10353-11 ch11 - lippincott williams &...
TRANSCRIPT

238
11
Motor activity is a hierarchically organized functionunder the control of reflex mechanisms and neural
networks in a rostral segment of the central nervous sys-tem (CNS) (Fig. 11-1). From the lowest to the highest,these arbitrarily identified anatomic levels are the spinalcord, cerebellum, brainstem, basal ganglia, and motorcortex. Each ascending level in the hierarchy makes a spe-cific contribution to the final motor activity, which also isinfluenced in part by the activity of higher motor centers.For example, the midbrain systems modulate reflexesorganized at the medullary or spinal level. The forebrainsystems regulate the midbrain and spinal motor activity.Motor responses begin in the spinal cord as simplereflexes, whereas the higher motor centers participate inthe regulation of skilled and patterned movements. Neu-ronal impulses from higher levels also initiate, inhibit,or facilitate motor functions at the brainstem and spinalcord, thus partially regulating all motor behavior.
This cortical control provides a type of parallel process-ing with rostral domination of hierarchical motor organiza-tion and simultaneous control of the segmental output itself.Because some time is involved in the conduction of afferentinformation, local (spinal) reflexes often are activated first,and motor mechanisms at higher hierarchic levels are acti-vated slightly later. The best example of reflexive move-ment is mistakenly stepping on a tack with your bare foot.This triggers a leg-withdrawal reflex, which is already inprogress before the forebrain can inhibit or modify theaction. However, the higher motor systems can inhibitsuch a withdrawal reflex if the stimulus is expected andanother response has been learned. For example, if some-one has to pick up a hot cup, the withdrawal reflex can beinhibited while the hot cup is being moved to a support-ing surface, even though the fingers are being burned.
The hierarchically defined motor functions and thenature of the specific contributions by each level are dis-cussed in Chapters 11–14. The spinal cord is the first levelin the regulation of sensorimotor functions. Stereotypicalmotor responses (reflexes), which are largely independentof voluntary motor control, are generated in the spinalcord and can be triggered by cortical and/or environmen-tal stimuli. The spinal cord also relays sensory informationto the cerebrum, which is vital to learning skilled andcoordinated motor activity.
SPINAL PREPARATION
Spinal cord functions are most easily demonstrated by aspinal preparation, which involves separating the cord fromthe regulating influence of the brain by cutting the cord.This separation allows for examination of spinal reflexesindependent of the inhibitory or facilitatory influenceexerted by higher motor centers. Immediately after thespinal cut is made, the flexor and extensor reflexes becomecompletely inactive, but they gradually re-emerge.
INNERVATION PATTERN
The general motor function in the spinal cord and brain-stem is organized ipsilateral to its output and reflex input.With the crossing of corticospinal fibers at the medulla,
Motor System 1: Spinal Cord
LEARNING OBJECTIVES
After studying this chapter, students should be able to:
• Discuss the anatomy of the spinal cord
• Describe the functions of the spinal structures
• Discuss the importance of the motor unit in movement
• List the major ascending and descending spinal tracts
• Describe the functions of the major sensorimotor tracts
• Explain the role of muscle spindles in reflexive motorfunctions
• Discuss the importance of the Golgi tendon organ
• Describe the physiology of basic spinal reflexes
• Explain the importance of the lower motor neurons
• Discuss lower motor neuron syndrome
• Explain the common pathologies affecting spinal cordfunctions
• Describe the sensorimotor symptoms associated withspinal cord injuries
10353-11_CH11.qxd 8/30/07 1:16 PM Page 238

CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 239
the spinal �-motor neurons and their axons (final com-mon pathway) extend to innervate the muscles ipsilaterally(Figs. 11-2 and 11-4). Consequently, lower motor neuron(LMN) signs in the brainstem and spinal cord are ipsilateralto the damage of the cell body or its axon.
GROSS ANATOMY OF THE SPINAL CORD
The spinal cord extends from the base of the skull to thelower back and is approximately 43.5 cm long and 1 cmin diameter. It is divided into five regions: cervical, tho-racic, lumbar, sacral, and coccygeal. A total of 31 pairs ofspinal nerves emerge from the spinal cord (see Fig. 2-33).These nerves have mixed functions; they carry sensoryinformation from peripheral receptors to the CNS and
transmit motor information from the CNS to the muscles.The nerves are named after the region of the spinal cordto which they are attached and are numbered in sequence.There are 8 pairs of cervical spinal nerves (C1–C8), 12 pairsof thoracic spinal nerves (T1–T12), 5 pairs of lumbar spinalnerves (L1–L5), 5 pairs of sacral spinal nerves (S1–S5), andusually 1 pair of coccygeal spinal nerves (see Chapter 2).
In adults, the spinal cord diminishes toward the lum-bar region and ends as the conus medullaris at the L2 ver-tebra. The nerve root fibers from the L3 to S5 spinalsegments stretch downward to reach the correspondingskeletal level before exiting the vertebral-spinal canal. Thepia mater covering of the spinal cord extends beyond theconus medullaris in a thin filum terminale, which extendsto and attaches to the coccyx. The spinal dura mater andsubarachnoid space, filled with cerebrospinal fluid (CSF),extend beyond the conus medullaris to terminate in thesacrum by attaching to the coccyx (see Fig. 2-33).
The fact that the spinal cord does not extend into thefluid-filled subarachnoid space of the lumbosacral area hasimportant clinical implications. This dural sac, at levelsbelow the L1 vertebra (much lower in children becausethey have less disparity between the length of the cordand vertebral canal), is used for extracting CSF for diagnos-tic purposes (spinal tap or puncture) and for administeringtherapeutic and anesthetic drugs without risking damageto the spinal cord (see Chapter 20).
Internal AnatomySeen in cross section, the spinal cord consists of an outerring of white matter and a butterfly-shaped central grayarea (Fig. 11-2). The white matter contains ascendingand descending fibers, whereas the gray matter containsnerve cell bodies and a small unmyelinated network offibers (neuropils). The dorsal horns contain the secondarysensory nerve cells that receive sensory information fromthe body through the dorsal root ganglia (DRG) fibers.The ventral horns contain motor nerve cells, which pro-ject through the anterior roots to activate muscles, glands,and joints. The gray columns on each side of the cord areconnected through the commissures, which consist ofcrossing fibers. The axonal processes of the motor neu-rons form the ventral nerve root fibers and send motorimpulses from the CNS to the muscles. After travelingthrough the pia mater, subarachnoid space, arachnoidmembrane, dura mater, and intervertebral foramina, thefibers from both the dorsal and anterior roots merge toform a spinal nerve (Fig. 11-2; see Fig. 2-32).
After a spinal nerve exits the intervertebral foramina,it divides into dorsal and ventral rami. The dorsal ramusfibers of each spinal nerve are concerned with the musclesand skin in the posterior part of the body. The ventralramus fibers of the spinal nerve supply anterior body parts,including the upper and lower limbs. Before projectingto target muscles and body parts (see Fig. 2-35), the ven-tral rami of the spinal nerves, except the nerves from T2 to
3
1
4
6
5
2
Figure 11-1 Six ascending levels of motor organization.1, spinal cord; 2, brainstem; 3, cerebellum; 4, diencephalon;5, basal ganglia; 6, cerebral cortex.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 239

240 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
T11, form a network of nerve fibers called plexuses. Ramiof the nerves from T2 to T11 directly innervate body parts.
The size and shape of the spinal cord are not uniformalong its length (Fig. 11-3). The cord in the cervical andlumbar areas is broad and flattened in contrast to its shapein the thoracic areas. Widening at the cervical and lumbarsegments of the cord results from the concentration ofnerve cells and their fibers, which is necessitated by thesensory and motor control of the upper and lower extrem-ity muscles. The upper extremities, especially the hands,possess more specialized sensory and motor functions thanthe legs, trunk, and viscera and therefore require a greaterconcentration of peripheral nerve fibers. Thus the size ofthe gray column at the cervical level is larger than at thelumbar level.
Segmental OrganizationThe spinal cord is anatomically and functionally organ-ized in transverse segments extending from the cervicalto the sacral region. This organization of the spinal cordindicates segmental sensorimotor innervation of the body(dermatomes and myotomes) with various distributionalshapes and sizes (see Fig. 2-34). The dermatome (areainnervated by the afferent fibers of the neurons in a sin-gle DRG) and myotome (muscle or muscles innervated bythe axons exiting the cord via a single ventral root) mayoverlap each other, but because of muscle migration dur-ing development, they are not always the same.
Motor UnitThe LMN and motor unit are two important componentsof spinal motor control circuitry (Figs. 11-2 and 11-4).The LMN cell body provides the output pathway via itsaxon, which travels through the ventral root and periph-
eral nerves to innervate a skeletal muscle, where it formsmultiple axon branches to activate many muscle fibers.The LMN is also the final common pathway because theefferent impulses from the motor cortex pass throughthis motor neuron before they can produce a musclemovement.
The motor unit consists of four components: motorcell body, efferent fibers, motor end plate (branching offthe axonal fiber in myoneural/neuromuscular junctions),and innervated muscle fibers. A motor unit can fire repeat-edly, resulting in sustained shortening of the muscle fiberelements. Damage to the LMN cell or the beginning axoneliminates the entire function of a motor unit. Damage toone of the terminal axon branches weakens the unit pro-jections. Generalized skeletal muscle disease (e.g., muscu-lar dystrophy) or reduced nerve–muscle transmission/activity (e.g., myasthenia gravis) weakens or eliminatesthe unit’s force generation and causes muscle degeneration.
Motor units can be small or large, and a muscle cancontain many motor units, depending on the nature of itsmotor control. For example, the small hand flexor muscleused for delicate and coordinated motor control can have10–30 muscle fibers per motor unit, whereas large muscleslike the quadriceps can have as many as 3,000 muscle fibersper motor unit.
TRACTS OF THE SPINAL CORD
The white matter of the spinal cord consists of three majorbundles of longitudinal axons (fasciculi): dorsal, lateral,and ventral columns (Fig. 11-2). In most instances, eachfasciculus contains bundles of ascending and descendingfibers. The tracts are difficult to pinpoint on a cross section
.Sensory
nerve
Dorsalhorn
Dorsal
VentralVentral
fasciculus
Dorsal fasciculus
Lateral fasciculus
Motor nerveto muscles
Gray matter
Centralcanal
Ventralhorn cell
Dorsal rootganglion
Spinalnerve
White matter
Motor neurons
Sensory cells
Figure 11-2 Internal anatomy of the spinal cord. The central gray matter is surrounded bywhite matter, which consists of three fasciculi (funiculi): dorsal, lateral, and ventral.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 240

of the spinal cord; however, their approximate locations canbe determined from clinical evidence. The dorsal fascicu-lus consists largely of ascending (sensory) fibers, whereasthe lateral and anterior fasciculi contain both descending(motor) and ascending (sensory) fiber bundles (Table 11-1).General locations of the various sensorimotor spinal tractsare demonstrated in Figure 11-5.
Descending TractsCorticospinal Tracts
Fibers of the corticospinal motor tract arise from pyrami-dal cells (the largest are Betz cells) in the cortex. Most ofthese cells are in the precentral gyrus (primary motor cor-
tex, Brodmann area 4); however, some of these cells are inother areas of the brain, including the premotor cortex,primary somesthetic cortex (Brodmann areas 3, 1, 2), andsupplementary motor cortex (Brodmann area 6). On theirway to the spinal cord, the corticospinal fibers cross themidline at the caudal end of the medulla and form the lat-eral corticospinal tract in the spinal cord. There are twocorticospinal tracts; the other one is the anterior corti-cospinal tract (Fig. 11-6; see Chapter 14).
Lateral Corticospinal Tract
Lateral corticospinal fibers provide a mechanism by whichthe cerebral cortex intervenes in the control of skeletal
Filumterminale
Caudaequina
Conusmedullaris
Lumbarenlargement
Dorsolateralsulcus
Cervicalenlargement
Dorsal mediansulcus
Coccygealnerve
Dorsal intermediatesulcus
Gray
White
Dorsalfasciculus
Dorsal (sensory) horn Lateral
fasciculus
Ventral (motor) horn
Ventralfasciculus
C1 Segment
C8 Segment
T2 Segment
L4 Segment
S3 Segment
Figure 11-3 The dorsal spinal cord and its internal anatomy in five crosssections. The increase in cord size at the cervical and lumbar levels corre-sponds to the nerve supply for the upper and lower extremities, respectively.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 241
10353-11_CH11.qxd 8/30/07 1:16 PM Page 241

242 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
muscles during delicate, skilled manipulation of the distalparts of the limbs, including the fingers and to some extentthe toes and forearms. This tract contains myelinated fibersand is the largest of the motor tracts. After emerging fromthe pyramidal cells (including the giant Betz cells) in the
motor cortex, the corticospinal fibers cross the midlineat the lower end of the medulla and enter with the spinalcord. Approximately 90% of the corticospinal fibers areknown to cross (decussate) the midline to form the lateralcorticospinal tract. Its fibers synapse on the ventral horn
Muscle
α-Motor neuron(final common path) Efferent fibers
Neuromuscularjunction
Spinal Pathways Function
Ascending tracts
Fasciculus gracilis Transmits discriminative touch from lower half of bodyFasciculus cuneatus Transmits discriminative touch from upper half of bodyDorsal spinocerebellar tract Mediates unconscious proprioception from distal lower limbsVentral spinocerebellar tract Mediates unconscious proprioception from muscles of lower extremities and proximal limbsCuneocerebellar tract Mediates unconscious proprioceptive information from upper limbsLateral spinothalamic tract Mediates sensations of pain and temperatureAnterior spinothalamic tract Mediates diffuse touch; backup sensory systemSpino-olivary tract Mediates proprioceptive information from limbsSpinoreticular tract Mediates sensory input to reflex networks of brainstemSpinotectal tract Mediates sensory input to eye and head orientation reflex networks of superior colliculus
Descending tracts
Lateral corticospinal tract Carries motor commands for muscle control during digital tasksAnterior corticospinal tract Carries uncrossed motor fibers responsible for motor precision of axial and girdle musclesTectospinal tract Carries visual and auditory impulses to regulate reflexive postural body movementsRubrospinal tract Carries cerebellar projections to spinal cord for regulating muscle tone to support body
against gravityVestibulospinal tract Relays vestibular input to muscles for regulating reflexive adjustments of body postureReticular descending tracts Relays brainstem reflex modulation to spinal reflex mechanism; regulates muscle preparedness
Table 11-1
Ascending and Descending Spinal Pathways
Figure 11-4 The components that form the final common pathway andmotor unit. The motor unit is composed of a motor neuron, efferent fibers,the neuromuscular junction, and muscle.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 242

α-motor neurons and regulate muscle activity. A lesionto the fibers of this tract causes profound weakness andloss of all individual digital manipulation skills. This con-dition called paralysis, or upper motor neuron (UMN)paralysis, and can be monoplegia (paralysis of one extrem-ity) or hemiplegia (paralysis of the arm and leg on one sideof the body).
Anterior Corticospinal Tract
The anterior corticospinal tract contains the 8–10% of thecorticospinal fibers that do not cross the midline at the medulla oblongata. Instead, these fibers continuedescending ipsilaterally. These uncrossed fibers eventu-ally cross the midline before synapsing on the ventralhorn �-motor neuron and internuncial cells. These axonsprovide a mechanism through which the cerebral cortexregulates precision in the movements of axial and girdlemuscles.
Extrapyramidal Tracts
Several extrapyramidal motor, reticular, and autonomictracts transmit information essential for smoothly coordi-nated motor function and upright, balanced posture. This
indicates that the motor system is not exclusively a corti-cospinal system. Rather, its functional organization is highlycomplex, involving many additional pathways. Some ofthe more notable extrapyramidal paths are the tectospinal,rubrospinal, and vestibulospinal tracts.
The tectospinal tract regulates neck and body twistingmovements with extensor support for the visual and audi-tory startle (astonish) reflexes. For example, in responseto a sudden, bright, moving light entering the visual fieldfrom one side or a loud sound to one side, a powerfulreflexive behavior involves the sudden turning of the head,neck, and/or body toward the stimulus. Originating fromthe superior colliculus, the tectospinal fibers cross themidline in the midbrain and descend to terminate in theventral horn motor nuclei of the cervical and lower regionsof the cord.
The rubrospinal tract originates from cell bodies ofthe red nucleus, which receive their afferents from thecontralateral cerebellum and ipsilateral primary motor(Brodmann area 4) and supplementary motor (Brodmannarea 6) cortices. This tract transmits impulses from thered nucleus to the spinal LMN to regulate muscle tone forlimb extension in support of the body against gravity.
Fasciculusgracilis Fasciculus
cuneatusFasciculus
dorsolateralis
Dorsalspinocerebellar
tract
Lateralspinothalamic
tract
Ventralspinocerebellar
tract
Spino-olivarytract
Spinotectal tract
Anteriorspinothalamic
tract
Anteriorcorticospinal
tract
TectospinaltractPontine
reticulospinaltract
Rubrospinaltract
Medullaryreticulospinal
tract
Vestibulospinaltract
Fasciculusproprius
Lateralcorticospinal
tract
➔➔
➔
➔
➔
➔
➔ ➔
Medial longitudinalfasciculus
(cervical extension)
Descending tracts Ascending tracts
Figure 11-5 Ascending (red) and descending (gray) spinal pathways.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 243
10353-11_CH11.qxd 8/30/07 1:16 PM Page 243

244 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
The vestibulospinal tract projects the vestibular im-pulses to the spinal LMNs. By regulating extensor mus-cle tone, these fibers control the reflexive adjustment ofthe body and limbs to keep the head stable. Originatingfrom the nerve cells in the vestibular nucleus and incor-porating projections from the inner ear and cerebellum,the vestibulospinal fibers extend throughout the lengthof the cord, giving off collaterals to α- and γ-motornuclei.
Originating from the pons (pontine reticulospinaltract) and medulla (medullary reticulospinal tract), thereticular projections regulate coordinated motor functions.Reticular stimulation has been found to facilitate or inhibitvoluntary and reflexive movements by altering muscle tonethrough the γ-motor system.
Autonomic Pathways
The hypothalamus is the central integrator and distributorof important autonomic projections to the brainstem andspinal visceral nuclei, and it regulates motor functions of thesympathetic and parasympathetic systems (see Chapter 16).Brainstem reticular nuclei, including the locus ceruleus,also relay projections that regulate spinal autonomic motorfunctions. Cells in the brainstem project to the cervical andthoracic segments, and they are primarily excitatory to thephrenic motor neurons and thoracic motor neurons par-ticipating in inspiration, vomiting, and coughing reflexes(see Chapter 16).
Ascending TractsThe ascending fibers of the spinal cord transmit sensoryinformation from various body parts (Fig. 11-5). The sen-sory impulses are pain, thermal sense, touch, propriocep-tion, and kinesthesia (see Chapter 7). Projections from thesensory cortex refine the cortical motor efferents to thespinal cord.
The fasciculus gracilis is composed of axons arisingfrom the spinal DRG in the sacral, lumbar, and lower sixthoracic levels (Fig. 11-5). The fibers of the fasciculus gra-cilis mediate the sensations of discriminative touch, jointmovement, and vibration from the lower half of the body.
Consisting of large fibers, the fasciculus cuneatus medi-ates the sensations of fine discriminative touch, joint move-ment, and vibration from the upper half of the body. Its fibersenter the cord from the upper six thoracic and all cervical lev-els (Fig. 11-5). Along with the fasciculus gracilis, the fascicu-lus cuneatus fibers terminate in the medullary relay nuclei ofcuneatus and gracilis. Secondary fibers from these nuclei(internal arcuate fibers) decussate to form the medial lem-niscus, which ascends to the ventral posterolateral nucleusof the thalamus and then to the primary sensory cortex (seeChapter 7).
The anterior spinothalamic tract is a backup sensorysystem. Its fibers mediate the sensation of crude and non-localizable touch (Fig. 11-5; see Fig. 7-8). After emergingfrom the cells in the spinal gray matter, these fibers crossthe midline through the anterior commissure within sev-eral segments of the cord before ascending in the anteriorspinothalamic tract. They terminate in the ventral postero-lateral nucleus of the thalamus, and from there they pro-ject to the primary sensory cortex.
The fibers of the lateral spinothalamic tract transmitpain and temperature sensation (Fig. 11-5). Fibers ofthis tract, like those of the anterior spinothalamic tract,arise from the spinal dorsal horn nuclei and ascend inthe lateral fasciculus to the thalamus and then to thecortex (see Chapter 7).
Spinocerebellar Tracts
The fibers of the ventral spinocerebellar tract mediate un-conscious proprioception from the muscles of the lower
Anteriorcorticospinal tract(uncrossed—10%)
Medullary pyramid
Lefthemisphere
Righthemisphere
Lateralcorticospinal tract (crossed—90%)
Figure 11-6 Corticospinal fibers predominantlyproject to the contralateral half of the body.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 244

extremities and proximal limbs, and they coordinate themovement and posture of the lower extremities (Fig. 11-5;see Fig. 7-10). The uncrossed fibers of the dorsal spino-cerebellar tract mediate unconscious proprioception fromthe distal regions of the lower limbs. The cuneocerebellarfibers mediate unconscious proprioceptive information fromthe upper limbs to the cerebellum (see Chapter 7); these areconcerned with fine and delicate control of the upper limbs.
MOTOR NUCLEI OF THE SPINAL CORD
The spinal gray matter contains thousands of motor nervecells, which are either anterior motor neurons or inter-neurons (internuncial cells). The anterior motor neuronsinclude �- and �-motor neurons (Fig. 11-7). The efferentfibers of α- and γ-motor nerve cells form the ventral rootsof spinal nerves that innervate skeletal muscles. Theseare called LMN to distinguish them from the corticospinalneurons of the motor (and sensory) cortex, which are calledUMN. The interneurons function as association cells inter-connecting cell bodies within sensory and motor neuronpools. The dendritic and axonal projections of the inter-nuncial cells connect adjacent spinal cord cells.
The cervical spinal cord gray matter also contains twospecialized motor nuclei. One supplies the phrenic nerve,which innervates the diaphragm and participates in respira-tion (see Chapter 16). The other motor nucleus contributesto the spinal roots of the spinal accessory (CN XI) nerve,which regulates head and shoulder movements (seeChapter 15).
The α- and γ-motor neurons receive motor impulsesdirectly from the motor centers in the forebrain andbrainstem. The supraspinal projections to α- and γ-motorneurons, which initiate voluntary motor activities, arerepresented by balanced inhibitory (−) and excitatory (+)cortical outputs to the spinal motor neurons.
�-Motor Neuronsα-Motor neurons are the major motor neurons of the spinalcord. Their axons, with a diameter of 9–16 µm, rapidly con-duct impulses. They pass through the ventral spinal rootbefore innervating the extrafusal fibers of the skeletalmuscles that are responsible for the voluntary and reflex-ive movements of the head, trunk, and extremities. Onaverage, one α-neuron fiber innervates > 200 musclefibers. These motor neurons are also the final commonpathway because all efferent impulses from the CNSmust pass through these cells before activating muscles.
�-Motor Neuronsγ-Motor neurons, which lie alongside the α-cells in thespinal ventral horns, are smaller, half as numerous, andhave smaller-diameter axonal processes. These neurons areslow impulse conductors. The primary role of γ-LMN neu-rons is to regulate the length of the spindle fibers and thusmodulate the excitability of the annulospiral primary (Ia)endings. This regulates the stretch reflex muscle tone andallows the CNS to regulate its own state of excitability.
γ-Motor neurons are controlled by synaptic input fromthe brainstem reticular formation and the vestibular sys-tem. The γ-efferent fibers leave through the ventral nerveroot and contract the end (contractile) portions of the intra-fusal muscle fibers, stretching the central parts of the mus-cle spindles. On being stretched, the muscle spindles send avolley of afferent projections to the corresponding α-motorneurons, causing the reflexive contraction of the extra-fusal fibers of the muscle.
InterneuronsAs functionally specialized cells, the interneurons are dif-fusely present throughout the spinal cord and brain. Thereare ~30 times as many interneurons as motor neurons. Withmost being inhibitory, the interneuron cells serve as filters,integrating all sensory and motor functions of the CNS.The Renshaw cell receives axonal collaterals from nearbymotor neurons, inhibiting the activity of the same or relatedadjacent motor neurons. This recurrent inhibition by theRenshaw cell facilitates and sharpens the activity of the pro-jecting motor neuron from which it receives the collaterals.
MOTOR FUNCTIONS OF THE SPINAL CORD
The basic motor function of the spinal cord is a reflexivemotor response. A reflex response is a stereotyped move-ment to sensory stimulation. The neuronal circuitry forreflexes is present at each segmental level throughout thespinal cord. It consists primarily of muscle spindles, affer-ent fibers, �-motor neurons, efferent fibers, and muscletissue (Fig. 11-8).
α-Motorneurons
γ -Efferentfibers
γ-MotorneuronsInterneurons(internuncial
cells)
α-Efferentfibers
Figure 11-7 α-Cells, γ-cells, and internuncial cells in theanterior gray column of the spinal cord.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 245
10353-11_CH11.qxd 8/30/07 1:16 PM Page 245

246 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
For the most part, reflex functions of the spinal cord areindependent of cortical voluntary control, although theyare influenced indirectly by descending impulses from themotor cortex and brainstem motor centers. The input fromhigher motor centers participates in providing a homeo-static state of motor control, resulting in smooth motormovements. If released from higher levels of motor control,as in the case of a lesion, the spinal cord reflexes becomehyperactive.
Muscle Spindles and Their Role in Motor ActivityThere are two types of specialized receptors in muscles:muscle spindles and Golgi tendon organs (Fig. 11-9).Muscle spindles detect the degree and rate of change in mus-cle length and help maintain muscle tone. Golgi tendonorgans monitor the degree of muscle tension during mus-cle contraction and reflexively inhibit muscle contraction,permitting the muscle to stretch and thus prevent injurycaused by excessive contraction.
Muscle Spindles
As a complex sensorimotor organ, the muscle spindle isa small structure and consists of three to five specializedintrafusal fibers that are parallel to the surroundingextrafusal (striate) muscle fibers. The center of an intra-fusal fiber is wrapped by a fast-conducting annulospiralprimary (Ia) sensory ending. If stretched, it generatesan afferent response in the afferent fiber from the spindle.The afferent impulses travel to α-motor neurons in thespinal cord via fast-conducting type Ia nerve fibers witha velocity of approximately 100 m/sec, causing the mus-cle to contract (Fig. 11-9A).
Muscles consist of extrafusal and intrafusal fibers.Extrafusal fibers make up the large mass of the skeletal
(striated) muscle. They are attached to bone by fibrous tis-sue extensions called tendons and are controlled by α-motorneurons. Striated muscles are composed of myosin fila-ments, which are responsible for the contractility (shorten-ing) of the muscle. Intrafusal fibers, which contain musclespindles, are attached to the extrafusal fibers and are con-trolled by γ-motor neurons. Both ends of the intrafusalfibers contract; but the central region, which is devoid ofmyosin filaments, does not contract.
The γ-motor neurons control both ends of the intra-fusal fibers. A contraction of both ends of these fibers causesthe central portion of the fiber to stretch passively. A sim-ilar stretch of the spindles can occur if the entire muscle(skeletal) mass is stretched. Whenever the central por-tion of the intrafusal fiber stretches, the annulospiral pri-mary sensory endings become depolarized and dischargeimpulses. These impulses travel in two types of sensorynerve fibers: type Ia (fast) fibers and type II (slow) fibers.Type Ia (primary) fibers innervate the central region of thespindles, whereas the type II (secondary) fibers innervateareas of the intrafusal fibers on each side of the primaryending (Fig. 11-9A).
As the spindles stretch, a surge of sensory input isdirected to the α-motor neurons, which in turn reflexivelycontract the muscle mass to progressively decrease the mus-cle length. The contraction of the entire muscle halts thestretch of the spindles. With diminished spindle stretching,sensory input from the intrafusal fibers to the α-motor neu-rons stops.
Stretching the central portion of the intrafusal fiberscan induce a stretch reflex. This can be elicited by eithercontracting the ends of the intrafusal fibers by activatingγ-nerve impulses or lengthening the surrounding striatemuscle. The latter is illustrated by the classic knee-jerk(stretch or myotatic) reflex. When the patient’s leg is in a
Muscle
Dorsal rootganglion
α-Motor neuron(final common path)
Afferent fibers
Efferent fibers
Spinal nerve
Neuromuscularjunction
Musclespindles
Figure 11-8 Neuronal circuitry for a spinal reflex.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 246

A
γ-Motorneuron
α-Motorneuron
Group Ia afferent
Group IIafferent
γ-Motornerve
Extrafusalmuscle fibers
Secondary endings
Primary endings
Nuclear chain fiber
Nuclear bag fiber
Golgi tendon organ➡
➡
➡
➡
γ-Motornerve
Intrafusalmuscle fibers
Secondary endings
Dynamic γ-fiber(efferent)
Group Ia fiber(primary afferent)
Group II fiber(secondary afferent)
Nuclear bag fiber
Nuclear chain fiber
B
Static γ-fiber(efferent)
Afferent fiber
Golgi tendonorgan
α-Efferent fiber
Inhibitoryinterneuron
α-Motorneuron
C
1b
Figure 11-9 A. The relation of extrafusal and intrafusal muscle fibers to musclestretch (muscle spindle) and tension (Golgi tendon) receptors. B. Nuclear bag andnuclear chain fibers. C. Golgi tendon organs, which mediate autogenic inhibition.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 247
10353-11_CH11.qxd 8/30/07 1:16 PM Page 247

248 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
relaxed state, a tap of the patellar tendon at the knee witha reflex hammer immediately stretches both the extrafusaland intrafusal fibers of the quadriceps muscle. After a briefdelay, the stretch is followed by a reflexive contraction ofthe same muscle. The response entails a spinal cord reflextriggered by sensory impulses in the annulospiral nerveendings of the intrafusal fibers that were stretched by theknee tap.
The intrafusal fibers are divided into nuclear bag andnuclear chain fibers (Fig. 11-9B). Nuclear bag fibers arelong and have many nuclei in their center. The nuclearchain fibers are smaller and have fewer nuclei. Primary(type Ia) sensory endings innervate the central part ofboth the nuclear bag and nuclear chain fibers. Secondary(type II) sensory endings innervate the nuclear chain fibersonly at sites beyond each end of the primary sensory end-ings. The intrafusal nuclear bag fibers mediate dynamicsensory responses, and nuclear chain fibers mediate staticresponses.
Dynamic Responses
Dynamic responses are mediated through primary (type Ia)sensory endings that terminate on the nuclear bag fibersof the muscle spindle. With a stretch of the muscle massand/or the muscle spindle, there is a simultaneous distor-tion of the primary sensory nerve endings. This causes asurge of dynamic sensory input to the α-motor neurons ofthe spinal cord. The α-motor impulses from the anteriorhorn cells activate a contraction of the muscle mass, short-ening the muscle. Sensory input from the type Ia nuclearbag fibers stops when intrafusal fibers cease stretching.
Static Responses
Static responses are generated in the secondary sensory end-ings of the nuclear chain fibers. Intrafusal fibers respondto stretch, a response which is proportional to the intensityof the stretch. The static sensory input from the intrafusalchain fibers maintains the muscle at the stretched positionlonger, from several minutes to hours.
Golgi Tendon Organs
Golgi tendon organs are the second type of sensory musclereceptors (Fig. 11-9C). They innervate the tough tissuesthat attach muscles to bones. The Golgi afferent impulsesregulate muscle tension and prevent damage from exces-sive muscle contraction. Whenever the muscle stretch orcontraction is excessive, type Ib projections from the Golgitendon organs have an effect opposite to that of type Iaprojections from muscle spindles. Excessive tension in themuscles stimulates the tendon organ type Ib fibers, whichactivate the intervening interneurons of the spinal cord.The interneurons in turn inhibit the spinal motor nuclei tothe muscle. This accounts for autogenic inhibition. TheGolgi-mediated reflex is protective, preventing the gener-ation of a too-sudden or excessive force that could damagethe muscle or its insertion.
Movement InitiationMuscle contractions can be initiated and modified througheither the γ-motor or the α-motor systems. Increased activ-ity of one system is accompanied by increased dischargesin the other system. This causes the muscle to assume anew, appropriate length. A situation in which both α- andγ-motor neurons are at subthreshold, with the muscle atits resting length, is demonstrated in Figure 11-10A. In theresting state, the spindles are stretched adequately. Anyfurther stretching of the spindles depolarizes them andtriggers a volley of action potentials to the α-motor neurons.
One way to alter this resting state and initiate a musclecontraction is through the stimulation of α-motor neurons.Activating them causes the extrafusal muscle fibers to con-tract. With contraction of the extrafusal fibers of the mus-cle, the intrafusal fibers become slack, and consequently,the spindles lose their sensitivity to muscle length. Tocorrect this impaired spindle sensitivity, the rubrospinal,vestibulospinal, and reticulospinal tracts reflexively dis-charge γ-motor neurons and contract the end portions of the intrafusal fibers, straightening the spindles. As aresult, the spindles regain their sensitivity to muscle length(Fig. 11-10A).
The other way to alter the resting state and initiate amuscle contraction is to contract the ends of the intrafusalfibers by way of the γ-motor neurons (Fig. 11-10B). The γ-mediated contraction of the ends of intrafusal fibersincreases the sensitivity of the spindles and their afferentfibers. The annulospiral endings send a volley of actionpotentials to the α-motor neurons on type Ia sensory fibers,shortening the extrafusal fibers. Once the muscle has con-tracted enough to decrease the stress on the center of theintrafusal fibers, the rate of the type Ia firing decreases, andthe extrafusal fibers cease contracting. The new desiredmuscle length permits maintenance of equilibrium inwhich the activity in type Ia fibers is below threshold.
SPINAL REFLEXES
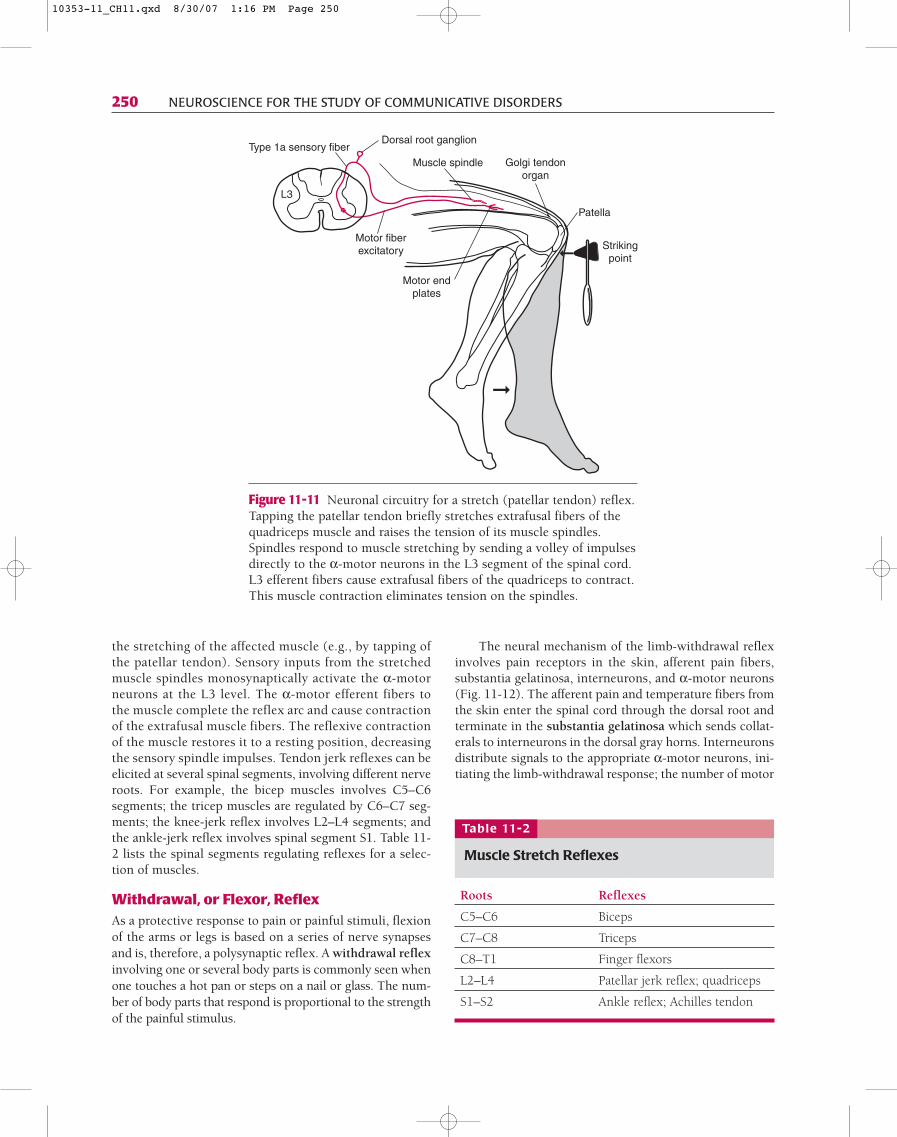
Stretch, or Myotatic, ReflexThe muscle stretch reflex, the simplest of all, involves asingle synapse in the spinal cord. The classic example ofthis monosynaptic, or two-neuron, reflex is the knee-jerk, which is initiated by tapping the patellar tendon ofthe quadriceps femoris muscle with a reflex hammer(Fig. 11-11). A tap of the tendon (input) leads to a briefstretch of the muscle, which stimulates the sensory end-ings of spindles. The muscle spindles send afferent pro-jections to α-motor neurons, the activation of whichcauses a quick contraction (muscle jerk) of the samemuscle (output).
The common element in all stretch reflexes is thatthe stretched muscle contracts after a brief delay. The prin-cipal receptors are the muscle receptors that respond to
10353-11_CH11.qxd 8/30/07 1:16 PM Page 248

A
1.
2. 3.
Rest length
Stimulated α-motorneuron contractsextrafusal fibers
Activation of γ-motor neuroncontracts intrafusal fibers, restoring
spindle sensitivity
α-Motorneuron
Primarysensory
γ-Stimulus
Extrafusal fibers
Intrafusal fiberscontract
α-Motorneuron
γ-Motorneuron
Extrafusal fibers
B
1.
2. 3.
α-Motorneuron
γ-Motorneuron
α-Motor Primary endings
→
γ-Motor
Primaryafferent
→
→γ-Efferent
→ →→
→→
γ-Motor
γ-Efferent
Figure 11-10 The α- and γ-motor neurons in movement initiation and themaintenance of sensitivity to stretch in the muscle spindles. A. α-Mediatedvoluntary contraction of the extrafusal muscle fibers leaves intrafusal fibersand their spindles without sensitivity to stretch. This condition is corrected bythe reticular and vestibulospinal projections to the γ-motor neurons, whichcontract intrafusal fibers and restore spindle stretch sensitivity. B. In γ-initiatedmovement, γ-cells activate intrafusal fiber ends, deforming the annulospiralendings of the spindle afferent fibers, which synaptically activate α-motorneurons, causing contraction of the extrafusal muscle cells.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 249
10353-11_CH11.qxd 8/30/07 1:16 PM Page 249
250 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
the stretching of the affected muscle (e.g., by tapping ofthe patellar tendon). Sensory inputs from the stretchedmuscle spindles monosynaptically activate the α-motorneurons at the L3 level. The α-motor efferent fibers tothe muscle complete the reflex arc and cause contractionof the extrafusal muscle fibers. The reflexive contractionof the muscle restores it to a resting position, decreasingthe sensory spindle impulses. Tendon jerk reflexes can beelicited at several spinal segments, involving different nerveroots. For example, the bicep muscles involves C5–C6segments; the tricep muscles are regulated by C6–C7 seg-ments; the knee-jerk reflex involves L2–L4 segments; andthe ankle-jerk reflex involves spinal segment S1. Table 11-2 lists the spinal segments regulating reflexes for a selec-tion of muscles.
Withdrawal, or Flexor, ReflexAs a protective response to pain or painful stimuli, flexionof the arms or legs is based on a series of nerve synapsesand is, therefore, a polysynaptic reflex. A withdrawal reflexinvolving one or several body parts is commonly seen whenone touches a hot pan or steps on a nail or glass. The num-ber of body parts that respond is proportional to the strengthof the painful stimulus.
The neural mechanism of the limb-withdrawal reflexinvolves pain receptors in the skin, afferent pain fibers,substantia gelatinosa, interneurons, and α-motor neurons(Fig. 11-12). The afferent pain and temperature fibers fromthe skin enter the spinal cord through the dorsal root andterminate in the substantia gelatinosa which sends collat-erals to interneurons in the dorsal gray horns. Interneuronsdistribute signals to the appropriate α-motor neurons, ini-tiating the limb-withdrawal response; the number of motor
Muscle spindle Golgi tendonorgan
Motor endplates
L3
Strikingpoint➞
➞
Type 1a sensory fiber
Motor fiberexcitatory
Dorsal root ganglion
Patella
Figure 11-11 Neuronal circuitry for a stretch (patellar tendon) reflex.Tapping the patellar tendon briefly stretches extrafusal fibers of thequadriceps muscle and raises the tension of its muscle spindles.Spindles respond to muscle stretching by sending a volley of impulsesdirectly to the α-motor neurons in the L3 segment of the spinal cord.L3 efferent fibers cause extrafusal fibers of the quadriceps to contract.This muscle contraction eliminates tension on the spindles.
Roots Reflexes
C5–C6 Biceps
C7–C8 Triceps
C8–T1 Finger flexors
L2–L4 Patellar jerk reflex; quadriceps
S1–S2 Ankle reflex; Achilles tendon
Table 11-2
Muscle Stretch Reflexes
10353-11_CH11.qxd 8/30/07 1:16 PM Page 250

neurons recruited by pain fibers depends on the strength ofthe stimulus. The reflex continues for several seconds afterthe stimulus ceases. A withdrawal reflex generally beginseven before one is aware of the painful stimulus becausethe afferent information triggers a spinal response beforethe ascending signal of pain reaches the forebrain.
Reciprocal inhibition of the motor neurons is a neu-ronal arrangement. The stimulation of one group of neu-rons causes inhibition of an associated group of neurons.This neuronal arrangement is essential for smooth motorfunction. For example, in arm flexion, the interneuronsactivating the motor neurons for the biceps simultane-ously inhibit the motor neurons for the paired triceps. Thisantagonistic effect—wherein one muscle contracts whilethe paired muscle extends because it is inhibited fromsimultaneous contraction—exemplifies reciprocal inhibi-tion (Fig. 11-12). Anatomically, this involves interneuronsthat are inhibitory to α-motor neurons of the antagonisticmuscle.
Crossed, or Intrasegmental, Extensor ReflexThe crossed extensor reflex is a complex movement pat-tern in which withdrawal of the limb (flexor response)on one side is accompanied by the extension of the oppo-site limb ~0.5 sec after the flexor response (Fig. 11-13).This multisynaptic reflex system, which moves limbs onthe opposite side of the body, is considered a geneticallyprogrammed protective behavior for survival because itmoves the entire body away from the painful stimulus.Its neural mechanism involves the crossing of sensoryinformation to the opposite side through polysynapticcircuits of interneurons recruiting the opposite limbs.The crossed limb extension follows the flexing action ofthe limb ipsilateral to the stimulus. Reverberating poly-synaptic circuits in interneuronal pools thus sustain thecomplex withdrawal behavior for long periods, even afterthe reflex-triggering stimulus has ceased. This is neces-sary to keep the body protected until the brain takes overbody control.
Dorsal rootganglion
α-Motorneuron
Internuncialcell Afferent fibers
Efferentfibers
Sensorynerve
WITHDRAWALREFLEX
Motornerve
Bicepscontraction
Triceps inhibitionrelaxation
Inhibitoryinterneuron
Tricepsmotor cell
Painful stimulus
Figure 11-12 Neuronal circuitry for the withdrawal (flexor) reflex. Apainful stimulus to the hand results in a reflexive flexing of the upperlimb, causing the withdrawal of the hand from the painful stimulus.The circuitry involves diverging interneuronal elements. This circuitrynot only withdraws the limb but also inhibits the antagonistic musclefrom contracting through reciprocal inhibition.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 251
10353-11_CH11.qxd 8/30/07 1:16 PM Page 251

252 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
NEUROTRANSMITTERS
Four important neurotransmitters are released by theactivity of brainstem projections to the spinal cord: acetyl-choline, norepinephrine, serotonin, and epinephrine.Acetylcholine is the major chemical messenger of theperipheral nervous system (PNS). Released by the efferentspinal fibers in the myoneural junction, acetylcholine regu-lates voluntary or reflexive motor movements. A diminishedeffect of acetylcholine occurs in myasthenia gravis and sim-ilar disorders associated with muscle weakness. Weaknesscan be caused by either excessive action of the enzymeacetylcholinesterase or a reduced number of acetylcholinereceptors at the myoneural junction. Acetylcholine alsoregulates autonomic functions. Except for the sympatheticpostganglionic cells, acetylcholine is the primary neuro-transmitter of the autonomic nervous system, which con-trols major visceral functions (see Table 16-1).
The pontine reticular nucleus ceruleus and lateralmedullary reticular formation transmit epinephrine andnorepinephrine to the spinal cord (see Fig. 13-1). Theirinfluence is thought to be inhibitory, enhancing the signal-
to-noise ratio in the spinal sensorimotor conduction sys-tem. These neurotransmitters are slow acting and have along-lasting effect.
Located in the brainstem, the caudal reticular raphenuclei send serotonin projections to the lower brain-stem and spinal cord. The projections to the spinal cordsynapse in the ventral and dorsal horns and in the sym-pathetic lateral columns. In addition, they synapse onspinal enkephalin interneurons and provide some con-trol over pain transmission.
CLINICAL CONCERNS
Trauma, tumors, infections, impaired blood circulation, anddegenerative conditions are common causes of spinal cordlesions. The testing of sensory and motor functions is themost reliable clinical method for determining the integrityof the spinal cord. The contraction of striate muscles dur-ing reflexes provides a clinician with important informa-tion about the complex internal mechanism of the entiremotor system. Muscle reflexes are examined clinically on
Dorsal rootganglion
Motor cell
Internuncialcell
Spinal nerve
CROSSEDEXTENSION REFLEX
FLEXOR REFLEX
Painful stimulus
➠
➠
Tricepsexcitation
Bicepsinhibition Biceps
facilitation
Tricepsinhibition
Inhibitoryinterneuron
Dorsal rootganglion
Afferentfibers
Efferentfibers
Afferent fibers
Efferentfibers
Figure 11-13 Neuronal circuitry for the crossed extensor reflex marked by thecontraction of the muscle on one side accompanied by the extension of theopposing muscle or muscles. This reflex involves interneurons that diverge sensoryinformation. This crossed sensory information activates motor neurons to extendthe agonistic muscle, whereas the interneurons inhibit the motor nucleus for theantagonistic muscle, ensuring a smooth extension of the limb.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 252

both sides of the body to determine whether muscular move-ments are symmetrical and the quality of movements isnormal. Reduced (hypoactive) or increased (hyperactive)quality of muscle reflexes indicates pathology in the nervoussystem. Spinal reflex functions are controlled via balancedexcitatory (+) and inhibitory (−) supraspinal projections tothe motor neurons. The corticospinal system is predomi-nantly excitatory to motor neurons in the spinal cord. Lossof this system reduces recruitment of motor neurons andthus produces weakness. This muscle paresis is graduallyconverted over several weeks into spasticity, which includeshyperexcitability of many stretch reflexes (UMN syndrome).In contrast to brisk (hyperactive) reflexes with interruptedsupraspinal fibers, lesions involving spinal motor neuronsand/or their efferent spinal fibers lead to reduced or absentmuscle reflexes (hyporeflexia or areflexia, respectively).
There are two types of spinal cord disorders: segmentaland longitudinal (pathway specific). Segmental disturbanceimplies a lesion at a specific spinal level; below that level,sensory and motor functions are impaired. The severity ofthe deficit depends on the site and extent of the lesion.Longitudinal disturbance selectively affects specific nervecells and their axons. The longitudinal involvement ofaxonal bundles may impair both sensory and motor systems.
Lower Motor Neuron SyndromeThe term lower motor neuron (LMN) refers to a motor neu-ron and its axon in the brainstem or spinal cord. A LMN cellbody provides the output pathway to peripheral functionvia its axon, which traverses the ventral root and peripheralnerves to innervate a skeletal muscle. LMN lesions of eitherthe spinal cell bodies in the ventral horn (e.g., poliomyelitis,
amyotrophic lateral sclerosis, vascular damage, and spinalcord tumor) or of the axon (in the ventral root or peripheralnerve) result in denervation of the skeletal muscle fibersand loss of muscle power (weakness) and precise control.
Small lesions can result in the loss of one or severalmotor units. Large lesions or peripheral nerve destructioncan result in complete muscle weakness and total flaccid-ity. Such a lesion results in LMN syndrome (Fig. 11-14;see Table 14-1), in which muscle fibers are disconnectedfrom motor efferents and thus cannot receive descendingcortical impulses and reflexive sensory input. Deprived oftheir trophic efferents, the affected muscle fibers graduallydegenerate. Clinical signs of LMN include flaccid paralysis,absent reflexes, muscular fibrillation, and eventual severeatrophy (wasting) of the muscle involved; these signs occurunilateral to the lesion.
With no projections of motor impulses from the motorneuron, the muscle fibers are completely paralyzed for bothreflexes and voluntary motor movements; this paralysis ischaracterized by flaccid muscle tone. Denervated musclefibers pass through several stages: a brief period of hyper-excitability and spontaneous firing (fibrillation), followedby silence of firing and shrinking of the muscle (atro-phy). Fibrillation is the contraction of individual dener-vated muscle cells, which contract under the influence ofacetylcholine circulating in the blood. If they are not rein-nervated within 6 months or so, the skeletal muscle cellsdie and are replaced permanently by scar tissue. This ismore severe than reduced muscle mass from disuse, whichoften is seen in UMN syndrome.
If the LMN survives, the normal membrane potentialeventually is restored and the unit returns to its normal
Corticospinalprojection
Muscle
Lower motor neuron syndrome • Flaccid paralysis • Muscular twitchings • Muscle atrophy • Hyporeflexia
Afferent fibers
Efferent fibers
Figure 11-14 Injury to the motor unit (lower motor neuron syndrome).
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 253
10353-11_CH11.qxd 8/30/07 1:16 PM Page 253

254 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
condition, except for when the motor system excites thecell body past threshold. Destruction of the LMN cell bodyor its axon results in denervation of the muscle fibers ormotor unit. Destroyed LMN cell bodies are not replaced.However, destroyed LMN axons (PNS) may regenerate, andif they are directed properly past scar tissue and throughtheir former peripheral nerve path, they can reinnervate totheir former muscle fiber targets. Skilled peripheral nervesurgery, inhibition of scar tissue formation, and effortsto maintain denervated muscle fibers until reinnervationoccurs are all necessary for the success of reinnervation.
Common Spinal SyndromesComplete Spinal Transection
Vertebral dislocations, myelitis (inflammation of the spinalcord), and tumors can cause spinal transection. Immediatelyafter the transection, all sensory and motor functions are lost bilaterally below the lesion but are spared abovethe lesion (Fig. 11-15; see “Lesion Localization” later inthis chapter). Thus the body regions affected depend on thespinal level of the lesion. Spinal shock abolishes sensori-motor functions and persists for weeks. After a while, the
A
B
Lateralspinothalamic tract
(crossed sensory fibers)
Lateralcorticospinal tract
(crossed motor fibers)
Dorsal lemniscal fibers(uncrossed discriminative
touch fibers)
Complete spinal sectionat T12
Figure 11-15 Complete transection of the thoracic spinal cord (A), resultingin bilateral paralysis and sensory loss below the level of the lesion withspared functions above the level of lesion (B).
10353-11_CH11.qxd 8/30/07 1:16 PM Page 254

Loss of pain and temperature
Upper motor neuron monoplegiaLoss of discriminative touch
B
Lateralspinothalamic tract(crossed pain and temperature fibers)
Lateralcorticospinal tract
(crossed motor fibers)
Dorsal lemniscal fibers(uncrossed discriminative
touch fibers)
Right Left
A
Figure 11-16 A spinal cord hemisection at T12 (A) results in a typicalsensorimotor deficit pattern characterized by ipsilateral paralysis,ipsilateral sensory loss (discriminative sensation), and contralateral painand temperature loss (B).
reflex activity gradually returns at levels below the lesion.However, with the interruption of the corticospinal tract,after several weeks the patient exhibits UMN syndrome,which includes loss of delicate manipulative capabilitiesin the forearm and fingers, hyperactive stretch reflexes,Babinski sign, and clonic movements in which a passivelymoved limb undergoes rapid and repeated contractionand relaxation. With spastic legs, the patient eventuallyassumes an extended posture, which is characterized by
extended lower limbs. If the lesion is above the level ofinnervation of the upper limbs, he or she often also assumesa flexed posture (see Chapter 14).
Brown-Séquard Syndrome: Spinal Hemisection
Lateral hemisection of the spinal cord produces the follow-ing three clinical conditions; understanding them requiresknowledge of the locations and crossing points of the sen-sorimotor pathways (Fig. 11-16).
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 255
10353-11_CH11.qxd 8/30/07 1:16 PM Page 255

256 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
• Signs of a lesion in the corticospinal tract on the ipsi-lateral half of the body. In the case of a right-sidedlesion at C4, there is spastic paralysis in the right armand leg (UMN symptoms). Babinski and Hoffman signsare also present. The Babinski sign is abnormal exten-sion of the big toe in response to a scraping stimulus tothe sole of the foot (Fig. 14-5). The Hoffmann sign iswhen the thumb and other fingers flex from the flickingof the nail or distal phalanx.
• Ipsilateral sensory loss. Destruction of the dorsal lem-niscal column, which contains tactile and proprioceptiveaxons ascending ipsilaterally to relay to the contralateralforebrain, results in loss of vibratory and discriminativesensation in the ipsilateral half of the body. The ascend-ing dorsal lemniscal fibers cross the midline in the cau-dal medulla (see Chapter 7).
• Contralateral pain and temperature sensation loss. Withfibers of the lateral spinal thalamic tract crossing imme-diately after entering the spinal cord, an interruption atthe right C4 level affects pain and temperature from theleft (contralateral) side of the body below the level ofthe lesion (see Chapter 7).
Syringomyelia
Syringomyelia is a developmental condition marked bya cyst or cavity within the central portion of the spinalcord. It is characterized by two clinical symptoms: lossof pain and temperature sensation and impaired motorcontrol. The central (gray) region is the site of degenera-tion, and the crossing of sensory (pain and touch) fibers isinterrupted, which results in a bilateral pattern of pain andtemperature loss (Fig. 11-17A). If the cyst extends into thespinal motor nuclei in the involved segments, bilateralsigns of LMN syndrome in the involved muscles includeflaccid paralysis, hyporeflexia, and hypotonia.
The sensorimotor symptoms depend on the level ofthe spinal cord implicated. For example, a cavitation inthe central spinal area at C4–C6 produces bilateral loss ofpain and temperature for the arms, forearms, thumbs, andindex fingers. A large cavitation involving C3–T4, in addi-tion to the loss of pain and temperature, affects the shoul-ders and chest (Fig. 11-17B).
Subacute Combined Degeneration
Subacute combined degeneration is associated with perni-cious anemia (when the number of red blood cells and theamount of hemoglobin in blood is less than normal), whichresults from malabsorption of vitamin B12. The bilateral sub-acute degeneration of the spinal cord mostly involves thefibers in the dorsal lemniscal column and corticospinaltract. Involvement of the dorsal column fibers results inbilateral loss of position and vibratory sense, whereas theinterruption of the corticospinal fibers produces a weaknessof limbs bilaterally. The presence of paresthesia (numb-ness and tingling) indicates peripheral nerve involve-
ment. As the disease progresses to the cerebral cortex,higher mental functions are affected.
LESION LOCALIZATION
Rule 5: Complete Spinal Cord LesionPresenting Symptoms
Paralysis and sensory loss bilaterally below the level of thelesion with spared functions above indicate a completespinal cord trans-sectional injury.
Rationale
The spinal cord contains all ascending (sensory) anddescending (motor) fibers. A complete lesion affects thetransmission of these functions below the lesion pointand spares such functions above the lesion point. Impairedbowel and bladder control and autonomic reflexes are com-monly seen in a spinal cord injury.
Rule 6: Spinal Hemisection: Brown-Séquard SyndromePresenting Symptoms
Ipsilateral loss of position and vibratory sensation belowthe level of the lesion, ipsilateral body paralysis, and con-tralateral loss of pain and temperature indicate a spinalhemisection.
Rationale
1. Transection of the fasciculus gracilis and fasciculuscuneatus results in the loss of position and vibrationsense along with discriminative touch on the side ofthe lesion.
2. Involvement of the fibers of the corticospinal tract belowits decussation produces spastic hemiplegia on the sideof the lesion.
3. Because the fibers mediating pain and temperature crossthe midline after entering the spinal cord, their inter-ruption results in loss of pain and temperature sensa-tion on the opposite side.
4. Impaired bowel and bladder control is commonly seenin spinal cord injury.
Rule 7: Peripheral or Central LesionPresenting Symptoms
Paralysis and sensory (pain and temperature) loss affectingthe same single limb suggest a lesion either in the periph-eral nerve or in the cortex.
Rationale
The descending and ascending sensory fibers supplyinga single limb are together only in the peripheral (nerve,plexus) area or in the somatosensory cortical area.
10353-11_CH11.qxd 8/30/07 1:16 PM Page 256

A
B
Lateralspinothalamic
tract
Figure 11-17 A syringomyelic cavity in the cervical gray matter (A) interruptsthe crossing of pain-mediating spinothalamic fibers, causing a bilateral loss ofpain and temperature and possibly bilateral flaccid paralysis of the muscles ofthe upper limbs (B).
C L I N I C A LC O N S I D E R A T I O N S
P A T I E N T O N E
A 3-year-old girl woke up with pain in her right leg.She had no control of her right leg and could not walk.She was taken to the hospital and examination revealedthe following:
• No voluntary movement in the right leg• Diminished tone in the leg muscles• Absent knee-jerk and Achilles reflexes
A spinal MRI study revealed no abnormality. Clinicalanalysis of the CSF withdrawn through spinal taprevealed increased white blood cells, indicating aninfection.
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 257
10353-11_CH11.qxd 8/30/07 1:16 PM Page 257

258 NEUROSCIENCE FOR THE STUDY OF COMMUNICATIVE DISORDERS
Question: How can you explain these clinical signs?
Discussion: An acute viral infection, which was diag-nosed by the chemical analysis of the CSF. The infec-tion had affected spinal LMNs from L1–L5 in the lumbar region and thereby caused weakness of the leg.
P A T I E N T T W O
A 30-year-old window washer fell from her ladder andinjured her back at the T11–T12 level. She had bilateralflaccid paralysis of the legs and was rushed to the hos-pital, where she was observed for a few days. In anexamination completed a few weeks later, she demon-strated the following symptoms:
• Spastic paralysis in the left leg• Positive Babinski sign in the left leg• Loss of position sense (proprioception) and discrim-
inative touch in the left leg• Loss of pain and temperature in the right foot
A spinal MRI study revealed a crushed appearance ofthe cord at T11–T12 on the left.
Question: How can you relate these symptoms to acrushed cord at the T11–T12 level?
Discussion: This is an example of Brown-Séquard syn-drome, which is associated with spinal hemisection andis characterized by three signs:
• Paralysis on the side of the lesion• Loss of fine discriminative touch and position sense
on the side of the lesion• Impaired sensation of pain and temperature on the
side opposite to the lesion
Flaccid paralysis followed by spastic paralysis indicatesspinal shock secondary to an UMN lesion, which wasconfirmed by the positive Babinski sign in the left foot.The loss of position sense and discriminative touch fromthe left toes and leg indicates the involvement of the dor-sal column–medial lemniscal fibers; the loss of pain fromthe right leg suggests interruption of the left spinothala-mic pathway. Mediating pain and temperature sensationfrom the right side of the body, this pathway crosses inthe spinal cord at the level of entry. In this case, the ini-tial loss of functions indicated injury to both sides of thespinal cord, but permanent injury was restricted to theleft side only.
P A T I E N T T H R E E
A 16-year-old gymnast lost control of her body whileexercising and fell flat on her back. Unconscious forseveral minutes, she was rushed to a nearby hospital.
After she awoke, she was unable to move. A neurologicexamination revealed the following:
• Flaccid paralysis of both lower limbs• No deep tendon reflexes in either leg• Loss of touch, pain, and temperature sensation below
the midthoracic (T6–T7) region• No bladder or bowel control
A spinal MRI study revealed a complete spinal sectionat the midthoracic level. The patient’s clinical picturechanged within a week and she began to exhibit thesesigns:
• Hyperreflexia in both lower limbs• Spastic paralysis of both lower limbs• Impaired control of bowel and bladder functions• Some return of touch and pressure sensation in the
lower limbs
Question: How can you account for this clinical pictureby relating the symptoms to a complete midthoracicspinal section?
Discussion: A complete midthoracic transection of thecord affected the motor control of the lower limbs. Theabsence of muscle reflexes for a few days immediatelyafter the injury is called spinal shock. The return ofhyperactive reflexes and spasticity, indicating an UMNlesion, reflects the loss of descending inhibitory influ-ences. In UMN involvement, Babinski sign is alsoexpected. The loss of consciousness in this case wasattributed to a cortical concussion because it is not afeature of spinal cord injury.
P A T I E N T F O U R
A 45-year-old man was taken to a neurologist with a complaint of a progressively worsening weakness ofboth lower limbs for the past 2 months. The neurol-ogist noted the following:
• Paralysis (flaccid) in both lower limbs and fascicula-tion in the muscles
• Reduced reflexes involving both limbs• Loss of pain and temperature involving both
extremities• Impaired bladder function
A spinal MRI study revealed a large intraspinal, centrallyplaced tumor at the level of the conus medullaris.
Question: How can you relate these symptoms to thelesion site?
Discussion: The location of the tumor within the spinalcentral gray matter had affected the following struc-
10353-11_CH11.qxd 8/30/07 1:16 PM Page 258

tures, which are associated with the reported clinicalsymptoms:
• The tumor blocked the crossing fiber tract that car-ries pain and temperature. This subsequently resultedin a bilateral loss of pain and temperature from bothlower extremities.
• The tumor also encroached on the adjacent LMNtracts and neurons bilaterally, affecting lower limbmuscle strength.
• A lumbar involvement is usually associated withbowl and bladder functions.
P A T I E N T F I V E
A 54-year-old man, who was involved in a head-onautomobile accident, experienced difficulty breathing;he was taken to the emergency room (ER), where thefollowing signs and symptoms were noted:
• Paralysis of both right upper and lower limbs• Loss of discriminative sensation from the right half
of the body• Profound breathing problem marked with inability
to inhale• No difficulty in exhaling• Unintelligible speech marked with weakly articu-
lated one-word utterances
A spinal MRI study revealed a partial spinal lesion on theright involving C3–C5. The attending neurologists con-cluded that the partial lesion had affected the epicriticsystem, corticospinal tract fibers, and phrenic nerve.
Question: How can you relate these clinical symptomsto the lesion site?
Discussion: A lesion anywhere in the neuraxis is likelyto affect the descending corticospinal fibers and ascend-ing sensory fibers. However, a lesion only at the cervi-cal spinal cord level is likely to interrupt the phrenicnerve fibers that innervate the diaphragm, the primarymuscle of inspiration. In this case, the phrenic nerveinjury had caused respiratory paralysis with marked dif-ficulty in inhalation. A right cervical lesion also causedipsilateral paralysis (subsequent to the interruption ofthe crossed fibers of the corticospinal tract) and ipsilat-eral loss of discriminative sensation (subsequent to theinvolvement of the uncrossed ascending fibers of thedorsal lemniscal system).
SUMMARY
Motor function is hierarchically organized at four arbitrarilyidentified neuraxial levels, with the spinal cord being thelowest. Motor nuclei of the spinal cord are the final commonpathway for both spinal reflex and cortical projections tomuscle fibers. Consequently, the spinal cord is crucial inreflex muscle contractions and voluntary movements. Theenvironmentally triggered reflex responses regulated by thespinal cord include the stretch (myotatic) reflex, withdrawal(flexor) reflex, and crossed extensor reflex. Even thoughthese reflexes are independent of voluntary motor control,intact cortical projections to the spinal cord are important inthe regulation of these reflexes. Constantly relayed sensoryinformation, which is vital to coordinated motor activity,is integrated at every level of the nervous system. Spinallesions that interrupt both cortical and spinal reflex projec-tions to limb muscles result in LMN syndrome. The clinicalpicture of this syndrome is characterized by flaccid paraly-sis, absent reflexes, and atrophy of muscle fibers.
QUIZ QUESTIONS
1. Define the following terms: atrophy, hyporeflexia, lowermotor neuron, muscle spindles, myoneural junction,stretch reflex.
2. Name the parts of a motor unit.
3. List the clinical characteristics of LMN syndrome.
4. List the spinal locations of the motor neurons that inner-vate the muscles of respiration. (Also see Chapter 2.)
TECHNICAL TERMS
afferentα-motor neuronatrophyaxoncrossed extensor reflexefferentextrafusal fibersγ-motor neuronGolgi tendon organhyporeflexiainterneuron
intrafusal fiberslower motor neuronmotor unitmuscle spindlesmyoneural junctionreciprocal inhibitionreflexskeletal musclestretch reflexwithdrawal reflex
CHAPTER 11 ■ MOTOR SYSTEM 1: SPINAL CORD 259
10353-11_CH11.qxd 8/30/07 1:16 PM Page 259