend. Plant Sci. 7(9):
405-410.
Mittler, R. and E. Blumwald. 2010. Genetic engineering for modern agriculture: challenges
and perspectives. Ann. Rev. Plant Bio. 61: 443-462.
Morales, F.n., A.n. Abada, J.n. Gzâ Aparisi and J. Abada. 1992. Effects of combined NaCl
and CaCl2 salinity on photosynthetic parameters of barley grown in nutrient solution.
Physiol. Plant. 86(3): 419-426.

167
Muhling, K.H. and A. Lauchli. 2002. Effect of salt stress on growth and cation
compartmentation in leaves of two plant species differing in salt tolerance. J. Plant
Physiol., 159(2): 137-146.
Munns, R. 1993. Physiological processes limiting plant growth in saline soils: some dogmas
and hypotheses. Plant Cell Environ. 16(1): 15-24.
Munns, R. 2002. Comparative physiology of salt and water stress. Plant Cell Environ. 25(2):
239-250.
Munns, R. 2005. Genes and salt tolerance: bringing them together. New Phytol. 167(3): 645-
663.
Munns, R., J.B. Passioura, J. Guo, O. Chazen and G.R. Cramer. 2000. Water relations and
leaf expansion: importance of time scale. J. Exp. Bot. 51(350): 1495-1504.
Munns, R. and M. Tester. 2008. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 59:
651-681.
Nadeem, S.M., Z.A. Zahir, M. Naveed, M. Arshad and S.M. Shahzad. 2006. Variation in
growth and ion uptake of maize due to inoculation with plant growth promoting
rhizobacteria under salt stress. Soil Environ. 25:78-84.
Nadeem, S.M., Z.A. Zahir, M. Naveed and M. Arshad. 2007. Preliminary investigations on
inducing salt tolerance in maize through inoculation with rhizobacteria containing
ACC deaminase activity. Can. J. Microbiol. 53(10): 1141-1149.
Nadeem, S.M., Z.A. Zahir, M. Naveed and M. Arshad. 2009. Rhizobacteria containing ACC-
deaminase confer salt tolerance in maize grown on salt-affected fields. Can. J.
Microbiol. 55(11): 1302-1309.
Nadeem, S.M., Z.A. Zahir, M. Naveed, H.N. Asghar and M. Arshad. 2010. Rhizobacteria
capable of producing ACC-deaminase may mitigate salt stress in wheat. Soil Sci. Soc.
Am. J. 74(2): 533-542.
Naqvi, S.M. and R. Ansari. 1974. Estimation of diffusible auxin under saline growth
conditions. Cell. Mol. Life Sci. 30(4): 350-350.

168
Nassar, A.H., K.A. El-Tarabily and K. Sivasithamparam. 2005. Promotion of plant growth by
an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (Zea
mays L.) roots. Bio. Fert. Soils. 42(2): 97-108.
Nautiyal, C.S., S. Bhadauria, P. Kumar, H. Lal and M.D. Verma. 2000. Stress induced
phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol.
Lett. 182: 291-296.
Nemati, I., F. Moradi, S. Gholizadeh, M.A. Esmaeili and M.R. Bihamta. 2011. The effect of
salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots
of rice (Oryza sativa L.) seedlings. Plant Soil Environ. 57(1): 26-33.
Neves-Piestun, B.G. and N. Bernstein. 2005. Salinity-induced changes in the nutritional
status of expanding cells may impact leaf growth inhibition in maize. Funct. Plant
Biol. 32(2): 141-152.
Nia, S.H., M.J. Zarea, F. Rejali and A. Varma. 2012. Yield and yield components of wheat as
affected by salinity and inoculation with Azospirillum strains from saline or non-
saline soil. J. Saud. Soc. Agric. Sci. 11(2): 113-121.
Nilsen, E.T. and D.M. Orcutt. 1996. Physiology of plants under stress. Abiotic factors.
Physiology of plants under stress. Abiotic factors.
Noctor, G. and C.H. Foyer. 1998. Ascorbate and glutathione: keeping active oxygen under
control. Ann. Rev. Plant Biol. 49(1): 249-279.
Noel, T.C., C. Cheng, C.K. Yost, R.P. Pharis, and M.F. Hynes. 1996. Rhizobium
leguminosarum as a plant-growth hizobacterium: direct growth promotion of canola
and lettuce. Can. J. Microbiol. 42: 279-283.
Okon, Y. and J.Vanderleyden. (1997). Root associated Azospirillum sp. can stimulate plants.
American Society for Microbiology News. 63: 366-370.
Omar, M.N.A., M.E.H. Osman, W.A. Kasim and I.A.A. El-Daim. 2009. Improvement of salt
tolerance mechanisms of barley cultivated under salt stress using Azospirillum
brasilense, Salinity and Water Stress. Springer. 133-147.
Owen, A.B. 2001. Empirical likelihood. CRC press.

169
Paknejad, F., M. Nasri, H.R.T. Moghadam, H. Zahedi and M.J. Alahmadi. 2007. Effects of
Drought Stress on Chlorophyll Fluorescence Parameters, Chlorophyll Content and
Grain Yield of Wheat Cultivars. J. Biol. Sci. 7(6)856-862.
Parasher, A. and S.K. Varma. 1988. Effect of pre-sowing seed soaking in gibberellic-acid on
growth of wheat (Triticum-aestivum) under different saline conditions. Ind. J. Exp.
Biol. 26(6): 473-475.
Parida, A.K. and A.B. Das. 2004. Effects of NaCl stress on nitrogen and phosphorous
metabolism in a true mangrove Bruguiera parviflora grown under hydroponic culture.
J. Plant Physiol. 161(8): 921-928.
Parida, A.K. and A.B. Das. 2005. Salt tolerance and salinity effects on plants: a review.
Ecotoxicol. Environ. Saf. 60(3): 324-349.
Parida, B.K., I.M. Chhibba and V.K. Nayyar. 2003. Influence of nickel-contaminated soils on
fenugreek (Trigonella corniculata L.) growth and mineral composition. Scientia
horticulturae, 98(2): 113-119.
Patten, C.L. and B.R. Glick. 1996. Bacterial biosynthesis of indole-3-acetic acid. Can. J.
Microbiol. 42(3): 207-220.
Patten, C.L. and B.R. Glick. 2002. Role of Pseudomonas putida indoleacetic acid in
development of the host plant root system. Appl. Environ. Microbiol. 68(8): 3795-
3801.
Pérez-López, U., A. Robredo, M. Lacuesta, C. Sgherri, A. Mena Petite, F. Navari Izzo and A.
Munoz Rueda. 2010. Lipoic acid and redox status in barley plants subjected to salinity
and elevated CO2. Physiol. Plantarum. 139(3): 256-268.
Pérez-López, U., A. Robredo, M. Lacuesta, C. Sgherri, A. Munoz Rueda, F. Navari Izzo and
A. Mena Petite. 2009. The oxidative stress caused by salinity in two barley cultivars is
mitigated by elevated CO2. Physiol plantarum. 135(1): 29-42.
Perez-Torres, C.-A., J. Lopez-Bucio, A. Cruz-Ramirez, E. Ibarra-Laclette, S. Dharmasiri, M.
Estelle and L. Herrera-Estrella. 2008. Phosphate availability alters lateral root

170
development in Arabidopsis by modulating auxin sensitivity via a mechanism
involving the TIR1 auxin receptor. The Plant Cell Online, 20(12): 3258-3272.
Perrine, F.M., B.G. Rolfe, M.F. Hynes and C.H. Hocart. 2004. Gas chromatography-mass
spectrometry analysis of indoleacetic acid and tryptophan following aqueous
chloroformate derivatisation of Rhizobium exudates. Plant Physiol. Biochem. 42(9):
723-729.
Pilet, P.E. and M. Saugy. 1987. Effect on Root Growth of Endogenous and Applied IAA and
ABA A Critical Reexamination. Plant Physiol. 83(1): 33-38.
Pitzschke, A. and H. Hirt. 2006. Mitogen-activated protein kinases and reactive oxygen
species signaling in plants. Plant Physiol. 141(2): 351-356.
Potters, G., T.P. Pasternak, Y. Guisez and M.A.K. Jansen. 2009. Different stresses, similar
morphogenic responses: integrating a plethora of pathways. Plant Cell Environ. 32(2):
158-169.
Potters, G., T.P. Pasternak, Y. Guisez, K.J. Palme and M.A.K. Jansen. 2007. Stress-induced
morphogenic responses: growing out of trouble? Tren. Plant Sci. 12(3): 98-105.
Prakash. and Parthapasenan. 1990. Interactive effect of NaCl salinity and gibberellic acid on
shoot growth, content of abscisic acid and gibberellin like substances and yield of rice
(Oryza sativa). . Plant Sci. 6(2): 447-453.
Prat, D. and R.A. Fathiae Ettai. 1990. Variation in organic and mineral components in young
Eucalyptus seedlings under saline stress. Physiol. Plantarum. 79(3): 479-486.
Principe, A., F. Alvarez, M.G. Castro, L. Zachi, S.E. Fischer, G.B. Mori, and E. Jofre. 2007.
Biocontrol and PGPR features in native strains isolated from saline soils of Argentina.
Curr. Microbiol. 55:314-322.
Prinsen, E., A. Costacurta, K. Michiels, J. Vanderleyden and H. Van Onckelen. 1993.
Azospirillum brasilense indole-3-acetic acid biosynthesis: evidence for a non-
tryptophan dependent pathway. Mol. Plant Microbe Interact. 6: 609-609.
Rabie, G.H. and A.M. Almadini. 2005. Role of bioinoculants in development of salt-tolerance
of Vicia faba plants under salinity stress.

171
Rahman, S.U. 1998. Theory of constraints: a review of the philosophy and its applications.
Int. J. Oper. Product. Manag. 18(4): 336-355.
Rai, S., R. Luthra and S. Kumar. 2003. Salt-tolerant mutants in glycophytic salinity response
(GSR) genes in Catharanthus roseus. Theor. Appl. Genet. 106(2): 221-230.
Reed, M.L. and B.R. Glick. 2005. Growth of canola (Brassica napus) in the presence of plant
growth-promoting bacteria and either copper or polycyclic aromatic hydrocarbons.
Can. J. Microbiol. 51(12): 1061-1069.
Reed, M.L.E. and B.R. Glick. 2004. Applications of free living plant growth-promoting
rhizobacteria. Antonie van Leeuwenhoek.
Remans, T., K. Smeets, K. Opdenakker, D. Mathijsen, J. Vangronsveld and A. Cuypers.
2008. Normalisation of real-time RT-PCR gene expression measurements in
Arabidopsis thaliana exposed to increased metal concentrations. Planta. 227(6): 1343-
1349.
Ribaut, J.M. and P.E. Pilet. 1994. Water stress and indol-3yl-acetic acid content of maize
roots. Planta. 193(4): 502-507.
Rodriguez, A., S. Griffiths-Jones, J.L. Ashurst and A. Bradley. 2004. Identification of
mammalian microRNA host genes and transcription units. Genome Res. 14(10a):
1902-1910.
Rodriguez, H. and R. Fraga. 1999. Phosphate solubilizing bacteria and their role in plant
growth promotion. Biotechnol. Adv. 17(4): 319-339.
Rogers, M.E., C.M. Grieve and M.C. Shannon. 2003. Plant growth and ion relations in
lucerne (Medicago sativa L.) in response to the combined effects of NaCl and P. Plant
Soil. 253(1): 187-194.
Sabri, A.N. and S. Hasnain. 1996. Spore germination analysis of Bacillus subtilis divIVA
mutant strain in ALA and AGFK systems. Pak. J. Zool. 28(3): 213-224.
Sadasivam, S. and A. Manickam. 1992. Biochemical methods for agricultural sciences.
Willey Eastern Limited, New Delhi.

172
Saimbhi, M.S. 1993. Growth regulators on vegetable crops. Advances in horticulture.
Malhotra Publishing House, New Delhi, India: 619-642.
Sairam, R.K. and A. Tyagi. 2004. Physiology and molecular biology of salinity stress
tolerance in plants. Curr. Sci. Bang. 86(3): 407-421.
Sakhabutdinova, A.R., D.R. Fatkhutdinova, M.V. Bezrukova and F.M. Shakirova. 2003.
Salicylic acid prevents the damaging action of stress factors on wheat plants. Bulg. J.
Plant Physiol. 29: 314-319.
Sandhya, V., S.Z. Ali, M. Grover, G. Reddy and B. Venkateswarlu. 2010. Effect of plant
growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and
plant growth of maize under drought stress. Plant Growth Reg. 62(1): 21-30.
Saravanakumar, D. and R. Samiyappan. 2007. ACC deaminase from Pseudomonas
fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl.
Microbiol. 102 (5); 1283-1292.
Sarwar, M., M. Arshad, D.A. Martens and W.T. Frankenberger Jr. 1992. Tryptophan-
dependent biosynthesis of auxins in soil. Plant Soil. 147(2): 207-215.
Sarwar, M. and R.J. Kremer. 1995. Determination of bacterially derived auxins using a
microplate method. Lett. Appl. Microbiol. 20(5): 282-285.
Sastry, E.V. and K.S. Shekhawat. 2001. Alleviatory effect of GA3 on the effects of salinity at
seedling stage in wheat (Triticum aestivum). Ind. J. Agricul. Res. 35(4): 226-231.
Sgherri, C., M.F. Quartacci and F. Navari-Izzo. 2007. Early production of activated oxygen
species in root apoplast of wheat following copper excess. J. Plant Physiol. 164(9):
1152-1160.
Sgroy, V.n., F. Cassán, O. Masciarelli, M.a.F. Del Papa, A. Lagares and V. Luna. 2009.
Isolation and characterization of endophytic plant growth-promoting (PGPB) or stress
homeostasis-regulating (PSHB) bacteria associated to the halophyte Prosopis
strombulifera. Appl. Microbiol. Biotechnol. 85(2): 371-381.

173
Shahbazi, E., S. Kiani, H. Gourabi and H. Baharvand. 2011. Electrospun nanofibrillar
surfaces promote neuronal differentiation and function from human embryonic stem
cells. Tissue Eng. 17(23-24): 3021-3031.
Shalata, A., V. Mittova, M. Volokita, M. Guy and M. Tal. 2001. Response of the cultivated
tomato and its wild salt tolerant relative Lycopersicon pennellii to salt-dependent
oxidative stress: The root antioxidative system. Physiologia Plantarum, 112(4): 487-
494.
Shetty, K., O.F. Curtis, R.E. Levin, R. Witkowsky and W. Ang. 1995. Prevention of
Vitrification associated with in vitro Shoot Culture of Oregano. (Origanum vulgare)
by Pseudomonas spp. J. Plant Physiol. 147(3): 447-451.
Shoebitz, M., C.M. Ribaudo, M.N.A. Pardo, M.a.L. Cantore, L. Ciampi and J.A. Curai. 2009.
Plant growth promoting properties of a strain of Enterobacter ludwigii isolated from
Lolium perenne rhizosphere. Soil Bio. Biochem. 41(9): 1768-1774.
Siddikee, M.A., P.S. Chauhan, R. Anandham, G.H. Han and T. Sa. 2010. Isolation,
characterization, and use for plant growth promotion under salt stress, of ACC
deaminase-producing halotolerant bacteria derived from coastal soil. J Microbiol.
Biotechnol. 20(11): 1577-1584.
Sindhu, S.S. and K.R. Dadarwal. 2001. Chitinolytic and cellulolytic Pseudomonas sp.
antagonistic to fungal pathogens enhances nodulation by Mesorhizobium sp. Cicer in
chickpea. Microbiol. Res. 156(4): 353-358.
Singh, L.P., S.S. Gill and N. Tuteja. 2011. Unraveling the role of fungal symbionts in plant
abiotic stress tolerance. Plant Signal Behav. 6(2): 175-191.
Singleton, P.W. and B.B. Bohlool. 1984. Effect of salinity on nodule formation by soybean.
Plant Physiol. 74(1): 72-76.
Somers, E., J. Vanderleyden, M. Srinivasan. 2004. Rhizosphere bacterial signaling:a love
parade beneath our feet. Crit. Rev. Microbiol. 30: 205-240.
Spaepen, S., S. Dobbelaere, A. Croonenborghs and J. Vanderleyden. 2008. Effects of
Azospirillum brasilense indole-3-acetic acid production on inoculated wheat plants.
Plant Soil. 312(1-2): 15-23.

174
Spaepen, S. and J. Vanderleyden. 2011. Auxin and plant-microbe interactions. Cold Spring
Harbor perspectives in biology, 3(4): 962-970.
Spaepen, S., J. Vanderleyden and Y. Okon. 2009. Plant growth-promoting actions of
rhizobacteria. Adv. Bot. Res. 51: 283-320.
Spaepen, S., J. Vanderleyden and R. Remans. 2007. Indole 3 acetic acid in microbial and
microorganism plant signaling. FEMS Microbiol. Rev. 31(4): 425-448.
Sridevi, M. and K.V. Mallaiah. 2007. Bioproduction of indole acetic acid by Rhizobium
strains isolated from root nodules of green manure crop, Sesbania sesban (L) Merr.
Iran. J. Biotechnol. 5(3): 178-182.
Sridevi, M. and K.V. Mallaiah. 2008. Production of Hydroxamate-Type of Siderophores by
Rhizobium strains from Sesbania sesban (L.) Merr. Int. J. Soil Sc. 3(1): 752-759.
Steenhoudt, O. and J. Vanderleyden. 2000. Azospirillum, a free-living nitrogen-fixing
bacterium closely associated with grasses: genetic, biochemical and ecological
aspects. FEMS Microbiol. Rev. 24(4): 487-506.
Swain, M.R., S.K. Naskar and R.C. Ray. 2007. Indole-3-acetic acid production and effect on
sprouting of yam (Dioscorea rotundata L.) minisetts by Bacillus subtilis isolated from
culturable cowdung microflora. Pol. J. Microbiol. 56(2): 103.
Synkova, H., A. A.R. Semoraidova¡iand L. Burketova. 2004. High content of endogenous
cytokinins stimulates activity of enzymes and proteins involved in stress response in
Nicotiana tabacum. Plant cell, tissue Org. Cult. 79(2): 169-179.
Szabados, L. and A. Savoure. 2009. Proline: a multifunctional amino acid. Trends in plant
science, 15(2): 89-97.
Taiz, L. and E. Zeiger, 1991. Plant Physiology. 1991. Benjamin Cummings Publishing, CA.
Tate, R.L., 1995. Soil microbiology. Wiley New York.
Teale, W.D., I.A. Paponov and K. Palme. 2006. Auxin in action: signalling, transport and the
control of plant growth and development. Nature Reviews Molecular Cell Biology,
7(11): 847-859.

175
Tejera, N.A., M. Soussi and C. Lluch. 2006. Physiological and nutritional indicators of
tolerance to salinity in chickpea plants growing under symbiotic conditions. Environ.
Exp. Bot. 58(1): 17-24.
Tester, M. and R. Davenport. 2003. Na+ tolerance and Na+ transport in higher plants. Ann.
Bot. 91(5): 503-527.
Thomashow, M.F. 1999. Plant cold acclimation: freezing tolerance genes and regulatory
mechanisms. Ann. Rev. Plant Bio. 50(1): 571-599.
Tilak, K., N. Ranganayaki and C. Manoharachari. 2006. Synergistic effects of plant-growth
promoting rhizobacteria and Rhizobium on nodulation and nitrogen fixation by
pigeonpea (Cajanus cajan). Eur. J. Soil Sci. 57(1): 67-71.
Tognetti, V.B., P.E.R. Muhlenbock and F. Van Breusegem. 2012. Stress homeostasis: the
redox and auxin perspective. Plant cell Environ. 35(2): 321-333.
Torres, M.A. and J.L. Dangl. 2005. Functions of the respiratory burst oxidase in biotic
interactions, abiotic stress and development. Curr. Opin. Plant Biol. 8(4): 397-403.
Torsethaugen, G., L.H. Pitcher, B.A. Zilinskas and E.J. Pell. 1997. Overproduction of
ascorbate peroxidase in the tobacco chloroplast does not provide protection against
ozone. Plant Physiol. 114(2): 529-537.
Tsavkelova, E.A., T.A. Cherdyntseva, S.Y. Klimova, A.I. Shestakov, S.G. Botina and A.I.
Netrusov. 2007. Orchid-associated bacteria produce indole-3-acetic acid, promote
seed germination, and increase their microbial yield in response to exogenous auxin.
Archives Microbiol. 188(6): 655-664.
Tsavkelova, E.A., S.Y. Klimova, T.A. Cherdyntseva and A.I. Netrusov. 2006. Microbial producers of plant growth stimulators and their practical use: a review. Appl. Biochem. Microbiol. 42(2): 117-126.
Valliyodan, B. and H.T. Nguyen. 2006. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Bio. 9(2): 189-195.
Vardharajula, S., S. Zulfikar Ali, M. Grover, G. Reddy and V. Bandi. 2011. Drought-tolerant
plant growth promoting Bacillus spp.: effect on growth, osmolytes, and antioxidant
status of maize under drought stress. J. Plant Inter. 6(1): 1-14.

176
Velagaleti, R.R. and S. Marsh. 1989. Influence of host cultivars and Bradyrhizobium strains
on the growth and symbiotic performance of soybean under salt stress. Plant Soil.
119(1): 133-138.
Verbruggen, N. and C. Hermans. 2008. Proline accumulation in plants: a review. Amino
acids, 35(4): 753-759.
Vinocur, B. and A. Altman. 2005. Recent advances in engineering plant tolerance to abiotic
stress: achievements and limitations. Curr. Opinion Biotechnol. 16(2): 123-132.
Vivas, A., A. Marulanda, J.M. Ruiz-Lozano, J.M. Barea and R. Azcon. 2003. Influence of a
Bacillus sp. on physiological activities of two arbuscular mycorrhizal fungi and on
plant response to PEGinduced drought stress. Mycorriza. 13:249-256.
Volpin, H., S. Burdman, S. Castro-Sowinski, Y. Kapulnik and Y. Okon. 1996. Inoculation
with Azospirillum increased exudation of rhizobial nocÃ--Gene inducers by alfalfa
roots.
Wang, T.L., E.A. Wood and N.J. Brewin. 1982. Growth regulators, Rhizobium and
nodulation in peas. Planta. 155(4): 345-349.
Wang, Y.-X.J., S.M. Hussain and G.P. Krestin. 2001. Superparamagnetic iron oxide contrast
agents: physicochemical characteristics and applications in MR imaging. Eur. Radiol.
11(11): 2319-2331.
Williams, M.N.V. and E.R. Signer. 1990. Metabolism of tryptophan and tryptophan analogs
by Rhizobium meliloti. Plant Physiol. 92(4): 1009-1013.
Winicov, I. 1998. New molecular approaches to improving salt tolerance in crop plants. Ann.
Bot. 82(6): 703-710.
Wolters, H. and G. Jurgens. 2009. Survival of the flexible: hormonal growth control and
adaptation in plant development. Nat. Rev. Genet. 10(5): 305-317.
Wyn Jones, R.G. and R. Storey. 1981. Betaines. The physiology and biochemistry of drought
resistance in plants. Academic Press, Sydney: 171-204.
Xue, J., S. Uchida, B.P. Rand and S.R. Forrest. 2004. Efficient organic photovoltaic cells
with low series resistances. Appl. Phy. Lett. 84(16): 3013-3015.

177
Yamaguchi, T. and E. Blumwald. 2005. Developing salt-tolerant crop plants: challenges and
opportunities. Trends Plant Sci. 10(12): 615-620.
Yang, J., J.W. Kloepper and C.M. Ryu. 2008. Rhizosphere bacteria help plants tolerateabiotic
stress. Trends Plant Sciences. 14: 1-4.
Yao, T. 2004. Associative nitrogen-fi xing bacteria in the rhizosphere of Avena sativa in an
alpine region: III. Phosphate-solubilizing power and auxin production. Acta
Prataculturae Sinica. 13:85-90.
Younis, M.E., O.A. El-Shahaby, M.M. Nemat Alla and Z.M. El-Bastawisy. 2003. Kinetin
alleviates the influence of waterlogging and salinity on growth and affects the
production of plant growth regulators in Vigna sinensis and Zea mays. Agronomie-
Sciences des Productions Vegetales et de l'Environnement. 23(4): 277-286.
Yue, H., W. Mo, C. Li, Y, Zheng, H. Li. 2007. The salt stress relief and growth promotion
eVect of RS-5 on cotton. Plant Soil. 297:139145.
Zahir, A.A., A.A. Rahuman, A. Bagavan, T. Santhoshkumar, R.R. Mohamed, C. Kamaraj, G.
Rajakumar, G. Elango, C. Jayaseelan and S. Marimuthu. 2010. Evaluation of
botanical extracts against Haemaphysalis bispinosa Neumann and Hippobosca
maculata Leach. Parasitol. Res. 107(3): 585-592.
Zahir, F., S.J. Rizwi, S.K. Haq and R.H. Khan. 2005. Low dose mercury toxicity and human
health. Environ. Toxicol. Pharmacol. 20(2): 351-360.
zahir, Z.A., H.M. Yasin, M. Naveed, M.A. Anjum and M. Khalid. 2010. L-tryptophan
application enhances the effectiveness of rhizobium inoculation for improving growth
and yield of mungbean (Vigna radiata L.). Pak. J. Bot. 42(3): 1771-1780.
Zahir, Z.A., M. Zafar-ul-Hye, S. Sajjad and M. Naveed. 2011. Comparative effectiveness of
Pseudomonas and Serratia sp. containing ACC-deaminase for coinoculation with
Rhizobium leguminosarum to improve growth, nodulation, and yield of lentil. Biol.
Fert. soils. 47(4): 457-465.
Zahran, H.H. 1999. Rhizobium-legume symbiosis and nitrogen fixation under severe
conditions and in an arid climate. Microbiol. Mol. Boil. Rev. 63(4): 968-989.

178
Zahran, H.H. and J.I. Sprent. 1986. Effects of sodium chloride and polyethylene glycol on
root-hair infection and nodulation of Vicia faba L. plants by Rhizobium
leguminosarum. Planta. 167(3): 303-309.
Zhao, B., J.S. Moore and D.J. Beebe. 2001. Surface-directed liquid flow inside
microchannels. Science. 291(5506): 1023-1026.
Zholkevich, V.N. and T.N. Pustovoytova. 1993. The role of Cucumis sativum L. leaves and
content of phytohormones under soil drought. Russ. J. Plant Physiol. 40: 676-680.