-;jnopr.niscair.res.in/bitstream/123456789/15389/1/ijbb 37(6) 433-440.pdf · transmembrane topology...
TRANSCRIPT

Indian Journal of Biochemistry & Biophysics VoL 37, Dece mber 2000, pp. 433-440
Addition 0
{ r
p ~ proteins of
Huado ng a ng and Parag R hitni s*
Department of Biochemistry, Bioplly siCS- and Molecular Biology, Iowa State University, Ames IA 5~ Accepted 30 May 2000
III vitro mutagenesi s was used to produce two photosys tem I mutants of the cyanobacter ium SYllechocystis sp. PCC 6803. le mutants HK and HL contai ned hexahi stidyl tags at the C-termini of the PsaK I and PsaL subunits, respecti ve ly. The HK mu tant contai ned wi ld-type amounts of trimeric PS I compl exes, but the level of hexahi st idine-tagged PsaK I was found onl y ten per cent in the PS I complexes and membranes of the wild type level. Therefore, attachment of a tag at the Cterminus interferes wi th the expression or assembly of PsaK 1. In contrast, the HL mutan t contained a simi lar level of tagged PsaL as that in the wi ld type. However, trimeric PS I complexes could not be obtained from thi s strain, indicating that the Cterminus of PsaL is in volved in the formation of PS I trimers. Hexahi stidine-tagged complexes of the HL and HK strains could not bc pu ri fied wi th Nickel-affini ty chromatography, unless photosystem 1 was denatured with urea. demonstrating that tagged C-termini of Psa K I and PsaL were e mbedded inside of the PS I complex. Protec tion of the C-termi nus from trypsi n cleavage fu rther sUP Ro rted thi s conclu sio n. Thus, hi stidine tagging all owed us to demo nstrate role of C-te'rmini of
two proteins of Photosyst::/ -;J C \, ~~
Introduction l participate in trimerization are not known . The Photosys tem I CPS I) is a multi subunit pi gment- complete sequence of Synechocystis sp. PCC 6803
protei n complex located in the thylakoid me mbranes genome revealed the presence of two unlinked psaK of cyanobacteria, green algae and higher plants . It genes l4
. The orf ssr0390 encodes an 86-amino ac id catalyzes elec tron transfer from reduced plastocyanin subunit, which has been identified as the PsaK 1 or cytoc hrome C6 to oxidized ferredoxin or prote in in the PS I pre parati ons. The orf ssr0629 flavodoxin
l-3 Cyanobac terial PS I consists of -100 encodes a prote in with 126 amino ac ids which was
chlorophyll a mol ecules, 10-12 B carotenes, 2 recently identified 15. Both proteins are di spensable for phylloquinones, three [4Fe-4S] clusters and 11 the function ofPS 115
.16
.
prote ins. The PsaA and PsaB subunits form the heterodimeric core of PS I complex, coordinating the majority of cofactors of the electron transfer sys tem and of the antenna system4
. PsaC binds the terminal e lec tron acceptors FA and FB and two [4Fe-4S] centers5
. PsaD provides a ferredoxin-docking site and is essential for the assembly of PsaC and PsaE into the PS I complex. PsaE facilitates the interaction between PS I and fe ITedoxin 6
.7
. Consequently, it is req uired for the cyc lic e lec tron transport8
.9
. PsaF provides the dock ing site for plastocyanin in chloropl asts , but not in cyanobacteria 10 . PsaJ may be required for the organi zation of PsaFII, whereas Psal is required for the normal organi zation of the PsaL subunit in PS ] 12. PsaL is essentia l for the formation of PS I trimers 13
. However, the regions of PsaL that
* Author for correspondence: Phonc 5 15 294 1657 Fax: 515 2940453: E-mail: chilni s@ iastate.edu AbbreviatiollS: OM . dodecyl ~-D-maltos ide; Hi s- tag, hexahi slidyl tag ; PCR, polymerase chain react ion: PS 1, photosystem I
Structure of PS I is known at 4A resolution l 7. This
structure reveals the locations of major e lectron transfer centers. However, the resolution in the peripheral part of PS I is poor. Therefore, the transmembrane topology of the sma ll proteins CPsaK, PsaL and PsaF) could not be described clearly from this structure. W e are us ing biochemical and molecular genetic techniques to probe topology of PS I . 41 8 H h . I ' I proteins'. ere we present an approac 111 w ll C 1
surface-exposure of the C-terminus of two integral membrane prote ins, PsaK and PsaL , was examined by attaching an oligopeptide tag. PsaK 1 contains two hydrophobic domains , which are predicted to form
membrane-spanning a-helices. The N-terminus of PsaKl may be in the lumen l 7
.19 and, therefore, may
have a function in me mbrane translocation of the protein . Therefore, the C-terminu~ of PsaKI was chosen to addition of six histidyl residues. The PsaL subunit may contain two membrane-spanning helices. with a large N-terminal domain that interacts with
434 INDIAN J BIOCI-IEM BIOPI-IYS., VOL. 37, DECEMBER 2000
PsaE protein20. We added histidyl res idues at the C
terminus of PsaL. In this paper, we report the effects of a Hi s-tag on the integrity of the PS I complex and the access ibility of a tag to proteases and nickel column , infer the structural roles of the C-termini .
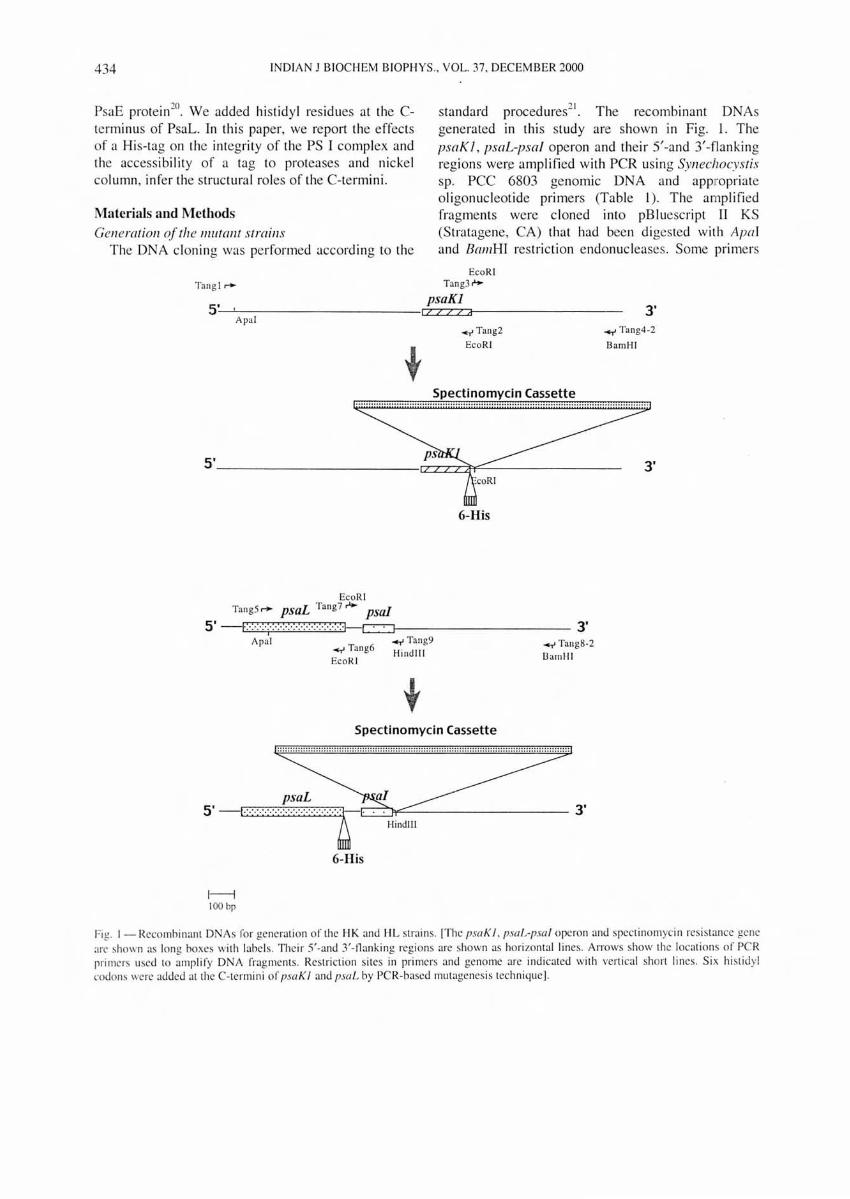
standard procedures21. The recombinant DNAs
generated in this study are shown in Fig. 1. The psaK J, psaL-psa/ operon and the ir 5'-and 3'-flanking regions wen:: amplified with PCR using Synechocystis sp. PCC 6803 genomic DNA and appropriate oligonucleotide primers (Table 1). The amplified fragments were c loned into pBluescript II KS (Stratagene, CA) that had been digested wi th Aprd and BamBI res triction endonucleases. Some primers
Materials and Methods Generation of the IIllItallt strains
The DNA cloning was performed according to the
Tangl ~
5' I
Apal
EcoRI Tang3 '"
psaKl I/ZZid
~Tang2
EcoR I
Spectinomycin Cassette
3' ~ Tang4.2
BamHI
5' ----------------C07~~--------- 3'
EcoR I TangS ~ psaL Tang? .... psal
6-His
5' E. ·:·.·:·;·:·:·:·:·:·:·:·:·:·:·:·:·:·:·I-c=J---------- 3' Apal ~ Tung9
~ Tang6 Hi ndlll EcoRI
Spectinomycin Cassette
~ Tang8-2
IJamH I
I ::: :: :: ::::::: ::: ::::::::::: ::: :::: :::::::::::: ::: :::::::::::::: ::::: :: ::::::::: ::: ::: ::::::::::: :::::::::::: ::::: 1
5 ' -1·········· 3'
1----1 100 bp
6-His
Hindlll
Fig. I - Recombi nant DNAs fo r ge neration of the I-IK and I-IL strains. [The psaK I . p.mL-psa l operon and spectinomycin resistance gene arc shown as long boxes with labels. Thei r 5'-and 3/-nanking regions are shown as hori zon tal lines. Arrows show the locations of PCR primers used to amplify D A fragments. Restriction sites in primers and genome are indicated with vertical short lines. Six hi stidyl codons were added at the C-termini of psaK 1 and psaL by PCR-based mutagenes is technique].

TANG & C HITNIS: ADD ITION OF C-TERMINAL HISTIDYL TAGS TO PsaL AND PsaKI PROTEINS 435
Table I -Oligonucleotide primers used in the constructi on of recombi nant DNAs
Oligonucleotide Sequence Position Purpose
Tangl 5'-CAAGGTTCGCAAATTTCTGG-3' 1265 bps upstream Amplification of the ofpsaKI 5'-flanking of psaK I
EcoRI Hexahistidine Before the stop codon Amplifi cat ion of the Tang2 5' -CGAA TTCTT AGTGGTGGTGGTGGTGGTGAAGT ACTCC-3' of psaKl psaKl and
introduction of an EcoRI site and a hexahistidyl codon sequence before the stop codon
EcoR I 6 bps downstream Amplifi cation of the Tang3 5' -CCGGAATTCATTCTGAATGGACAAGCTTTTATCC-3 ' of psaKI 3'-t1anking of psaKl
and introduction of an EcoR I site
BalllHI 734 bps downstream of Amplitication of the Tang4-2 5'-CGCGGATCCTCGGGATTGTATGGCCTAGTCC-3' psaKl 3' -t1anking of psa K I
and introduction of a BwnH I site
137 bps from start Amplification of the TangS 5'-CCATCCTCAGAGGTTTGGAAG-3' codon of psaL 3'-flanking region of
psaL Before the stop codon Amplification of the
Tang6 EcoRI Hexahi stidine of psaL 3'-flanking region of 5'-CCGAATTCTTAGTGGTGGTGGTGGTGGTGGTTAAATAG-3' psaL and int roduction
of an EcoRI site and a hexahistidyl codon sequence before the stop codon
Eco RI 6 bps downstream Amplifi cat ion ofpsal Tang7 5'-CCGGAATTCATTCTGAATGGACAAGCTTTTATCC-3' of psaL and the 3'-flanking
region of psaL and introduction of an EcoR I site
BamHI 693 bps downstream Ampli ficat ion of the Tang8-2 5'-CGCGGATCC GGCAAATCCAAAGACTGGG3 ' of psal 3'-flanking region of
psal and introduction ofaBalllH I site
! lind lll 26 bps downstream Introduction of a Tang9 5'-CCATTAGTCTTCAAGCTTTCCAGCCTAACC-3' of psal lIindllI site
included mutations to introduce appropriate restrict ion sites in the amplified DNA fragments. The mutagenic primers Tang2 and Tang6, contained sequences
complimentary to the 3' end of the coding region of the psaKJ and psaL genes , respectively. These two primers introduced six histidy l codons at the end of the psaK I and psaL genes.
The spectinomyc in-resistance gene was inserted in the 3'-flanking region of psaK I gene at EcoRI s ite and the 3'-flanking region of psa/ gene at HindIII site which was introduced by primer Tang9. The plasmid DNAs were used to transform the wild type of SYll echocystis sp. PCC 6803 (ref. 22). The spectinomycin resistant transformants were se lected and segregated for five generatioFls . Genomic DNA of mutant strain s was isolated , the psaK I and psaL genes
were amplified by PCR, and the amplified fragments were sequenced to confirm the mutations and to ensure the fide li ty ofTaq DNA polymerase.
Cyanobacterial strains and culture conditions The wild type, HK and HL stra in s of Synechocystis
sp. PCC 6803 (Table 2) were cultured in BG-I I medium with 5 mM glucose at 30°C under medium light intensity (50 flmol m,2s" ). Mutant strains were grown with 20 fl g/ml spect inomyci n. The growth of cultures was monitored by meas uring the absorption at 730 nm with a UV -160U spet rophotometer (Shi madzu, Tokyo, Japan). Cell s were harvested at the late exponential growth phase and resuspended in 0.4 M sucrose, 10 mM NaCI , 10 mM MOPS-HCI (3-N-morpholino-propane sulfonic acid, pH 7.0) and stored at -20°C until their use for membrane isolation.

436 INDIAN J BIOCHEM BIOPHYS., VOL. 37, DECEMB ER 2000
Strain
Table 2 - Strains of the Cyanobacterial SYlleciJocyslis sp. PCC 6803
Description Reference
Wi ld type glucose-tolerant strai n (4)
HL hexahistidyl tagged PsaL mutant strain Thi s work
HK hexahi stidyl tagged PsaK I mutant strain Thi s work
Biochemical methods Thylakoid membranes were isolated from
Synechocystis sp. PCC 6803 cell s after cell breakage with a bead-beater. Trimeric PS I complexes were puri fied from membranes by extraction with 0.3% OM fo llowed by sucrose gradi ent ultracentrifugation"3. Chl orophyll a concentrati ons in the thylakoid membranes and PS I complexes were determined in 80% acetone24
. In some experiments, thylakoid membranes were treated with trypsin (EC 3.4.21.4, Type XI from bov ine pancreas, Sigma). The membranes (200 Il g chi orophyll/ml ) in 50 mM Tri sHCl pH 8 and I mM CaCh were incubated with trypsin (40 pg/ml ) at 37°C for di ffe rent duration and reactions were terminated by adding 20 mM PMSF.
For analyti ca l gel electrophores is and immunodetec tion, thylakoid membranes and PS I complexes were solubili zed at 37°C for 2 hI' with 1% SOS and 0. 1 % OTT. Denatured proteins were resolved by modified Tricine-urea-SOS-PAOEG and were visuali zed by sil ver-s taining. Alternati vely, the proteins were examined by immunodetec ti on4 with polyclonal antibodies aga inst the PsaK I or PsaL proteins of Synechocystis sp. PCC 6803 or with India ™ HisProbe-HRP (Pierce, IL) that recogni zes a Hi s- tag.
Affinity chro111atography His-bind Ni 2
+ metal affi nity res in (Novagen) was washed sequentially with 3 volumes of deioni zed water, 3 volumes of the charge buffer (50 mM NiSO.j), and finall y with 3 volumes of the binding buffe r (5 mM imidazole, 0.5 M NaCI, 20 mM TrisHCl pH 7.9) to charge and equili brate the column. The thylakoid membranes of the wild type, HK and HL strains were dissolved in binding buffer at a fin al chl orophyll concentration of 100 Ilg/ml with 0.03 % OM or 0.3% OM and were applied on a column (5 ml bed volume) . In some experiments, the HL membrane samples were treated with 0.4 M , 1 M , 4 M , or 8 M urea or with 0.5% SOS after solubil iza ti on by OM. The column was washed with 10 volumes of binding buffer and was eluted by 1 xelution buffe r (1 M imidazole, 0.5 M NaCI, 20 mM Tris-HCI pH 7.9) and subsequentl y by 4 x elution buffer.
Results Th e His-tagged mutant strains
To examine the roles of the C-termini of PsaKI and PsaL of PS I, and to potentiall y use the tags for purification by His-tag affinity chromatography, we added six hi stidyl residues at the C-termini of PsaKI and PsaL. In the psaK] mutant strain (HK), spectinomyc in-res istance gene was inserted immediately to the 3'-terminu of the His-tag coding region. Since the psa] and psaL genes use the same promoter, inserti on of the resistant gene in the intergenic region between psaL and psa] could interrupt the transcription of psa], leading to a low amount of PsaI. Stability of the PS I trimers is dependent on the presence of PsaI in Synechocystis sp. PCC 6803 (ref. 12). Therefore, the spectinomyc inres istance cassette was inserted downstream from the psa! gene in the ONA construct used for generati on of the HL strain (Fig. 1). Seq uencing of the DNA fragments amplified by PCR confirmed the presence of tags in the mutant strains (data not shown). When the HK and HL mutant strains were cul tu red in BO 11 with glucose or without glucose, both mu tants showed similar growth rates as that of the wild type (data not shown). Therefore, functi on of mutant PS I complexes does not become a limiting factor for photoautotrophic or mixotrophic growth. Sucrose gradient ultracentri fugati on of OM-solubili zed thylakoids showed that the HK mutant contained similar level of trimeric PS I complexes in the membranes as in of the wild type (data not shown). In contrast, we could not obtain trimeric PS I complexes from the membranes of the HL mutant by sucrose gradient ultracentrifugation.
Accumulation of His-tagged proteins in mllltant membranes and PS I complexes
To examine express ion of tagged proteins In the mutant strains, we used i mmunodetection to investi gate the PsaK and PsaL prote ins. When membranes of the HK mutant strain were used fo r immunodetec ti on with anti-PsaK polyc lonal antibodies, the PsaK protein migrated at a larger apparent mass (5.7 kDa) than in the wild type (5. 1 kDa). This difference represented an addition of HHHHHH sequence at the end of the PsaKI protein , which was expected to increase mass of the tagged protein by 0.6 kDa. On an equal chl orophyll bas is, the level of His-tagged PsaKI in the HK membranes or trimeric PS I complexes was substantially lower than in the wild type trimers. We could not detect PsaK in the monomeric PS I complexes isolated from the HK

TA G & CHITN IS: ADDITION OF C-TERMINAL HISTIDYL TAGS TO PsaL AND PsaKI PROTEINS 437
mutant strain (Fig. 2A). To estimate the level of PsaK in the mutant trimeric complexes, we used different amounts of wi ld type complexes in an immunodetection experiment (Fig. 2B) . Comparison of relative intensities of the band reactive with PsaK antibodies showed that the PS I trimers from the HK strain contained approximately 10% of the wild type level of PS I complexes. A possible reason for observing low intensity for the PsaK band in immunodetection is that the tag interferes with the antibody-PsaK binding. To examine this possibility, we stained prote ins of wild type and mutant complexes with silver staining (Fig. 2C). The wild
A.
k Oa ,..--...,.-----;-..",,:;---;;:-;-=,;-----,
B.
119 <;hlorophyll
5.7 5.1
0.5
WT trimer HK trimer
1.0 1.7 5 5
kD9 5.7 5.1
L-______ _ _ ___ ~
p • .... c.
kOa
H;:,- I'){Jy(;'d f>!..<IK I -+ '.).1 ?s3KI-· .. ~.1
Fig. 2 - Accumulation of Hi s- tagged PsaK I prote ins in the HK strain . (A): hnmunoblotting analysis of the membrane and PS I complexes from HK strain . The proteins from sampl es containing 5 Ilg ch lorophyll were resolved by Tri ci ne/urealS DS-PAGE. The antibody agai nst SYllechocyslis sp. PCC 6803 PsaK I was used to detect the protein and the immunodetection was visuali zed by enhanced chemi luminescence. (B): Rel ati ve quantification of the levels of the Hi s-tagged PsaK I. Different amounts of wild type PS I complexes were used for gel electrophoresis and immunodetection with a polyclonal antibody against PsaK I. (C): Silver-stained polyacrylamide gel on which proteins from the membranes and PS I complexes of the HK strain were separated . Samples containing 5 Ilg chlorophyll were used for electrophoresi s.
membrane
WT HL kOa
15.0-J 14.4-1
~--------------~
Fig. 3 - Western blotting analysis of the membrane from HL strain. [Proteins of the HL membrane and WT membrane (5 Ilg chlorophyll) were separated by Tricine/SDS-PAGE and immunodetection was performed with the antibody against SYllechocyslis sp. PCC 6803 PsaL and enhanced
chemiluminescence).
type trimers contained PsaK as a 5.1 kDa protein . The HK trimers and membranes lacked this protein, but contained a 5.7 kDa protein, corresponding to the tagged PsaK. The relative intensity of this band was approximately 1/1Oth of the wild type. The monomeric PS I complexes did not contain the 5.7 kDa protein . The other proteins of PS I were present at similar levels in the wild type and mutant strains. The wild type PS I complexes contain equimolar ratio of PS I subunits. Therefore, 90% of PS I trimers in the HK strain do not contain PsaK proteins.
Immunodetection with a PsaL-specific antibody showed that the m.. membranes contained the wildtype level of PsaL (Fig. 3). The size of the His-tagged PsaL was larger than the wild-type PsaL, accounting for the addition of six histidyl residues at the Cterminus of the protein . These results showed that six histidyl residues at the C-terminus of PsaL did not affect the integration of PsaL into membranes, but disallowed formation of stable PS I trimers. Therefore , the C-terminus of the PsaL subunit is important for trimer formation.
Nickel-affinity chromatography of the His-tagged PS I complexes
The HK and m.. strain contained PS I complexes with His-tagged proteins. Therefore, we attempted to use Ni-affinity chromatography to purify these PS I complexes . When DM-solubilized thylakoid membranes of the HK and m.. mutants were applied on the nickel-column, none of the PS I complexes could bind to the column (data not shown). Thi s could result from inaccessibility of the hexahi stidine terminus to Ni on the column. To test this possibility we treated the PS I complexes of the m.. strain with SDS (Fig. 4A, lanes 1-3) or with urea (Fig. 4A, lanes 5-6) prior to chromatography. Denaturati on of complexes by an ionic detergent or a chaotrope a llowed binding of the His-tagged PsaL to nickelcolumn (Fig. 4A). The PS I complexes bound to the
438 INDIAN J BIOCHEM BIOPHYS. , VOL. 37, DECEMBER 2000
column could be eluted by addition of imidazole. With the increasing concentration of urea, the His-tag was more likely to be exposed to the surface due to the protein denaturation. Thus more PS I could be bound to nickel-column (Fig. 4B). This result indicated that the Hi s-tag in the PS I preparations of the HI... strain may not accessible for binding to nickel column in the intact complexes.
Exposure of the C-terminus of PsaL Inability of the intact PS I complexes from the HI...
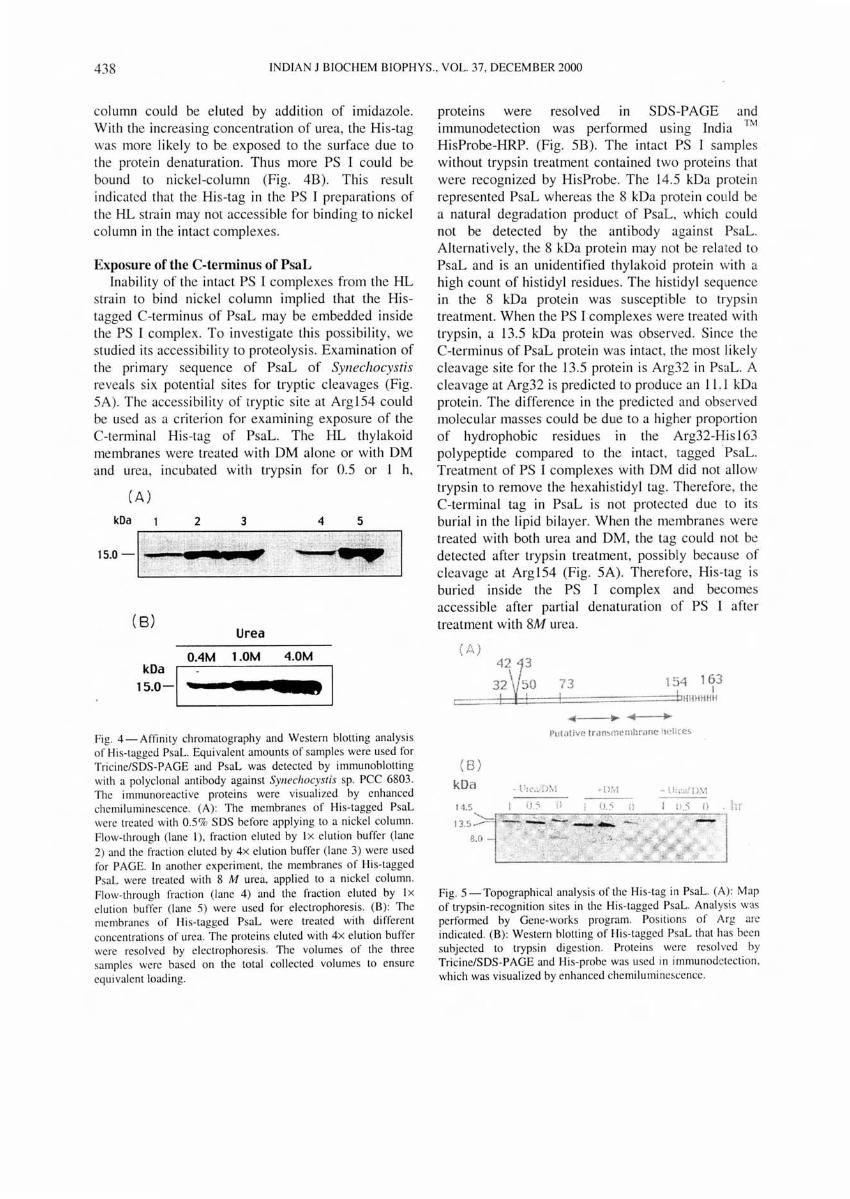
stra in to bind nickel column implied that the Histagged C-terminus of PsaL may be embedded inside the PS I complex. To investigate this possibility, we studied its accessibility to proteolysis. Examination of the primary sequence of PsaL of SYllechocystis reveals six potential sites for tryptic cleavages (Fig. SA). The accessibi li ty of tryptic site at ArglS4 cou ld be used as a criterion for examining exposure of the C-terminal His-tag of PsaL. The HI... thylakoid membranes were treated with DM alone or with DM and urea, incubated with trypsin for 0.5 or 1 h,
(A)
kOa 2 3 4 5
15.0 -I~~"'."_· I
( 8) Urea
DAM 1.0M 4.DM
kDa I 15.0-. I III
Fig. 4 - Aftinity chromatography and Western blotting analysis of His-tagged PsaL. Equivalent amounts of samples were used for Tricine/SDS-PAGE and PsaL was detected by immunoblotting with a polyclonal an tibody against SYllechocyslis sp. PCC 6803. The i mmunoreacti ve proteins were visualized by enhanced chemiluminescence. (A): The membranes of His-tagged PsaL were treated with 0.5% SDS before applying to a nickel column. Flow-th rough (lane I), fraction eluted by I x elution buffer (lane 2) and the fraction eluted by 4x elution buffer (lane 3) were used for PAGE. In another experiment, the membranes of His-tagged PsaL were treated with 8 M urea, applied to a nickel column. Flow-through fraction (lane 4) and the fraction eluted by Ix elution buffer (lane 5) were used for electrophoresis. (B): The membranes of His-tagged PsaL were treated with different concentrations of urea. The proteins elu ted with 4x elution buffer were resolved by e lectrophoresis. The volumes of the three samples were based on the total collected volumes to ensure equivalent loading.
proteins were resolved in SDS-PAGE and immunodetection was performed using India ™ HisProbe-HRP. (Fig. SB). The intact PS I samples without trypsin treatment contained two proteins that were recognized by HisProbe. The 14.5 kDa protein represented PsaL whereas the 8 kDa protein could be a natural degradation product of PsaL, wh ich could not be detected by the antibody against PsaL. Alternatively, the 8 kDa protein may not be related to PsaL and is an unidentified thylakoid protein with a high count of histidyl residues. The histidyl sequence in the 8 kDa protein was susceptible to trypsin treatment. When the PS I complexes were treated with trypsin, a 13.5 kDa protein was observed. Since the C-terminus of PsaL protein was intact , the most like ly cleavage site for the 13.5 protein is Arg32 in PsaL. A cleavage at Arg32 is predicted to produce an 11 .1 kDa protein. The difference in the predicted and observed molecular masses could be due to a higher proportion of hydrophobic residues in the Arg32-His 163 polypeptide compared to the intact, taggedPsaL. Treatment of PS I complexes with DM did not allow trypsin to remove the hexahi stidyl tag. Therefore, the C-terminal tag in PsaL is not protected due to its burial in the lipid bilayer. When the membranes were treated with both urea and DM, the tag cou ld not be detected after trypsin treatment , possibly because of cleavage at Arg lS4 (Fig. SA). Therefore, His-tag is buried inside the PS I complex and becomes accessible after partial denaturation of PS I after treatment with 8M urea.
( A)
(8) kDa 14.5
8.0
73 I
154 163 , I
... ·=:=:t"IHll 11 HH If
I'ulatilfc troll,memhrane iwllccs
. hI'
Fig. 5 - Topographical analysis of the His-tag in PsaL. (A) : Map of trypsin-recognition sites in the His-tagged PsaL. Analys is was performed by Gene-works program. Positions of Arg are indicated. (B) : Western blotting of Hi s- tagged PsaL that has been subjected to trypsin digestion. Proteins were resolved by Tricine/SDS-PAGE and His-probe was used in immunodetection , which was visualized by enhanced chemi luminescence.

TANG & CHITNIS: ADDITION OF C-TERMINA L HISTIDYL TAGS TO PsaL AND PsaKI PROTEINS 439
Discussion PsaKl contains two hydrophobic regions that could
fo rm transmembrane hel ices. This protein has been proposed to occupy a position near the region fac ing the adj oining monomerl? Function of PsaK has not been identified. The absence of PsaK 1 had no significant effect on photoautotropic growth, the amount of chl orophyll per cell , or the trimer formationl6. When we added a His-tag at the Cterminus of PsaK 1, it greatl y reduced level of PsaK 1 in the PS I complexes, but had no effect on cell growth and trimer formation. This result was consistent with the phenotype of the PsaK I-less mutantsl5. Therefore, the C-terminus of PsaKl is important for expression or assembl y of PsaKl. The polar Hi s-tag at the hydrophobic C-terminus could result in weaker interac ti ons between the His-tagged PsaK 1 and the PS I complex. Nickel-affinity chromatography of the solubili zed HK PS I complexes showed that Hi s-tagged PsaKI could not be bound to the column (data not shown). Since nickel chelate affinity constant fo r poly-hi stidine is Ka= IOIJ M, binding can occur even in ex treme cond it ions such as in 8 M urea or for insoluble proteins25
. The most likely reason for the affi nity bloc king of the Hi s-tag is that the C-terminus of assembl ed PsaK 1 is embedded inside PS I complex. If the C-tenninus is fo lded inside the PS I complex, the attached six hi stidyl res idues could interac t with the other proteins of PS I reducing its stability compared to the wild type PsaK I, res ulting in a lower percentage of assembled Hi s-tagged PsaK. Hi s-tagged PsaK 1 was not present in the monomeri c PS I complexes, also indicating the C-tenninus disturbs the in terac ti ons between the PsaK 1 and the PS I complex. On the other hand , the trimeric configuration may stabili ze PsaK I in the PS I complex.
Functi on of PsaL has been revealed by the inacti vati on of the psaL gene. Trimeric PS I complexes ca nnot be obtained from the PsaL-less
I, 0
mutant · . In 4A crys tal structure, there are three transmembrane helices near the C3 ax is26. Two helices in volved in the trimer contacts are ass igned to PsaL whereas the th ird heli x is ass igned to PsaI. Histagged PsaL acc umulated in PS I complex at a similar level as in the wild type, but it did not all ow fo rmati on of PS I trimers. This result showed that the C-terminus of PsaL is in volved in the trimer format ion. The six hi stidyl residues may change the hydrophobicity of the C-terminus or block certain residues of the C-terminus from interacting with the
other components of the PS I complex. The Histagged C-terminus in the solubilized membrane was inaccess ible to the digestion with trypsin and could not be bound to chelate-affinity column . These results showed that the C-terminus is located inside the PS I complex. Such an orientation may explain that the PS I monomers of a trimer are bridged, at least partiall y, by the C-terminus. Interestingly, fro m our results, arginyl residues at positions 42, 43, 50, 73, 154 of PsaL were inaccessible to tryps in digestion, indicating that the regions containing these residues, which are predicted to be in the extra membrane loops, are embedded inside the PS I complex.
In recent years, cyanobacteria have been used extensively to study structure-function relati ons in photosynthetic complexes, including PS 123 . Many mutations cause reduced levels of the mutant complexes in the cell. Consequentl y, puri fication of complexes is di ffi cult , pos ing severe restraints on the analytical techniques that can be used in characterizing the PS I complexes. In such cases, attachment of hi stidyl tag to the protein under study has prov ided an effecti ve solutionn . In thi s approach a stretch of 6-10 hi stidyl residues is added to a protei n using recombinant DNA techniques. The tagged protein is produced in a suitable host and purified using Ni-affinity chromatography. Small protein tags can also be used in topographical analysi s of proteins28. In thi s paper, we described generat ion of cyanobacterial st rains with a hexahi stidyl tag attac hed to PS I proteins. However, the tag was found to interfere with the express ion of assembl y of PsaK 1 and was buried in the complex when attached to the C-terminus of PsaL. Therefore, the strains generated here were not useful in puri fica tion of intact PS I complexes using nickel affinity chromatography.
To conclude, our res ults indicate that the C-termini of PsaK and PsaL are buried inside PS I complex. In additi on, we show that additi on of hexahi stidyl tag at the C-terminus of PsaK 1 reduces synthesis or assembly of PsaK 1. In contrast, the tag at the Cterminus of PsaL all ows assembl y of PsaL into PS I complexes but inhibits formati on of PS I trimers.
Acknowledgement This work was supported, in part , by grant from the
US Nati onal Science Foundation and the Nati onal Institutes of Health and by the Hatch Act and State of Iowa funds. This is Journal Paper No. J-XXXXX of the Iowa Agriculture and Home Economics Experiment Station, Ames, Iowa, Project No. 341 6.

440 INDIAN J BIOCHEM BIOPHYS ., VOL. 37, DECEMBER 2000
References I Chitnis P R, Xu Q, Chitnis V P & Nechushtai R ( 1995)
Photosynth Res 44, 23-40 2 Fromme P ( 1996) ClIrr Opin SII'IICI Bioi 6, 473-484 3 Chitnis P R ( 1996) Plalll Physiol I I 1,66 I -669 4 Sun J, Xu Q, Chitnis V P, Jin P & Chitnis P R ( 1997) J Bioi
Chelll 272 , 2 I 793-2 I 802 5 Oh-oka H, Takahashi Y, Kuriyama K, Saeki K & Matsubara
H ( 1988) J Biochelll 103, 962-968 6 XU Q, Jung Y S, Chitnis V P, Guikema J A, Golbeck ] H &
Chitnis P R ( 1994) J Bioi Chelll 269. 2 I 5 I 2-2 I 518 7 Rousseau F, Setif P & Lagoutte B ( 1993) EMBO J 12, 1755-
1765 8 Chitnis P R, Chitni s V P, Xu Q, Jung Y -S , Yu L & Golbeck,
J H ( 1995) in Photosynthesis: frolll Light 10 Biosphere (Mathis P, ed) Vol II , pp 17-22, Kluwer, Dodrecht
9 Yu L. Zhao J, Muhlenhoff U. Bryant D A & Golbeck , J H (1993 ) Planl Physio/ 103. 171 - 180
10 Chitnis P R. Purvis D & Nelson N ( 1991 ) J Bioi Chell' 266, 20 146-20151
II Xu Q. Odom W R. Guikema J A. Chitnis V P & Chitnis P R ( 1994) Plant Mol Bioi 26. 291-302
12 XU Q, Hoppe D, Chitnis V P, Odom W R, Guikema J A & Chitnis P R ( 1995) J Bioi Chelll 270, 16243-16250
13 Chitnis V P & Chitnis P R ( 1993) FEBS Le1l336. 330-334 14 Kaneko T, Sato S. Kotani . H. Tanaka A, Asamizu E.
Nakamura Y. Mi yajima N. Hirosawa M, Sugiura M. Sasamoto S. Kimura T. Hosouchi T. Matsuno A, Muraki A.
akazaki N, aruo K. Okumura S, Shimpo S, Takeuchi C,
Wada T, Watanabe A, Yamada M, Yasuda M & Tabata, S ( 1996) DNA Res 3,109-136
15 Naithani S, Hou J M & Chitnis P R (2000) Photosynlh Res (in Press)
16 Nakamoto H & Hasegawa M ( 1999) Planl Cell Physiol 40, 9-16
17 Schubert W -D, Klukas 0, KrauS N, Saenger W, Fromme P & Witt H T (1997) J Mol Bioi 272, 741-769
18 Xu Q & Chitnis P R ( 1995) Planl Physioll08, 1067-W75 19 Gavel Y, Steppuhn J, Herrman n R & Von Heijne, G ( 1991 )
FEBS Lell 282, 41-46 20 XU Q, Armbrust T S. Guikema J A & Chi tni s P R ( 1994)
Planl Physioll06. 1057-1063 2 1 Maniatis T, Fritsch E F & Sambrook, J ( 1982) MoleclIlar
cloning: A laboralory lIIal//((/ l, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
22 Wi lli ams J G K ( 1988) Methods EnZYlllol 167,766-778 23 Sun J, Ke A, Jin P, Chitnis V P & Chitnis P R ( 1998) Methods
EnZylllol 297, 124-139 24 Arnon D ( 1949) Plalll Physiol 24, 1- 14 25 Schmitt J H H & Stunnenberg H G ( 1993) Mol Bioi Rep 18,
223-30 26 KrauS N. Schubert W -D. Klukas O. Fromme P. Witt H T &
Saenger W ( 1996) Natllre Stmcl Bioi 3( 11 ).965-973 27 Bricker T. Morvant J. Masri N. Sutton H & Frankel L ( 1998 )
Biochilll Biophys Acta 1049, 50-57 28 Cramer W A, Martinez S E. Huang D, Tae G S, Everly, R M,
Heymann J B, Cheng R H, Baker T S & Smith J L ( 1994) J Bioenerg Biolllelllbr 26, 3 1-47