1-s2.0-s1056872715000082-main(1)
DESCRIPTION
WDQWDASDTRANSCRIPT

�������� ����� ��
Decreased expression of Heat Shock proteins may lead to compromisedwound healing in type 2 diabetes mellitus patients
Kanhaiya Singh, Neeraj K. Agrawal, Sanjeev K. Gupta, Gyanendra Mo-han, Sunanda Chaturvedi, Kiran Singh
PII: S1056-8727(15)00008-2DOI: doi: 10.1016/j.jdiacomp.2015.01.007Reference: JDC 6389
To appear in: Journal of Diabetes and Its Complications
Received date: 1 December 2014Revised date: 9 January 2015Accepted date: 11 January 2015
Please cite this article as: Singh, K., Agrawal, N.K., Gupta, S.K., Mohan, G., Chaturvedi,S. & Singh, K., Decreased expression of Heat Shock proteins may lead to compromisedwound healing in type 2 diabetes mellitus patients, Journal of Diabetes and Its Complica-tions (2015), doi: 10.1016/j.jdiacomp.2015.01.007
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Kanhaiya et al. : Heat shock proteins and impairment of wound healing in T2DM cases
Decreased expression of Heat Shock proteins may lead to compromised wound healing in
type 2 diabetes mellitus patients
Kanhaiya Singh1, Neeraj K Agrawal
2, Sanjeev K Gupta
3, Gyanendra Mohan
4, Sunanda
Chaturvedi4 and Kiran Singh
1*
1Department of Molecular & Human Genetics, Banaras Hindu University,
Varanasi-221005, India
2Department of Endocrinology and Metabolism, Institute of Medical Sciences,
Banaras Hindu University, Varanasi-221005, India
3Department of Surgery, Institute of Medical Sciences, Banaras Hindu University,
Varanasi-221005, India
4Indian Railway Cancer Hospital and Research Centre, N.E.R.,
Varanasi- 221002, India
1* Correspondence
Kiran Singh
Department of Molecular & Human Genetics
Banaras Hindu University
Varanasi-221005, India
e-mail: [email protected], [email protected]
Telefax: +91-542-670-2499
Telephone: +91-9454210058

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2
Abstract
Background: Heat shock proteins (HSPs) are inducible stress proteins expressed in cells
exposed to stress. HSPs promote wound healing by recruitment of dermal fibroblasts to the site
of injury and bring about protein homeostasis. Diabetic Wounds are hard to heal and inadequate
HSPs may be important contributors in the etiology of diabetic foot ulcers (DFU).
Objective: To analyze the differential expression of HSPs and their downstream molecules in
human diabetic wounds compared to control wounds.
Methods: Expressional levels of HSP27, HSP47 and HSP70 and their downstream molecules
like TLR4, p38-MAPK were seen in biopsies from 101 human diabetic wounds compared to 8
control subjects without diabetes using RT- PCR, western blot and immunohistochemistry.
Results: Our study suggested a significant down regulation of HSP70, HSP47 and HSP27 (p
value = <0.001 for HSP70; p value = 0.007 for HSP47; p value = 0.007 for HSP27) in DFU
along with their downstream molecules TLR4 and p38-MAPK (p value = 0.006 for p38-MAPK;
p value = 0.02 for TLR4). HSP70 levels were significantly lower in male subjects and their
levels increased significantly with the grades of wound on Wagner’s scale. Infection status of the
wounds was found to be significantly associated with the increased levels of HSP70 and HSP27
in infected diabetic wounds.
Conclusions: Our study demonstrates that the down regulation of HSPs in diabetic wounds is
associated with wound healing impairment in T2DM subjects.
KEY WORDS: Heat shock proteins; Wound healing impairment; type 2 diabetes mellitus
(T2DM); TLR4; MAPK; Diabetic foot ulcer

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
3
Introduction
Heat shock proteins (HSPs) are heterogeneous gene products of a highly conserved family of
stress proteins [1]. Their molecular weights vary roughly from 16 KDa to 110 KDa, and they are
rapidly expressed in cells exposed to a variety of stress [2]. All organisms ranging from
archeabacteria to eubacteria, or from plants to animals, respond to endogenous or exogenous
stress by inducing HSPs [1]. The synthesis and expression of these stress proteins is regulated
mainly by a transcription factor known as heat shock factor-1 (HSF-1) which bind to the heat
shock elements (HSE) present in the promoter region of specific genes. In addition to this
emergency response, they also serve as molecular chaperones in various physiological and
pathological conditions by bringing about the folding of nascent polypeptides and targeting
improperly folded proteins for degradation [3]. These proteins maintain a state of homeostasis
during normal cell growth as well as in pathological condition by maintaining the cellular
integrity [4]. Recent reports have suggested that HSPs after their release in the blood, also
participate in signal transduction [5]. These proteins have anti-inflammatory properties as they
inhibit nitric oxide synthase activity along with NF-κB dependent gene expression [6]. Emerging
evidences also suggest certain members of HSPs participate in both innate and adaptive
immunity [7]. HSPs can modulate cellular adaptive response by modulating CD8+ cytotoxic cell
receptor on one hand and can directly stimulate innate immune response by toll like receptor
(TLR) mediated signaling on the other [5, 8].
Wound healing is a dynamic process which includes 4 main overlapping stages ranging from (i)
acute inflammation through (ii) fibrin rich exudates organization, (iii) re-epithelialization to (iv)
granulation tissue formation [9]. The healing wound bed contains different inducible HSPs like
HSP90, HSP70, HSP47 and HSP27 which all together bring about protein homeostasis and cell

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
4
proliferation during wound healing [10]. HSP90 brings about wound healing by supporting the
differentiation of keratinocytes [11]. HSP70 lowers oxidative damage of fibroblasts and supports
cell proliferation in the wound area by inhibiting stress induced apoptosis and TLR activation
[12]. HSP70 is also a ligand for TLR4, which is shown to be an important contributor to wound
closure [13]. The functioning of HSP70 is aided by expression of HSP47, which is
predominantly involved in pro-collagen synthesis and binding with collagen type II and III [14].
Small HSPs like HSP27 also supports wound healing by stabilizing actin microfilaments,
supporting endothelial cell migration in the wound bed, protecting sensory neuron degeneration
and inhibiting stress induced apoptosis [15]. This function of HSP27 is governed by its MAPK
mediated phosphorylation [16].
Type 2 diabetes mellitus (T2DM) is characterized by chronic hyperglycemia and is a proximal
determinant of secondary complications like neuropathy, tissue ischemia and infection [17]. The
wounds in T2DM patients often turn into non healing micro-environment leading to chronic
ulcers due to less oxygen supply, death and deformity in migrating fibroblasts, abnormal matrix
degradation and oxidative stress [18, 19]. T2DM also regulates the HSP synthesis by modulation
of transcriptional or translational processes [20]. Deficient insulin signaling in diabetic
individuals lead to glycogen synthase kinase 3-β (GSK-3β) mediated inhibitory phosphorylation
and deactivation of HSF1.This result in the low levels of intracellular HSPs (iHSPs) especially in
insulin sensitive tissues like muscles and liver which further interfere with insulin signaling via
activation of inflammatory cytokines, c-Jun N-terminal kinase (JNK) and IkappaB kinase (pIKK-
β) in a cyclic manner [4]. This decreased level of iHSPs create an impaired stress response in
diabetic individuals manifested by glycation, oxidation and aggregation of cellular proteins
which in majority of cases lead to the disruption of homeostatic processes[4]. An array of

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
5
literature supports the deficiency of HSPs especially HSP70 or their transcriptional activators in
diabetic wounds in various model systems [21] but little is known about the role of HSPs in
human diabetic wounds. In the present work we have tried to see the expression levels of HSP27,
HSP47 and HSP70 and their downstream molecules like TLR4, p38 mitogen-activated protein
kinase (p38-MAPK) in biopsies from human diabetic wounds in comparison to non diabetic
wounds.
Materials and methods:
Subjects:
This case control study comprised of 109 subjects in which 101 were Diabetic foot ulcer (DFU)
cases and 8 were controls without having T2DM. All DFU patients included were Diabetics who
had non-healing wounds of > 4 weeks duration, thus qualifying as Diabetic wounds. Majority of
the patients had lower extremity wounds 90% of which were located on the foot alone and in the
remaining 10% foot + lower leg were involved. Both the plantar and dorsal aspects of the foot
were involved in the majority of the cases. The samples were collected at the time of their (the
patients) first visit to the Diabetic foot clinic. Samples were taken from the wound margins
during the debridement process and the histological analysis was performed to determine the cell
types (Sup. Figure1). Classification of wounds was made on the basis of the Wagner’s Grading
System [22]. The presence and absence of infection in the wounds were also recorded. Samples
were collected from the OPD clinics and operation theatres of Department of Endocrinology and
Metabolism and Department of Surgery, Institute of Medical Sciences, Banaras Hindu
University, Varanasi, India during the period of July 2010 to December 2013. Tissue samples
were collected in RNAlater solution (P/N AM7020, Ambion, Inc., Austin, TX, USA) and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
6
phosphate buffer saline (PBS) for RNA and Protein isolation respectively and kept frozen at -
80°C until use. For immunohistochemical staining, samples were collected in Boiun's fixative
solution and kept at room temperature. Patients underwent a standardized clinical and laboratory
evaluation. The T2DM patients having neuropathic, vascular or traumatic ulcers were included in
this study. Screening for neuropathy was done by taking a history of sensory loss and other
symptoms such as a burning sensation or paresthesias. Clinical neurological examination
included the assessment of the vibratory threshold perception using a 128 Hz tuning fork and
assessment of pain and fine touch with a pin and 10g monofilament respectively. The tendon
reflexes and muscle power were measured in patients with sensory neuropathy. Screening for
vascular involvement included a detailed history of vascular insufficiency, clinical examination
for signs of chronic ischemia and assessment of all lower limb pulses. A bed side hand held
Doppler study was carried out in all clinically suspicious cases and ABPI (ankle brachial
pressure index) of < 0.9 was considered indicative of peripheral vascular disease. Age and sex
matched control tissues were obtained by full thickness wound biopsies of post cellulitic chronic
ulcers of the foot and the distal leg. These were non-healing ulcers present for 4 weeks or more
following cellulitis of the lower limb. Each patient’s family history, habits (smoking, alcoholism
etc.), and disease were recorded through a questionnaire (Table 1). The exclusion criteria of the
study included presence of co morbid disorders such as thyroid dysfunction and patients not
belonging to north India. The study was approved by the Institutional Human Ethics Committee
of Institute of Medical Sciences, Banaras Hindu University, Varanasi, India. Informed written
consent was obtained from every participant of each group.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
7
Semi-quantitative RT-PCR
Total RNA was isolated from wounds samples using TRIzol reagent followed by DNase
treatment. cDNA was synthesized and semi-quantitative RT-PCR analysis of HSP70, HSP47
HSP27, p38-MAPK and TLR4 was done in 101 DFU cases and 8 controls. Sequences of the
implicated primers in the study are provided in Table 2. The PCR conditions were initial
denaturation step of 94°C for 5 min followed by 30 cycles of 30 sec at 94°C, 40 sec at 58°C, 40
sec at 72°C and then a final extension step of 10 min at 72°C. Glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) expression level was checked as an internal control to ascertain the
quality of cDNA. Expression of gene transcripts were quantified after normalizing samples using
GAPDH gene.
Quantitative Real-time PCR
A quantitative RT-PCR (RT-qPCR) experiment was also performed to validate the results
obtained by semi-quantitative RT-PCR in 88 DFU samples of different grades on Wagner Scale
and 8 controls. RT-qPCR experiment was performed according to the manufacturer’s protocol
(Applied Biosystem) using primers for HSP27, HSP47, HSP70 and GAPDH. Briefly, 20 μl total
reaction volume containing 10 μl SYBR Green, 0.1 μl each forward and reverse primer (10pm/
μl) and 2 μl cDNA was used in PCR using ABI 7500 instrument. PCR was performed with an
initial incubation at 50°C for 2 min, then followed by 10 min denaturation at 95°C and 40
cycles at 95°C for 15 s, 60°C for 1 min and 72°C for 15 s. Gene expression profiles were
normalized to the mRNA levels of housekeeping gene GAPDH. ΔΔCT and the relative fold
change of HSPs in DFU cases were calculated according to our previous report [23].

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
8
Immunohistochemical staining
Wound tissue samples obtained during the debridement process were fixed in Bouin’s solution,
embedded in paraffin, and sectioned into 3 µm thick sections. 5 DFU wounds and 5 control
wounds were randomly selected for the study. Anti HSP27 antibody (Catalog No. ab5579,
Abcam Inc., Cambridge, MA., 1:75 in PBS), Anti HSP47 antibody (Catalog No. ab88115,
Abcam Inc., Cambridge, MA., 1:75 in PBS) and Anti HSP70 antibody (Catalog No. ab47455,
clone C92F3A-5, Abcam Inc., Cambridge, MA., 1:75 in PBS) were applied separately to the
deparaffinized sections and incubated in a wet chamber at 4°C for 12 hours. Vectastain Elite
ABC Kit (Vector Laboratories, Burlingame, CA) was used for immunohistochemical staining.
Slides were counterstained with Hematoxylin (Himedia, India). Cells having brown-stained
cytoplasm were regarded as positive. Similar staining time and procedure was adopted for all
tissue samples. Expression patterns of HSP27, HSP47 and HSP70 in control and diabetic wounds
were done under the microscope (Nikon) using different magnifications (4, 10, 20 and 40).
Documentation of acquired images was done using a calibrated digital camera system (Nikon
eclipse 80i) together with the software evaluation package (NIS Elements software). The
expression density HSPs in wound biopsies were computed according to a previous study by
Souil et al. [24].
Western blot
Western blot analysis was performed for HSP27, HSP47 and HSP70 on whole-tissue extracts of
wound biopsies to verify the results of IHC. About 50 μg of protein was loaded on 12 % SDS-
PAGE gel, which was transferred to nitrocellulose membrane and then blocked with 5% of skim
milk in TBS. For HSP27, rabbit polyclonal anti-HSP27 antibody (Catalog No. ab5579, Abcam

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
9
Inc., Cambridge, MA., 1:1000 dilution), for HSP47, mouse polyclonal anti-HSP47 antibody
(Catalog No. ab88115, Abcam Inc., Cambridge, MA., 1:1000 dilution) and for HSP70, mouse
monoclonal anti-HSP70 antibody (Catalog No. ab47455, clone C92F3A-5, Abcam Inc.,
Cambridge, MA., 1:1000 dilution) were used and then incubated with the secondary antibody
linked to horseradish peroxidase. The immunoreactive bands were visualized by the Enhanced
Chemiluminescence System (Amersham Biosciences). Blots were stripped off and reprobed with
anti-GAPDH antibody.
Statistical analysis
The data were expressed as mean considering standard error of mean as error bars. Statistical
significance (P < 0.05) was determined by Student’s t test (two-tailed) and nonparametric
ANOVA. Statistical analysis of data was performed using Graph Pad Prism 5.01 and IBM SPSS
Statistics 20.0 software.
Results
Semi-quantitative RT-PCR analysis showed that there was significant down regulation of
HSP70, HSP47 and HSP27 in wounds of DFU patients compared to control cases (p value =
<0.001, t = 5.59, R squared = 0.22 for HSP70; p value = 0.007, t = 2.77, R squared = 0.06 for
HSP47; p value = 0.007, t = 2.72, R squared = 0.06 for HSP27) (Figure 1, Sup. Figure 2). This
down-regulation of analyzed HSPs message was again confirmed by RT-qPCR analysis (p-value
< 0.001, mean fold change = 2.47 ± 0.15, t = 4.73 for HSP70; p-value < 0.001, mean fold change
= 2.04 ± 0.14, t = 4.17 for HSP47; p-value = 0.03, mean fold change = 2.79 ± 0.15, t = 2.10 for
HSP27) (Figure 1.b). The mRNA level of HSP70 was higher in females as compared to males
(p-value = 0.017, t = 2.42, R squared = 0.06) while the expressions of HSP47 and HSP27

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
10
message were similar in both genders (p-value = 0.63, t = 0.47, R squared = 0.002 for HSP47
and p-value = 0.81, t = 0.23, R squared = 0.001 for HSP27) (Figure 2). HSP70 and HSP27 were
found to be significantly higher in the infected diabetic wounds compared to controls (p-value =
0.012, t = 0.2.55, R squared = 0.06 for HSP70 and p-value = 0.03, t = 2.17, R squared = 0.045
for HSP27) while HSP47 levels remained unaltered even in the presence of infection (p-value =
0.70, t = 0.38, R squared = 0.001 for HSP47) (Figure 3). HSP70 level was found to increased
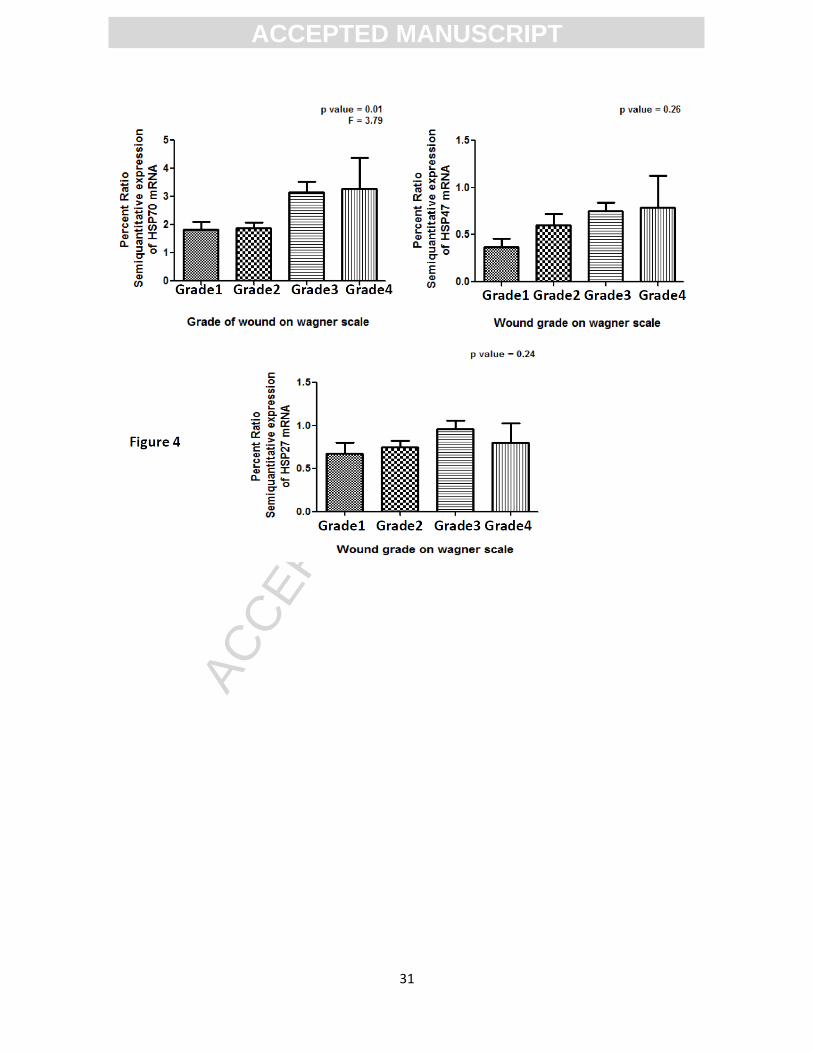
significantly with the severity of diabetic wounds on Wagner’s scale (p-value = 0.01, t = 3.79, R
squared = 0.105) while other two HSPs levels remained unaltered with the severity of diabetic
wounds (p-value = 0.26, t = 1.35, R squared = 0.041 for HSP47 and p-value = 0.24, t = 1.42, R
squared = 0.04 for HSP27) (Figure 4, Sup. Figure 3). Analysis of mRNA downstream molecules
to HSP27 and HSP70 were also done. Expression of p38-MAPK suggested its similar
expression patterns as HSP27 (Figure 5 a, 5 b, Sup. Figure 4) and TLR4 expression were similar
to the patterns of HSP70 (p value = 0.02, t = 2.30, R squared = 0.05 for TLR4; p value = <0.001,
t = 5.59, R squared = 0.22 for HSP70) (Figure 6 a, 6 b, 6 c, Sup. Figure 5). Western blot analysis
showed that these HSPs were significantly down regulated at translational levels also (p value =
0.001, t = 3.72, R squared = 0.42 for HSP70; p value = <0.0001, t = 5.88, R squared = 0.65 for
HSP47; p value = 0.01, t = 2.77, R squared = 0.31 for HSP27) (Figure 7). Figure 8.A shows the
positive and negative (no primary antibody) staining patterns of different antibodies of HSPs
used in IHC study. Absence of any signal in negative controls suggested that the signal in
positive control was specific signal of primary antibody binding to the antigens.
Immunohistochemical expression analysis between groups also suggested significant down
regulation of HSP70, HSP47 and HSP27 between the wound biopsies of DFU cases and controls

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
11
(Figure 8.B, 8.C, 8.D). The down regulation of TLR4 in DFU cases compared to control wounds
was in a similar pattern to HSP70 (Figure 9, Sup. Figure 6)
Conclusion
The cells of all organisms generally counteract sub lethal endogenous or exogenous stresses by
activating transcription of a specific set of genes termed as HSPs. The stresses may be
physiological (e.g., inflammation, ischemia), pathological (e.g., bacterial or viral infection) or
environmental (e.g. heat shock, oxidative stress or heavy metal poisoning) [25]. Some HSPs are
normally expressed in cells regulated by hormones like estrogens while others are produced
exclusively during stress periods by certain cells [26]. A cell dependent unique pattern of
induction and repression of other genes is followed after expression of HSPs. These HSPs are
highly conserved in amino acid sequences and contain a specific DNA motif called heat shock
element (HSE) [27]. These HSEs when occupied by HSFs bring about the transcription of HSPs.
HSPs are also shown to be protective against insulin resistance and obesity induced
inflammation, thereby abrogating T2DM [28]. Some members of HSPs like HSP90 and HSP70
are also shown to promote wound healing by promoting recruitment of dermal fibroblasts and
endothelial cells to the site of injury, thereby, supporting re-epithelialization in model animals
[29]. Wounds of T2DM patients are hard to heal and inadequate amount of inducible HSPs may
be one of the important contributors in the etiology of DFU [30]. The present work was designed
to correlate the expression levels of three HSPs namely HSP70, HSP47 and HSP27 along with
their downstream interacting partners like p38-MAPK and TLR4 with wound healing
impairment in human T2DM subjects. Several contributing factors like gender of the subjects,
infection status of the wounds and severity of wounds on Wagner’s scale were also taken into
account.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
12
Immediate release of anti-apoptotic and anti-oxidative HSP70 by keratinocytes prevailing in the
epidermis is a prerequisite of an acute wound [31]. This HSP70 after secretion in the wound
microenvironment bring about the regulation of the inflammatory phase by coordinating pro and
anti-inflammatory responses [32]. Appropriate levels of HSP70 in wounds often lead to abundant
granulation tissue synthesis and proper healing. The analysis of our data indicated that there was
significant down regulation of HSP70 in human diabetic wounds compared to non diabetic
wounds. Our result is supported by the findings of Oberringer et al. (1995), according to which
non-diabetic wounds lacking adequate HSP70 proteins may often develop into chronic decubitus
ulcers [33]. Another report by Vígh et al. (1997) suggested that the application of Bimoclomol, a
strong inducer of HSP70, significantly enhance wound closure in thermally wounded STZ
diabetic rats compared to controls [34]. Another finding that external HSP70 inductions in
chronic wounds of streptozotocin-induced diabetic mice bring about the rapid closure of wounds
also supported our hypothesis that decreased levels of HSP70 is one of the causes of DFU in
humans [35]. Lack of proper activation of innate immunity by TLR4 mediated signaling is also
one of the causes of developing DFU [23, 36, 37]. HSP70 being a ligand for TLR4 is also
required in proper amount in healing wounds [13]. Hence we simultaneously analyzed the
expression levels of HSP70 and TLR4 in DFU subjects and the results obtained indicated that
less expression of HSP70 and TLR4 combined together bring about compromised wound healing
in them. Levels of HSP70 and TLR4 both showed a gender dependency as female subjects had
comparatively higher amount of HSP70 and TLR4 compared to their male partners. Regulation
of HSP70 and TLR4 via estrogen is the reason behind this finding and it supports the higher
incidence of DFU in males compared to females [38, 39]. The levels of HSP70 along with TLR4
were found to be dependent on the grades of wounds and severe wounds were associated with

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
13
higher levels of HSP70 compared to normal wounds. The levels of HSP70 were also found to be
significantly associated with the infection status of the diabetic wounds with higher levels of
HSP70 transcripts in infected diabetic wound compared to non infected wounds. The reason for
this observation may be the finding that microbial infections bring about an increase in
expression of HSP70 in infected cells [40].
HSP27 plays an important role in modulating actin dynamics in response to various stimuli.
HSP27 participates in the wound healing process by regulating fibroblasts and endothelial cell
migration, adhesion and invasion in the wound microenvironment. The expression and spatial
distribution of HSP27 is shown to regulate normal wound healing [41]. This effect of HSP27 is
mainly orchestrated by p38MAPK mediated phosphorylation. HSP27 is only known substrate of
certain mitogen activated protein kinase-activated protein kinase (MAPKAPK) 2/3 [42, 43].
HSP27 is also a protective agent against diabetic neuropathy and hence diminished expression of
HSP27 may also compromise healing in diabetic neuropathic ulcers [44]. The findings of present
work suggested a significant down regulation of HSP27 both at the transcriptional and
translational levels in biopsies of DFU cases compared to controls. The p38-MAPK expression at
transcription levels also followed the similar trend in DFU subjects. This finding of ours was in
coordination with a recent report by Crowe et al. (2013) which suggests that mice deficient in
orthologue of HSP27 display delayed wound healing by lesser collagen deposition and prolonged
inflammatory stage [45]. HSP27 levels like HSP70 was also dependent upon the infection status
of diabetic wounds. This finding is supported by the observation of Wainberg et al. that
cumulative expression of HSP70 and HSP27 by CD4+ lymphocytic cells shoot up following
acute infection of DNA and RNA mediated viruses [46]. The expressional levels of HSP27

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
14
transcripts were found not be associated with grades of wounds and the gender of the DFU
subjects.
HSP47 is also called collagen binding protein 1 and is a molecular chaperone specific for
procollagen (47). Being localized to endoplasmic reticulum, the function of HSP47 is to bring
about the maturation of collagen by mediating post-transcriptional modification in procollagen
[48]. Wang et al. (2002) shown that HSP47 is inducible in wounds and is one of the positive
contributors in healing fetal wounds [49]. Our study showed that HSP47 expression was
significantly lower in diabetic wounds compared to control wounds. This down-regulation of
HSP47 in diabetic wounds may abrogate wound healing by inappropriate collagen synthesis and
less differentiation of fibroblasts into myofibroblasts [50]. This finding of ours is supported by
the report of Wang et al. (2009) which supports the wound healing enhancer capability of HSP47
in alloxan-induced diabetes rats [51]. Unlike HSP70 and HSP27, HSP47 transcripts did not show
any gender, wound grade or wound infection dependency. One limitation of the present study
was the less number of control samples compared to DFU cases, reason being unwillingness of
controls to provide tissue biopsy due to pain and fear associated with the process.
In conclusion, our study demonstrates the combined down regulation of HSP70, HSP27 and
HSP47 expression in diabetic wounds creates a sort of hostile microenvironment in the wound
bed and abrogates wound healing in T2DM subjects by less collagen availability, improper
migration of wound healing fibroblasts, sensory neuropathy promotion and persistent
inflammation. Further research is required to see the effect of activators of HSPs on healing
patterns of diabetic wounds in vitro for effective therapeutic intervention in this respect.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
15
Conflict of interest
The authors declare no conflict of interest.
Funding
This work was funded by Department of Science and Technology, New Delhi, India (SR/FT/LS-
101/2010). Financial assistance by Department of Biotechnology, Ministry of Science and
Technology, New Delhi, India in form of Senior Research fellowship to the first author is
thankfully acknowledged. We thank Nilu Prasad and Shanti Besra, Laboratory Superintendents,
Indian Railway Cancer Hospital and Research Centre, N.E.R., Varanasi, for their technical
assistance during IHC work.
Author contribution
Kanhaiya Singh designed research, performed experiments, collected and analyzed the data,
wrote the paper. Kiran Singh designed research, interpreted data, and wrote the paper. N.K.A.,
S.K.G., G.M. and S.C. provided samples and did the clinical evaluation of patients. Their critical
comments helped us in writing and approving the final manuscript. Dr. Kiran Singh is the
guarantor of this work, had full access to all the data, and takes full responsibility for the
integrity of data and the accuracy of data analysis.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
16
References
1. Lindquist S and Craig EA. The Heat-Shock Proteins. Annual Review of Genetics. 1988;
22: 631-677.
2. Kampinga HH, Hageman J, Vos MJ et al. Guidelines for the nomenclature of the human
heat shock proteins Cell Stress and Chaperones. 2009;14(1): 105-111.
3. Morimoto RI. Regulation of the heat shock transcriptional response: cross talk between a
family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev.
1998; 12(24): 3788-96.
4. Hooper PL, Balogh G, Rivas E, Kavanagh K, Vigh L. The importance of the cellular
stress response in the pathogenesis and treatment of type 2 diabetes. Cell Stress
Chaperones. 2014;19(4):447-64.
5. Calderwood SK, Mambula SS, Gray P.J. Jr. et al. Extracellular heat shock proteins in cell
signaling and immunity. Ann N Y Acad Sci. 2007; 1113: 28-39.
6. Wong HR. Potential protective role of the heat shock response in sepsis. New Horiz.
1998; 6(2): 194-200.
7. Javid B, MacAry PA, Lehner PJ. Structure and function: heat shock proteins and adaptive
immunity. J Immunol. 2007; 179(4): 2035-40.
8. Fang H, Wu Y, Huang X et al. Toll-like receptor 4 (TLR4) is essential for Hsp70-like
protein 1 (HSP70L1) to activate dendritic cells and induce Th1 response. J Biol Chem.
2011; 286(35): 30393-400.
9. Singh K, Agrawal NK, Gupta SK, Singh K. A functional SNP-1562C>T in the matrix
metalloproteinases-9 promoter is associated with type 2 diabetes and diabetic foot ulcers.
Int J Lower Extrem Wounds. 2013; 12(3): 199 –204.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
17
10. Laplante AF, Moulin V, Auger FA. Expression of Heat Shock Proteins in mouse skin
during wound healing. J Histochem Cytochem. 1998; 46: 1291.
11. Li W, Sahu D, Tsen F et al. Secreted heat shock protein-90 (Hsp90) in wound healing
and cancer. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 2012;
1823(3): 730–741.
12. Asea A, Rehli M, Kabingu E et al. Novel signal transduction pathway utilized by
extracellular HSP70: role of toll-like receptor (TLR) 2 and TLR4. J Biol Chem. 2002;
277: 15028-15034.
13. Luong et al. Stimulation of TLR4 by recombinant HSP70 requires structural integrity of
the HSP70 protein itself. Journal of Inflammation. 2012; 9: 11.
14. Kurkinen M, Taylor A, Garrels JI et al. Cell surface-associated proteins which bind
native type IV collagen or gelatin. J Bio Chem. 1984; 259: 5915–5922.
15. Hirano S, Shelden EA, and Gilmont RR. HSP27 regulates fibroblast adhesion, motility,
and matrix contraction. Cell Stress & Chaperones. 2004; 9(1): 29–37.
16. Rouse J, Cohen P, Trigon S et al. A novel kinase cascade triggered by stress and heat
shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock
proteins. Cell. 1994; 78: 1027–1037.
17. Fowler MJ. Microvascular and macrovascular complications of diabetes. Clin Diab.
2008; 26: 77-82.
18. Singh K, Singh VK, Agrawal NK, Gupta SK, Singh K. Association of toll-like receptor 4
polymorphisms with diabetic foot ulcers and application of artificial neural network in
DFU risk assessment in type 2 diabetes patients. Biomed Res Int. 2013; 2013: 318686.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
18
19. Singh K, Agrawal NK, Gupta SK, Singh K. Association of Variant rs7903146 (C/T)
Single nucleotide polymorphism of TCF7L2 gene with impairment in wound healing
among north Indian Type 2 Diabetes population, A Case–Control Study. The
International Journal of Lower Extremity Wounds. 2013; 12(4): 310-315.
20. McMurtry L, Cho K,Young LJT et al. Expression of HSP70 in Healing Wounds of
Diabetic and Nondiabetic Mice. Journal of Surgical Research. 1999; 86: 36–41.
21. Hooper PL and Hooper JJ. Loss of defense against stress, Diabetes and Heat Shock
Proteins. Diabetes technology & therapeutics. 2005; 7(1): 204-208.
22. Wagner FW. The dysvascular foot, a system for diagnosis and treatment. Foot Ankle.
1981; 2: 64-122.
23. Kanhaiya, Agrawal NK, Gupta SK, Singh K. Differential expression of Toll like
Receptor 4 in Type 2 Diabetic patients with impaired wound healing. Journal of Diabetes
and Metabolism. 2013; 4: 260. Doi:10.4172/2155-6156.1000260.
24. Souil E, Capon A et al. Treatment with 815-nm diode laser induces long-lasting
expression of 72-kDa heat shock protein in normal rat skin. Br. J. Dermatol. 2001;
144(2): 260–266.
25. Schlesinger MJ. Heat shock proteins. J Biol Chem. 1990; 265: 12111-12114.
26. Silva JAD. Heat shock proteins, the missing link between hormonal and reproductive
factors and rheumatoid arthritis. Annals of the Rheumatic Diseases. 1991; 50: 735-739.
27. Carper SW, Duffy JJ, Gerner EW. Heat shock proteins in thermotolerance and other
cellular processes. Cancer Res. 1987; 47: 5249-5255.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
19
28. Henstridge DC, Forbes JM, Penfold SA et al. The relationship between heat shock
protein 72 expression in skeletal muscle and insulin sensitivity is dependent on adiposity.
Metabolism. 2010; 59(11): 1556–1561.
29. Wagstaff MJ, Shah M, McGrouther DA. The heat shock proteins and plastic surgery. J
Plast Reconstr Aesthet Surg. 2007; 60(9): 974-82.
30. Atalay M, Oksala N, Lappalainen J et al. Heat shock proteins in diabetes and wound
healing. Curr Protein Pept Sci. 2009; 10(1): 85–95.
31. Shukla A, Dubey MP, Srivastava R et al. Differential expression of proteins during
healing of cutaneous wounds in experimental normal and chronic models. Biochem
Biophys Res Commun. 1998; 244(2): 434-9.
32. Klosterhalfen B, Klinge U, Tietze L et al. Expression of heat shock protein 70 (HSP70) at
the interface of polymer-implants in vivo. J Mater Sci Mater Med. 2000; 11(3): 175-81.
33. Oberringer M, Baum HP, Jung V et al. Differential expression of heat shock protein 70 in
well healing and chronic human wound tissue. Biochem Biophys Res Commun. 1995;
214(3): 1009-14.
34. Vígh L, Literáti PN, Horváth I, Török Z, Balogh G, Glatz A et al. Bimoclomol: a
nontoxic, hydroxylamine de- rivative with stress protein-inducing activity and
cytoprotective effects. Nat Med 1997; 3:1150-115
35. Bitar MS, Farook T, John B et al. Heat-shock protein 72/73 and impaired wound healing
in diabetic and hypercortisolemic states. Surgery. 1999; 125(6): 594-601.
36. Chen L, Guo S, Ranzer MJ, DiPietro LA. Toll-like receptor 4 has an essential role in
early skin wound healing. J Invest Dermatol. 2013; 133: 258-267.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
20
37. Singh K, Singh VK, Agrawal NK, Gupta SK, Singh K. Genetic alterations in Toll-Like
Receptor 4 signaling pathway and impairment of wound healing in patients with Type 2
Diabetes. Int J Lower Extrem Wounds. 2014; 13(2):162-163.
38. Rettew JA, Huet YM, Marriott I. Estrogens augment cell surface TLR4 expression on
murine macrophages and regulate sepsis susceptibility in vivo. Endocrinology. 2009;
150: 3877-3884.
39. Voss MR, Stallone JN, Li M et al. Gender differences in the expression of heat shock
proteins, the effect of estrogen. Am J Physiol Heart Circ Physiol. 2003; 285: H687–
H692.
40. Ohgitani E, Kobayashi K, Takeshita K et al. Biphasic translocation of a 70 kDa heat
shock protein in human cytomegalovirus-infected cells. Journal of General Virology.
1999; 80: 63–68.
41. Hirano S, Shelden EA, Gilmont RR. HSP27 regulates fibroblast adhesion, motility, and
matrix contraction. Cell Stress Chaperones. 2004; 9(1): 29-37.
42. Hirano S, Rees RS, Gilmont RR et al. MAP kinase pathways involving hsp27 regulate
fibroblast-mediated wound contraction. J Surg Res. 2002; 102(2): 77-84.
43. Rouse J, Cohen P, Trigon S et al. Novel kinase cascade triggered by stress and heat shock
that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins.
Cell. 1994; 78: 1027.
44. Korngut L, Ma CHE, Martinez JA et al. Overexpression of human HSP27 protects
sensory neurons from diabetes. Neurobiology of Disease. 2012; 47: 436–443.
45. Crowe J, Aubareda A, McNamee K et al. Heat Shock Protein B1-Deficient Mice Display
Impaired Wound Healing. PLoS One. 2013; 8(10): e77383.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
21
46. Wainberg Z, Oliveira M, Lerner S et al. Modulation of Stress Protein (hsp27 and hsp70)
Expression in CD4+ Lymphocytic Cells Following Acute Infection with Human
Immunodeficiency Virus Type-1. Virology. 1997; 233(2): 364–373.
47. Lamande SR, Bateman JF et al. Procollagen folding and assembly, the role of
endoplasmic reticulum enzymes and molecular chaperones. Semin Cell Dev Biol. 1999;
10: 455–464.
48. Dafforn TR, Della M, Miller AD. The molecular interactions of heat shock protein 47
(Hsp47) and their implications for collagen biosynthesis. J. Biol. Chem. 2001; 276(52):
49310e9.
49. Wang ZL, Inokuchi T, Ikeda H et al. Collagen-binding heat shock protein HSP47
expression during healing of fetal skin wounds. Int. J. Oral Maxillofac. Surg. 2002; 31:
179–184.
50. Hong S, Park K, Kim JH et al. Role of Heat Shock Protein 47 in transdifferentiation of
human tenon's fibroblasts to myofibroblasts. BMC Ophthalmology. 2012; 12: 49.
51. Wang Z, Li L et al. The plasmid encoding HSP47 enhances collagen expression and
promotes skin wound healing in an alloxan-induced diabetic model. Cell Biology
International. 2009; 33: 705-710.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
22
Figure Legends:
Figure.1 RT-PCR analysis indicates the decreased expression of HSP70, HSP47and
HSP27 transcripts in DFU patients. RNA was isolated from wound samples of DFU
patients and Controls, cDNA was synthesized and RT-PCR for HSP70, HSP47, HSP27
and GAPDH was performed.
(A) Bar Graph represents the percent ratio which was calculated for the expression of
HSPs and GAPDH in Controls and DFU respectively. Expression of HSP70, HSP47 and
HSP27 transcripts were found to be downregulated in DFU cases as compared to controls
(p value = <0.0001, t = 5.59, R squared = 0.22 for HSP70; p value = 0.007, t = 2.77, R
squared = 0.07 for HSP47; p value = 0.007, t = 2.72, R squared = 0.06 for HSP27.
(B) Bar Graph of qPCR analysis showing the lower expression of HSPs mRNA in the
wounds of DFU patients. Analysis was done in 88 DFU cases and 8 controls. Fold
change in the expression of genes was determined using the ΔΔCT method of relative
quantification. The graph was plotted using log (Relative average fold change) i.e. (log 2-
ΔΔCT). The graph clearly showed that HSP70, HSP47 and HSP27 were down regulated
significantly in the wounds of T2DM cases compared to controls (p-value < 0.001, mean
log (fold change) = - 2.47 ± 0.15, t = 4.73 for HSP70; p-value < 0.001, mean (log fold)
change = - 2.04 ± 0.14, t = 4.17 for HSP47; p-value = 0.03, mean log (fold change) = -
2.79 ± 0.15, t = 2.10 for HSP27). Log (10) = 1; log (100) = 2.
Figure2. Bar Graph showing the correlation of HSP70, 47 and 27 transcripts with
the Gender of DFU patients. Among the diabetic wounds, males showed relatively
lesser amount of HSP70 mRNA transcripts with respect to females (p-value = 0.017, t =
2.42, R squared = 0.06) while the expressions of HSP47 and HSP27 message were
similar in both genders (p-value = 0.63, t = 0.47, R squared = 0.002 for HSP47 and p-
value = 0.81, t = 0.23, R squared = 0.001 for HSP27).
Figure3. Bar Graph showing the correlation of HSP70, 47 and 27 transcripts with
the infection status of DFU wounds. HSP70 and HSP27 were found to be significantly
associated to the infection status of the diabetic wounds. Infected wounds contained
higher levels of HSP70 and HSP27 compared to sterile wounds (p-value = 0.012, t =

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
23
0.2.55, R squared = 0.06 for HSP70 and p-value = 0.03, t = 2.17, R squared = 0.045 for
HSP27). HSP47 levels remained unaltered even in the presence of infection (p-value =
0.70, t = 0.38, R squared = 0.001 for HSP47).
Figure4. Bar Graph showing the comparison of HSP70, 47 and 27 transcripts with
the wound grades on Wagner’s scale. HSP70 message was significantly dependent on
the grade of wounds on Wagner’s scale and its level were high in more severe wounds
compared to less severe wounds (p-value = 0.01, t = 3.79, R squared = 0.105). HSP27
and HSP47 levels remained unaltered with the severity of diabetic wounds (p-value =
0.26, t = 1.35, R squared = 0.041 for HSP47 and p-value = 0.24, t = 1.42, R squared =
0.04 for HSP27).
Figure5. Bar graph showing p38-MAPK having similar expression patterns as
HSP27. (A) Expression of p38-MAPK and HSP27 transcripts were found to be down
regulated in DFU cases as compared to controls (p value = 0.006, t = 2.79, R squared =
0.07 for p38-MAPK; p value = 0.007, t = 2.72, R squared = 0.06 for HSP27). (B) p38-
MAPK message was significantly dependent on the grade of wounds on Wagner’s scale
(p-value = 0.02, t = 3.34, R squared = 0.11) and shared a similar pattern followed by
HSP27 transcripts in DFU wounds.
Figure6. Bar graph showing TLR4 having similar expression patterns as HSP70. (A)
Expression of TLR4 and HSP70 transcripts were found to be significantly down
regulated in DFU cases as compared to controls (p value = 0.02, t = 2.30, R squared =
0.05 for TLR4; p value = <0.001, t = 5.59, R squared = 0.22 for HSP70). (B) Both
HSP70 and TLR4 message were found to be sharing a similar patterns dependent upon
grade of wounds on Wagner’s scale. (C) The mRNA levels of both HSP70 and TLR4
were significantly higher in females as compared to males (p-value = 0.017, t = 2.42, R
squared = 0.06 for HSP70; p value = 0.02, t = 2.3, R squared = 0.06).
Figure7. HSPs protein expression was analyzed in 15 DFU patients and 6 controls.
Tissue samples collected from control {C} and DFU patients {P} were homogenized and
Western blot analysis was performed for expression of HSP70, HSP47, HSP27 and

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
24
GAPDH protein. Bar Graph showing down regulation of HSPs in wounds of DFU
patients compared to controls (p value = 0.001, t = 3.72, R squared = 0.42 for HSP70; p
value = <0.0001, t = 5.88, R squared = 0.65 for HSP47; p value = 0.01, t = 2.77, R
squared = 0.31 for HSP27).
Figure8. Immunohistochemistry of HSP70, 47 and 27 in wound samples with insets
showing detail of staining (40 X magnifications). Microwave-induced antigen retrieval
used 0.01 M citrate buffer (pH 6). Figure 8.A shows the positive and negative (no
primary antibody) staining patterns of different antibodies of HSPs used in IHC study.
Absence of any signal in negative controls suggested that the signal in positive control
was specific signal of primary antibody binding to the antigens. Immunohistochemical
expression analysis among groups also suggested significant down regulation of HSP70,
HSP47 and HSP27 between the wound biopsies of DFU cases and controls (Figure 8.B,
8.C, 8.D).
Figure9. Immunohistochemistry of TLR4 in wound samples with insets showing
detail of staining (40 X magnifications) with Mouse monoclonal anti-TLR4 antibody.
Upper panel shows the negative (no primary) (left) and positive staining (right). Lower
panel shows the Immunohistochemistry for TLR4 in non-diabetic control wound (left)
and diabetic wound (right). TLR4 was found to be down regulated in DFU cases
compared to control wounds in a similar pattern to HSP70.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
25
Table 1 Biochemical and Demographic parameters of DFU patients (N = 101) and
controls (N = 8). Data are presented as mean ± SD or as number (percentage).
PARAMETERS DFU (N = 101) Control (N = 8) p-value
Age in years; mean ± SD 54.22 ± 8.94 years 56.27 ± 3.42 0.61
BMI in kg/m2; mean ± SD 21.69 ± 2.36 Kg/m2
23.45 ± 1.95 0.15
Duration of T2DM in years; mean ± SD 10.18 ± 4.46 years N/A --
Male 69 (68.32 %) 5 (62.5%) 0.88
Female 32 (31.68 %) 3 (37.5 %) 0.81
HbA1c levels (%) (Mean, range) 10.6 (8.7 to 13) % N/A --
Family history present; n (%) 13 (12.87 %) N/A --
Nephropathy present (Serum creatinine > 1.4 mg/dl); n (%) 30 (29.70%) N/A --
Neuropathy present (by monofilament test); n (%) 61(60.39 %) N/A -
Hypertension present (systolic BP > 140 mm of Hg); n (%) 35(34.65 %) N/A --
Retinopathy present; n (%) 11 (10.89%) N/A --
Dislipidimea present (Serum cholesterol and Tgy levels > 200
mg/dl); n (%)
15(14.85 %) N/A --
Infection present (Wound culture positive for microbes); n (%) 56(55.44 %) N/A --
Bone involvement (Osteomyelitis); n (%) 36 (35.64 %) N/A --

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
26
Table 2: Primers used for RT-PCR analysis of different genes studied and base pair products of
the amplicons.
GENE Forward Primer Reverse Primer Amplicon size
(bp)
HSP70 ACCAAGCAGACGCAGATCTTC CGCCCTCGTACACCTGGAT 73
HSP27 TCCCTGGATGTCAACCACTTCG GGGACAGGGAGGAGGAAACTTG 184
HSP47 CGCCATGTTCTTCAAGCCA CATGAAGCCACGGTTGTCC 70
p38-MAPK ATGCCGAAGATGAACTTTGC TCTTATCTGAGTCCAATACAAGCATC 94
TLR4 CAGAGTTTCCTGCAATGGATCA GCTTATCTGAAGGTGTTGCACAT 85

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
27

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
28

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
29

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
30
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
31

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
32

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
33

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
34

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
35

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
36

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
37

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
38

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
39