中本, 雅俊 - osaka prefecture university...
TRANSCRIPT

http://repository.osakafu-u.ac.jp/dspace/
Title Studies on essential roles of C24-ethylsterol in plant development
Author(s) 中本, 雅俊
Editor(s)
Citation
Issue Date 2016
URL http://hdl.handle.net/10466/15130
Rights

Studies on essential roles of C24-ethylsterol in plant development
(植物ステロールが担う未知生理機能の解明)
Masatoshi Nakamoto
2016

1
Contents
Introduction ………. 2
Chapter1 ………. 11
Establishment and characterization of an Arabidopsis C24-ethylsterol deficient
mutant line
Chapter2 ………. 43
Clarification of the disturbed polar localization of auxin efflux carrier protein
Chapter3 ………. 53
General discussion
Supplementary Information ………. 60
Materials and Methods
References ………. 72
Acknowledgements ………. 82

2
Introduction
Sterols are isoprenoid-derived lipids that have diverse and essential functions in all
eukaryotes. Sterols are integral components of the membrane lipid bilayer, where they
regulate membrane permeability and fluidity in conjunction with phospholipids. In
animals, cholesterol alone serves this structural function, whereas plant membranes
consist of several sterols. Plants and algae present distinct sterol biosynthetic features
compared to metazoan as they produce multiple pathway end-products. In addition to
relatively low amounts of cholesterol, their sterol profiles display several
24-alkyl-5-sterols as major products. This chemical diversity of the sterol-reinforcing
component of higher plant cellular membranes is considered the result of an
evolutionary process leading to optimal membrane stability in sessile organisms (Nes
and McKean, 1977; Ramgopal and Bloch, 1983; Dufourc, 2008). The question of that
chemical diversity has driven research on plant sterol biology. Recently, the power of
Arabidopsis thaliana genetics has clearly shown that breakdown of sterol homeostasis
led to severe damage throughout the plant life cycle, embryogenesis, cell elongation,
cell wall formation, vascular differentiation, and hormone signaling (Diener et al., 2000;
Carland et al., 2002; Schrick et al., 2004; Souter et al., 2004; Men et al., 2008).
Biosynthetic defects results in a lack of major membrane sterols that accompany the
accumulation of unusual sterols, a condition reminiscent of sterolosis syndromes well
described in humans (Kelley and Herman, 2001). Plant sterol profile disruption
furthermore leads to a dysfunctional brassinosteroid (BR) biosynthesis that in turn
causes dwarfisms (Schaller, 2004).
It is widely accepted that levels of 24-alkyl-5-sterol (phytosterols) must be

3
appropriately controlled for normal plant development (Schaeffer et al., 2001). This is
determined primarily by the ratio of C24-methylsterols (campesterol) to C24-ethylsterols
(-sitosterol, stigmasterol). This ratio is tuned by the sterol side chain C24-alkylation
reactions that add two exocyclic carbon atoms provided by S-adenosyl methionine to
sterol precursors and are catalyzed by two distinct sterol methyltransferases (SMTs)
(Fig. 1). The first-type SMT (SMT1) is conserved across kingdoms and is responsible for
the C24-methylation of cycloartenol (or for example, of lanosterol in fungi), and the
second-type SMT (SMT2) is specific for Charophycean green algae and plants, and is
responsible for the C28-methylation (or C241-methylation) of 24-methylene lophenol
(Fig. 2). Thus, SMT2 differentiate C24-ethylsterols from C24-methylsterols by adding
one extra methyl group to the side-chain.
Mutations in genes that act upstream of 24-methylene lophenol, the substrate of
SMT2 and SMT3 (Fig. 1), completely impair the production of major phytosterols
compatible with survival (Clouse, 2002). Such Arabidopsis mutants include the
cph/smt1/orc mutants defective in SMT1 (the first methyltransferase step producing
24-methylene-cycloartanol from cycloartenol) (Fig. 1), that exhibit incomplete cell wall
structures, abnormal embryonic development, low fertility rates, irregular vascular
system development, and aberrant membrane localization of the PIN1/PIN3 auxin efflux
transporter proteins (Diener et al., 2000; Lindsey et al., 2003; Willemsen et al., 2003;
Schrick et al., 2004). The Arabidopsis cpi1-1 mutant is defective in the cyclopropylsterol
isomerase gene, resulting in lack of major sterols and accumulation of atypical sterols.
cpi1-1 plants display disturbed root development, unsuccessful endocytosis, loss of
PIN2 polar localization and abnormal cytokinesis (Men et al., 2008). Other Arabidopsis
mutants such as fackel/hydra2 harboring mutations in the C14-reductase (Jang et al.,

4
2000; Schrick et al., 2002), and hydra1 (Souter et al., 2002), which defects in the
8,7-isomerase, exhibit defects in root development, cell wall structure, embryonic and
seedling cell patterning, and ethylene signaling (Souter et al., 2004). In general, these
sterol mutants are thought to be defective in the generation of the auxin concentration
gradients, which are primarily established through the cell-to-cell directional transport by
auxin carrier proteins (Dhonukshe et al., 2008; Petrásek and Friml, 2009; Feraru and
Friml, 2008). The sterol involvement in cell division has also been suggested from the
disrupted cytokinesis in smt1 (Schrick et al., 2002; Schrick et al., 2004) and cpi1-1 (Men
et al., 2008) mutants. The defective vascular patterning in the shoots of hydra1 and
fackel/hydra2 mutant seedlings is associated with ectopic cell divisions (Pullen et al.,
2010), whereas the cell division during early stages of lateral root initiation in the hydra1
and fackel/hydra2 mutants is not affected (Souter et al., 2004). In addition to these
mutations in the post-squalene biosynthetic segment leading to 24-methylene lophenol,
a 4-methyl sterol intermediate substrate of land plants SMTs (Fig. 1), the loss of function
of ERG28 in Arabidopsis induces the accumulation of 4-carboxy-4-methyl-24-methylene
cycloartanol (CMMC), a sterol biosynthetic intermediate yielded from
24-methylene-cycloartanol during the demethylation reaction at C4 (Fig. 1). The
accumulated 4-carboxy-4-methyl sterol derivative, otherwise undetectable in plant
tissues when ERG28 tethers the C4 demethylation enzymatic complex, interferes with
polar auxin transport via its binding to ABC transporters and this result in serious growth
disorder (Mialoundama et al., 2013). Taken together, these results indicate that
phytosterols play critical roles in a variety of molecular events involved in plant
development throughout the life cycle. However, little is known about the mechanism by
which the sterol compositions affect the polar recycling of PIN proteins, cell division, and

5
development. Most importantly, critical roles of individual sterols could not be clarified
from the characterization of a series of Arabidopsis biosynthetic mutants described
above.
Conversely, mutations of the multi-cellular plant specific sterol methyltransferase
genes (SMT2 and SMT3 in Arabidopsis, Fig. 1) affect the C24-methylsterol to
C24-ethylsterols ratio, and so modify specifically the pathway end-product profile. An
Arabidopsis SMT2 mutant (cvp1) displayed post-embryonic defects in vascular
patterning (Carland et al., 2002). In addition, SMT2 co-suppression lines harbored
higher levels of 24-methylcholesterol (campesterol and its epimer
22(23)-dihydrobrassicasterol) and lower levels of -sitosterol and exhibit reduced apical
dominance and fertility in brassinosteroid-independent manner (Schaeffer et al., 2001).
A double mutant line (cvp1 smt3) generated by crossing a cvp1-3 (a null mutation of
SMT2) mutant and a T-DNA mutant of SMT3 (SALK_085292) displayed severe growth
abnormalities affecting processes such as floral organ development and the auxin
response (Carland et al., 2010). The sterol profile of the cvp1 smt3 mutant almost
completely lacks the C24-ethylsterols instead of increased levels of C24-methylsterols,
without influencing the total sterol content and BR biosynthesis (Carland et al., 2010). In
Arabidopsis, the role of (24R)-24-methylcholesterol (campesterol) as a precursor of
brassinosteroids was demonstrated (Fujioka and Yokota, 2003). However, essential
roles of C24-ethylsterols beyond membrane reinforcement have never been clarified in
the multi-cellular plants expressing the SMT2 gene family or in other organisms that
produce C24-ethylsterols, albeit with a different 24-ethyl-orientation (Fig. 2).
While Qian et al. (2013) reported that exogenous application of stigmasterol
(C24-ethylsterol) and campesterol (C24-methylsterol) partially rescued the stomatal

6
development and patterning of the Arabidopsis fk-J3158 mutants, (fk-J3158 is a weak
allele of the sterol C-14 reductase gene FACKEL), exogenously added sterols have
been usually ineffective in the rescue of mutant phenotypes (Diener et al., 2000;
Willemsen et al., 2003; Schrick et al., 2002; Carland et al., 2002; Carland et al., 2010),
and thus it remains unknown whether C24-ethylsterols play a crucial role in plant
development. Aiming to clarify whether C24-ethylsterols may be play essential roles or
be involved in plant development, we studied the strikingly abnormal phenotype of an
Arabidopsis line (smt2/smt2-smt3/smt3) defective in both the SMT2 and SMT3 genes,
which encode the sterol methyltransferases in charge of the second methyltransfer
reaction at the C24 position of the side chain (Fig. 1). Here, we show that -sitosterol (a
C24-ethylsterol) is necessary and sufficient not only for auxin responses but also for the
cell division plane determination.

7
Figure 1. The sterol biosynthetic pathway in Arabidopsis
The biosynthetic pathway involves two methyltransfer reactions at C24 position. The first reaction is catalyzed by SMT1, leading to the production of C24-methylsterols, and the second reaction is catalyzed by SMT2 and SMT3, yielding C24-ethylsterols.
8

9
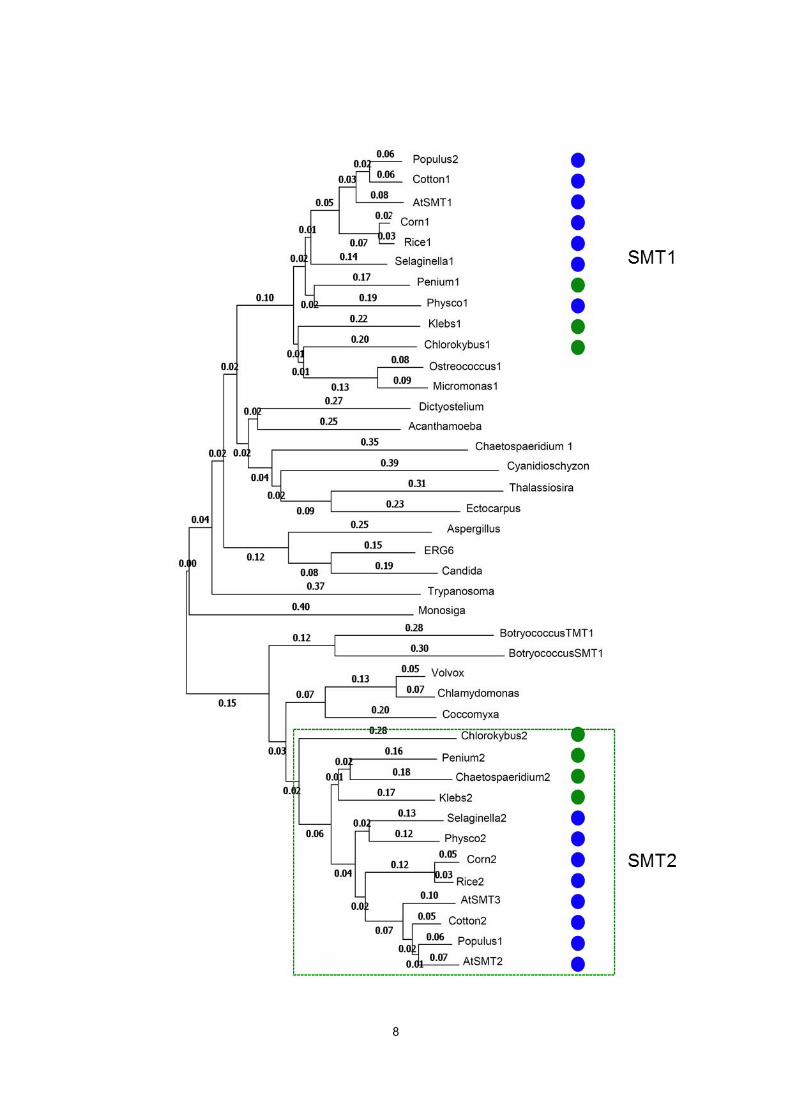
1; Maize2, 2; Maize1, 3; Volvox, 4; Trypanosoma, 5; Thalassiosira, 6; Selaginella1, 7; Selaginella2, 8; Populus2, 9; Populus1, 10; Penium2, 11; Penium1, 12; Physco2, 13; Physco1, 14;Ostreococcus1, 15; Rice1, 16; Rice2, 17; Monosiga, 18; Micromonas1, 19; Klebs2, 20; Klebs1, 21; Cotton2, 22; Cotton1, 23; ERG6, 24; Ectocarpus, 25; Dictyostelium, 26; Cyanidioschyzon, 27; Coccomyxa, 28; Chlorokybus2; Chlorokybus1, 30; Chlamydomonas, 31; Chaetospaeridium2, 32; Chaetospaeridium1, 33; Candida, 34; BotryococcusTMT1, 35; BotryococcusSMT2, 36; AtSMT3, 37;AtSMT2, 38; AtSMT1, 39; Aspergillus, 40; Acanthamoeba.

10
Figure 2. Phylogenetic tree for sterol methyltransferase proteins.
Green dots (algae) and blue dots (terrestrial plants) indicate species containing both SMT2 and SMT3 genes.
The Klebsromidium sequences were retrieved at the Klebsormidium genome project (http://www.plantmorphogenesis.bio.titech.ac.jp/~algae_genome_project/klebsormidium/ index.html). The methyltransferases in land plants are indicated with blue dots, and those in charophycean algae are shown with green dots. The second-type sterol methyltransferases (SMT2) are shown with in green dotted line. Nonredundant database using the TBLASTN search in the National Center for Biotechnology. Sterolmethyltransferase sequences were identified in the GenBank Information with the Arabidopsis SMT2 At1g20330 protein sequence as the query. Multiple alignments of sterolmethyltransferase protein sequences were aligned using ClustalW with default parameter values and analyzed using MEGA6 software version 6.0.6 (Tamura et al., 2013). The sterolmethyltransferase sequences used for the analysis were Acanthamoeba; Acanthamoeba castellanii XM_004336492.1, Aspergillus; Aspergillus nidulans XP_001211350, BotryococcusTMT1; Botryococcus braunii AEY68256.1, BotryococcusSMT1; B. braunii AEY68259.1, Candida; Candida tropicalis XP_002548099.1, Chaetospaeridium1; Chaetospaeridium globosum HO399328.1, Chaetospaeridium2; C.
globosum HO404922.1, Chlamydomonas; Chlamydomonas reinhardtii XP_001690775.1, Chlorokybus1; Chlorokybus atmophyticus HO414429, Chlorokybus2; C.atmophyticus HO408954.1, Coccomyxa; Coccomyxa subellipsoidea XP_005649905.1, Maize1; Zea mays P_002441576.1, Maize2; Zea mays NP_001131331.2, Cotton1; Gossypium hirsutum AAZ83345.1, Cotton2; Gossypium hirsutum ABY49844.1, Cyanidioschyzon; Cyanidioschyzon merolae XP_005537914.1, Dictyostelium; Dictyostelium discoideum XP_636481.1, Ectocarpus; Ectocarpus siliculosus FP261080.1, ERG6; Saccharomyces
cerevisiae erg6 NP_013706.1, Klebs2; Klebsormidium flaccidum kfl00449, Klebs1; K.
flaccidum kfl00283, Monosiga; Monosiga brevicollis XP_001748534.1, Micromonas1; Micromonas sp RCC299 XM_002499342.1, Ostreocossus1; Ostreococcus lucimarinus CCE9901 p XM_001416678.1, Penium1; Penium margaritaceum HO583831.1, Penium2; P.
margaritaceum HO608114.1, Populus1; Populus euphratica XM_011036654, Populus2; Populus euphratica XM_011027171, Physco2; Physcomitrella patens, XM_001772181.1, Physco1; P. patens XM_001785643.1, Rice1; Oryza sativa Os07g0206700, Rice2; O. sativa Os03g0136200, SMT1; Arabidopsis thaliana At5g13710, SMT2; A. thaliana At1g20330, SMT3; A. thaliana At1g76090, Selaginella1; Selaginella moellendorffii XM_002968054.1, Selaginella2; S. moellendorffii, XM_002975434.1, Thalassiosira; Thalassiosira pseudonana CCMP1335b XP_002292358.1, Trypanosoma; Trypanosoma cruzi XP_802864, Volvox; Volvox carteri XP_002948023.1.

11
Chapter1
Establishment and characterization of an Arabidopsis C24-ethylsterol
deficient mutant line
Sterol composition of smt2 smt3 plants
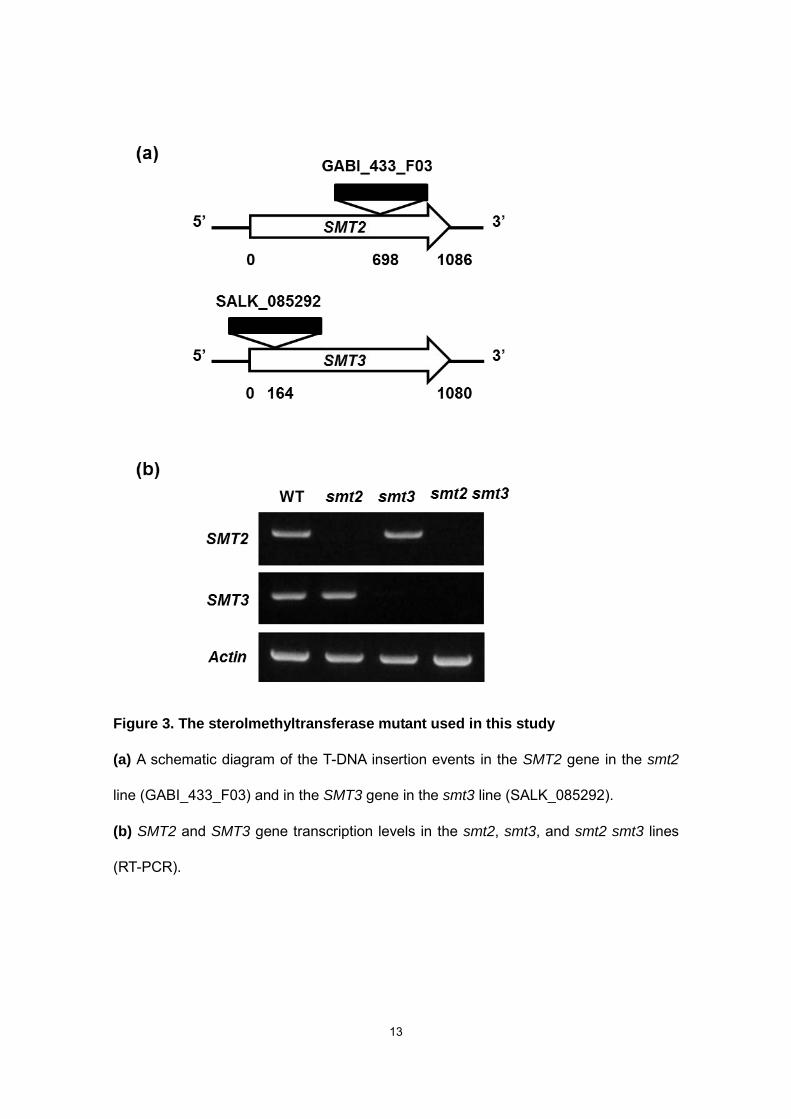
We crossed two T-DNA insertion lines (GABI_433_F03 for smt2 and SALK_085292
for smt3) with each other and maintained a +/smt2-smt3/smt3 line for self-pollination to
obtain homozygous smt2/smt2-smt3/smt3 (smt2 smt3, hereafter) plants, which are
self-sterile (described below). The smt2 smt3 plants lack both the SMT2 and SMT3
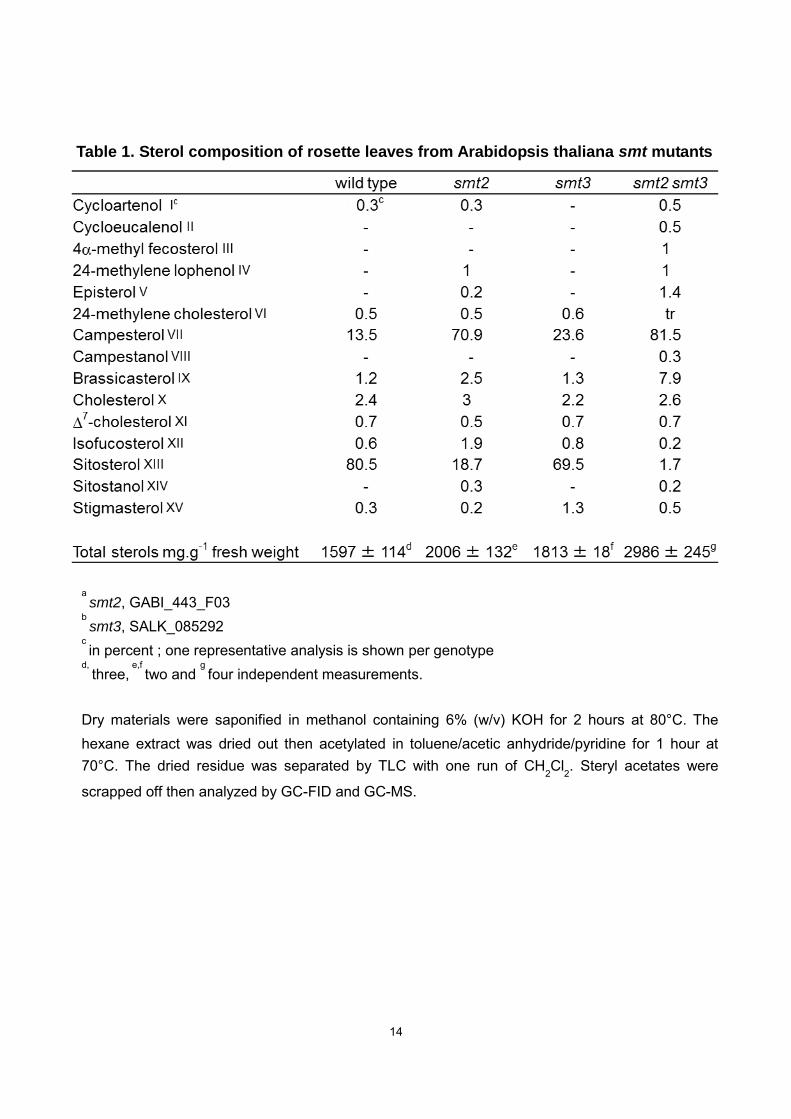
transcripts (Fig. 3) and exhibit a sterol profile highly distorted towards C24-methylsterol
accumulation accompanied by an almost complete loss of C24-ethylsterols (Table 1). In
smt2 and smt2 smt3, the faint C24-ethylsterol levels could be ascribed to the weak
substrate reactivity of SMT1 toward 24-methylenelophenol (Diener et al., 2000).
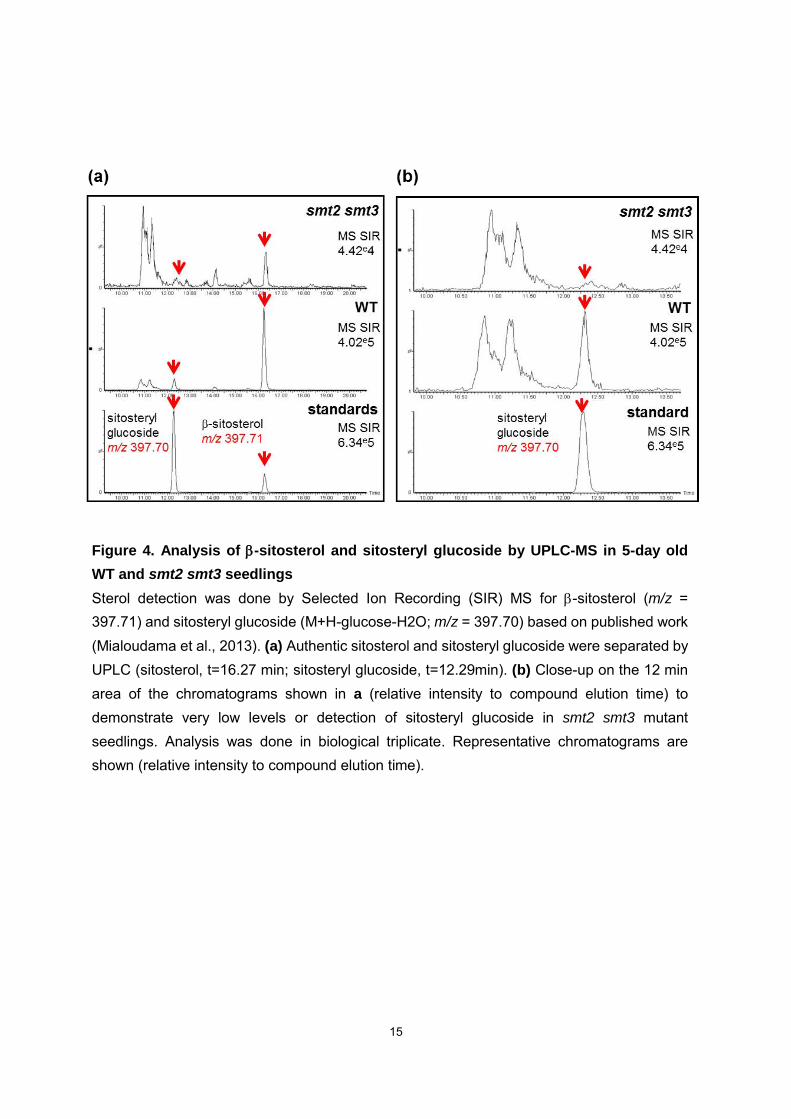
Drastically reduced amount of sitosterol (that accounted for 80% of the total sterol in the
wild-type and less than 2% in smt2 smt3) led also to a complete disappearance of
sitosterol glucoside (Fig. 4), a compound commonly found in the pool of sterol
conjugates in Arabidopsis (DeBolt et al., 2009). Moreover, plant sterol profiles usually
display minor amounts of 4,4-dimethyl- and 4-methyl sterol precursors in proportions
that may vary according to species (Nes and McKean, 1977; Benveniste, 2004; Schaller,
2010). We measured in the mutants considered here a relative increase in the content
of those precursors (cycloartenol, cycloeucalenol, 4-methylfecosterol, 24-methylene
lophenol, Table 1) that represented 0.3% of the total in the wild-type and 1.3% and 3%
in smt2 and smt2 smt3 mutants, respectively. We did not detect any accumulation of

12
unusual sterols reported in plants mutant affected in genes upstream of SMT2 step,
such as the cpi1-1 (Men et al., 2008). The slight increase in 4,4-methyl- and 4-methyl-
sterols prompted a search for 4-carboxy-4-methyl sterol biosynthetic intermediates
(Mialoundama et al., 2013) that might disturb polar auxin transport in smt2 smt3. Those
derivatives were not detected in either the WT or smt2 smt3 mutant seedlings (Fig. 5).
13
Figure 3. The sterolmethyltransferase mutant used in this study
(a) A schematic diagram of the T-DNA insertion events in the SMT2 gene in the smt2
line (GABI_433_F03) and in the SMT3 gene in the smt3 line (SALK_085292).
(b) SMT2 and SMT3 gene transcription levels in the smt2, smt3, and smt2 smt3 lines
(RT-PCR).
14
a smt2, GABI_443_F03
b smt3, SALK_085292
c in percent ; one representative analysis is shown per genotype
d, three,
e,f two and
g four independent measurements.
Dry materials were saponified in methanol containing 6% (w/v) KOH for 2 hours at 80°C. The hexane extract was dried out then acetylated in toluene/acetic anhydride/pyridine for 1 hour at 70°C. The dried residue was separated by TLC with one run of CH2Cl2. Steryl acetates were
scrapped off then analyzed by GC-FID and GC-MS.
Table 1. Sterol composition of rosette leaves from Arabidopsis thaliana smt mutants
15
Figure 4. Analysis of -sitosterol and sitosteryl glucoside by UPLC-MS in 5-day old
WT and smt2 smt3 seedlings Sterol detection was done by Selected Ion Recording (SIR) MS for -sitosterol (m/z = 397.71) and sitosteryl glucoside (M+H-glucose-H2O; m/z = 397.70) based on published work (Mialoudama et al., 2013). (a) Authentic sitosterol and sitosteryl glucoside were separated by UPLC (sitosterol, t=16.27 min; sitosteryl glucoside, t=12.29min). (b) Close-up on the 12 min area of the chromatograms shown in a (relative intensity to compound elution time) to demonstrate very low levels or detection of sitosteryl glucoside in smt2 smt3 mutant seedlings. Analysis was done in biological triplicate. Representative chromatograms are shown (relative intensity to compound elution time).
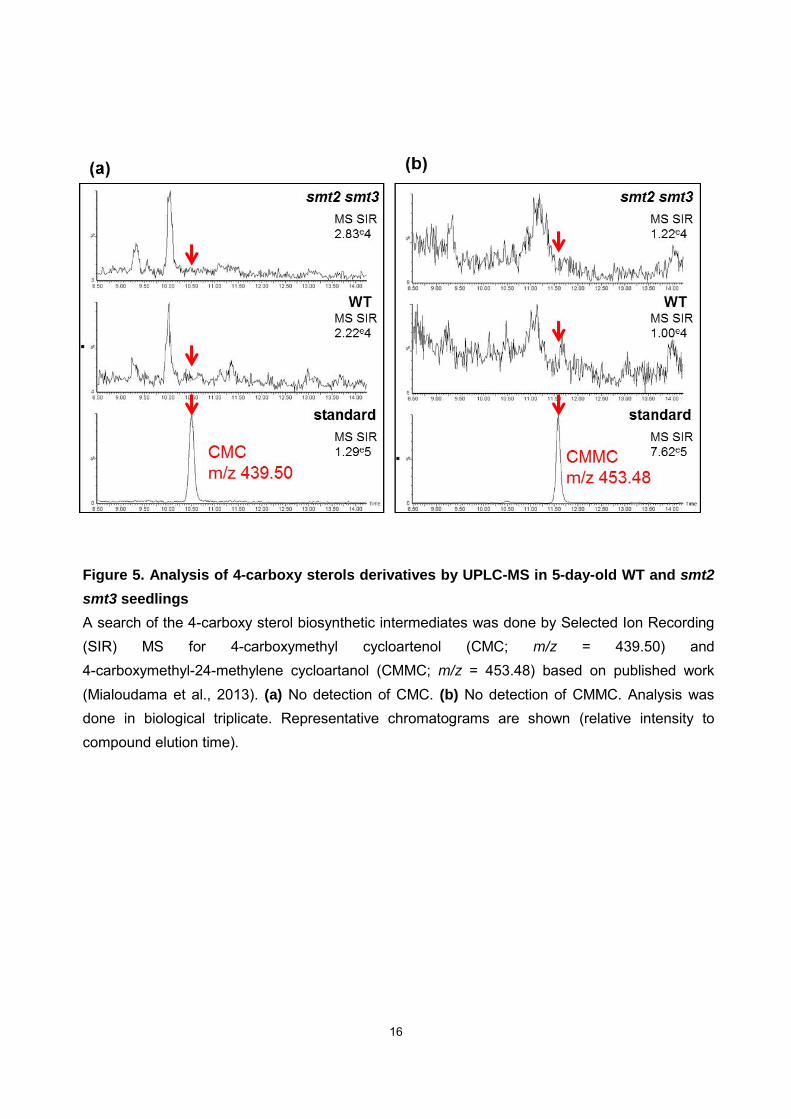
16
Figure 5. Analysis of 4-carboxy sterols derivatives by UPLC-MS in 5-day-old WT and smt2
smt3 seedlings
A search of the 4-carboxy sterol biosynthetic intermediates was done by Selected Ion Recording (SIR) MS for 4-carboxymethyl cycloartenol (CMC; m/z = 439.50) and 4-carboxymethyl-24-methylene cycloartanol (CMMC; m/z = 453.48) based on published work (Mialoudama et al., 2013). (a) No detection of CMC. (b) No detection of CMMC. Analysis was done in biological triplicate. Representative chromatograms are shown (relative intensity to compound elution time).

17
Shoot phenotypes of smt2 smt3 plants
The smt2 smt3 plants exhibit phenotypes similar to those reported for the cvp1-3
smt3 mutant (Carland et al., 2010), including severe growth retardation, abnormal
flowers, aberrant vascular development, and abnormal leaf development (Fig. 6a, 6b,
6c, 6d). In addition to those phenotypes, we observed that the smt2 smt3 mutant was
self-sterile (Fig. 6e), which could be ascribed to the dysfunctional development of the
female gametogenesis (Fig. 6f) and transmitting tract in pistils (Fig. 6g). The transmitting
tract is essential for the growth of pollen tubes that deliver sperms to the egg cells. The
smt2 smt3 pollens were viable and could cross-pollinate wild type (WT) flowers. A
detailed phenotypic characterization of SMT2 cosuppressed lines (smt2COSUP) has been
reported (Schaeffer et al., 2001). The developmental progression of integuments in
smt2COSUP ovules is impaired whereas the funiculus has the same pattern in both
genotypes. The formation of integuments that protect the embryo sac is most probably
affected by a dramatic reduction of SMT2 expression and subsequently -sitosterol
production (Schaeffer et al., 2001), at the early integument formation. Eventually, a few
mature seeds are formed in the smt2COSUP siliques suggesting that the embryo sac is
not affected by the suppressed SMT2 (Schaeffer et al., 2001). The overall
measurements of seed yield indicate a 15-fold reduction in smt2COSUP compared to WT.
The cellular phenotype of smt2COSUP ovules might suggest an action of SMT2 upstream
or during the initiation stage of integuments development (Gasser et al., 1998).
Embryonic phenotypes of smt2 smt3 plants
The embryogenesis in Arabidopsis, the cells of embryo acquires specific fates in
integrated manner to form plant body. The embryo undergoes characteristic shape

18
changes brought about by different orientations of cell divisions and directional cell
expansions. The smt2 smt3 mutant display abnormal embryonic phenotypes during
early stage of embryonic development (Fig. 7). During 8-cell stage and 16-cell stage,
the shrunken suspensor and abnormal cell morphologies at the base of embryo were
observed in smt2 smt3 (Fig. 7a), suggesting aberrant cell division in early embryos.
smt2 smt3 embryos showed irregular divisions in cells at the center of globular and
heart stage embryos (Fig. 7a). Moreover the DR5:GUS expression in smt2 smt3
embryos was strikingly intense not only in the columella cell but also the whole embryo
(Fig. 7b), suggesting that the directional auxin transport was defective. Mutations in
genes that act upstream of 24-methylene lophenol (smt1/cph, fk and hyd1) which lack
total sterols exhibit embryonic defects (Schrick et al., 2002; Clouse, 2002), but those
downstream do not (cvp1, dwf1, dwf5, dwf7) (Clouse, 2002). These results indicate that
C24-ethylsterols are required for embryo development.
Root phenotypes of smt2 smt3 plants
The smt2 smt3 displays extensive abnormalities in the roots (Fig. 8), whereas
SMT2 and SMT3 genes are specifically expressed in the root elongation zone (Carland
et al., 2002). A representative smt2 smt3 seedling phenotype is characterized by a
twisted short root (Fig. 8a, 8b). The root development is completely disrupted,
demonstrating abnormal tissue architecture, occasional swollen cells, and severe
growth inhibition (Fig. 8c). The abnormal tissue architecture in smt2 smt3 roots is
visualized by the staining of membranes (FM4-64; Fig. 9a), and cell walls (propidium
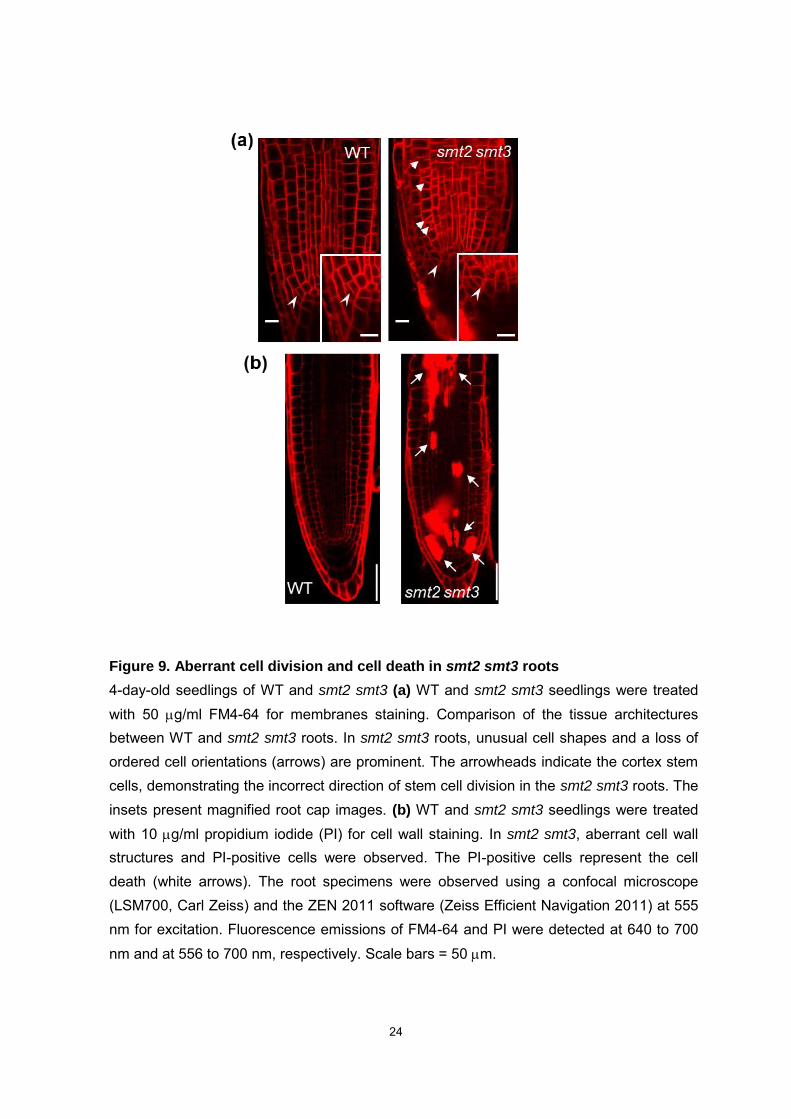
iodide, PI; Fig. 9b). Abnormal cell division was prominent in the smt2 smt3 root stem
cells. FM4-64 staining of smt2 smt3 revealed disruption of the formative division

19
followed by cell lineages with deformed shapes and irregular orientations as well as an
abnormally formed cell plate. In asymmetric divisions, the division planes are governed
by the polarity of the mother cell (Rasmussen et al., 2010). Thus, abnormal cytokinesis
in the smt2 smt3 root stem cell is likely responsible for the defects in subsequent cell
division that lead to the collapsed architecture of the subsequent cell lineages (Fig. 9a).
Furthermore, smt2 smt3 roots harbor a number of PI-positive cells, including root
meristem, vascular cells, and in epidermis and cortex cells (Fig. 9b). PI does not
penetrate into living cells across the plasma membrane (PM) (Curtis et al., 2007). Thus,
the PI-positive cells in smt2 smt3 roots demonstrate not only collapsed maintenance of
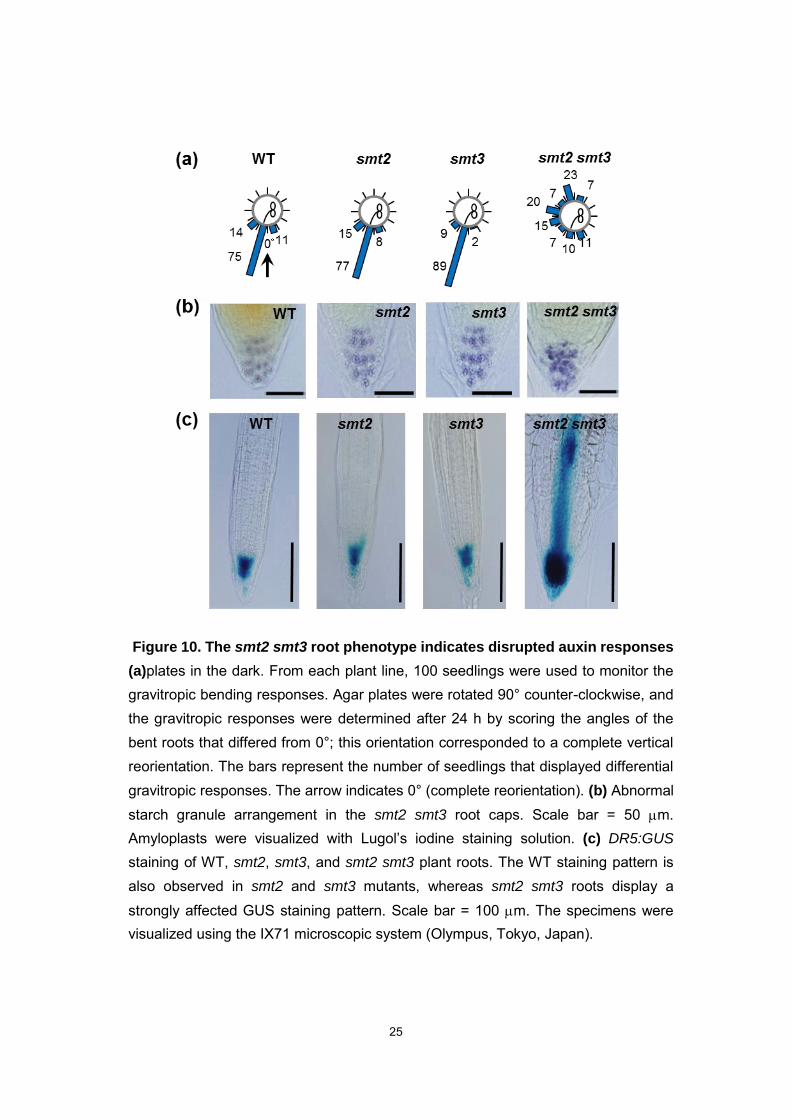
root meristem but also somatic cell death. In smt2 smt3 roots, the loss of gravitropism
and the disordered orientation of starch granules suggested that the auxin-related
cellular functions were impaired (Fig. 10a, 10b). DR5:GUS expression in the smt2 smt3
was strikingly intense in the root tip and throughout the root vascular system (Fig. 10c),
suggesting that the defect was in the directional auxin transport and/or due to excess
auxin accumulation. In WT plants, the PIN2 auxin efflux transporter localized correctly,
maintaining the downward and upward directions in the epidermis and the cortex cells,
respectively (Carland et al., 2010; Blilou et al., 2005). Multiple mislocalization patterns of
PIN2-GFP proteins were evident in smt2 smt3 on the apical side, basal side, and on the
disconnected cell wall (Fig. 11a, Fig. 12). PIN2-GFP protein levels were lower in smt2
smt3 than WT roots (Fig. 11b), and GFP-positive granular structures were observed in
smt2 smt3 root cells (Fig. 11c). Thus, these PIN2 mislocalization patterns were ascribed
to failure of cell division in smt2 smt3.

20
Auxin biosynthesis and auxin degradation
The DR5:GUS expression in smt2 smt3 roots was strikingly intense, suggesting
that auxin biosynthesis and/or auxin degradation may be disturbed in smt2 smt3.
However, indole-3-acetic acid (IAA) biosynthesis genes including TAA1, YUC1, YUC4,
YUC6, CYP79B2 and CYP79B3 expression levels in smt2 smt3 seedlings did not
increase as compared to those of WT (Fig. 13). And GH3 genes which are involved in
IAA degradation (GH3.2, GH3.5 and GH3.6) expression levels were also unaffected in
smt2 smt3 seedlings (Fig. 13).These results indicated that the lack of C24-ethylsterol
didn’t affect the auxin biosynthesis and the auxin degradation.
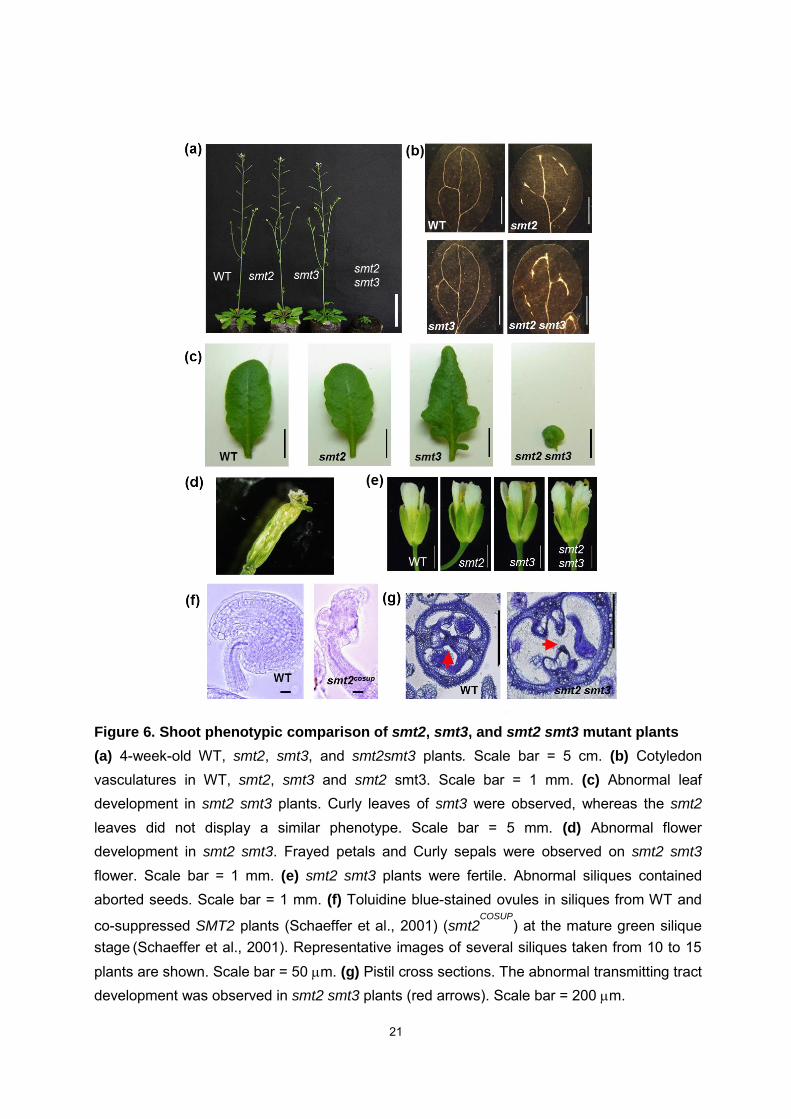
21
Figure 6. Shoot phenotypic comparison of smt2, smt3, and smt2 smt3 mutant plants
(a) 4-week-old WT, smt2, smt3, and smt2smt3 plants. Scale bar = 5 cm. (b) Cotyledon vasculatures in WT, smt2, smt3 and smt2 smt3. Scale bar = 1 mm. (c) Abnormal leaf development in smt2 smt3 plants. Curly leaves of smt3 were observed, whereas the smt2 leaves did not display a similar phenotype. Scale bar = 5 mm. (d) Abnormal flower development in smt2 smt3. Frayed petals and Curly sepals were observed on smt2 smt3
flower. Scale bar = 1 mm. (e) smt2 smt3 plants were fertile. Abnormal siliques contained aborted seeds. Scale bar = 1 mm. (f) Toluidine blue-stained ovules in siliques from WT and
co-suppressed SMT2 plants (Schaeffer et al., 2001) (smt2COSUP
) at the mature green silique stage
(Schaeffer et al., 2001). Representative images of several siliques taken from 10 to 15
plants are shown. Scale bar = 50 m. (g) Pistil cross sections. The abnormal transmitting tract development was observed in smt2 smt3 plants (red arrows). Scale bar = 200 m.
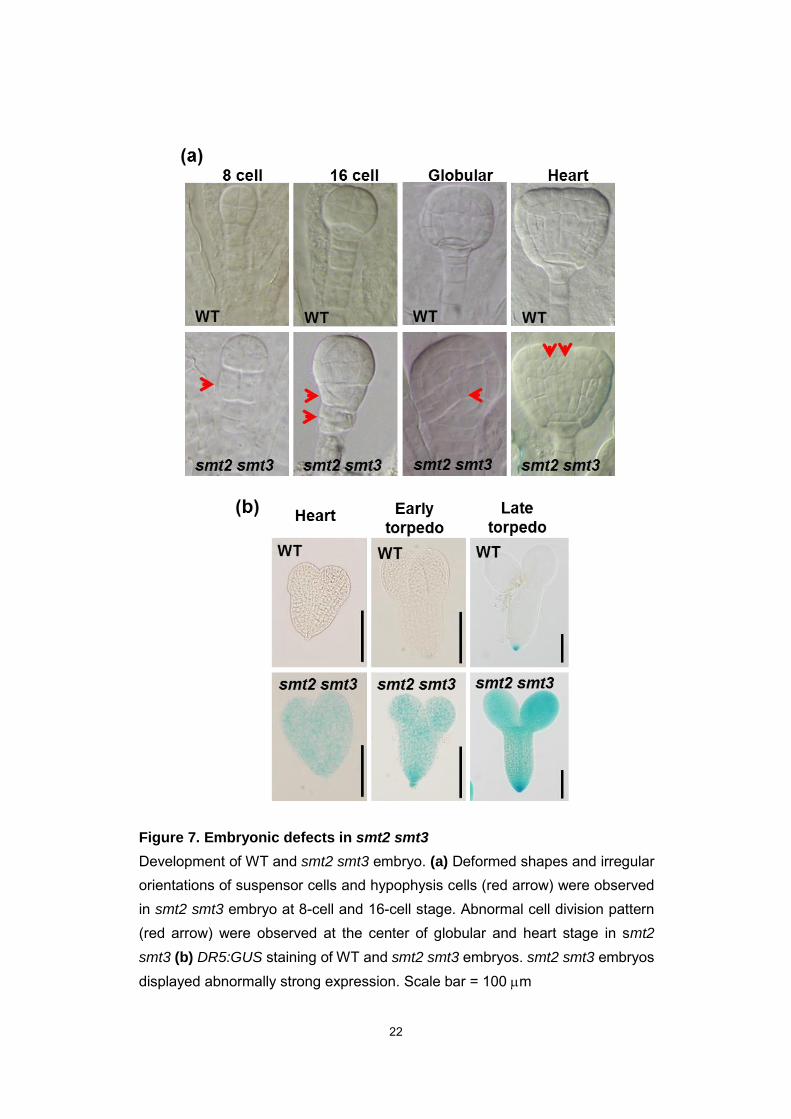
22
Figure 7. Embryonic defects in smt2 smt3
Development of WT and smt2 smt3 embryo. (a) Deformed shapes and irregular orientations of suspensor cells and hypophysis cells (red arrow) were observed in smt2 smt3 embryo at 8-cell and 16-cell stage. Abnormal cell division pattern (red arrow) were observed at the center of globular and heart stage in smt2
smt3 (b) DR5:GUS staining of WT and smt2 smt3 embryos. smt2 smt3 embryos displayed abnormally strong expression. Scale bar = 100 m
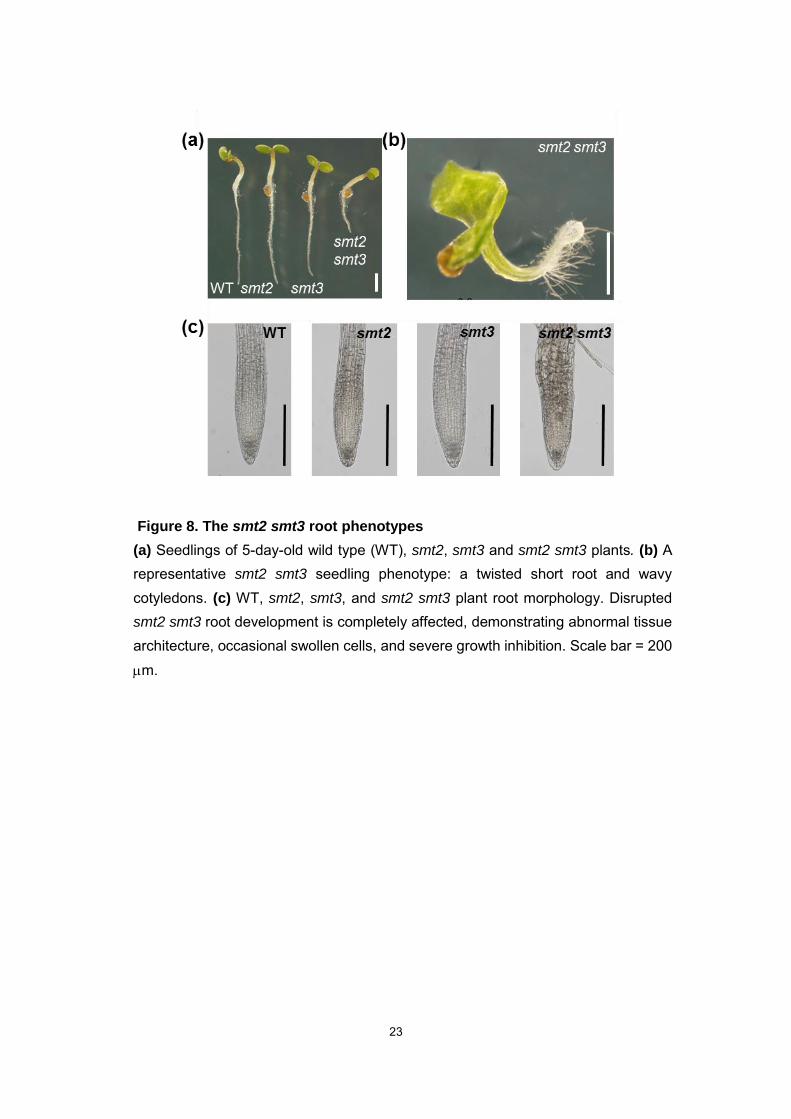
23
Figure 8. The smt2 smt3 root phenotypes
(a) Seedlings of 5-day-old wild type (WT), smt2, smt3 and smt2 smt3 plants. (b) A representative smt2 smt3 seedling phenotype: a twisted short root and wavy cotyledons. (c) WT, smt2, smt3, and smt2 smt3 plant root morphology. Disrupted smt2 smt3 root development is completely affected, demonstrating abnormal tissue architecture, occasional swollen cells, and severe growth inhibition. Scale bar = 200 m.
24
Figure 9. Aberrant cell division and cell death in smt2 smt3 roots
4-day-old seedlings of WT and smt2 smt3 (a) WT and smt2 smt3 seedlings were treated with 50 g/ml FM4-64 for membranes staining. Comparison of the tissue architectures between WT and smt2 smt3 roots. In smt2 smt3 roots, unusual cell shapes and a loss of ordered cell orientations (arrows) are prominent. The arrowheads indicate the cortex stem cells, demonstrating the incorrect direction of stem cell division in the smt2 smt3 roots. The insets present magnified root cap images. (b) WT and smt2 smt3 seedlings were treated with 10 g/ml propidium iodide (PI) for cell wall staining. In smt2 smt3, aberrant cell wall structures and PI-positive cells were observed. The PI-positive cells represent the cell death (white arrows). The root specimens were observed using a confocal microscope (LSM700, Carl Zeiss) and the ZEN 2011 software (Zeiss Efficient Navigation 2011) at 555 nm for excitation. Fluorescence emissions of FM4-64 and PI were detected at 640 to 700 nm and at 556 to 700 nm, respectively. Scale bars = 50 m.
25
Figure 10. The smt2 smt3 root phenotype indicates disrupted auxin responses
(a)plates in the dark. From each plant line, 100 seedlings were used to monitor the gravitropic bending responses. Agar plates were rotated 90° counter-clockwise, and the gravitropic responses were determined after 24 h by scoring the angles of the bent roots that differed from 0°; this orientation corresponded to a complete vertical reorientation. The bars represent the number of seedlings that displayed differential gravitropic responses. The arrow indicates 0° (complete reorientation). (b) Abnormal starch granule arrangement in the smt2 smt3 root caps. Scale bar = 50 m. Amyloplasts were visualized with Lugol’s iodine staining solution. (c) DR5:GUS
staining of WT, smt2, smt3, and smt2 smt3 plant roots. The WT staining pattern is also observed in smt2 and smt3 mutants, whereas smt2 smt3 roots display a strongly affected GUS staining pattern. Scale bar = 100 m. The specimens were visualized using the IX71 microscopic system (Olympus, Tokyo, Japan).

26
Figure 11. Abnormal cell file and loss of PIN2-GFP polar localization in smt2 smt3 roots
(a) The PIN2-GFP-expressing line was stained with 50 g/ml FM4-64. In WT plants, PIN2-GFP localizes upward in epidermal cells and downward in both cortical (c) and epidermal cells (e). In smt2 smt3 plants, abnormal PIN2-GFP localization is observed with occasional lateral side and upside-down localizations (white arrowheads). An aberrant cell file is also prominent in smt2 smt3 roots (circled by the dotted line). (b) PIN2-GFP localization in the WT and smt2 smt3 root epidermal cells. The PIN2-GFP level was significantly lower in smt2 smt3 than in the WT. (c) Small PIN2-GFP positive particles (white arrows) were observed in the smt2 smt3 root epidermal cells. Scale bar = 10 m
27
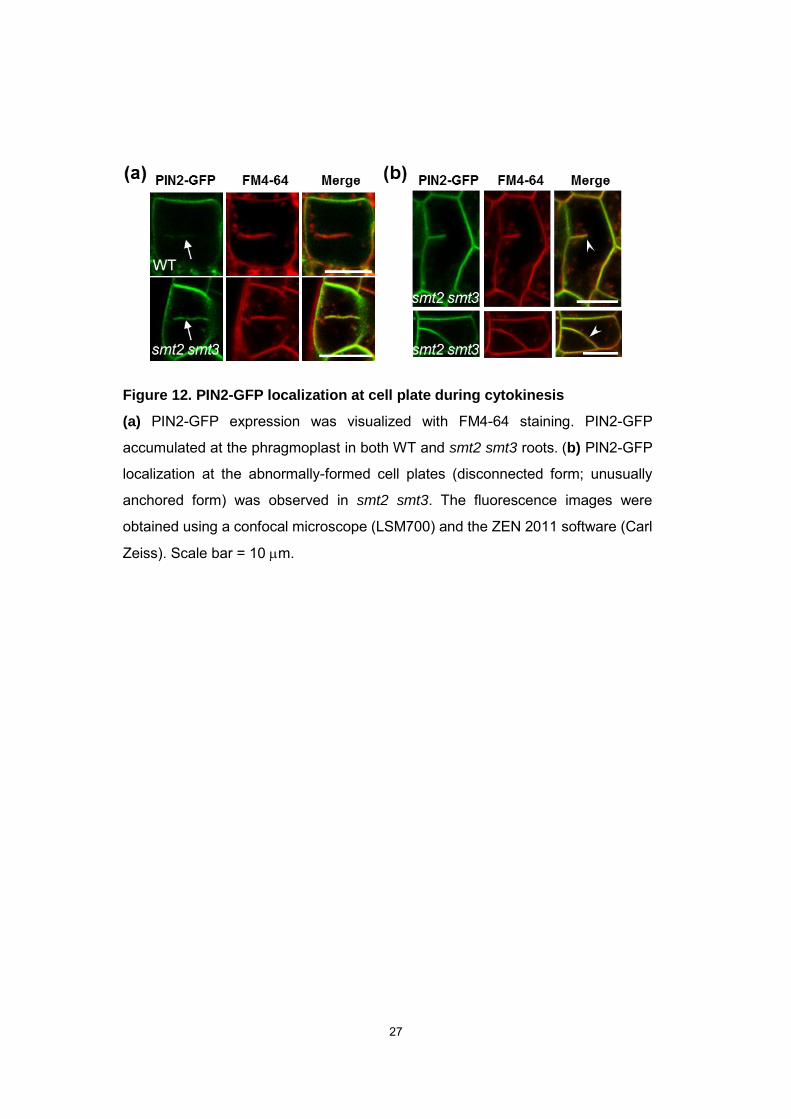
Figure 12. PIN2-GFP localization at cell plate during cytokinesis
(a) PIN2-GFP expression was visualized with FM4-64 staining. PIN2-GFP
accumulated at the phragmoplast in both WT and smt2 smt3 roots. (b) PIN2-GFP
localization at the abnormally-formed cell plates (disconnected form; unusually
anchored form) was observed in smt2 smt3. The fluorescence images were
obtained using a confocal microscope (LSM700) and the ZEN 2011 software (Carl
Zeiss). Scale bar = 10 m.

28
Figure 13. Auxin biosynthesis and degradation in smt2 smt3 seedlings
(a) IAA biosynthesis and degradation gene transcription levels in WT and smt2 smt3
seedling were determined by semi-quantitative RT-PCR. 7-day-old seedlings germinated on MS-agar plates. PCR reaction cycles are indicated to the top of the panels. (b) auxin biosynthesis pathway.
IAA: indole-3-acetic acid, IPA: indole-3-pyruvic acid, IAOx: indole-3-acetaldoxime, IAN: indole-3-acetonitrile, IAM: indole-3-acetamide, TAM: tryptamine, IAAld: indole-3-acetaldehyde

29
Cell division is disrupted in smt2 smt3 plants
Abnormal cell division is prominent in the smt2 smt3 root stem cells (Fig. 9a). The
FM4-64 staining of smt2 smt3 roots revealed the disruption of the formative division of
root stem cells followed by cell lineages with deformed shapes and irregular orientations
as well as abnormally formed cell plates (Fig. 9a, 12b). In the asymmetric divisions, the
division planes are governed by the polarity of the mother cell (Rasmussen et al., 2010).
Thus, the abnormal cytokinesis in the smt2 smt3 root stem cells may have caused the
defects in subsequent cell divisions that led to the collapsed architecture of the following
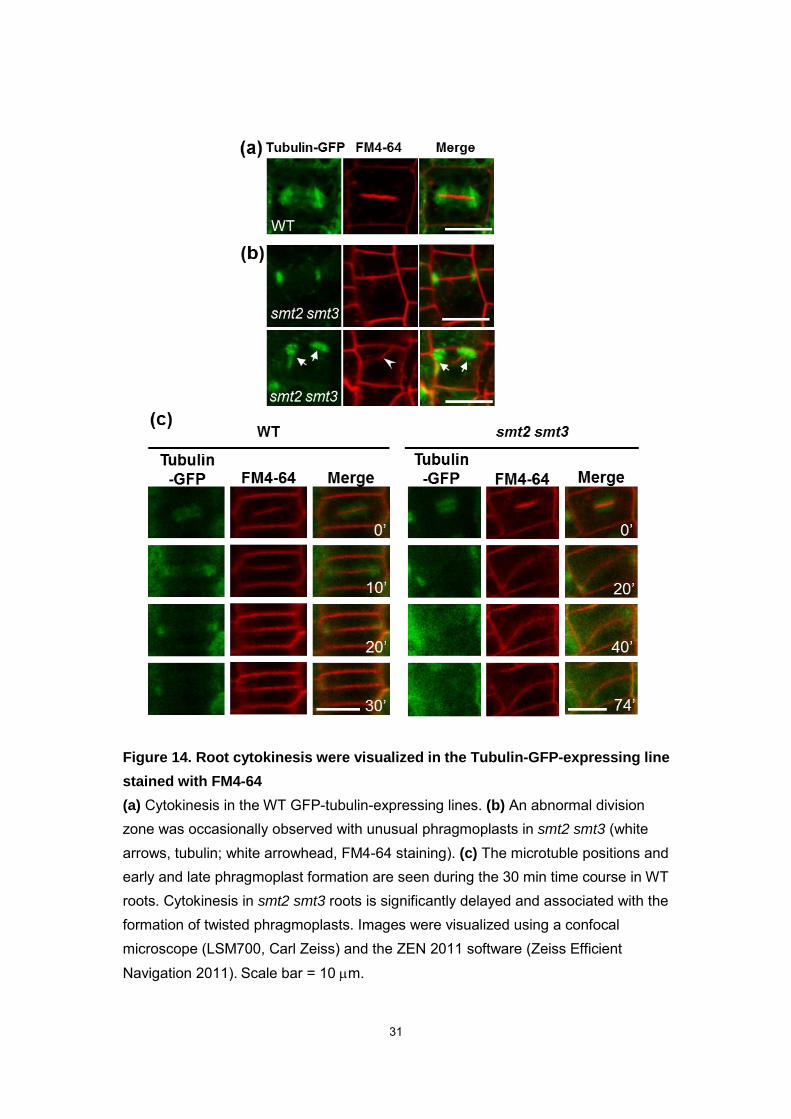
cell lineages (Fig. 9). The FM4-64 staining of the tubulin-GFP expression line revealed
both abnormal and normal cell division in smt2 smt3 roots (Fig. 14b). The time-lapse
imaging analysis revealed delayed cytokinesis, which resulted in improper division zone
formation in smt2 smt3 (Fig. 14c). The misorientation and perpendicular formation of the
preprophase band (PPB) in abnormal cell division were visualized by immunostaining
with anti-tubulin and anti-KNOLLE syntaxin antibodies (Fig. 15a). Arabidopsis KNOLLE
is a vesicle-docking syntaxin specifically expressed during mitosis and localizes to the
cell division plane (Lauber et al., 1997). Abnormal phragmoplast formation in smt2 smt3
was also indicated by the disjunctive distributions of KNOLLE and tubulin, as well as the
disorganized and fragmented spindles (Fig. 15b). Staining with DAPI
(4',6-diamidino-2-phenylindole) revealed the presence of multinucleated cells in the
smt2 smt3 roots (Fig. 15b, 16). Mislocalization of PIN2-GFP was demonstrated at the
unusually formed division zone in smt2 smt3 root cells (Fig. 12).
In smt2 smt3, tubulin-GFP signal was observed with FM4-64 fluorescence during
the phragmoplast formation stage in a similar way as visualized in WT roots (Fig. 14).
PIN2-GFP signal co-localized with FM4-64 (Fig. 12), irrespective of the normal and
abnormal cell division. Also, the immunostaining experiment demonstrated in smt2 smt3
that tubulin assembled at cell plate shown by KNOLLE localization (Fig. 15b). These

30
results indicate that the process of phragmoplast formation and the transport of newly
synthesized PIN2 protein are not defective in smt2 smt3 roots. We did not observe
KNOLLE mis-localization to lateral PM as reported with the upstream pathway mutants,
cpi1-1 and smt1orc, and roots treated with fenpropimorph (Boutté et al., 2010), a general
sterol biosynthetic inhibitor. KNOLLE localization in the cell division plane is maintained
through sterol-dependent endocytosis mediated by a clathrin- and DYNAMIN-RELATED
PROTEIN1A-dependent mechanism (Boutté et al., 2010; Lauber et al., 1997). These
observations could be due to the intrinsic differences in the sterol compositions between
the total defects in the sterol biosynthesis (cpi1-1 and smt1orc) and the specific loss of
C24-ethylsterols (smt2 smt3). In summary, the cytokinesis in smt2 smt3 plants was
delayed, as visualized by the FM4-64 staining of the tubulin-GFP expressing line (Fig.
14c). Thus, smt2 smt3 roots are defective in the determination of the cell division plane,
which is controlled by the consecutive events comprised of the positioning, anchoring,
and guidance of the division zone (Van Damme, 2009).
31
Figure 14. Root cytokinesis were visualized in the Tubulin-GFP-expressing line
stained with FM4-64
(a) Cytokinesis in the WT GFP-tubulin-expressing lines. (b) An abnormal division zone was occasionally observed with unusual phragmoplasts in smt2 smt3 (white arrows, tubulin; white arrowhead, FM4-64 staining). (c) The microtuble positions and early and late phragmoplast formation are seen during the 30 min time course in WT roots. Cytokinesis in smt2 smt3 roots is significantly delayed and associated with the formation of twisted phragmoplasts. Images were visualized using a confocal microscope (LSM700, Carl Zeiss) and the ZEN 2011 software (Zeiss Efficient Navigation 2011). Scale bar = 10 m.
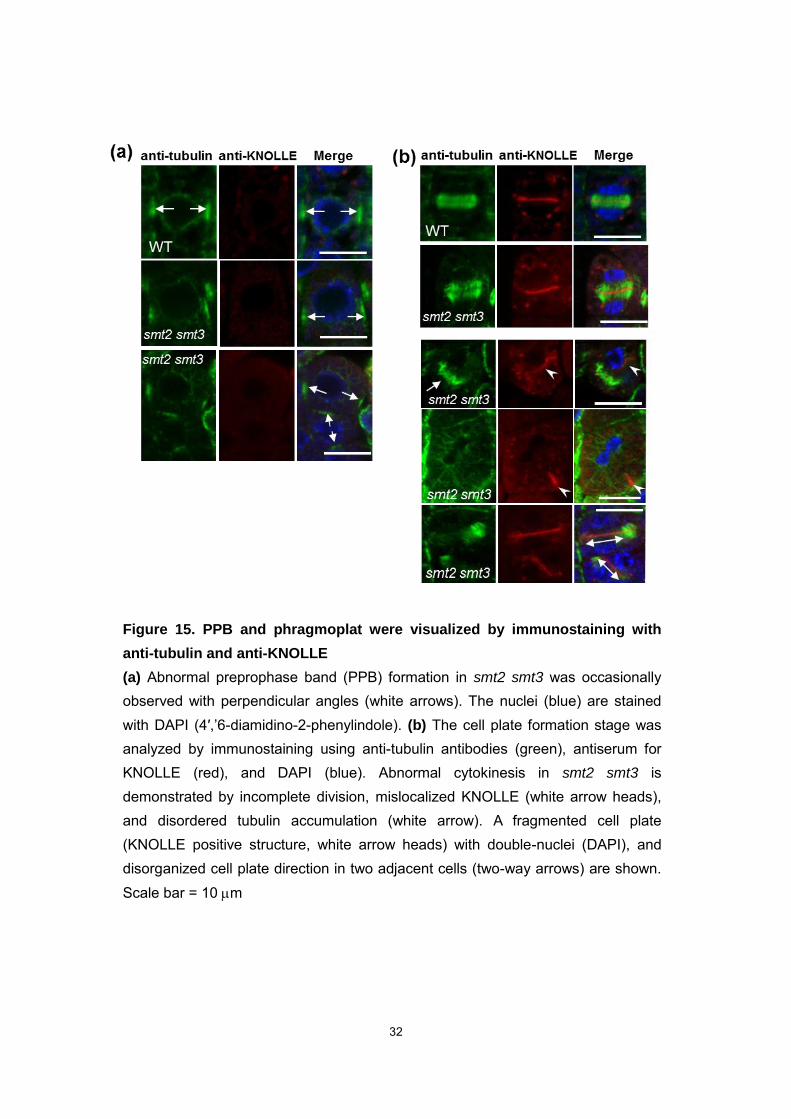
32
Figure 15. PPB and phragmoplat were visualized by immunostaining with
anti-tubulin and anti-KNOLLE
(a) Abnormal preprophase band (PPB) formation in smt2 smt3 was occasionally observed with perpendicular angles (white arrows). The nuclei (blue) are stained with DAPI (4′,’6-diamidino-2-phenylindole). (b) The cell plate formation stage was analyzed by immunostaining using anti-tubulin antibodies (green), antiserum for KNOLLE (red), and DAPI (blue). Abnormal cytokinesis in smt2 smt3 is demonstrated by incomplete division, mislocalized KNOLLE (white arrow heads), and disordered tubulin accumulation (white arrow). A fragmented cell plate (KNOLLE positive structure, white arrow heads) with double-nuclei (DAPI), and disorganized cell plate direction in two adjacent cells (two-way arrows) are shown. Scale bar = 10 m
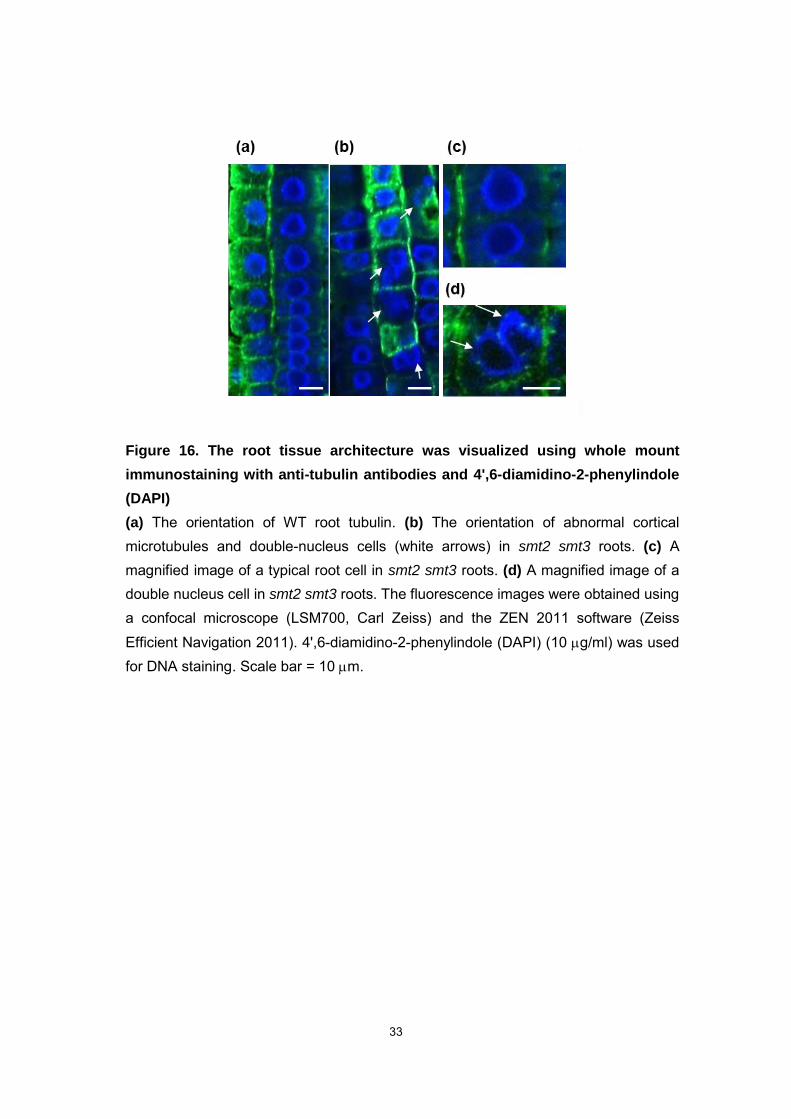
33
Figure 16. The root tissue architecture was visualized using whole mount
immunostaining with anti-tubulin antibodies and 4',6-diamidino-2-phenylindole
(DAPI)
(a) The orientation of WT root tubulin. (b) The orientation of abnormal cortical microtubules and double-nucleus cells (white arrows) in smt2 smt3 roots. (c) A magnified image of a typical root cell in smt2 smt3 roots. (d) A magnified image of a double nucleus cell in smt2 smt3 roots. The fluorescence images were obtained using a confocal microscope (LSM700, Carl Zeiss) and the ZEN 2011 software (Zeiss
Efficient Navigation 2011). 4',6-diamidino-2-phenylindole (DAPI) (10 g/ml) was used for DNA staining. Scale bar = 10 m.

34
Partial restoration of the disrupted cell division by exogenously supplied
-sitosterol
The roots were cultured in a liquid medium supplemented with synthetic auxins to
induce the differentiation of secondary roots. Under typical culture conditions, WT roots
successfully differentiated and developed secondary roots, while the smt2 smt3 roots
yielded only disorganized tissue bumps (Fig. 17a). Intriguingly, exogenous -sitosterol
administration partially rescued the failed secondary root development (Fig. 17a),
without restoring normal C24-ethylsterol levels, including steryl glycosides, acylated
steryl esters, and acylated steryl glycosides (Fig. 18, Table 2). The
-sitosterol-dependent recovery of the second root development was partly substituted
by stigmasterol (another significant C24-ethylsterol) but not by campesterol
(C24-methylsterol) (Fig. 17a, b). Secondary root development in WT plants is
associated with DR5:GUS expression that is localized to the secondary root tips, similar
to that in intact plant roots, which completely disappeared in smt2 smt3 plants (Fig. 17c).
The irregular DR5:GUS expression pattern in the smt2 smt3 roots was also rescued by
the addition of -sitosterol (Fig. 17c), suggesting that directional auxin transport and cell
polarity were amended to enable formative cell division in smt2 smt3 roots. -sitosterol
induced successful secondary root development (Fig. 17). The root culture experiment
may be a suitable system to study the requirements of sterols in plant development.
The disturbed cytokinesis in smt2 smt3 roots was further assessed at the single cell
level. Phragmoplast structures were visualized using GFP-tubulin and FM4-64 staining
in the late cytokinesis of cells from WT and smt2 smt3 cultured roots recovering from
protoplast isolation (Fig. 19). Protoplasts initiated cytokinesis at least 96 h after the
incubation, and almost 75% of WT protoplasts completed abscission at 102 h of
incubation. Conversely, most smt2 smt3 protoplasts failed to form normal
phragmoplasts during cytokinesis (Fig. 19a, b). Cytokinesis completion was scored by

35
visually discriminating between the telophase stage and the completion of cell division
(Fig. 19c). Only 10% of smt2 smt3 protoplasts completed cytokinesis within the same
time course, and further incubation did not increase the cytokinesis achievement rates
(Fig. 19d). However, when protoplasts were isolated from smt2 smt3 roots cultured with
-sitosterol, the cytokinetic abscission rate increased to 30% (Fig. 19d). The addition of
-sitosterol to the culture medium did not elicit acute effects on protoplasts in the
recovery of disrupted cytokinesis. Cellulose fiber formation by WT and smt2 smt3
protoplasts was indistinguishable under our experimental conditions (Fig. 20) The
involvement of sterols in cellulose biosynthesis (Schrick et al., 2004; Schrick et al.,
2012) was not observed in the surfaces of WT and smt2 smt3 protoplast-derived cells
(Fig. 20b). However, aberrant cell wall structures in smt2 smt3 (Fig. 9b) were consistent
with the suggestion that C24-ethylsterols are involved in cellulose biosynthesis. These
results demonstrate that the smt2 smt3 protoplasts cannot determine the cell division
plane and that the cell population capable of normal cytokinesis increased during the
secondary root development in the smt2 smt3 roots cultured with -sitosterol. The
expression of the DR5:GUS reporter suggested that the directional auxin transport in
the smt2 smt3 roots was restored upon -sitosterol supplementation (Fig. 17). The
supplemental -sitosterol may have rescued the dysfunctional root development by
restoring auxin transport and thereby promoting the auxin distribution-dependent plant
development (Dhonukshe et al., 2008).
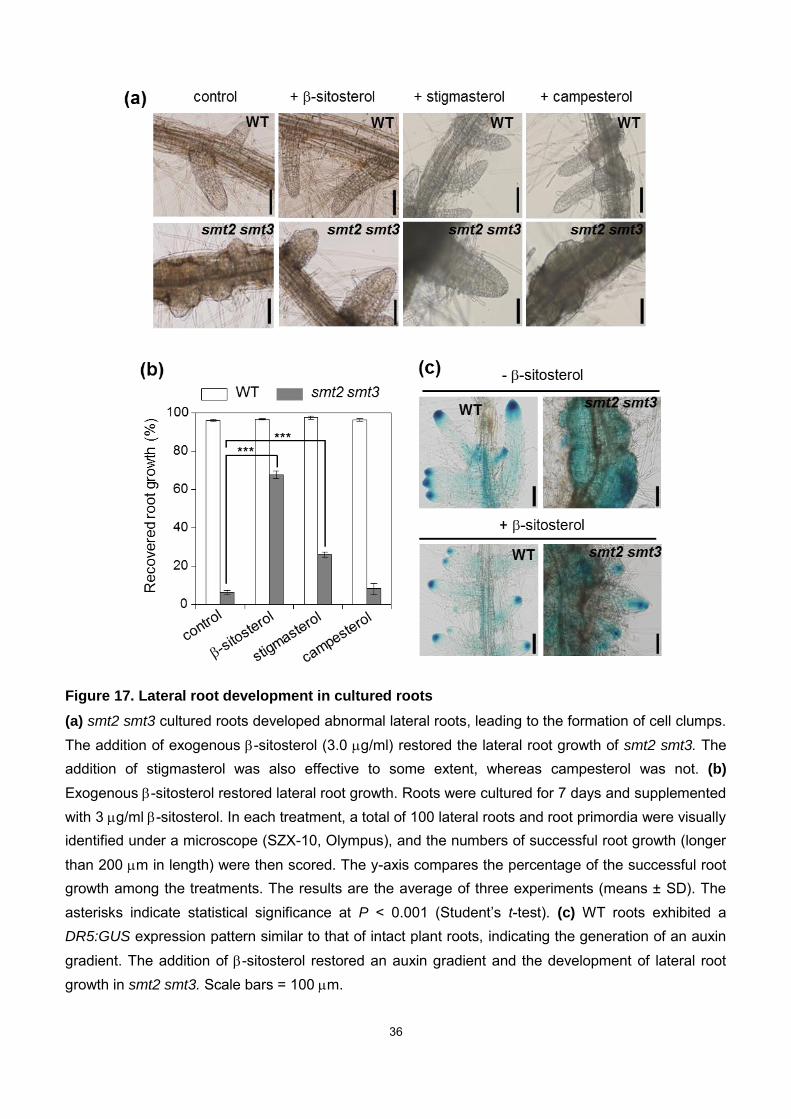
36
Figure 17. Lateral root development in cultured roots
(a) smt2 smt3 cultured roots developed abnormal lateral roots, leading to the formation of cell clumps. The addition of exogenous -sitosterol (3.0 g/ml) restored the lateral root growth of smt2 smt3. The addition of stigmasterol was also effective to some extent, whereas campesterol was not. (b)
Exogenous -sitosterol restored lateral root growth. Roots were cultured for 7 days and supplemented with 3 g/ml -sitosterol. In each treatment, a total of 100 lateral roots and root primordia were visually identified under a microscope (SZX-10, Olympus), and the numbers of successful root growth (longer than 200 m in length) were then scored. The y-axis compares the percentage of the successful root growth among the treatments. The results are the average of three experiments (means ± SD). The asterisks indicate statistical significance at P < 0.001 (Student’s t-test). (c) WT roots exhibited a DR5:GUS expression pattern similar to that of intact plant roots, indicating the generation of an auxin gradient. The addition of -sitosterol restored an auxin gradient and the development of lateral root growth in smt2 smt3. Scale bars = 100 m.
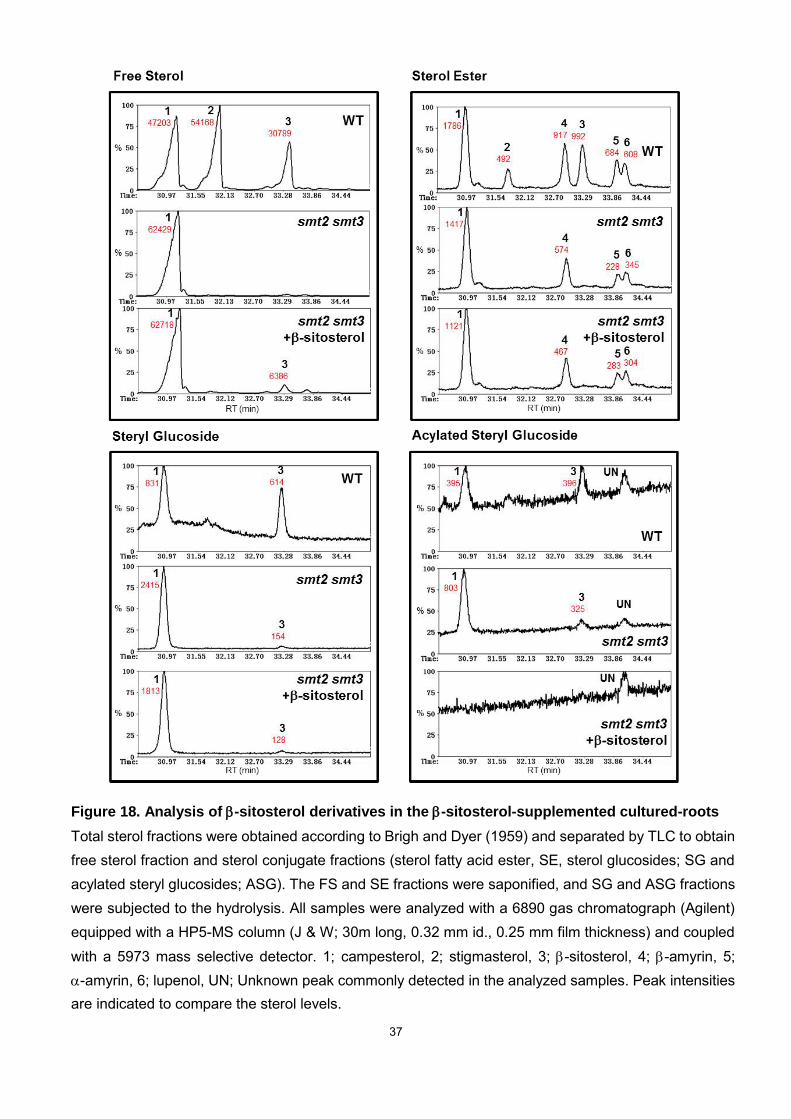
37
Figure 18. Analysis of -sitosterol derivatives in the -sitosterol-supplemented cultured-roots
Total sterol fractions were obtained according to Brigh and Dyer (1959) and separated by TLC to obtain free sterol fraction and sterol conjugate fractions (sterol fatty acid ester, SE, sterol glucosides; SG and acylated steryl glucosides; ASG). The FS and SE fractions were saponified, and SG and ASG fractions were subjected to the hydrolysis. All samples were analyzed with a 6890 gas chromatograph (Agilent) equipped with a HP5-MS column (J & W; 30m long, 0.32 mm id., 0.25 mm film thickness) and coupled with a 5973 mass selective detector. 1; campesterol, 2; stigmasterol, 3; -sitosterol, 4; -amyrin, 5; -amyrin, 6; lupenol, UN; Unknown peak commonly detected in the analyzed samples. Peak intensities are indicated to compare the sterol levels.

38
Table 2. Sterol profiles of the cultured roots
39
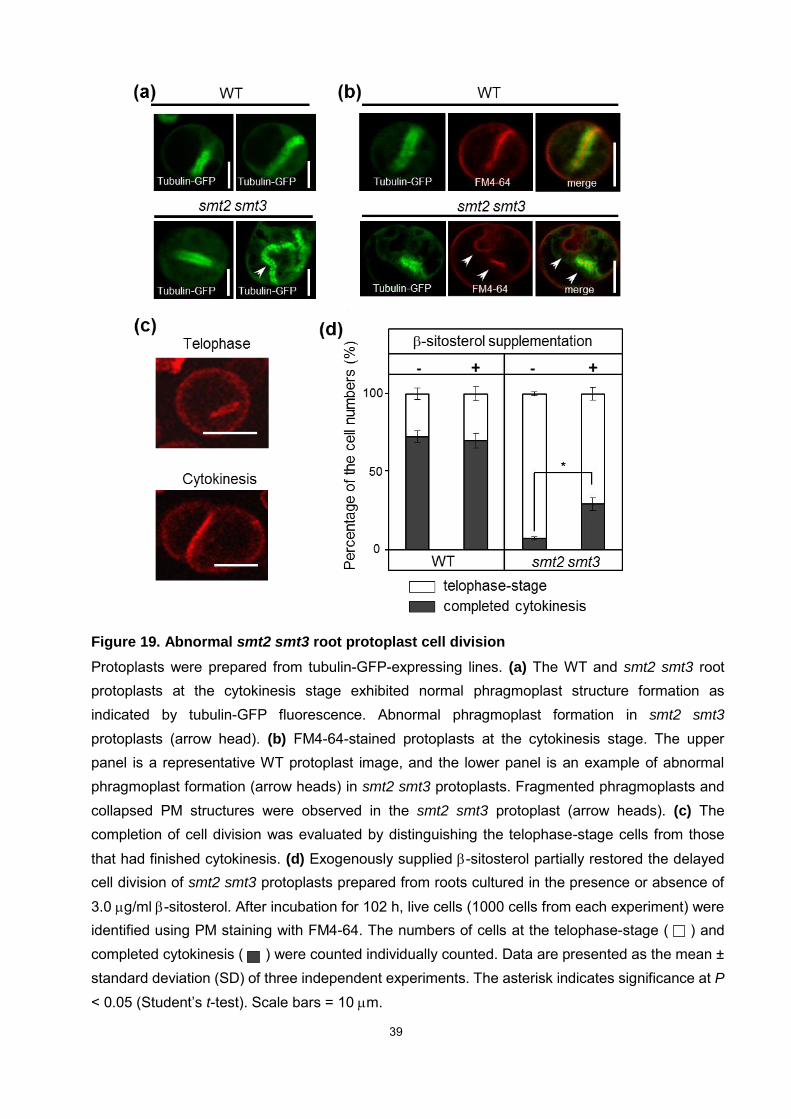
Figure 19. Abnormal smt2 smt3 root protoplast cell division
Protoplasts were prepared from tubulin-GFP-expressing lines. (a) The WT and smt2 smt3 root protoplasts at the cytokinesis stage exhibited normal phragmoplast structure formation as indicated by tubulin-GFP fluorescence. Abnormal phragmoplast formation in smt2 smt3
protoplasts (arrow head). (b) FM4-64-stained protoplasts at the cytokinesis stage. The upper panel is a representative WT protoplast image, and the lower panel is an example of abnormal phragmoplast formation (arrow heads) in smt2 smt3 protoplasts. Fragmented phragmoplasts and collapsed PM structures were observed in the smt2 smt3 protoplast (arrow heads). (c) The completion of cell division was evaluated by distinguishing the telophase-stage cells from those that had finished cytokinesis. (d) Exogenously supplied -sitosterol partially restored the delayed cell division of smt2 smt3 protoplasts prepared from roots cultured in the presence or absence of 3.0 g/ml -sitosterol. After incubation for 102 h, live cells (1000 cells from each experiment) were identified using PM staining with FM4-64. The numbers of cells at the telophase-stage ( ) and completed cytokinesis ( ) were counted individually counted. Data are presented as the mean ± standard deviation (SD) of three independent experiments. The asterisk indicates significance at P
< 0.05 (Student’s t-test). Scale bars = 10 m.
40
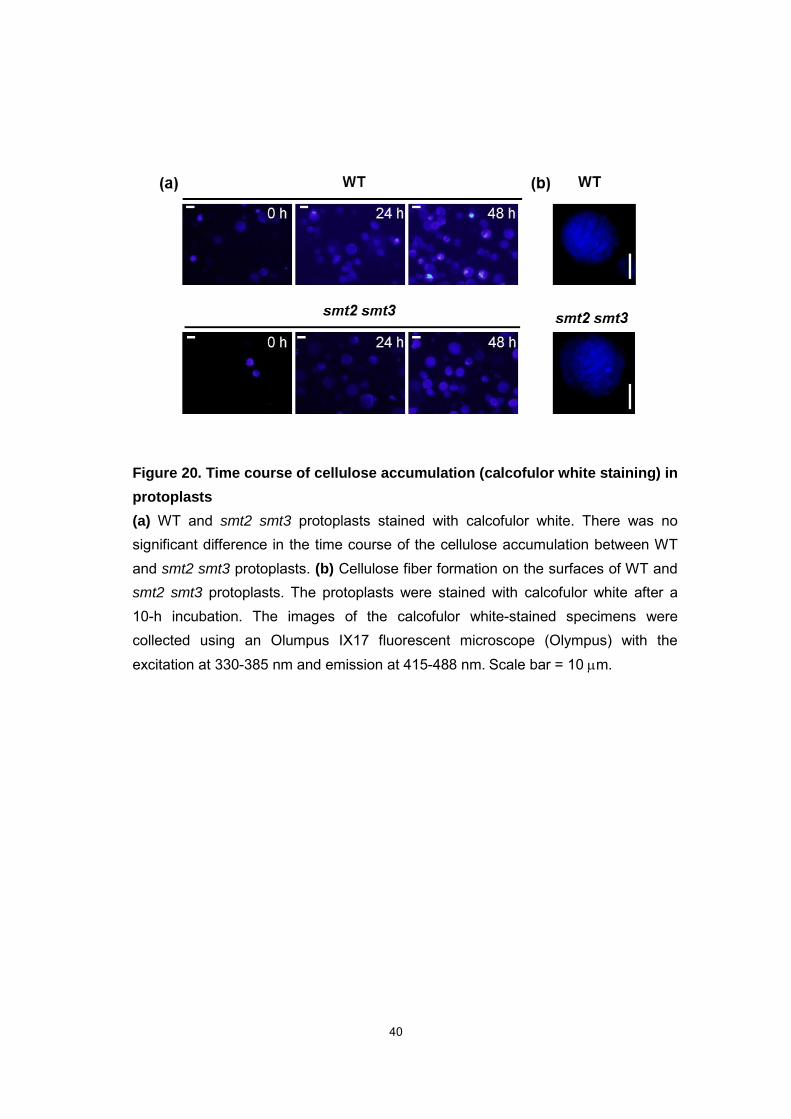
Figure 20. Time course of cellulose accumulation (calcofulor white staining) in
protoplasts
(a) WT and smt2 smt3 protoplasts stained with calcofulor white. There was no significant difference in the time course of the cellulose accumulation between WT and smt2 smt3 protoplasts. (b) Cellulose fiber formation on the surfaces of WT and smt2 smt3 protoplasts. The protoplasts were stained with calcofulor white after a 10-h incubation. The images of the calcofulor white-stained specimens were collected using an Olumpus IX17 fluorescent microscope (Olympus) with the excitation at 330-385 nm and emission at 415-488 nm. Scale bar = 10 m.

41
Lipid order of cell plate in smt2 smt3
The root culture experiments (Fig. 17, 18 and Table 2) demonstrated that a small
amount of -sitosterol (or stigmasterol) critically promoted the recovery of abnormal
growth of lateral roots of smt2 smt3. However, we were not able to identify metabolites
in very low amounts derived from the exogenously supplied -sitosterol in the cultured
roots. The exogenously supplied -sitosterol may accumulate at specific sites, such as
membrane microdomains in plants (Malinsky et al., 2013), in a temporary and spatially
precise manner at either the tissue or single cell level. Frescatada-Rosa et al. (2014)
monitored the membrane lipid packing state by recording generalized polarization (GP)
values (Owen et al., 2012) and observed that the lipid order increased at the cell plate
during cytokinesis in a sterol- and DRP1A- (DYNAMIN-RELATED PROTEIN1A)
dependent manner. The lipid order of cell plate in roots of cpi1-1 and roots treated with
fenpropimorph remain unchanged (Frescatada-Rosa et al. 2014). Their results
prompted us to examine whether the cell plate might be the specific site for
C24-ethylsterol accumulation. If C24-ethylsterols accumulate specifically at the cell
plate and are required for correct cytokinesis, the lipid order at the cell plate in smt2
smt3 lacking C24-ethylsterols should remain unchanged. Then we observed the lipid
order of cell plate in WT and smt2 smt3 root cells. The lipid order in smt2 smt3 shifted
toward a higher order, as observed in the WT, irrespective of correct or failed cytokinesis
(Fig. 28). By contrast, the GP values recorded in smt2 smt3 roots were generally higher
than those in the WT, regardless of cytokinetic stage, than those in the WT, suggesting
intrinsically different membrane properties between the WT and smt2 smt3 (Fig. 21).
42
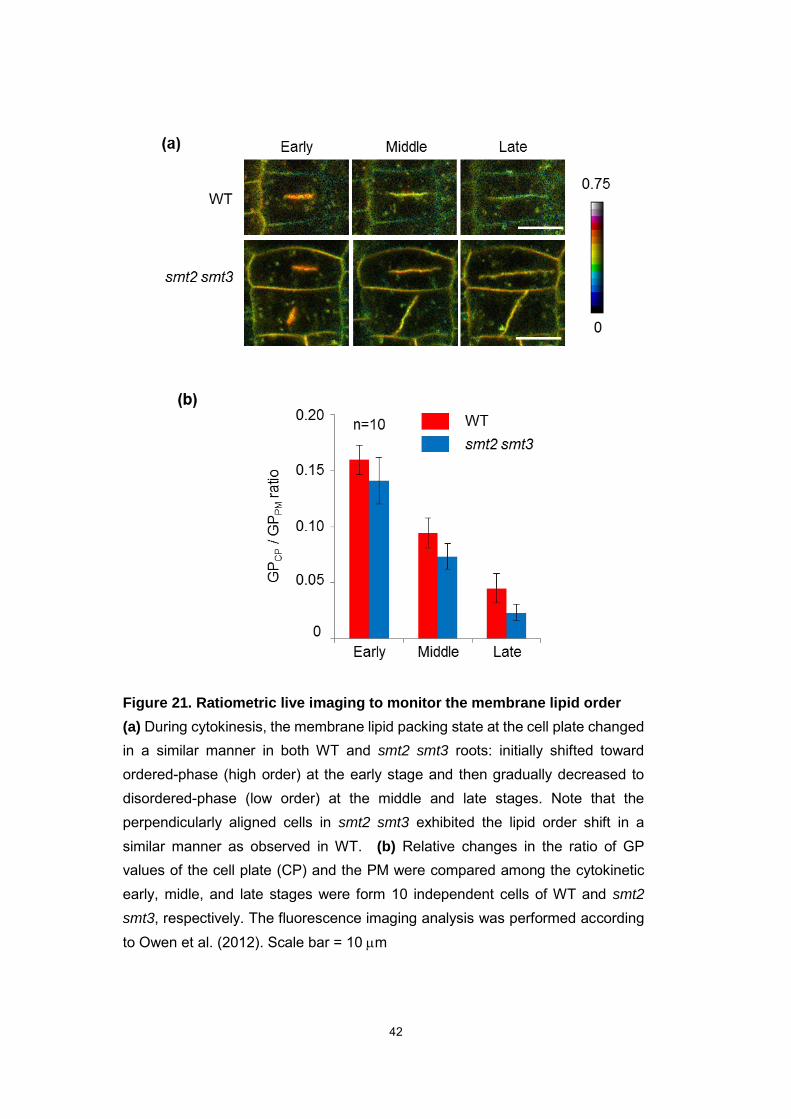
Figure 21. Ratiometric live imaging to monitor the membrane lipid order
(a) During cytokinesis, the membrane lipid packing state at the cell plate changed in a similar manner in both WT and smt2 smt3 roots: initially shifted toward ordered-phase (high order) at the early stage and then gradually decreased to disordered-phase (low order) at the middle and late stages. Note that the perpendicularly aligned cells in smt2 smt3 exhibited the lipid order shift in a similar manner as observed in WT. (b) Relative changes in the ratio of GP values of the cell plate (CP) and the PM were compared among the cytokinetic early, midle, and late stages were form 10 independent cells of WT and smt2
smt3, respectively. The fluorescence imaging analysis was performed according to Owen et al. (2012). Scale bar = 10 m

43
Chapter2
Clarification of the disturbed polar localization of auxin efflux protein
smt2 smt3 plants are impaired in the polar localization and endocytic recycling of
the PIN2 auxin efflux transporter protein
In WT plants, the PIN2 auxin efflux transporter localized correctly, maintaining the
downward and upward directions in the epidermis and the cortex cells, respectively
(Carland et al., 2010; Blilou et al., 2005). Multiple mislocalization patterns of PIN2-GFP
proteins were evident in smt2 smt3 on the apical side, basal side, and on the
disconnected cell wall (Fig. 11a). PIN2-GFP protein levels were lower in smt2 smt3 than
WT roots (Fig. 11b), and GFP-positive granular structures were observed in smt2 smt3
root cells (Fig. 11c). Immunohistochemical analysis revealed PIN2-GFP protein
mislocalization and the PIN2-GFP protein mislocalization was associated with abnormal
cell divisions (Fig. 22). Thus, these PIN2 mislocalization patterns were ascribed to
failure of cell division in smt2 smt3. Furthermore, the polar distribution of the PIN2
protein was not appropriately established in the epidermal cells of smt2 smt3 roots (Fig.
11). The correct PIN2 localization at the late cytokinesis stage (Fig. 12) and the cell
plate (Fig. 22) suggest that PIN2 protein trafficking operates normally in smt2 smt3.
It has been demonstrated that the intracellular asymmetric PIN2 distribution is a
controlled through a sterol-dependent clathrin-mediated endocytosis from the PM and
polar recycling to the PM as a post-cytokinetic event (Men et al., ellular 2008;
Dhonukshe et al., 2008; Blilou et al., 2005). Initial PIN2 internalization can be studied by
inhibiting endocytotic vesicular transport with brefeldin toxin A (BFA) (Xu and Scheres,
2005; Kleine-Vehn et al., 2008) in the presence of cycloheximide (CHX). The BFA
treatment induced the aggregate formation of PIN2-GFP and the endocytic marker

44
FM4-64 fluorescence within the cells (BFA compartments), indicating that the
endocytosed endomembrane aggregated where sterols co-accumulate with an
endocytic marker ARA6 (a Rab5-GTPase homolog) and PIN2 in Arabidopsis (Grebe et
al., 2003). FM4-64 pulse labeling demonstrated that the fluorescence signals of
PIN2-GFP and FM4-64 on the PM decreased with time in both the WT and smt2 smt3
root epidermal cells (Fig. 23). Compared with the WT, PIN2-GFP was internalized more
rapidly in smt2 smt3 (Fig. 23b), and prolonged residence of the BFA compartments was
apparent (Fig. 23a). By contrast, FM4-64 internalization was more rapid in the WT than
in smt2 smt3 (Fig. 23c). These observations are in clear contrast to the total endocytotic
inhibition observed in cpi1-1 plants (Men et al., 2008). The GFP-positive but
FM4-64-negative structures (Fig. 23a) may correspond to the PIN2-GFP-positive
particles present in smt2 smt3 before drug treatment (Fig. 11c, white arrows). Upon
BFA-washout, the BFA compartments in WT plants gradually disappeared (Fig. 24a),
whereas those in smt2 smt3 mutants remained within the cells, with a number of small
GFP-positive vesicles becoming apparent (Fig. 24a), and the PIN2-GFP signal on the
PM was dramatically reduced in smt2 smt3. These observations indicate that the
endocytosis and vesicular recycling are disturbed in smt2 smt3 plants, leading to the loss
of PIN2-GFP polar localization (Fig. 24). However, the disturbed endocytosis in smt2
smt3 roots was not restored by the addition of exogenous -sitosterol (Fig. 25). We
cannot rule out the possibility that exogenous -sitosterol may be unable to restore
endocytic recycling in the presence of the protein synthesis inhibitor CHX.
The unusual patterns of tubulin-GFP and ABD2-GFP (Fig. 26, 28) in smt2 smt3
roots suggested a possible involvement of C24-ethylsterols in the orientations of
cytoskeletal proteins. Early endocytic sterol transport has been demonstrated to depend
on actin (Grebe et al., 2003). The lack of efficient maintenance of PIN2 protein levels in
smt2 smt3 (Fig. 11b) may be due to disturbed endocytic recycling (Fig. 24). The

45
retromer component SNX1 (SORTING NEXIN 1) is involved in the endocytic recycling
of PIN2 to the PM (Jaillais et al. 2006; Kleine-Vehn et al. 2008). Ambrose et al. (2013)
demonstrated the involvement of microtubules (MTs) in PIN2 recycling through the
interaction of the MT-associated protein CLASP with SNX1. The aberrant localization of
GFP–CLASP as well as unusual MT orientations in smt2 smt3 (Fig. 26 and 27) suggest
that the retromer function is interrupted under C24-ethylsterol-deficient conditions,
leading to the retardation of PIN2-GFP recycling and PIN2 mislocalization.

46
Figure 22. Immunostaining of the PIN2 protein
PIN2-GFP localization was visualized by whole mount immunostaining using anti-GFP antibodies. The normal polarity of the PIN2-GFP localization (white arrows) was frequently lost in the smt2 smt3 root epidermal cells (blue arrows). The PIN2-GFP protein was detected at the division plane in the smt2 smt3 root as well as in the WT (white triangles), suggesting that the post-cytokinetic polar distribution of PIN2-GFP was disrupted in smt2 smt3. Abnormal PIN2 localization was also associated with perpendicular cell division (blue triangle). Scale bars = 10 m.

47
Figure 23. Internalization in the root epidermal cells
(a) Five-day-old seedlings were incubated for 30 min in 0.5× MS medium containing 50 M cycloheximide and stained for 10 min with 50 g/ml FM4-64 in the presence of 50 M CHX and 50 M brefeldin A (BFA). The tissue fluorescence images were recorded at indicated times (after BFA treatment). Endocytic PIN2-GFP internalization was not inhibited in the smt2 smt3 root epidermal cells, whereas PIN2-GFP signals became prominent over time in the intracellular space in smt2 smt3. GFP-positive structures without a FM4-64 signal were also observed in smt2
smt3.(b, c) The relative changes in the GFP and FM4-64 fluorescence intensities are presented in (b) and (c), respectively. Vertical axes show the relative changes in the fluorescence intensities from the PM (WT_PM, smt2 smt3_PM) and the intracellular space (WT_In, smt2 smt3_In). Fluorescence intensities were measured in 10 independent cells from 4 different seedlings, and the average values and standard errors are shown in (b) and (c) Scale bar = 10 m
48
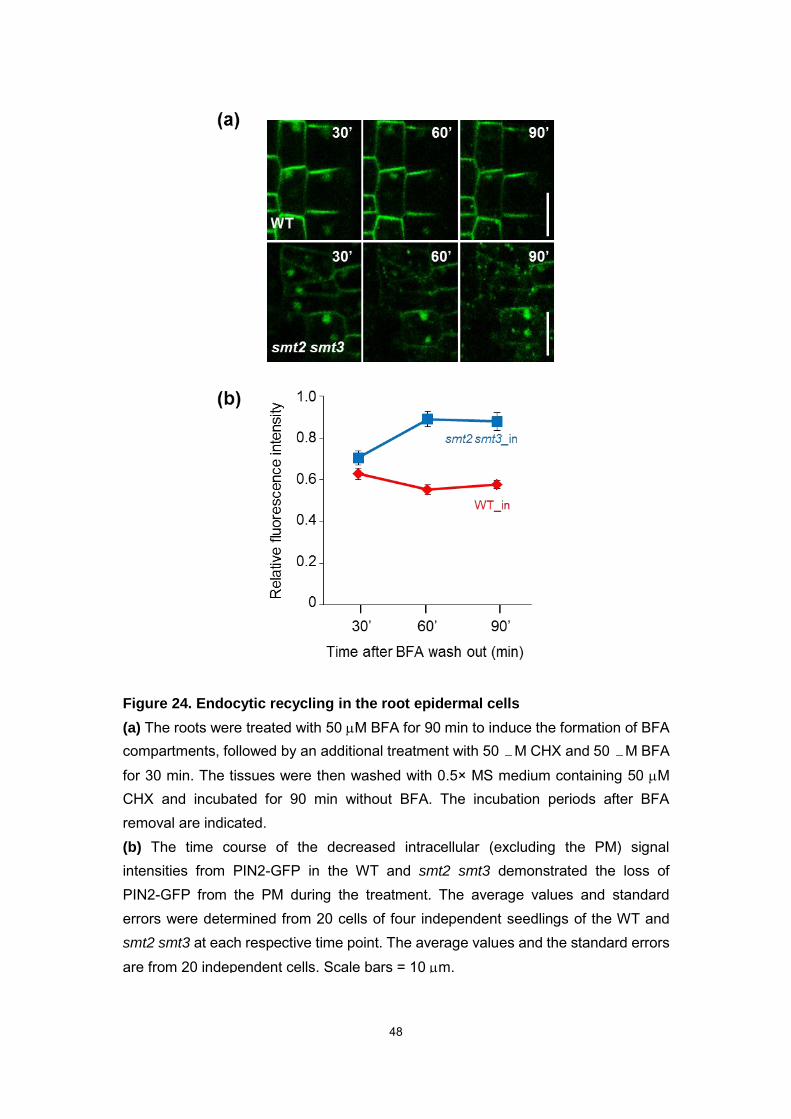
Figure 24. Endocytic recycling in the root epidermal cells
(a) The roots were treated with 50 M BFA for 90 min to induce the formation of BFA compartments, followed by an additional treatment with 50 M CHX and 50 M BFA
for 30 min. The tissues were then washed with 0.5× MS medium containing 50 M CHX and incubated for 90 min without BFA. The incubation periods after BFA removal are indicated.
(b) The time course of the decreased intracellular (excluding the PM) signal intensities from PIN2-GFP in the WT and smt2 smt3 demonstrated the loss of PIN2-GFP from the PM during the treatment. The average values and standard errors were determined from 20 cells of four independent seedlings of the WT and smt2 smt3 at each respective time point. The average values and the standard errors are from 20 independent cells. Scale bars = 10 m.
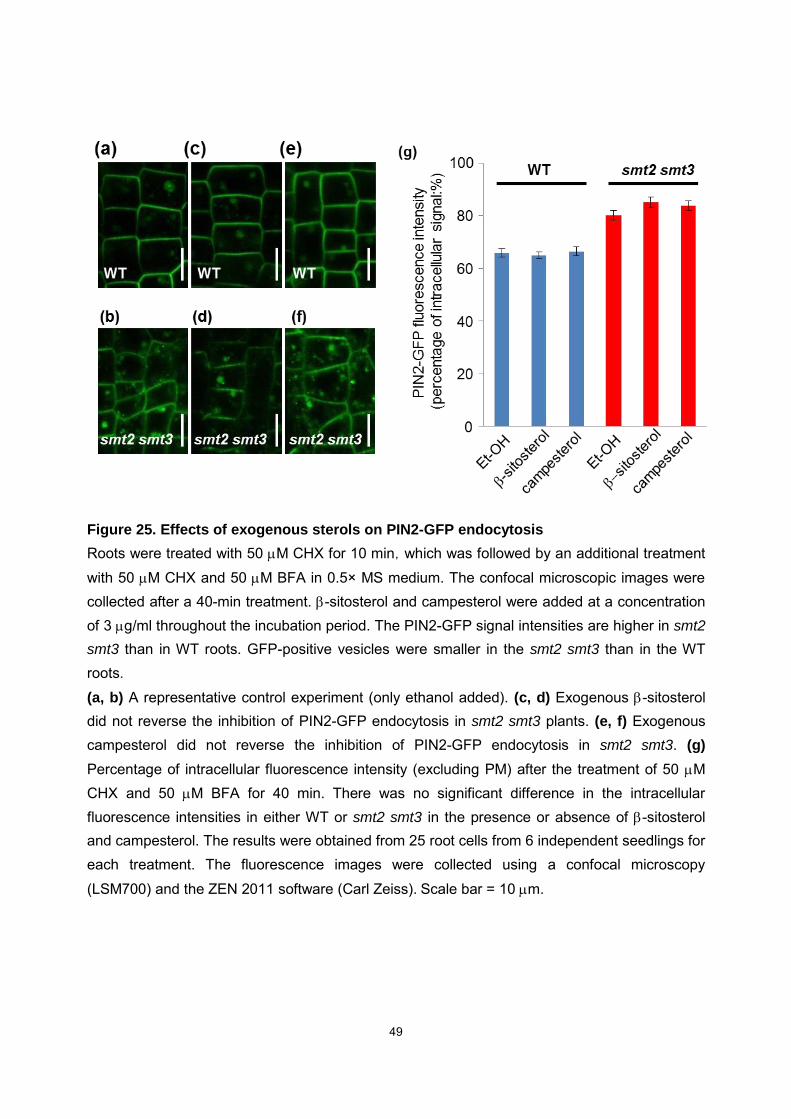
49
Figure 25. Effects of exogenous sterols on PIN2-GFP endocytosis
Roots were treated with 50 M CHX for 10 min,which was followed by an additional treatment with 50 M CHX and 50 M BFA in 0.5× MS medium. The confocal microscopic images were collected after a 40-min treatment. -sitosterol and campesterol were added at a concentration of 3 g/ml throughout the incubation period. The PIN2-GFP signal intensities are higher in smt2
smt3 than in WT roots. GFP-positive vesicles were smaller in the smt2 smt3 than in the WT roots.
(a, b) A representative control experiment (only ethanol added). (c, d) Exogenous -sitosterol did not reverse the inhibition of PIN2-GFP endocytosis in smt2 smt3 plants. (e, f) Exogenous campesterol did not reverse the inhibition of PIN2-GFP endocytosis in smt2 smt3. (g) Percentage of intracellular fluorescence intensity (excluding PM) after the treatment of 50 M CHX and 50 M BFA for 40 min. There was no significant difference in the intracellular fluorescence intensities in either WT or smt2 smt3 in the presence or absence of -sitosterol and campesterol. The results were obtained from 25 root cells from 6 independent seedlings for each treatment. The fluorescence images were collected using a confocal microscopy
(LSM700) and the ZEN 2011 software (Carl Zeiss). Scale bar = 10 m.
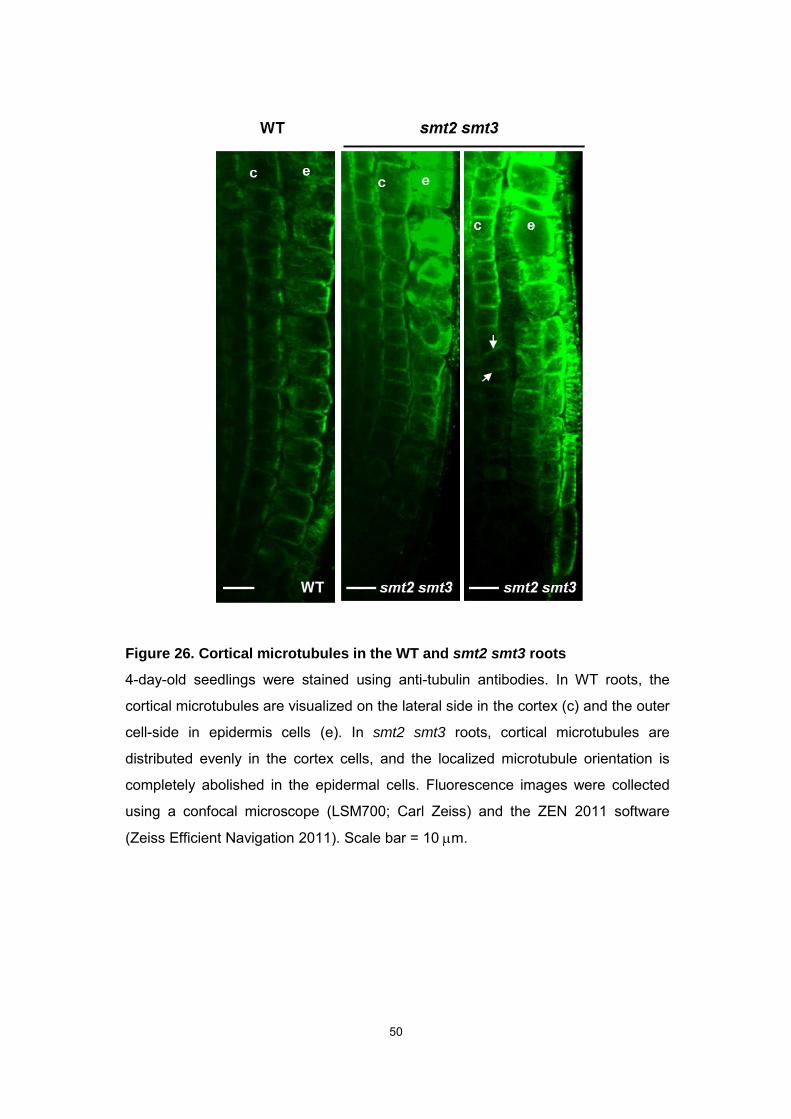
50
Figure 26. Cortical microtubules in the WT and smt2 smt3 roots
4-day-old seedlings were stained using anti-tubulin antibodies. In WT roots, the
cortical microtubules are visualized on the lateral side in the cortex (c) and the outer
cell-side in epidermis cells (e). In smt2 smt3 roots, cortical microtubules are
distributed evenly in the cortex cells, and the localized microtubule orientation is
completely abolished in the epidermal cells. Fluorescence images were collected
using a confocal microscope (LSM700; Carl Zeiss) and the ZEN 2011 software
(Zeiss Efficient Navigation 2011). Scale bar = 10 m.
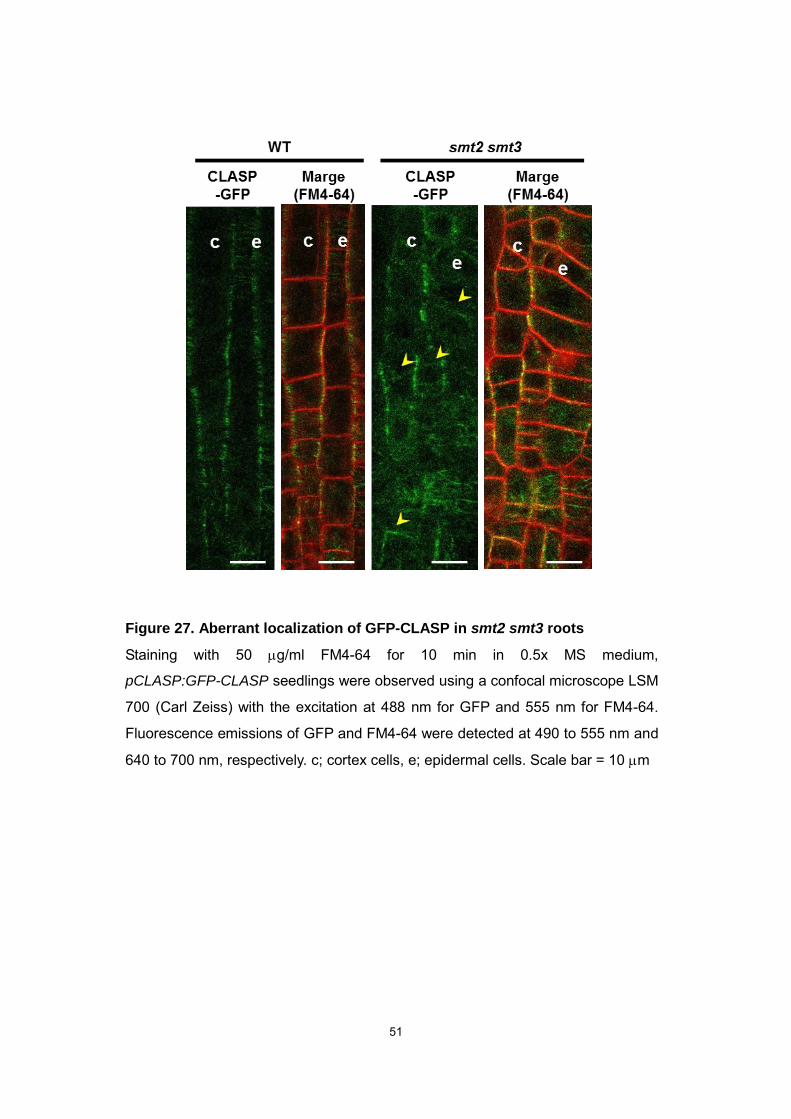
51
Figure 27. Aberrant localization of GFP-CLASP in smt2 smt3 roots
Staining with 50 g/ml FM4-64 for 10 min in 0.5x MS medium,
pCLASP:GFP-CLASP seedlings were observed using a confocal microscope LSM
700 (Carl Zeiss) with the excitation at 488 nm for GFP and 555 nm for FM4-64.
Fluorescence emissions of GFP and FM4-64 were detected at 490 to 555 nm and
640 to 700 nm, respectively. c; cortex cells, e; epidermal cells. Scale bar = 10 m
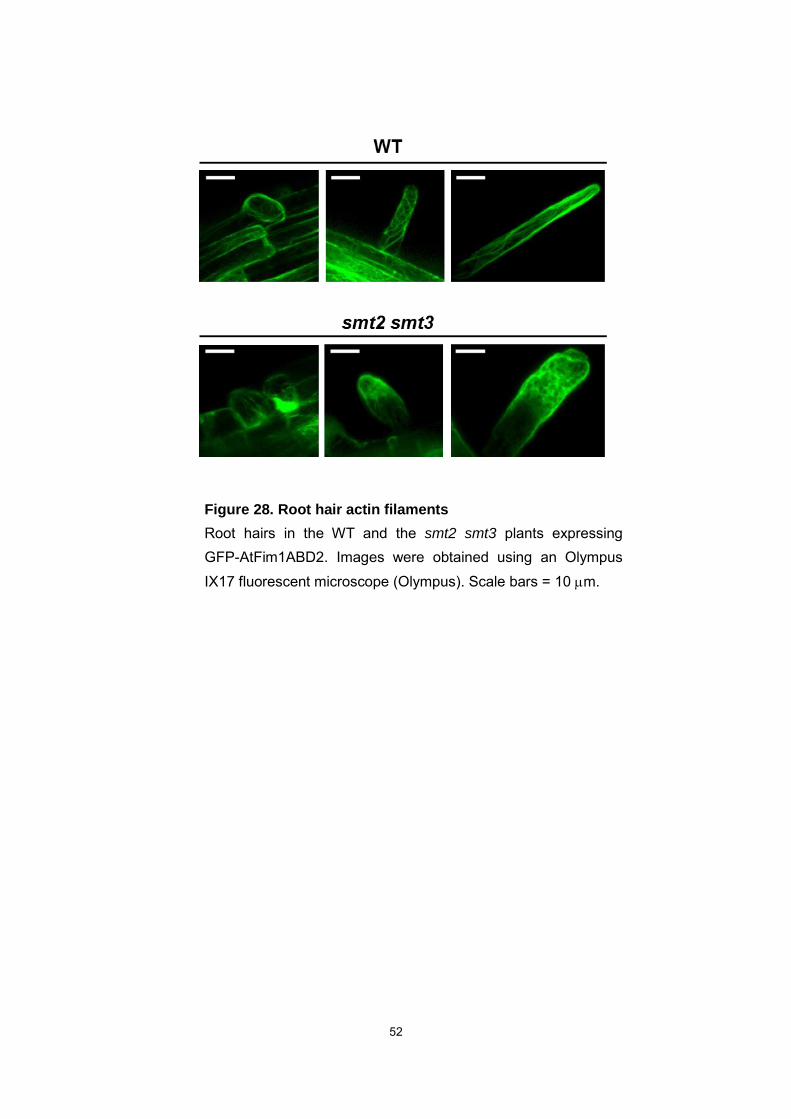
52
Figure 28. Root hair actin filaments
Root hairs in the WT and the smt2 smt3 plants expressing GFP-AtFim1ABD2. Images were obtained using an Olympus IX17 fluorescent microscope (Olympus). Scale bars = 10 m.

53
Chapter 3
General discussion
Sterols serve as essential membrane constituents, and therefore, there is no doubt
that the lack of major sterols or the abnormal sterol compositions that result from
genetic defects affect physicochemical properties of membranes and normal functions
of membrane proteins, thereby influencing diverse aspects of developmental processes
in eukaryotic life. Ample evidence supports the role of plant sterols in the generation of
the auxin concentration gradients achieved through the directional transport of auxin by
carrier proteins (Dhonukshe et al., 2008; Petrásek and Friml, 2009; Feraru and Friml,
2008). It has also been reported that sterols are required for correct auxin and ethylene
crosstalk to achieve vascular development (Pullen et al., 2010). Furthermore, disrupted
cell divisions in the sterol biosynthetic mutants smt1 (Schrick et al., 2004; Schrick et al.,
2002) and cpi1-1 (Men et al., 2008), or ectopic cell division leading to abnormal vascular
patterning in the hydra1 and fackel/hydra2 mutants reinforce the implication of sterols in
major cellular processes (Pullen et al., 2010). However, the possible relationships
between given plant sterol molecular species, cell division, and phytohormones have
not been clearly deduced from studies mentioned above (Fig. 1). In this report, we show
the essentiality of C24-ethylsterols using the Arabidopsis smt2 smt3 mutant that is
defective in the Charophycean green algae and land plant specific sterol
methyltransferase and that bear therefore a strongly modified distribution of
24-alkyl-5-sterols (Fig. 1). We present evidence that C24-ethylsterols are involved in
the endocytic recycling and the polarity control of the PIN2 protein as well as the

54
determination of the cell division plane to establish polarized growth.
Cell division
In smt2 smt3 roots, aberrant cell division plane formation was observed at the step
of PPB formation, and collapsed cell division was accompanied by the uncoordinated
localizations of KNOLLE and tubulin to the cell division plane. However, once PPB
formation occurred correctly, cytokinesis appeared to proceed normally. Thus,
C24-ethylsterols may be required for the determination of the cell division plane, which
is controlled by the consecutive events comprised of the positioning, anchoring, and
guidance of the division zone (Van Damme, 2009). The root culture experiments
demonstrated that a small amount of -sitosterol (or stigmasterol) critically promoted the
recovery of abnormal secondary root growth of smt2 smt3 roots. However, we were not
able to identify metabolites in very low amounts derived from the exogenously supplied
-sitosterol in the cultured roots (Fig. 18), and we cannot rule out the possibility that
-sitosterol may be further modified to yield novel bioactive sterol structures. We have
not eliminated the possibility that the exogenously supplied -sitosterol may have
accumulated at an extremely specific site, such as the membrane microdomain in plants
(Malinsky et al., 2013). Because the intracellular trafficking of the ABCB19 auxin
transporter in Arabidopsis was interrupted within 2 h of the treatment of fenpropimorph
(Yang et al., 2013), suggesting that spatiotemporal sterol synthesis, rather than total
sterol composition, may be required for endomembrane trafficking. The observation of
lipid order of cell plate demonstrated that the lipid order in smt2 smt3 shifted toward a
higher order, as observed in the WT, irrespective of correct or failed cytokinesis (Fig. 21).
By contrast, the GP values recorded in smt2 smt3 roots were generally higher than

55
those in the WT, regardless of cytokinetic stage, suggesting intrinsically different
membrane properties between the WT and smt2 smt3 (Fig. 21). The
C24-ethylsterol-deficient membrane in smt2 smt3 may represent an actual case of
membrane organization alteration by different phytosterol compositions in a model
system (Grosjean et al., 2015). Although the total disruption of sterol biosynthesis led to
the failure of normal cell plate formation via a sterol- and DRP1A-dependent mechanism
(Frescatada-Rosa et al., 2014), the vesicle transport responsible for cell plate formation
may not be reliant on C24-ethylsterols. In addition, KNOLLE maintenance in the plane
of cell division is also dependent on a sterol- and DRP1A-dependent mechanism, which
was not affected in smt2 smt3 (depletion of C24-ethylsterols) but is inhibited when sterol
biosynthesis is totally abolished (Boutté et al., 2010). These results indicate that the
totally disrupted sterol composition and the depletion of C24-ethylsterols may have
differentially influenced the processes of cell division.
PIN2 polarity and auxin transport
In plants, mutations in the sterol biosynthetic pathway (Fig. 1) generally affect polar
auxin transport. In fact, the mutants of smt1/cph/orc have reduced polar auxin transport
due to the mislocalization of PIN1 and PIN3 (Willemsen et al., 2003), and the cpi1-1
mutation results in the loss of polar localization of PIN1 and PIN2 (Men et al., 2008). The
interference of polar auxin transport in the sterol mutants are explainable by the
involvement of sterols in the cell polarity control and endosomal vesicle trafficking (Men
et al., 2008; Feraru and Friml, 2008; Grebe et al., 2003). The unaffected PIN1 and PIN2
polarity in sterol mutants such as hyd1 and hyd2/fk (Souter et al., 2002; Pullen et al.,
2010) and the unchanged polar auxin transport in cvp1/smt2 (Carland et al., 2002) may

56
implicate multiple roles of sterols in cellular events. Our results with smt2 smt3 mutant
indicate that C24-ethylsterols are involved in the control post-cytokinetic PIN2 polarity
and auxin distribution. The PIN2-GFP protein level was significantly lower in smt2 smt3
than that in the WT (Fig. 11b). The endocytosis experiments indicate that the substantial
decrease in the level of PIN2-GFP at the PM in smt2 smt3 was attributable, at least in
part to inhibited PIN2-GFP recycling (Fig. 24). Although auxin inhibits PIN endocytic
internalization to increase PIN protein levels at the PM, thus facilitating auxin efflux
(Paciorek et al., 2005), the lower PIN2 protein levels observed in smt2 smt3 are not
consistent with a potential increase in tissue auxin levels. The initial internalization of
PIN2-GFP and the endocytic tracer FM4-64 from the PM was not inhibited but rather
slightly facilitated in smt2 smt3. Interestingly, our results are not consistent with the
strong inhibition of the internalization of both FM4-64 and PIN2 in dividing and
post-cytokinetic root epidermal cells of the cpi1-1 mutant, an upstream sterol
biosynthetic mutant exhibiting a severely altered sterol composition (Men et al., 2008).
The intracellular asymmetric PIN2 distribution, which is essential for the directional
transport of auxin, is accomplished by clathrin-mediated endocytosis from the PM and
subsequent polar recycling as a post-cytokinetic event (Blilou et al., 2005; Xu and
Scheres, 2005; Kleine-Vehn and Friml, 2008; Men et al., 2008). PIN2 was localized to
the PM just after cytokinesis (Fig. 12), implying that the polar distribution of PIN2 may be
impaired in smt2 smt3. The interaction of CLASP with SNX1 stabilizes SNX1
endosomes, such as PIN2-carrying SNX1 vesicles on MTs that prevent PIN2
degradation (Ambrose et al., 2013). Thus, the aberrant localization patterns of CLASP
and MTs (Fig. 26, 27) suggest the involvement of C24-ethylsterols in PIN2 endocytic
recycling mediated by cytoskeletal proteins. Both the organization of the MTs and the

57
actin cytoskeletons were broadly affected in smt2 smt3 roots (Fig. 26, 28). Several lines
of evidence support the possible link between the smt2 smt3 phenotype and cytoskeletal
proteins.
Appropriate auxin concentration gradients are a prerequisite for normal
development. Auxin transport may be affected by a class of 4-methyl sterol derivatives
that accumulate upon disruption of the sterol-C4-demethylation enzymatic complex
(Mialoundama et al., 2013). Arabidopsis ERG28 acts as the scaffolding platform for the
functional complex of the sterol C4 demethylation enzymes (Gachotte et al., 2001;
Darnet and Rahier, 2004). Arabidopsis erg28 mutants exhibit serious growth
abnormalities associated with unusually high levels of CMMC, the substrate of the
carboxysterol-C3-dehydrogenase/C4-decarboxylase of the C4-demethylation complex
(Rahier et al., 2006). In the erg28 mutants, the accumulated CMMC is thought to inhibit
polar auxin transport without affecting the endocytosis-dependent control of the PIN1
protein localization (Mialoundama et al., 2013). The sterol substrate of SMT2, namely,
24-methylene lophenol, is also a substrate of the C4-demethylation complex (Fig 1).
Although smt2 smt3 mutant tissues contained small amounts of 24-methylene lophenol,
as well as other 4-methyl sterols (cycloartenol, cycloeucalenol), we did not detect
4-carboxy-4-methyl derivatives in smt2 smt3 seedlings (Fig. 5). Therefore, the
accumulation of the so-called cryptic sterol biosynthetic intermediates (Mialoundama et
al., 2013) appears not to be a factor in smt2 smt3 mutants. In addition, in smt2 smt3
plants, 24-methylene lophenol accumulated to a level comparable with that detected in
the smt2 mutant (Table 1), which exhibited only minor phenotypic defects of leaf
vascular development and petal morphology (Fig. 6b, 6c). Thus, the current
experimental observations in the smt2 smt3 mutant, such as the interference of the

58
endocytosis-dependent recycling of the PIN2 protein and the defective cell division, do
not conform to the suggested involvement of CMMC in the inhibition of polar auxin
transport mediated by ABCB1 and ABCB19 auxin efflux transporters (Mialoundama et
al., 2013).
SMT genes and C24-ethylsterols
We have demonstrated that -sitosterol, rather than sterol composition, plays a
critical role in plant development. -sitosterol differs from campesterol by the presence
of one additional methyl group in the side chain, which is added by the SMT2 specific to
Charophycean green algae and land plants (Neelakandan et al., 2009) (Fig. 2).
Concerning the side chain stereochemistry, land plants may produce (24R,
24S)-24-alkylsterols (McKean and Nes, 1977). The epimeric mixture of
24-methylcholesterol (24R, campesterol; 24S, dihydrobrassicasterol) was shown to be
ubiquitous whereas 24-ethylsterols are exclusively (24R)-24-ethylsterols such as
sitosterol and stigmasterol (Nes et al., 1977; Guo et al., 1995; Schaller, 2003) (Fig. 2)
with the exception of a few plant genus able to produce limited amounts of
(24S)-24-ethylsterols however restricted in the plant lifespan (Fenner and Patterson,
1992). The occurrence of C24-ethylsterols is not restricted to land plants. For example,
Chlamydomonas synthesizes and accumulates ergosterol as the major sterol together
with a C24-ethylsterol, 7-dehydroporiferasterol, as a minor component, and a single
sterol methyltransferase gene is thought to be responsible for both the C24-methylation
and subsequent C28-methylation reactions (Desmond and Gribaldo, 2009). Volvox and
Coccomyxa (a unicellular green algae) contain single SMT genes (Fig. 2); these algal
species may also contain a C24-ethylsterol as reported for Chlamydomonas.

59
Biosynthetic routes leading to these (24S)-24-alkylsterols have been reported in details
(Nes et al., 1990) as well as the identical features of yeast and algal sterol pathways
(Salimova et al., 1999). By contrast, sequence comparisons have suggested that
Charophycean green algae, which are thought to be most closely related to land plants
(Sørensen et al., 2012; Timme et al., 2012), contain two types of SMT genes
responsible for the two-step methyl transfer reactions similar to those operating in land
plants (Fig. 1 and 2). It is thus possible that -sitosterol is produced in these multicellular
Charophycean algae.
We propose that the acquisition of the two-step SMT reactions enabled the
production of -sitosterol, which was recruited to control cell division (correct formation
of PPB and phragmoplast) and the endocytotic recycling of PIN2 to generate an auxin
concentration gradient through directional auxin transport, and thus contributed to the
evolution of polarized plant growth. In addition, our results suggest different sterol
requirements among vesicle trafficking pathways, such as the secretion of newly
synthesized PIN proteins to the PM, endocytosis from the PM, and subsequent polar
endocytic recycling. Importantly, our results indicate that the presence of a very low
amount of -sitosterol, not the normal sterol composition, is critical for plant
developmental processes. Clarification of the spatiotemporal localization of -sitosterol
and its possible binding site, as well as the possible conversion of -sitosterol into an
unknown sterol-derived signaling molecule, should provide insights into the unique role
of this sterol in plants.

60
Supplementary information
Materials and methods
Plant growth conditions
All experiments were carried out using Arabidopsis thaliana, ecotype Columbia
(Col-0). Arabidopsis plants were grown in a growth chamber maintained at 23°C under
continuous light (140–160 μmol m-2s-1). Seeds were sterilized with 70% ethanol (1 min)
and 0.8% sodium hypochlorite (10 min), and then thoroughly washed with sterilized
water. Plants in soil were supplied with a culture solution composed of 1.0 mM NH4NO3,
2.5 mM KNO3, 0.5 mM KH3PO4, 0.5 mM CaCl2·2H2O, and 0.5 mM MgSO4·7H2O
supplemented with micronutirents (50 M H3BO3, 20 M MnCl2·4H2O, 20 M
CuSO4·7H2O, 2 M ZnSO4·7H2O, 0.5 M Na2MoO·4H2O, 100 M Fe-EDTA). For
experiments under sterile conditions, seeds were germinated on agar (0.8%) plates
containing a germination medium (GM) composed of 1× Murashige and Skoog (MS)
salts and 1% (w/v) sucrose (Valvekens et al., 1998).
Transgenic plants
The T-DNA insertion mutant lines of smt2 (GABI_443_F03) and smt3
(SALK_085292) were obtained from the Arabidopsis Biological Resource Center
(ABRC) (Alonso et al., 2003). Homozygotes with regard to the T-DNA insertion events
(smt2/smt2 and smt3/smt3) were identified by polymerase chain reaction (PCR) using
gene-specific primers and T-DNA-specific primers. The homozygous plants of
smt2/smt2 and smt3/smt3 were crossed with each other to establish a double mutant
line of +/smt2-smt3/smt3 for self-pollination to generate smt2/smt2-smt3/smt3 (smt2
smt3) plants. Since the smt2 smt3 plants were self-sterile, the current experiments were

61
performed by maintaining +/smt2-smt3/smt3 plants for self-pollination to obtain smt2
smt3 plants. smt2 smt3 seedlings were easily identified by their abnormal phenotype
(Fig. 8a). Homozygous plants for each T-DNA insertion event were identified by means
of PCR screening and segregation analyses. PCR was performed using primers
designed from the coding sequences for SMT2 and SMT3 (Table 3). For confirmation of
the smt2 mutation, an SMT2 gene-specific primer (SMT2F) and a reverse primer
(SMT2R) together with a primer for T-DNA sequence (GABI LB) were used. For the
smt3 mutation, an SMT3-specific reverse primer (SMT3R) and a forward primer
(SMT3F) together with a primer for T-DNA (LBa1) were used. A DR5:GUS expression
line of A. thaliana (Ulmasov et al., 1997) was crossed to the T-DNA insertion mutant
plants (smt2/smt2, smt3/smt3, and +/smt2-smt3/smt3). The Arabidopsis line
homozygous for the expression cassette of PIN2-GFP (Blilou et al., 2005) was crossed
to the mutant lines (+/smt2-smt3/smt3). The +/smt2-smt3/smt3 plants carrying either the
DR5:GUS cassette or the PIN2-GFP cassette were maintained. A tubulin-GFP
expression line and an actin-GFP line were generated by the transformation of A.
thaliana plants using the expression plasmids containing a coding sequence for either
GFP-NtTub fusion protein (Kumagai et al., 2001) or GFP-AtFim1ABD2 (Sano et al.,
2005) by the floral dip method (Clough et al., 1998). T1 seeds were germinated on GM
agar plates containing 25 g/ml kanamycin. After selecting T2 homozygous lines, T3
plants were crossed to the +/smt2-smt3/smt3 line to obtain F1 seeds, which were
allowed self-pollination to yield F2s (tubulin-GFP expression line of +/smt2-smt3/smt3
and actin-GFP expression line of +/smt2-smt3/smt3). The +/smt2-smt3/smt3 F2 plants
(expressing either tubulin-GFP or actin-GFP) were allowed self-pollination to generate
smt2/smt2-smt3/smt3 F3 plants expressing tubulin-GFP or actin-GFP. The presence of

62
the coding sequences of the GFP-fusion proteins was analyzed by PCR using a primer
set of 35sFw for the CaMV35S promoter and sGFPRv for the GFP sequence (Table 3).
RT-PCR
Reverse-transcription (RT)-PCR analyses were carried out to determine the
expression levels of SMT2 and SMT3 genes in Arabidopsis using the gene-specific
primer sets (Table 3). Arabidopsis actin gene was amplified under the same PCR
condition using a primer pair of Act2F and Act2R as the internal control (Table 3). Total
RNA was isolated using an RNeasy Plant Mini Kit (QIAGEN). Genomic DNA
contamination was eliminated using an RNase free DNase Set (QIAGEN, Tokyo,
Japan). First-strand cDNA was synthesized using a Takara RNA PCR Kit (AMV) Ver.3.0.
(Takara, Kyoto, Japan) in a 10 l reaction mixture containing 500 ng of total RNA using
an oligo(dT)16 as the reverse primer. The RT reactions were carried out at 30°C for 10
min, 50°C for 30 min, and 95°C for 5 min, and then chilled to 5°C for 5 min. The PCR
was done using 2 l of the RT products as the template in a 10 l of reaction mixture
containing 1 mM MgCl2, 0.2 mM dNTP mixture, 0.025 unit/l Takara Ex Taq HS
(Takara), and 0.2 M of primers using a Takara RNA PCR Kit (AMV) Ver.3.0 (Takara).
The PCR was programmed in 30 cycles of 94°C for 30 s, 50°C for 30 s, and 72°C for 1
min followed by an extension period of 10 min at 72°C.
GUS Staining of transgenic plants
Embryos and five-day-old seedlings were treated with 90% (v/v) ice-cold acetone
for 1 h and submerged in GUS staining buffer (Jefferson et al., 1987) containing 100
mM sodium phosphate (pH 7.0), 10 mM EDTA, 0.5 mM potassium ferrocyanide, 0.5 mM

63
potassium ferricyanide, 0.1% (w/v) Triton X-100, and 0.5 g/ml
5-bromo-4-chloro-3-indolyl--D-glucuronide (Nacalai Tesque Inc., Kyoto Japan). The
tissues were then infiltrated with staining buffer under vacuum and incubated overnight
at 37°C. After rinsing with water, tissues were cleared with chloral hydrate:distilled
water:glycerol (8:2:1, v/v), followed by washes using 90, 70, and 50% (v/v) diluted
ethanol.
Lugol staining
Starch granules in the columella root cap were visualized by the staining using 1%
Lugol’s solution (Nacalai Tesque Inc.) (Willemsen et al., 2003). Seedlings were stained
for 3 min, rinsed with water, and cleared with chloral hydrate. Microscopic images were
collected using an IX71 system (Olympus, Tokyo, Japan).
Sterol analysis
Samples (50 mg) were frozen and homogenized in liquid N2, and total sterol
fractions were extracted for 30 min at room temperature in 5 ml of chloroform/methanol
(1:2, v/v). By adding 2 ml of 1% (w/v) KCl and 1 ml of chloroform, sterols were recovered
in the organic phase. Two milliliters of methanol/H2O (10:9, v/v) were then added to the
organic phase and was evaporated to dryness under N2 stream. The saponification was
done in 2.5 ml of 1 M KOH in methanol at 90oC for 1h, and 2 ml chloroform and 2.5 ml
H2O were added to recover the organic phase. After adding 1.25 ml of 0.5 M KOH and 6
ml H2O, the organic phase was evaporated to dryness in vacuo and used for the
trimethylsilyl (TMS)-derivatization. Sterols were analyzed by gas chromatography-mass
spectrometry (GC-MS) on a Saturn 2200 GC/MS system (Varian Inc. Palo Alto, CA) with

64
a CP-SIL5 CB LOW Bleed/MS column (30 m×0.25 mm) (Varian). The port temperature
for split injections (a split ratio of 50%) was 270°C, and helium was used as the carrier
gas at a flow rate of 1.5 ml/min. The temperature program was started with a slow rise
from 230 to 285°C (2°C/min), and from 285 to 320°C (10°C/min), and finally 10 min at
320°C. Standards consisting of cholesterol, brassicasterol, campesterol, stigmasterol,
and -sitosterol were used for quantification and identification. The peak areas were
automatically calculated using a 2200 Workstation (Varian), and sterol amounts were
determined from the ratio of the peak area of each sterol to that of 5--cholestane as an
internal standard (Schaeffer et al., 2001; Dyer et al., 1995). The sterol structures were
identified by reference to relative retention time and mass spectra. The pattern of
fragment ions with the mass-to-charge (m/z) values of 484, 394, 255, and 129 were
attributed to stigmasterol, and the fragment ions (m/z = 470, 380, 365, and 129) were
used to identify brassicasterol/crinosterol. The 24-epimers (brassicasterol and
crinosterol) were not separately analyzed under our experimental conditions.
For analysis of sterol conjugates, cultured root samples (30 mg) were frozen and
homogenized in liquid N2, and sterols (including free sterol; FS, sterol fatty acid ester;
SE, sterol glucosides; SG and acylated steryl glucosides; ASG) were extracted with
dichloromethane/methanol (1:2, v/v). The FSs and sterol conjugates were separated by
TLC. The TLC plate was developed in solvent system, containing of
dichloromethane/methanol/water, 85:15:0.5 (v/v), FS fraction elutes at Rf 0.82-0.89, SE
fraction elutes at Rf 1.00, SG fraction elutes at 0.33, ASG fractions elutes at 0.61. The
plate was sprayed with 0.01% berberine in ethanol, and examined under UV light. The
separated individual bands were scraped and extracted using
dichloromethane/methanol (2:1). The saponification of SE fraction was done in 2 ml of

65
6% KOH in methanol at 70oC for 2h, and 1.5 ml hexane and 1 ml H2O were added to
recover the organic phase. Hydrolysis of SG and ASG fractions were done in 2 ml 1 %
sulfuric acid in ethanol at 70°C for 2 h and 1.5 ml hexane and 1 ml H2O were added to
recover the organic phase. Acetylation of 3’-OH-sterols in each fractions was done in 50
l toluene, 30 l acetic anhydride and 20 l pyridine at 70°C for 2 h. Sterols were
analyzed by GC-MS.
Chromatography and mass spectrometry of sterol derivatives using
UPLC-APPI+-MS
Plant lipids (including carboxysterols and glucoside conjugates) were extracted
with a mixture of dichloromethane/methanol (2:1, v/v). Samples of 3 l (of a total extract
of 200 l) were analyzed on an Acquity UPLC system (Waters Quattro Premier XE,
Waters, Mildorf, MA, USA) equipped with a diode array detector (DAD) and an
Atmospheric Pressure Photon Ionisation (APPI) source. UV spectra were recorded from
190 to 500 nm. Chromatographic separation was achieved using an Acquity UPLC BEH
C8 column (100 x 2.1 mm, 1.7µm; Waters) coupled to an Acquity UPLC BEH C8
pre-column (2.1 x 5 mm, 1.7µm; Waters). The column was operated at 46°C with a
flow-rate of 0.3 ml/min of the following mobile phases: solvent A (75% methanol in water
with 0.01% formic acid) and solvent B (99.99% isopropanol with 0.01% formic acid). The
separation (total run time of 48 min) started with 100% of solvent A hold for 2 min,
followed by 25 min of a gradient to reach 50% of solvent A and 50% of solvent B, and
then hold for 4 min. It was followed by a gradient to reach 100 % of solvent B in 8 min,
hold for 3 min. Then another 4 min gradient was applied to reach 100% of solvent A hold
for 3 min. The column was equilibrated with solvent A for 4 min before the next run. Both
of the drying gas and nebulising gas were nitrogen generated from pressurized air in an

66
N2G nitrogen generator (Mistral, Schmidlin-dbs-AG, Switzerland). The parameters for
the MS and MS/MS detection and APPI ionization were as follows. Nebulizer gas flow,
50 l/h; desolvation gas flow, 500 l/h; APPI probe temperature, 450°C; source
temperature, 120°C; capillary voltage, 1.5 kV; cone voltage after optimization for each
sterol derivative was set to 25V; positive ionization mode; low and high mass resolution,
15 for both mass analyzers; ion energies 1 and 2 were 0.2 and 1V; entrance and exit
potential, 30V; detector (multiplier) gain, 650V. The Selected Ion Recording (SIR) MS
mode was used to determine mass transitions from the parent ion [M+ H - H2O]:
4-carboxy-4-methyl-cycloartanol (CMC, m/z: 439.50) (Mialoundama et al., 2013),
4-carboxy-4-methyl-24-methylene cycloartanol (CMMC, m/z: 453.48) (Mialoundama et
al., 2013), sitosterol (m/z: 397.71), and sitosteryl glucoside [M+ H - glucose - H2O]: (m/z:
397.70). Measurements of sterol derivatives were based on specific chromatography
retention times and total parent ion counts by comparison with authentic standards.
Data acquisition and analysis were performed with the MassLynx software (version 5.1,
Windows XP professional, Pentium PC).
Protoplasts
Five-day-old seedlings germinated on GM agar plates were transferred to 0.5× MS
liquid medium and cultured for 20 days under 16 h light/8 h dark on a rotary shaker (100
rpm) at 24°C. The roots were cut to 5 to 10 mm in length and incubated for 5 days in a
secondary root-inducing medium consisting of 0.5× MS medium supplemented with 1%
(w/v) sucrose, 3.0 g/ml IBA (indolebutyric acid) 1.0 g/ml NAA (naphthaleneacetic acid)
on a rotary shaker (80 rpm). The secondary roots were induced in the presence or
absence of 3 g/ml phytosterols (-sitosterol, stigmasterol, or campesterol). Protoplasts
were prepared principally according to the methods described previously (Jaideep et al.,
1995; Yoo et al., 2007). Briefly, the induced secondary roots were incubated for 12 h in

67
0.5× MS liquid medium containing 0.4 M glucose, 1% (w/v) CELLULASE ONOZUKA
R-10 (Yakult Pharmaceutical Industry, Tokyo, Japan), 0.25% (w/v) MACEROZYME
R-10 (Yakult). Protoplasts were released from root tissues by incubating for 12 h on a
rotary shaker (80 rpm) and filtered through a 70 m mesh filter. The isolated protoplasts
were washed with 0.4 M mannitol and cultured in a glass- bottomed dish for 100 h in a
0.5× MS protoplast culture medium containing 0.4 M glucose, 2.0 g/ml 2,4-D, 0.5 g/ml
IAA and 6.25 g/ml meropenem hydrate. The protoplasts were stained with 50 g/ml
FM4-64 for the PM visualization. In order to monitor the cellulose accumulation,
protoplasts were stained with 1 g/ml calcofluor-white M2R (Sigma, Tokyo, Japan)
(Sato et al., 2001).
Endocytosis and endocytic recycling
Five-day-old seedlings were stained with 50 g/ml FM4-64 (Cosmo Bio, Tokyo,
Japan ) in 0.5× MS medium for 10 min on ice. After washing with 0.5× MS medium,
specimens were visualized using a confocal microscope (LSM 700). Five-day-old
seedlings were incubated for 30 min in 0.5× MS medium containing 50 mM
cycloheximide (CHX, Nacalai Tesque). Seedling specimens in 0.5× MS were mounted
on glass slides with coverslips and visualized using a confocal microscope (LSM700,
Carl Zeiss). The endocytic recycling inhibitor brefeldin A (BFA) induces the formation of
the BFA compartments consisting of aggregates derived from the vesicles and tubules
from the trans-Golgi network (Blilou et al., 2005; Geldner et al., 2001; Geldner et al.,
2003). After the pretreatment with 50 M CHX in 0.5× MS medium, the seedlings were
stained for 10 min with 50 g/ml FM4-64 in 0.5× MS medium containing 50 M CHX and
50 M BFA (Wako Pure Chemicals, Osaka, Japan). The seedlings were further
incubated in 0.5× MS medium containing 50 M CHX and 50 M BFA for 30 min. The

68
time course of the formation of BFA compartments was observed using a confocal
microscope LSM700 (Carl Zeiss). For the observation of endocytic recycling, the
seedlings were treated for 90 min with 50 M BFA in 0.5× MS medium, and further
incubated in 0.5× MS medium containing 50 M CHX and 50 M BFA for 30 min. The
seedlings were washed three times with 50 M CHX in 0.5× MS medium. The time
course of the PIN2-EGFP endocytic recycling was observed for 3 h using a confocal
microscope LSM 700 (Carl Zeiss) with the excitation at 488 nm for GFP and 555 nm for
FM4-64. Fluorescence emissions of GFP and FM4-64 were detected at 490 to 555 nm
and at 640 to 700 nm, respectively.
Immunolabeling
Whole-mount immunolabeling experiments principally followed the method
described previously (Men et al., 2008). Arabidopsis seedlings were fixed in 4%
paraformaldehyde for 1 h in MTSB (50 mM PIPES, 5 mM MgSO4, 5 mM EGTA, pH 7.0).
After washing 5 times for 5 min with 0.1% Triton X-100 in MTSB, seedlings were
extensively washed 5 times with sterilized H2O. For whole-mount immunolabeling, the
paraformaldehyde-fixed seedlings were treated with 2% (w/v) driselase (Sigma) for 45
min and washed with MTSB 5 times followed by 1 h incubation in MTSB containing 10%
(w/v) DMSO and 3% (w/v) NP-40. After washing with MTSB 5 times, specimens were
incubated in 2% (w/v) BSA in MTSB for 1 h and treated with primary antibodies in MTSB
containing 3% (w/v) BSA at 4°C for overnight. After washing with MTSB for 7 times,
secondary antibodies were applied at 37°C for 1.5 h and the samples were stained with
10 g/ml DAPI in MTSB for 10 min. Antibody dilutions were: rabbit anti-KNOLLE (Lauder
et al., 1997), 1:2000; mouse anti--tubulin IgG (Invitrogen), 1:500; rabbit anti-GFP

69
(Sigma), 1:1000; Alexa Fluor 488 goat anti-mouse IgG (Invitrogen), 1:500; Alexa Fluor
488 goat anti-rabbit IgG (Invitrogen), 1:500; Alexa Fluor 568 goat anti-mouse IgG
(Invitrogen), 1:500; and Alexa Fluor 568 goat anti-rabbit IgG (Invitrogen), 1:500. The
excitation wavelengths of DAPI, Alexa Fluor 488, and Alexa Fluor 568 were 405 nm,
488 nm, and 555 nm, respectively. Fluorescence detections were performed at 395 to
540 nm for DAPI, 490 to 635 nm for Alexa Fluor 488, and 569 to 700 nm for Alexa Flour
568 using a confocal microscope (LSM700) and images were incorporated using the
ZEN 2011 software (Zeiss Efficient Navigation 2011).
Toluidine blue staining.
Arabidopsis flowers and ovules in siliques were incubated in vacuo for 24 h in a
fixation medium (formalin:acetic acid:ethanol:H2O = 1:1:9:9). The specimens were
dehydrated in a graded alcohol series of n-butanol:absolute ethanol:H2O = 1:4:5, 2:5:3,
7:10:3, and 50:49:1. The samples were left for 1 h in each grade. The specimens were
then washed with n-butanol:absolute ethanol (3:1) for 1 h and washed twice with
n-buthanol and embedded in paraffin wax. For microscopic observation, the specimens
were cut into 5 to 10 mm sections using a microtome (MTH-1 NK system, Osaka,
Japan). Cut sections were placed on slides and subjected to deparaffinization with
xylene for 2× 60 min. Then the slides were transferred to a 1:1 mixture of xylene and
absolute ethanol for 15 min followed by a series of graded ethanol 95%, 85%, 70%,
50%, and 30%. The specimens were left for 5 min in each ethanol grade. After washing
with H2O, the specimens were air dried and used for the staining with 0.05% toluidine
blue O (Wako Pure Chemicals, Osaka, Japan). The stained specimens were examined
by the microscopic visualization using an IX71 system (Olympus).

70
Lipid order
The calculation of the GP images obtained from ratiometric di-4-ANEPPDHQ
fluorescence imaging was performed using ImageJ (http://imagej.nih.gov/ij/) following
the procedure described by Owen et al. (2012) and using their custom-written macro.
The ordered (490–580 nm) and disordered (615–750 nm) phase fluorescence channels
were assigned ch00 and ch01, respectively. The threshold value for the analysis was
fixed at 15, the color scale for the output GP images was set to ’16-color’, and no
immunofluorescence mask was selected. The ImageJ macro for GP analysis generates
GP images from ordered and disordered channel images based on the following
equation:
where I represents the intensity in each pixel in the image acquired in the indicated
spectral channel (numbers in nm). To compensate for differences in the efficiency of
collection in the two channels, GP values were corrected using a G factor. In order to
obtain the G factor, the same microscope set-up employed for imaging root samples
was used to image the fluorescence of a drop of undiluted di-4-ANEPPDHQ stock
solution (2 l) at three laser powers (0.3%, 0.5% and 1%). The mean pixel intensities of
the channels ch00 (ordered) and ch01 (disordered) were extracted in ImageJ, and
corresponding GPmes values were calculated (using the previous equation with G = 1).
The G factor was then calculated according to the equation:
GPref is a reference value for di-4-ANEPPDHQ in DMSO, here fixed at -0.85 (Owen et al.,
2012). In this study, the G factor was defined as G =0.68.
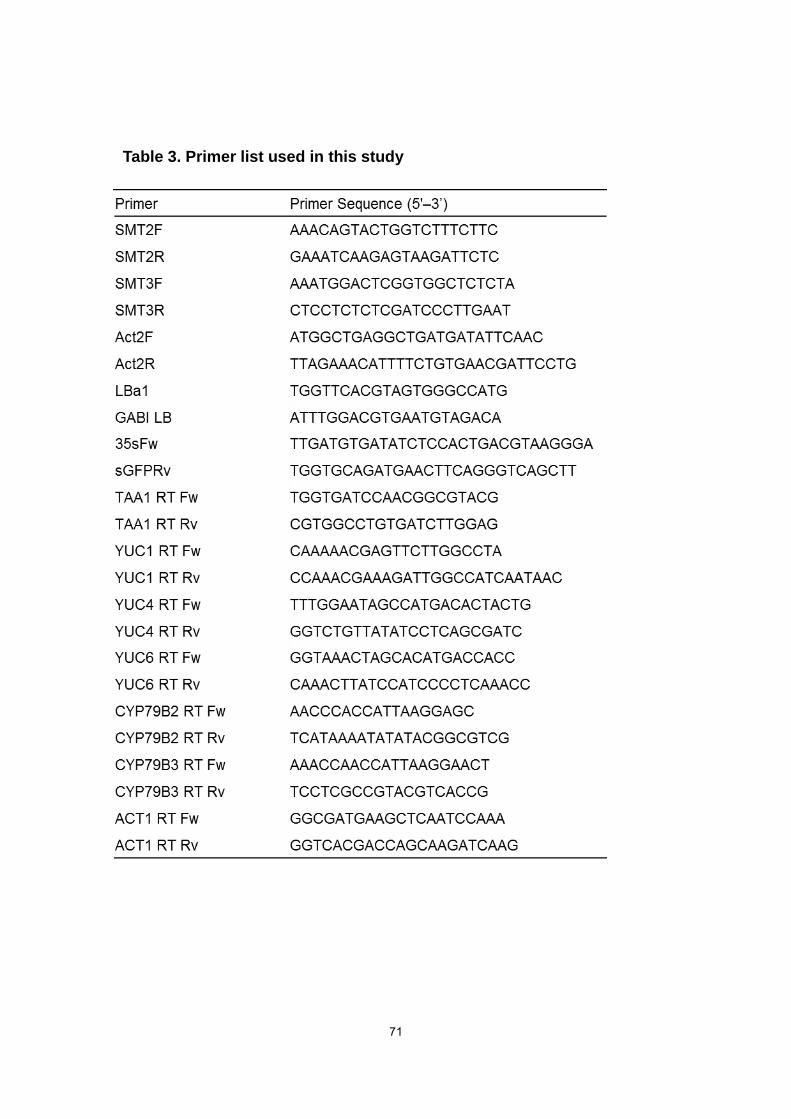
71
Table 3. Primer list used in this study

72
References
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P., Stevenson,
D.K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C., Heller, C., Jeske, A.,
Koesema, E., Meyers, C.C., Parker, H., Prednis, L., Ansari, Y., Choy, N., Deen, H.,
Geralt, M., Hazari, Formatted,N., Hom, E., Karnes, M., Mulholland, C., Ndubaku, R.,
Schmidt, I., Guzman, P., Aguilar-Henonin, L., Schmid, M., Weigel, D., Carter, D.E.,
Marchand, T., Risseeuw, E., Brogden, D., Zeko, A., Crosby, W., Berry, C.C. and
Ecker, J.R. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana.
Science, 301, 653-657.
Ambrose, C., Ruan, Y., Gardiner, J., Tamblyn, L.M., Catching, A., Kirik, V., Marc, J.,
Overall, R. and Wasteneys, G.O. (2013) CLASP interacts with sorting nexin 1 to link
microtubules and auxin transport via PIN2 recycling in Arabidopsis thaliana. Dev.
Cell, 24, 649-659.
Benveniste, P. (2004) Biosynthesis and accumulation of sterols. Annu Rev Plant Biol,
55, 429-457.
Blilou, I., Xu, J., Wildwater, M., Willemsen, V., Paponov, I., Friml, J., Heidstra, R., Aida,
M., Palme, K. and Scheres, B. (2005) The PIN auxin efflux facilitator network
controls growth and patterning in Arabidopsis roots. Nature, 433, 39-44.
Boutté, Y., Frescatada-Rosa, M., Men, S., Chow, C.M., Ebine, K., Gustavsson, A.,
Johansson, L., Ueda, T., Moore, I., Jürgens, G. and Grebe, M. (2010) Endocytosis
restricts Arabidopsis KNOLLE syntaxin to the cell division plane during late
cytokinesis. EMBO J., 29, 546-558.
Carland, F.M., Fujioka, S. and Nelson, T. (2010) The sterol methyltransferases SMT1,
SMT2, and SMT3 influence Arabidopsis development through nonbrassinosteroid
products. Plant Physiol., 153, 741-756.

73
Carland, F.M., Fujioka, S., Takatsuto, S., Yoshida, S. and Nelson, T. (2002) The
identification of CVP1 reveals a role for sterols in vascular patterning. Plant Cell, 14,
2045-2058.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J., 16,
735-743.
Clouse, S.D. (2002) Arabidopsis Mutants Reveal Multiple Roles for Sterols in Plant
Development. Plant Cell, 14, 1995-2000.
Curtis, M.J. and Hays, J.B. (2007) Tolerance of dividing cells to replication stress in
UVB-irradiated Arabidopsis roots: requirements for DNA translation polymerases eta
and zeta. DNA Repair., 6, 1341-1358.
Darnet, S. and Rahier, A. (2004) Plant sterol biosynthesis: identification of two distinct
families of sterol 4alpha-methyl oxidases. Biochem. J., 378, 889-898.
DeBolt, S., Scheible, W.R., Schrick, K., Schaller, H., Beisson, F., Bischoff, V.,
Bouvier-Nave, P., Li, Y., Nair, M., Carroll, A., Hematy, K., Li, Y., Milne, J., Nair, M.,
Schaller, H., Zemla, M. and Somerville, C. (2009) Mutations in UDP
glucose:sterol-glucosyltransferase in Arabidopsis cause transparent testa
phenotype and suberization defect in seeds. Plant Physiol., 151, 78-87.
Derksen, J., Janssen, G.J., Wolters-Arts, M., Lichtscheidl, I., Adlassnig, W., Ovecka, M.,
Doris, F. and Steer, M. (2011) Wall architecture with high porosity is established at
the tip and maintained in growing pollen tubes of Nicotiana tabacum. Plant J., 68,
495–506.
Desmond, E. and Gribaldo, S. (2009) Phylogenomics of Sterol Synthesis: Insights into
the Origin, Evolution, and Diversity of a key Eukaryotic Feature. Genome Biol. Evol.,
1, 364-381.

74
Dhonukshe, P., Tanaka, H., Goh, T., Ebine, K., Mähönen, A.P., Prasad, K., Blilou,
I., Geldner, N., Xu, J., Uemura, T., Chory, J., Ueda, T., Nakano, A., Scheres, B.
and Friml, J. (2008) Generation of cell polarity in plants links endocytosis, auxin
distribution and cell fate decisions. Nature, 456, 962-966.
Diener, A.C., Li, H., Zhou, W., Whoriskey, W.J., Nes, W.D. and Fink, G.R. (2000) Sterol
methyltransferase 1 controls the level of cholesterol in plants. Plant Cell, 12,
853-870.
Dufourc, E.J. (2008) Sterols and membrane dynamics. J. Chem. Biol., 1, 63-77.
Dyer, R.G., Hetherington, C.S., Alberti, K.G.M.M. and Laker, M.F. (1995) Simultaneous
measurement of phytosterols (campesterol and β-sitosterol) and 7-ketocholesterol in
human lipoproteins by capillary column gas chromatography. J. Chromatography B.,
663, 1-7.
Fenner, G.P. and Patterson, G.W. (1992) 24-ethyl-orientation of the 24-ethylsterols
during the life cycle of the squash. Phytochem., 31, 73-75.
Feraru, E. and Friml, J. (2008) PIN polar targeting. Plant Physiol., 147, 1553-1559.
Frescatada-Rosa, M., Stanislas, T., Backues, S.K,, Reichardt, I, Men, S., Boutté, Y.,
Jürgens, G., Moritz, T., Bednarek, S.Y., and Grebe, M. (2014) High lipid order of
Arabidopsis cell-plate membranes mediated by sterol and DYNAMIN-RELATED
PROTEIN1A function. Plant J., 80, 745-757.
Fujioka, S. and Yokota, T. (2003) Biosynthesis and metabolism of brassinosteroids.
Annu. Rev. Plant Biol., 54, 137-164.
Gachotte, D., Eckstein, J., Barbuch, R., Hughes, T., Roberts, C. and Bard, M. (2001) A
novel gene conserved from yeast to humans is involved in sterol biosynthesis. J.
Lipid Res., 42, 150-154.

75
Gasser, C.S., Broadhvest, J. and Hauser, B.A. (1998) Genetic analysis of ovule
development. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49, 1-24.
Geldner, N., Anders, N., Wolters, H., Keicher, J., Kornberger, W., Muller, P., Delbarre,
A., Ueda, T., Nakano, A. and Jürgens, G. (2003) The Arabidopsis GNOM ARF-GEF
mediates endosomal recycling, auxin transport, and auxin-dependent plant growth.
Cell, 112, 219-230.
Geldner, N., Friml, J., Stierhof, Y.D., Jürgens, G. and Palme, K. (2001) Auxin transport
inhibitors block PIN1 cycling and vesicle trafficking. Nature, 413, 425-428.
Grebe, M., Xu, J., Mobius, W., Ueda, T., Nakano, A., Geuze, H.J., Rook, M.B. and
Scheres, B. (2003) Arabidopsis sterol endocytosis involves actin-mediated
trafficking via ARA6-positive early endosomes. Curr. Biol., 13, 1378-1387.
Grosjean, K., Mongrand, S., Beney, L., Simon-Plas, F., and Gerbeau-Pissot, P. (2015)
Differential effect of plant lipids on membrane organization: specificities of
phytosphingolipids and phytosterols. J. Biol. Chem. 290, 5810-5825.
Guo, D., Venkatramesh, M. and Nes, W.D. (1995) Developmental regulation of sterol
biosynthesis in Zea mays. Lipids, 30, 203-219.
Jaideep, M., Csaba, K. and László, S. (1995) A simple method for isolation, liquid
culture, transformation and regeneration of Arabidopsis thaliana protoplasts. Plant
Cell Rep., 14, 221-226.
Jaillais, Y., Fobis-Loisy, I., Miège, C., Rollin, C. and Gaude, T. (2006) AtSNX1 defines
an endosome for auxin-carrier trafficking in Arabidopsis. Nature, 443, 106-109.
Jang, J.C., Fujioka, S., Tasaka, M., Seto, H., Takatsuto, S., Ishii, A., Aida, M., Yoshida,
S. and Sheen, J. (2000) A critical role of sterols in embryonic patterning and
meristem programming revealed by the fackel mutants of Arabidopsis thaliana.
Genes Dev., 14, 1485-1497.

76
Jefferson, R.A., Kavanagh, T.A. and Bevan, M.W. (1987) GUS fusion: -glucuronidase
as a sensitive and versatile gene fusion marker in higher plants. EMBO J., 16,
3901-3907.
Kazłowska, K., Lin, H.T., Chang, S.H. and Tsai, G.J. (2013) In Vitro and In Vivo
Anticancer Effects of Sterol Fraction from Red Algae Porphyra dentata. Evid. Based
Complement. Alternat. Med., 2013, 493869.
Kelley, R.I. and Herman, G.E. (2001) Inborn errors of sterol biosynthesis. Annu Rev
Genomics Hum Genet., 2, 299-341.
Kleine-Vehn, J. and Friml, J. (2008) Polar targeting and endocytic recycling in
auxin-dependent plant development. Annu. Rev. Cell. Dev. Biol., 24, 447-473.
Kleine-Vehn, J., Leitner, J., Zwiewka, M., Sauer, M., Abas, L., Luschnig, C. and Friml, J.
(2008) Differential degradation of PIN2 auxin efflux carrier by retromer-dependent
vacuolar targeting. Proc. Natl. Acad. Sci. U S A, 105, 17812-17817.
Kumagai, F., Yoneda, A., Tomida, T., Sano, T., Nagata, T. and Hasezawa, S. (2001)
Fate of nascent microtubules organized at the M/G1 interface, as visualized by
synchronized tobacco BY-2 cells stably expressing GFP-tubulin: time-sequence
observations of the reorganization of cortical microtubules in living plant cells. Plant
Cell Physiol., 42, 723-732.
Lauber, M.H., Waizenegger, I., Steinmann, T., Schwarz, H., Mayer, U., Hwang, I.,
Lukowitz, W. and Jürgens, G. (1997) The Arabidopsis KNOLLE protein is a
cytokinesis-specific syntaxin. J. Cell Biol., 139, 1485-1493.
Lindsey, K., Pullen, M.L. and Topping, J.F. (2003) Importance of plant sterols in pattern
formation and hormone signalling. Trends Plant Sci., 8, 521-525.
Malinsky, J., Opekarová, M., Grossmann, G. and Tanner, W. (2013) Membrane
microdomains, rafts, and detergent-resistant membranes in plants and fungi. Annu.
Rev. Plant Biol., 64, 501-529.

77
McKean, M. and Nes, W.R. (1977) Evidence for separate intermediates in the
biosynthesis of - -alkylsterols in tracheophytes. Phytochem., 16, 683-686.
Men, S., Boutté, Y., Ikeda, Y., Li, X., Palme, K., Stierhof, Y.D., Hartmann, M.A., Moritz,
T. and Grebe, M. (2008) Sterol-dependent endocytosis mediates post-cytokinetic
acquisition of PIN2 auxin efflux carrier polarity. Nat. Cell Biol., 10, 237-244.
Mialoundama, A.S., Jadid, N., Brunel, J., Di Pascoli, T., Heintz, D., Erhardt, M., Mutterer,
J., Bergdoll, M., Ayoub, D., Van Dorsselaer, A., Rahier, A., Nkeng, P., Geoffroy, P.,
Miesch, M., Camara, B. and Bouvier, F. (2013) Arabidopsis ERG28 Tethers the
Sterol C4-Demethylation Complex to Prevent Accumulation of a Biosynthetic
Intermediate That Interferes with Polar Auxin Transport. Plant Cell, 25, 4879-4893.
Neelakandan, A.K., Song, Z., Wang, J., Richards, M.H., Wu, X., Valliyodan, B., Nguyen,
H.T. and Nes, W.D. (2009) Cloning, functional expression and phylogenetic analysis
of plant sterol 24C-methyltransferases involved in sitosterol biosynthesis.
Phytochemistry, 70, 1982-1998.
Nes, W.R. and McKean, M.L. (1977) Biochemistry of steroids and other isopentenoids.
Baltimore, Md: University Park Press.
Nes, W.R., Krevitz, K., Joseph, J., Nes, W.D., Harris, B., Gibbons, G.F. and Patterson,
G.W. (1977) The phylogenetic distribution of sterols in tracheophytes. Lipids, 12,
511-527.
Nes, W.D., Norton, R.A., Crumley, F.G., Madigan, S.J. and Katz, E.R. (1990) Sterol
phylogenesis and algal evolution. Proc. Natl. Acad. Sci. U S A, 87, 7565-7569.
Nguyen, H.M., Baudet, M., Cuiné, S., Adriano, J.M., Barthe, D., Billon, E., Bruley, C.,
Beisson, F., Peltier, G., Ferro, M. and Li-Beisson, Y. (2011) Proteomic profiling of oil
bodies isolated from the unicellular green microalga Chlamydomonas reinhardtii:
with focus on proteins involved in lipid metabolism. Proteomics., 11, 4266-4273.

78
Owen, D.M., Rentero, C., Magenau, A., Abu-Siniyeh, A., and Gaus, K. (2012)
Quantitative imaging of membrane lipid order in cells and organisms. Nat. Protoc. 7,
24–35.
Paciorek, T., Zazímalová, E., Ruthardt, N., Petrásek, J., Stierhof, Y.D., Kleine-Vehn, J.,
Morris, D.A., Emans, N., Jürgens, G., Geldner, N. and Friml, J. (2005) Auxin inhibits
endocytosis and promotes its own efflux from cells. Nature, 435, 1251-1256.
Petrásek, J. and Friml, J. (2009) Auxin transport routes in plant development.
Development, 136, 2675-2688.
Pullen, M., Clark, N., Zarinkamar, F., Topping, J. and Lindsey, K. (2010) Analysis of
vascular development in the hydra sterol biosynthetic mutants of Arabidopsis. PLoS
One., 5, e12227.
Qian, P., Han, B., Forestier, E., Hu, Z., Gao, N., Lu, W., Schaller, H., Li, J. and Hou, S.
(2013) Sterols are required for cell-fate commitment and maintenance of the
stomatal lineage in Arabidopsis. Plant J., 74, 1029-1044.
Rahier, A., Darnet, S., Bouvier, F., Camara, B. and Bard, M. (2006) Molecular and
enzymatic characterizations of novel bifunctional 3beta-hydroxysteroid
dehydrogenases/C-4 decarboxylases from Arabidopsis thaliana. J. Biol. Chem., 281,
27264-27277.
Ramgopal, M. and Bloch, K. (1983) Sterol synergism in yeast. Proc. Natl. Acad. Sci. U S
A, 80, 712-715.
Rasmussen, C.G., Humphries, J.A. and Smith, L.G. (2010) Determination of symmetric
and asymmetric division planes in plant cells. Annu. Rev. Plant Biol., 62, 387-409.
Saeidnia, S., Permeh, P., Gohari, A.R. and Mashinchian-Moradi, A. (2012)
Gracilariopsis persica from Persian Gulf Contains Bioactive Sterols. Iran J. Pharm.
Res., 11, 845-849.

79
Salimova, E., Boschetti, A., Eichenberger, W. and Lutova, L. (1999) Sterol mutants of
Chlamydomonas reinhardtii: Characterisation of three strains deficient in C24(28)
reductase. Plant Physiol. Biochem., 37, 241-249.
Sano, T., Higaki, T., Oda, Y., Hayashi, T. and Hasezawa, S. (2005) Appearance of actin
microfilament ‘twin peaks’ in mitosis and their function in cell plate formation, as
visualized in tobacco BY-2 cells expressing GFP-fimbrin. Plant J., 44, 595-605.
Sato, S., Kato, T., Kakegawa, K., Ishii, T., Liu, Y.G., Awano, T., Takabe, K., Nishiyama,
Y., Kuga, S., Sato, S., Nakamura, Y., Tabata, S. and Shibata, D. (2001) Role of the
putative membrane-bound endo-1,4-beta-glucanase KORRIGAN in cell elongation
and cellulose synthesis in Arabidopsis thaliana. Plant Cell Physiol., 42, 251-263.
Schaeffer, A., Bronner, R., Benveniste, P. and Schaller, H. (2001) The ratio of
campesterol to sitosterol that modulates growth in Arabidopsis is controlled by
STEROLMETHYLTRANSFERASE 2;1. Plant J., 25, 605-615.
Schaller, H. (2003) The role of plant sterols in plant growth and development. Prog.
Lipid Res., 42, 163-175.
Schaller, H. (2004) New aspects of sterol biosynthesis in growth and development of
higher plants. Plant Physiol. Biochem., 42, 465-476.
Schaller, H. (2010) Sterol and steroid biosynthesis and metabolism in plants and
microorganisms. In Comprehensive Natural Products II Chemistry and Biology,
Mander, L. Lui, H.-W. eds, Elsevier: Oxford Vol. 1, pp 755-787.
Schrick, K., Debolt, S. and Bulone, V. (2012) Deciphering the molecular functions of
sterols in cellulose biosynthesis. Front. Plant Sci., 3, 84.
Schrick, K., Fujioka, S., Takatsuto, S., Stierhof, Y.D., Stransky, H., Yoshida, S. and
Jurgens, G. (2004) A link between sterol biosynthesis, the cell wall, and cellulose in
Arabidopsis. Plant J., 38, 227-243.

80
Schrick, K., Mayer, U., Horrichs, A., Kuhnt, C., Bellini, C., Dangl, J., Schmidt, J. and
Jürgens, G. (2000) FACKEL is a sterol C-14 reductase required for organized cell
division and expansion in Arabidopsis embryogenesis. Genes Dev., 14.1471-1484.
Schrick, K., Mayer, U., Martin, G., Bellini, C., Kuhnt, C., Schmidt, J. and Jürgens, G.
(2002) Interactions between sterol biosynthesis genes in embryonic development of
Arabidopsis. Plant J., 31, 61-73.
Sørensen, I., Rose, J.K., Doyle, J.J., Domozych, D.S. and Willats, W.G. (2012) The
Charophycean green algae as model systems to study plant cell walls and other
evolutionary adaptations that gave rise to land plants. Plant Signal Behav., 7,1-3.
Souter, M., Topping, J., Pullen, M., Friml, J., Palme, K., Hackett, R., Grierson, D. and
Lindsey, K. (2002) hydra mutants of Arabidopsis are defective in sterol profiles and
auxin and ethylene signaling. Plant Cell, 14, 1017-1031.
Souter, M.A., Pullen, M.L., Topping, J.F., Zhang, X. and Lindsey, K. (2004) Rescue of
defective auxin-mediated gene expression and root meristem function by inhibition
of ethylene signalling in sterol biosynthesis mutants of Arabidopsis. Planta., 219,
773-783.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. and Kumar, S. (2013) MEGA6:
Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and
Evolution, 30, 2725-2729.
Timme, R.E., Bachvaroff, T.R. and Delwiche, C.F. (2012) Broad phylogenomic sampling
and the sister lineage of land plants. PLoS One, 7, e29696.
Topping, J.F., May, V.J., Muskett, P.R. and Lindsey, K. (1997) Mutations in the
HYDRA1 gene of Arabidopsis perturb cell shape and disrupt embryonic and
seedling morphogenesis. Development. 124, 4415-4424.

81
Ulmasov, T., Murfett, J., Hagen, G. and Guilfoyle, T.J. (1997) Aux/IAA proteins repress
expression of reporter genes containing natural and highly active synthetic auxin
response elements. Plant Cell, 9, 1963-1971.
Valvekens, D., Montagu, M.V. and Lusebettens, M.V. (1998) Agrobacterium
tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using
kanamycin selection. Proc. Natl. Acad. Sci. U S A, 85, 5536-5540.
Van Damme, D. (2009) Division plane determination during plant somatic cytokinesis.
Curr Opin Plant Biol., 12, 745-751.
Van Damme, D. and Geelen, D. (2008) Demarcation of the cortical division zone in
dividing plant cells. Cell Biol. Int., 32, 178-187.
Willemsen, V., Friml, J., Grebe, M., van den Toorn, A., Palme, K. and Scheres, B.
(2003) Cell polarity and PIN protein positioning in Arabidopsis require STEROL
METHYLTRANSFERASE1 function. Plant Cell, 15, 612-625.
Xu, J. and Scheres, B. (2005) Dissection of Arabidopsis ADP-RIBOSYLATION
FACTOR 1 function in epidermal cell polarity. Plant Cell, 17, 525-536.
Yang, H., Richter, G.L., Wang, X., Młodzińska, E., Carraro, N., Ma, G., Jenness, M.,
Chao, D.Y., Peer, W.A. and Murphy, A.S. (2013) Sterols and sphingolipids
differentially function in trafficking of the Arabidopsis ABCB19 auxin transporter.
Plant J., 74, 37-47.
Yoo, S.D., Cho, Y.H. and Sheen, J. (2007) Arabidopsis mesophyll protoplasts: a
versatile cell system for transient gene expression analysis. Nat. Protocols, 2,
1565-1572.

82
ACNOREDGEMENTS
I am deeply grateful to my supervisor Professor Daisaku Ohta, Osaka Prefecture
University, for his guidance, invaluable discussion, continuous encouragement, and
critical review in all the time of research for and writing of this thesis.
I wish to express my since gratitude to Professor Nozomu Koizumi, Osaka
Prefecture University, and Professor Koh Aoki, Osaka Prefecture University, for their
invaluable advice and suggestions.
I also wish to express my thanks to Professor Hubert Schaller and member of
CNRS, for valuable guidance and stimulating discussion throughout this work.
I also wish to express my thanks to Dr. Masahiko Hurutani, NAIST, for valuable
guidance and stimulating discussion throughout this work.
I am thankful to Dr. Atsushi Okazawa, Osaka Prefecture University, for kind his kind
suggestions and encouragement throughout the work.
I am thankful to Dr. Takumi Ogawa, Osaka Prefecture University, for kind his kind
suggestions and encouragement throughout the work.
I also thank to member of Laboratory of Metabolomics, Osaka Prefecture University,
for their technical assistance and encouragement throughout the work.
Masatoshi Nakamoto